Corrigendum: Quantity and Quality of Inhaled Dose Predicts Immunopathology in Tuberculosis
 Kevin P. Fennelly
Kevin P. Fennelly Edward C. Jones-López
Edward C. Jones-López- 1Department of Medicine, Emerging Pathogens Institute, University of Florida, Gainesville, FL, USA
- 2Section of Infectious Diseases, Boston Medical Center, Boston University School of Medicine, Boston, MA, USA
Experimental animal models of tuberculosis (TB) have convincingly demonstrated that inhaled dose predicts immunopathology and survival. In contrast, the importance of inhaled dose has generally not been appreciated in TB epidemiology, clinical science, or the practice of TB control. Infectiousness of TB patients has traditionally been assessed using microscopy for acid-fast bacilli in the sputum, which should be considered only a risk factor. We have recently demonstrated that cough aerosol cultures from index cases with pulmonary TB are the best predictors of new infection among household contacts. We suggest that cough aerosols of M. tuberculosis are the best surrogates of inhaled dose, and we hypothesize that the quantity of cough aerosols is associated with TB infection versus disease. Although several factors affect the quality of infectious aerosols, we propose that the particle size distribution of cough aerosols is an important predictor of primary upper airway disease and cervical lymphadenitis and of immune responses in exposed hosts. We hypothesize that large droplet aerosols (>5 μ) containing M. tuberculosis deposit in the upper airway and can induce immune responses without establishing infection. We suggest that this may partially explain the large proportion of humans who never develop TB disease in spite of having immunological evidence of M. tuberculosis infection (e.g., positive tuberculin skin test or interferon gamma release assay). If these hypotheses are proven true, they would alter the current paradigm of latent TB infection and reactivation, further demonstrating the need for better biomarkers or methods of assessing TB infection and the risk of developing disease.
Introduction
In spite of the devastating impact of tuberculosis (TB) on mankind throughout history, scientists have struggled to decipher the determinants of the wide spectrum of infection and disease in TB. Data from over 50 years ago have suggested that the vast majority (90%) of persons infected with the causative agent, Mycobacterium tuberculosis, never manifest any symptoms or signs of disease. These persons are diagnosed with latent TB infection (LTBI) only by an immunological signature of infection using the traditional tuberculin skin test (TST) or a more modern interferon gamma release assay (IGRA) of the blood. It is estimated that one-third of the global population, or 2 billion people, are latently infected with TB. At the opposite end of the spectrum are persons who die of respiratory failure due to destruction of their lungs or who present with TB sepsis due to dissemination of M. tuberculosis in their bloodstream and to other organs. The variable balance between the immune response and virulence of the bacilli is responsible for the containment of M. tuberculosis in LTBI, for the destruction of lungs and other tissues due to an overly aggressive response, and for disseminated disease due to loss of control of M. tuberculosis. The actions of the immune system responsible for this variability are highly complex and are addressed by other authors in this issue. However, we present a simple concept that is poorly appreciated in both research and control of human TB: a major determinant of immunopathology in TB is the inhaled dose of M. tuberculosis. Our primary hypothesis is that small inhaled doses are more likely to result in LTBI, and that large inhaled doses are more likely to result in primary TB disease. In other words, we hypothesize that the quantity of inhaled dose predicts the immunopathology following infection. We propose that cough aerosol cultures of M. tuberculosis from human patients with active TB cases are better surrogates of inhaled dose than are sputa smears for acid-fast bacilli (AFB) or sputa cultures of M. tuberculosis.
We also propose that the quality of the infectious aerosol is an important determinant of immunopathology. The quality of an infectious aerosol includes its particle size distribution, virulence and infectivity of the pathogen, the duration of exposure to antimycobacterial drugs, desiccation, ultraviolet light, and other stressors. We will focus our discussion on the particle size distribution. We hypothesize that when M. tuberculosis is present in large droplet aerosols it deposits in the upper airways and produces immune responses but not necessarily infection. This would explain a large number of persons who are currently diagnosed as having LTBI but who never develop active TB disease. This idea also addresses the fundamental question of whether all those with positive TSTs or IGRAs are infected with viable M. tuberculosis (1). These hypotheses are based on our recent data on cough aerosols of M. tuberculosis; a new synthesis and interpretation of the literature on TB epidemiology, pathogenesis and clinical presentations; and principles of aerosol science. These concepts challenge current dogma in our understanding of TB, but we respectfully suggest that they help explain uncertainties in the science of TB and that they can be readily tested in the near future. If proven true, they would change the current paradigm of LTBI diagnosis and treatment. They would also emphasize the need for better biomarkers to confirm the presence of live bacilli within the human host and to predict the risk of reactivation of LTBI.
Hypothesis 1. The magnitude of cough aerosol cultures in fine (≤5 μ) aerosols of M. tuberculosis from index cases of pulmonary TB is positively associated with TB disease (Figure 1).
(a) Large magnitudes of cough aerosol cultures are associated with primary TB.
(b) Small magnitudes of cough aerosol cultures are associated with reactivation TB within the first few years of exposure.
FIGURE 1
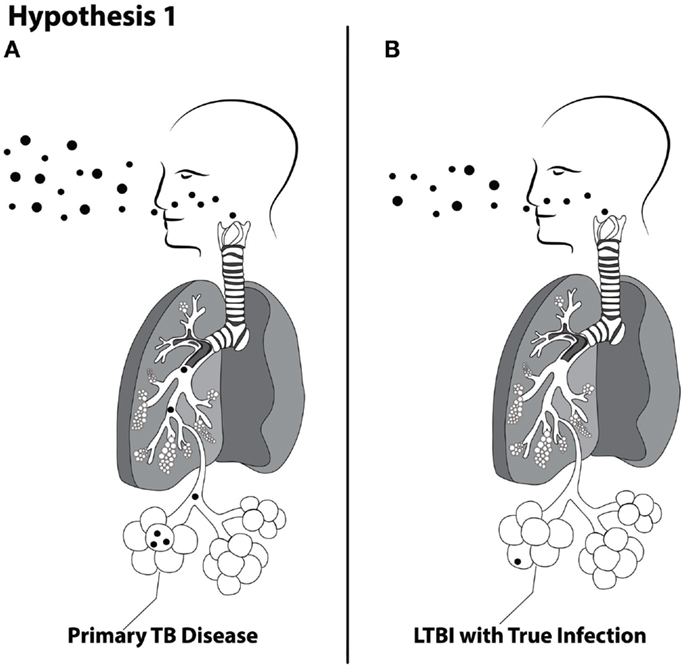
Figure 1. Schematic of Hypothesis 1 demonstrating the proposed concept that larger inhaled doses are more likely to produce primary TB disease (A) and lower doses are more likely to produce latent TB infection (B).
Inhaled Dose–Response in Animal Models of TB
General Principle of the Inoculum Effect
A general principle in the study of infectious diseases is that the size of the inoculum, or dose, and virulence of a given pathogen predict the establishment of infection and subsequent pathogenesis in the host (2). This principle has been demonstrated in bacteria (3–5), fungi (6, 7), viruses (8–10), and parasites (11). For example, classic experiments in which human volunteers ingested typhoid bacilli demonstrated the relationship between dose and the pathogenesis of typhoid fever (12, 13). However, such studies cannot ethically be done to understand the effect of inhaled dose and TB pathogenesis in humans. Our understanding of the effects of inhaled dose on the pathogenesis of TB is based largely on experimental work.
Brief Review of the History of the Science of Transmission of M. tuberculosis
Robert Koch conclusively demonstrated that M. tuberculosis is the etiologic agent of TB in 1882, but it was not until 1959 that Richard Riley and William Firth Wells and colleagues first demonstrated airborne transmission of M. tuberculosis to guinea pigs exposed to the air from humans with pulmonary TB in an experimental TB ward. Since then, TB has been considered the prototype disease of airborne transmission with a general acceptance that the other modes of transmission, i.e., large droplet and contact, do not occur. However, this is not absolute as there are reports indicating that the other modes of transmission rarely occur. Children can develop cutaneous TB in the form of chancres or warty TB from direct inoculation of bacilli onto damaged skin (14). A rare case of contact transmission occurred when a physician performed mouth-to-mouth resuscitation on a TB patient and acquired primary cutaneous TB in the nasolabial area (15). Also, direct injection of M. tuberculosis among pathologists and other laboratory workers can produce “prosectors warts,” i.e., granulomas at the site of accidental injection, first described by Laennec regarding a lesion on his own finger (16). Under our second hypothesis section later in this paper, we review the evidence for upper airway deposition of M. tuberculosis by large droplet aerosols.
Koch was also the first to demonstrate a dose–response effect of infection on pathogenesis (17). His observation of the association of higher mortality with increasing inoculum size was later confirmed by Glover (18) and Youmans (19). Glover’s semi-quantitative estimate of inhaled dose was also associated with lung pathology findings, ranging from small granulomatous lesions in the mice exposed to the lowest doses of M. bovis to gross suppurative consolidation in those exposed to the highest doses of M. bovis (Figure 2).
FIGURE 2
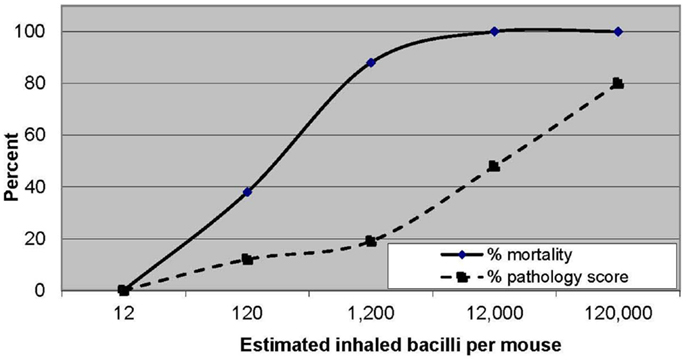
Figure 2. Increasing mortality and pathological changes in mice with increasing inhaled doses of Mycobacterium bovis. Data from Ref. (18).
Experimental aerogenic infections of the mouse, guinea pig, and rabbit models (20, 21) have demonstrated similarities in the kinetics of immunopathogenesis after infection. After infection in a given host that is not highly susceptible, growth occurs in the lung at a logarithmic rate for the first 2–3 weeks, and then plateaus at approximately 4 weeks when the infection is controlled by acquired immunity. In an immunologically naïve animal, more extensive replication of the bacteria is needed to induce the immune response, so the time course may be longer (approximately 3–4 weeks) than in the animal with pre-existing immunity (21). The genetic susceptibility of the animal determines the immune response to a given bacillary strain. For example, in Lurie’s studies of rabbits of varying susceptibility to infection, one resistant strain produced one grossly visible pulmonary tubercle per 600 viable H37Rv bacilli inhaled, whereas a susceptible strain developed one tubercle per 107 bacilli (20). However, for a given strain of host rabbit and a given strain of tubercle bacillus, intensity of airborne dose was associated with an increased number or rabbits infected and with shorter survival time. Similarly, natural resistance to TB in the rabbit model can be overcome by larger inhaled doses of tubercle bacilli (22), suggesting a strong dose-response relationship between inoculum size and risk of disease.
More recent studies using the same strain of mouse (C57BL/6) exposed to aerosols of three different strains of M. tuberculosis demonstrated that there was more rapid progression of disease associated with increasing bacillary concentrations in the aerosol within each strain, although a similar rapid progression of disease was observed when comparing more to less virulent strains. In other words, high inhaled doses of less virulent strains may produce similar lung pathology as lower doses of less virulent strains (23). In addition to the course of TB disease, the inhaled inoculum size has recently been shown to influence the therapeutic response to antituberculous medications (24).
These animal models all reflect acute infection, or what in humans is termed “primary TB disease” or “progressive primary disease.” This occurs most commonly in children <5 years of age (25) or in HIV infected persons (26). However, in one large study of 20,687 household contacts (HHCs) age 5 years or older), 598 (4.9%) had primary TB (25).
Models of LTBI
LTBI has been difficult to study due to the lack of good animal models until recently. The “Cornell Model” of LTBI involved the establishment of active TB disease that was then partially treated with antituberculous agents (27). The mice were then immunosuppressed to reactive TB disease in an effort to model the reactivation of TB in humans. Although some humans do reactivate LTBI due to immunosuppression, the vast majority of humans reactivate spontaneously without the medical intervention of immunosuppressive drugs. Why the majority of humans have LTBI but never reactivate has never been explained.
Non-human primate models
Although the experimental mouse model has provided valuable contributions to our understanding of the immunology of TB, it does not sufficiently model key aspects of human TB disease, notably necrotic lesions or cavitation or LTBI. Also, whereas the guinea pig and rabbit models have pathology more similar to humans, there are few immunologic reagents for these models. Some of the most exciting data from animal models in recent years has been generated using non-human primates (NHP) because of the similarity of TB disease to human disease and because of cross-reactivity of immunologic and molecular reagents with humans.
Early experiments used intratracheal administration of M. tuberculosis to Philippine cynomolgus monkeys (Macaca fascicularis) (28). Delivery of very high doses of 104 or 105 colony-forming units (CFU) of M. tuberculosis resulted in acute and rapidly progressive and fatal multilobar pneumonia. However, lower doses (<103 CFU) produced a more chronic and slowly progressive form of pulmonary TB that was similar to human disease sometimes with dissemination to other organs. Even lower doses (101 or 102 CFU) resulted in some monkeys containing the infection in a subclinical state.
Another group of investigators has used bronchoscopic administration of M. tuberculosis suspensions to infect 17 cynomolgus macaques with a low dose of approximately 25 CFU (29). They observed a range of pathology, with 7 classified as LTBI and eight remained alive and well, without evidence of disease after 15–21 months of observation. The other animals exhibited a range of pathology similar to primary human TB, and one animal may have reactivated with a small cavitary lesion.
Natural human infection with M. tuberculosis most often occurs after inhalation of infectious aerosols and yet another group of investigators have demonstrated that macaques infected by the head-only aerosol route develop disease that appears to be very similar to human TB. High doses of aerosols of M. tuberculosis produce a fulminant fatal pneumonic TB in rhesus macaques (30, 31). Lower doses of about 500 CFU of CDC 1551 strain produced asymptomatic, latent infections in 12 animals except for one with signs of active TB over 9 weeks of observation (32). All 11 appearing to have LTBI had no evidence of M. tuberculosis on bronchoalveolar lavage (BAL) at 9 weeks. Six animals were infected intravenously with simian immunodeficiency virus (SIV), and they all developed culture-positive BALs, and some had evidence of dissemination with positive blood cultures. The resurgence of the NHP model of TB is in part due to the availability of immunologic reagents for the study of AIDS, and reactivation of TB infection produced by either bronchoscopic instillation or by aerosol exposures, and has been demonstrated due to co-infection of macaques with SIV (32, 33). The NHP model has also demonstrated reactivation of LTBI with inhibition of tumor necrosis factor (TNF) alpha similar to that in humans (34).
In summary, the NHP model of TB has demonstrated the importance of inhaled (or instilled) dose in the establishment of LTBI versus primary (progressive) TB disease. What is not known is whether dose is a predictor of which patients with LTBI will reactivate in the future. We propose the concept that exposure of humans to large concentrations of infectious aerosol results in both (1) an increased risk of inhalation of live bacilli that can establish a focus of infection and (2) increased opportunities for multiple episodes of inhalation of one or more bacilli that may establish multiple foci of infection. Given recent evidence about the dynamic nature of the establishment of infection in a NHP model (35), this would increase the chances that one of those foci would result in a progressive infection acutely, as well as reactivation of a focus of infection at a later time point.
Epidemiological Evidence Supporting a Dose–Response Relationship
Epidemiologic studies have also suggested a dose–response relationship in TB immunopathogenesis. Epidemiologic studies of TB have been hampered by the inability to measure inhaled dose, or at least a measure of source strength from the index case with TB. This resulted in the use of the sputum smear as an indirect measure of bacillary load in the infectious aerosols generated by TB patients.
Household Contact Studies
Our understanding of transmission of M. tuberculosis to humans has been largely determined by epidemiological studies of HHCs of patients with pulmonary TB and by reports of nosocomial outbreaks. Close contacts of persons with sputum-smear-positive TB have the highest prevalence of LTBI compared to other populations, and they are second only to HIV infection for the highest risk of progressing from LTBI to active disease (36, 37). Increased transmission has been consistently associated with sputum smear-positive cases compared to that of sputum smear-negative cases. This has been most well-documented by comparing TSTs in HHCs who are exposed to patients with sputa that is smear-positive for AFB versus those with sputa that is smear-negative for AFB but culture-positive for M. tuberculosis. For example, in a classic study done in Canada, there was a gradient of higher rates of infection and secondary TB disease cases among HHCs exposed to sputum smear-positive cases, HHCs compared to smear-negative cases, and community controls (38). Among close contacts who developed active TB, 92% were exposed to a smear-positive case and only 8% to a smear-negative case. In another large study done in Chennai, India, the 15-year risk of progression to disease was highest among HHCs of sputum smear-positive/culture-positive cases (adjusted RR = 3.4; 95% CI 3.0–3.9) compared to contacts of sputum smear-negative/culture-positive cases (aRR 1.7; 1.4–2.0) and community controls without household exposure to TB (39). In a more recent study conducted in Galicia, Spain during 1996–2011 (40, 41), there were 3822 cases of smear-negative TB, most of them identified in contact tracing. The frequency of LTBI in HHC of these patients was similar to contacts of smear-positive cases (30.1 versus 33.4%). By contrast, the frequency of TB disease in HHC of smear-negative cases was <20% that of smear-positive cases.
The observed effect of AFB smear-positivity is most likely a simple quantitative difference in the concentration of bacilli in the respiratory secretions of the source cases, as approximately 5,000– 10,000 bacilli per milliliter must be present for detection by direct microscopy with Ziehl–Neelsen staining (42).
The percentage of TST-positive contacts of sputum smear-positive cases has varied from 1.1 to 73%, and that of contacts of smear-negative cases has ranged from 0 to 46%, with community rates from <1 to 41%. In a community-based study in England between 1948 and 52, positive TSTs were found in 65.2% of children exposed to a smear-positive case and 26.8% exposed to a smear-negative case (43). A strength of this study was that cases of active TB disease were also tracked. Positive TSTs were found in 14.4% of children exposed to a smear-positive case and in 3% of children exposed to a smear-negative case. However, interpretation of these data must include the background or community prevalence of a positive TST, and these data may vary widely (44). Although the TST has been a useful tool, it has many limitations, not the least of which is the inability to detect when infection occurred (44, 45).
Furthermore, epidemiological data suggest that there is marked variability of transmission from sputum-based studies (46). For example, in a Dutch study of contacts to smear-positive index TB cases in five clinics in Rotterdam from 1967 to 69, only 22 of 80 (27.5%) index cases were found to have infected their contacts. However, when the distribution was stratified by age, 57% of index cases under 40 years of age infected their contacts but only 9% of the index cases over 60 did so (47). Our preliminary data from Florida also suggest that <30% of sputum smear-positive index cases transmit to HHCs (see below), consistent with these data. Moreover, approximately 15% of transmission in three different populations has been associated with sputum smear-negative disease (46).
Intensity of Exposure and TB Outcomes
Intensity of exposure can be considered a surrogate measure of inhaled dose due to the likelihood of larger and more frequent exposures to infectious aerosols. Probably the best data regarding the effect of intensity of exposure to M. tuberculosis on TB infection and disease were the British Prophit Study conducted between 1933 and 1944, a time when TB was highly prevalent in the UK (48). As the data in Figure 3 demonstrate, 80% of the nurses in the highest exposure group demonstrated conversions of their TSTs compared to 26% in community controls. The nurse cohorts were more similar to each other and, therefore, more comparable than the community controls and medical student. Compared to nurses working in low exposure settings, those working with TB patients in high exposure settings had a 48% increased risk of TST conversion and a 41% increased risk of active TB.
FIGURE 3
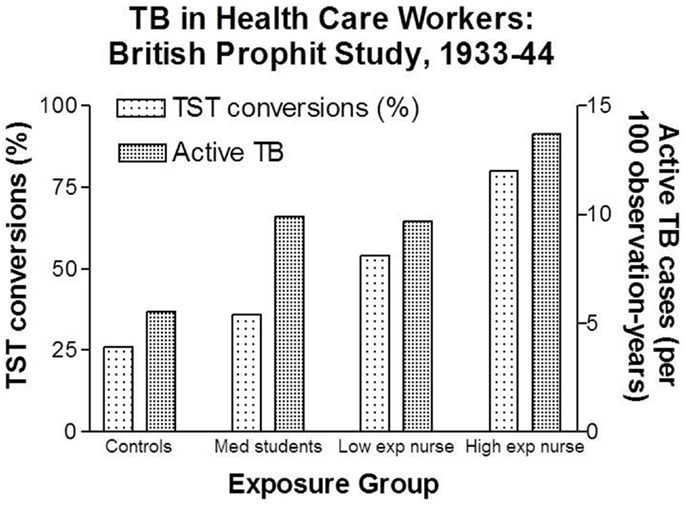
Figure 3. Increasing rates of latent TB infection and active TB among health care workers with increasing degrees of estimated exposure to airborne M. tuberculosis. Data from Ref. (48).
Proximity of Exposure and TB Outcomes
Proximity can also be considered a surrogate for dose, as concentrations of airborne hazards are higher the closer a person is to an infectious source. The association of proximity of exposure and TB infection has been demonstrated in various settings. HHCs sharing the same bed are at higher risk of infection than those living in another room in the same residence (49–51). Patients exposed to the infectious aerosols generated by a forceful irrigation of a tuberculous abscess were found to be infected along a clear spatial gradient, with more patients infected with TB the closer their rooms were along the hospital hallway (52). Similarly, airline passengers on a transoceanic flight were more likely to have evidence of TB infection the closer they were sitting to an infectious TB patient (53). In a classic outbreak of TB aboard a U.S. Navy ship, proximity and duration of contact were associated with both TB infection and disease (54, 55). In an unusual exposure to aerosols generated by water jet irrigation of a tuberculous abscess, there was a clear gradient of infection associated with proximity (52). A limitation of the epidemiological data on proximity of exposure is that it is not possible to determine if there were simply more frequent opportunities to become infected with similar inhaled doses or if there were greater chances of being exposed to larger concentrations of M. tuberculosis aerosols. Without a better surrogate measure for inhaled dose, such questions cannot be answered.
Transmission of M. tuberculosis and Cough
In addition to the bacillary load in the sputum, another known risk factor for transmission is the frequency of cough. In Oslo from 1940 to 53, the presence of cough in TB patients with smear-positive sputa increased the risk of infections in exposed children <5 years old (19.7 versus 5.5% in those without cough) (56). IN another study, Loudon recorded 8-h overnight cough frequency in newly admitted TB patients. Positive TSTs were found in 44% of contacts of patients who had more than 48 coughs per 8 h period compared to 28% of contacts of patients who had <12 coughs per night (57). In a more recent study in Brazil, we found that cough assessed by a visual analog scale or the Leicester Cough Questionnaire was associated with more transmission (58). Epidemiological studies have found that more secondary cases of TB are associated with men than with women, and that they decrease with advancing age (59). One explanation for this finding is that young patients tend to have more close contacts and more contacts in general than do older patients. However, we suggest that another explanation may be that younger persons and men may have higher forced expiratory volumes and thus be able to generate more cough aerosols, as we observed among patients with cystic fibrosis (CF) (60). Our preliminary data discussed below support the importance of cough in generating infectious aerosols of M. tuberculosis.
Cough Aerosol Cultures as a Surrogate for Inhaled Dose
While the inhaled dose can be manipulated in experimental animal models or in vitro systems, it is not possible to measure in human infections. It is obviously unethical to deliberately expose humans to varying concentrations of infectious aerosols of M. tuberculosis, as has been done with less virulent diseases. So any attempts to estimate inhaled dose must be observational and retrospective in nature, as human patients present either after they have become ill with active TB or after they have become exposed to someone with active TB. Thus, we must use markers that are surrogates for the inhaled dose.
Measurement of M. tuberculosis in Cough-Generated Aerosols
Although aerosol transmission of TB was convincingly established over 50 years ago, microscopy of sputum smears for AFB continues to be used as the best laboratory test of infectiousness due to the lack of better options. As discussed, whereas the sputum AFB smear has been a useful surrogate marker of inhaled dose for over 100 years, it clearly has its limitations and should only be considered a risk factor for infectiousness (61, 62). There is a need and capacity for a better surrogate marker of infectiousness.
During the resurgence of TB in the U.S. in the late 1980s and early 1990s, significant morbidity and mortality among health care workers (HCWs) was recognized. The use of personal respiratory protection (PRP, e.g., N95 respirators) was especially controversial, and the efficacy of PRP depends upon the particle size distribution of the inhalation hazard, in this case that of infectious aerosols containing M. tuberculosis. In addition, the size distribution of infectious aerosols determines how long they will remain airborne and where in the respiratory tract they are most likely to deposit. Although Wells had estimated the size range of infectious droplet nuclei to be 1–5 microns based on experimental nebulization in the laboratory and theoretical principles, the size of infectious aerosols of M. tuberculosis had never been directly measured. To address this need, we developed a “cough aerosol sampling system (CASS)” and initially aimed to study cough aerosols from patients with TB to determine the magnitude and particle size distribution of infectious aerosols of M. tuberculosis to improve the scientific basis for interventions to protect HCWs.
In our initial feasibility study using CASS in 16 AFB-positive patients at a referral hospital in the U.S., we collected culturable aerosols from only 4 (25%) of them (63). Aside from the small sample size and preliminary nature of that work, one criticism was that the sampling was done too close in proximity to the patient, and thus may not represent what infects human patients (64). However, this is probably not the case, as airborne droplets desiccate to the size of infectious droplet nuclei in <1 s (65, 66). Others more informally suggested that the method was too highly technical to be done in high incidence settings such as sub-Saharan Africa. We subsequently used the CASS to collect cough aerosols from sputum AFB-smear-positive TB patients in Kampala, Uganda (67). We cultured M. tuberculosis from the cough aerosols of 28 of 101 (27.7%; 95% confidence interval [CI], 19.9–37.1%) patients with culture-confirmed TB, with a median 16 CFU (range, 1–701) in 10 min of coughing. Nearly all (96.4%) cultivable particles were 0.65–4.7 μ in size, in agreement with Wells’ theoretical predictions decades earlier. Positive aerosol cultures were associated with higher physical performance (Karnofsky) scores (P < 0.016), higher sputum AFB smear microscopy grades (P < 0.007), lower days to positive in liquid culture (P < 0.004), stronger cough (P < 0.016), and fewer days on TB treatment prior to aerosol collection (P < 0.047). In multivariable analyses, cough aerosol cultures were associated with a salivary/mucosalivary (compared with purulent/mucopurulent) appearance of sputum (odds ratio, 4.42; 95% CI, 1.23–21.43) and low days to positive in liquid culture (per 1 day decrease; odds ratio, 1.17; 95%CI, 1.07–1.33). This study demonstrated the feasibility of using this research method in a resource-limited setting and again suggested considerable variability in the ability to produce infectious aerosols among TB patients. These data suggest that although the bacillary load in sputum is one of the predictors of cough aerosol cultures, sputum bacillary load is not the only predictor. As noted by CDC investigators 30 years ago, the bacillary load in the sputum should be considered a risk factor for transmission, not the sole definition of infectiousness (62).
We followed up this study with another at the same site, this time linking cough aerosol cultures from the index TB cases to new infections among their HHCs, the primary outcome being TST conversion (68). We enrolled 96 sputum culture-positive index TB cases and their 442 contacts. Contacts of patients with TB who produced high aerosols (>10 CFU) were more likely to have a new infection compared with contacts from low-aerosol (1–9 CFU) and aerosol-negative cases (83%, 32%, and 30%, respectively; P < 0.009). A high-aerosol patient with TB was the only predictor of new M. tuberculosis infection in unadjusted (odds ratio, 6.90; 95% confidence interval, 6.90, 16.42) and adjusted analyses (odds ratio, 4.81; 95% confidence interval, 1.20–19.23). In other words, when cough aerosols were included in models, the traditional indicators of infectiousness such as AFB sputum smear, culture, or cavitation were no longer significant (Figure 4).
FIGURE 4
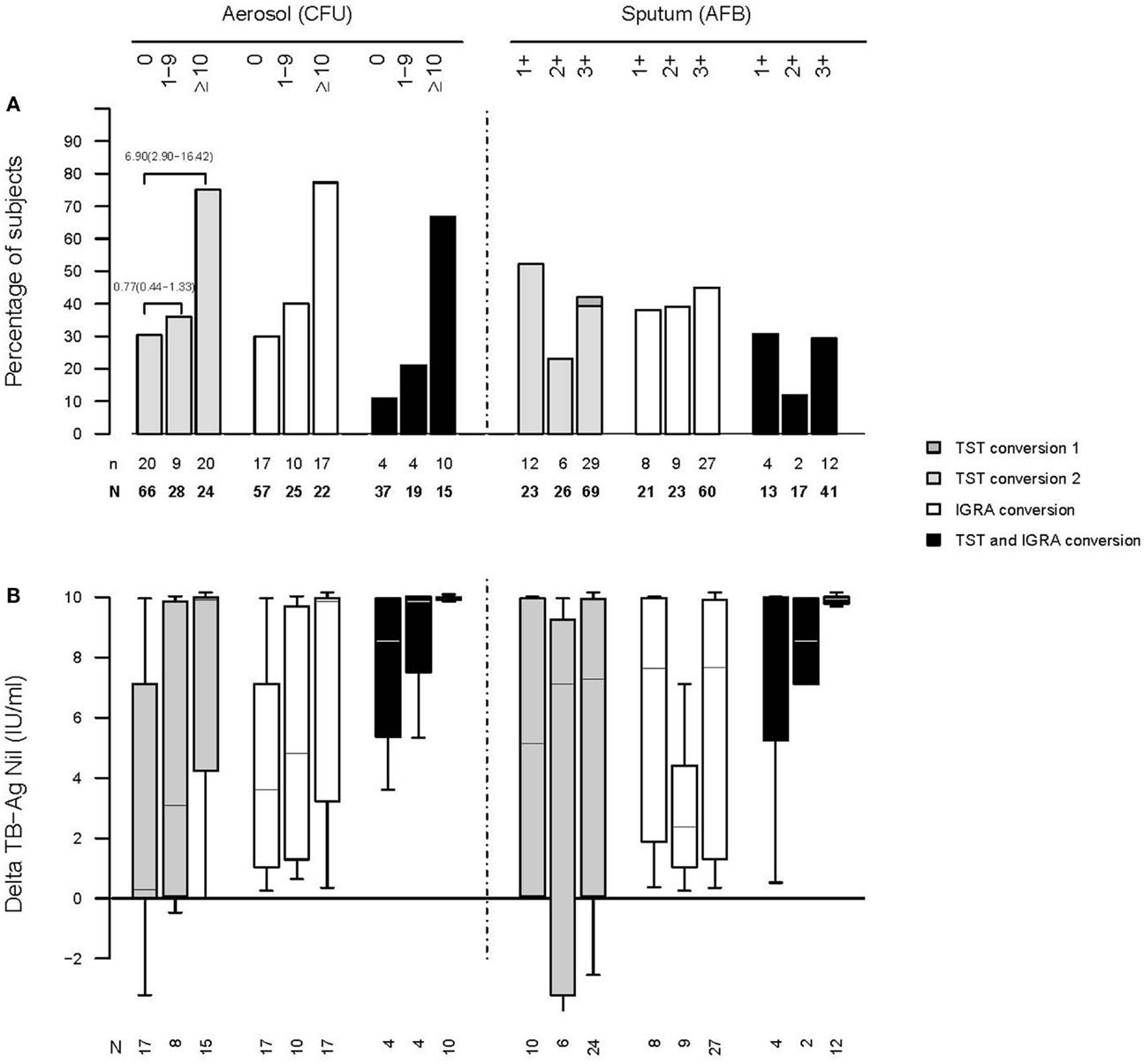
Figure 4. Increasing rates of infection among household contacts of active cases of TB who produced larger cough aerosols (A). Increasing amounts of interferon gamma output in an interferon-gamma release assay among household contacts exposed to active cases of TB who produce larger cough aerosols (B) [from Ref. (68); please see that paper for details regarding numbers of subjects listed].
In addition to these data, the use of cough aerosols as the best indicator of infectiousness is supported by Hill’s guidelines to strengthen the inference that a statistical association is also causal (69). There is clearly more biological plausibility favoring cough aerosols over sputum, as we know that pulmonary TB is not transmitted by sputum but rather through aerosols, i.e., infectious particles in the air. The experimental animal model evidence reviewed above favors cough aerosols over sputum, although a head-to-head comparison of the AFB sputum smear and cough aerosol cultures has not been done. However, this can be tested relatively easily using the approach of natural infection of guinea pigs (64). Temporality is a difficult criterion to satisfy for any surrogate, including sputum, necessarily obtained after infection has occurred. The strength of the association now favors cough aerosols given the data presented above, although it would be ideal for those data to be replicated in other settings by other investigators. Thus, there has not yet been enough data generated to evaluate consistency. Finally, we have argued above that there is a gradient, or dose–response, so we suggest that cough aerosols are the best surrogate for inhaled dose.
Why should M. tuberculosis be one of the few pathogens which is transmitted exclusively by the airborne route? We suggest that one reason is that airborne transmission of other pathogens has not been well studied. There is a growing body of literature indicating that influenza is probably at least partially transmitted by the airborne route as well as by large aerosols and direct contact (70). Pseudomonas aeruginosa and other Gram-negative bacilli (GNB) have been thought to be transmitted only by large droplet transmission, but coughing CF patients readily produce viable aerosols of these opportunistic pathogens (60, 71). However, transmission of disease via the fine aerosols of these GNB has not yet been demonstrated. It is most likely that many pathogens can be aerosolized from the human respiratory tract by coughing or sneezing. Whether or not transmission occurs is dependent on the concentration at the time of emission from the index case and subsequent dilution before inhalation by the exposed host as well as survival of the immediate desiccation and temperature change, exposure to ultraviolet light, and other environmental stresses during transport (72).
Hypothesis 2. Large droplet aerosols (>5 μ) of M. tuberculosis that deposit in the upper airway can induce immune responses without establishment of infection (Figure 5).
FIGURE 5
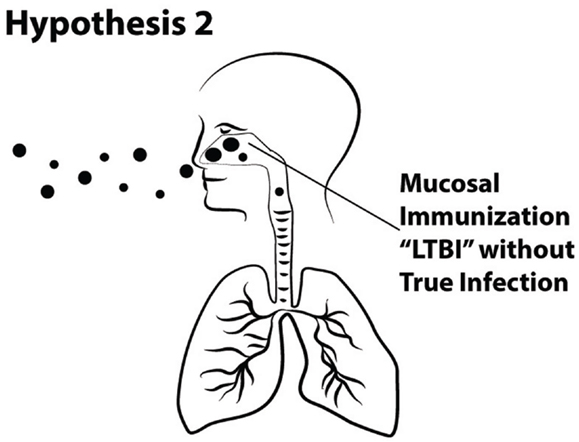
Figure 5. Schematic of Hypothesis 2 demonstrating immunization via the mucosa of the upper airway without the establishment of true TB infection in the alveolar region of the lung.
Although the discovery that TB is transmitted by the airborne route was brilliant, we respectfully suggest that the assumption that it is transmitted only by the airborne route is illogical. In this section, we argue that although TB disease is clearly preferentially transmitted by the airborne route, this is not exclusive or obligate. Furthermore, sensitization or immunization may occur due to deposition and presentation of tuberculous antigens in the upper airway, without causing persistent infection or disease.
A Short History of the Science of TB Transmission Mechanisms
Cornet (a student of Robert Koch) inoculated guinea pigs with bacilli from the dust from the rooms of TB patients and was able to produce infection in 40 of 118 samples in one experiment and in 15 of 21 samples from medical wards, demonstrating that tubercle bacilli can remain infectious for long periods of time. The fact that M. tuberculosis can be found in dust attests to the fact that large droplets from patients settle to the ground. Flugge doubted the role of dust and appears to be the first to consider cough aerosols as a primary mechanism of transmission of TB. He sprayed dried versus moist sputum of TB patients to create aerosol exposures of guinea pigs and it was only those exposed to the moist sprays that became infected, and he had patients cough into the air, exposing guinea pigs. On finding that droplets on settle plates between the patients and the guinea pigs grew M. tuberculosis, he concluded (we now know incorrectly) that these large droplets caused TB (73). However, these early experiments indicated that large droplet aerosols from TB patients contained viable M. tuberculosis.
Another early line of evidence indicating large droplet transmission was the observation that M. tuberculosis could be cultivated from the nasal cavities of 9 out of 29 healthy hospital workers with no evidence of TB disease (74). This is similar to another potentially lethal respiratory infection, inhalation anthrax, that was historically an occupational disease associated with exposures to wools and other materials contaminated with Bacillus anthracis. It was common to find anthrax spores in anterior nasal swabs and pharyngeal washings in exposed workers who had no clinical symptoms or signs of disease (75).
High Rates of Reactivation of LTBI
An accepted paradigm in the science of TB is that only 10% of persons infected with M. tuberculosis will develop TB disease, half of those developing disease within the first few years of exposure and the others at a later point in their lives. However, we are unable to find good explanations for why 90% of infected humans never develop disease. More recently, Horsburgh and colleagues evaluated data from the United States, and they found a low overall number of cases developing within the first 5 years after a positive TST (76). Overall, there were only 27 TB cases among 7742 persons tested, or 0.35% over 5 years. However, among the 106 HIV infected persons in that study, there were 12 cases, or 11.3% over 5 years. This represents the highest rate of reactivation studies.
Although HIV infection has been associated with high rates of reactivation of TB, the use of TNF-alpha inhibitor drugs has also been associated with a large amount of disease reactivation. Early reports were case series that did not include data on baseline TSTs or IGRAs (77). In a report from France on 69 cases of TB associated with the use of TNF-alpha inhibitors, there were 15 TB cases among the 38 persons (39.5%) with TSTs of 5 mm or more.
So if we accept TNF inhibition as the strongest inducer of reactivation TB, even more so than HIV, why do we not see more than 40% develop active TB disease, especially when nearly all NHPs who are experimentally infected and then treated with TNF-alpha inhibition reactivate (34)? We propose one explanation is that some people are exposed to large droplet aerosols of M. tuberculosis that deposit in their upper airway and induce an immune response but do not cause infection. The traditional test defining LTBI has been the TST, but it reflects delayed hypersensitivity to the antigens of M. tuberculosis and is only a surrogate for cell-mediated immunity (CMI). The more recently developed IGRAs are probably a better reflection of CMI. However, in the discussion to follow, we suggest that both delayed hypersensitivity and CMI may be stimulated without a true infection occurring.
Cough Aerosol Particle Size Distribution in Health and Disease
Aerosols are suspensions of particles in air (65, 78).The most important determinant of particle deposition within the respiratory tract is the mass median aerodynamic diameter (MMAD), or the diameter of a particle if it behaved as a sphere (65, 78). For simplicity, we will simply use the term “diameter.” Most aerosols in nature contain a wide range of sizes, or they are “polydisperse.” “Monodisperse aerosols” of a single diameter can generally only be produced in the laboratory.
Cough Aerosols from Healthy Humans
Most attempts to study the aerosols generated from the human respiratory tract have used healthy volunteers. In early studies, investigators applied dyes to the oral cavity and then measured the large droplets from the “mouth spray” on oiled slides held a few inches from the mouth; the size of smaller droplets were estimated by calculations from the larger ones. There was a wide range of particles from 1 to 2000 μ (79) In a more recent study, aerosols from five healthy human volunteers that were mouth breathing, nose breathing, coughing, and talking were measured using an optical particle counter and an analytical transmission electron microscope (80). Coughing produced the largest concentrations of droplets, but there was considerable variability. Most recently, Zayas and colleagues used a laser diffraction system to study 45 healthy subjects and found that voluntary coughs generated particles from 0.1 to 900 μ in size (81) (Figure 6).
FIGURE 6
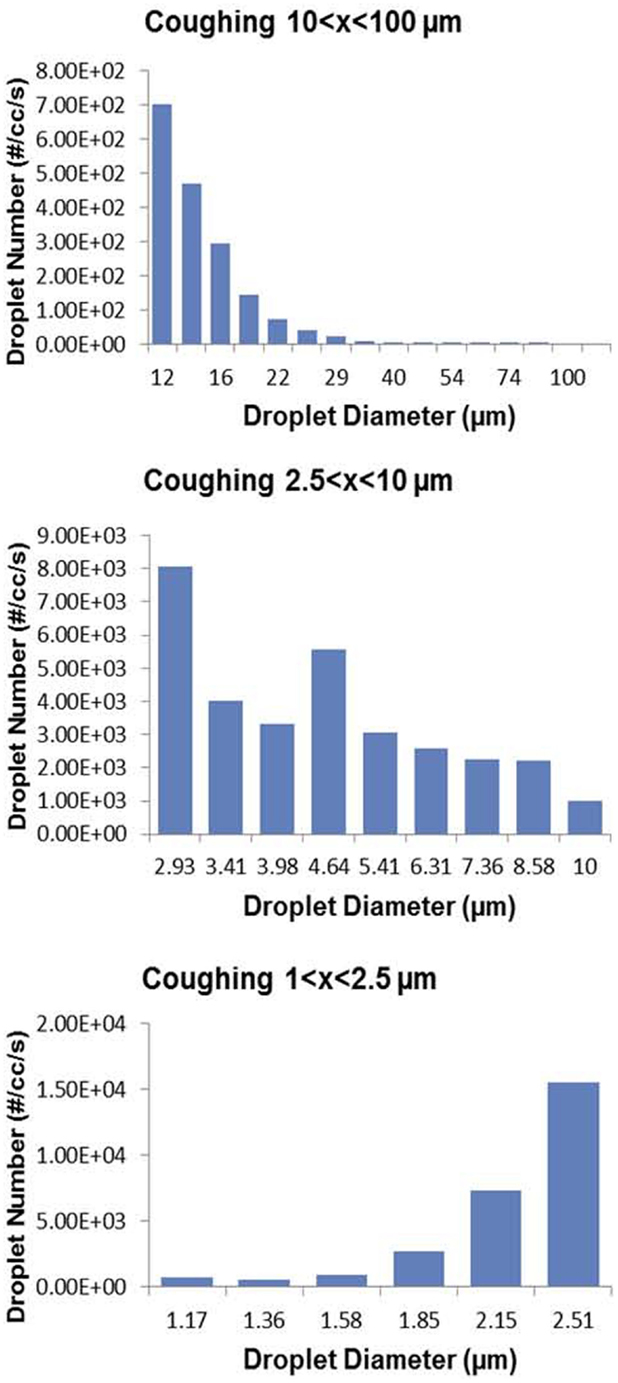
Figure 6. Wide range of particle size distributions of aerosols produced by voluntary coughing by healthy subjects. From Ref. (80).
Cough Aerosols during Pulmonary Infections
While the above data from healthy humans provide valuable insights into the particle sizes that can be emitted from the human respiratory tract, healthy humans do not cough and do not produce sputum. These data did not inform the size of the particles in which pathogens were emitted and transported. The classic and elegant early studies by Flugge, Riley, Wells, Lurie, and others determined that aerosols were generated from TB patients, but to our knowledge the particle size distribution had never been measured directly. The CASS apparatus discussed above is a method of quantifying and sizing cough aerosols (63), and we found that nearly all (96.4%) particles with culturable M. tuberculosis in cough aerosols from patients with active pulmonary TB were <4.7 μ (67). In a study of patients with CF who were infected with P. aeruginosa, we found that 72% of the cough aerosol particles containing culturable P. aeruginosa were 3.3 μ or less (60), with a distribution similar to that of TB patients (Figure 7). However, we also found culturable bacilli in the connector tubing between the patient’s mouth and the sampling chamber (60), suggesting carriage of bacilli in large droplets. In a follow-up study of 19 CF patients at the same center, they all produced culturable aerosols that were again markedly variable among the subjects and mostly in particles <3.3 μ. In addition, culturable P. aeruginosa was isolated at 4 meters and persisted in the air for 45 min (71).
FIGURE 7
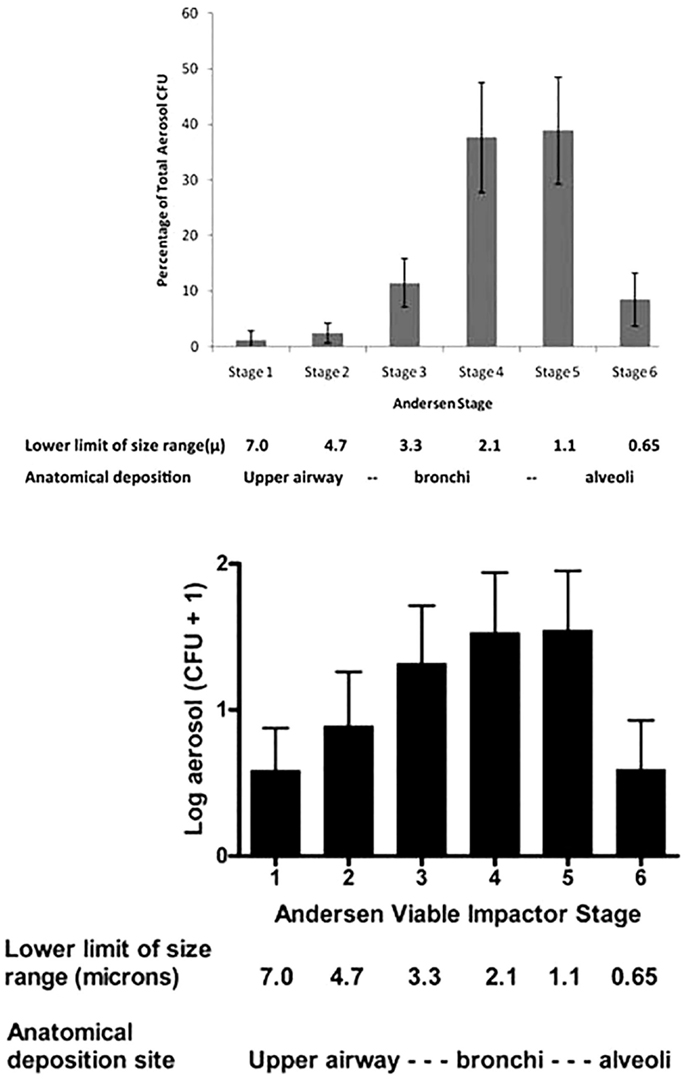
Figure 7. Particle size distributions of cough aerosols from patients with active pulmonary TB [top panel; from Ref. (67)] and from cystic fibrosis patients with chronic P. aeruginosa infections [bottom panel; from Ref. (60)].
Cough aerosols from patients with influenza have been measured more recently. In one study using Andersen cascade impactors as used for our TB and CF studies about, up to 89% of particles one foot away from patients were <4.7 μ (11% were larger) (82). In another study using different sampling methods of cough aerosols, influenza was found in 42% of particles <1 micron, in 23% of particles of 1–4 μ, and in 35% of particles >4 μ (83). In all of these studies, the sampling methods for viable pathogens are technically limited to the smaller size ranges, so our knowledge of the concentration of viable pathogens in large droplets is limited.
In summary, in both health and disease, humans generate cough aerosols in a broad range of particle sizes. Most pathogens are found in particles <4–5 μ, so predictably deposit in the lower respiratory tracts. However, pathogens, including M. tuberculosis, are also present in larger particles that probably deposit in the upper airways.
Primary TB of the Upper Airway
Given the above discussion about the size distribution of cough aerosols from patients with pulmonary TB, it should not be surprising that M. tuberculosis can deposit in the upper airways. Most nasal TB occurs in conjunction with pulmonary TB. Primary nasal TB, by definition without co-existing pulmonary infection, is rare, but there are multiple case reports (84–88). Laryngeal TB almost always occurs in tandem with pulmonary TB, but primary TB of the larynx occurs, probably more rarely than primary nasal TB (89–93).
Primary oral TB is also rare, although M. tuberculosis may be cultured from the dental plaque and saliva of patients with pulmonary TB (94). In a study of TB patients with periodontal disease, 92% had M. tuberculosis isolated in the saliva and 68% in the dental plaque. In this endemic setting in India, 12% of periodontitis patients not diagnosed with TB also had M. tuberculosis isolated from the saliva (95). In an unusual outbreak, a dentist with pulmonary TB had presumably coughed into the mouths of the cases or contaminated them with M. tuberculosis on his fingers, resulting in 13 cases of primary intraoral TB and involvement of regional lymph nodes, 1 case of both intraoral and pulmonary TB, and 1 case of erythema nodosum and pleural effusion (96). In several of these case reports, clinicians have suspected that nose-picking and smoking/alcohol use are risk factors for primary nasal and laryngeal TB, respectively. In the case of the outbreak of primary intraoral TB, dental manipulation or tooth extractions could obviously damage the oral mucosa. The suspected need for damaged mucosa is supported by a study using organ cultures of human tissue in which M. tuberculosis only adhered to extracellular matrix in areas of mucosal damage, but not to ciliated mucosa, intact extruded cells, basement membrane, or collagen fibers (97). We suspect that the rarity of primary TB of the upper airway is explained by the requirement of both damaged mucosa as well as deposition of large aerosol particles from a TB patient.
Cervical Lymphadenitis
The cervical lymph nodes are the most common site of TB lymphadenitis. Cervical lymphadenitis due to M. tuberculosis is thought by some authors to be a “local manifestation of a systemic disease,” presumably via lymphatic drainage from pulmonary lymph nodes (98). However, it is also clear that cervical nodes may become involved due to tuberculous infection in the head or neck (99). In a study of 100 cases of histologically confirmed TB cervical lymphadenitis, the most common site of disease was in the posterior triangle (51%), followed by the upper deep cervical (48%) and submandibular nodes (100). The supraclavicular and lower deep cervical nodes were involved in only 3 and 9%, respectively. These data suggest that an unappreciated source of infection of upper cervical lymph nodes is the upper airway. Cervical lymphadenitis from non-tuberculous mycobacteria, most common in children, is assumed to be due to a portal of infection in the oropharynx or conjunctiva (101). We propose that the same is likely to be true for M. tuberculosis if we accept that it is found in large aerosol droplets.
Mucosal Immunization via the Upper Airway
Patients who suffer from “hay fever” (allergic rhinitis) are all too familiar with the immunological hypersensitivity response after airborne particles of pollen (that are usually around 20 μ or larger, similar to large droplet aerosols) deposit in the nose. The mucosa of the upper airway is a rich connection between the environment and the immune system. There is a rapidly growing literature on the use of intranasal vaccination, also known as mucosal immunization. Intranasal administration of vaccines provides protection against a variety of respiratory pathogens (102). Intranasal vaccination with M. bovis BCG can elicit not only protection against aerosol challenge with M. tuberculosis H37Rv, but it can also elicit granuloma formation in the deep lung (103). Even heat-killed BCG delivered intranasally with an adjuvant can induce systemic immune responses (104). Intranasal administration of BCG to mice appears to offer short-term protection in the lung but persistent immune responses in the spleen for at least 10 months (105). We speculate that this may help explain the poor ability of BCG vaccination to protect humans from pulmonary TB but the documented protection against disseminated disease.
Summary
An inoculum effect is common in most infectious diseases, and we have reviewed data demonstrating the greater likelihood of more progressive TB disease with greater magnitudes of inhaled infectious aerosol or with surrogate measures for the same. We propose that cough aerosols of M. tuberculosis are the best surrogate measure to test our first hypothesis that inhaled dose predicts immunopathology in human TB. Our second hypothesis, that large droplet aerosols of M. tuberculosis can induce immune responses without establishing infection can be tested in both experimental animal models and perhaps in the near future in humans with attenuated live mycobacteria or with mycobacterial antigens. Such research would have the potential to discover better biomarkers of true persisting infection, and hopefully, to better identify those at risk for progression to active TB.
Author Contributions
KF and EJ-L both conceived of this work and contributed to the review of the literature; both contributed to drafts, revisions, and the final manuscript; both are accountable for all work and agree to the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Esmail H, Barry CE III, Wilkinson RJ. Understanding latent tuberculosis: the key to improved diagnostic and novel treatment strategies. Drug Discov Today (2012) 17(9–10):514–21. doi: 10.1016/j.drudis.2011.12.013
2. Osterholm MT, Hedberg CW. Epidemiologic principles. In: Mandell GL, Bennet JE, Dolin R, editors. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases. Philadelphia, PA: Churchill Livingstone (2009). p. 179–93.
3. Dance DA. Ecology of Burkholderia pseudomallei and the interactions between environmental Burkholderia spp. and human-animal hosts. Acta Trop (2000) 74(2–3):159–68. doi:10.1016/S0001-706X(99)00066-2
4. Duma RJ. Gram-negative bacillary infections. Pathogenic and pathophysiologic correlates. Am J Med (1985) 78(6A):154–64. doi:10.1016/0002-9343(85)90119-6
5. Rubin LG. Bacterial colonization and infection resulting from multiplication of a single organism. Rev Infect Dis (1987) 9(3):488–93. doi:10.1093/clinids/9.3.488
6. Bradsher RW. Histoplasmosis and blastomycosis. Clin Infect Dis (1996) 22(Suppl 2):S102–11. doi:10.1093/clinids/22.Supplement_2.S102
7. Wheat LJ. Diagnosis and management of histoplasmosis. Eur J Clin Microbiol Infect Dis (1989) 8(5):480–90. doi:10.1007/BF01964063
8. Alberti A, Chemello L, Benvegnu L. Natural history of hepatitis C. J Hepatol (1999) 31(Suppl 1):17–24.
9. Friedland GH, Klein RS. Transmission of the human immunodeficiency virus. N Engl J Med (1987) 317:1125–35. doi:10.1056/NEJM198710293171806
10. Quinn TC, Brookmeyer R, Kline R, Shepherd M, Paranjape R, Mehendale S, et al. Feasibility of pooling sera for HIV-1 viral RNA to diagnose acute primary HIV-1 infection and estimate HIV incidence. AIDS (2000) 14:2751–7.
11. Pereira BA, Alves CR. Immunological characteristics of experimental murine infection with Leishmania (Leishmania) amazonensis. Vet Parasitol (2008) 158:239–55. doi:10.1016/j.vetpar.2008.09.015
12. Hornick RB, Greisman SE, Woodward TE, DuPont HL, Dawkins AT, Snyder MJ. Typhoid fever: pathogenesis and immunologic control. 2. N Engl J Med (1970) 283:739–46. doi:10.1056/NEJM197010012831406
13. Hornick RB, Greisman SE, Woodward TE, DuPont HL, Dawkins AT, Snyder MJ. Typhoid fever: pathogenesis and immunologic control. N Engl J Med (1970) 283(13):686–91. doi:10.1056/NEJM197009242831306
14. Burgin S, et al. Mycobacteria and the skin. In: Rom WN, Garay SM, editors. Tuberculosis. Philadelphia, PA: Lippincott Williams and Wilkins (2004). p. 593–608.
15. Heilman KM, Muschenheim C. Primary cutaneous tuberculosis resulting from mouth-to-mouth respiration. N Engl J Med (1965) 273(19):1035–6. doi:10.1056/NEJM196511042731908
16. Marmelzat WL. Laennec and the “prosector’s wart”. Arch Dermatol (1962) 86(1):74–6. doi:10.1001/archderm.1962.01590070080014
17. Koch R. The etiology of tuberculosis (Die Aetiologie der Tuberculose in Berliner klinische Wochenschrift 1882;19:221-30). Essays of Robert Koch. Westport, CT: Greenwood Press, Inc. (1987). p. 221–30.
18. Glover RE. Infection of mice with Mycobact. tuberculosis (bovis) by the respiratory route. Br J Exp Pathol (1944) 25:141–9.
19. Youmans GP, Youmans AS. The relation between the size of the infecting dose of tubercle bacilli and the survival time of mice. Am Rev Tuberc (1951) 64:534–40.
20. Lurie MB. Resistance to Tuberculosis: Experimental Studies in Native and Acquired Defensive Mechanisms. Cambridge, MA: Harvard University Press (1964).
21. McMurray DN, Collins FM, Dannenberg AM Jr, Smith DW. Pathogenesis of experimental tuberculosis in animal models. Curr Top Microbiol Immunol (1996) 215:157–79.
22. Wells WF, Ratcliffe HL, Crumb C. On the mechanics of droplet nuclei infection: II. Quantitative experimental air-borne tuberculosis in rabbits. Am J Hyg (1948) 47:11–28.
23. Rhoades ER, Frank AA, Orme IM. Progression of chronic pulmonary tuberculosis in mice aerogenically infected with virulent mycobacterium tuberculosis. Tuber Lung Dis (1997) 78:57–66.
24. de Steenwinkel JE, ten Kate MT, de Knegt GJ, Verbrugh HA, van Belkum A, Hernandez-Pando R, et al. Course of murine tuberculosis and response to first-line therapy depends on route of infection and inoculum size. Int J Tuberc Lung Dis (2011) 15:1478–84. doi:10.5588/ijtld.11.0012
25. Ferebee SH, Mount FW. Tuberculosis morbidity in a controlled trial of the prophylactic use of isoniazid among household contacts. Am Rev Respir Dis (1962) 85:490–510.
26. Marks SM, Taylor Z, Burrows NR, Qayad MG, Miller B. Hospitalization of homeless persons with tuberculosis in the United States. Am J Public Health (2000) 90(3):435–8. doi:10.2105/AJPH.90.3.435
27. McCune RM Jr, McDermott W, Tompsett R. The fate of Mycobacterium tuberculosis in mouse tissues as determined by the microbial enumeration technique. II. The conversion of tuberculous infection to the latent state by the administration of pyrazinamide and a companion drug. J Exp Med (1956) 104(5):763–802. doi:10.1084/jem.104.5.763
28. Walsh GP, Tan EV, dela Cruz EC, Abalos RM, Villahermosa LG, Young LJ, et al. The Philippine cynomolgus monkey (Macaca fasicularis) provides a new nonhuman primate model of tuberculosis that resembles human disease. Nat Med (1996) 2(4):430–6. doi:10.1038/nm0496-430
29. Capuano SV III, Croix DA, Pawar S, Zinovik A, Myers A, Lin PL, et al. Experimental Mycobacterium tuberculosis infection of cynomolgus macaques closely resembles the various manifestations of human M. tuberculosis infection. Infect Immun (2003) 71(10):5831–44. doi:10.1128/IAI.71.10.5831-5844.2003
30. Mehra S, Pahar B, Dutta NK, Conerly CN, Philippi-Falkenstein K, Alvarez X, et al. Transcriptional reprogramming in nonhuman primate (rhesus macaque) tuberculosis granulomas. PLoS One (2010) 5(8):e12266. doi:10.1371/journal.pone.0012266
31. Dutta NK, Mehra S, Kaushal D. A Mycobacterium tuberculosis sigma factor network responds to cell-envelope damage by the promising anti-mycobacterial thioridazine. PLoS One (2010) 5(4):e10069. doi:10.1371/journal.pone.0010069
32. Mehra S, Golden NA, Dutta NK, Midkiff CC, Alvarez X, Doyle LA, et al. Reactivation of latent tuberculosis in rhesus macaques by coinfection with simian immunodeficiency virus. J Med Primatol (2011) 40(4):233–43. doi:10.1111/j.1600-0684.2011.00485.x
33. Diedrich CR, Mattila JT, Klein E, Janssen C, Phuah J, Sturgeon TJ, et al. Reactivation of latent tuberculosis in cynomolgus macaques infected with SIV is associated with early peripheral T cell depletion and not virus load. PLoS One (2010) 5(3):e9611. doi:10.1371/journal.pone.0009611
34. Lin PL, Myers A, Smith L, Bigbee C, Bigbee M, Fuhrman C, et al. Tumor necrosis factor neutralization results in disseminated disease in acute and latent Mycobacterium tuberculosis infection with normal granuloma structure in a cynomolgus macaque model. Arthritis Rheum (2010) 62(2):340–50. doi:10.1002/art.27271
35. Lin PL, Ford CB, Coleman MT, Myers AJ, Gawande R, Ioerger T, et al. Sterilization of granulomas is common in active and latent tuberculosis despite within-host variability in bacterial killing. Nat Med (2014) 20(1):75–9. doi:10.1038/nm.3412
36. Ferebee SH. Controlled chemoprophylaxis trials in tuberculosis. A general review. Bibl Tuberc (1970) 26:28–106.
37. Horsburgh CR Jr, Rubin EJ. Clinical practice. Latent tuberculosis infection in the United States. N Engl J Med (2011) 364(15):1441–8. doi:10.1056/NEJMcp1005750
38. Grzybowski S, Barnett GD, Styblo K. Contacts of cases of active pulmonary tuberculosis. Bull Int Union Tuberc (1975) 50:90–106.
39. Radhakrishna S, Frieden TR, Subramani R, Santha T, Narayanan PR. Additional risk of developing TB for household members with a TB case at home at intake: a 15-year study. Int J Tuberc Lung Dis (2007) 11:282–8.
40. Cruz-Ferro E, Ursúa-Díaz MI, Taboada-Rodríguez JA, Hervada-Vidal X, Anibarro L, Túñez V, et al. Epidemiology of tuberculosis in Galicia, Spain, 16 years after the launch of the Galician tuberculosis programme. Int J Tuberc Lung Dis (2014) 18(2):134–40. doi:10.5588/ijtld.13.0419
41. Cruz-Ferro E, Ursúa-Díaz MI, Taboada-Rodríguez JA, Hervada-Vidal X, Anibarro L, Túñez V, et al. In reply. Int J Tuberc Lung Dis (2014) 18(6):752–3. doi:10.5588/ijtld.14.0084-2
42. Rouillion A, Perdrizet S, Parrot R. Transmission of tubercle bacilli: the effects of chemotherapy. Tubercle (1976) 57:275–99. doi:10.1016/S0041-3879(76)80006-2
43. Shaw JB, Wynn-Williams N. Infectivity of pulmonary tuberculosis in relation to sputum status. Am Rev Tuberc (1954) 69:724–32.
44. Menzies RI. Tuberculin skin testing. In: Reichman LB, Hershfield ES, editors. Tuberculosis: A Comprehensive International Approach. New York, NY: Marcel Dekker, Inc. (2000). p. 279–322.
45. Huebner RE, Schein MF, Bass JBJ. The tuberculin skin test. Clin Infect Dis (1993) 17:968–75. doi:10.1093/clinids/17.6.968
46. Fennelly KP. Variability of airborne transmission of Mycobacterium tuberculosis: implications for control of tuberculosis in the HIV era. Clin Infect Dis (2007) 44(10):1358–60. doi:10.1086/516617
47. van Geuns HA, Meijer J, Styblo K. Results of contact examination in Rotterdam, 1967-1969. Bull Int Union Tuberc (1975) 50(1):107–21.
48. Sepkowitz KA. Tuberculin skin testing and the health care worker: lessons of the Prophit Survey. Tuber Lung Dis (1996) 77:81–5. doi:10.1016/S0962-8479(96)90081-7
49. Lienhardt C, Fielding K, Sillah J, Tunkara A, Donkor S, Manneh K, et al. Risk factors for tuberculosis infection in sub-Saharan Africa: a contact study in The Gambia. Am J Respir Crit Care Med (2003) 168(4):448–55. doi:10.1164/rccm.200212-1483OC
50. Lutong L, Bei Z. Association of prevalence of tuberculin reactions with closeness of contact among household contacts of new smear-positive pulmonary tuberculosis patients. Int J Tuberc Lung Dis (2000) 4(3):275–7.
51. Rathi SK, Akhtar S, Rahbar MH, Azam SI. Prevalence and risk factors associated with tuberculin skin test positivity among household contacts of smear-positive pulmonary tuberculosis cases in Umerkot, Pakistan. Int J Tuberc Lung Dis (2002) 6:851–7.
52. Hutton MD, Stead WW, Cauthen GM, Bloch AB, Ewing WM. Nosocomial transmission of tuberculosis associated with a draining abscess. J Infect Dis (1990) 161:286–95. doi:10.1093/infdis/161.2.286
53. Kenyon TA, Valway SE, Ihle WW, Onorato IM, Castro KG. Transmission of multidrug-resistant Mycobacterium tuberculosis during a long airplane flight. N Engl J Med (1996) 334:933–8. doi:10.1056/NEJM199604113341501
54. Houk VH, Kent DC, Baker JH, Sorensen K, Hanzel GD. The Byrd study. In-depth analysis of a micro-outbreak of tuberculosis in a closed environment. Arch Environ Health (1968) 16:4–6. doi:10.1080/00039896.1968.10665007
55. Houk VN, Baker JH, Sorensen K, Kent DC. The epidemiology of tuberculosis infection in a closed environment. Arch Environ Health (1968) 16(1):26–35. doi:10.1080/00039896.1968.10665011
56. Hertzberg G. The infectiousness of tuberculosis; an epidemiological investigation. Acta Tuberc Scand Suppl (1957) 38:1–146.
57. Loudon RG, Spohn SK. Cough frequency and infectivity in patients with pulmonary tuberculosis. Am Rev Respir Dis (1969) 99:109–11.
58. Jones-López EC, Kim S, Fregona G, Marques-Rodrigues P, Hadad DJ, Molina LP, et al. Importance of cough and M. tuberculosis strain type as risks for increased transmission within households. PLoS One (2014) 9(7):e100984. doi:10.1371/journal.pone.0100984
59. Borgdorff MW, Nagelkerke NJ, de Haas PE, van Soolingen D. Transmission of Mycobacterium tuberculosis depending on the age and sex of source cases. Am J Epidemiol (2001) 154:934–43. doi:10.1093/aje/154.10.934
60. Wainwright CE, France MW, O’Rourke P, Anuj S, Kidd TJ, Nissen MD, et al. Cough-generated aerosols of Pseudomonas aeruginosa and other Gram-negative bacteria from patients with cystic fibrosis. Thorax (2009) 64(11):926–31. doi:10.1136/thx.2008.112466
62. Snider DE Jr, Kelly GD, Cauthen GM, Thompson NJ, Kilburn JO. Infection and disease among contacts of tuberculosis cases with drug-resistant and drug-susceptible bacilli. Am Rev Respir Dis (1985) 132(1):125–32.
63. Fennelly KP, Martyny JW, Fulton KE, Orme IM, Cave DM, Heifets LB. Cough-generated aerosols of Mycobacterium tuberculosis: a new method to study infectiousness. Am J Respir Crit Care Med (2004) 169(5):604–9. doi:10.1164/rccm.200308-1101OC
64. Nardell EA. Catching droplet nuclei: toward a better understanding of tuberculosis transmission. Am J Respir Crit Care Med (2004) 169(5):553–4. doi:10.1164/rccm.2401003
65. Baron PA, Willeke K. Aerosol fundamentals. In: Baron PA, Willeke K, editors. Aerosol Measurement: Principles, Techniques, and Applications. New York, NY: Wiley-Interscience (2001). p. 45–60.
67. Fennelly KP, Jones-López EC, Ayakaka I, Kim S, Menyha H, Kirenga B, et al. Variability of infectious aerosols produced during coughing by patients with pulmonary tuberculosis. Am J Respir Crit Care Med (2012) 186(5):450–7. doi:10.1164/rccm.201203-0444OC
68. Jones-López EC, Namugga O, Mumbowa F, Ssebidandi M, Mbabazi O, Moine S, et al. Cough aerosols of Mycobacterium tuberculosis predict new infection: a household contact study. Am J Respir Crit Care Med (2013) 187(9):1007–15. doi:10.1164/rccm.201208-1422OC
69. Hill AB. The environment and disease: association or causation? Proc R Soc Med (1965) 58:295–300.
70. Tellier R. Review of aerosol transmission of influenza A virus. Emerg Infect Dis (2006) 12(11):1657–62. doi:10.3201/eid1211.060426
71. Knibbs LD, Johnson GR, Kidd TJ, Cheney J, Grimwood K, Kattenbelt JA, et al. Viability of Pseudomonas aeruginosa in cough aerosols generated by persons with cystic fibrosis. Thorax (2014) 69(8):740–5. doi:10.1136/thoraxjnl-2014-205213
72. Roy CJ, Milton DK. Airborne transmission of communicable infection – the elusive pathway. N Engl J Med (2004) 350:1710–2. doi:10.1056/NEJMp048051
73. Flugge C. Die verreitung der Phthise durch staubtormiges sputum und durch beim husten verspritzte tropfchen. Z Hyg Infektionskr (1898) 29:107–24.
74. Straus I. Sur la presence du bacille de la tuberculose dans les cavites nasales de l’homme sain. Archiv de medl experiment and d’anatomie path (1894) 4:633–9.
75. Brachman PS, Kaufman AF, Dalldorf FG. Industrial inhalation anthrax. Bacteriol Rev (1966) 30(3):646–59.
76. Horsburgh CR Jr, O’Donnell M, Chamblee S, Moreland JL, Johnson J, Marsh BJ, et al. Revisiting rates of reactivation tuberculosis: a population-based approach. Am J Respir Crit Care Med (2010) 182(3):420–5. doi:10.1164/rccm.200909-1355OC
77. Keane J, Gershon S, Wise RP, Mirabile-Levens E, Kasznica J, Schwieterman WD, et al. Tuberculosis associated with infliximab, a tumor necrosis factor alpha-neutralizing agent. N Engl J Med (2001) 345:1098–104.
78. Yeates DB, Mortensen J. Deposition and clearance. In: Murray JF, Nadel JA, editors. Textbook of Respiratory Medicine. Philadelphia, PA: W.B. Saunders Company (2000). p. 349–84.
79. Duguid JP. The size and duration of air carriage of respiratory droplets and droplet nuclei. J Hyg (Lond) (1946) 44:471–9. doi:10.1017/S0022172400019288
80. Papineni RS, Rosenthal FS. The size distribution of droplets in the exhaled breath of healthy human subjects. J Aerosol Med (1997) 10(2):105–16. doi:10.1089/jam.1997.10.105
81. Zayas G, Chiang MC, Wong E, MacDonald F, Lange CF, Senthilselvan A, et al. Cough aerosol in healthy participants: fundamental knowledge to optimize droplet-spread infectious respiratory disease management. BMC Pulm Med (2012) 12:11. doi:10.1186/1471-2466-12-11
82. Bischoff WE, Swett K, Leng I, Peters TR. Exposure to influenza virus aerosols during routine patient care. J Infect Dis (2013) 207(7):1037–46. doi:10.1093/infdis/jis773
83. Lindsley WG, Blachere FM, Thewlis RE, Vishnu A, Davis KA, Cao G, et al. Measurements of airborne influenza virus in aerosol particles from human coughs. PLoS One (2010) 5(11):e15100. doi:10.1371/journal.pone.0015100
84. Kamath MP, Bhojwani KM, Prabhu S, Naik R, Ninan GP, Chakravarthy Y. Multifocal tuberculosis of the nose and lymph nodes without pulmonary involvement: a case report. Ear Nose Throat J (2007) 86(5):284–6.
85. Kameswaran M, Anand Kumar RS, Murali S, Raghunandan S, Vijaya Krishnan P. Primary nasal tuberculosis – a case report. Indian J Otolaryngol Head Neck Surg (2007) 59(1):87–9. doi:10.1007/s12070-007-0029-y
86. Masterson L, Srouji I, Kent R, Bath AP. Nasal tuberculosis – an update of current clinical and laboratory investigation. J Laryngol Otol (2011) 125(2):210–3. doi:10.1017/S0022215110002136
87. Prstačić R, Jurlina M, Žižić-Mitrečić M, Janjanin S. Primary nasopharyngeal tuberculosis mimicking exacerbation of chronic rhinosinusitis. J Laryngol Otol (2011) 125(7):747–9. doi:10.1017/S0022215110002835
88. Ribeiro S, Ferreira B, Duarte R, Almeida I, Vasconcelos C. Primary nasal tuberculosis during anti-tumour necrosis factor alpha treatment of a patient with rheumatoid arthritis. Scand J Infect Dis (2013) 45(12):957–60. doi:10.3109/00365548.2013.818246
89. El Ayoubi F, Chariba I, El Ayoubi A, Chariba S, Essakalli L. Primary tuberculosis of the larynx. Eur Ann Otorhinolaryngol Head Neck Dis (2014) 131(6):361–4. doi:10.1016/j.anorl.2013.10.005
90. Fsadni P, Fsadni C, Caruana Montaldo B. Primary laryngeal tuberculosis: an unusual cause of hoarseness. Ear Nose Throat J (2014) 93(10–11):E15–7.
91. Gandhi S, Kulkarni S, Mishra P, Thekedar P. Tuberculosis of larynx revisited: a report on clinical characteristics in 10 cases. Indian J Otolaryngol Head Neck Surg (2012) 64(3):244–7. doi:10.1007/s12070-011-0333-4
92. Huon LK, Fang TY. Primary laryngeal tuberculosis. J Formos Med Assoc (2011) 110(12):792–3. doi:10.1016/j.jfma.2011.11.011
93. Nasti G, Tavio M, Rizzardini G, De Paoli P, Morassut S, Barzan L, et al. Primary tuberculosis of the larynx in a patient infected with human immunodeficiency virus. Clin Infect Dis (1996) 23(1):183–4. doi:10.1093/clinids/23.1.183
94. Eng HL, Lu SY, Yang CH, Chen WJ. Oral tuberculosis. Oral Surg Oral Med Oral Pathol Oral Radiol Endod (1996) 81(4):415–20. doi:10.1016/S1079-2104(96)80016-3
95. Palakuru SK, Lakshman VK, Bhat KG. Microbiological analysis of oral samples for detection of Mycobacterium tuberculosis by nested polymerase chain reaction in tuberculosis patients with periodontitis. Dent Res J (Isfahan) (2012) 9(6):688–93.
96. Smith WH, Davies D, Mason KD, Onions JP. Intraoral and pulmonary tuberculosis following dental treatment. Lancet (1982) 1(8276):842–4. doi:10.1016/S0140-6736(82)91886-4
97. Middleton AM, Chadwick MV, Nicholson AG, Dewar A, Groger RK, Brown EJ, et al. Interaction of Mycobacterium tuberculosis with human respiratory mucosa. Tuberculosis (Edinb) (2002) 82(2–3):69–78. doi:10.1054/tube.2002.0324
98. Sloane MF. Mycobacterial lymphadenitis. In: Rom WN, Garay SM, editors. Tuberculosis. Philadelphia, PA: Lippincott, Williams, and Wilkins (2004). p. 489–96.
99. Sikora AGet al. Tuberculosis of the head and neck. In: Rom WN, Garay SM, editors. Tuberculosis. Philadelphia, PA: Lippincott, Williams, and Wilkins (2004). p. 477–88.
100. Baskota DK, Prasad R, Kumar Sinha B, Amatya RC. Distribution of lymph nodes in the neck in cases of tuberculous cervical lymphadenitis. Acta Otolaryngol (2004) 124(9):1095–8. doi:10.1080/00016480410018089
101. Manolidis S, Frenkiel S, Yoskovitch A, Black M. Mycobacterial infections of the head and neck. Otolaryngol Head Neck Surg (1993) 109(3 Pt 1):427–33.
102. Baumann U. Mucosal vaccination against bacterial respiratory infections. Expert Rev Vaccines (2008) 7(8):1257–76. doi:10.1586/14760584.7.8.1257
103. Tree JA, Williams A, Clark S, Hall G, Marsh PD, Ivanyi J. Intranasal bacille Calmette-Guerin (BCG) vaccine dosage needs balancing between protection and lung pathology. Clin Exp Immunol (2004) 138(3):405–9. doi:10.1111/j.1365-2249.2004.02648.x
104. Haile M, Schröder U, Hamasur B, Pawlowski A, Jaxmar T, Källenius G, et al. Immunization with heat-killed Mycobacterium bovis bacille Calmette-Guerin (BCG) in Eurocine L3 adjuvant protects against tuberculosis. Vaccine (2004) 22(11–12):1498–508. doi:10.1016/j.vaccine.2003.10.016
Keywords: tuberculosis, inhaled dose, immunology, cough aerosol, latent infection, TB transmission
Citation: Fennelly KP and Jones-López EC (2015) Quantity and quality of inhaled dose predicts immunopathology in tuberculosis. Front. Immunol. 6:313. doi: 10.3389/fimmu.2015.00313
Received: 28 February 2015; Paper pending published: 29 March 2015;
Accepted: 02 June 2015; Published: 29 June 2015
Edited by:
Vishwanath Venketaraman, Western University of Health Sciences, USAReviewed by:
Anthony George Tsolaki, Brunel University London, UKXun Suo, China Agricultural University, China
Copyright: © 2015 Fennelly and Jones-López. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kevin P. Fennelly, Division of Infectious Diseases and Global Medicine, Department of Medicine, Emerging Pathogens Institute, University of Florida, Room 257, P.O. Box 103600, Gainesville, FL 32610-3600, USA, kevin.fennelly@medicine.ufl.edu