 Katrin Zurfluh1
Katrin Zurfluh1 Juan Wang
Juan Wang Magdalena Nüesch-Inderbinen
Magdalena Nüesch-Inderbinen Séamus Fanning
Séamus Fanning Roger Stephan
Roger Stephan- 1Institute for Food Safety and Hygiene, Vetsuisse Faculty, University of Zurich, Zürich, Switzerland
- 2UCD Centre for Food Safety, School of Public Health, Physiotherapy and Population Science, UCD Centre for Molecular Innovation and Drug Discovery, University College Dublin, Dublin, Ireland
- 3Institute of Food, Nutrition and Health, Swiss Federal Institute of Technology in Zürich, Zürich, Switzerland
Objectives: The purpose of this study was to characterize sets of extended-spectrum β-lactamases (ESBL)-producing Enterobacteriaceae collected longitudinally from different flocks of broiler breeders, meconium of 1-day-old broilers from theses breeder flocks, as well as from these broiler flocks before slaughter.
Methods: Five sets of ESBL-producing Escherichia coli were studied by multi-locus sequence typing (MLST), phylogenetic grouping, PCR-based replicon typing and resistance profiling. The blaCTX-M-1-harboring plasmids of one set (pHV295.1, pHV114.1, and pHV292.1) were fully sequenced and subjected to comparative analysis.
Results: Eleven different MLST sequence types (ST) were identified with ST1056 the predominant one, isolated in all five sets either on the broiler breeder or meconium level. Plasmid sequencing revealed that blaCTX-M-1 was carried by highly similar IncI1/ST3 plasmids that were 105 076 bp, 110 997 bp, and 117 269 bp in size, respectively.
Conclusions: The fact that genetically similar IncI1/ST3 plasmids were found in ESBL-producing E. coli of different MLST types isolated at the different levels in the broiler production pyramid provides strong evidence for a vertical transmission of these plasmids from a common source (nucleus poultry flocks).
Introduction
One of the currently most important antibiotic resistance mechanisms in Enterobacteriaceae is based on plasmid-mediated production of extended-spectrum β-lactamases (ESBLs) which inactivate β-lactam-antibiotics including cephalosporins and monobactams by hydrolyzing their β-lactam ring (Frère et al., 1992). ESBLs are classified according to their primary sequences and substrate profiles into different families such as the TEM-, the SHV-, the OXA-, and the CTX-M -family (Bush and Jacoby, 2010). Currently, CTX-M enzymes are the most widespread ESBLs (Naseer and Sundsfjord, 2011). Of the more than 150 different CTX-M sequence types (ST) that are known and filed in the Lahey database1, CTX-M-1 predominates in Escherichia coli isolated from food-producing animals and foods in Europe (EFSA, 2011).
Recently, several reports highlighted the dissemination of ESBL-producing E. coli in poultry in several countries across Europe (Smet et al., 2008; Cortés et al., 2010; Randall et al., 2011; Wasyl et al., 2012) or in chicken meat (Leverstein-van Hall et al., 2011; Overdevest et al., 2011; Kola et al., 2012). A high fecal prevalence of blaCTX-M-1-harboring E. coli has also been reported for poultry flocks in Switzerland (Endimiani et al., 2012; Geser et al., 2012). Moreover, CTX-M-1-producing E. coli have also been found both on local and imported retail poultry meat (Abgottspon et al., 2014; Vogt et al., 2014).
The high occurrence of ESBL-producing bacteria in Swiss poultry flocks, however, cannot be explained by the use of high amounts of antimicrobials for broilers, since on average only 1 of 10 poultry flocks need to be treated with antibiotics and when treatment is indicated, fluoroquinolones and not β-lactam-antibiotics are the drugs of choice (Büttner et al., 2013).
As hypothesized by others (Dierikx et al., 2013; Agersø et al., 2014; Nilsson et al., 2014), one possible explanation for this situation could be the existence of a common source (nucleus poultry flocks), disseminating ESBL-producers vertically along the poultry chain over large geographical regions. ESBL-producing Enterobacteriaceae might be imported by one-day-old broiler breeders and then spread from the top to the bottom of the production pyramid. We hypothesize that a specific E. coli type is unlikely to be responsible for this occurrence, but that a limited number of different plasmids transferred between different E. coli types by conjugation are involved.
The purpose of this study was to characterize sets of ESBL-producing Enterobacteriaceae collected longitudinally from different flocks of broiler breeders, meconium of one-day-old broilers from theses breeder flocks, as well as from these broiler flocks before slaughter, and to sequence plasmids harboring the blaESBL genes from one set of isolates.
Materials and Methods
Bacterial Isolates
From July 2013 through May 2014 boot socks (gauze socks walked over the length of a broiler chicken house) of parental broiler breeder flocks (from grandparent breeders from France), meconium of 1-day-old broilers from these breeder flocks and boot socks of these broilers flocks taken before slaughter were sampled longitudinally through the production pyramid. Boot socks and the meconium were enriched for 24 h at 37°C in 250 and 10 ml of EE Broth (BD, Franklin Lakes, NJ, USA), respectively. Thereafter, one loopful each of the enrichment cultures was inoculated onto chromogenic Brilliance ESBL agar (Oxoid, Hampshire, UK) and incubated at 37°C for 24 h under aerobic conditions. All colonies with different color and morphology were picked from the selective plates and sub-cultured on triple sugar iron-agar (TSI) agar (Oxoid) at 37°C for 24 h. Oxidase-negative isolates were subjected to identification by API ID 32 E (bioMérieux, Marcy l’Etoile, France). One isolate per sample was selected for further characterization.
Antibiotic Susceptibility Testing and Phenotypic ESBL Detection
Susceptibility testing was performed by agar diffusion methods, using antibiotic disks (Becton Dickinson and Company, Maryland, USA) and ESBL ellipsometer-test (E-test) strips (bioMérieux, Marcy l’Etoile, France), according to the manufacturers’ protocols. Results were interpreted according to the criteria of the Clinical and Laboratory Standards Institute (CLSI, 2011).
Identification of blaESBL Genes
Bacterial strains confirmed as producing ESBLs were further analyzed by PCR. DNA was purified using a standard heat lysis protocol (Sambrook and Russel, 2006). Thereafter, five specific primer sets (custom-synthesized by Microsynth, Balgach, Switzerland) were used to screen for β-lactamase-encoding genes belonging to the blaTEM, blaSHV, and blaCTX-M families (Pitout et al., 1998; Geser et al., 2012).
Resulting amplicons were purified using the GenEluteTM PCR Clean-Up (Sigma–Aldrich, Buchs, Switzerland) according to the manufacturer’s recommendations. Custom-sequencing was performed by Microsynth (Balgach, Switzerland) and the nucleotide- and translated protein-sequences were analyzed with CLC Main Workbench 7.0.2 (CLC bio, Aarhus, Denmark). For database searches the BLASTN program of NCBI2 was used.
MLST of ESBL Producers
Internal fragments of seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were sequenced (Wirth et al., 2006) and allele and ST were assigned in accordance with the E. coli multi-locus sequence typing (MLST) website3.
Determination of E. coli Phylogenetic Groups
Escherichia coli isolates were subjected to phylogenetic grouping by PCR as described previously (Clermont et al., 2000). Thereby, E. coli strains are assigned to four main phylogenetic groups (A, B1, B2, and D). Groups A and B1 contain commensal isolates and members of the group B2 and D are classified as virulent extra-intestinal strains which often carry pathogenicity associated genes.
PCR-Based Replicon Typing
Plasmid incompatibility (Inc) types were determined by PBRT (Carattoli et al., 2005; Villa et al., 2010), using genomic DNA as template.
Conjugation Mating Experiments
Conjugation experiments were performed with the plasmid-free recipient strain E. coli HK225 (Strepr, Rifr; Kayser et al., 1982). Briefly, single colonies of the donor and recipient were inoculated in LB broth (Difco Laboratories) and grown overnight at 37°C. Subsequently, equal volumes of the donor and recipient cultures were mixed and incubated overnight at 37°C without shaking. Serial dilutions were then plated on LB agar (Difco Laboratories) selection plates supplemented with a combination of 600 μg/ml streptomycin (Sigma–Aldrich, Buchs, Switzerland) 100 μg/ml rifampicin (Sigma–Aldrich) and 10 μg/ml cefotaxime (Sigma–Aldrich).
The conjugation frequency per donor was determined by plating serial dilutions of the mating on selective plates on which the donor strain and the transconjugant can grow (LB-agar supplemented with 10 μg/ml cefotaxime) as well as on plates on which only the transconjugants can grow (LB-agar supplemented with 600 μg/ml streptomycin, 100 μg/ml rifampicin, and 10 μg/ml cefotaxime). The transfer frequency was calculated as the quotient of the number of transconjugants over the number of transconjugants plus donors.
Plasmid Extraction and Sequencing
The plasmids pHV295.1, pHV114.1, and pHV292.1 were purified using the Qiagen Large-Construct Kit (Qiagen, Hombrechtikon, Switzerland) according the manufacturer’s protocol. The complete plasmid sequence was determined using the Pacific Biosciences SMRT sequencing approach (Functional Genomics Center, Zurich, Switzerland) applying P4/C2 chemistry on an RSII device. A 120 min movie was recorded from a 10 kb insert library per cell.
Bioinformatics
Sequence assembly was carried out using the SMRT-Analysis software version 2.0 with the RS_HGAP_Assembly.1 algorithm and default settings. The plasmid sequence was automatically annotated using the online Rapid Annotation Subsequencing Technology (RAST; Aziz et al., 2008) and CLC Main Workbench Version 7.0.2 (CLC bio, Aarhus, Denmark). Automated annotation was manually refined.
Nucleotide Sequence
The GenBank accession numbers for pHV295.1, pHV114.1, and pHV292.1 plasmids are KM377238, KM377239, and KM377240, respectively.
Results
Phenotypic and Genotypic Characteristics of the ESBL-Producing Isolates
Five sets of ESBL-producing Enterobacteriaceae collected longitudinally from five different flocks of broiler breeders, meconium of 1-day-old broilers from these breeder flocks as well as from these broiler flocks before slaughter were further characterized. Species identification revealed that all were E. coli and all harbored blaCTX-M-1 genes (Table 1). Phylogenetic grouping and MLST analysis showed that different E. coli types could be identified at the investigated production levels in all five sets, suggestive of a non-clonal bacterial composition. E. coli B1 ST1056 was isolated in all five sets either in the broiler breeder samples or in the meconium samples. Eleven of the 15 isolates belonged to commensal phylogenetic groups A and B1; only four isolates belonged to the virulent extra-intestinal E. coli group B2 and D. In total, 11 different MLST ST were detected (Table 1).
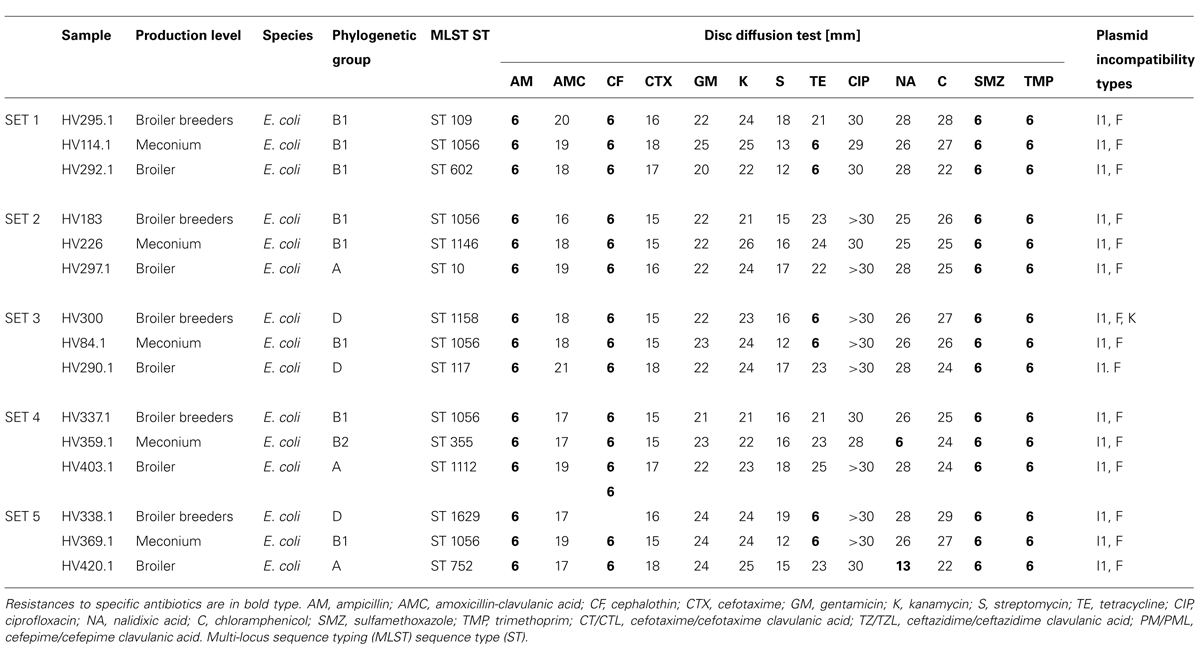
TABLE 1. Phenotypic and genotypic characteristics of the extended-spectrum β-lactamases (ESBL)-producing isolates from flocks of broiler breeder, meconium of 1-day-old broilers and broilers.
Phenotypic antibiotic resistance characterization indicated that all isolates were resistant to sulfamethoxazole and trimpethoprim in addition to the β-lactam antibiotics tested. Six isolates (two from the broiler breeders, three from meconium, one from a broiler) and two isolates (one each cultured from meconium and a broiler) were additionally resistant to tetracycline or nalidixic acid, respectively (Table 1).
PCR-based replicon typing showed that all isolates from the five sets possessed at least two plasmids: an IncI1 and an IncF replicon type plasmid. One isolate from broiler breeders (HV300) also carried an IncK plasmid (Table 1).
Conjugation Experiments, Plasmid Sequencing, and Comparative Analysis
Conjugation experiments were performed with the isolates from set 1 (comprising isolates HV295.1, HV114.1, and HV292.1). The blaCTX-M-1-carrying plasmids could be transferred to the recipient strain E. coli HK225 with conjugation frequencies of 1.35 × 10-3, 4.63 × 10-3, and 1.29 × 10-5 per donor cell, respectively. Whole plasmid sequencing revealed that all three plasmids belong to the incompatibility group IncI1 of plasmid pMLST ST3 type (García-Fernández et al., 2008). Plasmids pHV295.1, pHV114.1, and pHV292.1 were 105 076, 110 997, and 117 269 bp in size with overall G+C contents of 50.5, 51 and 51.3%, respectively. All three plasmids presented a similar backbone structure (nucleotide similarity 99%) that encoded for initiation of plasmid replication (oriT operon), conjugative transfer (traABCD), plasmid maintenance and stability (e.g., yefA, parAB, impABC, ssb,) and pil operon (Sampei et al., 2010). The latter encodes for a group of proteins forming a type IV pilus involved in the formation of donor-recipient cell aggregates (Kim and Komano, 1997). All of these modules were typical of those identified previously in IncI1 types and were also found on the IncI1 reference plasmid R64 (GenBank accession number AP005147.1; Figure 1).
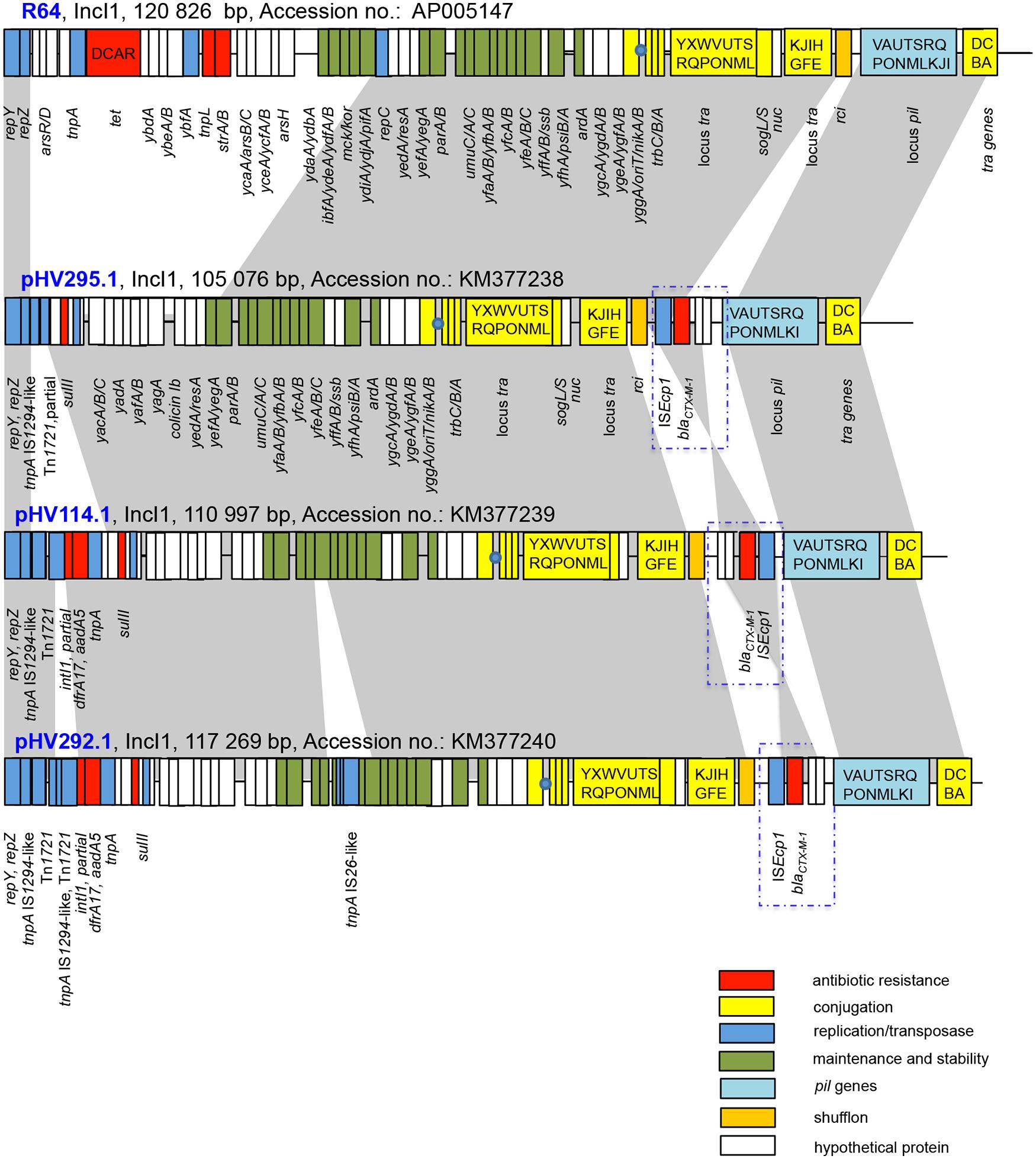
FIGURE 1. Comparative analysis of pHV295.1, pHV114.1, and pHV292.1. Major structural features of the IncI1 plasmids are shown in comparison with the IncI1 reference plasmid R64 (Accession no. AP005147). Gray shaded areas indicate homologies in the plasmid scaffold regions. Red boxes mark antibiotic resistance genes. Conjugation-related genes are shown with capital letters in yellow boxes. Blue boxes indicate transposon-, integron-, or replication associated genes. Green boxes denote maintenance- and stability-related genes. White boxes indicate hypothetical proteins and light blue boxes show pil genes. The origin of transfer oriT is shown as a blue circle. The figure is not drawn to scale.
Comparative analysis of the three plasmids sequenced in this study showed that they were very similar over the complete sequence length (Figure 1). Two regions are of main interest: (i) the region proximal of repZ (pHV295.1: 8.2 kb; pHV114.1: 14.1 kb, and pHV292.1: 17.6 kb) and (ii) a 2.7 kb module containing ISEcp1-blaCTX-M-1 encoding the resistance to broad-spectrum cephalosporins inserted between rci and pilVA.
The region between repZ and yacA is the most variable region in the three plasmids and it consists of a number of mobile elements such as (partial) transposases, (partial) integrons and IS-elements. The plasmid isolated on the level of broiler breeders (pHV295.1) carried in this region a sul2 gene that encodes for a protein leading to resistance to sulfonamides (Rådström and Swedberg, 1988). The other two plasmids (from meconium and broilers) contained in the corresponding region of the plasmid section, additionally to the sul2 gene, the resistance genes dfrA17 and aadA5 (trimethoprim and streptomycin/spectinomycin resistance; White et al., 2000) embedded in a partial class 1 integron devoid of the complete 3′-conserved structure.
A second antimicrobial resistance region identified in all three plasmids was the ISEcp1-blaCTX-M-1 module. It is inversely inserted in the plasmid isolated from the meconium sample (pHV114.1; see Figure 1).
Another difference between pHV295.1, pHV114.1, and pHV292.1 is the insertion of an IS66-like element into pHV292.1 leading to the disruption of umuC, a gene of the UV protection operon (Perry et al., 1985).
Discussion
The hypothesis of a non-clonal dissemination of ESBL-encoding genes in Swiss poultry flocks is strengthened by the fact that ESBL-producing E. coli with different MLST ST and belonging to different phylogroups were isolated in the poultry production pyramid. Furthermore, the sequenced plasmids isolated from these E. coli were all highly similar. Moreover, sequences of three further IncI1 plasmids from ESBL-producing E. coli isolated in 2012 from broiler flocks in Switzerland (Wang et al., 2014) showed remarkable similarities with those reported here. All had the same backbone structures as well as the identical ISEcp1-blaCTX-M-1 module and harbored also an incomplete class 1 integron containing a gene cassette composed of the trimethoprim and streptomycin/spectinomycin resistance genes.
When Leverstein-van Hall et al. (2011) investigated retail chicken meat and poultry samples, they identified a blaCTX-M-1 gene located mainly on IncI1 plasmids, but in contrast to our study, these plasmids belonged to the pMLST ST7. Similarly, in two Scandinavian studies vertical transmission of extended-spectrum cephalosporin resistant Enterobacteriaceae, mainly CMY-2-producing E. coli was reported (Agersø et al., 2014; Nilsson et al., 2014). These observed differences in the latter studies might be explainable by the different origin of the broiler breeder grandparents since the grandparents from the Swedish and Danish study originated from Sweden while those investigated in this study were from France. Dahmen et al. (2012) have confirmed earlier that IncI1/ST3 plasmids mainly contribute to the dissemination of blaCTX-M-1 in France. Furthermore, it has been shown, that IncI1 plasmids harboring blaCTX-M-1 show no or negligible fitness cost in E. coli and that these plasmids can persist in the absence of antimicrobial selection (Fischer et al., 2014). It is also known that plasmid-encoded elements are beneficial for stable inheritance of the plasmids, such as toxin/antitoxin systems or type IV pili which are beneficial to the bacteria for invasion and adhesion to the host gut (Carattoli, 2009).
Vertical transmission of ESBL-producing strains with these specific plasmids through the broiler production pyramid is only one possible explanation for their dissemination. Additionally, horizontal transmission of ESBL-producers and their plasmids within a poultry flock (coprophagic birds) plays another important role in their spread (Agersø et al., 2014; Nilsson et al., 2014). This factor might further explain the high diversity of different E. coli MLST types found in our study. The presence of E. coli of ST 1056 in all five sets, however, also highlights that some E. coli types might be more adapted to the poultry gut. This specific E. coli MLST has also been detected previously on Swiss retail poultry meat (Abgottspon et al., 2014). Moreover, we detected E. coli ST10 and ST117, two ST which are also considered poultry-associated (Leverstein-van Hall et al., 2011; Voets et al., 2013).
The final aspect to be considered focuses on the potential for co-selection via antimicrobial agents other than cephalosporins. Agersø et al. (2014) showed recently that to some extent extended-spectrum cephalosporinases-producing E. coli were selected by the use of aminopenicillins. The plasmids sequenced in this study harbored all of the sul2 gene, which encodes resistance to sulfonamides. However, since in Switzerland at an average only 1 of 10 poultry flocks have to be treated with antibiotics and if so, fluoroquinolones and not β-lactam-antibiotics or sulfonamides are applied, the co-selection aspect may not really contribute to a further spread of these plasmids.
In conclusion, this study provides strong evidence that highly similar IncI1/ST3 plasmids harboring the blaCTX-M-1 gene are transmitted vertically in the broiler production pyramid from the top to the bottom with little evidence of any antimicrobial selection pressure. Intervention actions to reduce the risk of spreading ESBL-producing E. coli containing these plasmids have mainly to be focused on the top of the poultry production pyramid (nucleus poultry flock level). This situation should now be re-assessed in light of these findings.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Alessandra Carattoli for providing the control strains for the PCR based replicon typing of plasmids and Helga Abgottspon and Gianna Jakobi for technical assistance. This work was partly funded by the Swiss poultry association (CH-IGG).
Footnotes
- ^http://www.lahey.org/studies/
- ^http://www.ncbi.nlm.nih.gov/blast/
- ^http://mlst.warwick.ac.uk/mlst/dbs/Ecoli
References
Abgottspon, H., Stephan, R., Bagutti, C., Brodmann, P., Hächler, H., and Zurfluh, K. (2014). Characteristics of extended-spectrum cephalosporin-resistant Escherichia coli isolated from Swiss and imported poultry meat. J. Food Prot. 77, 112–115. doi: 10.4315/0362-028X.JFP-13-120
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Agersø, Y., Jensen, J. D., Hasman, H., and Pedersen, K. (2014). Spread of extended spectrum cephalosporinase-producing Escherichia coli clones and plasmids from parent animals to broilers and to broiler meat in a production without use of cephalosporins. Foodborne Pathog. Dis. 11, 740–746. doi: 10.1089/fpd.2014.1742
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R.,et al. (2008). The RAST Server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bush, K., and Jacoby, G. A. (2010). Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 54, 969–976. doi: 10.1128/AAC.01009-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Büttner, S., Flurina, S., Müntener, C., Jäggi, M., and Overesch, G. (2013). Report on the Sales of Antibiotics for Veterinary Use and Antibiotic Resistance Monitoring of Livestock in Switzerland. Bern: Federal Veterinary Office.
Carattoli, A. (2009). Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 2227–2238. doi: 10.1128/AAC.01707-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Clermont, O., Bonacorsi, S., and Bingen, E. (2000). Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 66, 4555–4558. doi: 10.1128/AEM.66.10.4555-4558.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
CLSI. (2011). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-First Informational Supplement, Vol. 31. Wayne, PA: Clinical and Laboratory Standards Institute.
Cortés, P., Blanc, V., Mora, A., Dahbi, G., Blanco, J. E., Blanco, M.,et al. (2010). Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Environ. Microbiol. 76, 2799–805. doi: 10.1128/AEM.02421-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dahmen, S., Haenni, M., and Madec, J. (2012). IncI1/ST3 plasmids contribute to the dissemination of the blaCTX-M-1 gene in Escherichia coli from several animal species in France. J. Antimicrob. Chemother. 67, 3011–3012. doi: 10.1093/jac/dks308
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dierikx, C. M., van der Goot, J. A., Smith, H. E., Kant, A., and Mevius, D. J. (2013). Presence of ESBL/AmpC -producing Escherichia coli in the broiler production pyramid: a descriptive study. PLoS ONE 8:e79005. doi: 10.1371/journal.pone.0079005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
EFSA. (2011). Panel on biological hazards. Scientific opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 9, 2322. doi: 10.2903/j.efsa.2011.2322
Endimiani, A., Rossano, A., Kunz, D., Overesch, G., and Perreten, V. (2012). First countrywide survey of third-generation cephalosporin-resistant Escherichia coli from broilers, swine, and cattle in Switzerland. Diagn. Microbiol. Infect. Dis. 73, 31–38. doi: 10.1016/j.diagmicrobio.2012.01.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fischer, E. A. J., Dierikx, C. M., van Essen-Zandbergen, A., van Roermund, H. J. W., Mevius, D. J., Stegeman, A.,et al. (2014). The IncI1 plasmid carrying the blaCTX-M-1 gene persists in in vitro culture of a Escherichia coli strain from broilers. BMC Microbiol. 14:77. doi: 10.1186/1471-2180-14-77
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frère, J. M., Nguyen-Diste’ che, M., Coyette, J., and Joris, B. (1992). “Mode of action: interaction with the penicillian-binding proteins,” in The Chemistry of ß-Lactams, ed. M. I. Page (London: Chapman & Hall), 148–197.
García-Fernández, A., Chiaretto, G., Bertini, A., Villa, L., Fortini, D., Ricci, A.,et al. (2008). Multilocus sequence typing of IncI1 plasmids carrying extended-spectrum beta-lactamases in Escherichia coli and Salmonella of human and animal origin. J. Antimicrob. Chemother. 61, 1229–1233. doi: 10.1093/jac/dkn131
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Geser, N., Stephan, R., and Hächler, H. (2012). Occurrence and characteristics of extended-spectrum b-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 8:21. doi: 10.1186/1746-6148-8-21
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kayser, F. H., Morenzoni, G., and Homberger, F. (1982). Activity of cefoperazone against ampicillin-resistant bacteria in agar and broth dilution tests. Antimicrob. Agents Chemother. 22, 15–22. doi: 10.1128/AAC.22.1.15
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, S. R., and Komano, T. (1997). The plasmid R64 thin pilus identified as a type IV pilus. J. Bacteriol. 179, 3594–3603.
Kola, A., Kohler, C., Pfeifer, Y., Schwab, F., Kühn, K., Schulz, K.,et al. (2012). High prevalence of extended-spectrum-β-lactamase-producing Enterobacteriaceae in organic and conventional retail chicken meat, Germany. J. Antimicrob. Chemother. 67, 2631–2634. doi: 10.1093/jac/dks295
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Leverstein-van Hall, M. A., Dierikx, C. M., Cohen Stuart, J., Voets, G. M., van den Munckhof, M. P., van Essen-Zandbergen, A.,et al. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880. doi: 10.1111/j.1469-0691.2011.03497.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Naseer, U., and Sundsfjord, A. (2011). The CTX-M conundrum: dissemination of plasmids and Escherichia coli clones. Microb. Drug Resist. 17, 83–97. doi: 10.1089/mdr.2010.0132
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nilsson, O., Börjesson, S., Landén, A., and Bengtsson, B. (2014). Vertical transmission of Escherichia coli carrying plasmid-mediated AmpC (pAmpC) through the broiler production pyramid. J. Antimicrob. Chemother. 69, 1497–1500. doi: 10.1093/jac/dku030
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Overdevest, I., Willemsen, I., Rijnsburger, M., Eustace, A., Xu, L., Hawkey, P.,et al. (2011). Extended-spectrum β-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands. Emerg. Infect. Dis. 17, 1216–1222. doi: 10.3201/eid1707.110209
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Perry, K. L., Elledge, S. J., Mitchell, B. B., Marsh, L., and Walker, G. C. (1985). umuDC and mucAB operons whose products are required for UV light- and chemical-induced mutagenesis: UmuD, MucA, and LexA proteins share homology. Proc. Natl. Acad. Sci. U.S.A. 4, 1054–1058.
Pitout, J. D. D., Thomson, K. S., Hanson, N. D., Ehrhardt, A. F., Moland, E. S., and Sanders, C. C. (1998). Beta-lactamases responsible for resistance to expanded-spectrum cephalosporins in Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis isolates recovered in South Africa. Antimicrob. Agents Chemother. 42, 1350–1354.
Rådström, P., and Swedberg, G. (1988). RSF1010 and a conjugative plasmid contain sulII, one of two known genes for plasmid-borne sulfonamide resistance dihydropteroate synthase. Antimicrob. Agents Chemother. 32, 1684–1692. doi: 10.1128/AAC.32.11.1684
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Randall, L. P., Clouting, C., Horton, R. A., Coldham, N. G., Wu, G., Clifton-Hadley, F. A.,et al. (2011). Prevalence of Escherichia coli carrying extended-spectrum β-lactamases (CTX-M and TEM-52) from broiler chickens and turkeys in Great Britain between 2006 and 2009. J. Antimicrob. Chemother. 66, 86–95. doi: 10.1093/jac/dkq396
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sambrook, J., and Russel, D. W. (2006). The Condensed Protocols from Molecular Cloning: A Laboratory Manual. New York: Cold Spring Harbor Laboratory Press.
Sampei, G., Furuya, N., Tachibana, K., Saitou, Y., Suzuki, T., Mizobuchi, K.,et al. (2010). Complete genome sequence of the incompatibility group I1 plasmid R64. Plasmid 64, 92–103. doi: 10.1016/j.plasmid.2010.05.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Smet, A., Martel, A., Persoons, D., Dewulf, J., Heyndrickx, M., Catry, B.,et al. (2008). Diversity of extended-spectrum beta-lactamases and class C beta-lactamases among cloacal Escherichia coli isolates in Belgian broiler farms. Antimicrob. Agents Chemother. 52, 1238–1243. doi: 10.1128/AAC.01285-07
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Villa, L., García-Fernández, A., Fortini, D., and Carattoli, A. (2010). Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 65, 2518–2529. doi: 10.1093/jac/dkq347
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Voets, G. M., Fluit, A. C., Scharringa, J., Schapendonk, C., van den Munckhof, T., Leverstein-van Hall, M. A.,et al. (2013). Identical plasmid AmpC beta-lactamase genes and plasmid types in E. coli isolates from patients and poultry meat in the Netherlands. Int. J. Food Microbiol. 167, 359–362. doi: 10.1016/j.ijfoodmicro.2013.10.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vogt, D., Overesch, G., Endimiani, A., Collaud, A., Thomann, A., and Perreten, V. (2014). Occurrence and genetic characteristics of third-generation cephalosporin-resistant Escherichia coli in Swiss retail meat. Microb. Drug Resist. doi: 10.1089/mdr.2013.0210 [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, J., Stephan, R., Power, K., Yan, Q., Hächler, H., and Fanning, S. (2014). Nucleotide sequences of 16 transmissible plasmids identified in nine multidrug-resistant Escherichia coli isolates expressing an ESBL phenotype isolated from food-producing animals and healthy humans. J. Antimicrob. Chemother. 69, 2658–2668. doi: 10.1093/jac/dku206
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wasyl, D., Hasman, H., Cavaco, L. M., and Aarestrup, F. M. (2012). Prevalence and characterization of cephalosporin resistance in nonpathogenic Escherichia coli from food-producing animals slaughtered in Poland. Microb. Drug Resist. 18, 79–82. doi: 10.1089/mdr.2011.0033
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
White, P. A., McIver, C. J., Deng, Y., and Rawlinson, W. D. (2000). Characterisation of two new gene cassettes, aadA5 and dfrA17. FEMS Microbiol. Lett. 182, 265–269. doi: 10.1111/j.1574-6968.2000.tb08906.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L. H.,et al. (2006). Sex and virulence in Escherichia coli: an evolutionary perspective. Mol. Microbiol. 60, 1136–1151. doi: 10.1111/j.1365-2958.2006.05172.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: E. coli, plasmid sequencing, CTX-M-1, poultry production pyramid, IncI1, conjugation
Citation: Zurfluh K, Wang J, Klumpp J, Nüesch-Inderbinen M, Fanning S and Stephan R (2014) Vertical transmission of highly similar blaCTX-M-1-harboring IncI1 plasmids in Escherichia coli with different MLST types in the poultry production pyramid. Front. Microbiol. 5:519. doi: 10.3389/fmicb.2014.00519
Received: 07 August 2014; Accepted: 17 September 2014;
Published online: 30 September 2014.
Edited by:
Kornelia Smalla, Julius Kühn-Institut – Federal Research Centre for Cultivated Plants, GermanyReviewed by:
Yuji Morita, Aichi Gakuin University, JapanSiddharth Kaushal Tripathi, University of Mississippi, USA
Copyright © 2014 Zurfluh, Wang, Klumpp, Nüesch-Inderbinen, Fanning and Stephan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roger Stephan, Institute for Food Safety and Hygiene, Vetsuisse Faculty, University of Zurich, Winterthurerstr 272, CH-8057 Zurich, Switzerland e-mail: stephanr@fsafety.uzh.ch