
- Department of Cell Biology, School of Medicine of Ribeirão Preto, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, São Paulo, Brazil
The Nlrc4 inflammasome is triggered in response to contamination of the host cell cytoplasm with bacterial flagellin, which induces pyroptosis, a form of cell death that accounts for restriction of bacterial infections. Although induction of pyroptosis has been extensively investigated in response to Salmonella typhimurium and Legionella pneumophila, little is known regarding the role of the inflammasome for restriction of non-pneumophila Legionella species. Here, we used five species of the Legionella genus to investigate the importance of the inflammasome for restriction of bacterial infection in vivo. By infecting mice deficient for inflammasome components, we demonstrated that caspase-1 and Nlrc4, but not Asc, contribute to restriction of pulmonary infection with L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens. L. longbeachae, a non-flagellated bacterium that fails to trigger pyroptosis, was not restricted by the inflammasome and induced death in the infected mice. In contrast to L. longbeachae, flagellin mutants of L. pneumophila did not induce mice death; therefore, besides bypassing the Nlrc4 inflammasome, L. longbeachae may employ additional virulence strategies to replicate in mammalian hosts. Collectively, our data indicate that the Nlrc4 inflammasome plays an important role in host protection against opportunistic pathogenic bacteria that express flagellin.
Introduction
Innate immune cells employ germ line encoded pattern recognition receptors (PRRs) that are important for recognition of pathogen-associated molecular patterns (PAMPs), conserved structures present in microbes and pathogens (Janeway and Medzhitov, 2002). Extensively studied PRRs are the Toll-like receptors (TLRs), Rig-like receptors (RLRs), and Nod-like receptors (NLRs); this trinity of PRR plays an important role in recognition of bacterial, viruses, fungi, and parasitic infections (Kawai and Akira, 2009; Schroder and Tschopp, 2010). Whereas TLR are transmembrane proteins that signal for expression of inflammatory genes via transcriptional factors, the NLRs are cytosolic proteins that trigger diverse host cell processes.
Activation of NLRs, such as Nod1 and Nod2 turns on signaling cascades that culminate in activation of NF-κB and MAPK. Nod1 and Nod2 signaling requires the protein kinase Rip2, leading to expression of inflammatory genes, such as those of cytokines and chemokines (Franchi et al., 2008). Accordingly, mice deficient for Nod/Rip2 signaling were defective for recruitment of additional phagocytes for the sites of infection (Masumoto et al., 2006; Archer et al., 2010; Berrington et al., 2010; Frutuoso et al., 2010; Silva et al., 2010). Other members of the NLR family do not play an evident role in regulation of gene expression. Instead, they participate in the activation of the pro-caspase-1, which will lead to the formation of a multimeric complex called inflammasome (Schroder and Tschopp, 2010). Several inflammasomes have been described, among them the Nlrp3, Nlrc4, Nlrp1, and AMI2 inflammasomes (Pedra et al., 2009; Schroder and Tschopp, 2010). Nlrp3 inflammasome is possibly the most studied; it requires the adaptor protein Asc and leads to a strong inflammatory response mediated by the secretion of inflammatory cytokines, such as IL-1β and IL-18. The Nlrp3 inflammasome is triggered in response to diverse stimuli including several crystals, skin irritants, microbial PAMPs, microbial toxins, etc. (Schroder and Tschopp, 2010). The diverse range of stimulus that are able to trigger the Nlrp3 inflammasome led to the statement that Nlrp3 is a general sensor of host cell stress.
Regardless of the Nlrp3 inflammasome, the Nlrc4 inflammasome is another platform that also plays important roles for induction of inflammation and restriction of bacterial infection (Sutterwala and Flavell, 2009). The Nlrc4 inflammasome may be composed by another NLR called Naip5/Birc1e and may not require the adaptor protein Asc (Zamboni et al., 2006; Lightfield et al., 2008; Case et al., 2009). The Nlrc4 inflammasome was originally described as important for caspase-1 activation and host response against intracellular bacterial pathogens such as Salmonella enterica serotype Typhimurium (S. typhimurium) and Legionella pneumophila (Mariathasan et al., 2004; Amer et al., 2006; Franchi et al., 2006; Miao et al., 2006; Zamboni et al., 2006). It was shown that flagellin from S. typhimurium and L. pneumophila was required and sufficient to trigger the Nlrc4 inflammasome in macrophages (Franchi et al., 2006; Miao et al., 2006; Lightfield et al., 2008; Silveira and Zamboni, 2010; Whitfield et al., 2010). Furthermore, engagement of the Nlrc4 inflammasome contributed to pore formation, pyroptosis, and restriction of L. pneumophila replication both in macrophages and in vivo (Amer et al., 2006; Zamboni et al., 2006; Lightfield et al., 2008; Case et al., 2009; Miao et al., 2010; Silveira and Zamboni, 2010; Whitfield et al., 2010). Therefore, the Nlrc4 plays important role in recognition, response, and resolution of the infection by flagellated pathogens. Although activation of the Nlrc4 inflammasome has being extensively investigated in response to several bacteria, little is known about the role of the inflammasome in restriction of the infection by opportunistic bacterial pathogens such as non-pneumophila Legionella spp.
Besides L. pneumophila, a number of Legionella species are found parasitizing ameba in freshwater environments, soil, and man-made water distribution system. About 20 Legionella species have being described as human pathogens based in isolation from clinical material (Muder and Yu, 2002; Gobin et al., 2009a). L. pneumophila may account to more than 90% of the human infection by Legionella. However, this estimative may be biased because appropriate diagnostic kits for non-pneumophila species are not often used. It is estimated that non-pneumophila species, such as L. longbeachae, L. micdadei, and L. bozemanii, account for approximately 2–7% of the infections worldwide (Muder and Yu, 2002; Gobin et al., 2009a). Other species such as L. feeleii, L. dumoffii, L. wadsworthii, and L. anisa were not rarely reported, but their frequency may increase with the development of more accurate kits for diagnostic. We and others have previously demonstrated that the flagellin expression is variable among non-pneumophila species of Legionella (Cazalet et al., 2010; Kozak et al., 2010; Silveira and Zamboni, 2010; Whitfield et al., 2010). Whereas species, such as L. longbeachae and L. wadsworthii fail to express flagellin, some species behave similarly to L. pneumophila: are flagellated and trigger Nlrc4-dependent responses in macrophages (Cazalet et al., 2010; Kozak et al., 2010; Silveira and Zamboni, 2010). Herein, we used four flagellated Legionella spp. and the non-flagellated L. longbeachae to assess the role of the inflammasome in restriction of bacterial infection in vivo. We found that Nlrc4, but not Asc, accounts for restriction of in vivo infection by the flagellated bacteria such as L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens. In contrast, L. longbeachae bypasses the Nlrc4 inflammasome-mediated growth restriction and may employ additional virulence strategies to survive in mammalian hosts.
Materials and Methods
Bacterial Strains
Bacterial strains used were L. gratiana (ATCC 49413), L. micdadei (ATCC 33218), L. bozemanii (ATCC 33217), L. rubrilucens (ATCC 35304), and L. longbeachae (ATCC 33462). L. pneumophila used were JR32 and isogenic mutant for the flagellin (flaA) gene (Case et al., 2009). All bacteria grew in buffered charcoal-yeast extract (CYE) agar [1% yeast extract, 1% N-(2 acetamido)-2-aminoethanesulfonic (ACES), pH 6.9, 3.3 mM L-cysteine, 0.33 mM Fe(NO3)3, 1.5% Bacto agar, and 0.2% activated charcoal] at 37°C (Feeley et al., 1979). Before infection, bacteria were resuspended in sterile water and diluted in PBS to an appropriate MOI according to optical density (OD) at 600 nm.
Mice and in vivo Infections
Mice were bred and maintained in the Animal Facility of the University of São Paulo at Ribeirão Preto. C57BL/6 mice were from institutional animal facility and caspase-1−/−, Nlrc4−/−, and Asc−/− mice were previously described (Kuida et al., 1995; Lara-Tejero et al., 2006). Mice deficient for Nlrc4 or Caspase-1 were backcrossed to BL/6 mice for eight (caspase-1 and Nlrc4) or nine (Asc) generations. For in vivo infections, male or female mice from 10 to 14 weeks of age were anesthetized with 2,2,2-tribromoethanol (Sigma) by intraperitoneal administration followed by intranasal (i.n.) inoculation of 40 μl of PBS containing the bacteria at the indicated MOI. Mice were euthanized at the indicated time points for determination of CFU per lung. The lungs were harvested and homogenized in 10 ml of sterile water for 30 s using a tissue homogenizer (Fisher Scientific). Lung homogenates were diluted in water, plated on CYE agar plates and incubated at 37°C for determination of CFU. The limit of the detection of the assay was 102 CFU/lung. Survival rates were determined by daily examination of the infected animals. The mice protocols were previously approved by the institutional ethics committee for animal care and research (CETEA-FMRP/USP).
Statistical analysis
Statistical significance was calculated using the unpaired Student’s t-test or one-way ANOVA, followed by multiple comparisons according to Tukey’s procedure using GraphPad Prism v5 for Windows (GraphPad Software). The Kaplan–Meier method was used to compare survival curves. Differences were considered statistically significant if the P value was <0.05, indicated with an asterisk in the figures.
Results
Caspase-1 Contributes for Restriction of Pulmonary Infection with Legionella spp. that Trigger Pyroptosis
We have previously demonstrated that similarly to L. pneumophila, other Legionella species such as L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens express flagellin and trigger pyroptosis in macrophages (Silveira and Zamboni, 2010). To investigate whether the inflammasome contributes to growth restriction of these species in a murine model of pulmonary infection, we infected C57BL/6 and caspase-1−/− mice with L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens and measured the CFU in the lungs 4, 48, and 96 h after infection. By comparing mice infected for 4, 48, and 96 h, we found that whereas wild-type mice restricted bacterial growth, the caspase-1-deficient mice showed impaired restriction of bacterial replication in the mice lungs (Figure 1).
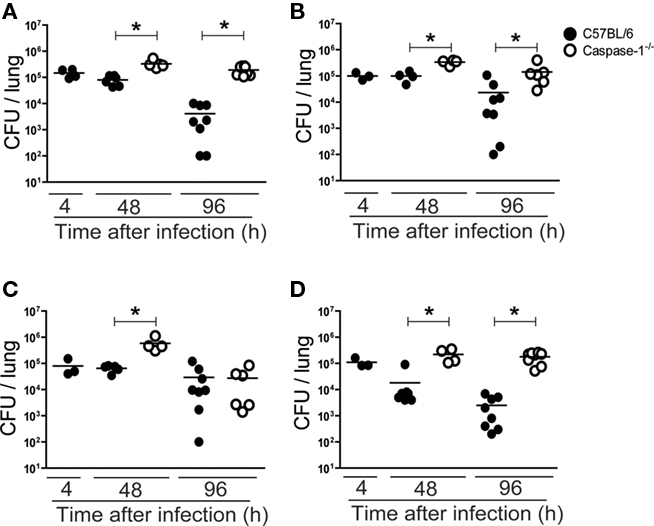
Figure 1. Caspase-1 contributes for restriction of in vivo infection with L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens. C57BL/6 (closed circles) and caspase-1−/− (open circles) mice were infected intranasally with 1.0 × 106 Legionella spp. per mouse. (A) L. micdadei. (B) L. bozemanii. (C) L. gratiana. (D) L. rubrilucens. Mice were sacrificed 4, 48, and 96 h after infection and dilutions of the lung homogenates were added to CYE agar plates for CFU determination. Each dot represents a single animal and the horizontal lines represent the average. Six to 10 mice were used per group. Data are representative of those found in four independent experiments. An asterisk indicates a P value of <0.05.
Nlrc4, but not Asc, Contributes for Growth Restriction of Legionella spp. that Trigger Pyroptosis
To investigate which inflammasome was responsible for caspase-1-dependent restriction of infection with L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens, we infected mice deficient for Nlrc4 or Asc and measured the CFU in the lungs 48 h after infection. We found that, for these four species, mice deficient for Nlrc4 restricted pulmonary infection less effectively than wild-type and mice deficient for Asc (Figure 2). These results indicate that the Nlrc4 inflammasome, but not an inflammasome-dependent on the adaptor protein Asc, contributed for growth restriction of flagellated Legionella species that trigger pyroptosis. Although statistically significant differences were not detected for L. rubrilucens, the data show a trend for increased bacterial numbers in the lungs of Nlrc4−/− mice (Figure 2D).
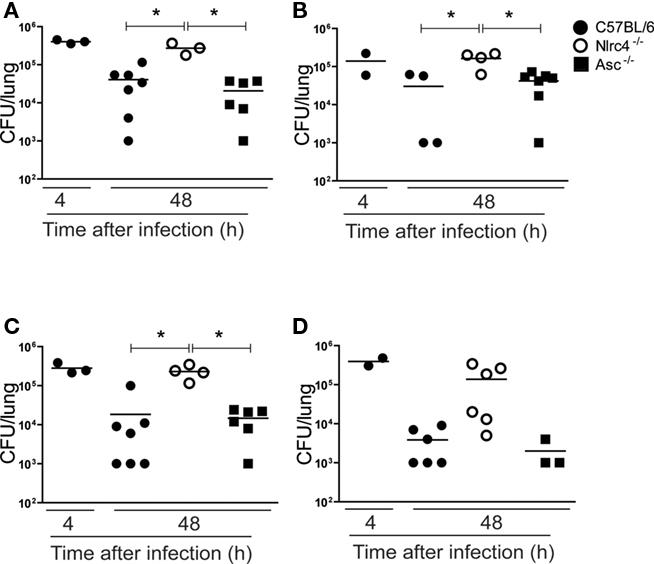
Figure 2. Nlrc4, but not Asc, contributes for restriction of in vivo infection with L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens. C57BL/6 (closed circles), Nlrc4−/− (open circles), and Asc−/− (closed squares) mice were infected intranasally with 1.0 × 106 Legionella spp. per mouse. (A) L. micdadei. (B) L. bozemanii. (C) L. gratiana. (D) L. rubrilucens. Mice were sacrificed 4 or 48 h after infection and dilutions of the lung homogenates were added to CYE agar plates for CFU determination. Each dot represents a single animal and the horizontal lines represent the average. Three to 10 mice were used per group. Data are representative of those found in three independent experiments. An asterisk indicates a P value of <0.05.
The Inflammasome does not Participate in the Restriction of Infection by L. longbeachae in vivo
Next, we investigated whether caspase-1 was important for in vivo restriction of the L. longbeachae replication, a non-flagellated bacteria that lacks genes important for flagellum biosynthesis (Cazalet et al., 2010; Kozak et al., 2010). To test if the inflammasome participated in the control of L. longbeachae infection in vivo, wild-type mice and those deficient for caspase-1 were infected for 48 h and the bacterial CFU in the lungs were counted. We found that caspase-1 deficiency did not influence bacterial replication in the lungs (Figure 3A). Furthermore, L. longbeachae was able to effectively multiply in the lungs of mice from 4 to 48 h of infection (Figure 3A). This information is consistent with previous reports showing that L. longbeachae is lethal for mice (Asare et al., 2007; Gobin et al., 2009b). To further evaluate the role of caspase-1 in mouse resistance, we performed infections in wild-type and caspase-1−/− mice with different amounts of L. longbeachae and evaluated the survival of the infected animals. We found that infections with 105 bacteria per mice is sub-lethal and induce mice death at variable rates (from 0 to approximately 60%). In contrast, infections with 106 or 107 L. longbeachae per mice induced death in 100% of the mice in less than 10 days after infection (Figures 3B–D). Importantly, in all the MOI used, caspase-1−/− mice died similarly to the wild-type mice, indicating that caspase-1 may not play an important role for host resistance against L. longbeachae infection (Figures 3B–D).
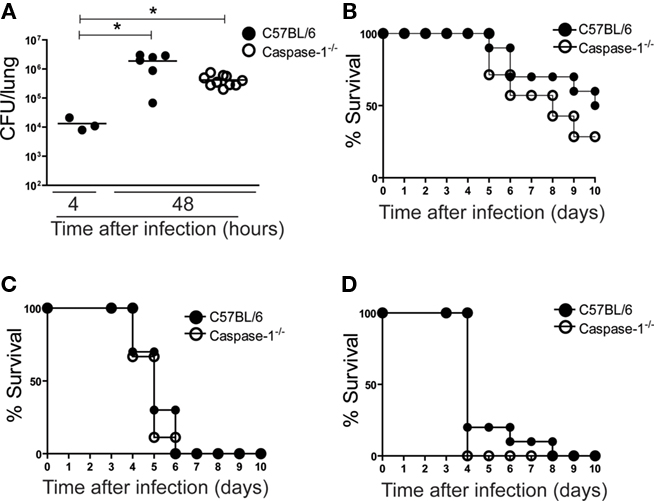
Figure 3. Caspase-1 does not influence pulmonary infection by L. longbeachae. C57BL/6 (closed circles) and caspase-1−/− (opened circles) mice were infected intranasally with Legionella longbeachae for determination of CFU in the lungs (A) and mice survival (B–D). (A) Mice were sacrificed after 4 or 48 h after infection and dilutions of the lung homogenates were added to CYE agar plates for CFU determination. Each dot represents a single animal and the horizontal lines represent the average. Six to 10 mice were used per group. (B–D) Survival of mice inoculated with 105 (B); 106 (C); or 107 (D) L. longbeachae per mice over a period of 10 days. Ten mice were used per group. Data are representative of those found in four independent experiments. An asterisk indicates a P value of <0.05.
Bypassing the Inflammasome is not Sufficient to Induce Mice Death in Response to Legionella Infection
Activation of the inflammasome is an important host defense against pulmonary infections by pathogenic bacteria including L. pneumophila (Amer et al., 2006; Molofsky et al., 2006; Zamboni et al., 2006; Miao et al., 2010). Since L. longbeachae bypasses the inflammasome activation and is lethal for mice, we investigated whether failure to trigger the Nlrc4 inflammasome is sufficient to Legionella to induce mice death. To test this hypothesis, we infected C57BL/6 mice with different MOIs of L. longbeachae and compared to infections using L. pneumophila and isogenic mutants for flagellin (flaA). Flagellin mutants were used because they are known to bypass Nlrc4-mediated growth restriction and are virulent for mice infections as the CFU numbers in the lungs increased over the first 48 h of mice infection (Amer et al., 2006; Molofsky et al., 2006; Ren et al., 2006; Case et al., 2009; Silveira and Zamboni, 2010). We found that whereas infections with L. longbeachae induced death of mice at MOIs higher than 105, flaA mutants and wild-type L. pneumophila did not induce mice death (Figure 4). These data suggests that bypassing the Nlrc4 inflammasome is not sufficient to induce mice death. Therefore, L. longbeachae may employ additional virulence strategies to multiply in mammalian hosts.
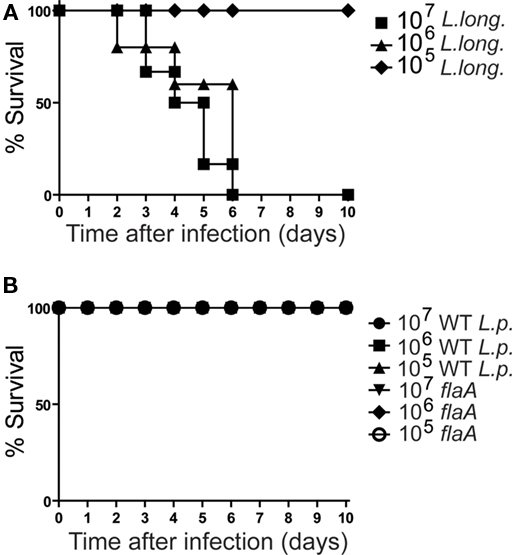
Figure 4. Legionella longbeachae, but not flaA mutants of L. pneumophila, is lethal for mice. (A) C57BL/6 mice were inoculated with 105 (diamonds); 106 (triangles); or 107 (squares) L. longbeachae (L. long.) per mouse and the survival was monitored over a period of 10 days. (B) C57BL/6 mice were inoculated 105, 106, or 107 CFUs of wild-type Legionella pneumophila (WT L.p.) or with flagellin mutants L. pneumophila (flaA). The survival was monitored over a period of 10 days. Ten mice were used per each group. Data are representative of those found in four independent experiments.
Discussion
The inflammasomes are important platforms that account for recognition and restriction of the infection by pathogenic microbes. Herein, we show that the inflammasome accounts for restriction of pulmonary infection of four species of non-pneumophila Legionella: L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens (Figure 1). Among the non-pneumophila Legionella that cause infection in humans, the most prevalent species are L. longbeachae, L. micdadei, and L. bozemanii, accounting for 2–7% of the infections worldwide (Muder and Yu, 2002). In this context, the demonstration that the Nlrc4 inflammasome accounts for restriction of in vivo infection by these species contributes to our understanding of the mechanisms by which host immune responses restrict infections by frequent pneumonia-causing bacteria such as L. micdadei and L. bozemanii.
It is worth noting that from the 52 species of the genus Legionella, only four species are prevalent causative agents of human infection (Gobin et al., 2009a). In this scenario, it is possible that the Nlrc4 inflammasome represents an important line of defense during pulmonary infection with these flagellated species of Legionella. Interestingly, although only L. pneumophila, L. longbeachae, L. bozemanii, and L. micdadei are frequent causative agents of pulmonary infections in humans, more than 25 species have been reported to induce disease in immunocompromised individuals (Gobin et al., 2009a). In this context, it is important to investigate whether the immunocompromised individuals that develop disease caused by infrequent Legionella spp. are defective for any component of the Nlrc4 inflammasome.
We have previously demonstrated that L. micdadei, L. bozemanii, L. gratiana, and L. rubrilucens express flagellin and trigger pyroptosis in isolated macrophages (Silveira and Zamboni, 2010). These previous observations are in agreement with the data presented here showing an important role of Nlrc4 inflammasome for restriction of pulmonary infection by these bacteria. Importantly, the Asc-containing inflammasome played no role in restriction of the infection by Legionella spp. (Figure 2). These data support the hypothesis that Nlrc4 and Asc proteins participate in distinct inflammasome (Case et al., 2009). However, this is still a controversial issue since macrophages from Asc−/− mouse are defective for caspase-1 activation, processing, and secretion of IL-1β in response to L. pneumophila infection (Zamboni et al., 2006; Case et al., 2009). These features had lead to the speculation that Asc protein does participate in the Nlrc4 inflammasome (Pedra et al., 2009). Alternatively, it is possible that Nlrc4 participates in the assembly of two distinct inflammasomes in response to L. pneumophila: one independent of Asc (which would be responsible for Legionella spp. growth restriction, pore formation, and pyroptosis) and one dependent on Asc (that would be required for robust caspase-1 activation and IL-1β secretion). In this context, further experiments may elucidate the complex interaction of Asc and Nlrc4 in the assembly of the pathogen-induced inflammasomes.
As opposed to flagellated Legionella spp., L. longbeachae does not encode genes for flagellum biosynthesis and thus does not express flagellin (Cazalet et al., 2010; Kozak et al., 2010; Silveira and Zamboni, 2010). Accordingly, experiments performed with L. longbeachae indicated that this bacterium does not trigger caspase-1 activation and fails to trigger pore formation and pyroptosis (Asare et al., 2007; Silveira and Zamboni, 2010). Here, we demonstrated that the inflammasome proteins might not substantially account for restriction of the infection by L. longbeachae in vivo (Figure 3). Importantly, L. longbeachae were lethal for mice infections in vivo, a feature not observed for other Legionella spp. (Alli et al., 2003; Asare et al., 2007; Gobin et al., 2009b). This feature has lead to speculation that L. longbeachae is highly virulent in mouse models because it bypasses the inflammasome-mediated growth restriction. To address this hypothesis we compared mortality curves of mice infected with L. longbeachae and L. pneumophila mutants for flaA, a gene that encodes flagellin. Although flaA mutants bypassed Nlrc4-dependent growth restriction, the flaA mutants failed to trigger death in infected mice even at high infection doses (Figure 4). These data suggests that bypassing the inflammasome is not sufficient to induce mouse mortality upon infection with virulent strains of L. pneumophila. Therefore, our data support the idea that L. longbeachae employ additional virulence strategies besides bypassing the activation of the Nlrc4 inflammasome. In this context, the recent elucidation of L. longbeachae genome revealed that the bacteria contain genes predicted to encode proteins for capsule formation and several genes encoding proteins with predicted eukaryotic-like motifs that are not shared with L. pneumophila (Cazalet et al., 2010; Kozak et al., 2010). We envisage that in the next few years, investigation of these putative virulence genes might reveal novel molecular mechanisms that explain the high virulence of L. longbeachae in mammalian models of experimental infection.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful for Maira Nakamura for technical assistance and for Larissa D. Cunha for her critical review of the manuscript. This work was supported by PEW, INCTV/CNPq, and FAPESP grant 2006/52867-4 (to DSZ). Marcelo S. F. Pereira is supported by a doctoral fellowship from FAPESP (2007/55852-0). Gabriel G. Marques and José Eduardo DelLama were supported by technical training fellowships from FAPESP (2009/14437-6) and PRP/USP respectively. Dario S. Zamboni is a research fellow from CNPq.
References
Alli, O. A., Zink, S., von Lackum, N. K., and Abu-Kwaik, Y. (2003). Comparative assessment of virulence traits in Legionella spp. Microbiology 149, 631–641.
Amer, A., Franchi, L., Kanneganti, T. D., Body-Malapel, M., Ozoren, N., Brady, G., Meshinchi, S., Jagirdar, R., Gewirtz, A., Akira, S., and Nunez, G. (2006). Regulation of Legionella phagosome maturation and infection through flagellin and host Ipaf. J. Biol. Chem. 281, 35217–35223.
Archer, K. A., Ader, F., Kobayashi, K. S., Flavell, R. A., and Roy, C. R. (2010). Cooperation between multiple microbial pattern recognition systems is important for host protection against the intracellular pathogen Legionella pneumophila. Infect. Immun. 78, 2477–2487.
Asare, R., Santic, M., Gobin, I., Doric, M., Suttles, J., Graham, J. E., Price, C. D., and Abu Kwaik, Y. (2007). Genetic susceptibility and caspase activation in mouse and human macrophages are distinct for Legionella longbeachae and L. pneumophila. Infect. Immun. 75, 1933–1945.
Berrington, W. R., Iyer, R., Wells, R. D., Smith, K. D., Skerrett, S. J., and Hawn, T. R. (2010). NOD1 and NOD2 regulation of pulmonary innate immunity to Legionella pneumophila. Eur. J. Immunol. 40, 3519–3527.
Case, C. L., Shin, S., and Roy, C. R. (2009). Asc and Ipaf Inflammasomes direct distinct pathways for caspase-1 activation in response to Legionella pneumophila. Infect. Immun. 77, 1981–1991.
Cazalet, C., Gomez-Valero, L., Rusniok, C., Lomma, M., Dervins-Ravault, D., Newton, H. J., Sansom, F. M., Jarraud, S., Zidane, N., Ma, L., Bouchier, C., Etienne, J., Hartland, E. L., and Buchrieser, C. (2010). Analysis of the Legionella longbeachae genome and transcriptome uncovers unique strategies to cause Legionnaires’ disease. PLoS Genet. 6, e1000851. doi: 10.1371/journal.pgen.1000851
Feeley, J. C., Gibson, R. J., Gorman, G. W., Langford, N. C., Rasheed, J. K., Mackel, D. C., and Baine, W. B. (1979). Charcoal-yeast extract agar: primary isolation medium for Legionella pneumophila. J. Clin. Microbiol. 10, 437–441.
Franchi, L., Amer, A., Body-Malapel, M., Kanneganti, T. D., Ozören, N., Jagirdar, R., Inohara, N., Vandenabeele, P., Bertin, J., Coyle, A., Grant, E. P., and Núñez, G. (2006). Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1beta in salmonella-infected macrophages. Nat. Immunol. 7, 576–582.
Franchi, L., Park, J. H., Shaw, M. H., Marina-Garcia, N., Chen, G., Kim, Y. G., and Nunez, G. (2008). Intracellular NOD-like receptors in innate immunity, infection and disease. Cell. Microbiol. 10, 1–8.
Frutuoso, M. S., Hori, J. I., Pereira, M. S., Junior, D. S., Sonego, F., Kobayashi, K. S., Flavell, R. A., Cunha, F. Q., and Zamboni, D. S. (2010). The pattern recognition receptors Nod1 and Nod2 account for neutrophil recruitment to the lungs of mice infected with Legionella pneumophila. Microbes Infect. 12, 819–827.
Gobin, I., Newton, P. R., Hartland, E. L., and Newton, H. J. (2009a). Infections caused by non-pneumophila species of Legionella. Rev. Med. Microbiol. 20, 1–11.
Gobin, I., Susa, M., Begic, G., Hartland, E. L., and Doric, M. (2009b). Experimental Legionella longbeachae infection in intratracheally inoculated mice. J. Med. Microbiol. 58, 723–730.
Janeway, C. A. Jr., and Medzhitov, R. (2002). Innate immune recognition. Annu. Rev. Immunol. 20, 197–216.
Kawai, T., and Akira, S. (2009). The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 21, 317–337.
Kozak, N. A., Buss, M., Lucas, C. E., Frace, M., Govil, D., Travis, T., Olsen-Rasmussen, M., Benson, R. F., and Fields, B. S. (2010). Virulence factors encoded by Legionella longbeachae identified on the basis of the genome sequence analysis of clinical isolate D-4968. J. Bacteriol. 192, 1030–1044.
Kuida, K., Lippke, J. A., Ku, G., Harding, M. W., Livingston, D. J., Su, M. S., and Flavell, R. A. (1995). Altered cytokine export and apoptosis in mice deficient in interleukin-1 beta converting enzyme. Science 267, 2000–2003.
Lara-Tejero, M., Sutterwala, F. S., Ogura, Y., Grant, E. P., Bertin, J., Coyle, A. J., Flavell, R. A., and Galan, J. E. (2006). Role of the caspase-1 inflammasome in Salmonella typhimurium pathogenesis. J. Exp. Med. 203, 1407–1412.
Lightfield, K. L., Persson, J., Brubaker, S. W., Witte, C. E., von Moltke, J., Dunipace, E. A., Henry, T., Sun, Y. H., Cado, D., Dietrich, W. F., Monack, D. M., Tsolis, R. M., and Vance, R. E. (2008). Critical function for Naip5 in inflammasome activation by a conserved carboxy-terminal domain of flagellin. Nat. Immunol. 9, 1171–1178.
Mariathasan, S., Newton, K., Monack, D. M., Vucic, D., French, D. M., Lee, W. P., Roose-Girma, M., Erickson, S., and Dixit, V. M. (2004). Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430, 213–218.
Masumoto, J., Yang, K., Varambally, S., Hasegawa, M., Tomlins, S. A., Qiu, S., Fujimoto, Y., Kawasaki, A., Foster, S. J., Horie, Y., Mak, T. W., Núñez, G., Chinnaiyan, A. M., Fukase, K., and Inohara, N. (2006). Nod1 acts as an intracellular receptor to stimulate chemokine production and neutrophil recruitment in vivo. J. Exp. Med. 203, 203–213.
Miao, E. A., Alpuche-Aranda, C. M., Dors, M., Clark, A. E., Bader, M. W., Miller, S. I., and Aderem, A. (2006). Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1beta via Ipaf. Nat. Immunol. 7, 569–575.
Miao, E. A., Leaf, I. A., Treuting, P. M., Mao, D. P., Dors, M., Sarkar, A., Warren, S. E., Wewers, M. D., and Aderem, A. (2010). Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 11, 1136–1142.
Molofsky, A. B., Byrne, B. G., Whitfield, N. N., Madigan, C. A., Fuse, E. T., Tateda, K., and Swanson, M. S. (2006). Cytosolic recognition of flagellin by mouse macrophages restricts Legionella pneumophila infection. J. Exp. Med. 203, 1093–1104.
Muder, R. R., and Yu, V. L. (2002). Infection due to Legionella species other than L. pneumophila. Clin. Infect. Dis. 35, 990–998.
Pedra, J. H., Cassel, S. L., and Sutterwala, F. S. (2009). Sensing pathogens and danger signals by the inflammasome. Curr. Opin. Immunol. 21, 10–16.
Ren, T., Zamboni, D. S., Roy, C. R., Dietrich, W. F., and Vance, R. E. (2006). Flagellin-deficient Legionella mutants evade caspase-1- and Naip5-mediated macrophage immunity. PLoS Pathog. 2, e18. doi: 10.1371/journal.ppat.0020018
Silva, G. K., Gutierrez, F. R., Guedes, P. M., Horta, C. V., Cunha, L. D., Mineo, T. W., Santiago-Silva, J., Kobayashi, K. S., Flavell, R. A., Silva, J. S., and Zamboni, D. S. (2010). Cutting edge: nucleotide-binding oligomerization domain 1-dependent responses account for murine resistance against Trypanosoma cruzi infection. J. Immunol. 184, 1148–1152.
Silveira, T. N., and Zamboni, D. S. (2010). Pore formation triggered by Legionella spp. is an Nlrc4 inflammasome-dependent host cell response that precedes pyroptosis. Infect. Immun. 78, 1403–1413.
Sutterwala, F. S., and Flavell, R. A. (2009). NLRC4/IPAF: a CARD carrying member of the NLR family. Clin. Immunol. 130, 2–6.
Whitfield, N. N., Byrne, B. G., and Swanson, M. S. (2010). Mouse macrophages are permissive to motile Legionella species that fail to trigger pyroptosis. Infect. Immun. 78, 423–432.
Zamboni, D. S., Kobayashi, K. S., Kohlsdorf, T., Ogura, Y., Long, E. M., Vance, R. E., Kuida, K., Mariathasan, S., Dixit, V. M., Flavell, R. A., Dietrich, W. F., and Roy, C. R. (2006). The Birc1e cytosolic pattern-recognition receptor contributes to the detection and control of Legionella pneumophila infection. Nat. Immunol. 7, 318–325.
Keywords: Legionella, pulmonary infection, caspase-1, Nlrc4, inflammasome
Citation: Pereira MSF, Marques GG, DelLama JE and Zamboni DS (2011) The Nlrc4 Inflammasome contributes to restriction of pulmonary infection by flagellated Legionella spp. that trigger pyroptosis. Front. Microbio. 2:33. doi: 10.3389/fmicb.2011.00033
Received: 17 November 2010;
Paper pending published: 28 December 2010;
Accepted: 07 February 2011;
Published online: 15 February 2011.
Edited by:
Amal Amer, The Ohio State University, USAReviewed by:
Amal Amer, The Ohio State University, USADaniel E. Voth, University of Arkansas for Medical Sciences, USA
Elizabeth L. Hartland, University of Melbourne, Australia
Copyright: © 2011 Pereira, Marques, DelLama and Zamboni. This is an open-access article subject to an exclusive license agreement between the authors and Frontiers Media SA, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Dario S. Zamboni, Department of Cell Biology, Medical School at Ribeirão Preto, Faculdade de Medicina de Ribeirão Preto, University of São Paulo, Avenida Bandeirantes, 3900, Ribeirão Preto, SP 14049-900, Brazil. e-mail: dszamboni@fmrp.usp.br