 Matty Knight1*
Matty Knight1* Halime D. Arican-Goktas2
Halime D. Arican-Goktas2 Wannaporn Ittiprasert3
Wannaporn Ittiprasert3 Edwin C. Odoemelam2
Edwin C. Odoemelam2 André N. Miller3
André N. Miller3 Joanna M. Bridger2
Joanna M. Bridger2- 1Department of Microbiology, Immunology and Tropical Medicine, The George Washington University, Washington, DC, USA
- 2Biosciences, Health Sciences and Social Care, Brunel University London, London, UK
- 3Schistosomiasis, Biomedical Research Institute, Rockville, MD, USA
Biomphalaria glabrata snails play an integral role in the transmission of Schistosoma mansoni, the causative agent for human schistosomiasis in the Western hemisphere. For the past two decades, tremendous advances have been made in research aimed at elucidating the molecular basis of the snail/parasite interaction. The growing concern that there is no vaccine to prevent schistosomiasis and only one effective drug in existence provides the impetus to develop new control strategies based on eliminating schistosomes at the snail-stage of the life cycle. To elucidate why a given snail is not always compatible to each and every schistosome it encounters, B. glabrata that are either resistant or susceptible to a given strain of S. mansoni have been employed to track molecular mechanisms governing the snail/schistosome relationship. With such snails, genetic markers for resistance and susceptibility were identified. Additionally, differential gene expression studies have led to the identification of genes that underlie these phenotypes. Lately, the role of schistosomes in mediating non-random relocation of gene loci has been identified for the first time, making B. glabrata a model organism where chromatin regulation by changes in nuclear architecture, known as spatial epigenetics, orchestrated by a major human parasite can now be investigated. This review will highlight the progress that has been made in using molecular approaches to describe snail/schistosome compatibility issues. Uncovering the signaling networks triggered by schistosomes that provide the impulse to turn genes on and off in the snail host, thereby controlling the outcome of infection, could also yield new insights into anti-parasite mechanism(s) that operate in the human host as well.
Introduction
The neglected tropical disease (NTD), schistosomiasis, ranks second only to malaria as one of the most persistent debilitating diseases in impoverished areas of the tropics and sub-tropics, mainly in sub-Saharan Africa (Jenkins-Holick and Kaul, 2013). Freshwater snails are the obligate intermediate hosts for the transmission of the causative schistosome parasite. Using a combination of mass drug administration (MDA), mainly in school-aged children, and molluscicides to eliminate the snail from fresh water bodies, such as lakes, rivers, and their tributaries, low prevalence of schistosomiasis has now been achieved in several endemic countries (Tchuem Tchuente et al., 2013; Olveda et al., 2014). Despite these control efforts, however, reduction of schistosomiasis for the long term remains elusive, especially in poorer countries where access to clean water and sanitation remains challenging. Increasingly, strong advocacy by organizations, such as the schistosomiasis control initiative (SCI) and schistosomiasis consortium organization research effort (SCORE) have called for a renewed effort to better detect the parasite in snail populations where low prevalence of schistosomiasis has been accomplished (Fenwick et al., 2009a,b; Garba et al., 2009; Musuva et al., 2014). It is believed that by using an integrated control approach with aggressive surveillance complete global elimination of schistosomiasis might even become possible (Rollinson et al., 2013).
To make this goal a reality, however, novel intervention tools targeting the intermediate snail host are needed, especially since only one drug, praziquantel, effectively treats schistosomiasis and no vaccine is available (Hotez et al., 2010; Chai, 2013). Left untreated, schistosome adult worms can survive for several years (between 5 and 30 has been recorded). Eggs from mating not passed with human excreta but remaining trapped in tissues are the main cause of the pathology associated with schistosomiasis. This depends on the infecting parasite species, for example Schistosoma haematobium, can in time cause bladder cancer expedited from eggs sitting in tissues of this organ. Most notably, chronic S. haematobium infection can also cause female genital schitosomiasis (FGS), a condition that results in infertility and predisposes this section of society to HIV infection (Kjetland et al., 2010; Downs et al., 2013; Owusu-Bempah et al., 2013; Sahu et al., 2013).
Given the current interest in eliminating schistosomiasis, frequent use of molluscicides can have a negative impact on the environment as well as damage delicate ecosystems. Therefore, as was suggested as early as in the 1950’s, a form of biological control can be adopted by using incompatible snails to replace resident susceptible ones in endemic foci (Ruiz-Tiben et al., 1969; Ferguson and Ruiz-Tiben, 1971; Jobin and Laracuente, 1979). Indeed, in a proof of concept study conducted in the Caribbean Island of St Lucia, the snail Biomphalaria straminae a secondary host of the parasite S. mansoni, not its compatible snail B. glabrata, was utilized to eliminate schistosomiasis in the part of the island where this form of control was adopted (Pointier, 1993). In a more recent Brazilian study, the introduction of parasite resistant strains of B. tenagophila into an endemic site was found to reduce transmission as cross hybridization between resident susceptible and introduced snails increased over time (De Almeida Marques et al., 2014). This alternative intervention method, focusing on reducing schistosomiasis by blocking the snail stage of the parasite life cycle although attractive, will require a better understanding of the molecular basis of the snail and parasite interaction. This review will highlight recent advances that have been made toward unraveling molecular mechanisms that underlie parasite development in the snail host, particularly in the role of spatial epigenetics in the outcome of schistosome infection in the snail host.
Genetics of the Snail Host and How the Parasite Shapes the Outcome of Infection
Over the past two decades a major shift occurred in studies investigating snail/schistosome compatibility issues, from a comparative immunological point of view to one with a more molecular focus. Facilitating these studies was the existence of resources enabling investigators to identify genes involved in the snail’s behavior toward the parasite. These include the B. glabrata embryonic cell line (Bge), and snail stocks that have been bred to display either resistance or susceptibility to S. mansoni.
Susceptibility of the snail host to the parasite has a genetic basis with genes of both the snail and parasite affecting the outcome (Spada et al., 2002; Theron and Coustau, 2005; Oliveira et al., 2008). For instance, resistance to infection in adult snail stocks, such as BS-90, 13-16-R1, and 10-R2 is a single dominant gene trait that is heritable by simple Mendelian genetics (Knight et al., 1991; Spada et al., 2002). Resistance in juvenile snails, however, is a complex trait that is controlled by several genes, five or six, each with multiple alleles (Ittiprasert et al., 2010).
Susceptibility to S. mansoni in B. glabrata snail stocks, such as M-line and NMRI is also a complex trait either as adults or juveniles. In some cases, snails can display susceptibility as juveniles, become resistant as adults, and revert to the susceptibility phenotype once they age (Knight et al., 1991; Richards et al., 1992). All these different outcomes of the parasite/snail encounter points to a relationship that is more variable and unpredictable in the snail than in the human host. Indeed, while a given parasite strain can only develop within a compatible snail, no such discrimination occurs in the human host. For this reason, it is highly plausible that discovery, in the snail host, of molecular pathways that either prevent or facilitate parasite development, if evolutionarily conserved, will help in designing tools that can be utilized for targeted control of schistosomiasis in both the snail vector and the human host.
Gene Discovery Has Been Accomplished by Examining Differential Gene Expression in Resistant, and Susceptible Snails
To determine the effect of parasite infection on B. glabrata gene expression, snail stocks that are either resistant (BS-90) or susceptible (NMRI, and M-line), exposed at various time points to S. mansoni, were examined by a variety of approaches to assess the degree of parasite-mediated modulation of the snail transcriptional profile. The variety of methods utilized to examine differential gene expression between these snails, included generating expressed sequence tags (ESTs) from either intact snails or specific tissues (hemocytes, ovotestis, mantle, hepatopancreas, and albumen gland), differential display reverse transcription-polymerase chain reaction (DD-RT-PCR), microarray analysis, and RNA-sequencing (RNA-seq; Knight et al., 2000; Raghavan et al., 2003; Mitta et al., 2005; Lockyer et al., 2007, 2008; Ittiprasert et al., 2009, 2010). In addition, a suppression subtractive hybridization (SSH)-cDNA cloning strategy was employed to enrich for transcripts featuring prominently between resistant and susceptible snails during the early response phase to the parasite (Ittiprasert et al., 2010).
From these various profiling experiments, results showed that transcripts involved in the snail’s innate defense system (IDS) were amongst those that were most significantly upregulated after early exposure to the parasite, especially in resistant snails. Thus in resistant snails, such as the BS-90 stock, compared to susceptible ones, e.g., the NMRI, and M-line snail stocks, immune-defense behavior of the snail/schistosome interplay, involving cellular, “macrophage-like” cells known as hemocytes, and the plasma, hemolymph, component, recognize the incoming parasite (larval miracidium) as non-self by an as yet unknown mechanism. This recognition results in the encapsulation of the parasite by hemocytes released into the open circulatory system from the snail’s hemopoietic tissue, the amoebocyte producing organ (APO) located in the pericardiac and renal regions (Souza Sdos and Andrade, 2006, 2012). The nature of the excretory-secretory products (ESPs) released from the parasite that stimulate hemocyte production, recruitment, and attachment to the parasite surface, remain unknown. However, hemocyte genes that are up-regulated in response to exposure to schistosomes and their ESPs are being identified (Zahoor et al., 2009, 2014; Lockyer et al., 2012). Aided by plasma products, including lectins, hemocytes encapsulate the parasite within a few hours post-exposure, and by a cytotoxic mechanism(s) that include the production of reactive oxygen and nitrogen intermediates, kill the parasite (Hahn et al., 2001).
Hemocytes passively transferred from a resistant to susceptible snail have been shown to be capable of rendering these snails less susceptible (Vasquez and Sullivan, 2001). This active anti-schistosome defense reaction in resistant snails does not occur in those that are compatible/susceptible. In this case, the prevailing hypothesis is that the parasite is capable of suppressing the snail IDS, leading to the successful establishment and development of the early mother sporocyst stage (Yoshino and Bayne, 1983; Mattos et al., 2011). What mechanism(s) are involved in the manipulation of the snail’s defense by the parasite has yet to be uncovered but from comprehensive transcriptomics, several lectins, including highly diversified transcripts encoding the fibrinogen encoding proteins, collectively known by the acronym fibrinogen-related proteins (FREPs) are upregulated in both resistant and susceptible snails. Studies using experimental dsRNA to knock down the transcript for FREP 3 in resistant snails rendered them susceptible, indicating that FREPs plays a role in anti-trematode defense not coagulation (Zhang and Loker, 2004; Hanington et al., 2012). The unusual diversification of FREPs, the first to be discovered for a defense-related molecule in invertebrates, points to an IDS that is more complex than was previously envisioned. Snail lectins, including FREPs also bind to released parasite molecules, such as mucin (Roger et al., 2008a; Zhang et al., 2008; Hanington et al., 2010). This interaction between snail and parasite molecules that have co-evolved sufficiently to “match” has been proposed as underlying snail/schistosome compatibility (Massoud et al., 2004; Oliveira et al., 2008; Roger et al., 2008a,b; Mitta et al., 2012). Lately, evidence for specific-genotype immune priming has also been detected in the B. glabrata/schistosome interaction (Portela et al., 2013). Thus far, however, interacting components known to trigger innate immunity in other invertebrates, specifically pathogen-associated molecular patterns (PAMPs), as well as host pattern recognition receptors (PRRs), including signaling PRRs, e.g., the membrane-bound Toll receptors remain to be characterized for the B. glabrata/S. mansoni relationship (Meurs et al., 2011; Paveley et al., 2011).
Considerable progress is being made on identifying shared molecular moieties in the parasite and snail host, and evidence is emerging that shows the existence of shared specific glycotypes (glycan epitopes) between the larval schistosome parasite and snail hemocytes (Yoshino et al., 2013). It is believed that the shared epitopes, also called molecular mimicry might be used to dampen immune recognition of the parasite thus evading immune detection.
Stress Induction by Schistosomes in the Snail Host Correlates with Susceptibility
From studies that compared the temporal regulation in gene expression between resistant and susceptible snails exposed to S. mansoni, it was detected that several stress related transcripts were up-regulated early and significantly in susceptible juvenile snails compared to their resistant counterparts (Figures 1A,B; Ittiprasert et al., 2010). For example, transcripts encoding heat shock protein (Hsp) 70, Hsp 90, and the reverse transcriptase (RT) domain of the B. glabrata non-LTR retro-transposable element, nimbus, were detected in susceptible NMRI snails at early time points (5 h) post-exposure compared to the resistant BS-90 snail. Furthermore, upregulation of the aforementioned stress related transcripts was observed in snails exposed to normal but not to irradiated miracidia (Ittiprasert et al., 2010).
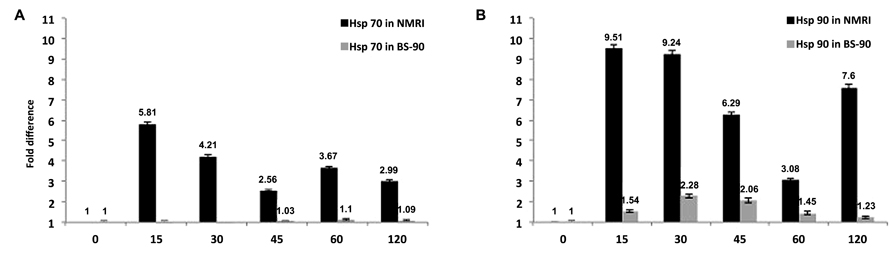
FIGURE 1. Temporal modulation of the transcript encoding (A) Hsp 70 or (B) Hsp 90 in susceptible NMRI and resistant (BS–90) B. glabrata snail stocks examined by qPCR after exposure (0 – 120 min) to S. mansoni miracidia. Note that expression of the transcripts occurs only in the susceptible but not in the resistant snail (Ittiprasert et al., 2010; Ittiprasert and Knight, 2012).
Genome Organisation in B. Glabrata is Similar to Higher Organisms
In order to understand how the parasite is able to induce and even regulate gene expression – such as the heat shock genes, it is imperative to understand the functional genome biology and its organization within the host cell nuclei. Furthermore, to fully appreciate how the parasite is controlling gene expression the structures that are involved in genome organization and behavior also need to be elucidated (see Figures 2A,B,F,G).
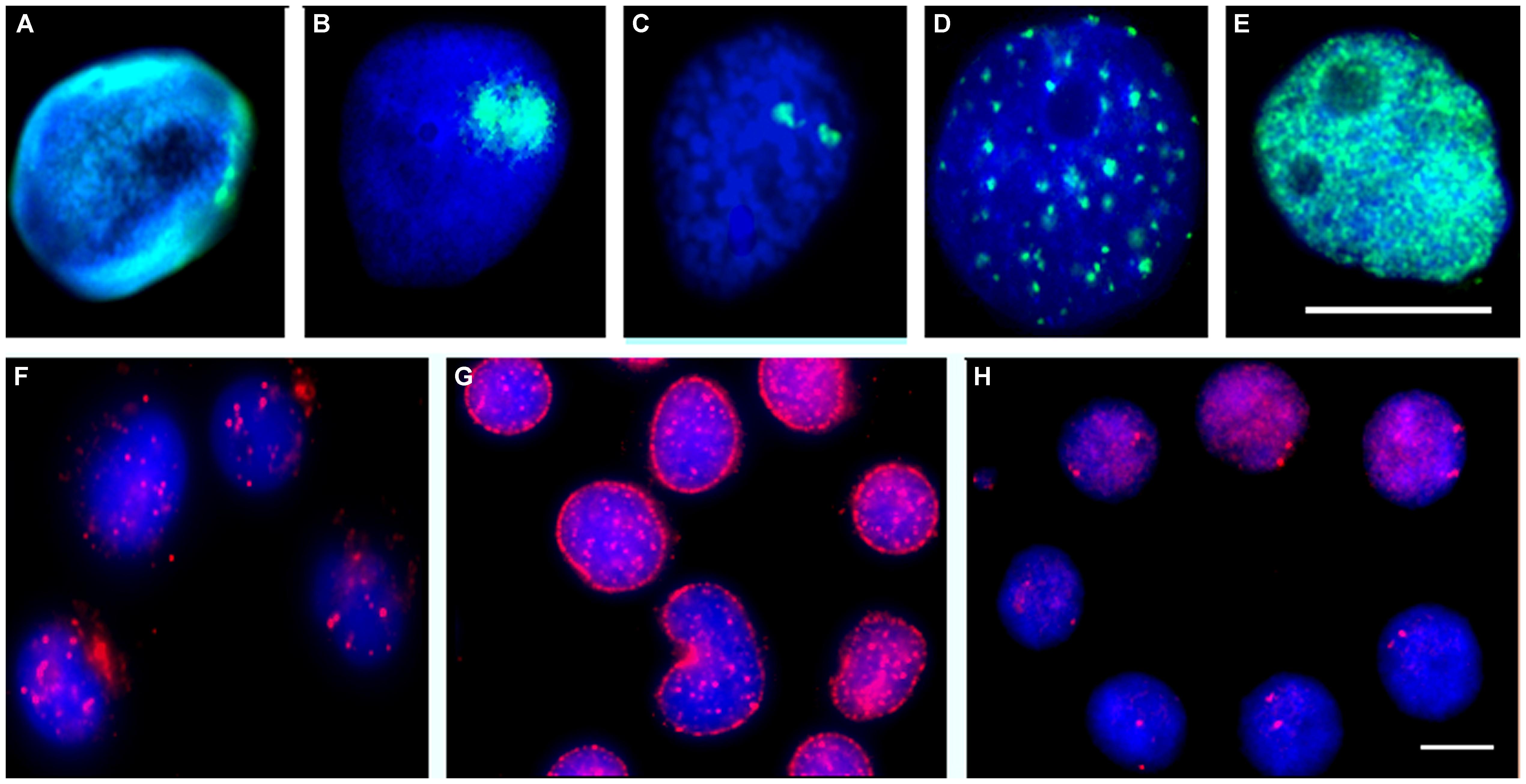
FIGURE 2. Biomphalaria glabrata embryonic (Bge) cells grown in vitro or ex vivo cells extracted from B. glabrata snails stained with antibodies or subjected to FISH. Antibodies raised against other species were used to reveal similar nuclear structures within snails and Bge cells; these include the nuclear lamina (A, green), fibrillarin in the nucleolus (B, green), PML bodies (F, red) and nuclear myosin (G, red). Chromatin modification was revealed using antibodies made against D. melanogaster modification that have a broad species range; histon 4 trimethylated K27 (D, green) and histone 3 trimethyl K4 (E, green). Individual chromosome territories were delineated by allowing 5-bromo-2′-deoxyuridine to be incorporated into replicating DNA and then allowing cells to divide seven times, followed by indirect immunofluorescence with an anti-5-bromo-2′-deoxyuridine antibody (C, red). Individual gene loci were revealed by FISH using labeled BAC probes (H, red). The DNA of the cells was stained with a DNA intercalating dye 4′6-diamidino-2-phenylindole. Scale bar 10 μm (Drs. Arican-Goktas, 2013 and Odoemelams′, 2009, Ph.D. thesis, Brunel University, UK) (Odoemelam, 2009; Arican-Goktas, 2013).
When originally analyzing the organization of the snail genome within cell nuclei it was hypothesized that it might be more similar to simpler organisms such as Drosophila melanogaster and Caenorhabditis elegans. Interestingly, it was found that the genome organization within the snail was similar to the mammalian genome. This was discovered by the preparation of a single chromosome painting probe that when used in fluorescence in situ hybridization (FISH) delineated individual chromosome territories (Figure 2C; Knight et al., 2009), these resembled territories found in higher organisms such as human, mouse (Foster et al., 2005) and pigs (Foster et al., 2012) and was also found to be non-randomly located within cell nuclei. Large and small chromosome territories were also revealed (Odoemelam et al., 2009), although it is as yet not possible to reveal gene density correlation with chromosome location since sequencing of the genome remains unfinished. However, the karyotype of B. glabrata has now been completed and several chromosomes have now been named and identified by utilizing bacterial artificial chromosomes (BAC) probes generated by the genome sequencing effort (Arican-Goktas et al., unpublished data). From these data, it is clear that B. glabrata chromosomes are not as previously described, i.e., being monomeric since they display differences in size, centromere position, and banding, implying there are gene-rich and gene-poor chromosome regions as well as gene-rich and -poor chromosomes. This advance will be very important for the genome sequencing effort as it will allow genes to be located to a specific chromosome, facilitating assembly, annotation, and organization of the sequence data. The protocol development for mapping onto B. glabrata chromosomes by FISH has taken a decade to refine and these methods will be published with the manuscript describing the genome.
The genome has some role in organizing itself within nuclei – through chromatin modification, i.e., the histone code (Figures 2D,E), but it also requires many different proteins within nuclei to help position and tether it. A concerted effort has been made to study B. glabrata cell nuclei to determine how similar they are to other organisms. As mentioned above, the nucleus of B. glabrata appears to be very similar to mammalian cell nuclei (Figure 2), containing many nuclear structures that were revealed using indirect immunofluorescence with antibodies reacting with human antigens, such as nuclear lamins (Figure 2A), nucleoli (Figure 2B), nuclear bodies (Figure 2F), nuclear motors (Figure 2G), transcription factories and splicing speckles, making this mollusk an interesting model to analyze genome behavior in since it has aspects of both simpler and more complex organisms.
Schistosomes Influence Repositioning of Gene Loci in Interphase Nuclear, Corresponding With Transcription
The complex relationship between the schistosome parasite and a given compatible species of snail developed over many 1000s of years, and is a co-evolved relationship.
Using labeled BAC probes and FISH analysis, gene loci position has been mapped in B. glabrata and found as small foci, very similar to other organisms (Figure 2H; Odoemelam et al., 2009). Using a bespoke computer analysis script (Croft, 1999), the positioning of these gene loci is found to be non-random with some genes being found at the nuclear periphery, some in the nuclear interior and some in an intermediate location as for other organisms (Szczerbal et al., 2009; Bourne et al., 2013). However, upon exposure to the parasite either in vitro with Bge cells (Knight et al., 2011) or in vivo in intact snails we found that specific genes change their nuclear location to a new non-random location correlating with their expression. For most genes attenuated parasite failed to elicit either a relocalization of gene loci, or change in expression. The in vivo studies have been most revealing since we were able to employ both susceptible and resistant snails. For gene loci corresponding to Hsp 70, we found overwhelming differences in behavior between the two snail lines. For example, in the susceptible NIMR snail, the gene loci for Hsp 70 moved to a new nuclear location before being expressed whereas no such movement and no gene expression for Hsp 70 were observed in the resistant BS-90 snail, corroborating previously reported differential gene expression data for Hsp 70 between resistant and susceptible snails (Knight et al., 2011). Additionally, when attenuated parasite was used to infect these snails, neither gene movement nor expression of Hsp 70 was observed. From these data, we believe the movement and expression of Hsp 70 is controlled by the parasite infection and may be used to help promote its survival/development in the snail (Arican-Goktas et al., 2014) parasite elicit a gene movement to a new nuclear location for gene expression to occur? We postulate that this requires remodeling of chromatin at specific genome locations and active directed movement to an area of active transcription in the cell nuclei using nuclear motor proteins such as nuclear myosin 1b (Figure 2G).
Conclusion
In the 20 years since a molecular approach to decipher mechanisms of the snail host/ schistosome interaction was undertaken – a relationship fashioned over centuries to give each organism a survival advantage over the other, we have discussed in this review how important advances have indeed been made. For example, the long held belief that innate defense molecules of invertebrates, unlike those of vertebrates (e.g., antibody) lack the ability to diversify in their structure has been decisively proven incorrect by the level of diversification revealed in B. glabrata FREP transcripts. Likewise, we now also know that schistosomes have evolved a mechanism that is capable of manipulating the genome of a compatible snail host, thereby orchestrating changes in the snail’s cell nuclei that alters gene expression in the parasite’s favor. While these important groundbreaking advances have been made, however, more work remains to be done. For example, we still do not know how schistosomes induce relocalization of gene loci to nuclear compartments for gene expression or silencing. These questions notwithstanding, it is clear that with the entire sequence of the genome soon becoming available, B. glabrata should fast become a new model system whereby the effect of a complex pathogen on genome behavior can be examined, enabling us to discover novel pathways that can be interfered with to disable snail transmission of schistosomiasis.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the NIH – NIAID for the grant award (R01AI63480) that funded the work. Halime D. Arican-Goktas was partially funded by the Malacological Society of London. We also thank Drs. Victoria Mann and Jacob Hodgson for their helpful critique of the manuscript.
References
Arican-Goktas, H. D. (2013). Parasitic Influences on the Host Genome Using the Molluscan Model Organism Biomphalaria glabrata. Ph.D. thesis, Doctor of Philosophy Biosciences, Brunel University, London.
Arican-Goktas, H. D., Ittiprasert, W., Bridger, J. M., and Knight, M. (2014). Differential spatio-epigenetic repositioning of activated genes in Biomphalaria glabrata resistant and susceptible snails infected with Schistosoma mansoni. PLoS Negl. Trop. Dis. (in press).
Bourne, G., Moir, C., Bikkul, U., Hassan Ahmed, M., Kill, I. R., Eskiw, C. H.,et al. (2013). Interphase chromosomes in diseases. Human interphase chromosomes: the Biomedical aspects. Springer (in press).
Chai, J. Y. (2013). Praziquantel treatment in trematode and cestode infections: an update. Infect. Chemother. 45, 32–43. doi: 10.3947/ic.2013.45.1.32
Croft, S. L. (1999). Pharmacological approaches to antitrypanosomal chemotherapy. Mem. Inst. Oswaldo Cruz 94, 215–220. doi: 10.1590/S0074-02761999000200017
De Almeida Marques, D. P., Rosa, F. M., Maciel, E., Negrao-Correa, D., Teles, H. M. S., Caldeira, R. L.,et al. (2014). Redueced susceptibility of a Biomphalaria tenagophila Population to Schistosoma mansoni after introducing the Taim/RS strain of B. tenagophila into Herivelton Martins stream. PLoS ONE 9:e99573. doi: 10.1371/journal.pone.0099573
Downs, J. A., Kabangila, R., Verweij, J. J., Jaka, H., Peck, R. N., Kalluvya, S. E.,et al. (2013). Detectable urogenital schistosome DNA and cervical abnormalities 6 months after single-dose praziquantel in women with Schistosoma haematobium infection. Trop. Med. Int. Health 18, 1090–1096. doi: 10.1111/tmi.12154
Fenwick, A., Webster, J. P., Bosque-Oliva, E., Blair, L., Fleming, F. M., Zhang, Y.,et al. (2009a). The Schistosomiasis Control Initiative (SCI): rationale, development and implementation from 2002-2008. Parasitology 136, 1719–1730. doi: 10.1017/S0031182009990400
Fenwick, A., Zhang, Y., and Stoever, K. (2009b). Control of the Neglected Tropical Diseases in sub-Saharan Africa: the unmet needs. Int. Health 1, 61–70. doi: 10.1016/j.inhe.2009.06.002
Ferguson, F. F., and Ruiz-Tiben, E. (1971). Review of biological control methods for schistosome-bearing snails. Ethiop. Med. J. 9, 95–104.
Foster, H. A., Abeydeera, L. R., Griffin, D. K., and Bridger, J. M. (2005). Non-random chromosome positioning in mammalian sperm nuclei, with migration of the sex chromosomes during late spermatogenesis. J. Cell Sci. 118, 1811–1820. doi: 10.1242/jcs.02301
Foster, H. A., Griffin, D. K., and Bridger, J. M. (2012). Interphase chromosome positioning in in vitro porcine cells and ex vivo porcine tissues. BMC Cell Biol. 13:30. doi: 10.1186/1471-2121-13-30
Garba, A., Toure, S., Dembele, R., Boisier, P., Tohon, Z., Bosque-Oliva, E.,et al. (2009). Present and future schistosomiasis control activities with support from the Schistosomiasis Control Initiative in West Africa. Parasitology 136, 1731–1737. doi: 10.1017/S0031182009990369
Hahn, U. K., Bender, R. C., and Bayne, C. J. (2001). Killing of Schistosoma mansoni sporocysts by hemocytes from resistant Biomphalaria glabrata: role of reactive oxygen species. J. Parasitol. 87, 292–299. doi: 10.1645/0022-3395(2001)087[0292:KOSMSB]2.0.CO;2
Hanington, P. C., Forys, M. A., Dragoo, J. W., Zhang, S. M., Adema, C. M., and Loker, E. S. (2010). Role for a somatically diversified lectin in resistance of an invertebrate to parasite infection. Proc. Natl. Acad. Sci. U.S.A. 107, 21087–21092. doi: 10.1073/pnas.1011242107
Hanington, P. C., Forys, M. A., and Loker, E. S. (2012). A somatically diversified defense factor, FREP3, is a determinant of snail resistance to schistosome infection. PLoS Negl. Trop. Dis. 6:e1591. doi: 10.1371/journal.pntd.0001591
Hotez, P. J., Bethony, J. M., Diemert, D. J., Pearson, M., and Loukas, A. (2010). Developing vaccines to combat hookworm infection and intestinal schistosomiasis. Nat. Rev. Microbiol. 8, 814–826. doi: 10.1038/nrmicro2438
Ittiprasert, W., and Knight, M. (2012). Reversing the resistance phenotype of the Biomphalaria glabrata snail host Schistosoma mansoni infection by temperature modulation. PLoS Pathog. 8:e1002677. doi: 10.1371/journal.ppat.1002677
Ittiprasert, W., Miller, A., Myers, J., Nene, V., El-Sayed, N. M., and Knight, M. (2010). Identification of immediate response genes dominantly expressed in juvenile resistant and susceptible Biomphalaria glabrata snails upon exposure to Schistosoma mansoni. Mol. Biochem. Parasitol. 169, 27–39. doi: 10.1016/j.molbiopara.2009.09.009
Ittiprasert, W., Nene, R., Miller, A., Raghavan, N., Lewis, F., Hodgson, J.,et al. (2009). Schistosoma mansoni infection of juvenile Biomphalaria glabrata induces a differential stress response between resistant and susceptible snails. Exp. Parasitol. 123, 203–211. doi: 10.1016/j.exppara.2009.07.015
Jobin, W. R., and Laracuente, A. (1979). Biological control of schistosome transmission in flowing water habitats. Am. J. Trop. Med. Hyg. 28, 916–917.
Kjetland, E. F., Kurewa, E. N., Mduluza, T., Midzi, N., Gomo, E., Friis, H.,et al. (2010). The first community-based report on the effect of genital Schistosoma haematobium infection on female fertility. Fertil. Steril. 94, 1551–1553. doi: 10.1016/j.fertnstert.2009.12.050
Knight, M., Brindley, P. J., Richards, C. S., and Lewis, F. A. (1991). Schistosoma mansoni: use of a cloned ribosomal RNA gene probe to detect restriction fragment length polymorphisms in the intermediate host Biomphalaria glabrata. Exp. Parasitol. 73, 285–294. doi: 10.1016/0014-4894(91)90100-B
Knight, M., Ittiprasert, W., Odoemelam, E. C., Adema, C. M., Miller, A., Raghavan, N.,et al. (2011). Non-random organization of the Biomphalaria glabrata genome in interphase Bge cells and the spatial repositioning of activated genes in cells co-cultured with Schistosoma mansoni. Int. J. Parasitol. 41, 61–70. doi: 10.1016/j.ijpara.2010.07.015
Knight, M., Ongele, E., and Lewis, F. A. (2000). Molecular studies of Biomphalaria glabrata, an intermediate host of Schistosoma mansoni. Int. J. Parasitol. 30, 535–541. doi: 10.1016/S0020-7519(99)00182-4
Knight, M., Raghavan, N., Goodall, C., Cousin, C., Ittiprasert, W., Sayed, A.,et al. (2009). Biomphalaria glabrata peroxiredoxin: effect of Schistosoma mansoni infection on differential gene regulation. Mol. Biochem. Parasitol. 167, 20–31. doi: 10.1016/j.molbiopara.2009.04.002
Lockyer, A. E., Emery, A. M., Kane, R. A., Walker, A. J., Mayer, C. D., Mitta, G.,et al. (2012). Early differential gene expression in haemocytes from resistant and susceptible Biomphalaria glabrata strains in response to Schistosoma mansoni. PLoS ONE 7:e51102. doi: 10.1371/journal.pone.0051102
Lockyer, A. E., Spinks, J., Kane, R. A., Hoffmann, K. F., Fitzpatrick, J. M., Rollinson, D.,et al. (2008). Biomphalaria glabrata transcriptome: cDNA microarray profiling identifies resistant- and susceptible-specific gene expression in haemocytes from snail strains exposed to Schistosoma mansoni. BMC Genomics 9:634. doi: 10.1186/1471-2164-9-634
Lockyer, A. E., Spinks, J. N., Walker, A. J., Kane, R. A., Noble, L. R., Rollinson, D.,et al. (2007). Biomphalaria glabrata transcriptome: identification of cell-signalling, transcriptional control and immune-related genes from open reading frame expressed sequence tags (ORESTES). Dev. Comp. Immunol. 31, 763–782. doi: 10.1016/j.dci.2006.11.004
Massoud, A., Metwally, D. M., Khalifa, K. E., and Habib, F. S. (2004). Compatibility of Biomphalaria alexandrina snails to infection with Schistosoma mansoni after exposure to sublethal concentrations of Myrrh. J. Egypt Soc. Parasitol. 34, 995–1008.
Mattos, A. C., Martins-Souza, R. L., Kusel, J. R., and Coelho, P. M. (2011). Interaction between primary and secondary sporocysts of Schistosoma mansoni and the internal defence system of Biomphalaria resistant and susceptible to the parasite. Mem. Inst. Oswaldo Cruz 106, 424–432. doi: 10.1590/S0074-02762011000400007
Meurs, L., Labuda, L., Amoah, A. S., Mbow, M., Ngoa, U. A., Boakye, D. A.,et al. (2011). Enhanced pro-inflammatory cytokine responses following Toll-like-receptor ligation in Schistosoma haematobium-infected schoolchildren from rural Gabon. PLoS ONE 6:e24393. doi: 10.1371/journal.pone.0024393
Mitta, G., Adema, C. M., Gourbal, B., Loker, E. S., and Theron, A. (2012). Compatibility polymorphism in snail/schistosome interactions: From field to theory to molecular mechanisms. Dev. Comp. Immunol. 37, 1–8. doi: 10.1016/j.dci.2011.09.002
Mitta, G., Galinier, R., Tisseyre, P., Allienne, J. F., Girerd-Chambaz, Y., Guillou, F.,et al. (2005). Gene discovery and expression analysis of immune-relevant genes from Biomphalaria glabrata hemocytes. Dev. Comp. Immunol. 29, 393–407. doi: 10.1016/j.dci.2004.10.002
Musuva, R. M., Awiti, A., Omedo, M., Ogutu, M., Secor, W. E., Montgomery, S. P.,et al. (2014). Community nowledge, attitudes and practices on Schistosomiasis in Western Kenya-The SCORE Project. Am. J. Trop. Med. Hyg. 90, 646–652. doi: 10.4269/ajtmh.13-0488
Odoemelam, E. (2009). Genomic Analysis of the Fresh Water Mollusc Biomphalaria glabrata to Understand Host-Parasite Interactions. Ph.D. thesis, Doctor of Philosophy Biosciences, Brunel University, London.
Odoemelam, E., Raghavan, N., Miller, A., Bridger, J. M., and Knight, M. (2009). Revised karyotyping and gene mapping of the Biomphalaria glabrata embryonic (Bge) cell line. Int. J. Parasitol. 39, 675–681. doi: 10.1016/j.ijpara.2008.11.011
Oliveira, A. L., Da Silva, D., Zanotti-Magalhaes, E. M., Abdel-Hamid, A. Z., and Ribeiro-Paes, J. T. (2008). Schistosome/mollusk: genetic compatibility. Genet. Mol. Res. 7, 518–526. doi: 10.4238/vol7-2gmr444
Olveda, D. U., Li, Y., Olveda, R. M., Lam, A. K., Mcmanus, D. P., Chau, T. N.,et al. (2014). Bilharzia in the Philippines: past, present, and future. Int. J. Infect. Dis. 18, 52–56. doi: 10.1016/j.ijid.2013.09.011
Owusu-Bempah, A., Odoi, A. T., and Dassah, E. T. (2013). Genital schistosomiasis leading to ectopic pregnancy and subfertility: a case for parasitic evaluation of gynaecologic patients in schistosomiasis endemic areas. Case Rep. Obstet. Gynecol. 2013, 634264. doi: 10.1155/2013/634264
Paveley, R. A., Aynsley, S. A., Turner, J. D., Bourke, C. D., Jenkins, S. J., Cook, P. C.,et al. (2011). The Mannose Receptor (CD206) is an important pattern recognition receptor (PRR) in the detection of the infective stage of the helminth Schistosoma mansoni and modulates IFNgamma production. Int. J. Parasitol. 41, 1335–1345. doi: 10.1016/j.ijpara.2011.08.005
Pointier, J. P. (1993). The introduction of Melanoides tuberculata (Mollusca: Thiaridae) to the island of Saint Lucia (West Indies) and its role in the decline of Biomphalaria glabrata, the snail intermediate host of Schistosoma mansoni. Acta Trop. 54, 13–18. doi: 10.1016/0001-706X(93)90064-I
Portela, J., Duval, D., Rognon, A., Galinier, R., Boissier, J., Coustau, C.,et al. (2013). Evidence for specific genotype-dependent immune priming in the lophotrochozoan Biomphalaria glabrata snail. J. Innate. Immun. 5, 261–276. doi: 10.1159/000345909
Raghavan, N., Miller, A. N., Gardner, M., Fitzgerald, P. C., Kerlavage, A. R., Johnston, D. A.,et al. (2003). Comparative gene analysis of Biomphalaria glabrata hemocytes pre- and post-exposure to miracidia of Schistosoma mansoni. Mol. Biochem. Parasitol. 126, 181–191. doi: 10.1016/S0166-6851(02)00272-4
Richards, C. S., Knight, M., and Lewis, F. A. (1992). Genetics of Biomphalaria glabrata and its effect on the outcome of Schistosoma mansoni infection. Parasitol. Today 8, 171–174. doi: 10.1016/0169-4758(92)90015-T
Roger, E., Grunau, C., Pierce, R. J., Hirai, H., Gourbal, B., Galinier, R.,et al. (2008a). Controlled chaos of polymorphic mucins in a metazoan parasite (Schistosoma mansoni) interacting with its invertebrate host (Biomphalaria glabrata). PLoS Negl. Trop. Dis. 2:e330. doi: 10.1371/journal.pntd.0000330
Roger, E., Mitta, G., Mone, Y., Bouchut, A., Rognon, A., Grunau, C.,et al. (2008b). Molecular determinants of compatibility polymorphism in the Biomphalaria glabrata/Schistosoma mansoni model: new candidates identified by a global comparative proteomics approach. Mol. Biochem. Parasitol. 157, 205–216. doi: 10.1016/j.molbiopara.2007.11.003
Rollinson, D., Knopp, S., Levitz, S., Stothard, J. R., Tchuem Tchuente, L. A., Garba, A.,et al. (2013). Time to set the agenda for schistosomiasis elimination. Acta Trop. 128, 423–440. doi: 10.1016/j.actatropica.2012.04.013
Ruiz-Tiben, E., Palmer, J. R., and Ferguson, F. (1969). Biological control of Biomphalaria glabrata by Marisa cornuarietis in irrigation ponds in Puerto Rico. Bull. World Health Organ. 41, 329–333.
Sahu, L., Tempe, A., Singh, S., and Khurana, N. (2013). Ruptured ectopic pregnancy associated with tubal schistosomiasis. J. Postgrad. Med. 59, 315–317. doi: 10.4103/0022-3859.123166
Souza Sdos, S., and Andrade, Z. A. (2006). On the origin of the Biomphalaria glabrata hemocytes. Mem. Inst. Oswaldo Cruz 101(Suppl. 1), 213–218. doi: 10.1590/S0074-02762006000900033
Souza Sdos, S., and Andrade, Z. A. (2012). The significance of the amoebocyte-producing organ in Biomphalaria glabrata. Mem. Inst. Oswaldo Cruz 107, 598–603. doi: 10.1590/S0074-02762012000500005
Spada, R. G., Da Silva, D., Abdel-Hamid, A. Z., Sobral-Hamaguchi, S. S., Zuim, N. R., Zanotti-Magalhaes, E. M.,et al. (2002). Genetic markers between Biomphalaria glabrata snails susceptible and resistant to Schistosoma mansoni infection. Mem. Inst. Oswaldo Cruz 97(Suppl. 1), 53–58. doi: 10.1590/S0074-02762002000900012
Szczerbal, I., Foster, H. A., and Bridger, J. M. (2009). The spatial repositioning of adipogenesis genes is correlated with their expression status in a porcine mesenchymal stem cell adipogenesis model system. Chromosoma 118, 647–663. doi: 10.1007/s00412-009-0225-5
Tchuem Tchuente, L. A., Dongmo Noumedem, C., Ngassam, P., Kenfack, C. M., Feussom Gipwe, N., Dankoni, E.,et al. (2013). Mapping of schistosomiasis and soil-transmitted helminthiasis in the regions of Littoral, North-West, South and South-West Cameroon and recommendations for treatment. BMC Infect. Dis. 13:602. doi: 10.1186/1471-2334-13–602
Theron, A., and Coustau, C. (2005). Are Biomphalaria snails resistant to Schistosoma mansoni? J. Helminthol. 79, 187–191. doi: 10.1079/JOH2005299
Vasquez, R. E., and Sullivan, J. T. (2001). Further characterization of passively transferred resistance to Schistosoma mansoni in the snail intermediate host Biomphalaria glabrata. J. Parasitol. 87, 1360–1365. doi: 10.1645/0022-3395(2001)087[1360:FCOPTR]2.0.CO;2
Yoshino, T. P., and Bayne, C. J. (1983). Mimicry of snail host antigens by miracidia and primary sporocysts of Schistosoma mansoni. Parasite Immunol. 5, 317–328. doi: 10.1111/j.1365-3024.1983.tb00747.x
Yoshino, T. P., Wu, X. J., Gonzalez, L. A., and Hokke, C. H. (2013). Circulating Biomphalaria glabrata hemocyte subpopulations possess shared schistosome glycans and receptors capable of binding larval glycoconjugates. Exp. Parasitol. 133, 28–36. doi: 10.1016/j.exppara.2012.10.002
Zahoor, Z., Davies, A. J., Kirk, R. S., Rollinson, D., and Walker, A. J. (2009). Nitric oxide production by Biomphalaria glabrata haemocytes: effects of Schistosoma mansoni ESPs and regulation through the extracellular signal-regulated kinase pathway. Parasit. Vectors 2, 18. doi: 10.1186/1756-3305-2-18
Zahoor, Z., Lockyer, A. E., Davies, A. J., Kirk, R. S., Emery, A. M., Rollinson, D.,et al. (2014). Differences in the gene expression profiles of haemocytes from schistosome-susceptible and -resistant Biomphalaria glabrata exposed to Schistosoma mansoni excretory-secretory products. PLoS ONE 9:e93215. doi: 10.1371/journal.pone.0093215
Zhang, S. M., and Loker, E. S. (2004). Representation of an immune responsive gene family encoding fibrinogen-related proteins in the freshwater mollusc Biomphalaria glabrata, an intermediate host for Schistosoma mansoni. Gene 341, 255–266. doi: 10.1016/j.gene.2004.07.003
Keywords: intermediate snail host, B. glabrata, S. mansoni, resistance, susceptibility, compatibility, gene-expression, gene loci re-localization
Citation: Knight M, Arican-Goktas HD, Ittiprasert W, Odoemelam EC, Miller AN and Bridger JM (2014) Schistosomes and snails: a molecular encounter. Front. Genet. 5:230. doi: 10.3389/fgene.2014.00230
Received: 24 April 2014; Accepted: 30 June 2014;
Published online: 21 July 2014.
Edited by:
Arnon Dias Jurberg, Oswaldo Cruz Institute – Oswaldo Cruz Foundation, BrazilReviewed by:
Anthony John Walker, Kingston University, UKPaulo Marcos Zech Coelho, Fundação Oswaldo Cruz, Brazil
Copyright © 2014 Knight, Arican-Goktas, Ittiprasert, Odoemelam, Miller and Bridger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matty Knight, Department of Microbiology, Immunology and Tropical Medicine, The George Washington University, Ross Hall, 2300 Eye Street, NW, Suite 502, Washington, DC 20037, USA e-mail: matty_knight@gwu.edu