 Faham Khamesipour
Faham Khamesipour Hassan Momtaz
Hassan Momtaz Morteza Azhdary Mamoreh3
Morteza Azhdary Mamoreh3- 1Young Researchers and Elite Club, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
- 2Department of Microbiology, Faculty of Veterinary Medicine, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
- 3Faculty of Veterinary Medicine, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
A total of 30 Pasteurella multocida strains isolated from 333 pneumonic and apparently health slaughter cattle were examined for capsule biosynthesis genes and 23 virulence-associated genes by polymerase chain reaction (PCR). The disc diffusion technique was used to determine antimicrobial resistance profiles among the isolates. Of the isolates, 23 belonged to capsular type A, 5 to capsular type D and two isolates were untypeable. The distribution of the capsular types in pneumonic lungs and in apparently health lungs was statistically similar. All virulence genes tested were detected among the isolates derived from pneumonic lungs; whereas isolates derived from apparently health lungs carried 16 of the 23 genes. The frequently detected genes among isolates from pneumonic lungs were exbD, hgbA, hgbB, ompA, ompH, oma87, and sodC; whereas tadD, toxA, and pmHAS genes occurred less frequently. Most of the adhesins and superoxide dismutases; and all of the iron acquisition and protectin proteins occurred at significantly (p ≤ 0.05) higher frequencies in isolates from pneumonic lungs. Isolates from apparently healthy lungs didn't carry the following genes; hsf-1, hsf-2, tadD, toxA, nanB, nanH, and pmHAS. One adhesion (hsf-1) and two iron acquisition (exbD and tonB) genes occurred at significantly (p ≤ 0.05) higher frequencies among capA isolates. All the P. multocida isolates were susceptible to ciprofloxacin, co-trimoxazole, doxycycline, enrofloxacin, nitrofurantoin, and tetracyclines. Different proportions of the isolates were however resistant to ampicillin, amoxicillin, erythromycin, lincomycin, penicillin, rifampin, streptomycin, and florfenicol. Our results reveal presence of virulence factors (VFs) in P. multocida strains isolated from symptomatic and asymptomatic bovids. A higher frequency of the factors among isolates from symptomatic study animals may suggest their role in pathogenesis of P. multocida-associated bovine respiratory disease (BRD). The results further reveal occurrence of antimicrobial resistance among some isolates. Control strategies for this pathogen, which could include development of an effective vaccine, are warranted so as to mitigate the social and economic consequences attributable to natural infections with this bacterium.
Introduction
Cattle rearing is one of the important sources of income in Iran, involving both dairy and beef breeds. The sector faces a number of constraints ranging from limited feed resources to diseases. Of the diseases, those caused by infectious agents are of great importance which include bacteria and viruses affecting the respiratory system (Hemmatzadeh et al., 2001; Haji Hajikolaei and Seyfi Abad Shapouri, 2007; Sakhaee et al., 2009). The most important bacteria that play a role in pneumonia include: Mannheimia haemolytica, Pasteurella multocida, and Haemophilus somnus pneumonia, which presence of these bacteria in pneumonia lesions of slaughtered cattle around Iran have also been reported (Haji Hajikolaei et al., 2010).
Bovine respiratory disease (BRD) is a significant cause of morbidity and mortality among beef cattle in the world (Dagleish et al., 2010; Hotchkiss et al., 2010; Portis et al., 2012). Among others, Pasteurella multocida has been identified as a major bacterial etiologic agent for this disease (Confer, 2009; Griffin et al., 2010). It is a zoonotic Gram negative bacterium responsible for a range of infections in domestic animals causing substantial economic losses (Steen et al., 2010). The organism causes fowl cholera in domestic and wild birds, bronchopneumonia and hemorrhagic septicemia in bovids, atrophic rhinitis in porcines and snuffles in rabbits (Mannheim, 1984; Hunt et al., 2000). Most human infections with P. multocida result from dog and cat bites, but infections through the respiratory tract may also occur (Hubbert and Rosen, 1970).
Several host and pathogen-specific attributes do determine the outcome of infections caused by P. multocida (Verma et al., 2013). Of the pathogen factors important ones include the capsular and virulence-associated genes (Katsuda et al., 2013). These virulence factors (VFs) and outer membrane proteins are important for pathogenesis, functionality, protective immunity and vaccine development against P. multocida infections (Harper et al., 2006; Hatfaludi et al., 2010). Based on capsular antigens, P. multocida strains are differentiated into five serogroups i.e., type A causing fowl cholera pathogen and bovine shipping fever, type B causing hemorrhagic fever in ungulates, type D causing atrophic rhinitis in swine, type E, an African serotype, infecting cattle and buffalo; and type F also causing fowl cholera (Carter, 1955, 1961, 1967; Rimler and Rhoades, 1987). Virulence associated genes described for P. multocida isolates and their examples include adherence and colonization factors (ptfA, fimA, hsf-1, hsf-2, pfhA, and tadD), iron-regulated and acquisition proteins(exbB, exbD, tonB, hgbA, hgbB, and Fur), extracellular enzymes such as neuraminidase (nanB and nanH), hyaluronidase (pmHAS) and superoxide dismutases (soda, sodC, and tbpA), toxins (toxA), lipopolysaccharides (LPS), capsule and a variety of outer membrane proteins such as protectins (ompA, omph, oma87, and plpB) (Katoch et al., 2014).
Increased use of antibiotics in modern animal production has been associated with emergence of antimicrobial resistant bacteria with potential for transfer of resistance from animals to humans (Witte, 1998). As a result, antimicrobial resistance among bacterial pathogens has of recent become a big problem in both the veterinary and human medicine fields (Levy, 1998; Caprioli et al., 2000; Kehrenberg et al., 2001; White et al., 2002; Shea, 2003). The implication of the problem is increased treatment cost, prolonged illness due to treatment failure and sometimes death (Kelly et al., 2004).
The present study was conducted with the aim to detect the occurrence of VFs in P. multocida isolated from pneumonic and apparently health lungs of slaughter cattle in Iran. It was also to determine the occurrence of antimicrobial resistance among the isolates.
Materials and Methods
Sample Collection
A total of 333 samples, from both pneumonic (219) and apparently healthy (114) lungs, were collected randomly from slaughter cattle in an industrial abattoir in Shahrekord province during the period of September 2013 to March 2014. The abattoir receives cattle from different herds within and outside the province. For the purpose of this study pneumonic lungs referred to those lungs with gross lesions such as consolidation, fibrin deposition on the pleura, pleurisy, and/or adhesion; and apparently healthy lungs was used to describe those lungs without gross lesions. A simple random procedure was used to select pre-identified pneumonic and apparently health lungs. Random numbers were generated in Microsoft excel®. Specimens were obtained aseptically using a sterile scalpel while taking precautions to prevent surface contamination. Following collection the samples were conveyed to the microbiology laboratory in special ice-filled containers within 6 h of sampling.
P. multocida Screening
Isolation of P. multocida was done using techniques described previously by other authors (Songer and Post, 2005). Briefly, swabs were obtained from the collected samples and were plated on tryptic soy agar (Difco, Detroit, MI) containing 10 μg/ml NAD (Sigma, St. Louis, MO) and 5% bovine serum, MacConkey agar, and blood agar (5% fresh sheep blood). All plates were incubated at 37°C in air for a minimum of 48 h.
Identification of Isolates
Preliminary identification of P. multocida isolates was carried out according to standard biochemical tests as described earlier (Songer and Post, 2005). The isolates were gram-negative coccobacilli and were indole, catalase and oxidase-positive. But, citrate, Methyl red (MR), Vogaes–Proskauer (VP), and gelatin liquefaction negative. They don't grow on MacConkey agar and do not show hemolysis on blood agar. Confirmation of the isolates was done by polymerase chain reaction (PCR) assay with primers specific for the amplification of the KMT1 gene, adopting the methodology previously described by Townsend et al. (1998). All confirmed isolates of P. multocida were subsequently characterized by capsular serotyping using PCR. Primers for amplification of hyaD-hyaC and DcbF genes were used for detection of capsular type A and capsular type D, respectively (Table 1). P. multocida isolates which didn't yield bands on PCR when the two primers were used were classified as untyped. Following confirmation and characterization all isolates were freeze-dried and kept at −20°C.

Table 1. Primers used for the detection of serogroups in strains of P. multocida.
Detection of Virulence Genes
The virulence genes of P. multocida isolates were detected by PCR. They included adhesins (ptfA, fimA, hsf-1, hsf-2, pfhA, and tadD), toxin (toxA), iron acquisition (exbB, exbD, tonB, hgbA, hgbB, and Fur), sialidases (nanB and nanH), hyaluronidase (pmHAS), protectins (ompA, omph, oma87, and plpB) and superoxide dismutases (soda, sodC, and tbpA) (Table 2). The base sequences and the predicted sizes of the amplified products for the specific oligonucleotide primers used in detection of the genes in this study are shown in Table 3. The bacterial lysates used as templates for the PCR were prepared as follows. A loopful of bacteria from a fresh overnight culture on a tryptic soy agar plate was resuspended homogeneously in 200 μl of sterile water, and the mixture was boiled at 100°C for 5 min to release the DNA and centrifuged. A 4 μl volume of the supernatant was used as a template for each 25 μl PCR mixture. The amplified products were analyzed in 1% agarose gels by electrophoresis, and the results were recorded with a gel documentation system. All tests were repeated three times in parallel with the relevant positive (P. multocida strains ATCC 15742, ATCC 12945, and ATCC 12946) and negative (distilled water) controls. Discrepant results for each VF were investigated further, and samples were sequenced for gene verification.

Table 2. Tested virulence-associated genes in strains of P. multocida.
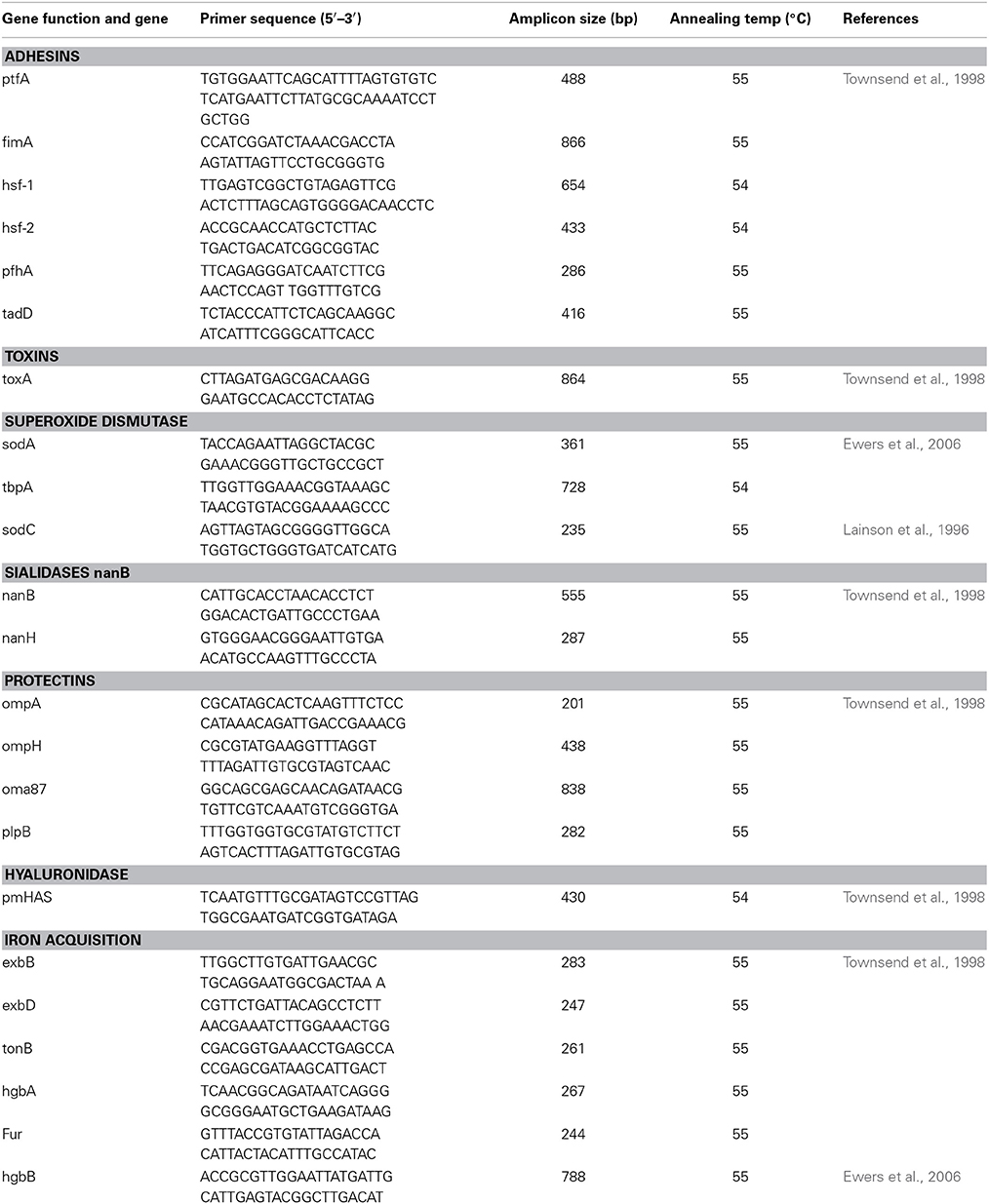
Table 3. Primers used for the detection of virulence-associated genes in strains of P. multocida.
Antimicrobial Resistance Test
Antimicrobial resistance profiles of the isolates to 20 antimicrobial agents were determined by the disc diffusion method on Muller Hinton agar with 5% blood (Carter and Subronto, 1978). The plates were inoculated with a cotton swab dipped into a 0.5 McFarland standard suspension of each isolates, according to the procedures outline in NCCLS (NCCLS, 2008). Then, the plates were incubated at 37°C for 24 h. The inhibition zones around each disc were measured and interpretation of results made according to the guidelines provided by manufacturers (Pattan-Teb, Tehran, Iran) and those provided by NCCLS (2008). The results were interpreted as resistant (R), intermediate (I), and susceptible (S).
Statistical Data Analysis
Data analysis was performed in SPSS software version 12.0 (SPSS Inc., Chicago, IL). Descriptive statistics were computed to determine the proportions of the different VFs among the isolates; and proportions of isolates resistant to different antimicrobial agents. Chi square test adopted for determination of statistical significance of differences between the proportions.
Results
Prevalence of P. multocida in Collected Samples
The prevalence of P. multocida in collected lung samples is indicated in Table 4. Overall 9.0% (30/333) of the sampled cattle were infected with the organism. The frequency of infection with the organism was higher in pneumonic lungs than in apparently health lungs and the difference was statistically significant at p ≤ 0.05.
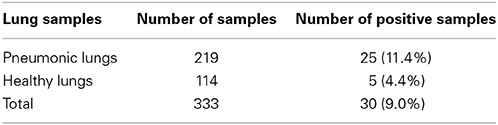
Table 4. Prevalence of P. multocida in collected cattle lung samples.
Distribution of VFs According to Capsular Serotypes
Two capsular types (A and D) were detected among 28 of the 30 isolates obtained as seen in Tables 5, 6. The majority (76.7%) of the isolates were of capsular type A. The distribution of the capsular types in pneumonic lungs and in apparently health lungs (Table 5) didn't show any statistically significant difference. The distribution of capsular serotypes for each individual isolate is displayed in Table 6.
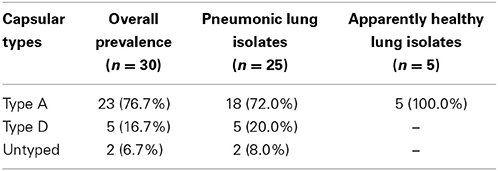
Table 5. Distribution of capsular serotypes among the isolates.
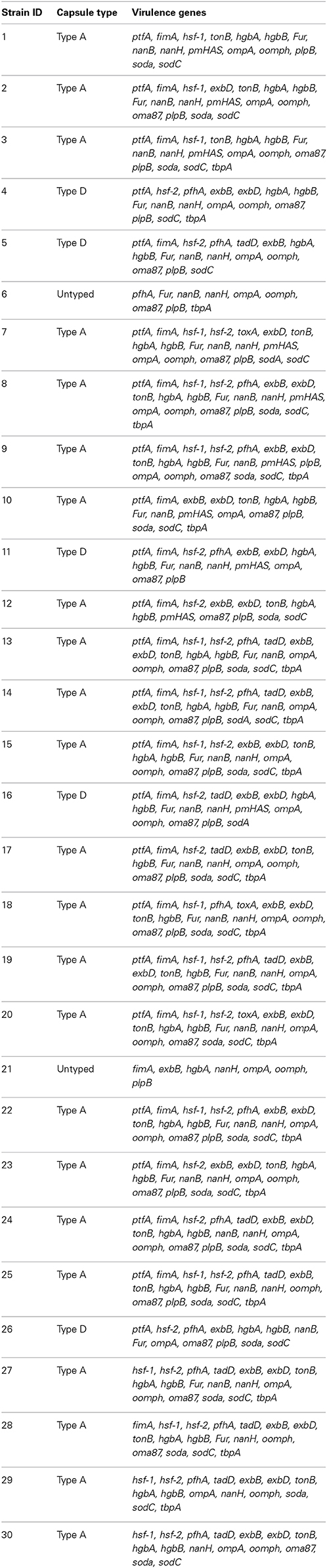
Table 6. Capsular types and virulence genes detected among P. multocida isolates obtained from cattle lungs in Iran.
Distribution of VFs According to Associated VF Genes
All isolates from pneumonic lungs harbored at least one virulence gene as displayed in Table 7. Table 8 shows the distribution of virulence genes by capsular serotypes. The detected virulence genes for each isolate obtained in this study is presented in Table 6. Most of the adhesins and superoxide dismutases; and all of the iron acquisition and protectin proteins occurred at significantly (p ≤ 0.05) higher frequencies in isolates from pneumonic lungs. One adhesion (hsf-1) and two iron acquisition (exbD and tonB) genes occurred at significantly (p ≤ 0.05) higher frequencies among capA isolates.
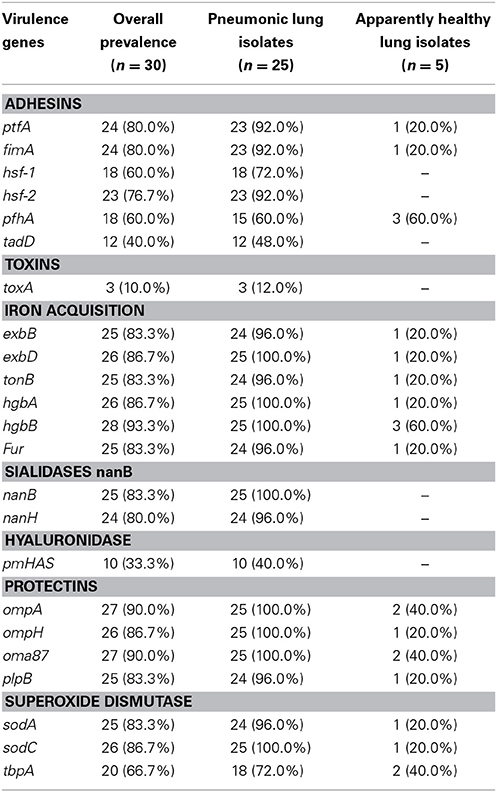
Table 7. Distribution of VFs according to associated VF genes.
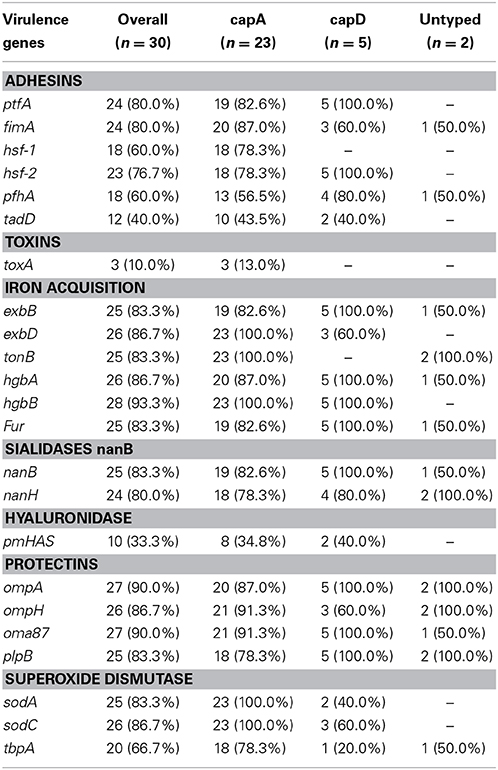
Table 8. Distribution of VFs according to capsule serotypes among 30 bovine isolates of P. multocida.
Antimicrobial Resistance Among the Isolates
Antimicrobial resistance profiles of P. multocida isolates obtained in this study are displayed in Table 9. All the isolates were susceptible to ciprofloxacin, co-trimoxazole, doxycycline, enrofloxacin, nitrofurantoin, and tetracyclines. Resistance to ampicillin, lincomycin, penicillin, rifampin, streptomycin, amoxicillin, erythromycin, and florfenicol was observed at different frequencies.
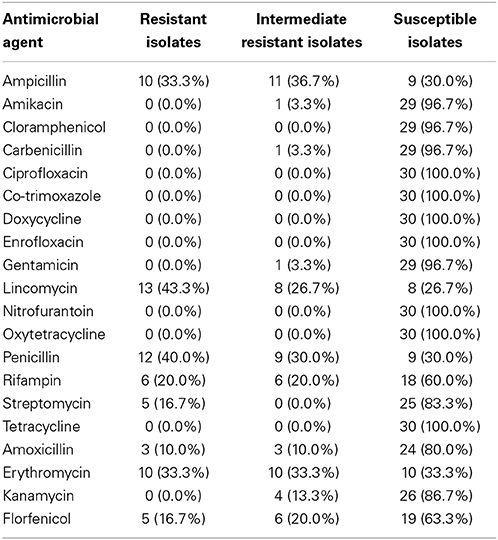
Table 9. Antimicrobial resistance profiles of P. multocida isolates against 20 antimicrobial agents.
Discussion
VFs play a key role in disease production by bacterial pathogens (Nanduri et al., 2009). Among others, their functions include competence, adherence, synthesis, and export of capsules; and evasion of host immune responses (Nanduri et al., 2009). In the present study the factors have been detected in P. multocida isolated from the lungs of slaughter cattle. The higher frequency of the factors among isolates from pneumonic lungs suggests the role of these factors in disease occurrence. It was pointed out that virulence gene occurrence in P. multocida has a strong positive association with the outcome of infection with the organism in cattle (Katsuda et al., 2013). On the other hand occurrence of the factors in apparently healthy lungs could possibly indicate early infection or contained infection which couldn't lead to disease. It was previously reported that this facultative anaerobic bacterium is commonly found in clinically healthy calves (Lainson et al., 2013).
In this study capsular types A and D were detected using PCR among the obtained P. multocida isolates. A small proportion (6.7%; 2/30) of P. multocida strains were untypeable, a similar observation to what was reported by Arumugam et al. (2011). Capsular type A was predominant among the strains accounting for 76.6%. Our observation is similar to a finding by Katsuda et al. (2013) who also detected capsular types A and D among cattle derived P. multocida isolates; with type A occurring at higher frequency. A higher frequency of capsular type A among cattle derived P. multocida isolates has also been reported in a study conducted earlier by Davies et al. (2004) who found that 99.3% of bovine P. multocida strains (n = 153) were of this capsular type. P. multocida isolates of serotype A are common in bovids occurring as normal flora in the nasopharynx; or as causes of disease including BRD and hemorraghic septicemia (Ewers et al., 2004; Dabo et al., 2007). The capsular type A is also most frequently described for rabbits (Ewers et al., 2006) and pigs (García et al., 2011).
Of the protectins; OmpA and oma87 were the most frequently detected genes particularly in the isolates from pneumonic lungs. Slightly higher frequencies of the two genes were noted for isolates of the capA serogroup than those belonging to the capD serogroup. The OmpA gene has a significant role in stabilizing the cell envelope structure by providing physical linkage between the outer membrane and peptidoglycan (Katoch et al., 2014). It mediates P. multocida host cells interaction through heparin and/or fibronectin binding and thus acts as an important invasive molecule which could determine the outcome of infection with the organism (Katoch et al., 2014).
The type 4 fimbria (ptfA gene) was described in 92.0% of the isolates tested in the current study. The gene plays a key role of fixing bacterial pathogens on the surface of the epithelial cells of hosts, a phenomenon which is more common in rabbits (Ewers et al., 2006). Consequently a study conducted on rabbits described a high prevalence of the ptfA gene (93.4%; 43/46).
Presence of adhesins on the bacterial surface is usually linked to virulence as these proteins are known to play a crucial role in facilitating host invasion and colonization (Kline et al., 2009). Studies by Ewers et al. (2000) and Tang et al. (2009) have demonstrated that, of the adhesins; fimA, hsf-2, and ptfA are of frequent occurrence among pathogenic isolates of P. multocida. In the current study the three adhesins were demonstrated at higher frequencies than others in both capA and capD serogroup isolates. On the other hand, gene tadD was the least frequently detected adhesin among P. multocida, occurring only in 48.0% of the isolates (n = 30). In these organisms the gene is known to be a putative non-specific tight adherence protein D (May et al., 2001). A more or less similar low frequency (43.3%; 100/233) of tadD was described in a field study involving pigs (Tang et al., 2009). A work on rabbits, however, observed a higher frequency (91.3%; 42/46) of this gene among P. multocida strains.
It is noteworthy that the dermonecrotoxin encoding toxA was the least frequently detected gene among the isolates; demonstrated only in those of capsular type A obtained from pneumonic lungs. Some other researchers indicated that this particular gene is more frequently expressed by strains of serogroup D and is responsible for the clinical symptoms associated with atrophic rhinitis in porcines (Harper et al., 2006; Ferreira et al., 2012). The observation in the current study could be attributed to the small sample size of capsular type D isolates. In a study conducted earlier the gene was detected in P. multocida isolates from avians, swine, shoats and cattle; but was only associated with disease in pigs (Ewers et al., 2006). Pullinger et al. (2004) points out that the toxA gene is not inserted into the bacterial chromosome but in a lysogenic bacteriophage that infects the agent.
The tbpA encoding gene is known to be of common occurrence among ruminant P. multocida strains (Ewers et al., 2006; Atashpaz et al., 2009). Its prevalence was however relatively low when compared to other superoxide dismutases (sodA and sodC) tested in this study. Ferreira et al. (2012) found a low frequency (8.6%; 4/46) of this gene in a study conducted on rabbits. Variable frequencies of the genes encoding proteins with different functions, such as adhesins (fimA, hsf-1, hsf-2, and pfhA), iron acquisition (exbB, exbD, tonB, hgbA, hgbB, and Fur), sialidases (nanB and nanH), hyaluronidase (pmHAS), and protectins (oomph and plpB) were found in the isolates. This finding is similar to what was reported in previous works which involved ruminants, porcine, poultry, and rabbits (Ewers et al., 2006; Tang et al., 2009; Ferreira et al., 2012).
Infections with P. multocida are commonly managed by broad-spectrum antimicrobials (Kehrenberg et al., 2001; Lion et al., 2006; Brogden et al., 2007). Studies have however reported occurrence of resistance to a large number of antimicrobial agents among P. multocida isolates (Hunt et al., 2001; Davies et al., 2004; Arashima and Kumasaka, 2005). In the current study all the P. multocida isolates were susceptible to ciprofloxacin, co-trimoxazole, doxycycline, enrofloxacin, nitrofurantoin, and tetracyclines. Similar observations for ciprofloxacin and co-trimoxazole (Mohamed et al., 2012); and for enrofloxacin, tetracycline, and doxycycline (Ferreira et al., 2012) have also been made earlier. These antibiotics can therefore be used for prevention and treatment of bovine P. multocida infections in the study area. Unlike other authors who reported poor (Gutiérrez Martin and Rodríguez Ferri, 1993; Yoshimura et al., 2001) and moderate (Mohamed et al., 2012) activity of aminoglycoside antibiotics against P. multocida, in the present study kanamycin, gentamicin, amikacin, and streptomycin exhibited high activity against the tested isolates. The frequencies of resistant isolates to other antibiotics varied greatly as reported by other researchers (Salmon et al., 1995; Kehrenberg et al., 2001; Yoshimura et al., 2001; Welsh et al., 2004).
The major limitation in the discussion of the findings of the current study was large differences in the sample sizes of comparison groups as seen for isolates between isolates from pneumonic lungs and those from apparently health lungs; and isolates of different capsular types. This made it difficult to infer on the observed variations as they could be attributed to chance.
In summary, our results reveal presence of VFs in P. multocida strains isolated from the lungs of symptomatic and asymptomatic slaughter cattle. Frequent detection of the factors among isolates from symptomatic study animals may suggest their role in pathogenesis of BRD caused by these organisms. Occurrence of antimicrobial resistance among some isolates is of great concern. Control strategies for this pathogen, which could include development of an effective vaccine, are warranted so as to mitigate the social and economic consequences attributable to natural infections with this bacterium. Further, the use of antimicrobial agents in modern livestock farming need to be controlled so as to minimize the emergence and eventually spread of resistance not only in target microbes but also in other important zoonotic pathogens.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to acknowledge the valuable contribution of Dr. Erick V. G. Komba of the Department of Veterinary Medicine and Public Health of Sokoine University of Agriculture, Morogoro, Tanzania; for reviewing this work before submission to the journal. Also the authors would like to thank Mr. M. Momeni at the Biotechnology Research Center of the Islamic Azad University of Shahrekord for technical support.
References
Arashima, Y., and Kumasaka, K. (2005). Pasteurellosis as zoonosis. Intern. Med. 44, 692–693. doi: 10.2169/internalmedicine.44.692
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Arumugam, N. D., Ajam, N., Blackall, P. J., Asiah, N. M., Ramlan, M., Maria, J., et al. (2011). Capsular serotyping of Pasteurella multocida from various animal hosts—a comparison of phenotypic and genotypic methods. Trop. Biomed. 28, 55–63.
Atashpaz, S., Shayegh, J., and Hejazi, M. S. (2009). Rapid virulence typing of Pasteurella multocida by multiplex PCR. Res. Vet. Sci. 87, 355–357. doi: 10.1016/j.rvsc.2009.04.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brogden, K. A., Nordholm, G., and Ackermann, M. (2007). Antimicrobial activity of cathelicidins BMAP28, SMAP28, SMAP29, and PMAP23 against Pasteurella multocida is more broad-spectrum than host species specific. Vet. Microbiol. 119, 76–81. doi: 10.1016/j.vetmic.2006.08.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Caprioli, A., Busani, L., Martel, J. L., and Helmuth, R. (2000). Monitoring of antibiotic resistance in bacteria of animal origin: epidemiological and microbiological methodologies. Int. J. Antimicrob. Agents 14, 295–301. doi: 10.1016/S0924-8579(00)00140-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Carter, G. R. (1955). Studies on Pasteurella multocida. A hemagglutination test for the identification of serological types. Am. J. Vet. Res. 16, 481–484.
Carter, G. R. (1961). A new serological type of Pasteurella multocida from Central Africa. Vet. Rec. 73, 1052.
Carter, G. R. (1967). Pasteurellosis: Pasteurella multocida and Pasteurella hemolytica. Adv. Vet. Sci. 11, 321–379.
Carter, G. R., and Subronto, P. (1978). Identification of type D strains of Pasteurella multocida with acriflavin. Am. J. Vet. Res. 34, 348–352.
Confer, A. W. (2009). Update on bacterial pathogenesis in BRD. Anim. Health Res. Rev. 10, 145–148. doi: 10.1017/S1466252309990193
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dabo, S. M., Taylor, J. D., and Confer, A. W. (2007). Pasteurella multocida and bovine respiratory disease. Anim. Health Res. Rev. 8, 129–150. doi: 10.1017/S1466252307001399
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dagleish, M. P., Finlayson, J., Bayne, C., MacDonald, S., Sales, J., and Hodgson, J. C. (2010). Characterization and time course of pulmonary lesions in calves after Intratracheal infection with Pasteurella multocida A:3. J. Comp. Pathol. 142, 157–169. doi: 10.1016/j.jcpa.2009.10.015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davies, R. L., MacCorquodale, R., and Reilly, S. (2004). Characterisation of bovine strains of Pasteurella multocida and comparison with isolates of avian, ovine and porcine origin. Vet. Microbiol. 99, 145–158. doi: 10.1016/j.vetmic.2003.11.013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Griffin, D., Chengappa, M. M., Kuszak, J., and McVey, D. S. (2010). Bacterial pathogens of the bovine respiratory disease complex. Vet. Clin. North Am. Food Anim. Pract. 26, 381–394. doi: 10.1016/j.cvfa.2010.04.004
Ewers, C., Lübke-Becker, A., Bethe, A., Kiebling, S., Filter, M., and Wieler, L. H. (2000). Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet. Microbiol. 114, 304–317. doi: 10.1016/j.vetmic.2005.12.012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ewers, C., Lübke-Becker, A., Bethe, A., Kiebling, S., Filter, M., and Wieler, L. H. (2006). Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet. Microbiol. 114, 304–317. doi: 10.1016/j.vetmic.2005.12.012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ewers, C., Lübke-Becker, A., and Wieler, L. H. (2004). Pasteurella: insights into the virulence determinants of a heterogenous bacterial type. Berliner und Münchener Tierärztliche Wochenschrift 117, 367–386.
Ferreira, T. S., Felizardo, M. R., Sena de Gobbi, D. D., Gomes, C. R., Nogueira Filsner, P. H., Moreno, M., et al. (2012). Virulence genes and antimicrobial resistance profiles of Pasteurella multocida strains isolated from rabbits in Brazil. Sci. World J. 2012:685028. doi: 10.1100/2012/685028
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
García, N., Fernández-Garayzábal, J. F., Goyache, J., Domínguez, L., and Vela, A. I. (2011). Associations between biovar and virulence factor genes in Pasteurella multocida isolates from pigs in Spain. Vet. Rec. 169, 362. doi: 10.1136/vr.d4869
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gutiérrez Martin, C. B., and Rodríguez Ferri, E. F. (1993). In vitro susceptibility of Pasteurella multocida subspecies multocida strains isolated from swine to 42 antimicrobial agents. Zentralbl. Bakteriol. 279, 387–393.
Haji Hajikolaei, M. R., Ghorbanpour, M., Seyfiabad Shapouri, M. R., Rasooli, A., Ebrahimkhani, D., and Jabbari, A. R. (2010). Bacteriological and serological studies on Mannheimia haemolytica infection in cattle slaughtered at Ahvaz (southwestern Iran) abattoir. J. Vet. Res. 11, 84–87.
Haji Hajikolaei, M. R., and Seyfi Abad Shapouri, M. R. (2007). Serological study of bovine diarrhea virus infection of cattle in Ahvaz. J. Vet. Res. 62, 21–26.
Harper, M., Boyce, J. D., and Adler, B. (2006). Pasteurella multocida pathogenesis: 125 years after Pasteur. FEMS Microbiol. Lett. 265, 1–10. doi: 10.1111/j.1574-6968.2006.00442.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hatfaludi, T., Al-Hasani, K., Boyce, J. D., and Adler, B. (2010). Outer membrane proteins of Pasteurella multocida. Vet. Microbiol. 14, 1–17. doi: 10.1016/j.vetmic.2010.01.027
Hemmatzadeh, F., Kojouri, G., Kargar Moakhar, R., and Rohany, M. (2001). A serological survey on bovine viral diarrhea virus infection in Chahar Mahal Bakhtiary province, Iran. J. Fac. Vet. Med. Univ. Tehran 56, 85–92.
Hotchkiss, E. J., Dagleish, M. P., Willoughby, K., McKendrick, I. J., Finlayson, J., Zadoks, R. N., et al. (2010). Prevalence of Pasteurella multocida and other respiratory pathogens in thenasal tract of Scottish calves. Vet. Rec. 167, 555–560. doi: 10.1136/vr.c4827
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hubbert, W. T., and Rosen, M. N. (1970). Pasteurella multocida infections. II: Pasteurella multocida infection in man unrelated to animal bite. Am. J. Public Health Nations Health 60, 1109–1117. doi: 10.2105/AJPH.60.6.1109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hunt, M. L., Adler, B., and Townsend, K. M. (2000). The molecular biology of Pasteurella multocida. Vet. Microbiol. 72, 3–25. doi: 10.1016/S0378-1135(99)00183-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hunt, M. L., Boucher, D. J., Boyce, J. D., and Adler, B. (2001). In vivo-expressed genes of Pasteurella multocida. Infect. Immun. 69, 3004–3012. doi: 10.1128/IAI.69.5.3004-3012.2001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Katoch, S., Sharma, M., Patil, R. D., Kumar, S., and Verma, S. (2014). In vitro and in vivo pathogenicity studies of Pasteurella multocida strains harbouring different ompA. Vet. Res. Commun. 38, 183–191. doi: 10.1007/s11259-014-9601-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Katsuda, K., Hoshinoo, K., Ueno, Y., Kohmoto, M., and Mikami, O. (2013). Virulence genes and antimicrobial susceptibility in Pasteurella multocida isolates from calves. Vet. Microbiol. 167, 737–741. doi: 10.1016/j.vetmic.2013.09.029
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kehrenberg, C., Schulze-Tanzil, G., Martel, J. L., Chaslus-Dancla, E., and Schwarz, S. (2001). Antimicrobial resistance in Pasteurella and Mannheimia: epidemiology and genetic basis. Vet. Res. 32, 323–339. doi: 10.1051/vetres:2001128
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kelly, L., Smith, D. L., Snary, E. L., Johnson, J. A., Harris, A. D., Wooldrige, M., et al. (2004). Animal growth promoters: to ban or not to ban? A risk assessment approach. Int. J. Antimicrob. Agents 24, 205–212. doi: 10.1016/j.ijantimicag.2004.04.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kline, K. A., Fälker, S., Dahlberg, S., Normark, S., and Henriques-Normark, B. (2009). Bacterial adhesins in host-microbe interaction. Cell Host Microbe 5, 580–592. doi: 10.1016/j.chom.2009.05.011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lainson, F. A., Dagleish, M. P., Fontaine, M. C., Bayne, C., and Hodgson, J. C. (2013). Draft genome sequence of Pasteurella multocida A:3 Strain 671/90. Genome Announc. 1, e00803–e00813. doi: 10.1128/genomeA.00803-13
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lainson, F. A., Thomson, N., Rowe, H. A., Langford, P. R., Aitchison, K. D., Donachie, W., et al. (1996). Occurrence of [copper, zinc]-cofactored superoxide dismutase in Pasteurella haemolytica and its serotype distribution. FEMS Microbiol. Lett. 142, 11–17. doi: 10.1111/j.1574-6968.1996.tb08400.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Levy, S. B. (1998). The challenge of antibiotic resistance. Sci. Am. 278, 46–53. doi: 10.1038/scientificamerican0398-46
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lion, C., Conroy, M. C., Carpentier, A. M., and Lozniewski, A. (2006). Antimicrobial susceptibilities of Pasteurella strains isolated from humans. Int. J. Antimicrob. Agents 27, 290–293. doi: 10.1016/j.ijantimicag.2006.02.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mannheim, W. (1984). “Family III. Pasteurellaceae,” in Bergey's Manual of Systematic Bacteriology, Vol. 1, eds N. R. Krieg, and J. G. Kolt (Baltimore, MD: Williams & Wilkins), 550–575.
May, B. J., Zhang, Q., Li, L. L., Paustian, M. L., Whittam, T. S., and Kapur, V. (2001). Complete genomic sequence of Pasteurella multocida, Pm70. Proc. Natl. Acad. Sci. U.S.A. 98, 3460–3465. doi: 10.1073/pnas.051634598
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mohamed, M. A., Mohamed, M. W., Ahmed, A. I., Ibrahim, A. A., and Ahmed, M. S. (2012). Pasteurella multocida in backyard chickens in Upper Egypt: incidence with polymerase chain reaction analysis for capsule type, virulence in chicken embryos and antimicrobial resistance. Vet. Ital. 48, 77–86.
Nanduri, B., Shack, L. A., Burgess, S. C., and Lawrence, M. L. (2009). The transcriptional response of Pasteurella multocida to three classes of antibiotics. BMC Genomics 10:S4 doi: 10.1186/1471-2164-10-S2-S4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
National Committee for Clinical Laboratory Standards. (2008). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standard, 2nd Edn. NCCLS Document M31-A3. Wayne, PA: National Committee for Clinical Laboratory Standards.
Portis, E., Lindeman, C., Johansen, L., and Stoltman, G. (2012). A ten-year (2000–2009) study of antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex—Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni—in the United States and Canada. J. Vet. Diagn. Invest. 24, 932–944 doi: 10.1177/1040638712457559
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pullinger, G. D., Bevir, T., and Lax, A. J. (2004). The Pasteurella multocida toxin is encoded within a lysogenic bacteriophage. Mol. Microbiol. 51, 255–269. doi: 10.1046/j.1365-2958.2003.03829.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rimler, R. B., and Rhoades, K. R. (1987). Serogroup F, a new capsule serogroup of Pasteurella multocida. J. Clin. Microbiol. 25, 615–618.
Sakhaee, E., Khalili, M., and Kazeminia, S. (2009). Serological study of bovine viral respiratory diseases in dairy herds in Kerman province, Iran. Iran. J. Vet. Res. 10, 49–53.
Salmon, S. A., Watts, J. L., Case, C. A., Hoffman, L. J., Wegener, H. C., and Yancey, R. J. Jr. (1995). Comparison of MICs of ceftiofur and other antimicrobial agents against bacterial pathogens of swine from the United States, Canada, and Denmark. J. Clin. Microbiol. 33, 2435–2444.
Shea, K. M. (2003). Antibiotic resistance. What is the impact of agricultural uses of antibiotics on children health? Pediatrics 112, 253–258.
Songer, J. G., and Post, K. W. (2005). Veterinary Microbiology, Bacterial and Fungal Agents of Animal Diseases. 1st Edn. Saint Louis, MO: Elsevier Saunders.
Steen, J. A., Steen, J. A., Harrison, P., Seemann, T., Wilkie, I., Harper, M., et al. (2010). Fis is essential for capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLOS Pathog. 6:e1000750. doi: 10.1371/journal.ppat.1000750
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tang, X., Zhao, Z., Hu, J., Wu, B., Cai, X., He, Q., et al. (2009). Isolation, antimicrobialm resistance, and virulence genes of Pasteurella multocida strains from swine in China. J. Clin. Microbiol. 47, 951–958. doi: 10.1128/JCM.02029-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Townsend, K. M., Frost, A. J., Lee, C. W., Papadimitriou, J. M., and Dawkins, H. J. S. (1998). Development of PCR assays for species and type-specific identification of Pasteurella multocida isolates. J. Clin. Microbiol. 36, 1096–1100.
Verma, S., Sharma, M., Katoch, S., Verma, L., Kumar, S., Dogra, V., et al. (2013). Profiling of virulence associated genes of Pasteurella multocida isolated from cattle. Vet. Res. Commun. 37, 83–89. doi: 10.1007/s11259-012-9539-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Welsh, R. D., Dye, L. B., Payton, M. E., and Confer, A. W. (2004). Isolation and antimicrobial susceptibilities of bacterial pathogens from bovine pneumonia: 1994–2002. J. Vet. Diagn. Invest. 16, 426–431. doi: 10.1177/104063870401600510
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
White, D. G., Zhao, S., Simjee, S., Wagner, D. D., and McDermott, P. F. (2002). Antimicrobial resistance of foodborne pathogens. Microbes Infect. 4, 405–412. doi: 10.1016/S1286-4579(02)01554-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Witte, W. (1998). Medical consequences of antibiotic use in agriculture. Science 279, 996–997. doi: 10.1126/science.279.5353.996
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yoshimura, H., Ishimaru, M., Endoh, Y. S., and Kojima, A. (2001). Antimicrobial susceptibility of Pasteurella multocida isolated from cattle and pigs. J. Vet. Med. B Infect. Dis. Vet. Public Health 48, 555–560. doi: 10.1046/j.1439-0450.2001.00468.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: Pasteurella multocida, virulence factors, antimicrobial resistance, cattle, Iran
Citation: Khamesipour F, Momtaz H and Azhdary Mamoreh M (2014) Occurrence of virulence factors and antimicrobial resistance in Pasteurella multocida strains isolated from slaughter cattle in Iran. Front. Microbiol. 5:536. doi: 10.3389/fmicb.2014.00536
Received: 18 June 2014; Accepted: 26 September 2014;
Published online: 14 October 2014.
Edited by:
Christina Maria Joseph Elisabeth Vandenbroucke-Grauls, VU University Medical Center, NetherlandsReviewed by:
Amanda L. Lewis, Washington University School of Medicine, USAAlasdair Cook, University of Surrey School of Veterinary Medicine, UK
Copyright © 2014 Khamesipour, Momtaz and Azhdary Mamoreh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hassan Momtaz, Department of Microbiology, Faculty of Veterinary Medicine, Shahrekord Branch, Islamic Azad University, PO Box 166, Shahrekord, Iran e-mail: hamomtaz@yahoo.com; hamomtaz@iaushk.ac.ir