Hypoxia signaling pathways: modulators of oxygen-related organelles
 Miriam J. Schönenberger
Miriam J. Schönenberger Werner J. Kovacs
Werner J. Kovacs- Department of Biology, Institute of Molecular Health Sciences, ETH Zurich, Zurich, Switzerland
Oxygen (O2) is an essential substrate in cellular metabolism, bioenergetics, and signaling and as such linked to the survival and normal function of all metazoans. Low O2 tension (hypoxia) is a fundamental feature of physiological processes as well as pathophysiological conditions such as cancer and ischemic diseases. Central to the molecular mechanisms underlying O2 homeostasis are the hypoxia-inducible factors-1 and -2 alpha (HIF-1α and EPAS1/HIF-2α) that function as master regulators of the adaptive response to hypoxia. HIF-induced genes promote characteristic tumor behaviors, including angiogenesis and metabolic reprogramming. The aim of this review is to critically explore current knowledge of how HIF-α signaling regulates the abundance and function of major O2-consuming organelles. Abundant evidence suggests key roles for HIF-1α in the regulation of mitochondrial homeostasis. An essential adaptation to sustained hypoxia is repression of mitochondrial respiration and induction of glycolysis. HIF-1α activates several genes that trigger mitophagy and represses regulators of mitochondrial biogenesis. Several lines of evidence point to a strong relationship between hypoxia, the accumulation of misfolded proteins in the endoplasmic reticulum, and activation of the unfolded protein response. Surprisingly, although peroxisomes depend highly on molecular O2 for their function, there has been no evidence linking HIF signaling to peroxisomes. We discuss our recent findings that establish HIF-2α as a negative regulator of peroxisome abundance and suggest a mechanism by which cells attune peroxisomal function with O2 availability. HIF-2α activation augments peroxisome turnover by pexophagy and thereby changes lipid composition reminiscent of peroxisomal disorders. We discuss potential mechanisms by which HIF-2α might trigger pexophagy and place special emphasis on the potential pathological implications of HIF-2α-mediated pexophagy for human health.
Introduction
Life with oxygen (O2) began around 2.4 billion years ago, when photosynthetic organisms prospered and multiplied, leading to a progressive increase of atmospheric O2. O2-related organelles, such as mitochondria, peroxisomes, and plastids, must have been acquired after that date (De Duve, 2007; Semenza, 2007). The appearance of O2 was one of the defining moments in evolution, as it offered organisms the advantage of generating energy more efficiently. Because of the high energy potential of O2, aerobic organisms have become dependent on this gas for their performance and survival. O2 is an essential substrate in cellular metabolism, bioenergetics, and signaling and as such inseparably linked to the survival and normal function of all metazoans. Hence, aerobic species developed mechanisms to sense O2 levels and regulate O2 consumption, in order to cope with conditions of insufficient O2 supply. This Review focuses on the role of hypoxia-inducible factors (HIFs) as master regulators of O2 homeostasis and, in particular, on recent advances in understanding their roles in regulating major O2-consuming organelles, namely mitochondria, the endoplasmic reticulum (ER), and peroxisomes.
Regulation of HIFs
Central to the molecular mechanisms underlying O2 homeostasis are HIF-1α and HIF-2α that function as master regulators of the adaptive response to hypoxia. HIFs form a heterodimer consisting of a constitutively expressed ARNT/HIF-1β subunit and O2-regulated α subunits (HIF-1α or EPAS1/HIF-2α) (Majmundar et al., 2010; Keith et al., 2012). A third HIF-α subunit (HIF-3α) has also been described. HIF3A mRNA is differentially spliced to produce multiple HIF-3α isoforms that either promote or inhibit the activity of other HIF complexes (Keith et al., 2012). Under normoxia, HIF-α subunits are hydroxylated by prolyl hydroxylases (PHD1-3) and recognized and targeted for proteasomal degradation by the von Hippel-Lindau (VHL) E3 ubiquitin ligase complex (Figure 1A). PHD enzymes are 2-oxoglutarate- and iron-dependent dioxygenases, whose activity is absolutely dependent on O2. Hence, the rate of HIF-α hydroxylation is suppressed under hypoxia. Hypoxia or loss of functional VHL stabilizes HIF-α subunits. HIF-α either dimerizes with HIF-1β and binds to hypoxia-responsive elements in promoters of target genes to promote a concerted transcriptional response (Keith et al., 2012) (Figure 1A) or it physically interacts with other non-HIF proteins (Uniacke et al., 2012; Hubbi et al., 2013), enabling convergence of HIF O2 sensing with other signaling pathways.
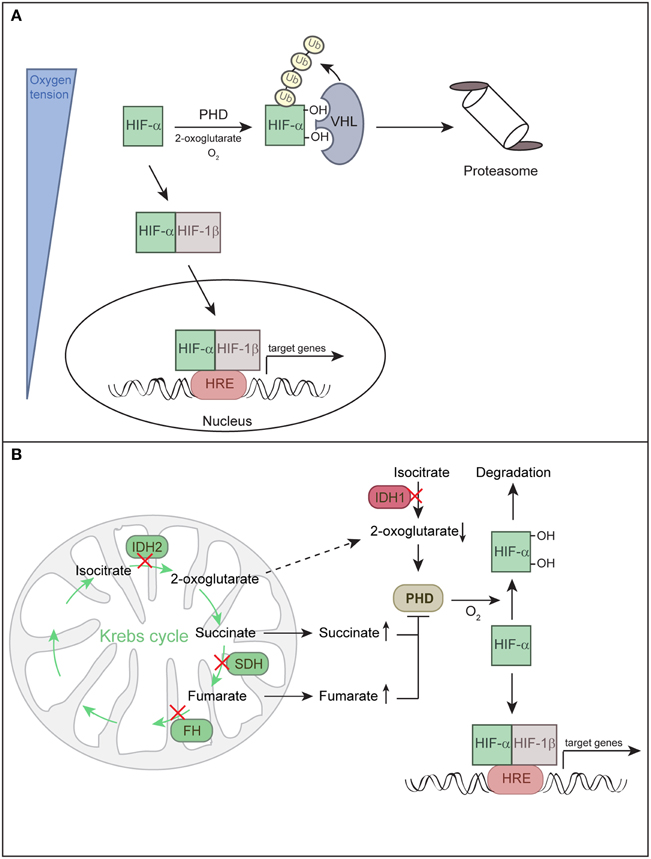
Figure 1. Regulation of HIF-α subunits. (A) Hypoxia-inducible factors (HIFs) are transcription factors composed of O2-regulated α subunits (HIF-1α or HIF-2α) and a constitutively expressed HIF-1β subunit. Together these subunits bind hypoxia response elements (HRE) to mediate adaptive responses to hypoxia. HIF-α activity is directly linked to oxygen partial pressure. Under normoxia, HIF-α is hydroxylated by prolyl hydroxylase domain protein (PHD) and targeted for proteasomal degradation by the von Hippel-Lindau (VHL) E3 ubiquitin ligase complex. Under hypoxia, hydroxylation is inhibited and HIF-α is stabilized, it dimerizes with HIF-1β and enters the nucleus to induce target gene transcription. (B) HIF-α can be stabilized irrespective of O2 tension due to inhibition of PHDs, a state defined as pseudohypoxia. Mutations in the Krebs cycle enzymes succinate dehydrogenase (SDH) and fumarate hydratase (FH) lead to accumulation of succinate and fumarate, respectively, whereas mutations in isocitrate dehydrogenases 1/2 (IDH1 and IDH2) lead to low levels of 2-oxoglutarate. Succinate and fumarate inhibit PHDs, while low levels of the co-substrate 2-oxoglutarate decrease the activity of PHDs. Decreased activity of PHDs leads to a low rate of HIF-α hydroxylation under normoxic conditions and stabilization of HIF-α.
HIF-α subunits can also be stabilized under non-hypoxic conditions, a phenomenon termed “pseudohypoxia.” In addition to O2, PHDs are sensitive to changes in certain Krebs cycle intermediates. Mutations in four genes involved in the metabolism of citrate have the potential to stabilize HIF-α by inhibiting HIF-α hydroxylation and are linked to various tumors (Figure 1B) (Raimundo et al., 2011; Losman and Kaelin, 2013). Succinate dehydrogenase and fumarate hydratase deficiencies lead to accumulation of succinate and fumarate, respectively, and these metabolites compete with 2-oxoglutarate to inhibit PHDs. Mutations in isocitrate dehydrogenases 1 and 2 could promote HIF-α stabilization as a result of low levels of 2-oxoglutarate, which is an essential co-substrate of PHDs (Thompson, 2009). Tumor-associated isocitrate dehydrogenase mutations cause a gain of function, leading to high-level production of (R)-2-hydroxyglutarate and depletion of 2-oxoglutarate (Losman and Kaelin, 2013). PHD enzymes were initially reported to be inhibited by (R)-2-hydroxyglutarate (Zhao et al., 2009). However, further studies have shown that (R)-2-hydroxyglutarate potentiates PHD activity and blunts the induction of HIF-α in response to hypoxia (Losman and Kaelin, 2013).
HIF-1α is expressed ubiquitously, whereas HIF-2α is selectively expressed in distinct cell populations of most organs (Majmundar et al., 2010). HIF-1α and HIF-2α have both overlapping and distinct target genes (Keith et al., 2012) and they are differentially regulated in various physiological settings (e.g., embryonic development) and function in pathophysiological conditions such as cancer and ischemic diseases (Semenza, 2012). They have also different roles in tumorigenesis dependent on specific tumor microenvironments (Majmundar et al., 2010; Keith et al., 2012). HIF-induced transcription promotes angiogenesis, erythropoiesis, metastasis and metabolic reprogramming, such as shifting cell metabolism from oxidative phosphorylation to glycolysis. HIF activation due to hypoxia or loss of VHL function also reprograms lipid metabolism leading to lipid accumulation (Huss et al., 2001; Boström et al., 2006; Rankin et al., 2009; Kucejova et al., 2011; Qu et al., 2011; Walter et al., 2014).
Mitochondria and HIF-α
Mitochondria, metabolism, and O2 are inextricably intertwined. In the following sections we will discuss the numerous mechanisms by which HIF signaling can affect mitochondrial function.
HIF-dependent Regulation of Mitochondrial Metabolism
The hypoxia-dependent increase in the abundance and activity of HIF-1α and the HIF-1α-dependent transcriptional program have three major effects on metabolism that serve to equilibrate O2 consumption with O2 supply. First, HIF-1α promotes glycolytic energy production by inducing genes that encode glucose transporters (e.g., GLUT1, GLUT3) and glycolytic enzymes (Figure 2A) (Semenza, 2010). HIF-1α also upregulates lactate dehydrogenase A (LDHA), which converts pyruvate to lactate and regenerates NAD+ for continuous supply for glycolysis, and monocarboxylate transporter 4 (MCT4), which transports lactate out of the cell (Figure 2A).
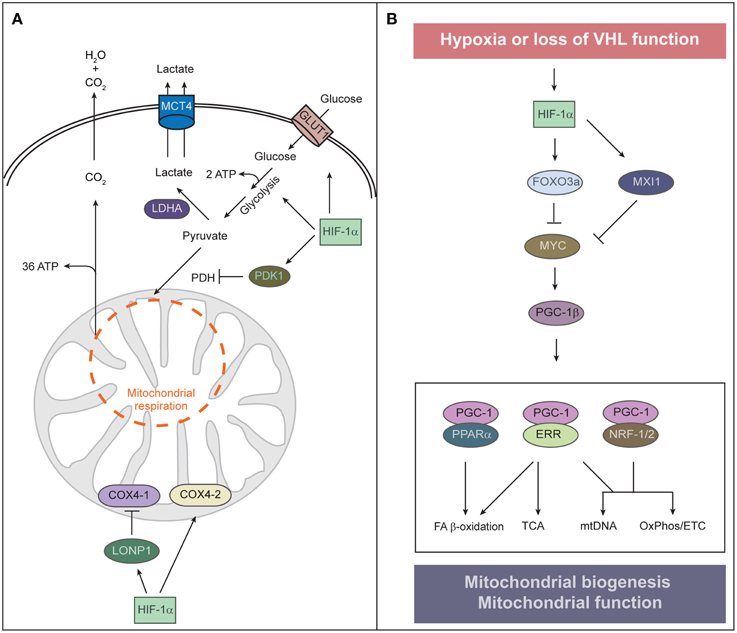
Figure 2. Regulation of mitochondrial function and abundance by HIF-α. (A) To adapt to low oxygen tension, cells undergo two HIF-1α-mediated alterations of cellular metabolism: O2-independent ATP production and reduction of mitochondrial O2 consumption. HIF-1α signaling also contributes to the Warburg effect of aerobic glycolysis—that is, an uncoupling of glycolysis from O2 levels—by stimulating the expression of the glucose transporter GLUT1 and glycolytic enzymes. Increased glycolysis generates increased levels of pyruvate, which is largely converted to lactate by HIF-inducible lactate dehydrogenase A (LDHA) and removed from the cell by the monocarboxylate transporter 4 (MCT4). HIF-1α induces pyruvate dehydrogenase kinase 1 (PDK1), which inhibits pyruvate dehydrogenase (PDH) and blocks conversion of pyruvate to acetyl-CoA, resulting in decreased flux through the tricarboxylic acid (TCA) cycle. Decreased TCA cycle activity attenuates oxidative phosphorylation and excessive mitochondrial ROS production. Under normoxia, COX4-1 is the predominant isoform of COX4 present in complex IV of the electron transport chain, which transfers electrons to O2. Under hypoxia, HIF-1α upregulates the expression of COX4-2 and the mitochondrial protease LONP1, which in turn degrades COX4-1. COX4-2 is more efficient at facilitating the electron transfer to O2 and thereby protects the cell from oxidative damage during hypoxia. (B) Control of mitochondrial biogenesis by HIF-α. HIF-1α induces the expression of MAX-interacting protein 1 (MXI1), a repressor of MYC activity, and thereby represses a subset of MYC target genes such as PGC-1β. HIF-1α-dependent activation of FOXO3a inhibits MYC activity by reducing MYC protein stability. Interaction between PGC-1 and transcription factors such as PPARα, ERR, and NRF-1/2 orchestrates the major functions of mitochondria. HIF-1α-mediated inhibition of MYC and PGC-1 results in reduced mitochondrial biogenesis.
Second, HIF-1α suppresses both the Krebs cycle and oxidative phosphorylation within mitochondria. HIF-1α induces pyruvate dehydrogenase kinase 1 (PDK1), which phosphorylates and inactivates the mitochondrial enzyme pyruvate dehydrogenase that catalyzes the conversion of pyruvate to acetyl-CoA, thereby shunting pyruvate away from mitochondria and diminishing hypoxic mitochondrial respiration (Kim et al., 2006; Papandreou et al., 2006). A benefit of attenuating mitochondrial respiration under hypoxia is reducing the amount of toxic mitochondrial reactive oxygen species (mtROS) generated by inefficient respiration. Mouse embryonic fibroblasts lacking Hif1α undergo cell death as a result of excess mtROS production due to a failure of PDK1 induction (Kim et al., 2006). HIF-1α also reduces mtROS production under hypoxic conditions by optimizing respiration efficiency through inducing cytochrome c oxidase (COX) subunit IV isoform 2 (COX4-2) and the mitochondrial protease LONP1, which degrades the less efficient COX4-1 (Fukuda et al., 2007). Mitochondrial ROS also modulate cell signaling through stabilization of HIF-1α, due to PHD inhibition (Sena and Chandel, 2012). Cells utilize an acute increase in mtROS to stabilize HIF under hypoxia and subsequently restrain ROS production under chronic hypoxia to avoid cellular damage. ROS have a relatively short diffusion distance and thus, mtROS signaling may rely on proximity of ROS-producing mitochondria to their sites of action. Intriguingly, perinuclear clustering of mitochondria triggered by hypoxia is accompanied by the accumulation of nuclear ROS and required for maximal HIF-1α binding to the VEGF promoter and VEGF expression (Al-Mehdi et al., 2012).
Third, hypoxia shifts mitochondrial glutamine metabolism from oxidation to reductive carboxylation (Sun and Denko, 2014). The activity of the α-ketoglutarate dehydrogenase complex is decreased under hypoxia, since HIF-1α induces the E3 ubiquitin ligase SIAH2 that mediates the proteasomal degradation of the E1 subunit of α-ketoglutarate dehydrogenase. Increased α-ketoglutarate levels drive the reverse reaction at isocitrate dehydrogenase, and the glutamine-derived citrate can be transported to the cytoplasm in order to generate acetyl-CoA for anabolic processes under hypoxia.
Cancer cells exhibit a high rate of glycolysis even in the presence of O2, a phenomenon known as aerobic glycolysis or the “Warburg effect” (Vander Heiden et al., 2009). HIF-1α activation as a result of VHL loss or pseudohypoxia contributes also to the development of the Warburg effect. Furthermore, although the shift from oxidative phosphorylation to glycolysis is attributed to the activity of HIF-1α, at least in the liver HIF-2α induces the same genes involved in this metabolic adaptation (Rankin et al., 2009; Walter et al., 2014).
Mitochondrial Biogenesis and HIF-α
A second key remodeling of mitochondria in hypoxia is suppression of mitochondrial biogenesis to decrease mitochondrial mass and O2 consumption. Mitochondrial biogenesis involves replication of the mitochondrial genome and coordinated expression of nuclear- and mitochondrial-encoded gene products. It depends upon the activity of a hierarchy of nuclear transcription factors that includes the peroxisome proliferator-activated receptor (PPAR) γ coactivator 1 family of transcriptional coactivators [PGC-1α, PGC-1β, and the PGC-related coactivator (PRC)], the nuclear respiratory factors (NRF1 and NRF2), and estrogen-related receptors (ERRα, ERRβ, and ERRγ) (Figure 2B) (Scarpulla et al., 2012). PGC-1α is the master regulator of all aspects of mitochondrial biogenesis, including activation of respiratory chain and fatty acid oxidation genes, increased mitochondrial number, mtDNA replication, and augmentation of mitochondrial respiratory capacity. It exerts these effects through direct interaction with and coactivation of PPARs, NRFs, and ERRs (Scarpulla et al., 2012). Nutrient supply and cellular energy balance regulate the activity of PGC-1α at both the transcriptional and posttranslational level (Dominy et al., 2010; Scarpulla et al., 2012). PGC-1β and PRC interact with and coactivate many of the same transcription factors as PGC-1α to promote mitochondrial biogenesis (Scarpulla et al., 2012).
The oncogenic transcription factor MYC promotes mitochondrial biogenesis through activation of PGC-1β (Li et al., 2005; Zhang et al., 2007). MYC dimerizes with MYC-associated factor X (MAX) and binds specific E-box sequences in target gene promoters to activate transcription. Heterodimers of MAX with MAX-interacting protein 1 (MXI1) antagonize MYC function and repress transcription by binding to the same promoter regions of MYC target genes. Cross-talk between HIF and MYC has been defined at a number of levels, however, HIF-1α and HIF-2α exert opposing roles on MYC interaction with its transcription cofactors (Dang et al., 2008; Keith et al., 2012). HIF-1α activation inhibits mitochondrial biogenesis by promoting MYC degradation and by inducing MXI1 expression (Zhang et al., 2007). This inhibition is mediated by the transcription factor FOXO3a (Forkhead-box protein O3a) which is activated in hypoxia downstream of HIF-1α (Peck et al., 2013). FOXO3a can also inhibit MYC activity by reducing MYC protein stability and by increasing the expression of miRNAs that perturb the translation of MYC mRNA (Peck et al., 2013). Huang et al. (2014) reported that HIF-1α, but not HIF-2α, represses MYC in human hepatoma cells and thereby decreases PGC-1β expression, leading to decreased expression of medium- and long-chain acyl-CoA dehydrogenases and subsequent inhibition of mitochondrial fatty acid β-oxidation. It has not been addressed if HIF-1α-mediated downregulation of PGC-1β also affects mitochondrial biogenesis and mass in hepatoma cells. HIF-2α actually enhances MYC transcriptional activity by binding to and stabilizing the MAX-MYC heterodimer (Gordan et al., 2007, 2008). The cooperation of HIF-2α with MYC increases MYC effects on various cell cycle regulators and drives tumorigenesis. Given the opposite effects of HIF-1α and HIF-2α on MYC, it remains to be established how MYC is modulated in cells that express both HIF isoforms.
However, with the exception of HIF-1α-mediated inhibition of PGC-1β, data about HIF-α mediated regulation of PGC-1α, the master regulator of mitochondrial biogenesis and function, are relatively scarce. Several studies demonstrated an induction of PGC-1α by hypoxia independently of HIF-1α activity (Shoag and Arany, 2010). Overexpression of PGC-1α under normoxia induces mitochondrial biogenesis which increases O2 consumption and decreases intracellular O2 levels, leading to HIF-1α protein stabilization and activation of HIF-1α target genes (O'Hagan et al., 2009). ROS accumulation in hypoxic cancer cells induces expression of PGC-1α/β to promote detoxification through induction of antioxidative enzymes (Austin and St-Pierre, 2012). The precise relationship between the PGC-1α and HIF-α pathways with respect to mitochondrial biogenesis obviously needs further clarification.
HIF-dependent Regulation of Mitophagy
The third mechanism by which HIF-α controls mitochondrial function is selective mitochondrial autophagy (mitophagy). Autophagy is an evolutionary conserved catabolic process for degradation of macromolecules and organelles. Both non-selective “bulk” autophagy and selective autophagy of specific proteins or organelles have been described (Mizushima et al., 2011; Schreiber and Peter, 2014). Selective and non-selective autophagy share a set of autophagy-related (Atg) proteins referred to as the core autophagic machinery (Stolz et al., 2014). Yeast Atg8 and its mammalian homologs of the microtubule-associated protein-1 light chain 3 family (LC3A, LC3B, LC3C) and γ-aminobutyric acid-receptor-associated (GABARAP, GABARAPL1, GABARAPL2) proteins are covalently conjugated to the lipid phosphatidylethanolamine upon induction of autophagy. Besides playing a pivotal role in different steps of autophagosome biogenesis, the LC3 family members are important for target recognition during selective autophagy. Selective autophagy requires specific receptors, which recognize cargo tagged with degradation signals, connect it to the autophagosomal membrane through their LC3-interacting regions (LIR), and are degraded together with their cargo within autolysosomes (Stolz et al., 2014).
Mitophagy can be initiated in several ways to arbitrate mitochondrial quality control via the selective removal of superfluous or damaged mitochondria. The best-studied mechanism for mitophagy in mammalian cells is the PINK1-Parkin-mediated pathway, which is elegantly reviewed elsewhere (Scarffe et al., 2014). Briefly, phosphatase and tensin homolog (PTEN)-induced putative protein kinase (PINK1), which is rapidly degraded in healthy mitochondria, accumulates upon mitochondrial membrane depolarization at the outer membrane, leading to the recruitment of the E3 ubiquitin ligase Parkin to mitochondria and ubiquitination of several outer mitochondrial membrane (OMM) proteins (Winklhofer, 2014). The recruitment of ubiquitinated mitochondria to autophagic structures is mediated by LC3 family members and ubiquitin-binding adaptor proteins such as sequestosome 1 (SQSTM1/p62), NBR1 (neighbor of BRCA1 gene 1), and optineurin (Rogov et al., 2014).
Although mitophagy has been extensively studied in mammals, mitophagy-specific factors still remain controversial. In yeast Atg32 has been identified as receptor protein for mitophagy (Kanki et al., 2009, 2015; Okamoto et al., 2009). Atg32 localizes to the OMM, harbors a classical LIR consensus sequence and interacts with Atg8 and the scaffold protein Atg11 to enable the assembly of the core autophagy machinery around the cargo (Figure 3A). Casein kinase 2 (CK2) regulates mitophagy by phosphorylating Atg32, which stabilizes the Atg32-Atg11 interaction and promotes mitophagy. So far no mammalian homolog has been identified for Atg32.
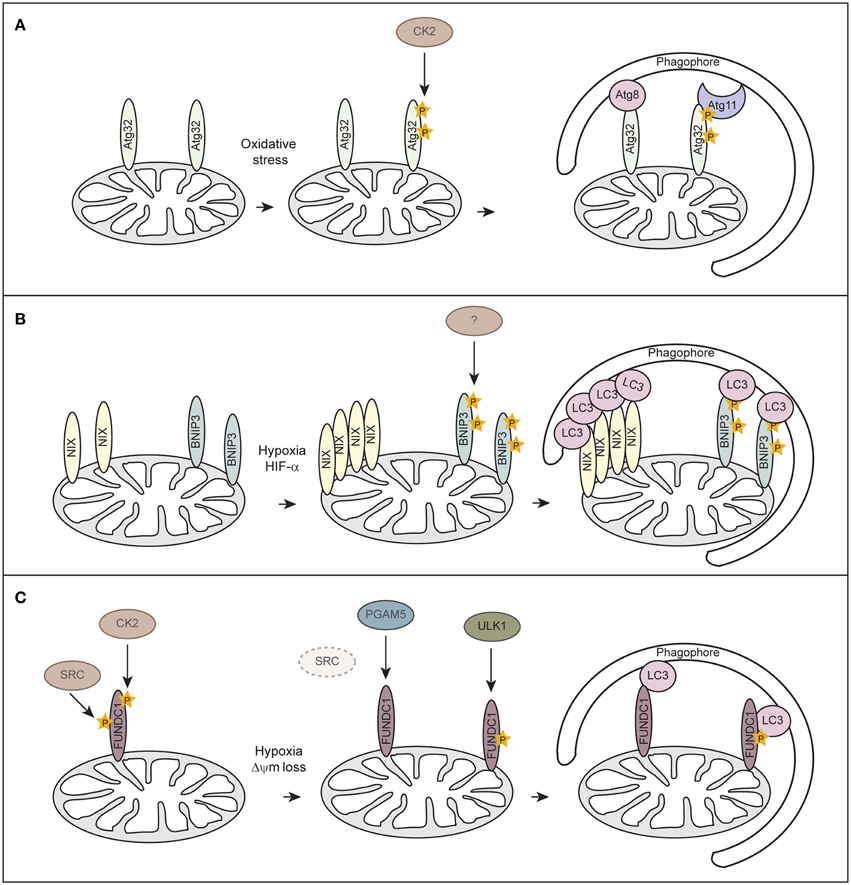
Figure 3. Receptor-mediated mitophagy. (A) Atg32-mediated mitophagy in S. cerevisiae. Atg32 is an outer mitochondrial membrane protein whose expression is induced upon mitophagy-inducing conditions. Atg32 interacts with Atg8 and Atg11 via distinct domains. Casein kinase 2 (CK2) phosphorylates Atg32 upon mitophagy-inducing conditions, which is essential for the Atg11 interaction without affecting Atg32-Atg8 binding. (B) NIX/BNIP3-mediated mitophagy in mammalian cells. NIX and BNIP3 are outer mitochondrial membrane proteins that interact with LC3 through LIR motifs in their N-terminal region. Upon hypoxia, NIX and BNIP3 are transcriptionally induced in a HIF-α-dependent manner. Phosphorylation of BNIP3 promotes its binding to LC3 and subsequent mitophagy. The kinase for BNIP3 phosphorylation is unknown. (C) FUNDC1-mediated mitophagy in mammalian cells. FUNDC1 is an outer mitochondrial membrane protein that interacts with LC3 through a LIR domain at its cytosol-exposed N-terminus. Under normal physiological conditions, FUNDC1 is phosphorylated by SRC and CK2, thereby preventing LC3 binding. Upon hypoxia or loss of mitochondrial membrane potential (Δψm), the expression of SRC is strongly suppressed and PGAM5 dephosphorylates FUNDC1. Dephosphorylation of FUNDC1 enhances the interaction between FUNDC1 and LC3 and promotes mitophagy. Phosphorylation of FUNDC1 by ULK1 enhances its binding to LC3.
Similar to Atg32 in yeast, the mammalian mitophagy receptors BNIP3 (Bcl-2 and adenovirus E1B 19-kDa-interacting protein 3), BNIP3-like (BNIP3L/NIX), and FUNDC1 (FUN14 domain containing 1) are OMM proteins which can directly bind to LC3 via their LIR motifs (Figures 3B,C). BNIP3 and NIX were originally thought to promote apoptosis or programmed necrosis (Zhang and Ney, 2009). They can activate autophagy by binding to Bcl-2 and thereby disrupting the interaction between Beclin-1 and Blc-2/Bcl-XL (Bellot et al., 2009; Boland et al., 2013). BNIP3 and NIX are hypoxia-inducible HIF-α target genes and it has been suggested that they act either in hypoxia-induced macroautophagy or mitophagy (Sowter et al., 2001; Tracy et al., 2007; Zhang et al., 2008; Bellot et al., 2009). While BH3 domains of BNIP3 and NIX are sufficient to induce the general autophagy response (Bellot et al., 2009), induction of mitophagy requires the LIR domain of NIX (Novak et al., 2010) and BNIP3 (Hanna et al., 2012; Zhu et al., 2013). Phosphorylation of BNIP3 at serines flanking its LIR domain promotes binding to LC3 family members and thereby increases mitophagy (Zhu et al., 2013), however, involved kinases are unknown (Figure 3B). It is not clear how phosphorylation of BNIP3 is regulated under hypoxia and if phosphorylation regulates NIX.
Recently, FUNDC1 has been implicated in mediating hypoxia-induced mitophagy (Liu et al., 2012). FUNDC1-mediated mitophagy is regulated at the posttranslational level by reversible phosphorylation. Under normal physiological conditions FUNDC1 is phosphorylated by Src kinase at Tyr18, which is located in the LIR motif, and at Ser13 by CK2 (Figure 3C) (Chen et al., 2014a). In response to hypoxia or loss of mitochondrial membrane potential the mitochondrially localized phosphoglycerate mutase family member 5 (PGAM5), a Ser/Thr phosphatase, dephosphorylates FUNDC1 at Ser13, whereas Tyr18 phosphorylation seems to be prevented before mitophagy-induction due to inactivation of Src kinase (Figure 3C) (Chen et al., 2014a). Dephosphorylated FUNDC1 displays a higher binding affinity to LC3, resulting in selective autophagosome incorporation and autophagic degradation of mitochondria (Liu et al., 2012; Chen et al., 2014a). Moreover, a study showed that hypoxia or mitochondrial uncouplers elevate protein levels of the autophagy-initiating kinase ULK1 (UNC51-like kinase 1) and target ULK1 to damaged mitochondria where it phosphorylates FUNDC1 at Ser17 and thereby enhances FUNDC1 binding to LC3 (Figure 3C) (Wu et al., 2014). However, it remains to be determined how BNIP3, NIX and FUNDC1 are interconnected or even needed during hypoxia-induced mitophagy.
Hepatic HIF-α Signaling and Mitochondrial Abundance
We examined the effect of HIF-α signaling on hepatic mitochondrial abundance in control and liver-specific Vhl−∕−, Vhl−∕−/Hif1α−∕−, and Vhl−∕−/Hif2α−∕− mice. Mitochondrial protein levels are similar in control and knockout livers (Walter et al., 2014), suggesting that constitutive HIF-α signaling does not affect hepatic mitochondrial mass. However, subcellular fractionation of livers from control and Vhl−∕− mice shows that mitochondrial protein levels are decreased in the heavy mitochondrial fraction of Vhl−∕− livers compared with controls, whereas their levels are increased in the light mitochondrial fraction (Walter et al., 2014). Further purification of the light mitochondrial fraction by density gradient centrifugation and immunoblots of gradient fractions show that the OMM protein VDAC shifts toward lower density fractions. In summary, HIF-α signaling in the liver alters the ratio between heavy and light mitochondria, whereas mitochondrial protein levels are not changed in whole liver homogenates. It remains to be clarified if and how hepatic HIF-α signaling affects mitochondrial size, ultrastructure and function. Mitochondrial morphology and ultrastructure depend on mitochondria-shaping proteins that regulate organellar fusion and fission (Mishra and Chan, 2014). Rates of mitochondrial fission and fusion respond to changes in metabolism, and mitochondria regulate their shape to adjust their activity with metabolic conditions (Mannella, 2006).
Endoplasmic Reticulum and Hypoxia
ER Stress and Hypoxia
The ER is an extensive intracellular membrane network that extends throughout the cytoplasm and is essential for the translation and folding of membrane and secretory proteins (Gidalevitz et al., 2013). It is also a critical site of lipid and glucose metabolism, calcium homeostasis, and detoxification of drugs and metabolic byproducts. A biochemical process that is crucial for ER protein homeostasis is the formation of disulfide bridges, which is referred to as oxidative protein folding (Eletto et al., 2014). Disulfide bond generation by ER-localized protein disulfide isomerases is an oxidative process, and molecular O2 and H2O2 are the principal electron acceptors for oxidative folding in the ER (Eletto et al., 2014). Protein disulfide isomerases catalyze oxidation by coupling de novo disulfide formation to the reduction of O2 to H2O2.
The term “endoplasmic reticulum stress” defines any perturbation that compromises the protein folding functionality of the ER (Walter and Ron, 2011). A number of biochemical and physiologic stimuli, such as perturbation in calcium homeostasis or redox status, elevated secretory protein synthesis, expression of misfolded proteins, glucose deprivation, altered glycosylation, viral infection, and excess lipids can disrupt ER homeostasis and impose stress to the ER. These disturbances trigger an adaptive signaling pathway known as the unfolded protein response (UPR) that aims to restore ER homeostasis and function. Hypoxia is a physiologically important ER stress common to solid tumors. Several lines of evidence point to a strong relationship between hypoxia and the accumulation of misfolded proteins in the ER (Koumenis et al., 2007). In tumors, hypoxia is also associated with other conditions that can cause ER stress, such as glucose and amino acid deprivation, and oxidative stress.
The Unfolded Protein Response Signaling Pathways
Activation of the three canonical branches of the UPR is mediated by three stress-sensing ER transmembrane proteins: protein kinase RNA-like ER kinase (PERK), inositol-requiring protein 1α (IRE1α), and activating transcription factor 6 (ATF6) (Figure 4) (Walter and Ron, 2011; Faust and Kovacs, 2014). In a stress-free ER, these sensors are bound by the ER-resident chaperone glucose-regulated protein of 78 kDa (GRP78) in their intraluminal domains and rendered inactive. Upon ER stress, GRP78 dissociates from PERK, IRE1, and ATF6, leading to their activation (Bertolotti et al., 2000; Shen et al., 2002).
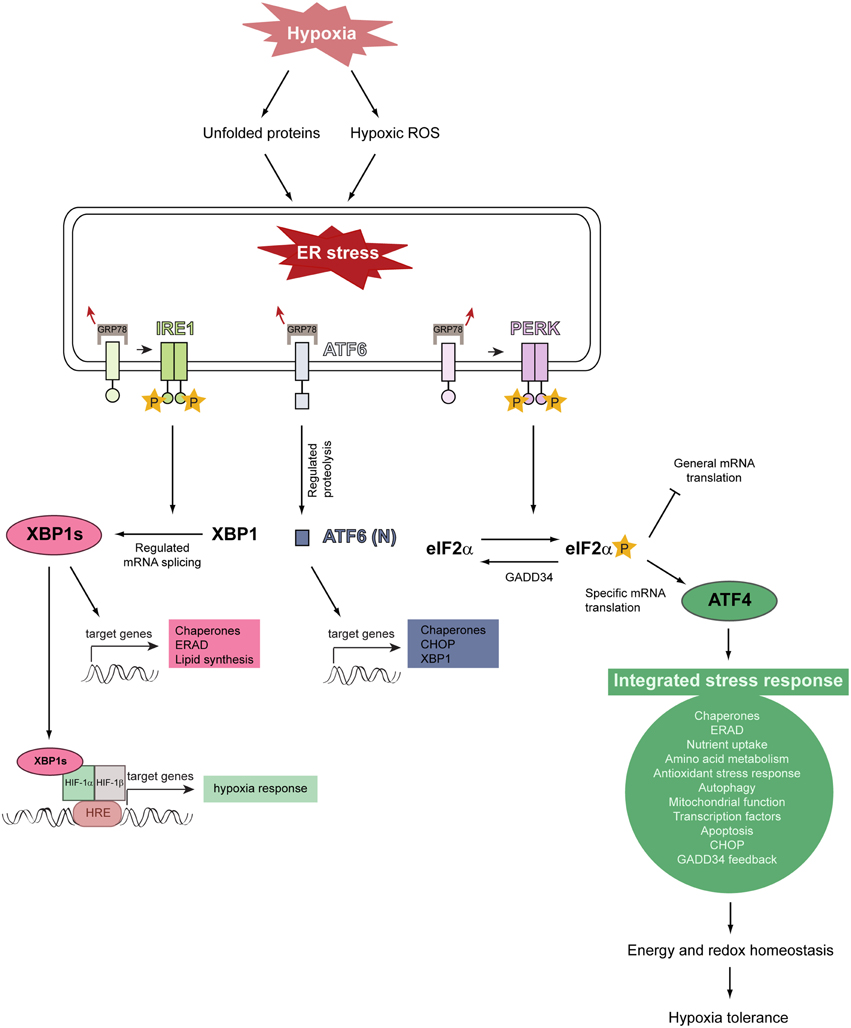
Figure 4. Model illustrating the relationship between hypoxia, ER stress, and activation of the unfolded protein response. Severe hypoxic stress perturbs and reduces O2-dependent protein folding capacity, resulting in the accumulation and aggregation of misfolded proteins in the ER lumen. Hypoxia increases intracellular ROS production and ROS stimulate multiple biological responses during O2 deprivation. Unfolded proteins and hypoxic ROS trigger ER stress which leads to HIF-α-independent activation of the UPR. The UPR is initiated by the stress-sensing ER transmembrane proteins PERK, IRE1, and ATF6. The chaperone GRP78 is normally bound to these ER stress sensors and keeps them inactive, but dissociates from them under ER stress conditions. This dissociation leads to the activation of the three UPR pathways. GRP78 dissociation allows PERK to homodimerize, which facilitates autotransphosphorylation and kinase domain activation. Activated PERK phosphorylates eIF2α which decreases general translation while increasing the preferential translation of specific proteins, such as the transcription factor ATF4. ATF4 triggers the activation of a gene expression program referred to as the integrated stress response. ATF4 induces the expression of GADD34, which acts as a negative regulator of the PERK pathway by dephosphorylating eIF2α. The integrated stress response promotes energy and redox homeostasis and is an important prosurvival mechanism under moderate hypoxia. IRE1 homodimerization, followed by autotransphosphorylation, triggers its RNase activity. IRE1-mediated splicing of full-length XBP1 mRNA generates XBP1s, which encodes an active transcription factor. ER stress leads to the translocation of ATF6 to the Golgi, where it is cleaved by regulated intramembrane proteolysis to produce the active transcription factor. Modified from Faust and Kovacs (2014).
IRE1 is a serine-threonine kinase and endoribonuclease, which catalyzes the splicing of full-length XBP1 (X-box-binding protein 1) mRNA to generate an active transcription factor, termed spliced XBP1s. XBP1s activates genes encoding proteins involved in protein folding, ER-associated degradation (ERAD) of misfolded ER proteins, and lipid synthesis.
ATF6 is comprised of two isoforms, ATF6α and ATF6β, and resides as transcriptionally inactive precursor protein in the ER membrane. ER stress leads to ATF6 translocation from the ER to the Golgi apparatus where it is cleaved sequentially by Site-1 and Site-2 proteases to produce an active transcription factor. ATF6 induces XBP1 and genes mainly encoding ER chaperones and proteins involved in ERAD.
Dissociation of GRP78 from PERK leads to its homodimerization and activating autophosphorylation. PERK phosphorylates the α-subunit of the eukaryotic translation initiation factor 2 (eIF2α) on serine 51. This phosphorylation event attenuates general translation, resulting in a reduced protein folding load of the ER (Baird and Wek, 2012), but it stimulates selective translation of the activating transcription factor 4 (ATF4), which plays a crucial role for the adaptation to stress. ATF4 target genes are involved in protein folding and assembly, metabolism, nutrient uptake, gene expression, alleviation of oxidative stress, autophagy, and the regulation of apoptosis. In addition to PERK, three other kinases induce eIF2α phosphorylation and preferential translation of ATF4: GCN2 (general control non-derepressible kinase 2), PKR (double-stranded RNA-activated protein kinase), and HRI (heme-regulated inhibitor kinase) (Baird and Wek, 2012). The PERK-eIF2α-ATF4 pathway is referred to as the integrated stress response (ISR), because divergent signals activate the four eIF2α kinases and the ISR, which seeks to remediate stress and restore cellular homeostasis.
Hypoxia and the Unfolded Protein Response
As protein synthesis and O2-dependent protein folding are energy-intensive processes and chronic hypoxia markedly reduces intracellular ATP levels (Kim et al., 2006; Liu et al., 2006), control of mRNA translation is an important cellular response to hypoxia. Hypoxia activates PERK and thereby leads to eIF2α phosphorylation and global translation inhibition, whereas translation of ATF4 is increased in a PERK/eIF2α-dependent manner (Koumenis et al., 2002, 2007; Blais et al., 2004; Bi et al., 2005; Koritzinsky et al., 2006; Wouters and Koritzinsky, 2008). This is a rapid HIF-1α-independent response, occurring within minutes when cells are exposed to anoxic conditions and somewhat more slowly during moderate hypoxia. eIF2α phosphorylation is transient due to the negative feedback loop initiated by ATF4-dependent upregulation of GADD34 (growth arrest DNA-inducible gene 34), which dephosphorylates eIF2α (Figure 4).
Hypoxia increases intracellular ROS production in various cells to stimulate multiple biological responses, and mitochondria appear to be the primary source of hypoxic ROS (Liu et al., 2008). Mitochondrial hypoxic ROS activate the ISR to promote energy and redox homeostasis and to constitute an early adaptive response to hypoxia. Enzymatic antioxidants such as catalase and glutathione peroxidase reduce eIF2α phosphorylation caused by hypoxia, suggesting that H2O2 is a key biologically active form of ROS during hypoxia (Liu et al., 2008). ATF4 augments HIF-1α-mediated upregulation of its downstream targets to promote cell survival (Pereira et al., 2014).
Transient exposure to ER stress can condition and prepare cells for survival during a subsequent, more severe stress. This preconditioning is likely due to induction of pro-survival genes, and the ISR is an important prosurvival mechanism under hypoxia. Tumor cells in the primary tumor are exposed to hypoxia and might be preconditioned to survive the subsequent metastatic process. Indeed, cells with compromised PERK-eIF2α-ATF4 signaling are more sensitive to hypoxic stress in vitro and they form slower growing tumors in vivo, indicating that the PERK-eIF2α-ATF4 pathway confers a survival advantage for tumor cells under hypoxia (Fels and Koumenis, 2006). Severe hypoxia (<0.01%) and ER stress induce in cancer cells the transcription of ULK1, LC3B, and ATG5 through the activity of ATF4 (Rouschop et al., 2010; Pike et al., 2013), and this up-regulation is crucial for maintaining high levels of autophagic flux to survive intratumoral hypoxia, metabolic stress, and starvation.
Analysis of gene expression changes during hypoxia indicated that UPR genes, including genes specifically regulated by XBP1, were most robustly induced during severe hypoxia/anoxia (Romero-Ramirez et al., 2004). Hypoxia induced in a HIF-1α-independent manner XBP1 expression and activated splicing of its mRNA, resulting in increased levels of XBP1s (Romero-Ramirez et al., 2004). XBP1s colocalizes with hypoxia markers in tumors, and loss of XBP1 increases the sensitivity of transformed cells to hypoxia-induced apoptosis and inhibits tumor growth (Wouters and Koritzinsky, 2008; Spiotto et al., 2010).
XBP1 is activated in triple-negative breast cancer (TNBC)—a type of breast cancer that does not have estrogen, progesterone and HER2 receptors—and has a pivotal role in the tumorigenicity and progression of TNBC (Chen et al., 2014b). HIF-1α is hyperactivated in TNBCs, but XBP1 splicing is not directly regulated by HIF-1α. XBP1 drives TNBC tumorigenicity by assembling a transcriptional complex with HIF-1α that augments HIF-1α activity and regulates the HIF-1α transcriptional program, and XBP1 knockdown reduces mammosphere formation in hypoxic conditions (Chen et al., 2014b). The XBP1 gene expression signature of TNBC patients correlates with HIF-1α and hypoxia-driven signatures and is associated with poor prognosis.
Although it is expected, a connection between ATF6 and hypoxia has not been reported yet and remains largely unexplored. One study showed that ATF6 is activated independently of HIF-1α by simulated ischemia (0.1% O2) in a primary cardiac myocyte model system and inactivated upon reperfusion (Doroudgar et al., 2009). The absence of HIF-1α activation at 0.1% O2 is consistent with other studies showing that HIF-α activation is maximal at 0.5% O2 but decreases to nearly basal levels at lower O2 concentrations (Jiang et al., 1996). Furthermore, while PERK and XBP1 activation occur in a HIF-1α-independent manner, a possible involvement of HIF-2α in UPR activation has not been addressed yet.
Peroxisomes and HIF Signaling
Peroxisomal Metabolism and Oxygen
Peroxisomes are ubiquitous and highly dynamic organelles whose number, size, and function are dependent on cell type and metabolic needs. They play key roles in the degradation of fatty acids [i.e., very long-chain fatty acids (VLCFAs), branched-chain FAs, polyunsaturated FAs (PUFAs)], ether lipid synthesis, cholesterol and bile acid synthesis, and metabolism of ROS (Figure 5A) (Van Veldhoven, 2010; Fransen et al., 2012; Faust and Kovacs, 2014). They also act as intracellular signaling platforms in redox, lipid, inflammatory, and innate immunity signaling (Dixit et al., 2010; Nordgren and Fransen, 2014; Odendall et al., 2014). The importance of peroxisomes for cellular metabolism is illustrated by the marked abnormalities in brain and systemic organs in peroxisome biogenesis disorders of the Zellweger spectrum in which functional peroxisomes are absent and disorders caused by single peroxisomal enzyme deficiencies (Raymond et al., 2009). Lack of peroxisomal metabolism creates severe biochemical abnormalities, leading to a variety of clinical symptoms both in patients with peroxisomal disorders as well as peroxisome-deficient mice (Kovacs et al., 2002; Raymond et al., 2009; Baes and Van Veldhoven, 2012; Faust and Kovacs, 2014).
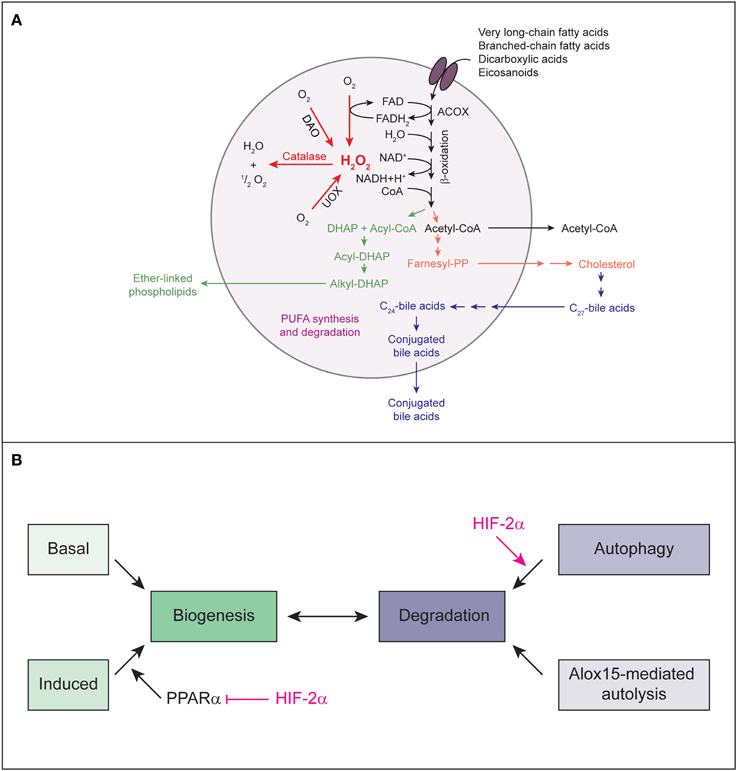
Figure 5. (A) The major metabolic pathways in peroxisomes of the mammalian liver. Various lipids are transported by PMPs (e.g., the ABC transporter proteins ABCD1, ABCD2, ABCD3) into the peroxisomal matrix, where they are oxidized by the β-oxidation enzymes. The products of the β-oxidation can serve as substrates for the biosynthesis of ether-linked phospholipids, cholesterol and bile acids or may exit the peroxisome for further oxidation in mitochondria. With regard to PUFAs, peroxisomes not only degrade these compounds but are also involved in their formation through retroconversion of PUFAs by catalyzing the chain-shortening steps. Peroxisomal function depends highly on molecular O2 due to its oxidative type of metabolism. Peroxisomal β-oxidation and the activity of other peroxisomal oxidases (e.g., UOX, DAO) result in the production of H2O2, which is decomposed by catalase. Modified from Schrader and Fahimi (2008). (B) Model for HIF-2α-mediated decrease in peroxisome abundance. Peroxisome homeostasis is achieved by balancing biogenesis and degradation of peroxisomes. HIF-2α signaling promotes degradation of peroxisomes by pexophagy. Reduced peroxisome abundance and the ensuing deficiency in peroxisomal function leads to major changes in the lipid profile, such as accumulation of VLCFAs. VLCFAs are activating ligands for the transcription factor PPARα. HIF-2α represses ligand-induced PPARα-mediated peroxisome proliferation and consequential restoration of peroxisome homeostasis. Thus, by simultaneously inducing pexophagy and counteracting PPARα, HIF-2α ensures efficient depletion of the peroxisome pool.
Peroxisomal function depends highly on molecular O2 due to its oxidative type of metabolism (Figure 5A). In fact, peroxisomes may be responsible for as much as 20% of O2 consumption and 35% of H2O2 production in tissues such as the liver (Fransen et al., 2012). The number of peroxisomes is approximately 10–15 times less than that of mitochondria (De Duve and Baudhuin, 1966); therefore, on a per unit basis, peroxisomes may consume a significant amount of O2 as compared to mitochondria. However, so far there has been no evidence linking HIF signaling to peroxisomes. We hypothesized that to minimize O2 consumption under hypoxic conditions, HIF-α signaling may inhibit O2-dependent peroxisomal metabolism and/or decrease peroxisome abundance. Since peroxisomes are highly abundant in the liver and liver-specific loss of Vhl causes severe lipid accumulation, we investigated peroxisome homeostasis and metabolism in the liver of control and liver-specific Vhl, Vhl/Hif1α, and Vhl/Hif2α knockout mice and explored the role of HIF-1α and HIF-2α in this context.
Peroxisome Biogenesis and Hepatic HIF-α Signaling
Peroxisome homeostasis is maintained by balancing biogenesis and degradation of peroxisomes. Peroxisomes can either multiply by growth and fission of pre-existing ones (Schrader et al., 2012) or develop de novo from the ER (Tabak et al., 2013). Proteins involved in peroxisome biogenesis, the peroxins, are encoded by PEX genes (Hasan et al., 2013; Smith and Aitchison, 2013). In mammalian cells, peroxisome proliferation is triggered by lipids which are substrates of peroxisomal metabolism and ligands of PPARα (Schrader et al., 2012). The majority of peroxins is not induced transcriptionally through peroxisome proliferators. The peroxins PEX11α, β, and γ are involved in the regulation of peroxisome size and number in mammalian cells (Schrader et al., 2012), but only PEX11α is a PPARα target gene. Their overexpression increases peroxisome number in the absence of extracellular stimuli or peroxisome metabolism (Li and Gould, 2002; Schrader et al., 2012). Recently, we showed that activation of HIF-α signaling in the liver does not affect the expression of Pex genes (Walter et al., 2014). Pex11α is the only peroxin that is transcriptionally induced in response to HIF-2α activation in Vhl−∕− and Vhl−∕−/Hif1α−∕− livers (Walter et al., 2014). The peroxisome biogenesis machinery in Vhl−∕− livers is functional, because peroxisome proliferation can be induced by treatment with PPARα-dependent and -independent peroxisome proliferators. However, HIF-2α signaling has a repressive effect on ligand- and fasting-induced PPARα activation (Figure 5B) (Walter et al., 2014).
Pexophagy
Three mechanisms for mammalian peroxisome degradation have been described, which include selective autophagy (pexophagy), proteolysis by peroxisomal Lon protease 2 (LONP2), and 15-lipoxygenase-1 (ALOX15)-mediated autolysis (Till et al., 2012). Studies using liver-specific Atg7−∕− mice suggest that 70–80% of excess liver peroxisomes are degraded by pexophagy, while the remaining 20–30% are degraded via the action of LONP2 and ALOX15 (Till et al., 2012). A general rule applicable for both yeast and mammalian cells is that environmental conditions that require peroxisomal metabolism lead to peroxisome proliferation, followed by pexophagic degradation when the organelles are no longer required (Iwata et al., 2006; Farré et al., 2008; Motley et al., 2012).
The pexophagy receptors Atg30 and Atg36 were identified in P. pastoris and S. cerevisiae, respectively, and their overexpression stimulates pexophagy even under peroxisome-inducing conditions (Farré et al., 2008; Motley et al., 2012). Their synthesis is upregulated in peroxisome proliferation conditions, they localize to the peroxisome membrane and bind to Pex3, and they depend on phosphoregulation for their interactions with components of the autophagy machinery (i.e., Atg8, Atg11) during pexophagy conditions (Farré et al., 2008, 2013). Pex3 acts as a docking station for several proteins involved in peroxisomal biogenesis and its interaction with Atg30 regulates the phosphorylation status of Atg30 by a yet unknown kinase (Burnett et al., 2015). Atg30 interacts also with Pex14, the scaffold protein Atg17, and the acyl-CoA binding protein Atg37 (Nazarko et al., 2014). The human ortholog of Atg37, acyl-CoA-binding domain containing protein 5 (ACBD5), is also peroxisomal and required for pexophagy (Nazarko et al., 2014). Despite their functional similarities Atg30 and Atg36 do not display any significant sequence homology and they are conserved only among a few yeast species.
There are no orthologous genes of Atg30 and Atg36 in mammals. Overexpression of NBR1 and SQSTM1, which are autophagy receptors of ubiquitinated targets, induces clustering and degradation of peroxisomes in cell lines (Deosaran et al., 2013). SQSTM1 is not required for pexophagy when NBR1 is in excess, but its binding to NBR1 increases the efficiency of NBR1-mediated pexophagy (Deosaran et al., 2013). Artificial mono-ubiquitination of peroxisomal membrane proteins (PMPs) in mammalian cells causes peroxisome turnover by pexophagy in a SQSTM1-dependent manner (Kim et al., 2008). However, it is unknown if a PMP is ubiquitinated under pexophagy-inducing conditions and whether subsequent interaction with NBR1 and/or SQSTM1 links ubiquitinated peroxisomes to the autophagic machinery.
In Chinese hamster ovary (CHO) cells, it has been suggested that under nutrient starvation PEX14 is involved in pexophagy by interacting with the lipidated form of LC3 (Hara-Kuge and Fujiki, 2008). Cell-free synthesized lipidated LC3 interacts in an in vitro assay with the transmembrane domain of recombinant PEX14, although PEX14 does not contain a LIR sequence that could ensure LC3 binding (Jiang et al., 2015). PEX14 is an essential component of the peroxisomal translocon complex (Hasan et al., 2013), and it has been proposed that the PEX14-LC3 and PEX14-PEX5 interactions are mutually exclusive (Hara-Kuge and Fujiki, 2008). This competitive interaction might ensure functional segregation of metabolically active and degradation-prone peroxisomes.
Overexpression of Pex3 in CHO cells and mouse embryonic fibroblasts induced clustering of peroxisomes and NBR1-mediated pexophagy, albeit no direct interaction between PEX3 and NBR1 could be detected (Yamashita et al., 2014). Interestingly, ubiquitin signals were observed on peroxisomes upon Pex3 overexpression, suggesting that a currently unidentified PMP is ubiquitinated in PEX3-mediated pexophagy and might function in NBR1 recruitment (Yamashita et al., 2014).
HIF-2α-mediated Pexophagy in the Liver
We examined the effect of HIF-α signaling on hepatic peroxisome abundance in control and liver-specific Vhl−∕−, Vhl−∕−/Hif1α−∕−, and Vhl−∕−/Hif2α−∕− mice. Peroxisome abundance is significantly decreased in livers of Vhl−∕− mice (Walter et al., 2014). Reduction of peroxisome abundance is mediated by HIF-2α, because, with respect to the peroxisomal phenotype, we observe a striking rescue in Vhl−∕−/Hif2α−∕− but not Vhl−∕−/Hif1α−∕− mice. HIF-2α promotes pexophagy because peroxisome abundance is increased after inhibition of autophagy with 3-methyladenine (3-MA) in Vhl−∕− mice. In addition, expression of a non-degradable active HIF-2α variant fails to decrease peroxisome abundance in liver-specific, autophagy-deficient Atg7−∕− mice (Figure 5B) (Walter et al., 2014). In support of this finding super-resolution and electron microscopy demonstrated that both single and multiple peroxisomes, but no other cytoplasmic organelles, are sequestered in autophagosomes in Vhl−∕− livers.
Peroxisome abundance and protein levels of NBR1 and SQSTM1 are concomitantly decreased in Vhl−∕− and Vhl−∕−/Hif1α−∕− livers (Walter et al., 2014). Neither peroxisome abundance nor NBR1 and SQSTM1 levels decline in 3-MA-treated Vhl−∕− mice, showing that the abundance of these receptors and peroxisomes are interconnected. Expression of a constitutively active HIF-2α variant results also in a concomitant decrease of peroxisome abundance and NBR1 levels. NBR1 and SQSTM1 colocalize with peroxisomes in Vhl−∕− livers, but surprisingly NBR1 already localizes to peroxisomes in control livers (Walter et al., 2014). An intriguing feature of autophagy receptors is their tendency to oligomerize, which facilitates sequestration and clustering of the autophagic cargo. Indeed, treatment of Vhl−∕− mice with 3-MA leads to a significant clustering of NBR1- and SQSTM1-positive peroxisomes, suggesting that binding of multiple NBR1 and SQSTM1 to peroxisomes and oligomerization of these receptors might induce peroxisome clustering and prime peroxisomes for pexophagy.
In summary, by simultaneously inducing pexophagy and counteracting PPARα, HIF-2α ensures efficient depletion of the peroxisome pool (Figure 5B). Our data show that the autophagy receptors NBR1 and SQSTM1 localize to peroxisomes and are degraded together with peroxisomes by HIF-2α-mediated pexophagy. However, it remains an open question how HIF-2α induces pexophagy at the molecular level, but several possibilities exist and are discussed below.
Peroxisome Abundance in Tumors
Peroxisome proliferation is a unique phenomenon generated by a broad spectrum of structurally diverse compounds, such as lipid-lowering drugs and plasticizers (Pyper et al., 2010; Misra et al., 2013). These compounds induce peroxisome proliferation in liver parenchymal cells of rodents, whereas no effects have been observed in non-human primates and humans. Prolonged exposure to peroxisome proliferators leads to the development of hepatocellular carcinomas in rodents (Misra et al., 2013). The mechanism(s) by which these non-mutagenic peroxisome proliferators induce liver tumors remains controversial. Evidence strongly implicates that hyperactivation of PPARα leads to disproportionate large increases in H2O2-generating enzymes, whereas the expression of H2O2-degrading enzymes is only moderately increased (Pyper et al., 2010). This imbalance increases the levels of H2O2 and other ROS in hepatocytes and contributes to oxidative stress, lipid peroxidation, and oxidative DNA damage (Pyper et al., 2010; Misra et al., 2013).
Information on the role of peroxisomes in human tumor development is scarce. It has been shown that the protein levels of peroxisomal branched-chain FA β-oxidation enzymes (i.e., α-methylacyl-CoA racemase, peroxisomal multifunctional protein 2) are upregulated in human prostate cancer (Zha et al., 2005) and that this pathway is essential for optimal proliferation of some prostate cancer cell lines (Zha et al., 2003). Recently, it has been shown that monocarboxylate transporter 2 localizes to peroxisomes in prostate cancer cells and that its expression increases from non-malignant to malignant cells (Valença et al., 2015). It would be important to know if these proteins are selectively upregulated or if peroxisome abundance is also increased in prostate cancer.
A decrease in peroxisome abundance has been observed in various tumor cells, including hepatocellular carcinoma (Litwin et al., 1999), colon carcinoma (Cable et al., 1992; Lauer et al., 1999), breast cancer (el Bouhtoury et al., 1992; Keller et al., 1993), and in renal cell carcinoma (Frederiks et al., 2010). However, so far the mechanism leading to reduced peroxisome abundance was unknown. We explored the relevance of HIF-2α-dependent pexophagy in human clear cell renal cell carcinomas (ccRCC), because loss of VHL function occurs in up to 90% of sporadic human ccRCC and HIF-2α is considered to be a driver oncoprotein for ccRCC. Analysis of more than 200 ccRCC tissue samples revealed that peroxisome abundance is reduced in VHL-deficient ccRCC characterized by high HIF-2α levels (Walter et al., 2014), suggesting that HIF-2α-mediated pexophagy is relevant to human disease. Interestingly, peroxisome abundance is reduced more frequently in well-differentiated tumors, however, it remains to be determined if induction of pexophagy and subsequent loss of peroxisomes promotes or slows down tumor growth. Since HIF-2α stabilization is observed in the vast majority of solid tumors (Franovic et al., 2009; Qing and Simon, 2009), we propose that in addition to ccRCC HIF-2α-mediated pexophagy might also lead to reduced peroxisome abundance in other cancer types.
Metabolic Consequences of Reduced Peroxisome Abundance
Livers of Vhl−∕− and Vhl−∕−/Hif1α−∕− mice are enlarged and display severe steatosis (Rankin et al., 2009; Walter et al., 2014). HIF-2α-mediated reduced peroxisome abundance leads to major changes in the lipid profile of Vhl−∕− and Vhl−∕−/Hif1α−∕− livers, like accumulation of VLCFAs and VLC-PUFAs and depletion of docosahexaenoic acid (DHA) and arachidonic acid (Walter et al., 2014). Furthermore, the levels of the C27-bile acid intermediates 3α,7α-dihydroxycholestanoic acid and 3α,7α,12α-trihydroxycholestanoic acid are significantly increased in the plasma of Vhl−∕− and Vhl−∕−/Hif1α−∕− mice. These lipid changes are characteristic features of human patients and mice lacking peroxisomes (Raymond et al., 2009; Van Veldhoven, 2010; Wanders et al., 2010; Baes and Van Veldhoven, 2012). β-oxidation of VLCFAs and C27-bile acid intermediates occurs only in peroxisomes, peroxisomes play a role in synthesis and degradation of PUFAs, and DHA synthesis requires one cyle of peroxisomal β-oxidation (Van Veldhoven, 2010). Since peroxisomes are essential for plasmalogen biosynthesis and substrates for peroxisomal β-oxidation also include branched-chain FAs, dicarboxylic FAs, and eicosanoids (Van Veldhoven, 2010), it is likely that additional changes in lipid metabolism result from reduced peroxisome abundance in response to HIF-2α activation.
Indirect consequences of reduced peroxisomal metabolism like activation of ER stress pathways and mitochondrial dysfunction might also contribute to alterations in lipid metabolism (Baumgart et al., 2001; Dirkx et al., 2005; Kovacs et al., 2009, 2012). Toxic effects of accumulation of peroxisomal β-oxidation substrates might damage the mitochondrial compartment by altering the lipid composition of mitochondrial membranes. It is well-known that free fatty acids act as potent detergents that can damage cellular membranes (Ho et al., 1995). Membrane properties (e.g., acyl chain order, fluidity, permeability, fusion events, lipid raft microdomains, protein activity) are affected by changes in VLCFAs, VLC-PUFAs, DHA, and plasmalogen levels, influencing secretory and vesicular trafficking pathways (Whitcomb et al., 1988; David et al., 1998; Gleissman et al., 2010; Obara et al., 2013). Hypoxia or loss of VHL function has been shown to delay endocytosis and thereby to enhance receptor tyrosine kinase-mediated signaling (Wang et al., 2009). Thus, lipid alterations as a result of HIF-2α-mediated pexophagy might affect endosomal trafficking and signaling pathways downstream of membrane receptors.
A hallmark of cancer is the reprogramming of metabolism, and recent data suggest that alterations in lipid metabolism play an important role in tumor development (Hirsch et al., 2010; Santos and Schulze, 2012; Currie et al., 2013). Fatty acids support cancer growth by providing substrates for energy production or by generating building blocks for membranes and signaling lipids in proliferating cells (Carracedo et al., 2013). VLC-PUFAs could be converted to eicosanoids, biologically active lipids involved in various pathological processes such as inflammation and cancer. Eicosanoids are degraded in peroxisomes, and loss of peroxisomes affects eicosanoid signaling. Peroxisomes are essential for the synthesis of ether lipids, which represent up to 20% of the total phospholipid mass in humans (Braverman and Moser, 2012; Lodhi and Semenkovich, 2014). Aggressive cancers have high levels of ether lipids, and the expression of the peroxisomal ether lipid synthetic enzyme alkylglyceronephosphate synthase (AGPS) is increased in various cancer cell lines and primary tumors (Benjamin et al., 2013). AGPS knockdown impairs cancer pathogenesis through not only lowering the levels of ether lipids, but also by altering fatty acid, eicosanoid, and glycerophospholipid metabolism, resulting in an overall reduction in the levels of several oncogenic signaling lipids (Benjamin et al., 2013).
Several environmental challenges including ischemia-reperfusion injury, obstructive sleep apnea, viral hepatitis, and alcohol-mediated liver injury are known to induce hepatic hypoxia signaling and are associated with changes in lipid metabolism (Nath and Szabo, 2012). It is tempting to speculate that HIF-2α-mediated pexophagy contributes, at least in some of these pathophysiological conditions, to alterations in lipid metabolism. Decreased plasma and hepatic levels of arachidonic acid and DHA have been observed in patients with non-alcoholic fatty liver disease and non-alcoholic steatohepatitis (Puri et al., 2007, 2009), suggesting impaired peroxisomal metabolism in their pathogenesis. An increasing number of studies suggest that peroxisome dysfunction may be a specific marker for Alzheimer disease. Kou et al. (2011) noted extensive peroxisome-related alterations in Alzheimer disease brains such as increased VLCFAs and decreased plasmalogens containing PUFAs. The question remains if the general loss of peroxisome functions in AD brains is due to pexophagy.
Models How HIF-2α Might Trigger Pexophagy
Since HIF-2α is a transcription factor, the most likely possibility would be that HIF-2α induces the expression of an autophagy receptor and subsequent clustering of peroxisomes via oligomerization of receptor-bound organelles, however, neither Nbr1 nor Sqstm1 are HIF-2α target genes (Walter et al., 2014). Ubiquitination of cargo prone for selective autophagic degradation is the most prevalent autophagy-targeting signal in mammals, and most of the currently known autophagy receptors harbor both ubiquitin-binding domains and LIRs (Kirkin et al., 2009; Stolz et al., 2014). HIF-2α might induce an E3 ubiquitin ligase that mediates the ubiquitination of a PMP (Figure 6A). We propose that HIF-2α signaling increases in this manner NBR1 accumulation on peroxisomes, which in turn serves as a platform for the recruitment of SQSTM1 to achieve a critical mass of autophagy receptors on peroxisomes required for pexophagy (Schönenberger et al., 2015). This might concentrate sufficient ubiquitin-like modifiers (e.g., LC3 and GABARAPs) in close proximity to peroxisomes to prime phagophore assembly. The peroxisomal membrane harbors three E3 ligases (PEX2, PEX10, PEX12) that are essential for peroxisome biogenesis and involved in PEX5 receptor ubiquitination. Their transcriptional induction and concomitant increase of protein levels could increase their ubiquitination capability leading to enhanced ubiquitination of PMPs, but HIF-2α does not induce the expression of those E3 ligases (Walter et al., 2014). Thus, further studies are required to identify and characterize putative E3 ligases involved in HIF-2α-mediated pexophagy.
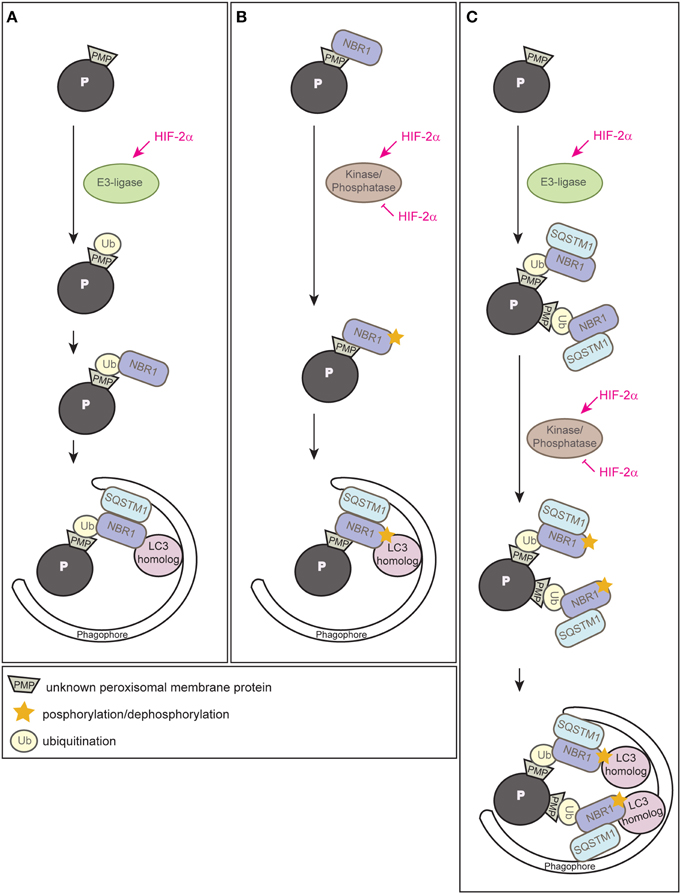
Figure 6. Three alternative models illustrating how HIF-2α might trigger pexophagy. (A) HIF-2α might induce an E3 ubiquitin ligase that ubiquitinates a PMP that enhances the recruitment of the autophagy receptor NBR1 to the peroxisome surface. Accumulation of NBR1 on peroxisomes likely recruits SQSTM1, which was suggested to act as pexophagy co-receptor, and subsequently leads to clustering of peroxisomes via oligomerization of receptor-bound organelles. Accumulation of a critical mass of autophagy receptors might prime phagophore assembly at peroxisomes. (B) NBR1 could be recruited to peroxisomes independently of ubiquitin via its membrane-interacting amphipathic α-helical J domain. HIF-2α might induce or inhibit a kinase/phosphatase that leads to a change in the posttranslational modification of peroxisome-bound NBR1 and thereby triggers recruitment of the autophagic machinery. (C) HIF-2α-dependent activation of pexophagy might be a 2-step process and HIF-2α functions as master regulator that combines two layers of posttranslational modifications to trigger pexophagy. First, it induces an E3 ubiquitin ligase leading to an increased ubiquitination of PMP(s) and subsequent accumulation of NBR1 and SQSTM1 on peroxisomes. Second, HIF-2α activates or inhibits a kinase/phosphatase that leads to a change in the posttranslational modification of peroxisome-bound NBR1 and thereby enhances its binding affinity to a LC3 homolog that finally results in pexophagy. Modified from Schönenberger et al. (2015).
Why does NBR1 localize to peroxisomes in control livers where pexophagy is not induced? In fact, yeast Atg30 and Atg36 also localize to peroxisomes under peroxisome proliferation conditions, but they depend on phosphoregulation for their interactions with components of the autophagy machinery during pexophagy conditions (Farré et al., 2008, 2013). Posttranslational modifications (e.g., phosphorylation, ubiquitination, acetylation) of autophagy proteins are crucial for induction, inhibition, cargo-recognition, and fine-tuning of autophagy. We propose that additional protein modifications are very likely necessary to ultimately drive pexophagy by recruiting and tailoring the autophagic machinery to peroxisomes. For example, phosphorylation as an inducing event of autophagy is conserved from yeast to mammals and has already been discussed above in the context of pexophagy in yeast and mitophagy. Phosphorylation of SQSTM1 and optineurin increases affinity to ubiquitin chains and LC3 (Stolz et al., 2014), and NBR1 phosphorylation by glycogen synthase kinase 3 prevents the aggregation of ubiquitinated proteins and their selective autophagic degradation (Nicot et al., 2014). We propose that HIF-2α governs pexophagy by promoting posttranslational modifications of PMPs and/or autophagy receptors that enhance interactions of receptor-labeled peroxisomes with the autophagic machinery (Figure 6B) (Schönenberger et al., 2015).
Finally, one could envision that HIF-2α-dependent activation of pexophagy is a 2-step process that involves interplay between ubiquitination of a PMP(s) as well as phosphoregulation of autophagy receptors or a PMP(s) as a trigger of pexophagy (Figure 6C) (Schönenberger et al., 2015). Interestingly, a similar interplay promotes PINK1-Parkin-mediated mitophagy whereby phosphorylation of ubiquitin contributes to a feedforward mechanism for ubiquitination events on dysfunctional mitochondria (Ordureau et al., 2014).
A receptor protein complex (RPC) model has been proposed that encompasses the receptor protein as the key player that establishes interactions with ligands, scaffold, and phagophore proteins (Nazarko et al., 2014). The question remains which components of the RPC are involved in HIF-2α-driven pexophagy. Is there an Atg11 homolog in the mammalian liver that would act as a scaffold for HIF-2α-mediated pexophagy? Little is known about mammalian autophagy adaptor proteins that bind to LC3 family members and serve as an anchor point to regulate autophagosome formation around the specific cargo (Stolz et al., 2014). Similar to Atg11, ALFY (autophagy-linked FYVE protein) is a scaffolding protein implicated in aggrephagy that links cargo to the autophagic machinery (Isakson et al., 2013). Moreover, Huntingtin (HTT) has been proposed to serve as adaptor for any type of selective autophagy, because the domain of HTT shares structure similarities and binding activity with the yeast Atg11 protein and interacts with autophagic effector proteins (Ochaba et al., 2014). It is tempting to speculate that ALFY or HTT are part of the RPC mediating HIF-2α-induced pexophagy and thus, functioning as scaffold protein(s). In summary, the identification of HIF-2α as an inducer of pexophagy opens new avenues for studying the underlying molecular mechanism.
Concluding Remarks
We have described hypoxia signaling pathways that regulate function and abundance of mitochondria, ER, and peroxisomes under hypoxia or in response to loss of VHL function. There is emerging evidence that these O2-related organelles exhibit a close functional interplay, and peroxisomal alterations influence mitochondrial and ER functions and vice versa. Although peroxisomal function depends highly on molecular O2, there has been no evidence linking their abundance to O2 availability and HIF-α signaling. In a recent study we identified a unique function of HIF-2α as promoter of pexophagy. An open question that remains to be answered is how HIF-2α induces pexophagy, and we discussed in this review alternative models for how it might trigger pexophagy. Posttranslational modification of autophagy-related proteins and receptors has emerged as an essential regulatory mechanism of selective autophagy. Future studies should address which posttranslational modifications regulate HIF-2α-mediated pexophagy and which components of the receptor protein complex are involved in HIF-2α-mediated pexophagy. In addition, it remains to be determined how HIF-α signaling affects mitochondrial size and ultrastructure and consequently their activity. PPARα modulates metabolic and inflammatory pathways by responding to nutritional signals through ligand activation of transcription, and it is a target of drugs in use and in development to treat diseases. We showed that HIF-α signaling has a repressive effect on ligand-dependent PPARα transcriptional activity, but the mechanism by which HIF-α exerts its inhibitory effect requires further studies. In the past the role of peroxisomes in the cell and in human disease apart from peroxisomal disorders has been grossly underestimated, but this might change given increasing appreciation for the complexity of their interactions with other organelles and the recent discovery of novel functions for peroxisomes. Reduction in peroxisome abundance by pexophagy might positively and negatively impact human disorders including cancer, inflammation, metabolic and neurodegenerative diseases. Along with mechanistic studies of HIF-2α-dependent regulation of pexophagy, the identification of pharmacological regulators of pexophagy might have practical health benefits.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We sincerely apologize to colleagues whose work we could not include due to space limitations. We thank P. Kim, H. Stangl and C. Röhrl for helpful discussions. This work was supported by the Swiss National Science Foundation grant 31003A_132982 to WJK.
Abbreviations
ALFY, autophagy-linked FYVE protein; ALOX15, 15-lipoxygenase-1; ATF, activating transcription factor; Atg, autophagy-related protein; BNIP3, Bcl-2 and adenovirus E1B 19-kDa-interacting protein 3; BNIP3L/NIX, BNIP3-like; ccRCC, clear cell renal cell carcinoma; CHO, Chinese hamster ovary; CHOP, C/EBP homologous protein; CK2, casein kinase 2; COX, cytochrome c oxidase; DAO, D-amino acid oxidase; DHA, docosahexaenoic acid; ER, endoplasmic reticulum; ERAD, ER-associated degradation; ERR, estrogen-related receptor; FA, fatty acid; FOXO3a, forkhead-box protein O3a; FUNDC1, FUN14 domain containing 1; GABARAP, γ-aminobutyric acidreceptor-associated protein; GADD34, growth arrest DNA-inducible gene 34; GLUT, glucose transporter; GRP, glucose-regulated protein; HIF, hypoxiainducible factor; HTT, Huntingtin; IRE1, inositol-requiring protein 1; LC3, microtubule-associated protein-1 light chain 3; LDHA, lactate dehydrogenase A; LIR, LC3-interacting region; LONP, Lon protease; MAX, MYC-associated factor X; MCT4, monocarboxylate transporter 4; mtROS, mitochondrial reactive oxygen species; MXI1, MAX-interacting protein 1; NBR1, neighbor of BRCA1 gene; NRF, nuclear respiratory factor; OMM, outer mitochondrial membrane; OxPhos/ETC, oxidative phosphorylation and electron transport chain; PDH, pyruvate dehydrogenase; PDK1, pyruvate dehydrogenase kinase 1; PERK, protein kinase RNA-like ER kinase; PEX, peroxin; PGAM5, phosphoglycerate mutase family member 5; PHD, prolyl hydroxylase; PGC-1, PPARγ coactivator 1; PINK1, PTEN-induced putative protein kinase 1; PMP, peroxisomal membrane protein; PPAR, peroxisome proliferator-activated receptor; PTEN, phosphatase and tensin homolog; PUFA, polyunsaturated fatty acid; ROS, reactive oxygen species; RPC, receptor protein complex; SQSTM1/p62, sequestosome 1; TNBC, triple-negative breast cancer; ULK1, UNC51-like kinase 1; UOX, urate oxidase; UPR, unfolded protein response; VDAC, voltage-dependent anion channel; VEGF, vascular endothelial growth factor; VHL, von Hippel-Lindau; VLC, very long-chain; XBP1, X-box-binding protein 1.
References
Al-Mehdi, A.-B., Pastukh, V. M., Swiger, B. M., Reed, D. J., Patel, M. R., Bardwell, G. C., et al. (2012). Perinuclear mitochondrial clustering creates an oxidant-rich nuclear domain required for hypoxia-induced transcription. Sci. Signal. 5, ra47. doi: 10.1126/scisignal.2002712
Austin, S., and St-Pierre, J. (2012). PGC1α and mitochondrial metabolism – emerging concepts and relevance in ageing and neurodegenerative disorders. J. Cell Sci. 125, 4963–4971. doi: 10.1242/jcs.113662
Baes, M., and Van Veldhoven, P. P. (2012). Mouse models for peroxisome biogenesis defects and β-oxidation enzyme deficiencies. Biochim. Biophys. Acta 1822, 1489–1500. doi: 10.1016/j.bbadis.2012.03.003
Baird, T. D., and Wek, R. C. (2012). Eukaryotic initiation factor 2 phosphorylation and translational control in metabolism. Adv. Nutr. 3, 307–321. doi: 10.3945/an.112.002113
Baumgart, E., Vanhorebeek, I., Grabenbauer, M., Borgers, M., Declercq, P. E., Fahimi, H. D., et al. (2001). Mitochondrial alterations caused by defective peroxisomal biogenesis in a mouse model for Zellweger Syndrome (PEX5 knockout mouse). Am. J. Pathol. 159, 1477–1494. doi: 10.1016/S0002-9440(10)62534-5
Bellot, G., Garcia-Medina, R., Gounon, P., Chiche, J., Roux, D., Pouysségur, J., et al. (2009). Hypoxia-induced autophagy is mediated through hypoxia-inducible factor induction of BNIP3 and BNIP3L via their BH3 domains. Mol. Cell. Biol. 29, 2570–2581. doi: 10.1128/MCB.00166-09
Benjamin, D. I., Cozzo, A., Ji, X., Roberts, L. S., Louie, S. M., Mulvihill, M. M., et al. (2013). Ether lipid generating enzyme AGPS alters the balance of structural and signaling lipids to fuel cancer pathogenicity. Proc. Natl. Acad. Sci. U.S.A. 110, 14912–14917. doi: 10.1073/pnas.1310894110
Bertolotti, A., Zhang, Y., Hendershot, L. M., Harding, H. P., and Ron, D. (2000). Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2, 326–332. doi: 10.1038/35014014
Bi, M., Naczki, C., Koritzinsky, M., Fels, D., Blais, J., Hu, N., et al. (2005). ER stress-regulated translation increases tolerance to extreme hypoxia and promotes tumor growth. EMBO J. 24, 3470–3481. doi: 10.1038/sj.emboj.7600777
Blais, J. D., Filipenko, V., Bi, M., Harding, H. P., Ron, D., Koumenis, C., et al. (2004). Activating transcription factor 4 is translationally regulated by hypoxic stress. Mol. Cell. Biol. 24, 7469–7482. doi: 10.1128/MCB.24.17.7469-7482.2004
Boland, M. L., Chourasia, A. H., and Macleod, K. F. (2013). Mitochondrial dysfunction in cancer. Front. Oncol. 3:292. doi: 10.3389/fonc.2013.00292
Boström, P., Magnusson, B., Svensson, P.-A., Wiklund, O., Borén, J., Carlsson, L. M. S., et al. (2006). Hypoxia converts human macrophages into triglyceride-loaded foam cells. Arterioscler. Thromb. Vasc. Biol. 26, 1871–1876. doi: 10.1161/01.ATV.0000229665.78997.0b
Braverman, N. E., and Moser, A. B. (2012). Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 1822, 1442–1452. doi: 10.1016/j.bbadis.2012.05.008
Burnett, S. F., Farré, J.-C., Nazarko, T. Y., and Subramani, S. (2015). Peroxisomal Pex3 activates selective autophagy of peroxisomes via interaction with pexophagy receptor, Atg30. J. Biol. Chem. 290, 8623–8631. doi: 10.1074/jbc.M114.619338
Cable, S., Keller, J. M., Colin, S., Haffen, K., Kedinger, M., Parache, R. M., et al. (1992). Peroxisomes in human colon carcinomas. A cytochemical and biochemical study. Virchows Arch. B Cell Pathol. 62, 221–226. doi: 10.1007/BF02899685
Carracedo, A., Cantley, L. C., and Pandolfi, P. P. (2013). Cancer metabolism: fatty acid oxidation in the limelight. Nat. Rev. Cancer 13, 227–232. doi: 10.1038/nrc3483
Chen, G., Han, Z., Feng, D., Chen, Y., Chen, L., Wu, H., et al. (2014a). A regulatory signaling loop comprising the PGAM5 phosphatase and CK2 controls receptor-mediated mitophagy. Mol. Cell 54, 362–377. doi: 10.1016/j.molcel.2014.02.034
Chen, X., Iliopoulos, D., Zhang, Q., Tang, Q., Greenblatt, M. B., Hatziapostolou, M., et al. (2014b). XBP1 promotes triple-negative breast cancer by controlling the HIF1α pathway. Nature 508, 103–107. doi: 10.1038/nature13119
Currie, E., Schulze, A., Zechner, R., Walther, T. C., and Farese, R. V. Jr. (2013). Cellular fatty acid metabolism and cancer. Cell Metab. 18, 153–161. doi: 10.1016/j.cmet.2013.05.017
Dang, C. V., Kim, J. W., Gao, P., and Yustein, J. (2008). The interplay between MYC and HIF in cancer. Nat. Rev. Cancer 8, 51–56. doi: 10.1038/nrc2274
David, D., Sundarababu, S., and Gerst, J. E. (1998). Involvement of long chain fatty acid elongation in the trafficking of secretory vesicles in yeast. J. Cell Biol. 143, 1167–1182. doi: 10.1083/jcb.143.5.1167
De Duve, C. (2007). The origin of eukaryotes: a reappraisal. Nat. Rev. Genet. 8, 395–403. doi: 10.1038/nrg2071
De Duve, C., and Baudhuin, P. (1966). Peroxisomes (microbodies and related particles). Physiol. Rev. 46, 323–357.
Deosaran, E., Larsen, K. B., Hua, R., Sargent, G., Wang, Y., Kim, S., et al. (2013). NBR1 acts as an autophagy receptor for peroxisomes. J. Cell Sci. 126, 939–952. doi: 10.1242/jcs.114819
Dirkx, R., Vanhorebeek, I., Martens, K., Schad, A., Grabenbauer, M., Fahimi, D., et al. (2005). Absence of peroxisomes in mouse hepatocytes causes mitochondrial and ER abnormalities. Hepatology 41, 868–878. doi: 10.1002/hep.20628
Dixit, E., Boulant, S., Zhang, Y., Lee, A. S., Odendall, C., Shum, B., et al. (2010). Peroxisomes are signaling platforms for antiviral innate immunity. Cell 141, 668–681. doi: 10.1016/j.cell.2010.04.018
Dominy, J. E. Jr Lee, Y., Gerhart-Hines, Z., and Puigserver, P. (2010). Nutrient-dependent regulation of PGC-1α's acetylation state and metabolic function through the enzymatic activities of Sirt1/GCN5. Biochim. Biophys. Acta 1804, 1676–1683. doi: 10.1016/j.bbapap.2009.11.023
Doroudgar, S., Thuerauf, D. J., Marcinko, M. C., Belmont, P. J., and Glembotski, C. C. (2009). Ischemia activates the ATF6 branch of the endoplasmic reticulum stress response. J. Biol. Chem. 284, 29735–29745. doi: 10.1074/jbc.M109.018036
el Bouhtoury, F., Keller, J. M., Colin, S., Parache, R. M., and Dauca, M. (1992). Peroxisomal enzymes in normal and tumoral human breast. J. Pathol. 166, 27–35. doi: 10.1002/path.1711660106
Eletto, D., Chevet, E., Argon, Y., and Appenzeller-Herzog, C. (2014). Redox controls UPR to control redox. J. Cell Sci. 127, 3649–3658. doi: 10.1242/jcs.153643
Farré, J.-C., Burkenroad, A., Burnett, S. F., and Subramani, S. (2013). Phosphorylation of mitophagy and pexophagy receptors coordinates their interaction with Atg8 and Atg11. EMBO Rep. 14, 441–449. doi: 10.1038/embor.2013.40
Farré, J.-C., Manjithaya, R., Mathewson, R. D., and Subramani, S. (2008). PpAtg30 tags peroxisomes for turnover by selective autophagy. Dev. Cell 14, 365–376. doi: 10.1016/j.devcel.2007.12.011
Faust, P. L., and Kovacs, W. J. (2014). Cholesterol biosynthesis and ER stress in peroxisome deficiency. Biochimie 98, 75–85. doi: 10.1016/j.biochi.2013.10.019
Fels, D. R., and Koumenis, C. (2006). The PERK/eIF2α/ATF4 module of the UPR in hypoxia resistance and tumor growth. Cancer Biol. Ther. 5, 723–728. doi: 10.4161/cbt.5.7.2967
Franovic, A., Holterman, C. E., Payette, J., and Lee, S. (2009). Human cancers converge at the HIF-2α oncogenic axis. Proc. Natl. Acad. Sci. U.S.A. 106, 21306–21311. doi: 10.1073/pnas.0906432106
Fransen, M., Nordgren, M., Wang, B., and Apanasets, O. (2012). Role of peroxisomes in ROS/RNS-metabolism: implications for human disease. Biochim. Biophys. Acta 1822, 1363–1373. doi: 10.1016/j.bbadis.2011.12.001
Frederiks, W. M., Bosch, K. S., Hoeben, K. A., van Marle, J., and Langbein, S. (2010). Renal cell carcinoma and oxidative stress: the lack of peroxisomes. Acta Histochem. 112, 364–371. doi: 10.1016/j.acthis.2009.03.003
Fukuda, R., Zhang, H., Kim, J. W., Shimoda, L., Dang, C. V., and Semenza, G. L. (2007). HIF-1 regulates cytochrome oxidase subunits to optimize efficiency of respiration in hypoxic cells. Cell 129, 111–122. doi: 10.1016/j.cell.2007.01.047
Gidalevitz, T., Stevens, F., and Argon, Y. (2013). Orchestration of secretory protein folding by ER chaperones. Biochim. Biophys. Acta 1833, 2410–2424. doi: 10.1016/j.bbamcr.2013.03.007
Gleissman, H., Johnsen, J. I., and Kogner, P. (2010). Omega-3 fatty acids in cancer, the protectors of good and the killers of evil? Exp. Cell Res. 316, 1365–1373. doi: 10.1016/j.yexcr.2010.02.039
Gordan, J. D., Bertout, J. A., Hu, C.-J., Diehl, J. A., and Simon, M. C. (2007). HIF-2α promotes hypoxic cell proliferation by enhancing c-Myc transcriptional activity. Cancer Cell 11, 335–347. doi: 10.1016/j.ccr.2007.02.006
Gordan, J. D., Lal, P., Dondeti, V. R., Letrero, R., Parekh, K. N., Oquendo, C. E., et al. (2008). HIF-α effects on c-Myc distinguish two subtypes of sporadic VHL-deficient clear cell renal carcinoma. Cancer Cell 14, 435–446. doi: 10.1016/j.ccr.2008.10.016
Hanna, R. A., Quinsay, M. N., Orogo, A. M., Giang, K., Rikka, S., and Gustafsson, Å. B. (2012). Microtubule-associated protein 1 light chain 3 (LC3) interacts with Bnip3 protein to selectively remove endoplasmic reticulum and mitochondria via autophagy. J. Biol. Chem. 287, 19094–19104. doi: 10.1074/jbc.M111.322933
Hara-Kuge, S., and Fujiki, Y. (2008). The peroxin Pex14p is involved in LC3-dependent degradation of mammalian peroxisomes. Exp. Cell Res. 314, 3531–3541. doi: 10.1016/j.yexcr.2008.09.015
Hasan, S., Platta, H. W., and Erdmann, R. (2013). Import of proteins into the peroxisomal matrix. Front. Physiol. 4:261. doi: 10.3389/fphys.2013.00261
Hirsch, H. A., Iliopoulos, D., Joshi, A., Zhang, Y., Jaeger, S. A., Bulyk, M., et al. (2010). A transcriptional signature and common gene networks link cancer with lipid metabolism and diverse human diseases. Cancer Cell 17, 348–361. doi: 10.1016/j.ccr.2010.01.022
Ho, J. K., Moser, H., Kishimoto, Y., and Hamilton, J. A. (1995). Interactions of a very long chain fatty acid with model membranes and serum albumin. Implications for the pathogenesis of adrenoleukodystrophy. J. Clin. Invest. 96, 1455–1463. doi: 10.1172/JCI118182
Huang, D., Li, T., Zhang, L., Sun, L., He, X., Zhong, X., et al. (2014). HIF-1-mediated suppression of acyl-CoA dehydrogenases and fatty acid oxidation is critical for cancer progression. Cell Rep. 8, 1930–1942. doi: 10.1016/j.celrep.2014.08.028
Hubbi, M. E., Kshitiz, Gilkes, D. M., Rey, S., Wong, C. C., Luo, W., et al. (2013). A nontranscriptional role for HIF-1α as a direct inhibitor of DNA replication. Sci. Signal. 6, ra10. doi: 10.1126/scisignal.2003417
Huss, J. M., Levy, F. H., and Kelly, D. P. (2001). Hypoxia inhibits the peroxisome proliferator-activated receptor α/retinoid X receptor gene regulatory pathway in cardiac myocytes: a mechanism for O2-dependent modulation of mitochondrial fatty acid oxidation. J. Biol. Chem. 276, 27605–27612. doi: 10.1074/jbc.M100277200
Isakson, P., Holland, P., and Simonsen, A. (2013). The role of ALFY in selective autophagy. Cell Death Differ. 20, 12–20. doi: 10.1038/cdd.2012.66
Iwata, J., Ezaki, J., Komatsu, M., Yokota, S., Ueno, T., Tanida, I., et al. (2006). Excess peroxisomes are degraded by autophagic machinery in mammals. J. Biol. Chem. 281, 4035–4041. doi: 10.1074/jbc.M512283200
Jiang, B. H., Semenza, G. L., Bauer, C., and Marti, H. H. (1996). Hypoxia-inducible factor 1 levels vary exponentially over a physiologically relevant range of O2 tension. Am. J. Physiol. 271, C1172–C1180.
Jiang, L., Hara-Kuge, S., Yamashita, S., and Fujiki, Y. (2015). Peroxin Pex14p is the key component for coordinated autophagic degradation of mammalian peroxisomes by direct binding to LC3-II. Genes Cells 20, 36–49. doi: 10.1111/gtc.12198
Kanki, T., Furukawa, K., and Yamashita, S. I. (2015). Mitophagy in yeast: molecular mechanisms and physiological role. Biochim. Biophys. Acta. doi: 10.1016/j.bbamcr.2015.01.005. [Epub ahead of print].
Kanki, T., Wang, K., Cao, Y., Baba, M., and Klionsky, D. J. (2009). Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 17, 98–109. doi: 10.1016/j.devcel.2009.06.014
Keith, B., Johnson, R. S., and Simon, M. C. (2012). HIF1α and HIF2α: sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 12, 9–22. doi: 10.1038/nrc3183
Keller, J. M., Cable, S., el Bouhtoury, F., Heusser, S., Scotto, C., Armbruster, L., et al. (1993). Peroxisome through cell differentiation and neoplasia. Biol. Cell 77, 77–88. doi: 10.1016/S0248-4900(05)80177-7
Kim, J. W., Tchernyshyov, I., Semenza, G. L., and Dang, C. V. (2006). HIF-1-mediated expression of pyruvate dehydrogenase kinase: a metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 3, 177–185. doi: 10.1016/j.cmet.2006.02.002
Kim, P. K., Hailey, D. W., Mullen, R. T., and Lippincott-Schwartz, J. (2008). Ubiquitin signals autophagic degradation of cytosolic proteins and peroxisomes. Proc. Natl. Acad. Sci. U.S.A. 105, 20567–20574. doi: 10.1073/pnas.0810611105
Kirkin, V., McEwan, D. G., Novak, I., and Dikic, I. (2009). A role for ubiquitin in selective autophagy. Mol. Cell 34, 259–269. doi: 10.1016/j.molcel.2009.04.026
Koritzinsky, M., Magagnin, M. G., van den Beucken, T., Seigneuric, R., Savelkouls, K., Dostie, J., et al. (2006). Gene expression during acute and prolonged hypoxia is regulated by distinct mechanisms of translational control. EMBO J. 25, 1114–1125. doi: 10.1038/sj.emboj.7600998
Kou, J., Kovacs, G., Höftberger, R., Kulik, W., Brodde, A., Forss-Petter, S., et al. (2011). Peroxisomal alterations in Alzheimer's disease. Acta Neuropathol. 122, 271–283. doi: 10.1007/s00401-011-0836-9
Koumenis, C., Bi, M., Ye, J., Feldman, D., and Koong, A. C. (2007). Hypoxia and the unfolded protein response. Methods Enzymol. 435, 275–293. doi: 10.1016/s0076-6879(07)35014-3
Koumenis, C., Naczki, C., Koritzinsky, M., Rastani, S., Diehl, A., Sonenberg, N., et al. (2002). Regulation of protein synthesis by hypoxia via activation of the endoplasmic reticulum kinase PERK and phosphorylation of the translation initiation factor eIF2α. Mol. Cell. Biol. 22, 7405–7416. doi: 10.1128/MCB.22.21.7405-7416.2002
Kovacs, W. J., Charles, K. N., Walter, K. M., Shackelford, J. E., Wikander, T. M., Richards, M. J., et al. (2012). Peroxisome deficiency-induced ER stress and SREBP-2 pathway activation in the liver of newborn PEX2 knock-out mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1821, 895–907. doi: 10.1016/j.bbalip.2012.02.011
Kovacs, W. J., Olivier, L. M., and Krisans, S. K. (2002). Central role of peroxisomes in isoprenoid biosynthesis. Prog. Lipid Res. 41, 369–391. doi: 10.1016/S0163-7827(02)00002-4
Kovacs, W. J., Tape, K. N., Shackelford, J. E., Wikander, T. M., Richards, M. J., Fliesler, S. J., et al. (2009). Peroxisome deficiency causes a complex phenotype because of hepatic SREBP/Insig dysregulation associated with endoplasmic reticulum stress. J. Biol. Chem. 284, 7232–7245. doi: 10.1074/jbc.M809064200
Kucejova, B., Sunny, N. E., Nguyen, A. D., Hallac, R., Fu, X., Pena-Llopis, S., et al. (2011). Uncoupling hypoxia signaling from oxygen sensing in the liver results in hypoketotic hypoglycemic death. Oncogene 30, 2147–2160. doi: 10.1038/onc.2010.587
Lauer, C., Völkl, A., Riedl, S., Fahimi, H. D., and Beier, K. (1999). Impairment of peroxisomal biogenesis in human colon carcinoma. Carcinogenesis 20, 985–989. doi: 10.1093/carcin/20.6.985
Li, F., Wang, Y., Zeller, K. I., Potter, J. J., Wonsey, D. R., O'Donnell, K. A., et al. (2005). Myc stimulates nuclearly encoded mitochondrial genes and mitochondrial biogenesis. Mol. Cell. Biol. 25, 6225–6234. doi: 10.1128/MCB.25.14.6225-6234.2005
Li, X., and Gould, S. J. (2002). PEX11 promotes peroxisome division independently of peroxisome metabolism. J. Cell Biol. 156, 643–651. doi: 10.1083/jcb.200112028
Litwin, J. A., Beier, K., Völkl, A., Hofmann, W. J., and Fahimi, H. D. (1999). Immunocytochemical investigation of catalase and peroxisomal lipid β-oxidation enzymes in human hepatocellular tumors and liver cirrhosis. Virchows Arch. 435, 486–495. doi: 10.1007/s004280050432
Liu, L., Cash, T. P., Jones, R. G., Keith, B., Thompson, C. B., and Simon, M. C. (2006). Hypoxia-induced energy stress regulates mRNA translation and cell growth. Mol. Cell 21, 521–531. doi: 10.1016/j.molcel.2006.01.010
Liu, L., Feng, D., Chen, G., Chen, M., Zheng, Q., Song, P., et al. (2012). Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 14, 177–185. doi: 10.1038/ncb2422
Liu, L., Wise, D. R., Diehl, A. J., and Simon, M. C. (2008). Hypoxic reactive oxygen species regulate the integrated stress response and cell survival. J. Biol. Chem. 283, 31153–31162. doi: 10.1074/jbc.M805056200
Lodhi, I. J., and Semenkovich, C. F. (2014). Peroxisomes: a nexus for lipid metabolism and cellular signaling. Cell Metab. 19, 380–392. doi: 10.1016/j.cmet.2014.01.002
Losman, J. A., and Kaelin, W. G. (2013). What a difference a hydroxyl makes: mutant IDH, (R)-2-hydroxyglutarate, and cancer. Genes Dev. 27, 836–852. doi: 10.1101/gad.217406.113
Majmundar, A. J., Wong, W. J., and Simon, M. C. (2010). Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 40, 294–309. doi: 10.1016/j.molcel.2010.09.022
Mannella, C. A. (2006). The relevance of mitochondrial membrane topology to mitochondrial function. Biochim. Biophys. Acta 1762, 140–147. doi: 10.1016/j.bbadis.2005.07.001
Mishra, P., and Chan, D. C. (2014). Mitochondrial dynamics and inheritance during cell division, development and disease. Nat. Rev. Mol. Cell Biol. 15, 634–646. doi: 10.1038/nrm3877
Misra, P., Viswakarma, N., and Reddy, J. (2013). Peroxisome proliferator-activated receptor-α signaling in hepatocarcinogenesis. Subcell. Biochem. 69, 77–99. doi: 10.1007/978-94-007-6889-5_5
Mizushima, N., Yoshimori, T., and Ohsumi, Y. (2011). The role of Atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol. 27, 107–132. doi: 10.1146/annurev-cellbio-092910-154005
Motley, A. M., Nuttall, J. M., and Hettema, E. H. (2012). Pex3-anchored Atg36 tags peroxisomes for degradation in Saccharomyces cerevisiae. EMBO J. 31, 2852–2868. doi: 10.1038/emboj.2012.151
Nath, B., and Szabo, G. (2012). Hypoxia and hypoxia inducible factors: diverse roles in liver diseases. Hepatology 55, 622–633. doi: 10.1002/hep.25497
Nazarko, T. Y., Ozeki, K., Till, A., Ramakrishnan, G., Lotfi, P., Yan, M., et al. (2014). Peroxisomal Atg37 binds Atg30 or palmitoyl-CoA to regulate phagophore formation during pexophagy. J. Cell Biol. 204, 541–557. doi: 10.1083/jcb.201307050
Nicot, A.-S., Lo Verso, F., Ratti, F., Pilot-Storck, F., Streichenberger, N., Sandri, M., et al. (2014). Phosphorylation of NBR1 by GSK3 modulates protein aggregation. Autophagy 10, 92–109. doi: 10.4161/auto.28479
Nordgren, M., and Fransen, M. (2014). Peroxisomal metabolism and oxidative stress. Biochimie 98, 56–62. doi: 10.1016/j.biochi.2013.07.026
Novak, I., Kirkin, V., McEwan, D. G., Zhang, J., Wild, P., Rozenknop, A., et al. (2010). Nix is a selective autophagy receptor for mitochondrial clearance. EMBO Rep. 11, 45–51. doi: 10.1038/embor.2009.256
Obara, K., Kojima, R., and Kihara, A. (2013). Effects on vesicular transport pathways at the late endosome in cells with limited very long-chain fatty acids. J. Lipid Res. 54, 831–842. doi: 10.1194/jlr.M034678
Ochaba, J., Lukacsovich, T., Csikos, G., Zheng, S., Margulis, J., Salazar, L., et al. (2014). Potential function for the Huntingtin protein as a scaffold for selective autophagy. Proc. Natl. Acad. Sci. U.S.A. 111, 16889–16894. doi: 10.1073/pnas.1420103111
Odendall, C., Dixit, E., Stavru, F., Bierne, H., Franz, K. M., Durbin, A. F., et al. (2014). Diverse intracellular pathogens activate type III interferon expression from peroxisomes. Nat. Immunol. 15, 717–726. doi: 10.1038/ni.2915
O'Hagan, K. A., Cocchiglia, S., Zhdanov, A. V., Tambuwala, M. M., Cummins, E. P., Monfared, M., et al. (2009). PGC-1α is coupled to HIF-1α-dependent gene expression by increasing mitochondrial oxygen consumption in skeletal muscle cells. Proc. Natl. Acad. Sci. U.S.A. 106, 2188–2193. doi: 10.1073/pnas.0808801106
Okamoto, K., Kondo-Okamoto, N., and Ohsumi, Y. (2009). Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy. Dev. Cell 17, 87–97. doi: 10.1016/j.devcel.2009.06.013
Ordureau, A., Sarraf, S. A., Duda, D. M., Heo, J.-M., Jedrychowski, M. P., Sviderskiy, V. O., et al. (2014). Quantitative proteomics reveal a feedforward mechanism for mitochondrial PARKIN translocation and ubiquitin chain synthesis. Mol. Cell 56, 360–375. doi: 10.1016/j.molcel.2014.09.007
Papandreou, I., Cairns, R. A., Fontana, L., Lim, A. L., and Denko, N. C. (2006). HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 3, 187–197. doi: 10.1016/j.cmet.2006.01.012
Peck, B., Ferber, E. C., and Schulze, A. (2013). Antagonism between FOXO and Myc regulates cellular powerhouse. Front. Oncol. 3:96. doi: 10.3389/fonc.2013.00096
Pereira, E. R., Frudd, K., Awad, W., and Hendershot, L. M. (2014). Endoplasmic reticulum (ER) stress and hypoxia response pathways interact to potentiate hypoxia-inducible factor 1 (HIF-1) transcriptional activity on targets like vascular endothelial growth factor (VEGF). J. Biol. Chem. 289, 3352–3364. doi: 10.1074/jbc.M113.507194
Pike, L. R., Singleton, D. C., Buffa, F., Abramczyk, O., Phadwal, K., Li, J. L., et al. (2013). Transcriptional up-regulation of ULK1 by ATF4 contributes to cancer cell survival. Biochem. J. 449, 389–400. doi: 10.1042/BJ20120972
Puri, P., Baillie, R. A., Wiest, M. M., Mirshahi, F., Choudhury, J., Cheung, O., et al. (2007). A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 46, 1081–1090. doi: 10.1002/hep.21763
Puri, P., Wiest, M. M., Cheung, O., Mirshahi, F., Sargeant, C., Min, H.-K., et al. (2009). The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 50, 1827–1838. doi: 10.1002/hep.23229
Pyper, S. R., Viswakarma, N., Yu, S., and Reddy, J. K. (2010). PPARalpha: energy combustion, hypolipidemia, inflammation and cancer. Nucl. Recept. Signal. 8, 1–21. doi: 10.1621/nrs.08002
Qing, G., and Simon, M. C. (2009). Hypoxia inducible factor-2α: a critical mediator of aggressive tumor phenotypes. Curr. Opin. Genet. Dev. 19, 60–66. doi: 10.1016/j.gde.2008.12.001
Qu, A., Taylor, M., Xue, X., Matsubara, T., Metzger, D., Chambon, P., et al. (2011). Hypoxia-inducible transcription factor 2α promotes steatohepatitis through augmenting lipid accumulation, inflammation, and fibrosis. Hepatology 54, 472–483. doi: 10.1002/hep.24400
Raimundo, N., Baysal, B. E., and Shadel, G. S. (2011). Revisiting the TCA cycle: signaling to tumor formation. Trends Mol. Med. 17, 641–649. doi: 10.1016/j.molmed.2011.06.001
Rankin, E. B., Rha, J., Selak, M. A., Unger, T. L., Keith, B., Liu, Q., et al. (2009). Hypoxia-inducible factor 2 regulates hepatic lipid metabolism. Mol. Cell. Biol. 29, 4527–4538. doi: 10.1128/MCB.00200-09
Raymond, G. V., Watkins, P., Steinberg, S., and Powers, J. (2009). “Peroxisomal disorders,” in Handbook of Neurochemistry and Molecular Neurobiology: Neural Lipids, eds A. Lajtha, G. Tettamanti, and G. Goracci (Berlin: Springer Science+Business Media), 631–670. doi: 10.1007/978-0-387-30378-9_26
Rogov, V., Dötsch, V., Johansen, T., and Kirkin, V. (2014). Interactions between autophagy receptors and ubiquitin-like proteins form the molecular basis for selective autophagy. Mol. Cell 53, 167–178. doi: 10.1016/j.molcel.2013.12.014
Romero-Ramirez, L., Cao, H., Nelson, D., Hammond, E., Lee, A.-H., Yoshida, H., et al. (2004). XBP1 is essential for survival under hypoxic conditions and is required for tumor growth. Cancer Res. 64, 5943–5947. doi: 10.1158/0008-5472.CAN-04-1606
Rouschop, K. M. A., van den Beucken, T., Dubois, L., Niessen, H., Bussink, J., Savelkouls, K., et al. (2010). The unfolded protein response protects human tumor cells during hypoxia through regulation of the autophagy genes MAP1LC3B and ATG5. J. Clin. Invest. 120, 127–141. doi: 10.1172/JCI40027
Santos, C. R., and Schulze, A. (2012). Lipid metabolism in cancer. FEBS J. 279, 2610–2623. doi: 10.1111/j.1742-4658.2012.08644.x
Scarffe, L. A., Stevens, D. A., Dawson, V. L., and Dawson, T. M. (2014). Parkin and PINK1: much more than mitophagy. Trends Neurosci. 37, 315–324. doi: 10.1016/j.tins.2014.03.004
Scarpulla, R. C., Vega, R. B., and Kelly, D. P. (2012). Transcriptional integration of mitochondrial biogenesis. Trends Endocrinol. Metab. 23, 459–466. doi: 10.1016/j.tem.2012.06.006
Schönenberger, M. J., Krek, W., and Kovacs, W. J. (2015). EPAS1/HIF-2α is a driver of mammalian pexophagy. Autophagy 11, 967–969. doi: 10.1080/15548627.2015.1045180
Schrader, M., Bonekamp, N. A., and Islinger, M. (2012). Fission and proliferation of peroxisomes. Biochim. Biophys. Acta 1822, 1343–1357. doi: 10.1016/j.bbadis.2011.12.014
Schrader, M., and Fahimi, H. D. (2008). The peroxisome: still a mysterious organelle. Histochem. Cell Biol. 129, 421–440. doi: 10.1007/s00418-008-0396-9
Schreiber, A., and Peter, M. (2014). Substrate recognition in selective autophagy and the ubiquitin–proteasome system. Biochim. Biophys. Acta 1843, 163–181. doi: 10.1016/j.bbamcr.2013.03.019
Semenza, G. L. (2010). HIF-1: upstream and downstream of cancer metabolism. Curr. Opin. Genet. Dev. 20, 51–56. doi: 10.1016/j.gde.2009.10.009
Semenza, G. L. (2012). Hypoxia-inducible factors in physiology and medicine. Cell 148, 399–408. doi: 10.1016/j.cell.2012.01.021
Sena, L. A., and Chandel, N. S. (2012). Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 48, 158–167. doi: 10.1016/j.molcel.2012.09.025
Shen, J., Chen, X., Hendershot, L., and Prywes, R. (2002). ER stress regulation of ATF6 localization by dissociation of BiP/GRP78 binding and unmasking of golgi localization signals. Dev. Cell 3, 99–111. doi: 10.1016/S1534-5807(02)00203-4
Shoag, J., and Arany, Z. (2010). Regulation of hypoxia-inducible genes by PGC-1α. Arterioscler. Thromb. Vasc. Biol. 30, 662–666. doi: 10.1161/ATVBAHA.108.181636
Smith, J. J., and Aitchison, J. D. (2013). Peroxisomes take shape. Nat. Rev. Mol. Cell Biol. 14, 803–817. doi: 10.1038/nrm3700
Sowter, H. M., Ratcliffe, P. J., Watson, P., Greenberg, A. H., and Harris, A. L. (2001). HIF-1-dependent regulation of hypoxic induction of the cell death factors BNIP3 and NIX in human tumors. Cancer Res. 61, 6669–6673.
Spiotto, M. T., Banh, A., Papandreou, I., Cao, H., Galvez, M. G., Gurtner, G. C., et al. (2010). Imaging the unfolded protein response in primary tumors reveals microenvironments with metabolic variations that predict tumor growth. Cancer Res. 70, 78–88. doi: 10.1158/0008-5472.CAN-09-2747
Stolz, A., Ernst, A., and Dikic, I. (2014). Cargo recognition and trafficking in selective autophagy. Nat. Cell Biol. 16, 495–501. doi: 10.1038/ncb2979
Sun, R. C., and Denko, N. C. (2014). Hypoxic regulation of glutamine metabolism through HIF1 and SIAH2 supports lipid synthesis that is necessary for tumor growth. Cell Metab. 19, 285–292. doi: 10.1016/j.cmet.2013.11.022
Tabak, H. F., Braakman, I., and van der Zand, A. (2013). Peroxisome formation and maintenance are dependent on the endoplasmic reticulum. Annu. Rev. Biochem. 82, 723–744. doi: 10.1146/annurev-biochem-081111-125123
Thompson, C. B. (2009). Metabolic enzymes as oncogenes or tumor suppressors. N. Engl. J. Med. 360, 813–815. doi: 10.1056/NEJMe0810213
Till, A., Lakhani, R., Burnett, S. F., and Subramani, S. (2012). Pexophagy: the selective degradation of peroxisomes. Int. J. Cell Biol. 2012:512721. doi: 10.1155/2012/512721
Tracy, K., Dibling, B. C., Spike, B. T., Knabb, J. R., Schumacker, P., and Macleod, K. F. (2007). BNIP3 is an RB/E2F target gene required for hypoxia-induced autophagy. Mol. Cell. Biol. 27, 6229–6242. doi: 10.1128/MCB.02246-06
Uniacke, J., Holterman, C. E., Lachance, G., Franovic, A., Jacob, M. D., Fabian, M. R., et al. (2012). An oxygen-regulated switch in the protein synthesis machinery. Nature 486, 126–129. doi: 10.1038/nature11055
Valença, I., Pértega-Gomes, N., Vizcaino, J. R., Henrique, R. M., Lopes, C., Baltazar, F., et al. (2015). Localization of MCT2 at peroxisomes is associated with malignant transformation in prostate cancer. J. Cell. Mol. Med. 19, 723–733. doi: 10.1111/jcmm.12481
Vander Heiden, M. G., Cantley, L. C., and Thompson, C. B. (2009). Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324, 1029–1033. doi: 10.1126/science.1160809
Van Veldhoven, P. P. (2010). Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. J. Lipid Res. 51, 2863–2895. doi: 10.1194/jlr.R005959
Walter, K. M., Schönenberger, M. J., Trötzmüller, M., Horn, M., Elsässer, H.-P., Moser, A. B., et al. (2014). Hif-2α promotes degradation of mammalian peroxisomes by selective autophagy. Cell Metab. 20, 882–897. doi: 10.1016/j.cmet.2014.09.017
Walter, P., and Ron, D. (2011). The unfolded protein response: from stress pathway to homeostatic regulation. Science 334, 1081–1086. doi: 10.1126/science.1209038
Wanders, R. J. A., Ferdinandusse, S., Brites, P., and Kemp, S. (2010). Peroxisomes, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1801, 272–280. doi: 10.1016/j.bbalip.2010.01.001
Wang, Y., Roche, O., Yan, M. S., Finak, G., Evans, A. J., Metcalf, J. L., et al. (2009). Regulation of endocytosis via the oxygen-sensing pathway. Nat. Med. 15, 319–324. doi: 10.1038/nm.1922
Whitcomb, R. W., Linehan, W. M., and Knazek, R. A. (1988). Effects of long-chain, saturated fatty acids on membrane microviscosity and adrenocorticotropin responsiveness of human adrenocortical cells in vitro. J. Clin. Invest. 81, 185–188. doi: 10.1172/JCI113292
Winklhofer, K. F. (2014). Parkin and mitochondrial quality control: toward assembling the puzzle. Trends Cell Biol. 24, 332–341. doi: 10.1016/j.tcb.2014.01.001
Wouters, B. G., and Koritzinsky, M. (2008). Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat. Rev. Cancer. 8, 851–864. doi: 10.1038/nrc2501
Wu, W., Tian, W., Hu, Z., Chen, G., Huang, L., Li, W., et al. (2014). ULK1 translocates to mitochondria and phosphorylates FUNDC1 to regulate mitophagy. EMBO Rep. 15, 566–575. doi: 10.1002/embr.201438501
Yamashita, S., Abe, K., Tatemichi, Y., and Fujiki, Y. (2014). The membrane peroxin PEX3 induces peroxisome-ubiquitination-linked pexophagy. Autophagy 10, 1549–1564. doi: 10.4161/auto.29329
Zha, S., Ferdinandusse, S., Denis, S., Wanders, R. J., Ewing, C. M., Luo, J., et al. (2003). α-Methylacyl-CoA racemase as an androgen-independent growth modifier in prostate cancer. Cancer Res. 63, 7365–7376.
Zha, S., Ferdinandusse, S., Hicks, J. L., Denis, S., Dunn, T. A., Wanders, R. J., et al. (2005). Peroxisomal branched chain fatty acid β-oxidation pathway is upregulated in prostate cancer. Prostate 63, 316–323. doi: 10.1002/pros.20177
Zhang, H., Bosch-Marce, M., Shimoda, L. A., Tan, Y. S., Baek, J. H., Wesley, J. B., et al. (2008). Mitochondrial autophagy is an HIF-1-dependent adaptive metabolic response to hypoxia. J. Biol. Chem. 283, 10892–10903. doi: 10.1074/jbc.M800102200
Zhang, H., Gao, P., Fukuda, R., Kumar, G., Krishnamachary, B., Zeller, K. I., et al. (2007). HIF-1 inhibits mitochondrial biogenesis and cellular respiration in VHL-deficient renal cell carcinoma by repression of C-MYC activity. Cancer Cell 11, 407–420. doi: 10.1016/j.ccr.2007.04.001
Zhang, J., and Ney, P. A. (2009). Role of BNIP3 and NIX in cell death, autophagy, and mitophagy. Cell Death Differ. 16, 939–946. doi: 10.1038/cdd.2009.16
Zhao, S., Lin, Y., Xu, W., Jiang, W., Zha, Z., Wang, P., et al. (2009). Glioma-derived mutations in IDH1 dominantly inhibit IDH1 catalytic activity and induce HIF-1α. Science 324, 261–265. doi: 10.1126/science.1170944
Keywords: endoplasmic reticulum, ER stress, hypoxia, HIF-α, mitochondria, mitophagy, peroxisomes, pexophagy
Citation: Schönenberger MJ and Kovacs WJ (2015) Hypoxia signaling pathways: modulators of oxygen-related organelles. Front. Cell Dev. Biol. 3:42. doi: 10.3389/fcell.2015.00042
Received: 21 April 2015; Accepted: 15 June 2015;
Published: 21 July 2015.
Edited by:
Michael Schrader, University of Exeter, UKReviewed by:
J. Kalervo Hiltunen, University of Oulu, FinlandChangle Ma, Shandong Normal University, China
Copyright © 2015 Schönenberger and Kovacs. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Werner J. Kovacs, Department of Biology, Institute of Molecular Health Sciences, ETH Zurich, Otto-Stern-Weg 7, HPL H16, CH-8093 Zurich, Switzerland, werner.kovacs@biol.ethz.ch
†Present Address: Miriam J. Schönenberger, Institute of Physiology, University of Zurich, Zurich, Switzerland