 Michel Chrétien
Michel Chrétien- Functional Endoproteolysis Laboratory, Institut de Recherches Cliniques de Montréal, Ottawa Hospital Research Institute, Montreal, QC, Canada
This Prohormone Theory was simultaneously proposed in 1967 by two independent groups using two different approaches and two experimental models. Donald Steiner, in elegant pulse-chase experiments, proposed the existence of proinsulin when he observed that a human insulinoma was producing higher MW forms of immunoreactive insulin, subsequently transformed into insulin-like material (1). Simultaneously and independently, Michel Chrétien, based on amino acid sequence homologies between three pituitary peptides, β-lipotropic hormone (β-LPH), γ-LPH, and β-melanocyte-stimulating hormone (β-MSH), concluded that active peptide hormones are derived from endoproteolytic cleavages of inactive precursors, apparently at pairs of basic amino acids (2). One year later, Donald Chance confirmed that the cleavage sites in proinsulin were also made of paired basic amino acids (3). This novel paradigm solved two major controversies on the biosynthesis of both insulin and neuropeptides. This short review describes how.
The Insulin Saga
In the mid 1950s and early 1960s, many scientists wandered how insulin was synthesized in the pancreatic β-cells. Fred Sanger had established that insulin is made of two peptide chains linked by disulfide bridges (4). A prevailing view was that insulin was biosynthesized as two separate peptide chains “zipped” post-transcriptionally by interchain disulfide bridges. Preceding that period, Oliver Smithies, as mentioned in his 2007 Nobel Lecture, was looking for a precursor to insulin “which I never found” (5). Many groups in the US, Canada, China, and Germany, using separate insulin A and B chains, had tried to reconstitute insulin in vitro, with minimal yield (6). In the mid-1960s, some in vitro studies of pancreatic islet tissue had led to the conclusion that the two insulin chains were biosynthesized as separate entities. The controversy was definitely solved, when the amino acid sequence of proinsulin unequivocally proved that insulin is made as a single polypeptide, subsequently modified to its active form by endoproteolysis at pairs of basic residues (3).
The Neuropeptide Saga
The β-LPH/γ-LPH/β-MSH model of biosynthesis, initially proposed in 1967 for pituitary peptides, contained elements that would solved the upcoming controversy on neuropeptide biosynthesis. In 1969, the field of neuroendocrinology underwent a revolution when Roger Guillemin published the astonishing discovery that thyrotropin releasing factor (TRF) was a tripeptide (7). It was suggested that this tripeptide, like glutathione, was produced by a soluble non-ribosomal enzymatic mechanism and the existence of a TRH synthetase was seriously considered.
The first indication that β-LPH could be a neuropeptide precursor came when Hughes and Kosterlitz published the amino acid sequence of met-enkephalin and noted that it corresponded to residues 61–65 of β-LPH (8). Shortly thereafter, many groups revealed that the main opioid secretory product was the fragment 61–91 of β-LPH, now universally known as β-endorphin, a strong indication that β-LPH (1–91) was its most plausible precursor candidate. Definite proof that β-LPH is the precursor of β-endorphin came about when it was unequivocally demonstrated that β-endorphin is produced by endoproteolytic cleavage of β-LPH at pair basic residues 59–60 (9).
Coincidently, it was realized that β-LPH itself is part of a larger precursor containing ACTH. The existence of this precursor was confirmed with the cloning of its cDNA; in it, the sequences of its active end products (β-endorphin, MSHs, and ACTH) are flanked by the canonical pairs of basic residues (10). The precursor, now named proopiomelanocortin (POMC) (11), has become the gold standard of endocrine and neuroendocrine precursors. Soon after, the cloning of cDNAs for the other neuropeptides confirmed that all of them were produced through a similar mechanism (12). The non-ribosomal enzymatic concept of TRH production thus became obsolete. The endoproteolysis of polyproteins like ProTRF and POMC (Figure 1) greatly amplifies the multiple active end products of large precursor molecules (13).
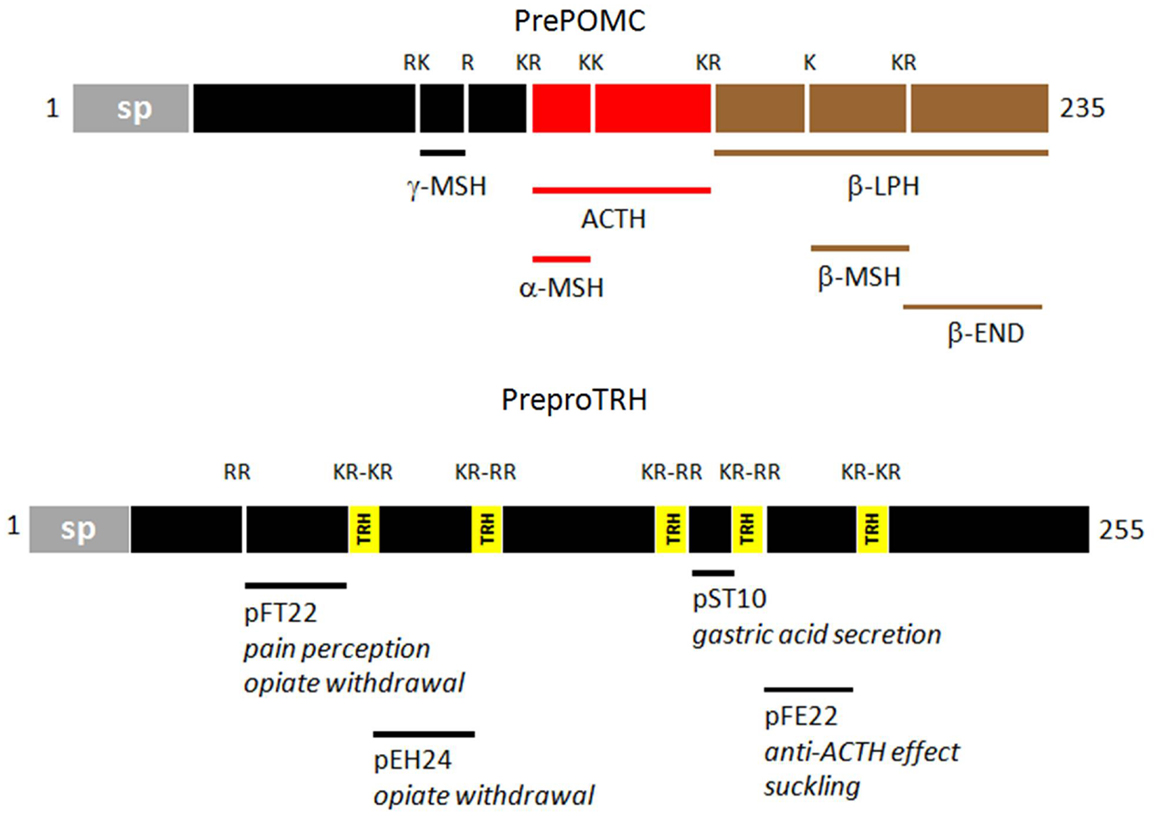
Figure 1. Diagrammatic representations of prePOMC and preproTRH. The diagrams are based on rat sequences. The additional functional peptides of proTRH have been reviewed by Nillni and Sevarino (21). Single and paired basic residues (K/R) flanking the functional peptides are shown. (sp: signal peptide).
During the following decades, post-translational endoproteolytic activation became applicable to numerous other polyproteins, including precursors to neurotrophins, growth factors, transcription factors, receptors, extracellular matrix proteins, bacterial toxins, viral glycoproteins, etc. It is now recognized as a fundamental cellular process, affecting many biological functions and opened a new chapter in biology (13, 14).
The 1967 Prohormone Theory and its biological ramifications implied the existence of endoproteolytic enzymes dedicated to the process (15). These were discovered 23 years later (16, 17). Collectively, they are called proprotein convertases (PCs) or proproteins convertases subtilisin/kexin type (PCSKs). They are calcium-dependent serine endoproteases, structurally related to bacterial Subtilisin and to yeast Kexin (13, 18, 19). The first two, PC1/3-PC2 are considered the prototypical convertases for prohormones and proneuropeptides.
Conclusion
In solving two major controversies concerning the biosynthetic pathways of insulin and neuropeptides, the 1967 prohormone theory has become a tenet of the peptidergic systems in endocrinology and neuroendocrinology. This is one of many other examples in biology whereby incompatible hypotheses are clarified by one type of results. One of the most famous is certainly the 1943 fluctuation test of Salvador Luria and Max Delbruck (20). Although less spectacular than the genetics of bacterial resistance, the prohormone concept ended two scientific debates and led to new horizons which surpassed all the most elaborate expectations.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from the Canadian Institutes of Health Research, the Richard and Edith Strauss Canada Foundation, and the J. Louis Lévesque Foundation.
References
1. Steiner DF, Cunningham D, Spigelman L, Aten B. Insulin biosynthesis: evidence for a precursor. Science (1967) 157:697–700. doi:10.1126/science.157.3789.697
2. Chrétien M, Li CH. Isolation, purification, and characterization of gamma-lipotropic hormone from sheep pituitary glands. Can J Biochem (1967) 45:1163–74. doi:10.1139/o67-133
3. Chance RE, Ellis RM, Bromer WW. Porcine proinsulin: characterization and amino acid sequence. Science (1968) 161:165–7. doi:10.1126/science.161.3837.165
4. Sanger F. Chemistry of insulin; determination of the structure of insulin opens the way to greater understanding of life processes. Science (1959) 129:1340–4. doi:10.1126/science.129.3359.1340
5. Smithies O. Turning pages (Nobel lecture). Chembiochem (2008) 9:1342–59. doi:10.1002/cbic.200800205
6. Cahill GF Jr. Insulin and proinsulin. N Engl J Med (1970) 283:762. doi:10.1056/NEJM197010012831412
7. Guillemin R. Peptides in the brain: the new endocrinology of the neuron. Science (1978) 202:390–402. doi:10.1126/science.212832
8. Hughes J, Smith TW, Kosterlitz HW, Fothergill LA, Morgan BA, Morris HR. Identification of two related pentapeptides from the brain with potent opiate agonist activity. Nature (1975) 258:577–80. doi:10.1038/258577a0
9. Crine P, Benjannet S, Seidah NG, Lis M, Chrétien M. In vitro biosynthesis of beta-endorphin, gamma-lipoprotein, and beta-lipotropin by the pars intermedia of beef pituitary glands. Proc Natl Acad Sci U S A (1977) 74:4276–80.
10. Nakanishi S, Inoue A, Kita T, Nakamura M, Chang AC, Cohen SN, et al. Nucleotide sequence of cloned cDNA for bovine corticotropin-beta-lipotropin precursor. Nature (1979) 278:423–7. doi:10.1038/278423a0
11. Chrétien M, Benjannet S, Gossard F, Gianoulakis C, Crine P, Lis M, et al. From beta-lipotropin to beta-endorphin and ‘pro-opio-melanocortin’. Can J Biochem (1979) 57:1111–21. doi:10.1139/o79-143
12. Douglass J, Civelli O, Herbert E. Polyprotein gene expression: generation of diversity of neuroendocrine peptides. Annu Rev Biochem (1984) 53:665–715. doi:10.1146/annurev.bi.53.070184.003313
13. Chrétien M. My road to Damascus: how I converted to the prohormone theory and the proprotein convertases. Biochem Cell Biol (2012) 90:750–68. doi:10.1139/o2012-031
14. Seidah NG, Chretien M. Proprotein and prohormone convertases: a family of subtilases generating diverse bioactive polypeptides. Brain Res (1999) 848:45–62. doi:10.1016/S0006-8993(99)01909-5
15. Lazure C, Seidah NG, Pelaprat D, Chretien M. Proteases and posttranslational processing of prohormones: a review. Can J Biochem Cell Biol (1983) 61:501–15. doi:10.1139/o83-066
16. Seidah NG, Gaspar L, Mion P, Marcinkiewicz M, Mbikay M, Chrétien M. cDNA sequence of two distinct pituitary proteins homologous to Kex2 and furin gene products: tissue-specific mRNAs encoding candidates for pro-hormone processing proteinases. DNA Cell Biol (1990) 9:789. doi:10.1089/dna.1990.9.415
17. Smeekens SP, Steiner DF. Identification of a human insulinoma cDNA encoding a novel mammalian protein structurally related to the yeast dibasic processing protease Kex2. J Biol Chem (1990) 265: 2997–3000.
19. Seidah NG, Prat A. The biology and therapeutic targeting of the proprotein convertases. Nat Rev Drug Discov (2012) 11:367–83. doi:10.1038/nrd3699
20. Luria SE, Delbruck M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics (1943) 28:491–511.
Keywords: prohormone theory, proprotein convertases, peptide hormones, neuropeptides, biosynthesis
Citation: Chrétien M (2013) How the prohormone theory solved two important controversies in hormonal and neural peptide biosynthesis. Front. Endocrinol. 4:148. doi: 10.3389/fendo.2013.00148
Received: 09 July 2013; Paper pending published: 03 September 2013;
Accepted: 02 October 2013; Published online: 23 October 2013.
Edited by:
Hubert Vaudry, University of Rouen, FranceReviewed by:
José A. Muñoz-Cueto, University of Cadiz, SpainBernard Pierre Roques, Université Paris Descartes, France
Copyright: © 2013 Chrétien. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michel Chrétien, Functional Endoproteolysis Laboratory, Institut de Recherches Cliniques de Montréal, Ottawa Hospital Research Institute, 110 Avenue des Pins Ouest, Montreal, QC H2W 1R7, Canada e-mail: mchretien@ohri.ca