
- 1 CBRN Defence and Security, FOI Swedish Defence Research Agency, UmeÅ, Sweden
- 2 UmeÅ Centre for Microbial Research (UCMR), Laboratory for Molecular Infection Medicine Sweden (MIMS), Department of Molecular Biology, UmeÅ University, UmeÅ, Sweden
Francisella tularensis is a highly virulent intracellular human pathogen that is capable of rapid proliferation in the infected host. Mutants affected in intracellular survival and growth are highly attenuated which highlights the importance of the intracellular phase of the infection. Genomic analysis has revealed that Francisella encodes all genes required for expression of functional type IV pili (Tfp), and in this focused review we summarize recent findings regarding this system in the pathogenesis of tularemia. Tfp are dynamic adhesive structures that have been identified as major virulence determinants in several human pathogens, but it is not obvious what role these structures could have in an intracellular pathogen like Francisella. In the human pathogenic strains, genes required for secretion and assembly of Tfp and one pilin, PilA, have shown to be required for full virulence. Importantly, specific genetic differences have been identified between the different Francisella subspecies where in the most pathogenic type A variants all genes are intact while several Tfp genes are pseudogenes in the less pathogenic type B strains. This suggests that there has been a selection for expression of Tfp with different properties in the different subspecies. There is also a possibility that the genetic differences reflect adaptation to different environmental niches of the subspecies and plays a role in transmission of tularemia. This is also in line with recent findings where Tfp pilins are found to be glycosylated which could reflect a role for Tfp in the environment to promote survival and transmission. We are still far from understanding the role of Tfp in virulence and transmission of tularemia, but with the genomic information and genetic tools available we are in a good position to address these issues in the future.
Introduction
Francisella tularensis, the causative agent of tularemia, has attracted significant attention over the years. A major reason is that the most pathogenic variant, subspecies tularensis also known as type A, causes severe infections that without rapid therapeutic intervention shows high mortality rates. These strains have also been recognized to have potential for development of biological weapons. Type A strains are found exclusively in North America while the less pathogenic subspecies holarctica, also known as type B strains, is more broadly distributed in the Northern hemisphere (Petersen and Schriefer, 2005). Still, Francisella remained an understudied pathogen and this did not really change until the first genome sequence became available and genetic systems were developed (Golovliov et al., 2003; Larsson et al., 2005). When the first genome sequence became accessible it was somewhat of a disappointment to note that the number of genes with homology to known virulence determinants in other pathogens were relatively few (Larsson et al., 2005). One of the exceptions was the gene clusters predicted to encode a type IV pili (Tfp) system. Tfp have been identified as a major virulence determinant in many different pathogens even if it was not obvious what role a pilus adhesin could have for an intracellular pathogen like Francisella. In this focused review we summarize and discuss the main findings regarding the biological role of genes encoding the Tfp system in F. tularensis and the significance of the distinct differences seen for specific Tfp genes between different subspecies.
Type IV pili – Dynamic Adhesive Surface Structures
Type IV pili are multifunctional, flexible filamentous appendages that have been assigned specific virulence traits in several important pathogens. These properties include adhesion, twitching motility, biofilm formation, and competence for DNA transformation and are important for host colonization and virulence in pathogens like Pseudomonas aeruginosa, Neisseria spp, Vibrio cholerae, and Moraxella catarrhalis (Mathis and Scocca, 1984; Bergström et al., 1986; Taylor et al., 1987; Sato et al., 1988; Catlin, 1990; Marrs and Weir, 1990; O’Toole and Kolter, 1998). The nomenclature of the Tfp gene clusters have not been harmonized between systems and pathogens (Craig and Li, 2008), and also for F. tularensis different research groups use different nomenclature. Here we have chosen to mainly use the nomenclature adapted for P. aeruginosa. In Table 1 the nomenclature used for the Tfp genes discussed in this review are listed.

Table 1. Nomenclature and presence of functional Tfp genes in different strains.
Type IV pili biogenesis is a process whereby a single protein subunit, the so called major pilin, is processed and translocated across the inner membrane where it forms a dynamic multimeric filament. Multiple proteins sharing structural similarities with this major pilin subunit, known as minor pilins, are also required for proper Tfp function and/or assembly, but their exact role is not completely understood (Alm and Mattick, 1996; Winther-Larsen et al., 2005; Helaine et al., 2007). Tfp are further divided into two subclasses, type IVa and type IVb pilins, based on the presence of specific conserved motifs (Strom and Lory, 1993; Kachlany et al., 2001; Craig et al., 2004). Type IVb pili are commonly found in pathogens colonizing the human intestine like V. cholerae, Salmonella typhi, and enteropathogenic Escherichia coli (EPEC) (Faast et al., 1989; Girón et al., 1991; Zhang et al., 2000).
The major pilin, PilA, is processed by a specific peptidase, PilD, and thereafter translocated across the inner membrane, followed by assembly into a multimeric pilus fiber on the periplasmic side of the inner membrane. The pilus fiber is then secreted across the outer membrane via the secretin pore PilQ (Figure 1; Strom and Lory, 1993; Drake and Koomey, 1995). The assembly and extension of Tfp is facilitated by the PilB ATPase and pilB mutants are negative for Tfp (Turner et al., 1993). In several of the bacteria expressing Tfp a second ATPase PilT promotes disassembly and retraction of Tfp and in this case pilT mutants are hyperpiliated (Wolfgang et al., 2000). PilT is also required for motility on solid surfaces – a phenomenon denoted twitching motility seen in several bacteria expressing PilT (Whitchurch et al., 1991; Maier et al., 2004). Another key component of the Tfp biogenesis is PilC, an inner membrane protein of unknown function (Nunn et al., 1990). Several F. tularensis Tfp related genes also show homology to genes involved in type II secretion system (T2SS) (Peabody et al., 2003). These include the inner membrane associated proteins PilB and PilC, the secretin PilQ, and the pilin peptidase PilD (Nunn and Lory, 1991). In addition, Tfp pilins show homology to T2SS pseudopilins (Peabody et al., 2003). It has been shown that the Tfp subunit PilA of P. aeruginosa is also required for efficient secretion of T2S substrates (Lu et al., 1997). In addition, there is evidence that Tfp in some cases can promote protein secretion by a mechanism similar to T2SS (Kennan et al., 2001; Kirn et al., 2003; Han et al., 2007).
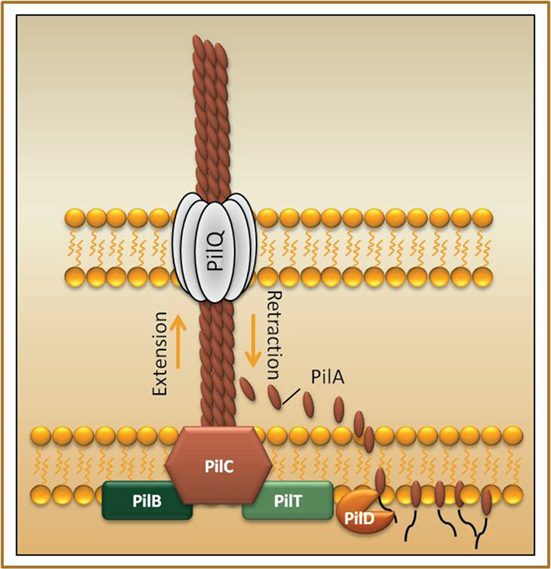
Figure 1. Schematic overview of the type IV pili system. The pilus fiber is mainly composed of the major pilin subunit (denoted PilA in P. aeruginosa), which is expressed as a prepilin that upon cleavage by the prepilin peptidase PilD allows for proper pilus assembly and function (Strom et al., 1993). The major pilin is translocated across the inner membrane where it forms a dynamic multimeric filament that is secreted via the pore-forming secretin PilQ to the bacterial surface (Marceau et al., 1998). PilC is a transmembrane protein found in the inner membrane (Nunn et al., 1990). The two ATPases, PilB and PilT, mediates extension and retraction respectively of the pilus (Whitchurch et al., 1991; Maier et al., 2004).
F. tularensis Subspecies Show Distinct Genetic Differences in Tfp Genes
The genomes of the different F. tularensis subspecies; tularensis (type A), holarctica (type B), and novicida, all encode Tfp clusters including six putative pilin genes; pilA, pilE, pilV, FTT0861, FTT0230, and FTT1314 (Gil et al., 2004; Larsson et al., 2005; Forslund et al., 2006). Interestingly, there are some distinct differences in these pilin genes between the subspecies (Larsson et al., 2005). In virulent type B strains, pilE and pilV are non-functional due to non-sense mutations, and the type IVb pilin gene, FTT0861, harbors a mutation in the stop codon resulting in a longer gene. Furthermore, some type B strains harbor an additional frame-shift mutation in the FTT0861 gene resulting in an even longer open reading frame. The three remaining pilin genes; pilA, FTT0230, and FTT1314, are essentially identical between type A and type B strains. In contrast, all six pilin genes are intact and functional in type A strains and F. novicida. Interestingly, the pilA gene in F. novicida differs in the 3′-end compared to pilA in type A and type B strains (Zogaj et al., 2008). The Tfp genes encoded by the different subspecies are listed in Table 1.
Genomic analysis early revealed the presence of several regions of difference (RDs), flanked with direct repeat sequences that could mediate deletions of certain genes or regions (Broekhuijsen et al., 2003; Svensson et al., 2005). One of these regions, RD19, encodes the pilin gene pilA. Interestingly, several attenuated type B strains, like the live vaccine strain (LVS) and an isolate from a hare, have lost the pilA gene due to homologous recombination involving the direct repeats (Svensson et al., 2005; Forslund et al., 2006; Salomonsson et al., 2009b). Another distinct difference between the subspecies is that there is a unique, single non-sense mutation that truncates the pilT gene in type B strains (Gil et al., 2004). Taken together, all these distinct genetic differences indicate that the biological properties of Tfp expressed by the different subspecies could be significantly different.
Tfp Genes are Required for Virulence of F. tularensis
Among the different pilin genes only pilA has been found to be required for virulence in both type A and type B strains. The above-mentioned hare type B isolate possessing the spontaneous pilA deletion showed reduced virulence at levels comparable to that of the LVS strain, which also lacks pilA and several pilus assembly genes (Table 1; Forslund et al., 2006; Salomonsson et al., 2009b). Furthermore, pilA has been demonstrated to be important for full virulence in the pathogenic type B strain FSC200 and the highly virulent type A strain SCHU S4 (Forslund et al., 2010; Näslund Salomonsson et al., unpublished results). When we compared the importance of pilA for virulence in different strains and subspecies, we found that pilA had less impact on virulence in the more virulent strains (Table 2). As seen in column 3 of Table 2, the infection dose is very low in virulent type A and type B strains, making it problematic to measure small differences in attenuation of strains. Hence, the infection dose experiments are not sensitive enough for the highly virulent strains. Therefore, when using strains that are highly virulent to mice we performed competitive infection experiments where mice were simultaneously infected with the pilA mutant and the isogenic wildtype in order to be able to verify small differences in virulence. The competitive index (CI) was calculated by dividing the ratio of mutant/wt after infection with the mutant/wt ratio before infection. The low CI value, 0.004, for the infection with the type B strain FSC200, is consistent with the result from the single infection experiment where the infection dose was 40-fold higher for mice infected with the pilA negative strain. When studying the highly virulent type A strain and the influence of pilA in virulence, we were unable to measure any difference regarding the infection dose in the single infection study, but the relatively low CI value verifies that pilA is also needed for full virulence in the most pathogenic subspecies (Table 2).
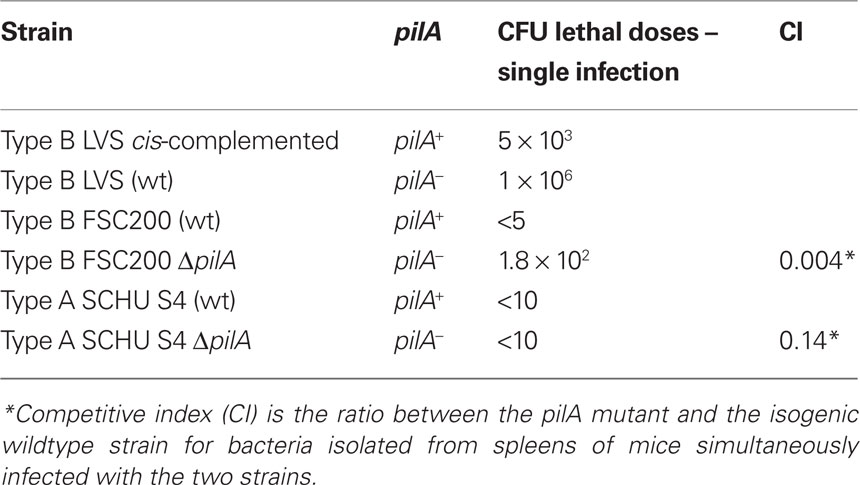
Table 2. Comparison in infection doses and CI values for pilA positive and pilA negative strains.
Regarding F. novicida, the Tfp encoding genes are, overall very similar to type A strains and therefore it is interesting to note that virulence data between these subspecies are so diverse. There are also conflicting results between different F. novicida studies. In one study a F. novicida pilA mutant was found to be even more virulent than the wildtype strain (Hager et al., 2006), while Zogaj et al. (2008) found that pilA mutants were attenuated compared to the wildtype. Hager and colleagues suggested the enhanced virulence to be a result of abolished secretion of the protein PepO, a protease involved in vasoconstriction which limits the spread of F. novicida. Since pepO is non-functional or lacking in type A and type B strains, it is not possible to directly compare the different subspecies regarding these defects. In addition, there are genetic findings indicating that F. novicida displays Tfp-mediated secretion, similar to the type IV toxin coregulated pilus-dependent secretion of TcpF in V. cholerae, rather than a T2SS mediated secretion (Kirn et al., 2003; Hager et al., 2006). In conclusion, there is convincing evidence for a role of a functional Tfp in virulence where most data so far support a role for PilA as well as some of the assembly/secretion factors. The evidence for Tfp-mediated secretion in F. novicida is convincing, while it is somewhat less clear if secretion is common to all subspecies or what the role of functional Tfp-mediated secretion could play in virulence.
So far, FTT0861 is the only pilin besides PilA that has been reported to be virulence associated. Zogaj et al. (2008) showed that FTT0861 was required for full virulence in mice infected with F. novicida via the intradermal route, while the pilin had no influence on intracellular survival and growth in macrophages. Of particular interest is that this pilin belongs to the type IVb pilin family found in enteropathogens like EPEC and V. cholerae. Our lab has not found a link to virulence for any of the other pilin genes in type B strains but we have data suggesting that the pseudopilins/minor pilins encoded by FTT1621 and/or FTT1622 are required for full virulence of type B strains (Näslund Salomonsson et al., unpublished).
Several genes are involved in secretion and assembly of Tfp and there are evidence supporting that some of these genes are also associated with virulence of F. tularensis. The Tfp biogenesis genes pilC and pilQ both contribute to virulence in the highly virulent type A strain SCHU S4 (Forslund et al., 2010). These proteins are of fundamental importance for biogenesis and function of Tfp in other pathogens and provide indirect evidence suggesting that Francisella expresses functional Tfp. Furthermore, PilT has previously been shown to be of importance of virulence in other pathogens, e.g., P. aeruginosa and Dichelobacter nodosus (Comolli et al., 1999; Han et al., 2008). Chakraborty et al. (2008) demonstrated that mutations in either pilB or pilT in LVS resulted in attenuation in a mouse infection model. These results are somewhat puzzling as pilT is a pseudogene in type B strains where there is a missense mutation that would be expected to result in a truncated protein of only one-third in size compared to the full length protein. In an attempt to verify this hypothesis we introduced a FLAG-tag at the 3′-end of both an intact pilT and pilT that harbors the stop codon, and looked for a read-through. The FLAG-tag was expressed from the intact pilT gene but not from the gene carrying the stop codon, suggesting that suppression of this non-sense mutation does not occur and pilT is therefore non-functional in type B strains (Näslund Salomonsson et al., unpublished). The influence of pilT in virulence has also been studied in the type A strain SCHU S4 where pilT is intact, but in this case pilT was found not to be involved in mouse virulence (Forslund et al., 2010).
Even if it has been established that pilA contributes to virulence in F. tularensis, the overall picture is that the pilin is not required for intracellular survival or replication in type A or type B strains (Forslund et al., 2006), with the exception of F. novicida where it was found that pilA mutants were impaired for intracellular growth (Zogaj et al., 2008). The mechanism whereby PilA promotes infection has not been elucidated, but there is evidence suggesting that PilA is required for optimal spread of the bacterium from the initial peripheral site of infection to cause a systemic infection (Forslund et al., 2006). This suggests that PilA mediates specific interactions with host cells/tissue that facilitates spread of the infection.
Physical Evidence for Tfp
In a study published by Gil et al. (2004), LVS was demonstrated to express Tfp-like structures on the surface (Figure 2). In a following study, mutations in either the pilB gene, involved in pilus extension, or the pilT gene, involved in pilus retraction, resulted in loss of surface fibers as well as a defect in adherence to host epithelial cells, arguing that both proteins are involved in pilus assembly (Chakraborty et al., 2008). The pilB results are in agreement with other Tfp expressing pathogens while the pilT results were somewhat unexpected as in other pathogens, like Neisseria spp, deletion of pilT results in a hyperpiliated phenotype (Wolfgang et al., 2000). Interestingly, in Clostridium spp., the only known Gram-positive that harbors Tfp genes, it has been shown that a pilT mutant can still produce pilins, but are unable to assemble the pili on the surface of the cell (Varga et al., 2006). The reason for this unexpected phenotype of the pilT mutant suggests that PilT may have a different role than retraction for Tfp in Francisella. However, the F. tularensis PilT results were even more surprising due to the fact that the pilT gene is only intact in type A strains while it is non-functional in all type B strains (Gil et al., 2004; Näslund Salomonsson et al., unpublished). Overall, the role of PilT in Tfp biogenesis and virulence in different subspecies is still unclear.
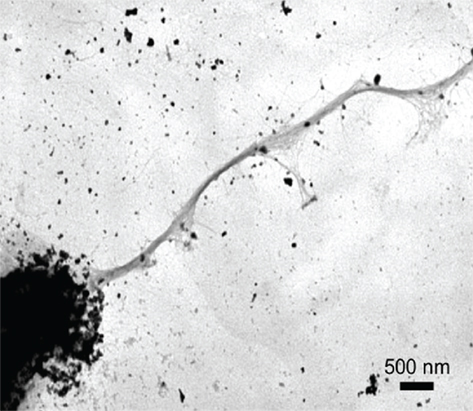
Figure 2. The live vaccine strain was demonstrated to express Tfp-like structures on the bacterial surface with negative stained electron microscopy (Gil et al., 2004). Reprinted with permission. Copyright 2004, ASM press.
In a different study, F. novicida was also confirmed to assemble filamentous structures on the bacterial surface (Zogaj et al., 2008). In this case, a mutation in the secretin-encoding gene pilQ resulted in decreased but not complete lack of pili. Furthermore, a pilC mutant had no impact on piliation. This is again surprising, since both PilQ and PilC are expected to be essential for secretion and assembly of Tfp (Tønjum et al., 1995; Helm et al., 2007). Similar to what was found in LVS, pilB and pilT mutants were totally deficient for Tfp-like structures in F. novicida, indicating that PilB and PilT are required for Tfp assembly. Interestingly, Zogaj et al. (2008) suggested the type IVb pilin FTT0861 to be the major Tfp subunit in F. novicida, as they showed that a FTT0861 mutant lack surface fibers, which was not seen for any of the other pilin mutants. Still, some of the other pilin mutants expressed significantly fewer surface fibers indicating that the mutated genes could encode minor pilins that somehow influence the assembly and expression of the pilus. PilA, on the other hand, was shown to be required for secretion in F. novicida but had no impact on pili expression.
Indeed, even if Tfp-like structures have been identified on the surface of LVS and F. novicida, these structures have still not been verified to be composed of one of the Tfp pilins. In one of our studies the PilA protein was FLAG-tagged in the C-terminus and analyzed with electron microscopy in order to facilitate detection on the surface of type B strains (Forslund et al., 2006). However, in that study no filamentous structures were detected, but the finding that PilA is exported to the bacterial surface was confirmed (Figure 3), though we could not correlate pilA to adhesion to epithelial cells (Forslund et al., 2006). In a further study we used a different strategy to verify if any of the F. tularensis pilin genes could form pili structures. Here a trans-species complementation assay was used where F. tularensis pilins were expressed in N. gonorrhoeae (Salomonsson et al., 2009a). Interestingly, out of all pilins tested only PilA was able to complement Tfp related functions in the heterologous system. Both PilA derived from the type A strain SCHU S4 and from F. novicida promoted assembly of Tfp-like structures in the N. gonorrhoeae background (Figure 4; Salomonsson et al., 2009a). Further support for the ability of these two PilA proteins to form pilus-like multimers was their capacity to restore competence for DNA uptake in a N. gonorrhoeae pilin mutant strain. According to these findings, PilA seems to be the only F. tularensis pilin able to form functional filaments. The type II secretion pseudopilins have also been demonstrated to form pilus-like structures when overexpressed in the T2S pathway (Sauvonnet et al., 2000). Hence, the Tfp-like structures seen in N. gonorrhoeae could be built up by pseudopilins. Importantly, the Francisella specific appendages in N. gonorrhoeae were dependent on Tfp biogenesis factors and pseudopilins does not to support genetic transformation which provides further support for the idea that PilA indeed functions as a Tfp pilin (Salomonsson et al., 2009a).
Figure 3. Expression of FLAG-tagged PilA on the bacterial surface of a type B strain visualized by immunogold electron microscopy (Forslund et al., 2006). Reprinted with permission. Copyright 2006, Blackwell publishing ltd.
Figure 4. Piliation of a gonococcal strain expressing F. novicida derived PilA visualized by immunogold electron microscopy (Salomonsson et al., 2009a). Reprinted with permission. Copyright 2009, Society for General Microbiology.
In conclusion, there are physical evidence for Tfp-like structures in Francisella, still, conclusive evidence as to which protein constitutes the major structural subunit has yet to be presented.
Posttranslational Modification of Tfp
In our first study of Tfp in Francisella, where we showed that loss of pilA also resulted in virulence attenuation, we also presented evidence for posttranslational modification of PilA (Forslund et al., 2006). PilA expressed by Francisella displayed significantly lower mobility in SDS-PAGE compared to PilA expressed by a non- glycosylating strain of P. aeruginosa. Recently we have shown that a gene encoding a glycosyltransferase with homology to transferases known to be required for glycosylation of Tfp in other pathogens (Aas et al., 2007; Faridmoayer et al., 2007; Qutyan et al., 2007), is required for glycosylation of PilA (unpublished results). In addition we have preliminary evidence that other pilins expressed by the different Francisella subspecies also are posttranslationally modified by a modification that requires the same glycosyltransferase (Näslund Salomonsson et al., unpublished). Recently published results indicate that glycosylation of proteins in Francisella is not limited to Tfp related proteins, but includes other surface proteins (Balonova et al., 2010). Genome information, as well as our own preliminary data, suggests that glycosylation of pilins is common to all subspecies (Brotcke et al., 2006; Weiss et al., 2007; Näslund Salomonsson et al., unpublished). It is also possible that the degree of modification at specific glycosylation sites may vary depending on growth conditions which are reflected in the ladder like appearance of PilA in Western blot analysis (Forslund et al., 2006). Posttranslational modification of Tfp has been shown to affect twitching motility and properties of the pilus by rendering the pilus fiber less hydrophobic and more stable (Smedley et al., 2005). For P. aeruginosa there is also a strong correlation between glycosylation of Tfp and clinical isolates, and Tfp glycosylation has been found to provide a competitive advantage in macrophage and lung infection models (Smedley et al., 2005). This opens up the exciting possibility that glycosylation of Tfp may also be of relevance for tularemia either at the level of infection or at the level of transmission.
Prospects
The F. tularensis subspecies show great diversity with respect to virulence in humans from the highly pathogenic subsp. tularensis (type A) to the essentially non-pathogenic subspecies novicida. This is intriguing as the genome sequences have revealed that they are highly homologous with >97% identity at DNA level between the most and least virulent subspecies (Larsson et al., 2009). When it comes to the gene clusters encoding Tfp there are specific differences at genetic level, where type A strains and F. novicida only show significant differences in one gene, pilA, while type B strains have acquired non-sense mutations in several Tfp genes. These mutations are remarkably conserved in the strains analyzed so far, suggesting that loss of function of these genes has occurred in order to adapt to a specific environmental and/or host niche.
One interesting functional difference between the subspecies is that Tfp can promote secretion of a subset of proteins in F. novicida (Hager et al., 2006; Zogaj et al., 2008), but so far there is no evidence for Tfp-mediated secretion of proteins in vitro in the human pathogenic subspecies. While it remains a possibility that the in vitro conditions that promotes secretion may differ, it is still a possibility that evolution of the highly virulent subspecies included loss of Tfp-mediated secretion and that the key molecule in this development is the differences seen in the C-terminal part of PilA between F. novicida and the pathogenic subspecies.
When it comes to functional analysis and virulence, PilA has been found to be required for virulence in the human pathogenic subspecies (Forslund et al., 2006, 2010; Salomonsson et al., 2009b). In addition PilA has been shown to function as a pilin subunit when expressed in a heterologous Tfp expressing system in N. gonorrhoeae, both with respect to forming a Tfp filament but also in the Tfp-mediated DNA uptake assay (Salomonsson et al., 2009a). Several other pilin proteins that were evaluated in this assay were also expressed in N. gonorrhoeae but did not promote formation of filaments or DNA uptake.
PilA has been shown to localize to the bacterial surface and there is also evidence that export/assembly is important for the role of PilA in virulence as mutants in the assembly/secretion genes pilC and pilQ, similar to pilA mutants, were attenuated in a mouse infection model (Forslund et al., 2006, 2010). Several studies have also revealed the presence of Tfp-like structures in various Francisella strains even if no one yet have been able to identify the major subunit in pili expressed by Francisella (Gil et al., 2004; Chakraborty et al., 2008; Zogaj et al., 2008). The only way to validate if different subspecies express different Tfp is to use biochemical or immunological approaches to identify the main component of the Tfp filaments in different strains. However, it is fair to say that Tfp expressed by Francisella appear to be different in many aspects and properties compared to what is known from studies in other pathogens.
Another significant finding regarding the role of PilA in virulence is that the attenuation of pilA mutants in the mouse infection model for tularemia is less pronounced the more virulent the strain is (Forslund et al., 2006, 2010; Salomonsson et al., 2009b). The interpretation and significance of these results is somewhat difficult as all these studies were conducted in the mouse infection model which fails to discriminate between strains that differ significantly in virulence for humans. This makes it difficult to assess if the differences in Tfp genes between type A and type B strains also results in differences in human virulence.
Our recent finding, that PilA and also other pilin proteins are glycosylated, opens other possibilities and raises new questions regarding the role of Tfp in tularemia. Glycosylation is known to influence the properties of Tfp by lowering hydrophobicity, increasing stability and motility. In addition to its role in virulence it is possible that Tfp glycosylation is important for survival and transmission of tularemia.
We are still far from understanding the role of Tfp in virulence and transmission of tularemia, but the identified highly conserved differences strongly suggest that the different subspecies may express Tfp with different properties with respect to filament subunits as well as ability to promote motility. Future work to resolve the role of Tfp in tularemia, need to include both infection models which better reflect the human infection as well as studies on how Tfp genes affect survival and transmission in different natural environments.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aas, F., Vik, A., Vedde, J., Koomey, M., and Egge-Jacobsen, W. (2007). Neisseria gonorrhoeae O-linked pilin glycosylation: functional analyses define both the biosynthetic pathway and glycan structure. Mol. Microbiol. 65, 607–624.
Alm, R., and Mattick, J. (1996). Identification of two genes with prepilin-like leader sequences involved in type 4 fimbrial biogenesis in Pseudomonas aeruginosa. J. Bacteriol. 178, 3809–3817.
Balonova, L., Hernychova, L., Mann, B. F., Link, M., Bilkova, Z., Novotny, M. V., and Stulik, J. (2010). Multimethodological approach to identification of glycoproteins from the proteome of Francisella tularensis, an intracellular microorganism. J. Proteome Res. 9, 1995–2005.
Bergström, S., Robbins, K., Koomey, J. M., and Swanson, J. (1986). Piliation control mechanisms in Neisseria gonorrhoeae. Proc. Natl. Acad. Sci. U.S.A. 83, 3890–3894.
Broekhuijsen, M., Larsson, P., Johansson, A., Byström, M., Eriksson, U., Larsson, E., Prior, R., Sjöstedt, A., Titball, R., and Forsman, M. (2003). Genome-wide DNA microarray analysis of Francisella tularensis strains demonstrates extensive genetic conservation within the species but identifies regions that are unique to the highly virulent F. tularensis subsp. tularensis. J. Clin. Microbiol. 41, 2924–2931.
Brotcke, A., Weiss, D., Kim, C., Chain, P., Malfatti, S., Garcia, E., and Monack, D. (2006). Identification of MglA-regulated genes reveals novel virulence factors in Francisella tularensis. Infect. Immun. 74, 6642–6655.
Catlin, B. W. (1990). Branhamella catarrhalis: an organism gaining respect as a pathogen. Clin. Microbiol. Rev. 3, 293–320.
Chakraborty, S., Monfett, M., Maier, T., Benach, J., Frank, D., and Thanassi, D. (2008). Type IV pili in Francisella tularensis: roles of pilF and pilT in fiber assembly, host cell adherence, and virulence. Infect. Immun. 76, 2852–2861.
Comolli, J. C., Hauser, A. R., Waite, L., Whitchurch, C. B., Mattick, J. S., and Engel, J. N. (1999). Pseudomonas aeruginosa gene products PilT and PilU are required for cytotoxicity in vitro and virulence in a mouse model of acute pneumonia. Infect. Immun. 67, 3625–3630.
Craig, L., and Li, J. (2008). Type IV pili: paradoxes in form and function. Curr. Opin. Struct. Biol. 18, 267–277.
Craig, L., Pique, M., and Tainer, J. (2004). Type IV pilus structure and bacterial pathogenicity. Nat. Rev. Microbiol. 2, 363–378.
Drake, S., and Koomey, M. (1995). The product of the pilQ gene is essential for the biogenesis of type IV pili in Neisseria gonorrhoeae. Mol. Microbiol. 18, 975–986.
Faast, R., Ogierman, M., Stroeher, U., and Manning, P. (1989). Nucleotide sequence of the structural gene, tcpA, for a major pilin subunit of Vibrio cholerae. Gene 85, 227–231.
Faridmoayer, A., Fentabil, M. A., Mills, D. C., Klassen, J. S., and Feldman, M. F. (2007). Functional characterization of bacterial oligosaccharyltransferases involved in O-linked protein glycosylation. J. Bacteriol. 189, 8088–8098.
Forslund, A., Kuoppa, K., Svensson, K., Salomonsson, E., Johansson, A., Byström, M., Oyston, P., Michell, S., Titball, R., Noppa, L., Frithz-Lindsten, E., Forsman, M., and Forsberg, A. (2006). Direct repeat-mediated deletion of a type IV pilin gene results in major virulence attenuation of Francisella tularensis. Mol. Microbiol. 59, 1818–1830.
Forslund, A., Salomonsson, E., Golovliov, I., Kuoppa, K., Michell, S., Titball, R., Oyston, P., Noppa, L., Sjöstedt, A., and Forsberg, A. (2010). The type IV pilin, PilA, is required for full virulence of Francisella tularensis subspecies tularensis. BMC Microbiol. 10, 227. doi: 10.1186/1471-2180-10-227
Gil, H., Benach, J., and Thanassi, D. (2004). Presence of pili on the surface of Francisella tularensis. Infect. Immun. 72, 3042–3047.
Girón, J., Ho, A., and Schoolnik, G. (1991). An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 254, 710–713.
Golovliov, I., Sjöstedt, A., Mokrievich, A., and Pavlov, V. (2003). A method for allelic replacement in Francisella tularensis. FEMS Microbiol. Lett. 222, 273–280.
Hager, A., Bolton, D., Pelletier, M., Brittnacher, M., Gallagher, L., Kaul, R., Skerrett, S., Miller, S., and Guina, T. (2006). Type IV pili-mediated secretion modulates Francisella virulence. Mol. Microbiol. 62, 227–237.
Han, X., Kennan, R., Parker, D., Davies, J., and Rood, J. (2007). Type IV fimbrial biogenesis is required for protease secretion and natural transformation in Dichelobacter nodosus. J. Bacteriol. 189, 5022–5033.
Han, X., Kennan, R. M., Davies, J. K., Reddacliff, L. A., Dhungyel, O. P., Whittington, R. J., Turnbull, L., Whitchurch, C. B., and Rood, J. I. (2008). Twitching motility is essential for virulence in Dichelobacter nodosus. J. Bacteriol. 190, 3323–3335.
Helaine, S., Dyer, D., Nassif, X., Pelicic, V., and Forest, K. (2007). 3D structure/function analysis of PilX reveals how minor pilins can modulate the virulence properties of type IV pili. Proc. Natl. Acad. Sci. U.S.A. 104, 15888–15893.
Helm, R. A., Barnhart, M. M., and Seifert, H. S. (2007). pilQ Missense mutations have diverse effects on PilQ multimer formation, piliation, and pilus function in Neisseria gonorrhoeae. J. Bacteriol. 189, 3198–3207.
Kachlany, S. C., Planet, P. J., Desalle, R., Fine, D. H., Figurski, D. H., and Kaplan, J. B. (2001). flp-1, the first representative of a new pilin gene subfamily, is required for non-specific adherence of Actinobacillus actinomycetemcomitans. Mol. Microbiol. 40, 542–554.
Kennan, R. M., Dhungyel, O. P., Whittington, R. J., Egerton, J. R., and Rood, J. I. (2001). The type IV fimbrial subunit gene (fimA) of Dichelobacter nodosus is essential for virulence, protease secretion, and natural competence. J. Bacteriol. 183, 4451–4458.
Kirn, T., Bose, N., and Taylor, R. (2003). Secretion of a soluble colonization factor by the TCP type 4 pilus biogenesis pathway in Vibrio cholerae. Mol. Microbiol. 49, 81–92.
Larsson, P., Elfsmark, D., Svensson, K., Wikström, P., Forsman, M., Brettin, T., Keim, P., and Johansson, A. (2009). Molecular evolutionary consequences of niche restriction in Francisella tularensis, a facultative intracellular pathogen. PLoS Pathog. 5, e1000472. doi: 10.1371/journal.ppat.1000472
Larsson, P., Oyston, P., Chain, P., Chu, M., Duffield, M., Fuxelius, H., Garcia, E., Hälltorp, G., Johansson, D., Isherwood, K., Karp, P., Larsson, E., Liu, Y., Michell, S., Prior, J., Prior, R., Malfatti, S., Sjöstedt, A., Svensson, K., Thompson, N., Vergez, L., Wagg, J., Wren, B., Lindler, L., Andersson, S., Forsman, M., and Titball, R. (2005). The complete genome sequence of Francisella tularensis, the causative agent of tularemia. Nat. Genet. 37, 153–159.
Lu, H. M., Motley, S. T., and Lory, S. (1997). Interactions of the components of the general secretion pathway: role of Pseudomonas aeruginosa type IV pilin subunits in complex formation and extracellular protein secretion. Mol. Microbiol. 25, 247–259.
Maier, B., Koomey, M., and Sheetz, M. (2004). A force-dependent switch reverses type IV pilus retraction. Proc. Natl. Acad. Sci. U.S.A. 101, 10961–10966.
Marceau, M., Forest, K., Béretti, J., Tainer, J., and Nassif, X. (1998). Consequences of the loss of O-linked glycosylation of meningococcal type IV pilin on piliation and pilus-mediated adhesion. Mol. Microbiol. 27, 705–715.
Mathis, L. S., and Scocca, J. J. (1984). On the role of pili in transformation of Neisseria gonorrhoeae. J. Gen. Microbiol. 130, 3165–3173.
Nunn, D., Bergman, S., and Lory, S. (1990). Products of three accessory genes, pilB, pilC, and pilD, are required for biogenesis of Pseudomonas aeruginosa pili. J. Bacteriol. 172, 2911–2919.
Nunn, D., and Lory, S. (1991). Product of the Pseudomonas aeruginosa gene pilD is a prepilin leader peptidase. Proc. Natl. Acad. Sci. U.S.A. 88, 3281–3285.
O’Toole, G., and Kolter, R. (1998). Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 30, 295–304.
Peabody, C., Chung, Y., Yen, M., Vidal-Ingigliardi, D., Pugsley, A., and Saier, M. J. (2003). Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. Microbiology 149, 3051–3072.
Qutyan, M., Paliotti, M., and Castric, P. (2007). PilO of Pseudomonas aeruginosa 1244: subcellular location and domain assignment. Mol. Microbiol. 66, 1444–1458.
Salomonsson, E., Forsberg, A., Roos, N., Holz, C., Maier, B., Koomey, M., and Winther-Larsen, H. (2009a). Functional analyses of pilin-like proteins from Francisella tularensis: complementation of type IV pilus phenotypes in Neisseria gonorrhoeae. Microbiology 155, 2546–2559.
Salomonsson, E., Kuoppa, K., Forslund, A. L., Zingmark, C., Golovliov, I., Sjostedt, A., Noppa, L., and Forsberg, A. (2009b). Reintroduction of two deleted virulence loci restores full virulence to the live vaccine strain of Francisella tularensis. Infect. Immun. 77, 3424–3431.
Sato, H., Okinaga, K., and Saito, H. (1988). Role of pili in the pathogenesis of Pseudomonas aeruginosa burn infection. Microbiol. Immunol. 32, 131–139.
Sauvonnet, N., Vignon, G., Pugsley, A., and Gounon, P. (2000). Pilus formation and protein secretion by the same machinery in Escherichia coli. EMBO J. 19, 2221–2228.
Smedley, J. G. III, Jewell, E., Roguskie, J., Horzempa, J., Syboldt, A., Stolz, D. B., and Castric, P. (2005). Influence of pilin glycosylation on Pseudomonas aeruginosa 1244 pilus function. Infect. Immun. 73, 7922–7931.
Strom, M., and Lory, S. (1993). Structure-function and biogenesis of the type IV pili. Annu. Rev. Microbiol. 47, 565–596.
Strom, M., Nunn, D., and Lory, S. (1993). A single bifunctional enzyme, PilD, catalyzes cleavage and N-methylation of proteins belonging to the type IV pilin family. Proc. Natl. Acad. Sci. U.S.A. 90, 2404–2408.
Svensson, K., Larsson, P., Johansson, D., Byström, M., Forsman, M., and Johansson, A. (2005). Evolution of subspecies of Francisella tularensis. J. Bacteriol. 187, 3903–3908.
Taylor, R. K., Miller, V. L., Furlong, D. B., and Mekalanos, J. J. (1987). Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc. Natl. Acad. Sci. U.S.A. 84, 2833–2837.
Tønjum, T., Freitag, N., Namork, E., and Koomey, M. (1995). Identification and characterization of pilG, a highly conserved pilus-assembly gene in pathogenic Neisseria. Mol. Microbiol. 16, 451–464.
Turner, L. R., Lara, J. C., Nunn, D. N., and Lory, S. (1993). Mutations in the consensus ATP-binding sites of XcpR and PilB eliminate extracellular protein secretion and pilus biogenesis in Pseudomonas aeruginosa. J. Bacteriol. 175, 4962–4969.
Varga, J., Nguyen, V., O’Brien, D., Rodgers, K., Walker, R., and Melville, S. (2006). Type IV pili-dependent gliding motility in the Gram-positive pathogen Clostridium perfringens and other Clostridia. Mol. Microbiol. 62, 680–694.
Weiss, D., Brotcke, A., Henry, T., Margolis, J., Chan, K., and Monack, D. (2007). In vivo negative selection screen identifies genes required for Francisella virulence. Proc. Natl. Acad. Sci. U.S.A. 104, 6037–6042.
Whitchurch, C., Hobbs, M., Livingston, S., Krishnapillai, V., and Mattick, J. (1991). Characterisation of a Pseudomonas aeruginosa twitching motility gene and evidence for a specialised protein export system widespread in eubacteria. Gene 101, 33–44.
Winther-Larsen, H., Wolfgang, M., Dunham, S., van Putten, J., Dorward, D., Løvold, C., Aas, F., and Koomey, M. (2005). A conserved set of pilin-like molecules controls type IV pilus dynamics and organelle-associated functions in Neisseria gonorrhoeae. Mol. Microbiol. 56, 903–917.
Wolfgang, M., van Putten, J., Hayes, S., Dorward, D., and Koomey, M. (2000). Components and dynamics of fiber formation define a ubiquitous biogenesis pathway for bacterial pili. EMBO J. 19, 6408–6418.
Zhang, X., Tsui, I., Yip, C., Fung, A., Wong, D., Dai, X., Yang, Y., Hackett, J., and Morris, C. (2000). Salmonella enterica serovar typhi uses type IVB pili to enter human intestinal epithelial cells. Infect. Immun. 68, 3067–3073.
Keywords: Francisella tularensis, type IV pili, virulence, type II secretion
Citation: Näslund Salomonsson E, Forslund A-L and Forsberg Å (2011) Type IV pili in Francisella – a virulence trait in an intracellular pathogen. Front. Microbio. 2:29. doi: 10.3389/fmicb.2011.00029
Received: 02 December 2010;
Accepted: 04 February 2011;
Published online: 15 February 2011.
Edited by:
Anders Sjostedt, Umeå University, SwedenReviewed by:
David Thanassi, Stony Brook University, USALisa Craig, Simon Fraser University, Canada
Copyright: © 2011 Näslund Salomonsson, Forslund and Forsberg. This is an open-access article subject to an exclusive license agreement between the authors and Frontiers Media SA, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Åke Forsberg, Department of Molecular Biology, UmeÅ University, 901 87 UmeÅ, Sweden. e-mail: ake.forsberg@molbiol.umu.se