Comparing marine primary production estimates through different methods and development of conversion equations
 Aurore Regaudie-de-Gioux
Aurore Regaudie-de-Gioux Sébastien Lasternas
Sébastien Lasternas Susana Agustí
Susana Agustí Carlos M. Duarte1,2
Carlos M. Duarte1,2- 1Department of Global Change Research, Instituto Mediterráneo de Estudios Avanzados (Consejo Superior de Investigaciones Científicas - Universidad de las Islas Baleares), Esporles, Spain
- 2The University of Western Australia Oceans Institute and School of Plant Biology, University of Western Australia, Perth, WA, Australia
Numerous studies have compared the rates of primary production using various techniques at specific locations and times. However, these comparisons are local and cannot be used to compare or scale rates of primary production using different methods across ocean basins or seasonal time scales. Here, we quantify the range in rates of primary production derived using different techniques and provide equations that allow conversions of estimates between different methods. We do so on the basis of a compilation of data on volumetric estimates of primary production rates concurrently estimated with at least two different methods. We observed that the comparison of estimates of marine phytoplankton primary production derived from different methods reveals very large variations between methods. The highest primary production estimates are derived using the 18O method, which may provide the best and more generally applicable estimate of gross primary production (GPP). The regression equations presented in this work provide the best available approach to convert data across methods and therefore integrate and synthesize available and future data derived using different methods.
Introduction
Plankton photosynthesis, responsible for about half of the primary production in the biosphere (Field et al., 1998), is a fundamental process at the global and the ecosystem scale. At the global scale, phytoplankton primary production affects oxygen and CO2 fluxes, constraining gas exchange with the atmosphere and thus the gaseous composition of the atmosphere. Furthermore, phytoplankton primary production is the main source of organic matter fueling marine food webs (Duarte et al., 1999). The measurement of phytoplankton primary production is so a fundamental property of the ocean ecosystem, receiving considerable effort that has resulted in several million estimates available to-date (del Giorgio and Williams, 2005).
Photosynthetic rates of marine phytoplankton were first measured using the oxygen evolution method in phytoplankton communities in the Oslo Fjord by Gaarder and Gran (1927). This method originally suffered from poor resolution, being unable to resolve the low primary production rates in the less productive regions of the ocean (Truesdale et al., 1955; Mortimer, 1956; Richards and Corwin, 1956). These limitations were resolved with the development of high-precision Winkler analyses using automatic titrators and end-point detection of the Winkler reaction (Carpenter, 1965; Carrit and Carpenter, 1966), which allow low primary production rates to be resolved. However, the 14C method was developed before improved oxygen-based techniques were available (Steeman Nielsen, 1952) and rapidly became the standard for the oceanographic community, used to calibrate remote sensing algorithms, despite recurrent caveats as to what exactly the 14C incorporation method measures (Dring and Jewson, 1982; Behrenfeld and Falkowski, 1997a; Banse, 2002). Since then, other approaches have been derived, such as the use of tracer methods based on stable isotope additions, such as 13C (Slawyk et al., 1977) and 18O (Bender et al., 1987), as well as a suite of incubation-free techniques, including the use of active fluorescence, FRRF (fast repetition rate fluorometry) (Kolber and Falkowski, 1993), the non-intrusive bio-optical (OPT) method (Loisel and Morel, 1998; Claustre et al., 1999), the triple oxygen isotopes method (Luz and Barkan, 2000) or the analysis of oxygen records from buoys and gliders (Nicholson et al., 2008). These methods differ, however, in assumptions and the particular process through which primary production is represented and thus yield different results when applied to any one community (Marra, 2002). Indeed, resolving estimates derived from different methods can be a challenging exercise and the differences in estimates among various methods continue to fuel discussion (Quay et al., 2010; Marra, 2012). Awareness of the fundamental differences in the specific component of primary production addressed by the different methods to assess phytoplankton primary production has generated a demand to derive conversion equations allowing rates derived with different methods to be compared and eventually integrated. An approach to do so is the development of equations using available concurrent, paired measurements of primary production using two or more techniques (Marra, 2002).
Whereas the majority of published studies evaluate the primary production of planktonic community using a single method, a number of reports using two or more techniques have now become available. Different approaches to measure the volumetric primary production have been compared in a series of papers examining the rates delivered by different methods for specific locations, cruises or sampling events (Williams et al., 1996; Marra, 2002; Corno et al., 2005; Juranek and Quay, 2005; Gazeau et al., 2007; González et al., 2008; Robinson et al., 2009). Most of these comparisons revealed differences among methods, varying in magnitude, attributable to differences in the specific components of primary production addressed by each method as well as to their inherent assumptions. However, these comparisons have been limited in scope to date and cannot be used with confidence to allow interconversions across estimates derived with methods. A set of different equations allowing comparisons among primary production estimates derived with different methods would allow the estimation of, for instance, gross primary production (GPP) from satellite-based primary production estimates, which are currently calibrated against the particulate 14C primary production method. This difficulty is not minor, as current estimates of primary production in the global ocean do not necessarily represent either gross or net primary production and cannot be readily reconciled with estimates of terrestrial primary production into estimates of global photosynthesis, introducing uncertainty into our understanding of global carbon and oxygen budgets.
Here we compare estimates of volumetric rates of marine phytoplankton primary production derived concurrently using different methods and describe the scaling among different metrics of phytoplanktonic primary production, providing equations that allow conversion of estimates among different methods. We do so based on a compilation of data on volumetric estimates of primary production rates concurrently estimated with at least two different methods, thereby allowing the comparison among the derived rates. We first provide a summary of the major assumptions, strengths and weaknesses for the various methods, and then compare concurrent estimates derived from different methods to derive conversion equations.
Materials and Methods
We searched the published literature for volumetric estimates of primary production of natural marine plankton communities produced using at least two different methods applied concurrently. The O2:Ar method evaluates the net primary production while in this study, we only use GPP of plankton communities. Although the 14C method estimates values of primary production closer to net primary production than GPP, we considered it essential to include those data to our study due to its widespread use by the scientific community. Most in situ methods (i.e., non-intrusive bio-optical (OPT) method, the triple oxygen isotopes method, and estimates derived from the analysis of oxygen records obtained by gliders surveys or buoys) yield estimates of phytoplanktonic primary production integrating across the mixed layer and variable time scales and were thus not included here as they have only occasionally paired with estimates derived from other methods, therefore precluding quantitative comparisons among estimates derived from these and other methods. The FRRF method is the only in situ method that allows for volumetric estimates to be derived and has been paired with other methods, thereby allowing comparisons and is therefore included in the evaluation presented here.
In this paper, we present and compare the following methods: the oxygen evolution in dark and light incubations, the FRRF method and methods based in tracer additions (18O, 14C, and 13C).
An Overview of Methods to Measure Marine Phytoplanktonic Primary Production
During photosynthesis, carbohydrates are synthesized and O2 produced from CO2 and H2O, respectively, using solar radiation as the energy source. Approaches to measure primary production include in vitro methods based on either oxygen production or inorganic carbon incorporation and methods based on the analysis of oxygen and gas field in situ. In vitro oxygen-based estimates of primary production involve the evaluation of the release of oxygen during photosynthesis from change in the bulk O2 concentration, corrected for respiratory O2 consumption of communities enclosed in bottles (i.e., dark-light method), or that of 18O2 when 18O-labeled H2O was added (i.e., 18O method). In vitro carbon-based estimates of primary production use tracer additions as 14C or 13C labeled bicarbonate followed by the measurement of the assimilation of the tracer onto organic carbon (particulate or total) following incubation of communities enclosed in bottles for a variable time in the light (i.e., 14C and 13C methods). In situ methods involve the evaluation of oxygen fields retrieved from time series of oxygen derived from moored sensors (Dickey, 1991) or sensors mounted in gliders (Nicholson et al., 2008), corrected for atmospheric exchange, or the synoptic evaluation of triple oxygen isotopes (16O2, 17O2, 18O2) to extract GPP from the anomalies relative to the values expected from atmospheric equilibrium.
The in vitro methods are prone to errors due to the confinement which may affect the organisms involved (e.g., excluding zooplankton or enhancing trophic interactions within the bottle) or the environmental conditions (e.g., temperature, light and nutrient fields), which are not affected by the in situ methods (Maske and Garcia-Mendoza, 1994; Karl et al., 1998; Robinson and Williams, 2005; Duarte et al., 2013; Williams et al., 2013). For instance, modifications of the light field (Kirk, 1994) are observed as the incubations have been conducted with borosilicate bottles, a material that excludes UVB radiation. Indeed, recent analyses comparing estimates derived using quartz and glass bottles have shown that estimates of net community production are affected by the removal of the ambient UVB radiation when glass or plastic materials that filter out UV-B radiation are used, as has been the case for most analyses (Godoy et al., 2012; Agustí et al., 2014; Regaudie-de-Gioux et al., 2014). Furthermore, it is rare for in vitro incubations to be carried out at the correct corresponding environmental temperature. Considering that temperature is a fundamental parameter for respiration rates, the oxygen-based primary production may be over- or underestimated if the incubation temperatures differs from that in situ. Moreover, incubation periods for in vitro methods range from a few hours (0.5–6 h) for 14C or 13C methods to 24 h for the dark-light method. The different incubation time may bias the estimation of PP and the comparison between the different in vitro methods. In contrast, in situ methods are very sensitive to assumptions regarding mixing within the mixed layer and exchanges of oxygen with the atmosphere and the waters below the mixed layer (Duarte et al., 2013).
In this study, we compared estimates of volumetric primary production rates and considered therefore only estimates derived from the dark-light method, the FRRF method and methods based in tracer additions (18O, 14C, and 13C).
The 14C-labeled method (Steeman Nielsen, 1952) consists of measuring the photosynthetic incorporation of 14C labeled inorganic C, added as a NaH14CO3 solution, into particulate and total pools of organic carbon. Whereas the bulk of the measurements available (>90%) report the 14C incorporated into particles retained in filters, as originally proposed Steeman Nielsen (1952), the technique also allows the measurement of total organic carbon (TOC) production [i.e., 14C incorporated into the dissolved organic carbon (DOC), and the particulate organic carbon (POC)], from measurements of the 14C activity in the water sample following removal of the NaH14CO3by purging the sample after acidification. The use of the 14C-labeled method to resolve total organic carbon (TOC) production requires use of high NaH14CO3activity in the sample as to yield a sufficient signal in the water tested. This method has the advantage of allowing to differentiate photosynthetic carbon retained into particulate and dissolved (as total - particulate production) fractions, and allows precise estimates to be derived over short time intervals (Table 1). Furthermore, the high sensitivity of the 14C-labeled method allows the determination of the photosynthesis production in unproductive oceans (Ichimura et al., 1962). However, it is also subject to bottle effects and the underestimation of primary production as it does not include any organic carbon produced that has been respired by the plankton community during the incubation (Table 1). Furthermore, the question of whether the 14C method measures net or gross photosynthesis has been addressed several time in the literature (Ryther, 1954, 1956a; Steemann Nielsen and Al Kholy's, 1956; Peterson, 1980; Dring and Jewson, 1982). The incubation period and nutrient availability may bias the PP estimation. Indeed, Rodhe (1958) and Vollenweider (1969) compared the summation of a series of short 4 h incubations to one long 14C incubation. They reported that the sum of five 4 h incubation exceeded the results of a 20 h incubation by 9–35% depending on depth. They both recommended using short incubations. As described by Morán and Estrada (2002) and other authors (Steeman Nielsen, 1952; Marra, 2002; Halsey et al., 2010; Lasternas and Agustí, 2013) a short incubation period should be selected when measuring DOC produced to match a compromise between the times needed to obtain a significant signal in the PP phase, but at the same time, minimize the loss of 14C-labeled DOC due to assimilation by heterotrophic prokaryotes. Short-time incubations are recommended to optimize the measurements and minimize the contribution of trophic-related processes to DOC production. Health concerns about the use of 14C, international regulation and the potential of contamination on ships may be a real issue for the ongoing use of this method. Indeed, radioisotope legislation in some countries restricts (e.g., Spain) or prohibits altogether (e.g., Japan) use of 14C on research vessels.
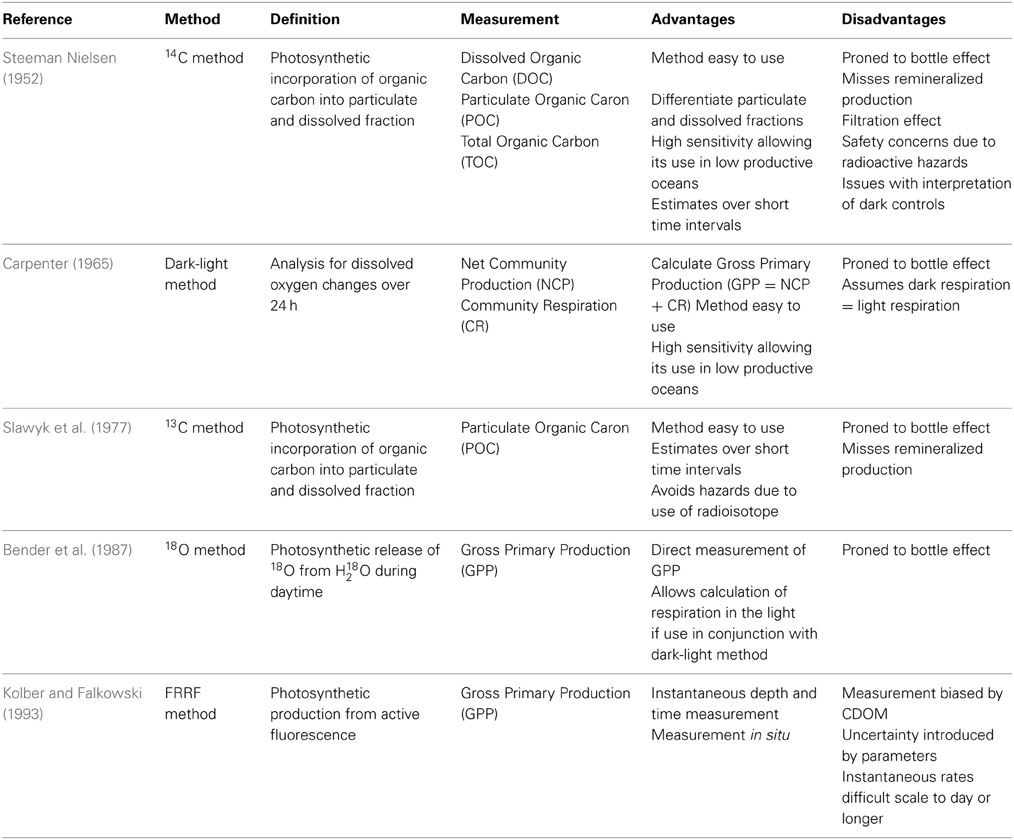
Table 1. Summary of the main characteristics, advantages, and limitations of different approaches measuring volumetric phytoplanktonic primary production.
The 13C-method has also been used to evaluate particulate organic carbon production (Slawyk et al., 1977). This method is similar to the 14C-labeled method except that bicarbonate is labeled with 13C rather than 14C. The main advantages compared to the 14C-labeled method are that the 13C-method allows primary production incorporated by different components of the food web to be resolved using compound-specific analyses (Boschker and Middelburg, 2002), and that 13C is a natural stable carbon isotope, which does not involve any risks to the operator.
The dark-light (or bulk oxygen evolution) method has been used to assess primary production for nearly one century (Gaarder and Gran, 1927). The dark-light method consists of the evaluation of changes in oxygen concentration using high-precision Winkler method, allowing 0.1% precision in oxygen determinations (Carpenter, 1965; Carrit and Carpenter, 1966), following the incubation, typically for 24 h, of natural plankton communities enclosed in clear and dark bottles. Primary production is calculated as the sum of the rate of change in oxygen concentration in clear bottles, the net community production, and that in dark bottles, the dark respiration. This estimate, which is calculated rather than derived directly, is used as a metric of GPP, defined as the total photosynthetic oxygen production prior to any losses, but relies on the assumption that respiration in the dark does not differ from that in the light (Table 1). Indeed, Grande et al. (1989) showed that natural populations show equal respiration rates during day and night. Furthermore, Marra and Barber (2004) assumed that virtually all CO2 respired during the day is re-fixed during the photosynthesis and concluded that twice the dark loss of carbon equals the 24 h rate of phytoplankton respiration. However, there is evidence that respiratory losses are often enhanced in the light (Harris and Lott, 1973), so that the dark-light method is likely to underestimate GPP.
The 18O method measures the GPP using the stable isotope 18O as a tracer of molecular oxygen production through photosynthesis (Bender et al., 1987). The sample water is enriched in 18O derived by the photosynthetic release of 18O from added H218O (Table 1), and thus provides an estimate of GPP free of assumptions on the effect of light on respiration, but still subject to the potential bottle effects indicated above (Table 1). The error in GPP estimates derived from this method is considered lower than 2% (Bender et al., 1999). One advantage of this method is that considering the daily turnover time of phytoplankton, recycling of labeled O2 will be weak (2%) during 24 h in comparison with that of labeled PO14C, which can be very large (Bender et al., 1999). A second advantage of the 18O method is that it can measure directly the gross photosynthetic O2 production in comparison with the dark-light or 14C method that measure the NCP and CR or TOC and POC production, respectively. Lastly, there are no health hazards associated with using 18O because it is a stable isotope. One disadvantage of this method is that gross oxygen production determined by the 18O method evaluates total oxygen production and it is unclear if this is directly linked to carbon assimilation (Bender et al., 1999; Robinson et al., 2009). Indeed, four different metabolic pathways mainly involve oxygen consumption in the light, respiration through the cytochrome oxidase pathway, respiration by the alternative oxidase pathway, photorespiration and the Mehler reaction (Robinson et al., 2009). Although the GPP measured by the 18O method could be corrected by a factor of 20% for the Mehler reaction and photorespiration, it is expected that the GPP would be overestimated by a factor up to 20–50% (Laws et al., 2000; Hendricks et al., 2004) when it is converted into carbon units using a photosynthetic quotient. The 18O method is superior in assessing GPP to all other in vitro methods (Marra, 2002), but it does account for a small fraction of all in vitro measurements available.
The FRRF method measures phytoplanktonic production from active chlorophyll fluorescence (Kolber and Falkowski, 1993). This method evaluates the instantaneous depth and time dependent value of primary production. Indeed, it resolves primary production at spatial (<1 m) and temporal (~1 s) resolutions that cannot be achieved by in vitro approaches (Robinson et al., 2009). The FRRF method has the potential to quantify rapid changes in productivity and make instantaneous measurements of certain physiological parameters (Sakshaug et al., 1997). It also provides a better signal-to-noise ratio and allows more robust measurements in oligotrophic ecosystems. However, assumptions and uncertainties have been reported in the FRRF method (Suggett et al., 2001, 2004; Moore et al., 2003). The maximum light utilization efficiency (∝*) can be overestimated by the FRRF method due to the decoupling of the electron transport rate (ETR) by the cyclic electron flow around the photosystem II (PSII) (Falkowski et al., 1986; Prášil et al., 1996), photorespiration (Raven and Johnston, 1991) and the Mehler reaction (Kana, 1992). Furthermore, Suggett et al. (2001, 2004) reported that uncertainties remain in the estimation of the photosynthetic unit size of the photosystem II (PSURCII), in the assumption of equal distribution of excitation energy between RCI and RCII within the wavelength of the FRRF light source, in the evaluation of the fraction of photochemically active RCIIs from 1.8/(Fv/Fa), in the measurements of the absorption of light by photosynthetic pigments, and in the use of the assumed values of the ratio of PSII reaction centers to the chlorophyll a concentration (nPSII) for prokaryotes and eukaryotes. The FRRF method may be prone to errors when the measured sample contains Colored Dissolved Organic Matter (CDOM) that can affect the spectral absorption and so the accurate in situ measurement of active fluorescence. In addition, the quantification of primary production in terms of carbon involves significant uncertainties as the use of appropriate controls remains a challenge and the method is prone to a number of sources of bias (Laney, 2003).
Comparing Across Primary Production Estimates
Our search of the literature yielded 19 studies measuring primary production concurrently using 14C and dark-light methods (Table 2) including 188 different stations and 692 individual primary production estimates. Twelve studies measured primary production concurrently using 14C and 18O methods including 65 different stations and 367 individual production estimates (Table 2). Six studies reported estimates derived using both the 18O and dark-light method including 45 stations and 232 individual rate estimates. Two reports determined the primary production using FRRF method with 14C (4 stations and 70 individual estimates) and only one with dark-light and 18O methods concurrently including 3 stations and 15 individual production estimates (Table 2). Finally, only one report presented primary production measured concurrently by 14C and 13C methods including 198 primary production estimates (Table 2).
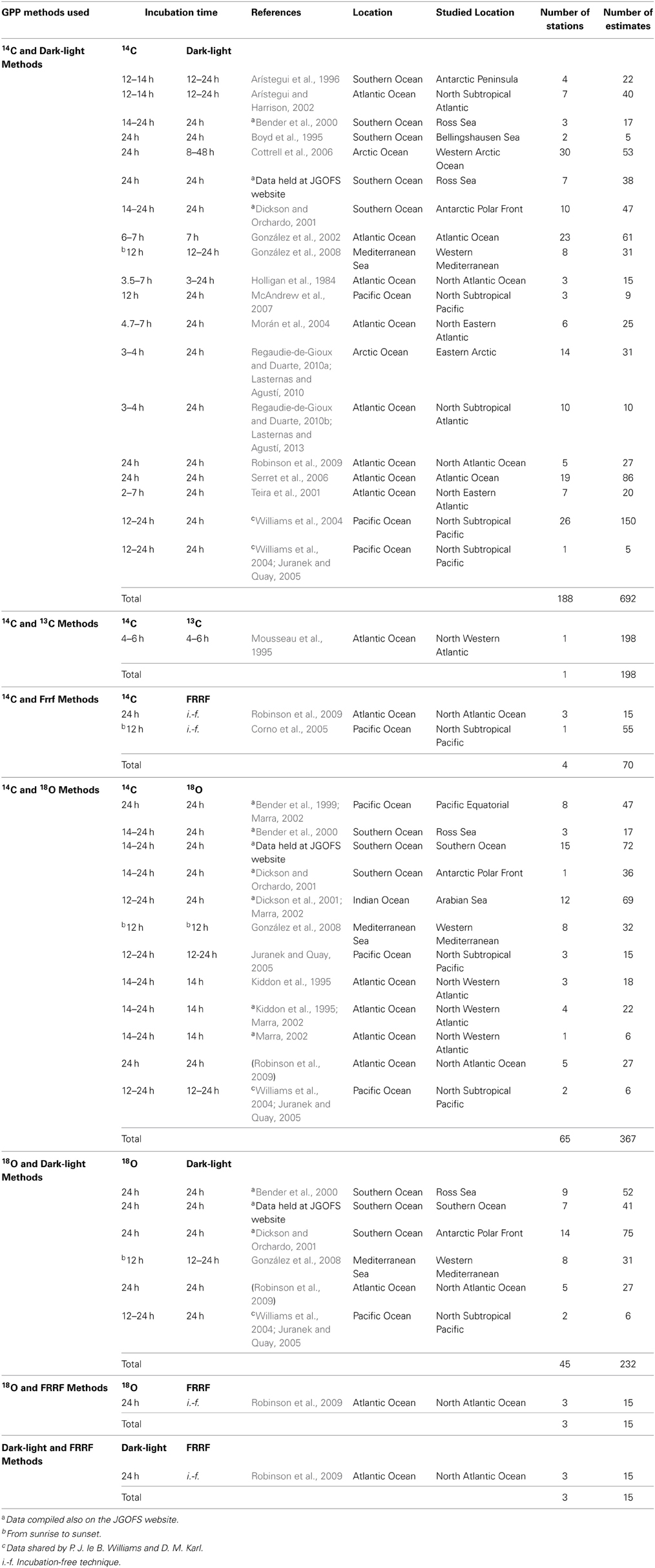
Table 2. Incubation period, references, description of the location, number of stations and of estimates of the different studies analyzed here measuring GPP rate by two different methods.
The compiled 14C primary production data (14C-TOC, 14C-POC, or 14C-DOC) have been estimated according to different cited reports from 3 to 24 h (from dawn to dawn) of incubation (Table 2). In general, the phytoplankton community receives 12 h of sunlight per day. When the primary production has been measured after 2–4 h of incubation, we estimated the hourly rate and then scaled to 12 h of light. We considered that the primary production estimated after 12 or 24 h of incubation received the same amount of light (12 h) and so, are comparable.
The relationship between paired primary production estimates derived using different methods (x and y) were described by fitting power equations of the type,
using reduced major axis (RMA) regression analysis on log-transformed data where log transformation was found necessary to address the problem of heteroscadicity affecting the untransformed relationships between variables.
The predictive power of the relationship between paired primary production was tested by evaluating the errors derived from bootstrap analyses. Bootstrapping analysis is a statistical approach for assigning measures of accuracy to sample estimates. In this study, we selected randomly 90% of a paired-method dataset. From those 90% selected, we estimate the relationship between paired primary production estimates. Using the conversion equations derived, we predict the primary production of one type of method from values obtained using another method for the 10% left of the paired-method dataset. Error (as absolute error, mean ± SE prediction error; and relative error, as the mean ± SE percent error) were derived from 10 bootstrapping iterations of the conversion equations obtained.
Results
The geographic distribution of the different stations which the primary production has been evaluated by at least two distinct methods (Figure 1, Table 2) shows that the studies are scattered across the ocean. Indeed, the studies where primary production was evaluated by the 14C and dark-light methods, the 14C and 18O methods or 18O and dark-light methods took place across contrasting oceanic regions such as low productive oceans (Pacific Ocean and the Mediterranean Sea), high productive oceans (Southern and Arctic Ocean) and the Atlantic Ocean, thereby spanning a broad range of communities and environmental conditions. However, the study using 13C and 14C methods concurrently was confined to individual oceanic region (North Western Atlantic Ocean), thereby limiting the ability to develop conversion equations.
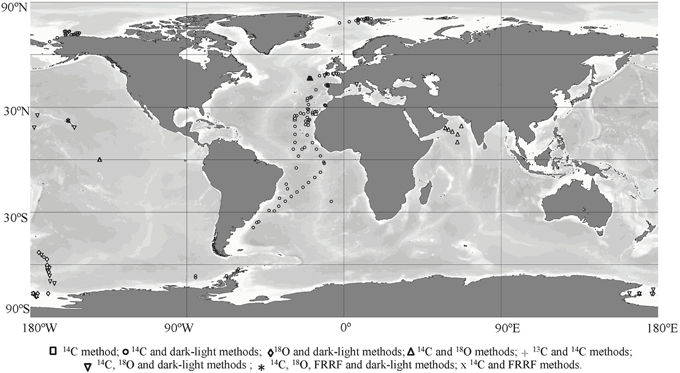
Figure 1. Geographic distribution of the stations where the primary production has been evaluated by two different methods at least.
The ratios between pairs of metrics ranged greatly, typically >10-fold, and up to 200-fold (Figures 2, 3). The ratios presented here are orders of magnitude greater than the ratio observed in laboratory cultures studies (Halsey et al., 2010) and in field studies (Bender et al., 1999; Marra, 2002). This broad range indicates that (a) the relation between the different components resolved by primary production methods and their sources of error are highly variable (Table 1), and (b) the use of simple ratios as conversion factors between two different methods can lead to very large errors. Some of these differences are expected, as the processes resolved by the methods differ. For instance, GPP-DO, GPP-18O, GPP-O2FRRF, and 14C-TOC intend to measure GPP whereas 14C-POC and 13C-POC attempt to resolve specific components of primary production. The comparison between the in vitro metrics indicated that the GPP measured by the 18O method (GPP-18O) tends to produce the highest estimates of primary production, followed by the GPP measured by the dark-light method (GPP-DO), the TOC measured by the 14C method (14C-TOC), the POC measured by the 13C and 14C methods respectively (13C-POC and 14C-POC) (i.e. GPP-18O > GPP-DO > 14C-TOC > 13C-POC > 14C-POC, Figure 2, Table 3).
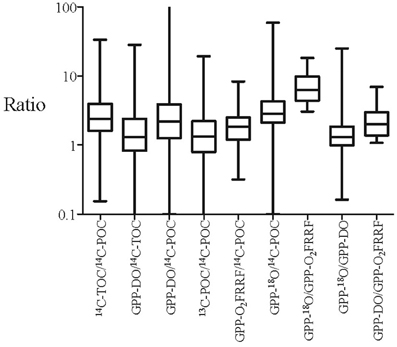
Figure 2. Box plot showing the variation of the primary production ratios measured by two different methods concurrently. The boxes encompass the lower (25%) and upper (75%) quartiles, and the central line shows the median ratio, and the whiskers go down to the smallest value and up to the largest.
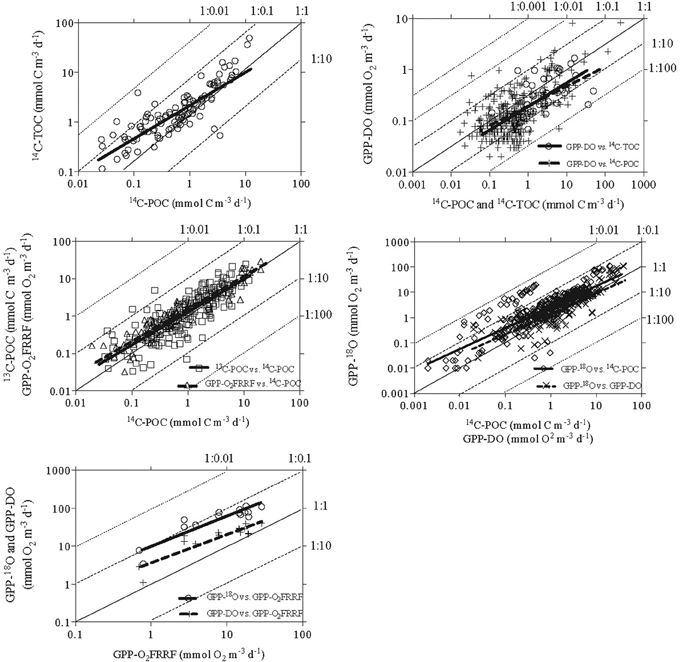
Figure 3. Relationships between the log-log transformed primary production rates measured by different methods concurrently. The bold solid lines represent the linear regressions, the thin solid lines represent the 1:1 lines and the thin dotted lines represent the 1:10, 1:100, 1:0.1, 1:0.01, 1:0.001. Details of linear regressions parameters in Table 4.
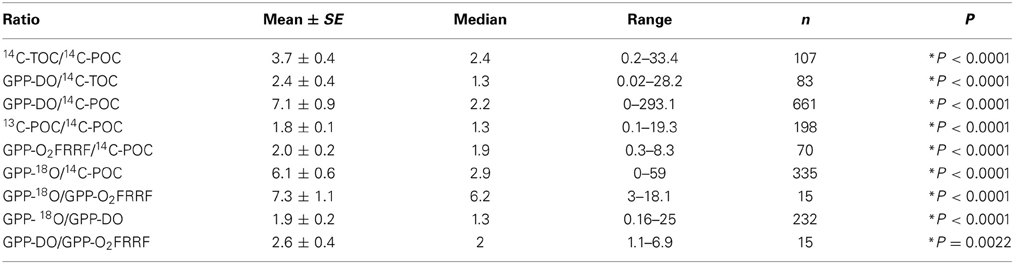
Table 3. Mean (±SE), median, range (minimum–maximum), and number of observations (n) of the primary production ratio between different methods used concurrently, and the probability P, statistics t, and degrees of freedom df of the t-test testing if primary production rates measured by two methods concurrently are significantly different (*represents the significant difference).
The only in situ technique reported used to measure GPP reported here, the FRRF method (GPP-O2FRRF) yielded estimates significantly higher than 14C-POC estimates (t-test, t = 6.7, df = 69, P < 0.0001; Figure 2, Table 3). In contrast, GPP-O2FRRF was 2-times lower than GPP-DO (t-test, t = 3.7, df = 14, P = 0.0022; Figure 2, Table 3) and 7-times lower than GPP-18O (t-test, t = 6, df = 14, P < 0.0001; Figure 2, Table 3). Comparisons among in situ and in vitro methods were limited to a subset of all possible metrics. Indeed, we found two studies reporting the concurrent use of FRRF and 14C methods (Corno et al., 2005; Robinson et al., 2009), and only one study reporting the concurrent use of FRRF, 18O and dark-light to measure GPP (Robinson et al., 2009).
The power equation between estimates of PP derived from different methods allows for ratios between methods shifting with PP (i.e., slope ≠ 1, Figure 3, Table 4). Slopes <1 imply that the ratio of y/x (i.e., a PPy/PPx) declines with increasing x (PPx), and slopes >1 implies that the ratio of y/x increases with increasing x (Table 4). All of these pairwise relationships between methods were significant (P < 0.05) and strong (R2 > 0.35). The error associated with the prediction of primary production in one type of method from values obtained by another method improved the predictive power of those relationships (Table 5).
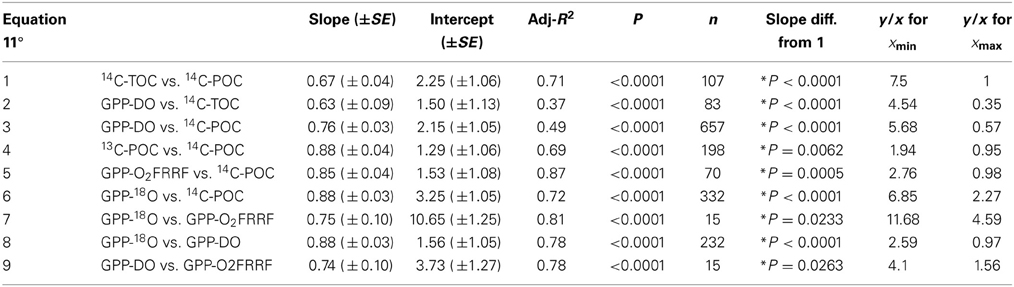
Table 4. Principal component RMA regression equations of the form PPy = aPPbx showing the relationship between the log primary production estimates derived in parallel using different methods, along with the corresponding adjusted coefficient of determination (Adj-R2), the associated probability (P), the probability P of the F-test testing if the slope is different from 1 (* represents the significant difference of the slope from 1), and the y/x (i.e., aPPbx/PPx) ratio for the minimum and the maximum values of x.
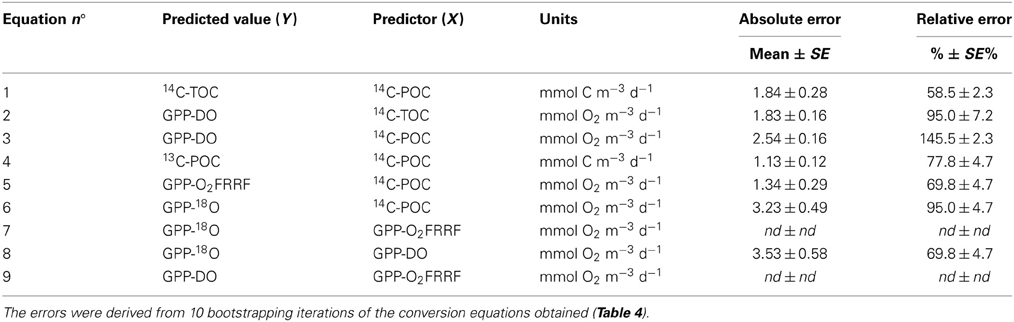
Table 5. Error (absolute, as mean ± SE prediction error; and relative, as the mean ± SE% error) associated with the prediction of primary production in one type of method from values obtained using another method.
The power slopes between the pairs of metrics were statistically different from 1 for most relationships, indicating that the ratios between these metrics changed systematically as primary production increased (Figure 3, Table 4). The departure from 1 indicates that the ratios between estimates derived from various methods change with x, and indeed the ratios predicted from the fitted regression equations varied greatly (Table 4). For instance, the ratio of y/x varied greatly when the minimum or maximum value of x is used (xmin or xmax) ranging about 10-fold across the range of PPx estimates (Table 4). In contrast, the greater departure from a power slope of 1, indicative of a uniform ratio between the metrics, was observed between GPP-DO and 14C-TOC (Table 4).
Discussion
The comparison of estimates of marine phytoplankton primary production derived from different methods provided above reveals very large differences between methods. Some of these differences are expected, as the processes resolved by the methods differ (GPP measurement or specific components of PP measurement) or the methodology itself may be different (i.e., incubation time, temperature, light, or nutrient availability restricted by the incubation process). Indeed, GPP-18O provided the highest estimates of primary production and the 14C-POC provided the smallest ones, as expected, consistent with previous reports. Indeed, Grande et al. (1989) showed that the 18O content of the dissolved oxygen pool increased with photosynthesis and is 2 to 3 times larger than the pool of POC labeled by the 14C. Juranek and Quay (2005) observed that the GPP-18O rates were 1.5–2 times higher than 14C-POC rates. Furthermore, we observed here that GPP-O2FRRF estimates were much lower than the GPP-18O. These results support the observation by Robinson et al. (2009) that the main source of uncertainty in the calculation of GPP-O2FRRF is the use of fixed values for nPSII (the photosynthetic unit size of PSII), which future applications of this method should aim at resolving for natural plankton communities. The 13C method was expected to yield estimates of particulate primary production similar to those of the 14C method, as the only difference is that 13C is a stable isotope. However, our results indicated that the 13C method yields estimates of particulate primary production higher than those of the 14C method (Figure 2, Table 3). Indeed, Mousseau et al. (1995) observed that 70% of PP determined by the 13C method was higher than PP determined by 14C method. Although they were unable to give a complete explanation of the differences between the two methods, they related it to biological and/or environmental conditions (biomass and/or irradiance).
The 18O method should be considered as the best approach to resolve GPP with the greatest precision (here, median 9% error for GPP-18O). Although this method is also robust against changes in respiration over the natural light/dark cycle, it seems to overestimate the gross organic carbon production as it measures all oxygen production without taking into account if it is directly linked to carbon fixation (Laws et al., 2000). Although a correction of 20% for the Mehler reaction and the photorespiration (Laws et al., 2000; Hendricks et al., 2004) could be applied to the GPP-18O, a constant correction factor may not be adequate. Indeed, Steeman Nielsen (1975) argued that photorespiration is correlated with the internal O2 concentration, which is highest at high photosynthetic rates. In this study, GPP-18O rates were higher than 14C-POC rates, the GPP-DO rates and the GPP-O2FRRF measured concurrently (Tables 3, 4, Figures 2, 3). These discrepancies may be explained by the changes in respiration over the natural day–night cycle. Indeed, Grande et al. (1989) indicated that the difference between 14C-POC and GPP-18O rates was due to respiratory 14C losses by both autotrophs and heterotrophs. Furthermore, although some studies reported a lack of differences between light and dark respiration (Marra and Barber, 2004; González et al., 2008), other reports found that light respiration may be higher than dark respiration leading to an underestimation of GPP measured by the dark-light method (Bender et al., 1987; Dickson and Orchardo, 2001; Dickson et al., 2001; Pringault et al., 2007). Moreover, we observed in this study that the difference between production estimates derived using GPP-DO and GPP-18O increases with increasing primary production, as expected. The discrepancy observed here between GPP-18O and GPP-DO rates suggests that the assumption that dark and light respiration is similar maybe questioned. Our results tend to reject the assumption embedded in the dark-light method that dark respiration is equal to light respiration (Table 1), as there is a wealth of indications that respiratory processes are enhanced in the light (Harris and Lott, 1973). In contrast to GPP, NCP estimates should not be influenced by this effect, and the dark-light method should still yield reliable estimates of NCP (Duarte et al., 2013).
Estimates derived using 14C-POC are often indicated to provide a metric close to net primary production, NPP, which is defined as the production available to support phytoplankton growth (i.e., after accounting for losses due to respiration and excretion; Ryther and Vaccaro, 1954; Antia et al., 1963; Eppley and Sharp, 1975). However, 14C-POC represents strictly the production recovered in particulate form after the incubation time, includes the effects of trophic interactions within the incubated sample and does not account for DOC release nor respiratory losses by the community (Bender et al., 1987). Indeed, 14C-POC probably underestimates NPP, as heterotrophic respiration of PP consumed by microzooplankton grazers also affects 14C-POC. The measurement of 14C incorporation into total organic matter partially overcomes this problem by accounting for the 14C recovered in the DOC pool, which can be substantial (González et al., 2008). Yet, this estimate of primary production falls short of accounting for respiratory losses, both by autotrophs and heterotrophs (Bender et al., 1987). Oxygen-based estimates are believed to derive estimates closely approaching GPP (González et al., 2008). Moreover, Ryther (1956b) concluded that respired CO2 could be reutilized into photosynthetic pathways, whereas the O2 released is not similarly consumed in respiration. For that reason, O2 based-methods provide estimates of GPP while 14C-POC provides estimates closer to NPP, relative to that measured by O2 (Marra, 2002). Where 14C needs be used, 14C-TOC, which better approximates GPP, should be measured in parallel to 14C-POC measurements. In this study, 14C-TOC was significantly lower than GPP-DO (Table 3). These differences may be explained by several factors. First, 14C incubation time varied in this study from 2 to 12 h and may result in a 14C-TOC closer to NPP (<4 h of incubation) or to GPP (incubations up to 6 h). Furthermore, some uncertainties regarding the magnitude of extracellular release of newly fixed carbon may explain these differences (González et al., 2008).
We argue that none of the methods tested here resolve NPP with confidence in natural communities. We suggest that the use of NPP to assess primary production by phytoplankton communities should be replaced with a measure of GPP or NCP. Unfortunately, the global primary production of the ocean, a property of interest for multiple applications, has been derived from remote-sensing ocean color calibrated with 14C-POC (Behrenfeld and Falkowski, 1997b). As indicated above it is unclear what these values actually measure and what is, therefore, represented by these global primary production estimates. An option may be to convert these estimates into GPP. However, Peterson (1980) reported that the 14C method underestimated GPP rates by about a factor of 2–100. At the ocean time series HOT, GPP-18O was reported to be 2.4 at surface to 1.1-fold at 100 m greater than 14C-POC, which was interpreted to represent NPP (Nicholson et al., 2012). These observations confirm the results presented here that use of a single conversion factor to estimate GPP from 14C-POC yields biased estimates, as the ratio is highly variable (Table 4). Instead, the conversion equations reported here should be applied to the individual estimates prior to integration at the basin or global scale. Furthermore, several authors assessed the uncertainty in NPP models derived from remote sensing by comparison with in situ 14C uptake and observed a significant systematic bias (Westberry et al., 2008; Friedrichs et al., 2009; Saba et al., 2010, 2011). We were able to evaluate here the integrated PP using the vertically generalized production model (VGPM, Behrenfeld and Falkowski, 1997a) from remote sensing (irradiance and temperature) and in situ data (chlorophyll a) for each oceanographic station of our dataset where in situ integrated 14C-POC rates were available (data not shown here). As expected, we observed that the integrated PPPredicted was very weakly related to the integrated PPObserved (Figure 4, R2 = 0.28, P = 0.03). Milutinović and Bertino (2011) reported that Pb opt (maximum PP per unit of chlorophyll) contributes the most to the random uncertainty in VGPM NPP. Previous results that suggest that remotely sensed estimates of primary production are not precise and until new models are derived and tested widely, the estimates of NPP derived from remote sensing should be considered as approximations only.
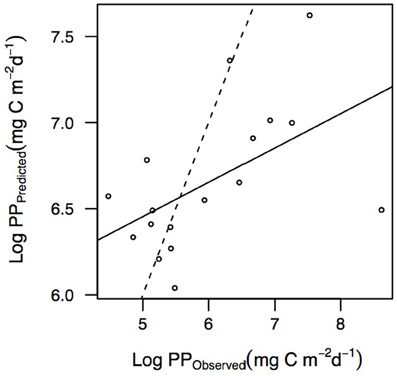
Figure 4. Relationship between the log-log transformed integrated PPPredicted and PPObserved rates (mg C m−2 d−1) for oceanographic stations where integrated 14C-POC was available. The dotted line represents the 1:1 lines and the solid line represents the linear regression with equation: Log PPPredicted = 0.20 (±0.08) log PPObserved + 5.46 (±0.50), R2 = 0.28, P = 0.02775, n = 17.
Following a comparison of various methods, Marra (2002) concluded that there is probably no single method able to provide an absolute estimate of primary production in the ocean and that various methods should be combined in any research programme. However, the scientific community predominantly uses a single method, typically the 14C-POC method, possibly the method with highest uncertainties as to the process actually measured, for the assessment of primary production, and data on primary production estimates derived concurrently with various methods are limited. Moreover, many factors, such as irradiance, temperature, nutrient concentrations, plankton community structure and others may affect the variability in the ratios between estimates derived from different methods. Unfortunately, few studies publish information related to the environmental conditions or community structure so their impact on the ratios between techniques could not be assessed here.
Our study shows that the 18O method provides the most accurate measure of GPP with the fewest assumptions required during estimate of the rate. The remaining flaw may be the presence of bottle effects, which can be partially mitigated using quartz bottles to avoid modifications to the light field (Godoy et al., 2012) and simulating correctly the in situ temperature. While the scientific community embrace a new standard, the regression equations presented here (Table 4) provide the best available approach to convert data across methods, and, therefore, integrate and synthesize available and future data derived using different methods.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This is a contribution to the “Malaspina 2010” CONSOLIDER project funded by the Spanish Ministry of Science and Innovation, the Metaoceans Marie Curie Early Stage Research Network (019678-2), funded by the Framework Program 6 of the EU and the Marine and Coastal Carbon Biogeochemistry Flagship Cluster, funded by the CSIRO. We thank J. Arístegui, N. Cassar, M. Cottrell, D. Kirchman, X. Morán, D. Nicholson, N. González, V. S. S. Sarma, P. Serret, E. Fernández, L. Mousseau, K. Westwood, P. J. le B Williams, and D. M. Karl for providing published data. Aurore Regaudie-de-Gioux was supported by the METAOCEANS project.
References
Agustí, S., Regaudie-de-Gioux, A., Arrieta, J. M., and Duarte, C. M. (2014). Consequences of UV-enhanced community respiration for plankton metabolic balance. Limnol. Oceanogr. 59, 223–232. doi: 10.4319/lo.2014.59.1.0223
Antia, N. J., McAllister, C. D., Parsons, T. R., Stephens, K., and Strickland, J. D. H. (1963). Further measurements of primary production using a large-volume plastic sphere. Limnol. Oceanogr. 8, 166–183. doi: 10.4319/lo.1963.8.2.0166
Arístegui, J., and Harrison, W. G. (2002). Decoupling of primary production and community respiration in the ocean: implications for regional carbon studies. Aquat. Microb. Ecol. 29, 199–209. doi: 10.3354/ame029199
Arístegui, J., Montero, M. F., Ballesteros, S., Basterretxea, G., and van Lenning, K. (1996). Planktonic primary production and microbial respiration measured by 14C assimiliation and dissolved oxygen changes in coastal waters of the Antarctic Peninsula during austral summer: implications for carbon flux studies. Mar. Ecol. Prog. Ser. 132, 191–201.
Banse, K. (2002). Should we continue to measure 14C-uptake by phytoplankton for another 50 years. Limnol. Oceanogr. 11, 45–46.
Behrenfeld, M. J., and Falkowski, P. G. (1997a). A consumer's guide to phytoplankton primary productivity models. Limnol. Oceanogr. 42, 1479–1491.
Behrenfeld, M. J., and Falkowski, P. G. (1997b). Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 42, 1-20.
Bender, M., Orchardo, J., Dickson, M.-L., Barber, R., and Lindley, S. (1999). In vitro O2 fluxes compared with 14C production and other rate terms during the JGOFS Equatorial Pacific experiment. Deep Sea Res. 46, 637–654.
Bender, M. L., Dickson, M.-L., and Orchardo, J. (2000). Net and gross production in the Ross Sea as determined by incubation experiments and dissolved O2 studies. Deep Sea Res. 47, 3141–3158. doi: 10.1016/S0967-0645(00)00062-X
Bender, M. L., Grande, K., Johnson, K., Marra, J., Williams, P. J. le B., Sieburth, J., et al. (1987). A comparison of four methods for determining planktonic community production. Limnol. Oceanogr. 32, 1085–1098. doi: 10.4319/lo.1987.32.5.1085
Boschker, H. T. S., and Middelburg, J. J. (2002). Stable isotopes and biomarkers in microbial ecology. FEMS Microbiol. Ecol. 40, 85–95. doi: 10.1111/j.1574-6941.2002.tb00940.x
Boyd, P. W., Robinson, C., Savidge, G., and Williams, P. J. le B. (1995). Water column and sea-ice primary production during Austral spring in the Bellingshaussen Sea. Deep Sea Res. 42, 1177–1200. doi: 10.1016/0967-0645(95)00070-7
Carpenter, J. H. (1965). The accuracy of the Winkler method for dissolved oxygen analysis. Limnol. Oceanogr. 10, 135–140. doi: 10.4319/lo.1965.10.1.0135
Carrit, D. E., and Carpenter, J. H. (1966). Comparison and evaluation of currently employed modifications of the Winkler method for determining dissolved oxygen in sea-water. J. Mar. Res. 24, 286–318.
Claustre, H., Morel, A., Babin, M., Cailliau, C., Marie, D., Marty, J. C., et al. (1999). Variability in particle attenuation and chlorophyll fluorescence in the Tropical Pacific: scales, patterns, and biogeochemical implications. J. Geophys. Res. 104, 3401–3422. doi: 10.1029/98JC01334
Corno, G., Letelier, R. M., and Abbott, M. R. (2005). Assessing primary production variability in the North Pacific Subtropical gyre: a comparison of fast repetition rate fluorometry and 14C measurements. J. Phycol. 42, 51–60. doi: 10.1111/j.1529-8817.2006.00163.x
Cottrell, M. T., Malmstrom, R. R., Hill, V., Parker, A. E., and Kirchman, D. L. (2006). The metabolic balance between autotrophy and heterotrophy in the western Arctic Ocean. Deep Sea Res. 53, 1831–1844. doi: 10.1016/j.dsr.2006.08.010
del Giorgio, P. A., and Williams, P. J. le B. (2005). Respiration in Aquatic Ecosystems. Oxford: University Press.
Dickey, T. D. (1991). The emergence of concurrent high-resolution physical and bio-optical measurements in the upper ocean and their applications. Rev. Geophys. 29, 383–413. doi: 10.1029/91RG00578
Dickson, M. J., and Orchardo, J. (2001). Oxygen production and respiration in the Antarctic Polar Front region during the austral spring and summer. Deep Sea Res. 48, 4101–4126. doi: 10.1016/S0967-0645(01)00082-0
Dickson, M. J., Orchardo, J., Barber, R. T., Marra, J., McCarthy, J. J., and Sambrotto, R. N. (2001). Production and respiration rates in the Arabian Sea during the 1995 Northeast and Southwest Monsoons. Deep Sea Res. 48, 1199–1230. doi: 10.1016/S0967-0645(00)00136-3
Dring, M. J., and Jewson, D. H. (1982). What does 14C uptake by phytoplankton really measure? A theoretical modelling approach. R. Soc. Publish. 214, 351–368. doi: 10.1098/rspb.1982.0016
Duarte, C. M., Agustí, S., Kennedy, H., and Vaqué, D. (1999). The Mediterranean climate as a template for Mediterranean marine ecosystems: the example of the northeast Spanish littoral. Prog. Oceanogr. 44, 245–270. doi: 10.1016/S0079-6611(99)00028-2
Duarte, C. M., Regaudie-de-Gioux, A., Arrieta, J. M., Delgado-Huertas, A., and Agustí, S. (2013). The oligotrophic ocean is heterotrophic. Ann. Rev. Mar. Sci. 5, 551–569. doi: 10.1146/annurev-marine-121211-172337
Eppley, R. W., and Sharp, J. H. (1975). Photosynthetic measurements in the central North Pacific: the dark loss of carbon in 24-h incubations. Limnol. Oceanogr. 20, 981–987. doi: 10.4319/lo.1975.20.6.0981
Falkowski, P. G., Wyman, K., Ley, A. C., and Mauzerall, D. C. (1986). Relationship of steady state photosynthesis to fluorescence in eukaryotic algae. Biochim. Biophys. Acta 849, 183–192. doi: 10.1016/0005-2728(86)90024-1
Field, C. B., Behrenfeld, M. J., Randerson, J. T., and Falwoski, P. G. (1998). Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240. doi: 10.1126/science.281.5374.237
Friedrichs, M. A. M., Carr, M.-E., Barber, R. T., Scardi, M., Antoine, D., Armstrong, R. A., et al. (2009). Assessing the uncertainties of model estimates of primary productivity in the tropical Pacific Ocean. J. Mar. Syst. 76, 113-133. doi: 10.1016/j.jmarsys.2008.05.010
Gaarder, T., and Gran, H. H. (1927). Investigations of the production of plankton in the Oslo Fjord. Rapp. P. v. Reun. Cons. Perm. Int. Explor. Mer. 42, 1–48.
Gazeau, F., Middelburg, J. J., Loijens, M., Vanderbroght, J.-P., Pizay, M.-D., and Gattuso, J.-P. (2007). Planktonic primary production in estuaries: comparison of 14C, O2 and 18O methods. Aquat. Microb. Ecol. 46, 95–106. doi: 10.3354/ame046095
Godoy, N., Canepa, A., Lasternas, S., Mayol, E., Rúiz-Halpern, S., Agustí, S., et al. (2012). Experimental assessment of the effect of UVB radiation on plankton community metabolism along the Southeastern Pacific off Chile. Biogeosciences 9, 1267–1276. doi: 10.5194/bg-9-1267-2012
González, N., Anadón, R., and Marañón, E. (2002). Large-scale variability of planktonic net community metabolism in the Atlantic Ocean: importance of temporal changes in oligotrophic subtropical waters. Mar. Ecol. Prog. Ser. 233, 21–30. doi: 10.3354/meps233021
González, N., Gattuso, J.-P., and Middelburg, J. J. (2008). Oxygen production and carbon fixation in oligotrophic coastal bays and the relationship with gross and net primary production. Aquat. Microb. Ecol. 52, 119–130. doi: 10.3354/ame01208
Grande, K. D., Williams, P. J. leB., Marra, J., Purdie, D. A., Heinemann, K., Eppley, R. W., et al. (1989). Primary production in the North Pacific gyre: a comparison of rates determined by the 14C, O2 concentration and 18O methods. Deep Sea Res. 36, 1621–1634. doi: 10.1016/0198-0149(89)90063-0
Halsey, K. H., Milligan, A. J., and Behrenfeld, M. J. (2010). Physiological optimization underlies growth rate-independent chlorophyll-specific gross and net primary production. Photosynth. Res. 103, 125–137. doi: 10.1007/s11120-009-9526-z
Harris, G. P., and Lott, J. N. A. (1973). Light intensity and photosynthetic rates in phytoplankton. J. Fish. Res. Board Can. 30, 1771–1778. doi: 10.1139/f73-286
Hendricks, M. B., Bender, M. L., and Barnett, B. A. (2004). Net and gross O2 production in the southern ocean measurements of biological O2 saturation and its triple isotope composition. Deep Sea Res. 51, 1541–1561. doi: 10.1016/j.dsr.2004.06.006
Holligan, P. M., Williams, P. J. leB., Purdie, D., and Harris, R. P. (1984). Photosynthesis, respiration and nitrogen supply of plankton populations in stratified, frontal and tidally mixed shelf waters. Mar. Ecol. Prog. Ser. 17, 201–213.
Ichimura, S., Saijo, Y., and Aruga, Y. (1962). Photosynthetic characteristics of marine phytoplankton and their ecological meaning in the chlorophyll method. Bot Mag. 75, 212–220.
Juranek, L. W., and Quay, P. D. (2005). In vitro and in situ gross primary and net community production in the North Pacific Subtropical Gyre using labelled and natural abundance isotopes of dissolved. Global Biogeochem. Cycles 19, 1–15. doi: 10.1029/2004GB002384
Kana, T. M. (1992). Relationship between photosynthetic oxygen cycling and carbon assimilation in Synechococcus WH7803 (Cyanophyta). J. Phycol. 28, 304–308. doi: 10.1111/j.0022-3646.1992.00304.x
Karl, D. M., Hebel, D. V., Björkman, K., and Letelier, R. M. (1998). The role of dissolved organic matter release in the productivity of the oligotrophic North Pacific Ocean. Limnol. Oceanogr. 43, 1270–1286. doi: 10.4319/lo.1998.43.6.1270
Kiddon, J., Bender, M. L., and Marra, J. (1995). Production and respiration in the 1989 North Atlantic spring bloom: an analysis of irradiance-dependent changes. Deep Sea Res. 42, 553–576. doi: 10.1016/0967-0637(95)00008-T
Kirk, J. T. O. (1994). Light and Photosynthesis in Aquatic Ecosystems, 2nd Edn. Cambridge: Univ. Press. doi: 10.1017/CBO9780511623370
Kolber, Z., and Falkowski, P. G. (1993). Use of active fluorescence to estimate phytoplankton photosynthesis in situ. Limnol. Oceanogr. 38, 1646–1665. doi: 10.4319/lo.1993.38.8.1646
Laney, S. R. (2003). Assessing the error in photosynthetic properties determined by fast repetition rate fluorometry. Limnol. Oceanogr. 48, 2234–2242. doi: 10.4319/lo.2003.48.6.2234
Lasternas, S., and Agustí, S. (2010). Phytoplankton community structure during the record Arctic ice-melting of summer 2007. Polar Biol. 33, 1709–1717. doi: 10.1007/s00300-010-0877-x
Lasternas, S., and Agustí, S. (2013). Bacterial survival governed by organic carbon release from senescent oceanic phytoplankton. Biogeosci. Discuss 10, 16973–16999. doi: 10.5194/bgd-10-16973-2013
Laws, E. A., Landry, M. R., Barber, R. T., Campbell, L., Dickson, M.-L., and Marra, J. (2000). Carbon cycling in primary production bottle incubations: inferences from grazing experiments and photosynthetic studies using 14C and 18O in the Arabian Sea. Deep Sea Res. 47, 1339–1352. doi: 10.1016/S0967-0645(99)00146-0
Loisel, H., and Morel, A. (1998). Light scattering and chlorophyll concentration in case 1 waters: a reexamination. Limnol. Oceanogr. 43, 847-858. doi: 10.4319/lo.1998.43.5.0847
Luz, B., and Barkan, E. (2000). Assessment of oceanic productivity with the triple-isotope composition of dissolved oxygen. Science 288, 2028–2031. doi: 10.1126/science.288.5473.2028
Marra, J. (2002). “Approaches to the measurement of plankton production,” in Phytoplankton Productivity: Carbon Assimilation in Marine and Freshwater Ecosystems, eds P. J. le B. Williams, D. N. Thomas, and C. S. Reynolds (Oxford, UK: Blackwell Science), 78–108.
Marra, J. (2012). Comment on “Measuring primary production rates in the ocean: Enigmatic results between incubation and non-incubation methods at Station ALOHA” by Quay, P. D., et al. Glob. Biogeochem. Cycles 26, GB2031, doi: 10.1029/2011GB004087
Marra, J., and Barber, R. T. (2004). Phytoplankton and heterotrophic respiration in the surface layer of the ocean. Geophys. Res. Lett. 31, 19–24. doi: 10.1029/2004GL019664
Maske, H., and Garcia-Mendoza, E. (1994). Adsorption of dissolved organic matter to the inorganic filter substrate and its implications for 14C uptake measurements. Appl. Environ. Microb. 60, 3887–3889.
McAndrew, P. M., Björkman, K. M., Church, M. J., Morris, P. J., Jachowski, N., Williams, P. J. B., et al. (2007). Metabolic response of oligotrophic plankton communities to deep water nutrient enrichment. Mar. Ecol. Prog. Ser. 332, 63–75. doi: 10.3354/meps332063
Milutinović, S., and Bertino, L. (2011). Assessment and propagation of uncertainties in input terms through an ocean-color-based model of primary production. Remote Sens. Environ. 115, 1906–1917. doi: 10.1016/j.rse.2011.03.013
Moore, C. M., Suggett, D. J., Holligan, P. M., Sharples, J., Abraham, E. R., Lucas, M. I., et al. (2003). Physical controls on phytoplankton physiology and production at a shelf sea front: a fast repetition-rate fluorometer based field study. Mar. Ecol. Prog. Ser. 259, 29–45. doi: 10.3354/meps259029
Morán, X. A., and Estrada, M. (2002). Phytoplanktonic DOC and POC production in the Bransfield and Gerlache Straits as derived from kinetic experiments of 14C incorporation. Deep Sea Res. 49, 769–786. doi: 10.1016/S0967-0645(01)00123-0
Morán, X. A. G., Fernández, E., and Pérez, V. (2004). Size-fractionated primary production, bacterial production and net community production in subtropical and tropical domains of the oligotrophic NE Atlantic in autumn. Mar. Ecol. Prog. Ser. 274, 17–29. doi: 10.3354/meps274017
Mortimer, C. H. (1956). The oxygen content of air-saturated fresh waters, and aids in calculating percentage saturation. Intern. Assoc. Theoret. Appl. Limnol. Commun. 6, 1–20.
Mousseau, L., Dauchez, S., Legendre, L., and Fortier, L. (1995). Photosynthetic carbon uptake by marine phytoplankton: comparison of the stable (13C) and radioactive (14C) isotope methods. J. Plank. Res. 17, 1449–1460. doi: 10.1093/plankt/17.7.1449
Nicholson, D., Emerson, S., and Erisken, C. C. (2008). Net community production in the deep euphotic zone of the subtropical North Pacific gyre from glider surveys. Limnol. Oceanogr. 53, 2226–2236. doi: 10.4319/lo.2008.53.5_part_2.2226
Nicholson, D. P., Stanley, R. H., Barkan, E., Karl, D. M., Luz, B., Quay, P. D., et al. (2012). Evaluating triple oxygen isotope estimates of gross primary production at the Hawaii Ocean Time-series and Bermuda Atlantic Time-series Study sites. J. Geophys. Res. 117, C05012. doi: 10.1029/2010JC006856
Peterson, B. J. (1980). Aquatic primary production and the 14C-CO2 method: a history of the productivity problem. Annu. Rev. Ecol. Syst. 11, 359–385. doi: 10.1146/annurev.es.11.110180.002043
Prášil, O., Kolber, Z., Berry, J. A., and Falkowski, P. G. (1996). Cyclic electron flow around photosystem II in vivo. Photosynth. Res. 48, 395–410. doi: 10.1007/BF00029472
Pringault, O., Tassas, V., and Rochelle-Newall, E. (2007). Consequences of respiration in the light on the determination of production in pelagic systems. Biogeosciences 3, 1367–1389. doi: 10.5194/bgd-3-1367-2006
Quay, P. D., Peacock, C., Björkman, K., and Karl, D. M. (2010). Measuring primary production rates in the ocean:Enigmatic results between incubation and non-incubation methods at Station ALOHA. Glob. Biogeochem. Cycle 24:GB3014. doi: 10.1029/2009GB003665
Raven, J. A., and Johnston, A. M. (1991). Mechanisms of inorganiccarbon acquisition in marine phytoplankton and their implications for the use of other resources. Limnol. Oceanogr. 36, 1701–1714. doi: 10.4319/lo.1991.36.8.1701
Regaudie-de-Gioux, A., Agustí, S., and Duarte, C. M. (2014). UV sensitivity of planktonic net community production in ocean surface waters. J. Geophys. Res. 119, 929–936. doi: 10.1002/2013JG002566
Regaudie-de-Gioux, A., and Duarte, C. M. (2010a). Plankton metabolism in the Greenland Sea during the polar summer of 2007. Polar Biol. 33, 1651–1660. doi: 10.1007/s00300-010-0792-1
Regaudie-de-Gioux, A., and Duarte, C. M. (2010b). The compensation irradiance for planktonic community metabolism in the ocean. Glob. Biogeochem. Cycle. 24:GB4013. doi: 10.1029/2009GB003639
Richards, F. A., and Corwin, N. (1956). Some oceanographic applications of recent determinations of the solubility of oxygen in sea water. Limnol. Oceanogr. 1, 263–267. doi: 10.4319/lo.1956.1.4.0263
Robinson, C., Tilstone, G. H., Rees, A. P., Smyth, T. J., Fishwick, J. R., Tarran, G. A., et al. (2009). Comparison of in vitro and in situ plankton production determinations. Aquat. Microb. Ecol. 54, 13–34. doi: 10.3354/ame01250
Robinson, C., and Williams, P. J. le B. (2005). “Respiration and its measurement in surface marine waters,” in Respiration in Aquatic Ecosystems, eds P. A. del Giorgio and P. J. le B. Williams (Oxford, UK: Oxford University Press), 147–181.
Rodhe, W. (1958). The primary produc-tion in lakes: some results and restric-tions of the 14C method. Rapp. Proc. Verb. Cons. Perm. Int. Expl. Mer. 144, 122–128.
Ryther, J. H. (1954). The ratio of photosynthesis to respiration in marine plankton algae and its effect upon the measurement of productivity. Deep Sea Res. 2, 134–139. doi: 10.1016/0146-6313(55)90015-0
Ryther, J. H. (1956a). Interrelation be-tween photosynthesis and respiration in the marine flagellate, Dunaliella eu-chlora. Nature 178, 861–862.
Ryther, J. H., and Vaccaro, R. F. (1954). A comparison of the oxygen and 14C methods of measuring marine photosynthesis. J. Cons. Int. Expl. Mer. 20, 25–34. doi: 10.1093/icesjms/20.1.25
Saba, V. S., Friedrichs, M. A. M., Carr, M.-E., Antoine, D., Armstrong, R. A., Asanuma, I., et al. (2010). Challenges of modeling depth-integrated marine primary productivity over multiple decades: a case study at BATS and HOT. Glob. Biogeochem. Cycle 24, GB3020. doi: 10.1029/2009GB003655
Saba, V. S., Friedrichs, M. A. M., Antoine, D., Armstrong, R. A., Asanuma, I., Behrenfeld, M. J., et al. (2011). An evaluation of ocean color model estimates of marine primary productivity in coastal and pelagic regions across the globe. Biogeosciences 8, 489–503. doi: 10.5194/bg-8-489-2011
Sakshaug, E., Bricaud, A., Dandonneau, Y., Falkowski, P. G., Kiefer, D. A., Legendre, L., et al. (1997). Parameters of photosynthesis: definitions, theory and interpretation of results. J. Plankton Res. 19, 1637–1670. doi: 10.1093/plankt/19.11.1637
Serret, P., Fernández, E., Robinson, C., Woodward, E. M. S., and Pérez, V. (2006). Local production does not control the balance between plankton photosynthesis and respiration in the open Atlantic Ocean. Deep Sea Res. 53, 1611–1628. doi: 10.1016/j.dsr2.2006.05.017
Slawyk, G., Collos, Y., and Auclair, J.-C. (1977). The use of the 13C and 15N isotopes for the simultaneous measurement of carbon and nitrogen turnover rates in marine phytoplankton. Limnol. Oceanogr. 22, 925–932. doi: 10.4319/lo.1977.22.5.0925
Steeman Nielsen, E. (1952). The use of radioactive carbon (14C) for measuring production in the sea. J. Cons. Perm. Int. Explor. Mer. 18, 117–140. doi: 10.1093/icesjms/18.2.117
Steeman Nielsen, E. (1975). Marine Photosynthesis with Special Emphasis on the Ecological Aspects. Amsterdam: Elsevier.
Steemann Nielsen, E., and Al Kholy, A. A. (1956). Use of 14C technique in measuring photosynthesis of phosphorus or nitrogen deficient algae. Physiol. Plant. 9, 144–145. doi: 10.1111/j.1399-3054.1956.tb08996.x
Suggett, D. J., MacIntyre, H. L., and Geider, R. J. (2004). Evaluation of biophysical and optical determinations of light absorption by photosystem II in phytoplankton. Limnol. Oceanogr. Methods 2, 316–332. doi: 10.4319/lom.2004.2.316
Suggett, D. J., Kraay, G., Holligan, P., Davey, M., Aiken, J., and Geider, R. J. (2001). Assessment of photosynthesis in a spring cyanobacterial bloom by use of a fast repetition rate fluorometer. Limnol. Oceanogr. 46, 802–810. doi: 10.4319/lo.2001.46.4.0802
Teira, E., Pazó, M. J., Serret, P., and Fernández, E. (2001). Disolved organic carbon production by microbial populations in the Atlantic Ocean. Limnol. Oceanogr. 46, 1370–1377. doi: 10.4319/lo.2001.46.6.1370
Truesdale, G. A., Downing, A. L., and Lowden, G. F. (1955). The solubility of oxygen in pure water and sea-water. J. Appl. Chem. Lond. 5, 53–62. doi: 10.1002/jctb.5010050201
Vollenweider, R. A. (1969). A Manual on Methods for Measuring Primary Production in Aquatic Environments. International Biology Program Handbook 12. Oxford: Blackwell Scientific.
Westberry, T., Behrenfeld, M. J., Siegel, D. A., and Boss, B. (2008). Carbon-based primary productivity modelling with vertically resolved photoacclimation. Glob. Biogeochem. Cycle 22, GB2024. doi: 10.1029/2007GB003078
Williams, P. J. B., Morris, P. J., and Karl, D. M. (2004). Net community production and metabolic balance at the oligotrophic ocean site, station ALOHA. Deep Sea Res. 51, 1563–1578. doi: 10.1016/j.dsr.2004.07.001
Williams, P. J. B., Quay, P. D., Westberry, T. K., and Behrenfeld, M. J. (2013). The oligotrophic ocean is autotrophic. Annu. Rev. Mar. Sci. 5, 535–549. doi: 10.1146/annurev-marine-121211-172335
Keywords: marine, phytoplankton, GPP, methods, conversion
Citation: Regaudie-de-Gioux A, Lasternas S, Agustí S and Duarte CM (2014) Comparing marine primary production estimates through different methods and development of conversion equations. Front. Mar. Sci. 1:19. doi: 10.3389/fmars.2014.00019
Received: 19 February 2014; Accepted: 17 June 2014;
Published online: 03 July 2014.
Edited by:
Claire Mahaffey, University of Liverpool, UKReviewed by:
Martin Johnson, University of East Anglia, UKNatalia Gonzalez Benitez, Universidad Rey Juan Carlos, Spain
Copyright © 2014 Regaudie-de-Gioux, Lasternas, Agustí and Duarte. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurore Regaudie-de-Gioux, C/Salud 38 Ático, 07015 Palma de Mallorca, Baleares, Spain e-mail: auroreregaudie@yahoo.fr