 Luis G. C. Pacheco1,2 Thiago L. P. Castro1 Rodrigo D. Carvalho1 Pablo M. Moraes1 Fernanda A. Dorella1 Natália B. Carvalho1 Susan E. Slade3 James H. Scrivens3 Martin Feelisch4 Roberto Meyer2 Anderson Miyoshi1 Sergio C. Oliveira1 Christopher G. Dowson3 Vasco Azevedo1*
Luis G. C. Pacheco1,2 Thiago L. P. Castro1 Rodrigo D. Carvalho1 Pablo M. Moraes1 Fernanda A. Dorella1 Natália B. Carvalho1 Susan E. Slade3 James H. Scrivens3 Martin Feelisch4 Roberto Meyer2 Anderson Miyoshi1 Sergio C. Oliveira1 Christopher G. Dowson3 Vasco Azevedo1*- 1 Institute of Biological Sciences, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 2 Institute of Health Sciences, Universidade Federal da Bahia, Salvador, Brazil
- 3 School of Life Sciences, University of Warwick, Coventry, UK
- 4 Warwick Medical School, University of Warwick, Coventry, UK
Pathogenic intracellular bacteria can respond to antimicrobial mechanisms of the host cell through transient activation of stress-responsive genes by alternative sigma (σ) factors of the RNA polymerase. We evaluated the contribution of the extracytoplasmic function sigma factor σE for Corynebacterium pseudotuberculosis resistance to stress conditions resembling those found intracellularly during infection. A sigE-null mutant strain (ΔsigE) of this bacterium was more susceptible in vitro to acidic pH, cell surface stressors, and biologically relevant concentrations of nitric oxide (NO). The same mutant strain was unable to persist in C57BL/6 mice but remained infective in mice lacking inducible nitric oxide synthase (iNOS), confirming the significance of σE for resistance to nitric oxide/peroxide stress in vivo. High-throughput proteomic analysis identified NO-responsive extracellular proteins of C. pseudotuberculosis and demonstrated the participation of σE in composition of this bacterium’s exoproteome.
Introduction
Corynebacterium pseudotuberculosis, a pathogenic bacterium belonging to the so-called CMN-group of Actinobacteria, is the etiological agent of various disease manifestations in different hosts, including humans (Dorella et al., 2006; Trost et al., 2010). The high infectious potential of C. pseudotuberculosis is dependent on its ability to resist diverse stressful conditions, both as a free-living organism and within the host. Previous reports have suggested that this bacterium can survive in the environment for several months before infecting an animal (Baird and Fontaine, 2007). Once inside the host, C. pseudotuberculosis survives phagocytosis to remain as an intracellular parasite within phagocytic cells (Stefańska et al., 2010). This is achieved by overcoming the innate antimicrobial defense mechanisms of the host cells. These cells create a harsh environment inside the phagolysosome by generating reactive oxygen and nitrogen intermediates, via phagocyte oxidase (Phox) and inducible nitric oxide synthase (iNOS), respectively (Nathan and Shiloh, 2000).
Bacteria can respond to different environmental stimuli through switching of the primary sigma (σ) factor subunit that is associated with core RNA polymerase by alternative σ factors. This confers novel promoter-recognition specificities to the polymerase and provides a mechanism for rapid regulation of different sets of stress-responsive genes (Staron et al., 2009). Proteins of the extracytoplasmic function (ECF) family of alternative σ factors, such as σE, mostly regulate response to cell surface-stresses and have already been shown to control virulence-associated genes in different pathogenic bacteria (Helmann, 2002; Kazmierczak et al., 2005).
Previous studies on Mycobacterium tuberculosis transcriptional responses to the stress conditions found within phagocytic cells have suggested the involvement of the ECF sigma factor σE in bacterial intracellular survival (Ohno et al., 2003; Schnappinger et al., 2003; Talaat et al., 2004; Fontán et al., 2008). Moreover, sigE has recently been shown to be part of a small set of M. tuberculosis genes that are selectively up-regulated in response to nitrosative stress (Voskuil et al., 2011).
In this study with C. pseudotuberculosis, we show that σE is indeed required for resistance to in vitro-generated, biologically relevant concentrations of nitric oxide (NO). Moreover, we also demonstrate that this regulatory protein plays a role in resistance to combined NO/peroxide stress faced by bacteria during infection.
Materials and Methods
Bacterial Strains and Growth Conditions
Corynebacterium pseudotuberculosis strains were routinely maintained in Brain Heart Infusion broth or in BHI 1.5% bacteriological agar plates, at 37°C. Tween-80 was added to the broth cultures at 0.05% to prevent cell clumping. When necessary, the antibiotic kanamycin was used at 25 μg/ml. Escherichia coli strains were maintained in Luria–Bertani medium and employed in DNA manipulation experiments, according to standard protocols. Ampicillin (100 μg/ml) and kanamycin (50 μg/ml) were used where appropriate.
A chemically defined medium (CDM), previously optimized for growth of C. pseudotuberculosis (Moura-Costa et al., 2002), was used in all the experiments that included stress-generating agents. The composition of the CDM was as follows: autoclaved phosphate buffer pH = 7.4 [Na2HPO4·7H2O (12.93 g/l), KH2PO4 (2.55 g/l), NH4Cl (1 g/l), MgSO4·7H2O (0.20 g/l), CaCl2 (0.02 g/l), and 0.05% (v/v) Tween-80]; 4% (v/v) MEM Vitamins Solution 100× (Invitrogen); 1% (v/v) MEM Amino Acids Solution 50× (Invitrogen); 1% (v/v) MEM Non-Essential Amino Acids Solution 100× (Invitrogen); and 1.2% (w/v) filter-sterilized glucose.
Construction of a sigE-Null Mutant Strain of C. pseudotuberculosis
A conserved region of the sigE gene of C. pseudotuberculosis (GenBank: ADL20681.1) was initially isolated by a strategy employing degenerate primers. Basically, the amino acid sequences of the σE factors of the bacteria M. tuberculosis (Rv1221), C. diphtheria (DIP0994), C. glutamicum (NCgl1075), and C. efficiens (CE1177) were retrieved from public databases and aligned using the ClustalW tool. Highly conserved regions were identified in the σE factors of these bacteria and degenerate primers were designed according to the coding sequences of these factors, such that it would be possible to amplify a partial region of the sigE gene in all genomes analyzed. The primer pair sigE#1: GGMACCGCAGCDTTCGACGC and sigE#2: CGTCCRCGGTGRATWCGGGA was used for amplification of an expected 490 bp internal fragment of the sigE gene of C. pseudotuberculosis. PCR products around the expected size were purified from agarose gels, ligated into the vector pCR2.1 TOPO (Invitrogen), and sequenced according to standard protocols.
A plasmid carrying a fragment of the sigE gene was introduced into the wild-type strain 1002 of C. pseudotuberculosis by electroporation. This plasmid functions as a suicide vector for this bacterium, as it does not carry a functional corynebacterial replication origin. C. pseudotuberculosis clones that underwent recombination were selected by kanamycin resistance (25 μg/ml). Confirmation of recombination events in the C. pseudotuberculosis genome was obtained by Southern blot analysis using a [α32-P]dCTP radiolabeled probe for the sigE gene, or through PCR reactions employing different combinations of primers that align in the start and stop codons of sigE with primers that align within the inserted plasmid: sigE#3-ATGACATCGAACAGTGGTTC/sigE#4-TTAGTGGGACATCGGTAGG; Kan#1-ATGATTGAACAAGATGGATTG/Kan#2-TTAATAATTCAGAAGAACTC; M13F/M13R (Invitrogen; data not shown).
In vitro Resistance to Stress-Generating Agents
Corynebacterium pseudotuberculosis strains were grown in CDM to early exponential phase (OD540nm = ∼0.1), cultures were split into several aliquots and then incubated separately with different concentrations of various stress-generating agents, as follows: osmotic stress (NaCl, 0.5 and 1 M); acidic stress (HCl, to pH 4.5 and 5.5); detergent stress (SDS, 0.01% w/v and 0.05%); lysozyme, 500 and 750 μg/ml; alcoholic stress (ethanol, 2.5 and 5%); oxidative stress (H2O2, 1, 10, and 50 mM); nitric oxide stress (DETA/NO – diethylenetriamine NON-Oate, 0.1 and 1 mM). For starvation stress, CDM was prepared with low concentrations of glucose: 0.3 and 0.15%. For heat and cold shocks, cultures were incubated at 50 or 4°C for 30 min, and then returned to normal growth at 37°C.
Growth of the control and stressed cultures of C. pseudotuberculosis, of both wild-type (1002) and mutant (ΔsigE) strains, was monitored for 24 h in a LabSystems iEMS Absorbance Plate Reader (Thermo Fisher), at OD540nm. Results were plotted using the GraphPad Prism software (GraphPad Software, Inc); the integrals of the areas under the curves (AUC) were determined, and a growth index (GI) was then calculated, as follows: GI (%) = [(AUCTreated culture/AUCControl culture) × 100]. Experiments were performed at least in triplicate.
Nitric Oxide Detection
Steady-state concentrations of NO generated by the NO-donor DETA/NO in the cell culture medium were determined amperometrically using an NO-specific electrode (ISO-NOP, World Precision Instruments, Inc.) attached to an NO meter (ISO-NO Mark II, WPI). Briefly, the electrode was immersed in CDM and the electrode current allowed to stabilize under conditions identical to those used in the cell culture experiments before addition of different concentrations of DETA/NO and recording of the electrode response. Stock solutions of DETA/NO (Sigma-Aldrich) were prepared fresh and kept on ice in the dark, as described (Feelisch, 1998). Currents in pA were converted into NO concentrations by comparison to a standard calibration curve generated from either copper(I)-mediated decomposition of S-nitroso-N-acetyl-D,L-penicillamine or from iodide-mediated nitrite reduction, under otherwise identical conditions (Davies and Zhang, 2008).
Infection of Mouse Macrophages with C. pseudotuberculosis
Bone-marrow derived macrophages (BMMs) were obtained from the femurs and tibias of C57BL/6 or iNOS knockout mice (iNOS−/−), according to a standardized protocol (Carvalho et al., 2011). Gamma (γ)-interferon was added at 20 U/well for macrophage activation. A Neutral Red assay was used to evaluate macrophage viability following C. pseudotuberculosis infection (MOI 5:1), as described (McKean et al., 2007).
Corynebacterium pseudotuberculosis Growth and Persistence in C57BL/6 and iNOS−/− Mice
C57BL/6 or iNOS−/− mice were infected intraperitoneally with 106 colony forming units (CFU) of the wild-type (1002) or mutant (ΔsigE) strain of C. pseudotuberculosis. On days 1 and 3 post-infection, animals were sacrificed and bacterial loads in the spleens were enumerated.
Extraction of C. pseudotuberculosis Extracellular Proteins Following NO-Treatment and Comparative Exoproteome Profiling
The 1002 (wt) and ΔsigE strains of C. pseudotuberculosis were grown in CDM to mid-exponential phase, cultures were split into two aliquots and 100 μM of the NO-donor DETA/NO were added to one aliquot of each strain. NO-treated cultures along with control cultures were further incubated for 1 h at 37°C. At this point, extracellular proteins were extracted by the three-phase partitioning technique and identified/quantified by a high-throughput proteomic strategy, based on a recently introduced method of liquid chromatography–mass spectrometry acquisition (LC–MSE), exactly as previously described (Pacheco et al., 2011). Biological function annotations for the identified proteins were retrieved from the gene ontology (GO) database, using the web tool AmiGO1 (Carbon et al., 2009). Protein regulation data were obtained from the CoryneRegNet v6.0 Database2 (Pauling et al., 2011).
Results
A sigE-Null Mutant strain of C. pseudotuberculosis is more Sensitive to Cell Surface Stressors, Acidic pH, and in vitro-Generated Nitric Oxide
In order to evaluate the role played by the ECF sigma factor σE in C. pseudotuberculosis resistance to stress conditions faced during intracellular infection, a sigE-null mutant strain of this bacterium (ΔsigE) was generated by homologous recombination in the parental strain 1002 (wt). Both strains were submitted to a series of stress-generating agents in vitro, which aimed at mimicking conditions found by bacterial pathogens in the intraphagosomal environment (Rohde et al., 2007; Ehrt and Schnappinger, 2009; Schaible, 2009).
Mutation of sigE in C. pseudotuberculosis did not alter cell growth and morphology under normal conditions (Figure 1). Growth of the ΔsigE strain was also comparable to the 1002 (wt) strain under conditions of nutrient starvation (limiting glucose), and under osmotic, thermal, alcoholic, and oxidative stresses (Figure 1). On the other hand, the mutant strain was more sensitive to an acidic pH that resembles that found within an activated macrophage (pH = 5.5), and to cell surface stressors, namely SDS and lysozyme treatments (Figure 1), corroborating previous studies on the role of σE in resistance to environmental-stresses in other corynebacteria and mycobacteria (Manganelli et al., 2001; Park et al., 2008). Interestingly, growth of the ΔsigE strain was also more affected than that of the wt strain following exposure to different concentrations of the NO-donor DETA/NO (Figures 1 and 2A). NO electrode measurements indicated that the concentrations of the NO-donor used in our experiments, 0.1 and 1 mM, resulted in steady-state NO concentrations of ∼500 nM and 5 μM, respectively. The effect of these concentrations on growth of the two C. pseudotuberculosis strains was apparently more due to a bacteriostatic rather than bactericidal action of NO (Figure 2C), as reported for other bacterial pathogens (Ogawa et al., 2001; Voskuil et al., 2011). Notably, the ΔsigE strain was much more sensitive to the combination of the NO-donor and H2O2 (Figures 1 and 2B), reinforcing the requirement of σE for C. pseudotuberculosis resistance to combined NO/peroxide stress.
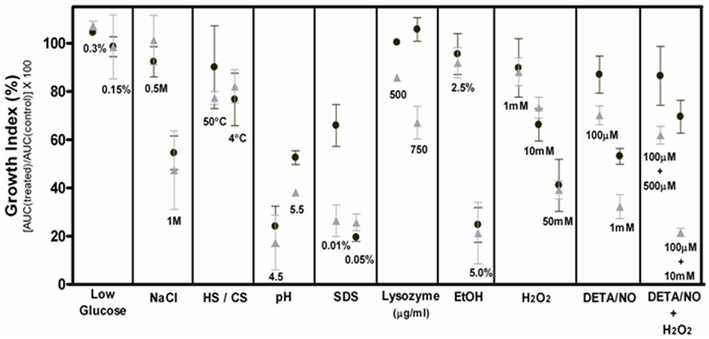
Figure 1. Differential susceptibilities of the parental (1002) and mutant (ΔsigE) strains of C. pseudotuberculosis to different stress conditions in vitro. Average percent growth ± SD of 1002 (black circles) and ΔsigE (gray triangles) strains exposed to various stress conditions in vitro (see text for details), in comparison to non-treated controls. HS, heat shock; CS, cold shock.
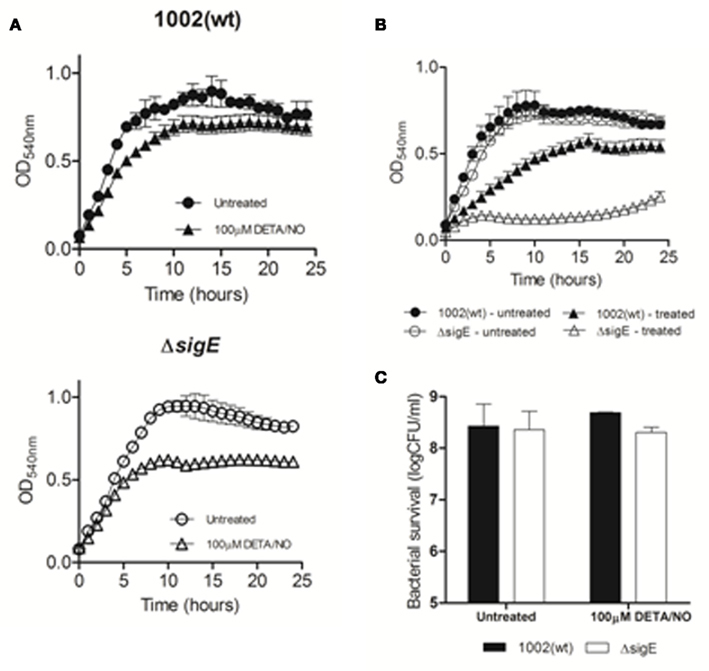
Figure 2. Growth of the wt and mutant strains of C. pseudotuberculosis following exposure to nitric oxide/peroxide stress. (A) Representative growth curves of 1002 and ΔsigE strains exposed to 100 μM DETA/NO (B) Representative growth curves of 1002 (filled symbols) and ΔsigE (open symbols) strains treated with a combination of 10 mM H2O2 + 100 μM DETA/NO. Untreated (control) cultures are represented by circles. (C) Colony forming units counts of 1002 or ΔsigE strains following 1 h treatment by 100 μM DETA/NO.
σE is Required for C. pseudotuberculosis Resistance to NO-Stress during Infection
The participation of σE in C. pseudotuberculosis resistance to the conditions found within the host was first evaluated by infection of C57BL/6 mice with the wt and mutant strains of this bacterium, followed by analysis of bacterial persistence in mouse spleens. After 3 days of infection, the ΔsigE strain of C. pseudotuberculosis was virtually undetectable in the spleens of C57BL/6 mice, whereas the 1002 (wt) strain still persisted (Figure 3A). We then evaluated whether nitrosative stress was a determining condition for the lowered persistence of the ΔsigE mutant in the host. For this, we infected C57BL/6 or iNOS knockout (iNOS−/−) mice, which are unable to produce NO in the intraphagosomal environment, and compared bacterial loads in mouse spleens 3 days post-infection. Again, the C. pseudotuberculosis strain lacking σE was unable to persist in C57BL/6 wild-type mice (Figure 3B); nevertheless, the ΔsigE mutant displayed a surprisingly high ability to survive in the iNOS−/− animals, following an initial 106 CFU inoculum (Figure 3B).
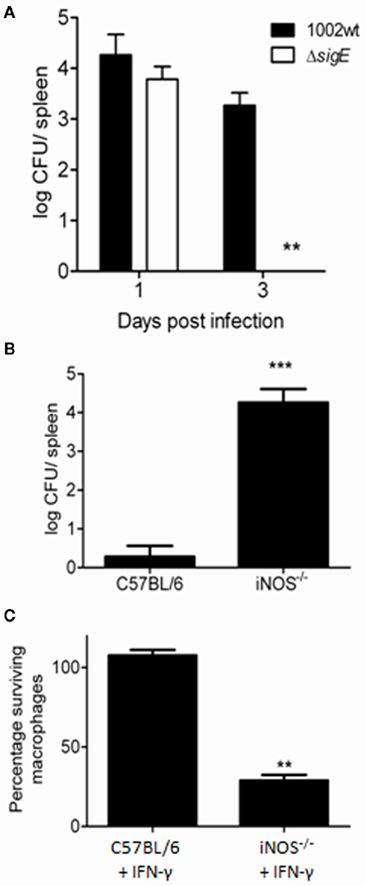
Figure 3. Differential persistence of the ΔsigE mutant of C. pseudotuberculosis during infection of wild-type and iNOS−/− mice. (A) The ΔsigE strain is rapidly eliminated, becoming almost undetectable in the spleens of C57BL/6 mice 3 days after inoculation with 106 CFU. (B) Bacterial loads in spleens of C57BL/6 or iNOS−/− mice, 3 days post-infection with the ΔsigE strain of C. pseudotuberculosis. (C) Viability of macrophages obtained from either C57BL/6 or iNOS−/− mice, 4 h post-infection with the ΔsigE strain of C. pseudotuberculosis. **t-Test p < 0.001; ***t-test p < 0.0001.
Wild-type C. pseudotuberculosis has been shown to possess a profound capability to kill macrophages in culture (Stefańska et al., 2010), but this natural ability is also affected in the ΔsigE strain (Figure 3C). However, only ca. 30% of iNOS−/− macrophages survived after 4 h infection with this mutant (Figure 3C). This corroborated the importance of σE in C. pseudotuberculosis resistance to NO generated intracellularly.
Alterations of the C. pseudotuberculosis Exoproteome in Response to NO-Stress
The extracellular proteomes of the 1002 (wt) and ΔsigE strains of C. pseudotuberculosis were profiled by a high-throughput proteomics method (LC–MSE), before and after treatment by 100 μM of the NO-donor DETA/NO.
In total, 281 extracellular proteins were confidentially identified by this strategy, in the four groups studied: (i) 1002 (wt) untreated; (ii) 1002 (wt) + DETA/NO; (iii) ΔsigE untreated; (iv) ΔsigE + DETA/NO. This represented 104 different extracellular proteins of C. pseudotuberculosis (Figure 4A). Proteins exported after NO-treatment could be detected in both 1002 (wt) and ΔsigE strains (Figure 4A). The mutant strain exported a higher number of proteins in comparison to the wt strain, under normal growth conditions, and after NO-stress (Figure 4A). The majority (67%) of these differential proteins of the ΔsigE strain are predicted in silico to be truly exported proteins (data not shown). Moreover, the proteins commonly identified in the four groups presented very similar concentrations in all samples (Figure 4B) showing a high reproducibility of the methods used for extraction and identification of the exoproteins. This demonstrates that the qualitative differences observed between the exoproteomes are in fact due to biological variations of the extracellular proteomes.
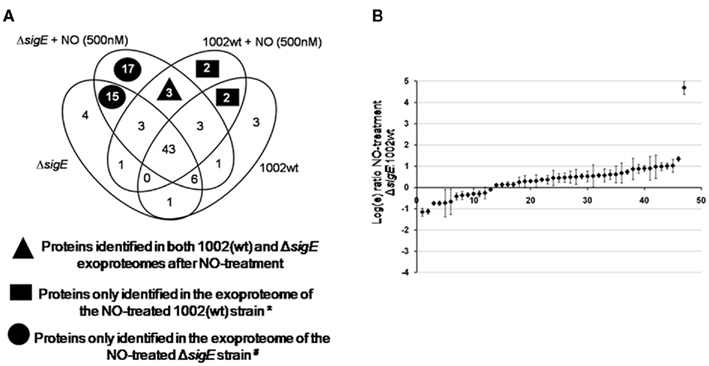
Figure 4. Extracellular proteins identified in the 1002 (wt) and ΔsigE strains following NO-stress. (A) Venn diagram showing the distribution of the 104 different C. pseudotuberculosis extracellular proteins identified by LC–MSE analysis throughout the four groups studied: (i) 1002 (wt) untreated; (ii) 1002 (wt) + 100 μM DETA/NO; (iii) ΔsigE untreated; (iv) ΔsigE + 100 μM DETA/NO. *Compared to the NO-treated ΔsigE strain. #Compared to the NO-treated 1002 (wt) strain. (B) Relative expression analysis of exoproteins commonly identified between the wt and mutant strains of C. pseudotuberculosis after NO-treatment. Protein expression data was obtained by label-free quantification using LC–MSE. A single overrepresented protein (putative secreted protein; ADL21925.1) was identified in the exoproteome of the ΔsigE strain following NO-stress.
Three proteins were commonly identified in the 1002 (wt) and ΔsigE strains exclusively after treatment by NO (Figure 4A). Only a few proteins were differentially exported in the 1002 (wt) strain in response to NO, compared to the mutant strain (Figure 4A). These included proteins predicted to be involved in metal ion transport and cell redox homeostasis. A putative dioxygenase was also specifically detected in the exoproteome of the wt strain (Table 1). On the other hand, 17 differential proteins could be detected in the extracellular proteome of the ΔsigE-null strain following NO-stress (Figure 4A). Metal ion transport and cell redox homeostasis were again highly represented predicted biological functions in this protein set (Table 1). Some of these proteins have already been seen to participate in C. pseudotuberculosis responses to other environmental stress conditions, according to the CoryneRegNet Database (Table 1).
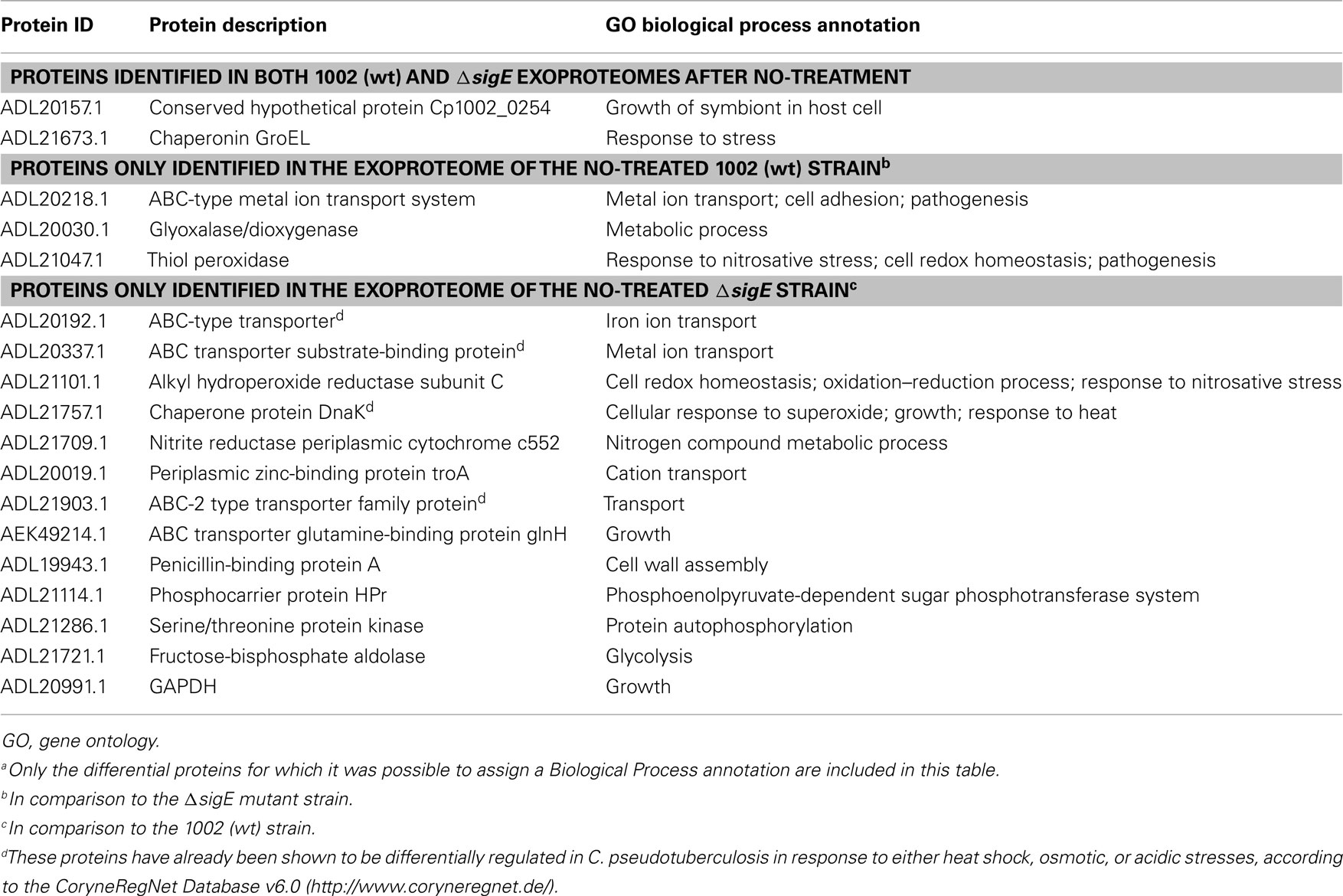
Table 1. Proteins identified in the exoproteomes of the 1002 (wt) and ΔsigE strains of C. pseudotuberculosi s following NO-stressa.
A few proteins primarily considered to have a cytoplasmic location have been identified following NO-stress (Table 1). These included the chaperonin GroEL, which was found in the exoproteomes of both wt and ΔsigE strains of C. pseudotuberculosis, and the proteins glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and dnaK, differentially exported only in the mutant strain (Table 1). These cytoplasmic proteins have been consistently identified in recent exoproteome studies of various bacteria (Sengupta et al., 2010; Dreisbach et al., 2011). Besides, environmental cues such as osmotic stress and host contact have been shown to induce exportation of some of these proteins (Chitlaru et al., 2007; Mattinen et al., 2007; Pumirat et al., 2009). However, the molecular mechanisms that account for non-classical (leaderless) secretion of specific cytoplasmic proteins is still to be completely understood (Pasztor et al., 2010; Yang et al., 2011). Moonlighting roles have already been demonstrated for these proteins while in the extracellular environment, including adhesion to host cells and evasion of host’s immune mechanisms (Matta et al., 2010; Jin et al., 2011).
Noticeably, while most of the proteins commonly identified between the exoproteomes of the NO-treated strains presented very similar concentrations (Figure 4B), a single protein annotated as a “putative secreted protein” (ADL21925.1) was highly secreted only in the ΔsigE strain. This protein has also been shown to be differentially regulated during osmotic stress, according to CoryneRegNet.
Discussion
Pathogenic bacteria that reside within phagocytic cells face nitrosative stress due to the action of iNOS, which delivers NO to the intraphagosomal compartment (Nathan and Shiloh, 2000; Ehrt and Schnappinger, 2009). NO is a highly reactive radical which in high concentrations can lead to metabolic arrest and damage of cellular components, including membranes and DNA, thus inhibiting bacterial growth (Ogawa et al., 2001). Through combination with reactive oxygen species, considerably more reactive antimicrobial agents can be generated, such as peroxynitrite (ONOO−; Ehrt and Schnappinger, 2009). To survive such stressful conditions found in the intracellular environment, bacteria need to be able to rapidly respond to environmental cues through global changes in gene expression; this role is played by alternative sigma factors of the bacterial RNA polymerase (Helmann, 2002).
Previous studies with M. tuberculosis have consistently shown that the gene coding for the extracytoplasmic function sigma factor σE is up-regulated during intracellular infection (Ohno et al., 2003; Schnappinger et al., 2003; Talaat et al., 2004; Fontán et al., 2008). More recently, the sigE gene was shown to participate in a specific transcriptional response of M. tuberculosis following NO-treatment in vitro (Voskuil et al., 2011). Taken together, these studies strongly suggest that σE may be necessary for M. tuberculosis to cope with reactive nitrogen species within the host cell, but this possibility has not been specifically investigated yet.
In this study, we show that σE is indeed required for full resistance to nitrosative stress in the intracellular actinobacterium C. pseudotuberculosis. Biologically relevant concentrations of NO, generated here by the NO-donor DETA/NO, importantly affected growth of this bacterium, similarly to what has been reported for M. tuberculosis (Voskuil et al., 2003, 2011). Growth impairment was much more pronounced in a sigE-null mutant strain of C. pseudotuberculosis, following exposure to both low and high concentrations of NO (Figure 1). The fact that the strain lacking σE displayed lowered persistence in mice following experimental infection (Figure 3A) raises the possibility that impaired growth in the host might be due, at least in part, to the inability of this strain to resist NO-stress during infection. This notion was confirmed by our experiments in mice unable to mount an intracellular NO response during infection (iNOS knockout mice). In these animals, the ΔsigE strain of C. pseudotuberculosis persists much longer than in wild-type mice (Figure 3B); this demonstrated that σE is in fact necessary for resistance to nitrosative stress in vivo. Moreover, the significant effect of this mutant on viability of iNOS−/− macrophages in culture (Figure 3C), which is expected to be related to the ability of the bacteria to replicate in the intraphagosomal compartment, further corroborates the involvement of σE in bacterial survival to reactive nitrogen species.
Since the ECF sigma factor σE seems to participate in response to cell envelope stresses in different bacteria, we hypothesized that mutation of its coding gene in C. pseudotuberculosis could lead to alterations of this bacterium’s extracellular proteome in response to NO-stress. In fact, we could identify differentially regulated exoproteins of C. pseudotuberculosis following NO-treatment. Besides, marked differences were observed between the exoproteomes of the 1002 (wt) and ΔsigE strains (Figure 4).
Exportation of proteins involved in metal ion acquisition and redox homeostasis seems to be a primary response of C. pseudotuberculosis to biologically relevant concentrations of NO (Table 1). Recent studies on the transcriptional alterations in M. tuberculosis and M. bovis BCG following treatment by NO or H2O2 also demonstrated that these bacteria up-regulate iron acquisition genes as a specific response to nitrosative and oxidative stresses (Jang et al., 2009; Voskuil et al., 2011). Additional proteins found to be exported in the ΔsigE mutant strain of C. pseudotuberculosis in response to NO, compared to the 1002 (wt) strain, might be indicative of a compensatory, more general response to stress (Table 1). This is reinforced by the fact that some of these proteins have already been shown to be part of other stimulons in C. pseudotuberculosis, regulated by heat shock, osmotic, or acidic stresses (Table 1).
In conclusion, our results demonstrate a role for the ECF sigma factor σE in resistance of an intracellular bacterium to nitrosative stress. While deletion of this sigma factor in C. pseudotuberculosis did contribute for altering the composition of this bacterium’s exoproteome in response to environmental cues, further studies remain necessary to better define SigE-responsive genes that account for the observed nitric oxide-susceptible phenotype of the ΔsigE strain.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from the Funding Agencies CNPq (Universal; Caprinos e Ovinos; Repensa) and FAPEMIG (Repensa; Pronex), in Brazil; and by The Medical Research Fund, in the UK.
Footnotes
References
Baird, G. J., and Fontaine, M. C. (2007). Corynebacterium pseudotuberculosis and its role in ovine caseous lymphadenitis. J. Comp. Pathol. 137, 179–210.
Carbon, S., Ireland, A., Mungall, C. J., Shu, S., Marshall, B., Lewis, S., Hub, AmiGO, and Web Presence Working Group. (2009). AmiGO: online access to ontology and annotation data. Bioinformatics 25, 288–289.
Carvalho, N. B., Oliveira, F. S., Durães, F. V., de Almeida, L. A., Flórido, M., Prata, L. O., Caliari, M. V., Appelberg, R., and Oliveira, S. C. (2011). Toll-like receptor 9 is required for full host resistance to Mycobacterium avium infection but plays no role in induction of Th1 responses. Infect. Immun. 79, 1638–1646.
Chitlaru, T., Gat, O., Grosfeld, H., Inbar, I., Gozlan, Y., and Shafferman, A. (2007). Identification of in vivo-expressed immunogenic proteins by serological proteome analysis of the Bacillus anthracis secretome. Infect. Immun. 75, 2841–2852.
Dorella, F. A., Pacheco, L. G., Oliveira, S. C., Miyoshi, A., and Azevedo, V. (2006). Corynebacterium pseudotuberculosis: microbiology, biochemical properties, pathogenesis and molecular studies of virulence. Vet. Res. 37, 201–218.
Dreisbach, A., van Dijl, J. M., and Buist, G. (2011). The cell surface proteome of Staphylococcus aureus. Proteomics 11, 3154–3168.
Ehrt, S., and Schnappinger, D. (2009). Mycobacterial survival strategies in the phagosome: defense against host stresses. Cell. Microbiol. 11, 1170–1178.
Feelisch, M. (1998). The use of nitric oxide donors in pharmacological studies. Naunyn Schmiedebergs Arch. Pharmacol. 358, 113–122.
Fontán, P. A., Aris, V., Alvarez, M. E., Ghanny, S., Cheng, J., Soteropoulos, P., Trevani, A., Pine, R., and Smith, I. (2008). Mycobacterium tuberculosis sigma factor E regulon modulates the host inflammatory response. J. Infect. Dis. 198, 877–885.
Helmann, J. D. (2002). The extracytoplasmic function (ECF) sigma factors. Adv. Microb. Physiol. 46, 47–110.
Jang, H. J., Nde, C., Toghrol, F., and Bentley, W. E. (2009). Microarray analysis of Mycobacterium bovis BCG revealed induction of iron acquisition related genes in response to hydrogen peroxide. Environ. Sci. Technol. 43, 9465–9472.
Jin, H., Agarwal, S., Agarwal, S., and Pancholi, V. (2011). Surface export of GAPDH/SDH, a glycolytic enzyme, is essential for Streptococcus pyogenes virulence. MBio 2, e00068–11.
Kazmierczak, M. J., Wiedmann, M., and Boor, K. J. (2005). Alternative sigma factors and their roles in bacterial virulence. Microbiol. Mol. Biol. Rev. 69, 527–543.
Manganelli, R., Voskuil, M. I., Schoolnik, G. K., and Smith, I. (2001). The Mycobacterium tuberculosis ECF sigma factor sigmaE: role in global gene expression and survival in macrophages. Mol. Microbiol. 41, 423–437.
Matta, S. K., Agarwal, S., and Bhatnagar, R. (2010). Surface localized and extracellular Glyceraldehyde-3-phosphate dehydrogenase of Bacillus anthracis is a plasminogen binding protein. Biochim. Biophys. Acta 1804, 2111–2120.
Mattinen, L., Nissinen, R., Riipi, T., Kalkkinen, N., and Pirhonen, M. (2007). Host-extract induced changes in the secretome of the plant pathogenic bacterium Pectobacterium atrosepticum. Proteomics 7, 3527–3537.
McKean, S. C., Davies, J. K., and Moore, R. J. (2007). Expression of phospholipase D, the major virulence factor of Corynebacterium pseudotuberculosis, is regulated by multiple environmental factors and plays a role in macrophage death. Microbiology 153, 2203–2211.
Moura-Costa, L. F., Paule, B. J., Azevedo, V., Freire, S. M., Nascimento, I., Schaer, R., Regis, L. F., Vale, V. L., Matos, D. P., Bahia, R. C., Carminati, R., and Meyer, R. (2002). Chemically defined synthetic medium for Corynebacterium pseudotuberculosis culture. Rev. Bras. Saúde e Produção Anim. 3, 1–9.
Nathan, C., and Shiloh, M. U. (2000). Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. U.S.A. 97, 8841–8848.
Ogawa, R., Pacelli, R., Espey, M. G., Miranda, K. M., Friedman, N., Kim, S. M., Cox, G., Mitchell, J. B., Wink, D. A., and Russo, A. (2001). Comparison of control of Listeria by nitric oxide redox chemistry from murine macrophages and NO donors: insights into listericidal activity of oxidative and nitrosative stress. Free Radic. Biol. Med. 30, 268–276.
Ohno, H., Zhu, G., Mohan, V. P., Chu, D., Kohno, S., Jacobs, W. R., and Chan, J. (2003). The effects of reactive nitrogen intermediates on gene expression in Mycobacterium tuberculosis. Cell. Microbiol. 5, 637–648.
Pacheco, L. G., Slade, S. E., Seyffert, N., Santos, A. R., Castro, T. L., Silva, W. M., Santos, A. V., Santos, S. G., Farias, L. M., Carvalho, M. A., Pimenta, A. M., Meyer, R., Silva, A., Scrivens, J. H., Oliveira, S. C., Miyoshi, A., Dowson, C. G., and Azevedo, V. (2011). A combined approach for comparative exoproteome analysis of Corynebacterium pseudotuberculosis. BMC Microbiol. 11, 12. doi:10.1186/1471-2180-11-12
Park, S., Youn, J., Kim, Y., Lee, S., Kim, Y., and Lee, H. (2008). Corynebacterium glutamicum sigmaE is involved in responses to cell surface stresses and its activity is controlled by the anti-sigma factor CseE. Microbiology 154, 915–923.
Pasztor, L., Ziebandt, A. K., Nega, M., Schlag, M., Haase, S., Franz-Wachtel, M., Madlung, J., Nordheim, A., Heinrichs, D. E., and Götz, F. (2010). Staphylococcal major autolysin (Atl) is involved in excretion of cytoplasmic proteins. J. Biol. Chem. 285, 36794–36803.
Pauling, J., Röttger, R., Tauch, A., Azevedo, V., and Baumbach, J. (2011). CoryneRegNet 6.0 – updated database content, new analysis methods and novel features focusing on community demands. Nucleic Acids Res. 40, 610–614.
Pumirat, P., Saetun, P., Sinchaikul, S., Chen, S. T., Korbsrisate, S., and Thongboonkerd, V. (2009). Altered secretome of Burkholderia pseudomallei induced by salt stress. Biochim. Biophys. Acta 1794, 898–904.
Rohde, K., Yates, R. M., Purdy, G. E., and Russell, D. G. (2007). Mycobacterium tuberculosis and the environment within the phagosome. Immunol. Rev. 219, 37–54.
Schaible, U. (2009). “Mycobacterium tuberculosis and his comrades,” in Intracellular Niches of Microbes, eds U. E. Schaible and A. Haas (Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA), 327–354.
Schnappinger, D., Ehrt, S., Voskuil, M. I., Liu, Y., Mangan, J. A., Monahan, I. M., Dolganov, G., Efron, B., Butcher, P. D., Nathan, C., and Schoolnik, G. K. (2003). Transcriptional adaptation of Mycobacterium tuberculosis within macrophages: insights into the phagosomal environment. J. Exp. Med. 198, 693–704.
Sengupta, N., Alam, S. I., Kumar, B., Kumar, R. B., Gautam, V., Kumar, S., and Singh, L. (2010). Comparative proteomic analysis of extracellular proteins of Clostridium perfringens type A and type C strains. Infect. Immun. 78, 3957–3968.
Staron, A., Sofia, H. J., Dietrich, S., Ulrich, L. E., Liesegang, H., and Mascher, T. (2009). The third pillar of bacterial signal transduction: classification of the extracytoplasmic function (ECF) sigma factor protein family. Mol. Microbiol. 74, 557–581.
Stefańska, I., Gierynska, M., Rzewuska, M., and Binek, M. (2010). Survival of Corynebacterium pseudotuberculosis within macrophages and induction of phagocytes death. Pol. J. Vet. Sci. 13, 143–149.
Talaat, A. M., Lyons, R., Howard, S. T., and Johnston, S. A. (2004). The temporal expression profile of Mycobacterium tuberculosis infection in mice. Proc. Natl. Acad. Sci. U.S.A. 101, 4602–4607.
Trost, E., Ott, L., Schneider, J., Schröder, J., Jaenicke, S., Goesmann, A., Husemann, P., Stoye, J., Dorella, F. A., Rocha, F. S., Soares, S. C., D’Afonseca, V., Miyoshi, A., Ruiz, J., Silva, A., Azevedo, V., Burkovski, A., Guiso, N., Join-Lambert, O. F., Kayal, S., and Tauch, A. (2010). The complete genome sequence of Corynebacterium pseudotuberculosis FRC41 isolated from a 12-year-old girl with necrotizing lymphadenitis reveals insights into gene-regulatory networks contributing to virulence. BMC Genomics 11, 728. doi:10.1186/1471-2164-11-728
Voskuil, M. I., Bartek, I. L., Visconti, K., and Schoolnik, G. K. (2011). The response of Mycobacterium tuberculosis to reactive oxygen and nitrogen species. Front. Microbiol. 2:105. doi:10.3389/fmicb.2011.00105
Voskuil, M. I., Schnappinger, D., Visconti, K. C., Harrell, M. I., Dolganov, G. M., Sherman, D. R., and Schoolnik, G. K. (2003). Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program. J. Exp. Med. 198, 705–713.
Keywords: Corynebacterium pseudotuberculosis, sigma factor, nitric oxide, inducible nitric oxide synthase
Citation: Pacheco LGC, Castro TLP, Carvalho RD, Moraes PM, Dorella FA, Carvalho NB, Slade SE, Scrivens JH, Feelisch M, Meyer R, Miyoshi A, Oliveira SC, Dowson CG and Azevedo V (2012) A role for sigma factor σE in Corynebacterium pseudotuberculosis resistance to nitric oxide/peroxide stress. Front. Microbio. 3:126. doi: 10.3389/fmicb.2012.00126
Received: 09 November 2011; Accepted: 17 March 2012;
Published online: 03 April 2012.
Edited by:
Martin G. Klotz, University of North Carolina at Charlotte, USAReviewed by:
Donald A. Bryant, The Pennsylvania State University, USAJohann Heider, Philipps-Universität Marburg, Germany
Copyright: © 2012 Pacheco, Castro, Carvalho, Moraes, Dorella, Carvalho, Slade, Scrivens, Feelisch, Meyer, Miyoshi, Oliveira, Dowson and Azevedo. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Vasco Azevedo, Laboratório de Genética Celular e Molecular (Q3-259), Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha, Belo Horizonte, Minas Gerais 31270-901, Brazil. e-mail: vasco@icb.ufmg.br