 Vito Valiante
Vito Valiante Juliane Macheleidt
Juliane Macheleidt Martin Föge
Martin Föge Axel A. Brakhage
Axel A. Brakhage- 1 Molecular Biotechnology of Natural Products, Department of Molecular and Applied Microbiology, Leibniz Institute for Natural Product Research and Infection Biology – Hans Knöll Institute, Jena, Germany
- 2 Department of Microbiology and Molecular Biology, Institute of Microbiology, Friedrich Schiller University, Jena, Germany
Aspergillus fumigatus is the most important airborne fungal pathogen, causing severe infections with invasive growth in immunocompromised patients. The fungal cell wall (CW) prevents the cell from lysing and protects the fungus against environmental stress conditions. Because it is absent in humans and because of its essentiality, the fungal CW is a promising target for antifungal drugs. Nowadays, compounds acting on the CW, i.e., echinocandin derivatives, are used to treat A. fumigatus infections. However, studies demonstrating the clinical effectiveness of echinocandins in comparison with antifungals currently recommended for first-line treatment of invasive aspergillosis are still lacking. Therefore, it is important to elucidate CW biosynthesis pathways and their signal transduction cascades, which potentially compensate the inhibition caused by CW- perturbing compounds. Like in other fungi, the central core of the cell wall integrity (CWI) signaling pathway in A. fumigatus is composed of three mitogen activated protein kinases. Deletion of these genes resulted in severely enhanced sensitivity of the mutants against CW-disturbing compounds and in drastic alterations of the fungal morphology. Additionally, several cross-talk interactions between the CWI pathways and other signaling pathways are emerging, raising the question about their role in the CW compensatory mechanisms. In this review we focused on recent advances in understanding the CWI signaling pathway in A. fumigatus and its role during drug stress response and virulence.
Overview About Cell Wall Organization in Aspergillus fumigatus
The fungal cell wall (CW) is the exoskeleton of fungal cells. Besides its starkness and firmness, it has a very flexible structure, which, therefore, can alter rapidly and efficiently in response to external and internal stimuli. In particular, filamentous fungi adapt their internal pressure and turgor in concert with CW-biosynthesis enzymes, in order to direct hyphal growth, following gradients of nutrients, and chemo attractants (e.g., hormones), or to avoid adverse habitats (Brand and Gow, 2009; Lew, 2011).
In recent years, the composition of the fungal CW was extensively studied. It varies among different species but is mostly composed of polymers of sugars, which show a high degree of branching. Although the CW composition varies among different fungal species, there are conserved parts such as a common core composed of branched β-1,3-glucan-chitin. An exception was found in zygomycetes, which contain chitosan instead of chitin (Singh et al., 2011). The polysaccharide-based three-dimensional network in Aspergillus fumigatus is completed by the addition of sugar-chains composed of α-1,3-glucan, galactofuran, and mannan, which make the structure rather variable compared to other fungi (Latge and Beauvais, 2014). Besides the polysaccharide structure, the CW is adorned with a variety of proteins. In A. fumigatus dormant conidia, the cell surface is covered by a rodlet layer, which is composed of regularly arranged RodA hydrophobin proteins. This hydrophobin envelope was found to be essential to immunologically mask the conidia, which are continuously inhaled by humans (Aimanianda et al., 2009). Other hydrophobins, which are produced at different developmental stages in A. fumigatus, were associated with drug response (Gautam et al., 2008) and biofilm-growth conditions (Bruns et al., 2010).
In the CW biosynthesis, the β-1,3-glucan synthase (Fks1) plays the major role (Beauvais et al., 2001). Fks1 is an integral plasma membrane protein having 16 putative trans-membrane helices. The genome of the majority of fungi normally contains only a single β-1,3-glucan synthase gene. Consequently, the deletion of this gene was supposed to be lethal (Firon et al., 2002; Henry et al., 2012). However, a recent publication reported the characterisation of an A. fumigatus Δfks1 mutant, which appeared to be viable besides showing a severe growth phenotype (Dichtl et al., 2015).
The A. fumigatus genome harbors many putative genes responsible for CW modifications. Different β-glucanases and branching enzymes were identified (Mouyna et al., 2013). These genes are more difficult to analyze by classical reverse genetics, because some of them are apparently functionally redundant. An example is given by the β-1,3-glucanosyltransferase (Gel) family, which, in A. fumigatus, is composed of seven different members. Among them, only the deletion of gel4 was reported to be lethal (Gastebois et al., 2010), whereas the Δgel2, Δgel1, and Δgel7 deletion mutant strains were viable (Mouyna et al., 2005; Zhao et al., 2014).
For chitin, which is the second most abundant polysaccharide of the CW, a very similar situation was found. Among the eight putative chitin synthase genes identified in A. fumigatus, only the deletion of csmA and csmB led to a significantly altered phenotype, i.e., reduction of the colony radial growth rate and decrease in chitin content (Aufauvre-Brown et al., 1997; Jimenez-Ortigosa et al., 2012), while the mutation of the remaining genes did not result in a significant phenotype, and obvious phenotypical changes of mutants were only reported upon multiple simultaneous gene disruptions (Rogg et al., 2011; Muszkieta et al., 2014).
The knowledge about genes involved in the biosynthesis of the CW is steadily increasing. In total, more than 30% of the A. fumigatus genome still encodes genes with unknown function (Cerqueira et al., 2014). Thus, it is reasonable to assume that in the future further enzymes involved in CW biosynthesis will be discovered, likely having novel enzymatic functions. As an example, it was predicted that A. fumigatus potentially expresses more than 100 different glycosylphosphatidylinositol (GPI) anchored proteins (Cao et al., 2009), which are likely to form a bridge between the membrane lipid bilayer and the CW. The majority of these proteins are supposed to play a role in the CW formation, but only very few have been characterized so far (Li et al., 2007).
Activity of Antifungal Drugs Targeting the Fungal Cell Wall and Resistance Mechanisms
From all the different classes of potential antifungal drugs, only three of them have a wide clinical use against invasive and systemic infections caused by A. fumigatus. These compounds belong to the polyenes, azoles, and echinocandins (Ullmann and Cornely, 2006).
Polyenes and azoles are mainly targeting ergosterol and its biosynthesis, respectively, while echinocandins specifically inhibit β-1,3-glucan formation (Figure 1). However, it is increasingly acknowledged that defense mechanisms against these drugs have common elements. Consistently, a genome-wide screen conducted in Schizosaccharomyces pombe indicated that different ergosterol biosynthesis deficient mutants were also sensitive to β-1,3-glucan synthase inhibitor echinocandins and β-glucanase (Fang et al., 2012). A similar observation was made for Candida albicans biofilms, which normally exhibit higher resistance to polyenes depending on ergosterol and β-1,6-glucan synthesis (Khot et al., 2006).
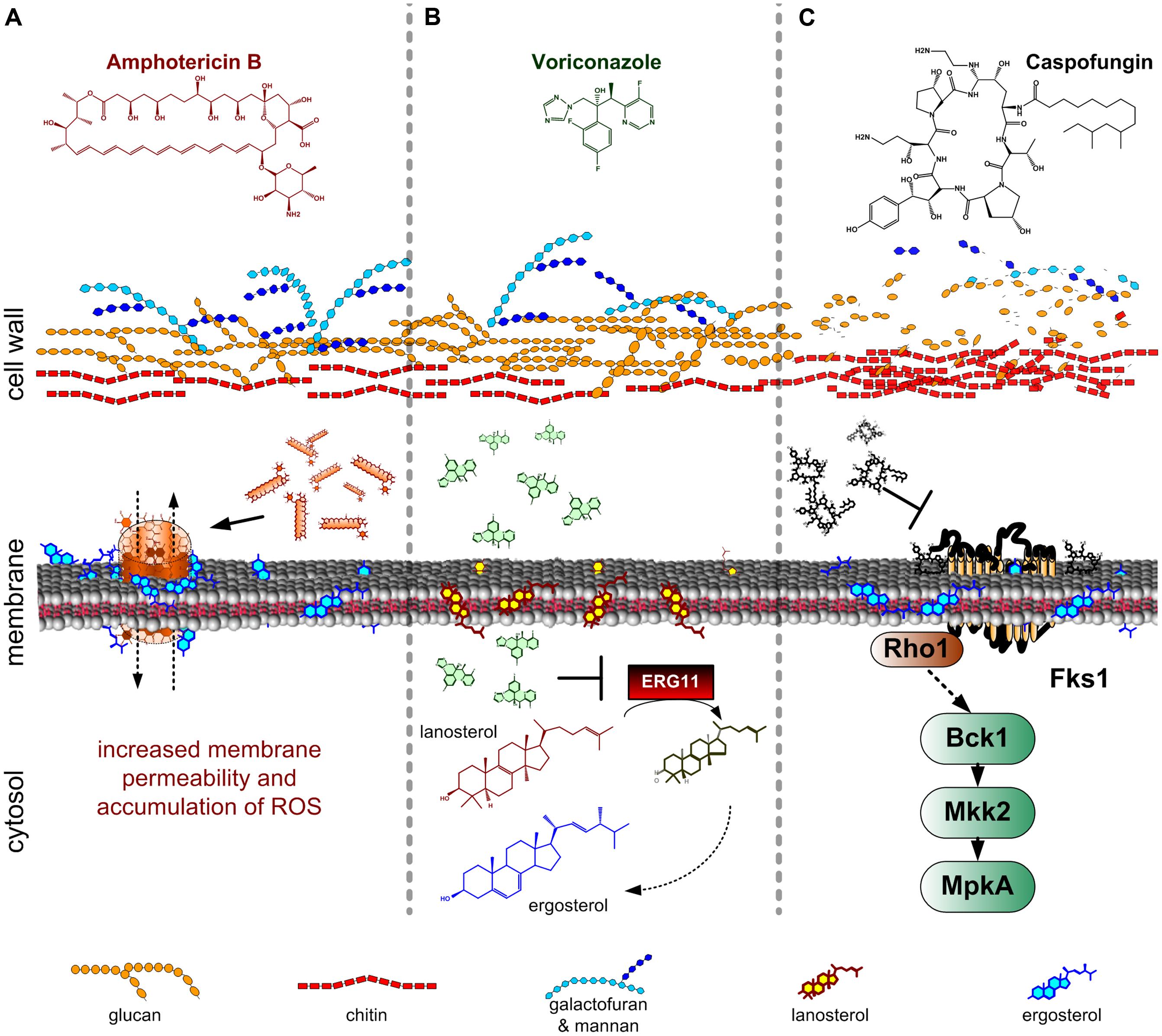
FIGURE 1. Mode of actions of antifungal drugs commonly used against invasive and systemic Aspergillus fumigatus infection (Ullmann and Cornely, 2006). (A) Polyenes (in this case amphotericin B) bind to sterols, forming pores in the cell membrane. The formation of these pores leads to increased membrane permeability. Additionally, amphotericin B induces the accumulation of reactive oxygen species (ROS), which have multiple toxic effects on fungal cells; (B) Azoles (in this case voriconazole) inhibit the ERG11 enzyme thereby blocking the ergosterol biosynthesis. ERG11 catalyses the formation of 4,4-dimethylcholesta-8,14,24-trienol from lanosterol. The lack of ergosterol and the subsequent accumulation of lanosterol, results in high toxicity for the cell; (C) Echinocandins (in this case caspofungin) inhibit the β-1,3-glucan synthesis. As a response to the reduction of glucan content there is an increase in chitin biosynthesis.
Polyenes belong to the oldest known antifungal compounds. Amphothericin B (AmB), first isolated from Streptomyces nodosus, is the most commonly used polyene against systemic infections (Blum et al., 2013). AmB possesses a name-giving amphoteric character, which allows the binding of the lipophilic compound to the cytoplasmic membrane. The mode of action of polyenes is not fully understood. They theoretically bind to all kinds of sterols, with a significant higher affinity to ergosterol (Bolard, 1986; Figure 1A). The fungicidal activity of AmB is potentially based on the formation of channels in the cell membrane, which lead to increased membrane permeability and leakage of small molecules, inhibition of aerobic and anaerobic respiration, and accumulation of reactive oxygen species (ROS) (Mesa-Arango et al., 2014; Figure 1A). It was suggested that eight AmB molecules are assembled to a ring-like structure, showing both a hydrophobic interaction to ergosterol with its conjugated double-bond system and forming a hydrophilic pore with its inward directed hydroxyl groups (Baginski et al., 1997). Thus, the sensitivity toward polyenes is dependent on the composition of membranes, with regard to sterol content, and chemical structure of phospholipid fatty acyl chains.
Surprisingly, despite the long-term use of polyenes, resistance to AmB and others polyene drugs remains rare. Nevertheless, there are resistant species of Candida (C. glabrata, C. tropicalis, C. lusitaniae), which are characterized by an intrinsically low ergosterol content (Young et al., 2003; Vandeputte et al., 2008; Gray et al., 2012; Eddouzi et al., 2013). The resistance to AmB in Aspergillus terreus was attributed to a higher level of catalase expression that counteracts the AmB-induced formation of ROS (Blum et al., 2013). Additionally, resistant environmental Aspergillus flavus strains were isolated, which were characterized by an alteration of CW composition and higher levels of α-1,3-glucan (Seo et al., 1999).
The group of azoles comprise a plethora of synthetic compounds, characterized by the presence of either a 2-N-containing imidazole ring or a 3-N-containing triazole ring with complex side-chains of at least one halogenated phenyl group (Bruggemann et al., 2009). In general, azoles show a broad spectrum of antifungal activity, a good toleration by humans and a relative low cytotoxicity, making them the major agents for treatment, and prevention of Aspergillus infections. There are many licensed azole antifungal drugs, but only four, namely fluconazole, itraconazole, posaconazole, and voriconazole, are mainly used for treatment of fungal infections (Bruggemann et al., 2009). They all act by inhibiting the fungal cytochrome P450 14α-sterol demethylase, which leads to a decrease in ergosterol in the fungal cytoplasmic membrane and thus to inhibition of growth (Figure 1B). In A. fumigatus, two genes have been assigned to this enzymatic function: cyp51A and cyp51B (erg11) (Mellado et al., 2001). The deduced proteins catalyze the oxidative removal of the 14α-methyl group of lanosterol or eburicol, respectively, (Figure 1B). However, single deletions of genes involved in either demethylation or desaturation of ergosterol intermediates such as cyp51A, cyp51B, and erg3A, erg3B and erg3C, respectively, showed differences in relative sterol composition, but further tests indicated that neither of these genes is individually essential for A. fumigatus survival and virulence (Mellado et al., 2005; Hu et al., 2007; Alcazar-Fuoli and Mellado, 2012).
The main mechanism of resistance of A. fumigatus against azole antifungal drugs is based on a mutation of the cyp51A gene locus with a conversion of leucine at position 98 into histidine (L98H) in combination with the appearance of a 34-bp tandem repeat in cyp51A promoter region (TR34). In addition to TR34/L98H, also other hot spots for point mutations in the cyp51A gene have been reported, e.g., the exchange of glycine at pos. 54, with different impact on azole resistance in clinical isolates (Lelievre et al., 2013). Recently, a novel azole resistance mechanism was reported, which was caused by a mutation in the CCAAT-binding transcription factor (TF) complex subunit HapE (Camps et al., 2012). HapE interacts with the Hap-complex, which is, besides other functions, also important for adaptation to iron starvation and iron excess stress (Hortschansky et al., 2007).
The third and most recent class of clinically used anti-mycotic drugs are echinocandins. These molecules are composed of a cyclic hexapeptide core linked to a lipid side chain. Echinocandins act as non-competitive inhibitors of the β-1,3-glucan synthase (Figure 1C; Perez et al., 1983; Sawistowska-Schroder et al., 1984). Caspofungin was the first clinically applied echinocandin (CANCIDAS®, caspofungin acetate), which specifically targets the fungal CW assembly (Bowman et al., 2002). Inhibition of β-1,3-glucan synthesis results in inhibition of growth, increased osmotic sensitivity, and can even lead to lysis of cells. Besides their specificity, the use of echinocandins in clinical therapy is influenced by two drawbacks: the emergence of resistant strains, and the occurrence of the so-called paradoxical effect, which makes the drug less effective when used at high concentrations (Rocha et al., 2007; Wiederhold, 2009).
Until today, only little is known about naturally occurring resistance mechanisms against echinocandins. The most common mechanism is over-expression of the fks1 gene in clinical isolates of A. fumigatus (Arendrup et al., 2009). Additionally, mutation of the Fks1 protein by substitution of a serine in position 678 by a proline (S678P) makes A. fumigatus resistant to caspofungin (Rocha et al., 2007). Apart from these changes, the adaptation of A. fumigatus to caspofungin is accompanied by a change in the CW sugar composition. As reported for other fungi, the exposition of A. fumigatus to caspofungin led as well to a decrease in β-glucan content and to an increase of chitin (Cowen and Steinbach, 2008; Verwer et al., 2012). In particular, these mechanisms seem to be related to the paradoxical effect exerted by this drug (Fortwendel et al., 2010). Potential mechanisms that have been suggested to induce chitin synthesis include manipulation of signaling pathways, such as the up-regulation of protein kinase C-encoding gene pkcA, a key component of the cell wall integrity (CWI) pathway, and elements acting in the calcineurin pathway (Wiederhold, 2007; Fortwendel et al., 2010).
The MAPK Cell Wall Integrity Signaling Pathway in A. fumigatus
Several of the major findings about signaling pathways were firstly reported in the model fungus Saccharomyces cerevisiae (Levin, 2011). The advances in genome sequencing allowed the identification of similar or identical signaling pathways in different fungal species, revealing the presence of highly conserved proteins that can be potentially used as antifungal drug targets (Horn et al., 2012). In particular, general signaling cascades such as the mitogen activated protein kinases (MAPKs), calcineurin, cAMP, and target of rapamycin (Tor) pathways are highly conserved in the fungal kingdom (Grosse et al., 2008; Rispail et al., 2009; Baldin et al., 2015).
Among the different signaling pathways, the MAPK CWI signaling plays a major role in CW maintenance. The CWI signal pathway in A. fumigatus is composed of a highly conserved module formed by three MAPKs, namely Bck1 (MAPK kinase kinase), Mkk2 (MAPK kinase), and MpkA (MAPK), which sequentially phosphorylate each other (Figure 2; Valiante et al., 2009; Dirr et al., 2010). Upon phosphorylation, MpkA moves into the nucleus, where it likely activates transcriptional regulators (Jain et al., 2011). The deletion of one of the three kinases of the MpkA module led to the lack of MpkA phosphorylation, which is the bottleneck in the activation of the entire pathway (Valiante et al., 2008). The three mutants appear phenotypically identical (Figure 2).
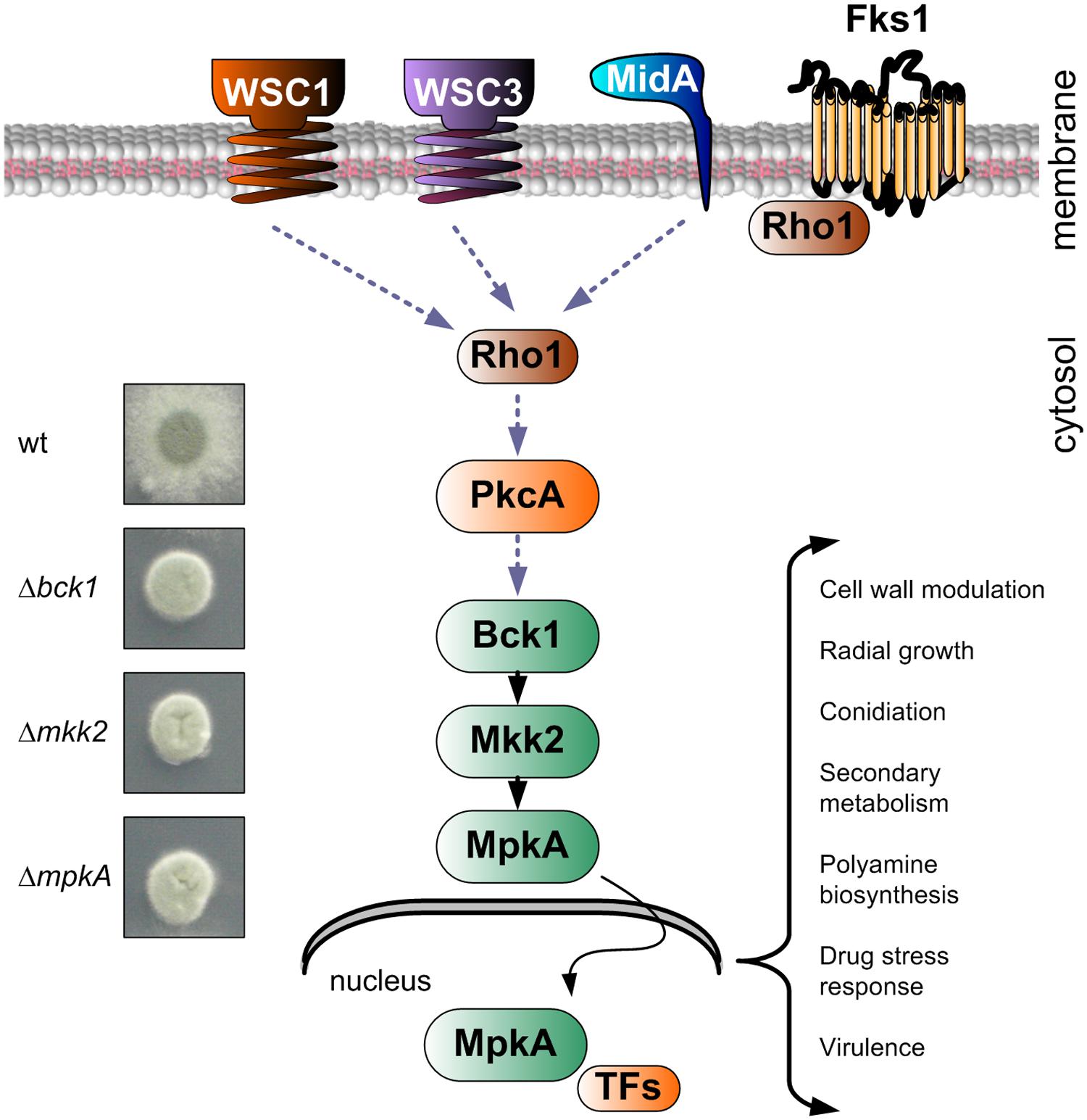
FIGURE 2. Schematic representation of the cell wall integrity (CWI) signaling pathway in A. fumigatus and colony growth of indicated deletion mutants on agar plates in comparison to the wild type. The picture shows those elements that were experimentally proven to be part of the A. fumigatus CWI pathway, plus some important elements, whose action was predicted based on data reported in other organisms (Fks1, Rho1, and PkcA). Once phosphorylated, MpkA translocates in the nucleus, where specific transcription factors (TFs) are activated. The deletion of genes constituting the central MAPK module have identical phenotypes, resulting in a compact colony and sensitivity to all cell wall (CW)-acting compounds (Valiante et al., 2008, 2009; Jain et al., 2011; Dichtl et al., 2012).
The lack of phosphorylation of MpkA yields mutants with a typically impaired CW, showing compact colonies and reduced filamentation (Valiante et al., 2009). Additionally, mutants of all three MAPKs are equally sensitive to all known CW-disturbing compounds, which highlight their incompetence to recover from CW stress (Valiante et al., 2008, 2009).
RNA-seq analysis performed on the ΔmpkA mutant of A. fumigatus showed that the deletion of this important kinase globally changed the expression of genes putatively involved in primary metabolism. Many genes involved in sugar and amino acid metabolism were differentially regulated, whereas many well-known CW biosynthesis genes were not affected (Müller et al., 2012). This unexpected finding suggested that the CWI pathway regulates CW biosynthesis by directly affecting the synthesis of sugars, which are the essential bricks for the CW formation. As a consequence, the role of the CWI pathway seems to be not only the mere structuring of the CW, but it is likely involved in fine-tuning the balance between stress responses and energy consumption in cellular processes required for growth and development. The latter statement is strengthened by the involvement of the CWI pathway in stress responses that are not directly connected to the CW biosynthesis. As an example, MpkA was activated during iron depletion (Jain et al., 2011). When A. fumigatus grows under iron starvation conditions, the fungus produces secondary metabolites named siderophores, which are derived from ornithine metabolism. Siderophores are secreted to bind external iron from the environment. The production of such metabolites is rather energy-consuming (Haas, 2014). The lack of mpkA in A. fumigatus increased the global siderophore production during iron depletion by positively regulating the metabolic flux toward the ornithine metabolic pathway. Living cells use ornithine and its precursor arginine for polyamine production. The functions of these small aliphatic molecules like putrescine and spermidine are still a matter of debate, but it was shown that their concentration accompanies certain developmental transitions or exposure to stress conditions (Minguet et al., 2008). In fact, the synthesis of putrescine and spermidine has been proven to be essential for life (Chattopadhyay et al., 2002). As a result, the increased production of siderophores occurring during iron starvation conditions negatively influenced the global polyamine production and storage (Jain et al., 2011). These results suggested that during iron starvation, a functional MpkA is required to act as a central regulator to balance essential polyamine formation and de novo siderophore production. Additionally, the latter mechanism seems to be independent from the well-known HapX-SreA iron regulation system in A. fumigatus (Hortschansky et al., 2007; Gsaller et al., 2014), suggesting that the role of this kinase is epistatic to HapX-SreA in the stress response process.
Besides siderophore production, the CWI pathway seems to be an important regulator of secondary metabolism in A. fumigatus. Transcriptome data revealed that more than 50% of the identified secondary metabolite gene clusters were affected by the lack of mpkA (Müller et al., 2012). This also included the production of important virulence determinants such as gliotoxin and melanins (Jain et al., 2011). Melanins are a class of brown-pigmented molecules, which are often associated with the CW. Their main role is to protect the organisms from exogenous stresses, contributing to the first line of defense against external hazards (Heinekamp et al., 2012). A. fumigatus produces two kinds of melanins: the dihydroxynaphthalene (DHN) melanin, which is responsible for the characteristic gray-greenish color of the conidia (Langfelder et al., 2003), and pyomelanin, which is produced during tyrosine catabolism (Schmaler-Ripcke et al., 2009). The expression of the DHN-melanin biosynthesis gene cluster was found to be decreased in the ΔmpkA mutant strain. However, these mutants were still able to produce DHN-melanin, suggesting that the role of the CWI pathway in the regulation of the cluster is significant but not decisive. Concerning pyomelanin, a higher production of this compound was found in the ΔmpkA mutant strain (Valiante et al., 2009). These data confirmed that pyomelanin exerts a protective role, potentially compensating CW stress effects in a dysfunctional CWI background.
As summarized here, the A. fumigatus CWI pathway is required for many physiological states, and the CW maintenance is not its only function. It is still a matter of debate whether the CWI signaling is actively involved in the regulation of such a variety of processes or whether the observed phenotypes of corresponding deletion mutant strains are a consequence of the strong physiological impairment derived by the lack of a functional phosphorylation cascade. The meaning of the CWI signaling in A. fumigatus for virulence is unclear. The lack of mpkA did not affect virulence in a cyclophosphamide-cortisone-acetate murine infection model (Valiante et al., 2008). By contrast, the Δmkk2 mutant strain showing the same phenotypes as the ΔmpkA mutant in vitro, displayed reduced virulence in a murine systemic infection model (Dirr et al., 2010). These results suggest that in spite of the strongly reduced growth, the presence of neutrophil granulocytes is needed to clear the infection caused by these mutants.
Comparative analysis of signaling cascades between different fungi highlighted that the central core of signaling, i.e., kinases, are conserved, while upstream (receptors), and downstream (TFs) elements are more diverse (Rispail et al., 2009). In particular, receptor families are conserved, but vary in number. This suggests that activation of signaling is species-specific.
One of the most studied class of receptors acting upstream to the CWI signaling are the so-called WSC receptors (CWI and stress response components) (Nanduri and Tartakoff, 2001), which are characterized by the presence of highly repetitive domains containing serine and threonine residues. The WSC receptors also contain a putative carbohydrate-binding domain, which suggests that they could function as a bridge between the CW itself and the cytoplasmic membrane. Studies on the S. cerevisiae Wsc1 receptor suggested that these receptors work as a nanospring, able to detect/sense pulses derived from mechanical stress (Dupres et al., 2009). BLAST analysis revealed that the genome of A. fumigatus contains four putative genes coding for WSC receptors (wsc1, wsc2, wsc3, and mid2; Dichtl et al., 2012). All these proteins were found in the cytoplasmic membrane. However, their activity in A. fumigatus appears to be partially redundant, and a clearer CW phenotype was observed when multiple genes were simultaneously deleted. Additionally, the function of wsc2 could not be related to the CWI signaling.
Other receptors putatively acting upstream of MAPK signaling pathways are the G protein coupled receptors (GPCRs). The number of these receptors also varies between different species (Grice et al., 2013). Deletion of the gprC and gprD genes in A. fumigatus produced mutant strains impaired in CW-related stress response (Gehrke et al., 2010).
In S. cerevisiae, receptors activating the MAPK signaling cascade are connected to the MAPK module via different signaling elements. BLAST analysis revealed that those elements are also conserved in the A. fumigatus genome (Rispail et al., 2009). Two of them, Rho1 and PkcA, are supposed to be essentials (Dichtl et al., 2012; De Souza et al., 2013); thus, their action should not only be related to the CWI signaling (Figure 2; Dichtl et al., 2010). Rho1 belongs to the Ras homolog family, which is a group of important signaling proteins (Levin, 2011). In A. fumigatus, Rho1 is localized in the hyphal tip, and it is misplaced by addition of farnesol (Dichtl et al., 2010), which suggests that this Ras protein interacts with putative membrane receptors to transduce external stimuli and to activate signaling cascades. A more evident phenotype was reported for the Rho guanyl-nucleotide exchange factor Rom2, which clearly localizes to the hyphal tips and septa, and its reduced expression strongly effects CW shape and stability (Samantaray et al., 2013).
Concerning PkcA (protein kinase C), there are no studies describing its function in A. fumigatus, but the repression of this gene in the closely related fungus Aspergillus nidulans affected the production of penicillin, which supports the involvement of the CWI pathway in the regulation of secondary metabolism (Herrmann et al., 2006). However, although these results suggested that the role of Rho1 and PkcA is conserved in fungi, the link between membrane receptors and the activation of the CWI pathway was shown in model yeasts, but not experimentally confirmed in Aspergillus species until now (Levin, 2011).
As mentioned before, the MAPK module is supposed to modulate the activity of transcriptional factors, associated with CWI signaling. Transcriptional regulators that act downstream of the CWI signaling pathway in S. cerevisiae were identified by BLAST analysis also in the A. fumigatus genome (Rispail et al., 2009; Levin, 2011), but were mostly not analyzed in detail yet. Deletion of two genes coding for putative zinc-finger TFs, named dvrA and ace2, resulted in phenotypes affecting the CW. In particular, mutants lacking dvrA were more resistant to nikkomycin Z, a well-known chitinase inhibitor, and more virulent compared to the wild-type strain (Ejzykowicz et al., 2010; Verwer et al., 2012). A similar phenotype was observed for the Δace2 strain, which displayed abnormal pigmentation, as well as increased virulence (Ejzykowicz et al., 2009). Nonetheless, besides these phenotypes, it remains to be shown whether these TFs act downstream of the CWI signaling pathway in a MAPK-dependent manner.
Cross Talk Between Different Signaling Pathways
Aspergillus fumigatus is challenged by a multitude of external stimuli, each of them needing an appropriate response. Although several different signal transduction pathways exist to initiate the required transcriptional changes, they would not be able to respond to environmental signals in a balanced way if they only acted separately in a linear manner. Therefore, an interaction between the pathways is likely. For S. cerevisiae, cross talk between the CWI pathway and other signal transduction pathways under different conditions have been described (Fuchs and Mylonakis, 2009), but in A. fumigatus this interesting and complex field of research is just emerging.
In S. cerevisiae, the connection between the high osmolarity glycerol (HOG) and the CWI signaling pathway has been elucidated in detail. Under CW stress, the pathways can display co-regulatory roles depending on the stress-inducing agent. It was shown that Hog1 (the yeast SakA ortholog) was activated in the absence of Slt2 (the yeast MpkA ortholog) under conditions causing CW stress (Bermejo et al., 2008; Garcia et al., 2009), indicating an inhibitory effect of the CWI signaling on the HOG pathway.
Candida albicans also shows the previously mentioned paradoxical effect in response to high dose exposure to caspofungin. When analyzing the response to this antifungal drug, the involvement of several signal transduction cascades was discovered, including CWI, HOG, and calcium/calmodulin-dependent calcineurin signaling, suggesting an interaction of these three pathways (Wiederhold et al., 2005; Munro et al., 2007; Walker et al., 2008). For A. fumigatus the participation of calcium-mediated signaling in the paradoxical growth in response to caspofungin has been described as well (Fortwendel et al., 2010). Calcium signaling involves the Ca2+-binding protein calmodulin and the serine/threonine protein phosphatase calcineurin (Carafoli, 2005). Deletion of the calcineurin subunit A-encoding gene cnaA (also named calA) in A. fumigatus resulted in the loss of the paradoxical growth phenotype under caspofungin stress, which was attributed to the transcriptional regulation of chitin synthase-encoding genes by this signal transduction pathway (Fortwendel et al., 2010). This finding shows that the caspofungin response in A. fumigatus, apart from CWI and HOG signaling, also involves the calcium signal transduction cascade suggesting an interaction between all three pathways in this filamentous fungus, as it was described for C. albicans. This conclusion was supported by the finding that in A. nidulans the constitutive over-expression of protein kinase C (PkcA) can in part restore the wild-type phenotype of a ΔcnaA deletion mutant (Colabardini et al., 2014). In addition, CnaA affects CWI signaling by regulating MpkA phosphorylation. Moreover, PkcA has an influence on the transcription of calcium-related processes as well as on the maintenance of normal intracellular calcium levels (Colabardini et al., 2014). For the basidiomycete Cryptococcus neoformans, an interaction of the CWI pathway and calcium signaling has also been described (Kraus et al., 2003). This suggests that the cross talk between both pathways could be common to different fungal species.
Another survey shows that not only inhibitors of the calcineurin signal transduction cascade but also rapamycin, the inhibitor of the Tor signal transduction pathway, dramatically increased the effect of caspofungin on A. fumigatus (Kontoyiannis et al., 2003). As another component involved in the response to caspofungin, TOR might as well interact with CWI, HOG, or calcium signaling to coordinate the response to this drug. Altogether, this model raises the question whether in A. fumigatus the cross talk between signaling pathways is more pronounced than it is thought today.
In the last years, several transcriptomics studies of A. fumigatus signal transduction mutants were published (Malavazi et al., 2009; Jain et al., 2011; Müller et al., 2012; Macheleidt et al., 2015). The generated data suggest potential interactions of the CWI signaling cascade with other signal transduction pathways. For example, in a microarray analysis of the ΔmpkA strain, two calcium/calmodulin dependent kinases were found to be differentially regulated in the mutant compared to the wild type under stress conditions induced by glucanex, which lyses the CW (Jain et al., 2011). Furthermore, a microarray hybridisation approach comparing the transcriptional profile of the calcineurin mutant ΔcalA with the wild type, found the two MAP kinase kinase-encoding genes mkk2, involved in CWI signaling, and pbs2, involved in the HOG pathway, to be significantly down-regulated in the mutant strain (Malavazi et al., 2009). Data indicate once more a potential co-regulation of central signaling pathways in A. fumigatus. The increasing number of transcriptome analyses will lead to the identification of further interactions between signaling cascades, and most likely reveal the complexity of signal transduction in A. fumigatus.
Cell Wall Impaired Mutants and Virulence
It is common praxis in infection biology to test the virulence of mutant strains in mouse infection models, in order to define whether genes have a potential function as virulence determinant. Among the different A. fumigatus mutant strains having a defect in the CW, more than 20 were affected in virulence (Table 1). Surprisingly, many mutant strains with defects in CW biosynthesis enzymes were still virulent in the applied infection models. As an example, both the α-1,3-glucan synthase mutant strains, Δags1, and Δags2, were still pathogenic, although their α-1,3-glucan content was reduced by 50% compared to the wild-type strain (Beauvais et al., 2005). Moreover, the Δags3 mutant was reported to be even hyper-virulent (Maubon et al., 2006). To our knowledge, only the deletion of the genes encoding for a β-1,3-glucanosyltransferase (gel2) and a α-1,2-mannosyltransferase (mnt1) resulted in a decrease of virulence (Mouyna et al., 2005; Wagener et al., 2008).
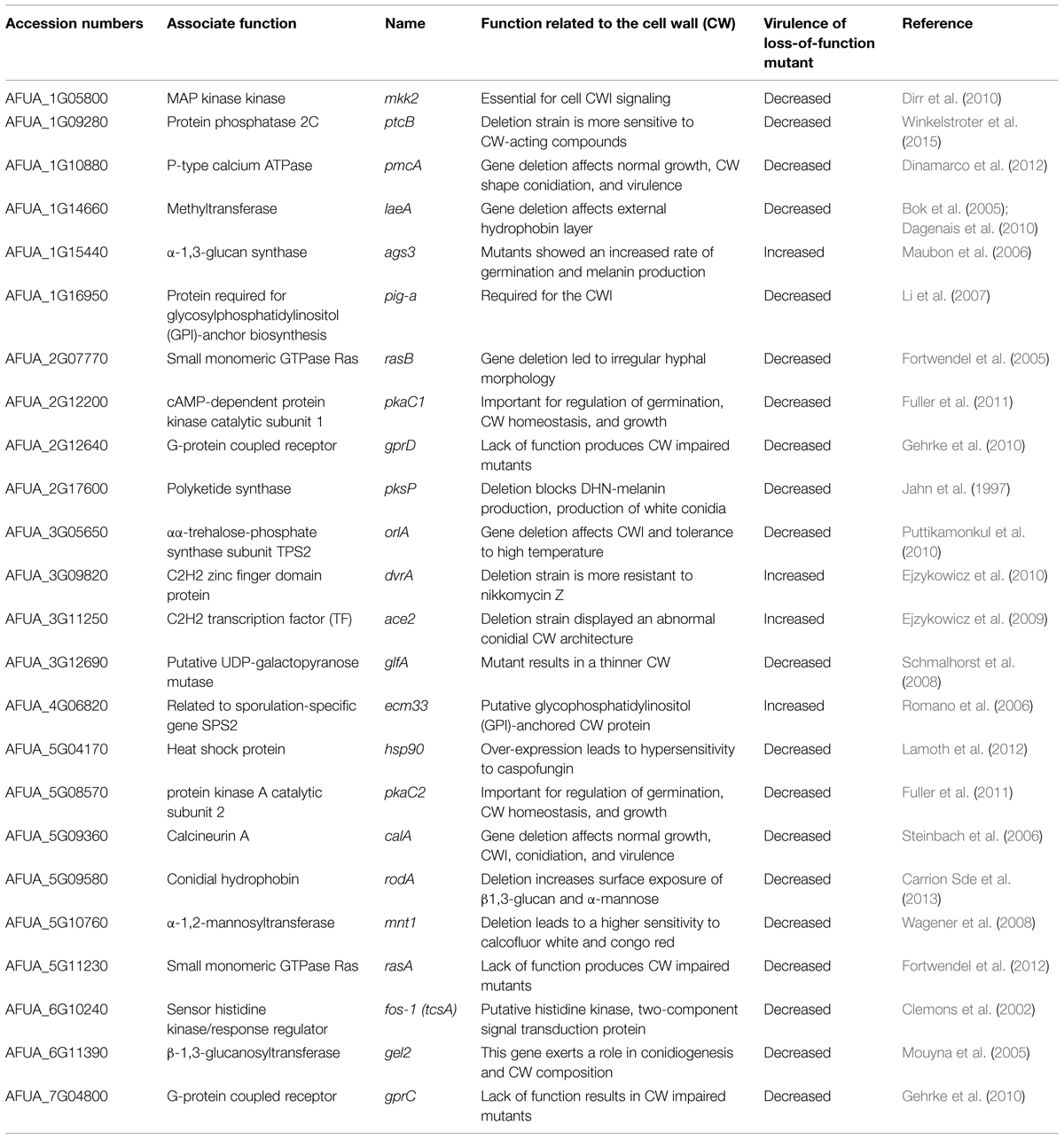
TABLE 1. The table lists all A. fumigatus mutant strains that show an altered CW structure and were reported to be involved in virulence.
As reported above, structuring of the CW depends on different signaling pathways. The observation that these pathways are connected to each other is increasingly acknowledged. Deletion of a single gene often affected more than one specific signaling pathway. In fact, many of the reported mutants with impaired CW, which displayed decreased virulence, were obtained by deleting genes putatively involved in signaling, such as ΔcalA, ΔrasA, and B, and ΔpkaC mutants (Fortwendel et al., 2005, 2012; Steinbach et al., 2006; Fuller et al., 2011). Recently, it was reported that the deletion of ptcB, a putative protein phosphatase 2C, positively affected the phosphorylation status of both MpkA and SakA, resulting in decreased virulence (Winkelstroter et al., 2015). Similarly, blocking of CWI signaling led to reduced virulence (Dirr et al., 2010). However, blocking of CWI signaling also resulted in various physiological alterations, which are apparently not directly connected to the CW (e.g., alteration of secondary metabolism; Jain et al., 2011).
Perspective
A major question concerns the selection of suitable and effective targets for future antifungal drugs. The three clinically important classes of antifungal drugs used against A. fumigatus target either enzymatic steps involved in cell membrane/CW biosynthesis or ergosterol as part of the cytoplasmic membrane. These drugs show some limitations either because they cannot really clear A. fumigatus infection, or because resistant strains were isolated and are therefore of major clinical concern. The increase of life-threatening mycoses accompanied with the lack of effective drugs, has fostered the search for new, broad-spectrum fungicidal agents. Drug efficacy could also be increased by the reformulation of existing antifungals as well as the search for synergistically acting compounds that can be used for treatment and prophylaxis. As an example, the effectiveness of caspofungin in prophylaxis is still a matter of debate, and there are studies indicating that caspofungin did not decrease the mortality rate of patients with diagnosed invasive aspergillosis (Karthaus, 2011). In future, because of their low toxicity, echinocandins might well be used in a combinatorial antifungal therapy with other synergistically acting drugs (Deresinski and Stevens, 2003). In line, it was already reported that the combination of caspofungin with azoles or AmB increased caspofungin activity in vitro (Dannaoui et al., 2004; Liu et al., 2012). Additionally, the combination of these drugs improved the efficacy of treatments of patients with severe fungal infections, in which the first line therapy failed (Nivoix et al., 2006).
The fungal CW still remains a very powerful target for antifungal drugs. However, recent studies suggest that the signaling pathways responsible for CW formation have not been completely elucidated. It remains to be shown which signaling pathways act as compensatory pathways that decrease the effectiveness of drug treatments. The identification of such pathways could lead to the discovery of new targets and new modes of action that can be exploited to potentiate the efficiency of known drugs, and to improve prophylaxis against invasive mycoses.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the excellence graduate school Jena School for Microbial Communication (JSMC) and the ERA-NET PathoGenoMics program ANTIFUN (The CW as a target to improve antifungal therapy against Aspergillosis), BFU2008-04709-E/BMC (German Federal Ministry of Education and Research: BMBF FKZ 0315439).
References
Aimanianda, V., Bayry, J., Bozza, S., Kniemeyer, O., Perruccio, K., Elluru, S. R., et al. (2009). Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature 460, 1117–1121. doi: 10.1038/nature08264
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Alcazar-Fuoli, L., and Mellado, E. (2012). Ergosterol biosynthesis in Aspergillus fumigatus: its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 3:439. doi: 10.3389/fmicb.2012.00439
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Arendrup, M. C., Garcia-Effron, G., Buzina, W., Mortensen, K. L., Reiter, N., Lundin, C., et al. (2009). Breakthrough Aspergillus fumigatus and Candida albicans double infection during caspofungin treatment: laboratory characteristics and implication for susceptibility testing. Antimicrob. Agents Chemother. 53, 1185–1193. doi: 10.1128/AAC.01292-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Aufauvre-Brown, A., Mellado, E., Gow, N. A., and Holden, D. W. (1997). Aspergillus fumigatus chsE: a gene related to CHS3 of Saccharomyces cerevisiae and important for hyphal growth and conidiophore development but not pathogenicity. Fungal. Genet. Biol. 21, 141–152. doi: 10.1006/fgbi.1997.0959
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Baginski, M., Resat, H., and Mccammon, J. A. (1997). Molecular properties of amphotericin B membrane channel: a molecular dynamics simulation. Mol. Pharmacol. 52, 560–570.
Baldin, C., Valiante, V., Krüger, T., Schafferer, L., Haas, H., Kniemeyer, O., et al. (2015). Comparative proteomics of a tor inducible Aspergillus fumigatus mutant reveals involvement of the Tor kinase in iron regulation. Proteomics doi: 10.1002/pmic.201400584 [Epub ahead of print].
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beauvais, A., Bruneau, J. M., Mol, P. C., Buitrago, M. J., Legrand, R., and Latge, J. P. (2001). Glucan synthase complex of Aspergillus fumigatus. J. Bacteriol. 183, 2273–2279. doi: 10.1128/JB.183.7.2273-2279.2001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beauvais, A., Maubon, D., Park, S., Morelle, W., Tanguy, M., Huerre, M., et al. (2005). Two alpha(1-3) glucan synthases with different functions in Aspergillus fumigatus. Appl. Environ. Microbiol. 71, 1531–1538. doi: 10.1128/AEM.71.3.1531-1538.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bermejo, C., Rodriguez, E., Garcia, R., Rodriguez-Pena, J. M., Rodriguez De La Concepcion, M. L., Rivas, C., et al. (2008). The sequential activation of the yeast HOG and SLT2 pathways is required for cell survival to cell wall stress. Mol. Biol. Cell 19, 1113–1124. doi: 10.1091/mbc.E07-08-0742
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Blum, G., Hortnagl, C., Jukic, E., Erbeznik, T., Pumpel, T., Dietrich, H., et al. (2013). New insight into amphotericin B resistance in Aspergillus terreus. Antimicrob. Agents Chemother. 57, 1583–1588. doi: 10.1128/AAC.01283-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bok, J. W., Balajee, S. A., Marr, K. A., Andes, D., Nielsen, K. F., Frisvad, J. C., et al. (2005). LaeA, a regulator of morphogenetic fungal virulence factors. Eukaryot. Cell 4, 1574–1582. doi: 10.1128/EC.4.9.1574-1582.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bolard, J. (1986). How do the polyene macrolide antibiotics affect the cellular membrane properties? Biochim. Biophys. Acta 864, 257–304. doi: 10.1016/0304-4157(86)90002-X
Bowman, J. C., Hicks, P. S., Kurtz, M. B., Rosen, H., Schmatz, D. M., Liberator, P. A., et al. (2002). The antifungal echinocandin caspofungin acetate kills growing cells of Aspergillus fumigatus in vitro. Antimicrob. Agents Chemother. 46, 3001–3012. doi: 10.1128/AAC.46.9.3001-3012.2002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brand, A., and Gow, N. A. (2009). Mechanisms of hypha orientation of fungi. Curr. Opin. Microbiol. 12, 350–357. doi: 10.1016/j.mib.2009.05.007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bruggemann, R. J., Alffenaar, J. W., Blijlevens, N. M., Billaud, E. M., Kosterink, J. G., Verweij, P. E., et al. (2009). Clinical relevance of the pharmacokinetic interactions of azole antifungal drugs with other coadministered agents. Clin. Infect. Dis. 48, 1441–1458. doi: 10.1086/598327
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bruns, S., Seidler, M., Albrecht, D., Salvenmoser, S., Remme, N., Hertweck, C., et al. (2010). Functional genomic profiling of Aspergillus fumigatus biofilm reveals enhanced production of the mycotoxin gliotoxin. Proteomics 10, 3097–3107. doi: 10.1002/pmic.201000129
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Camps, S. M., Dutilh, B. E., Arendrup, M. C., Rijs, A. J., Snelders, E., Huynen, M. A., et al. (2012). Discovery of a HapE mutation that causes azole resistance in Aspergillus fumigatus through whole genome sequencing and sexual crossing. PLoS ONE 7:e50034. doi: 10.1371/journal.pone.0050034
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cao, W., Maruyama, J., Kitamoto, K., Sumikoshi, K., Terada, T., Nakamura, S., et al. (2009). Using a new GPI-anchored-protein identification system to mine the protein databases of Aspergillus fumigatus, Aspergillus nidulans, and Aspergillus oryzae. J. Gen. Appl. Microbiol. 55, 381–393. doi: 10.2323/jgam.55.381
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Carafoli, E. (2005). Calcium–a universal carrier of biological signals. Delivered on 3 July 2003 at the special FEBS meeting in Brussels. FEBS J. 272, 1073–1089. doi: 10.1111/j.1742-4658.2005.04546.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Carrion Sde, J., Leal, S. M. Jr., Ghannoum, M. A., Aimanianda, V., Latge, J. P., and Pearlman, E. (2013). The RodA hydrophobin on Aspergillus fumigatus spores masks dectin-1- and dectin-2-dependent responses and enhances fungal survival in vivo. J. Immunol. 191, 2581–2588. doi: 10.4049/jimmunol.1300748
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cerqueira, G. C., Arnaud, M. B., Inglis, D. O., Skrzypek, M. S., Binkley, G., Simison, M., et al. (2014). The Aspergillus genome database: multispecies curation and incorporation of RNA-Seq data to improve structural gene annotations. Nucleic Acids Res. 42, D705–D710. doi: 10.1093/nar/gkt1029
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chattopadhyay, M. K., Tabor, C. W., and Tabor, H. (2002). Absolute requirement of spermidine for growth and cell cycle progression of fission yeast (Schizosaccharomyces pombe). Proc. Natl. Acad. Sci. U.S.A. 99, 10330–10334. doi: 10.1073/pnas.162362899
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Clemons, K. V., Miller, T. K., Selitrennikoff, C. P., and Stevens, D. A. (2002). fos-1, a putative histidine kinase as a virulence factor for systemic aspergillosis. Med. Mycol. 40, 259–262. doi: 10.1080/714031112
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Colabardini, A. C., Ries, L. N., Brown, N. A., Savoldi, M., Dinamarco, T. M., Von Zeska, M. R., et al. (2014). Protein kinase C overexpression suppresses calcineurin-associated defects in Aspergillus nidulans and is involved in mitochondrial function. PLoS ONE 9:e104792. doi: 10.1371/journal.pone.0104792
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cowen, L. E., and Steinbach, W. J. (2008). Stress, drugs, and evolution: the role of cellular signaling in fungal drug resistance. Eukaryot. Cell 7, 747–764. doi: 10.1128/EC.00041-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dagenais, T. R., Giles, S. S., Aimanianda, V., Latge, J. P., Hull, C. M., and Keller, N. P. (2010). Aspergillus fumigatus LaeA-mediated phagocytosis is associated with a decreased hydrophobin layer. Infect. Immun. 78, 823–829. doi: 10.1128/IAI.00980-09
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dannaoui, E., Lortholary, O., and Dromer, F. (2004). In vitro evaluation of double and triple combinations of antifungal drugs against Aspergillus fumigatus and Aspergillus terreus. Antimicrob. Agents Chemother. 48, 970–978. doi: 10.1128/AAC.48.3.970-978.2004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Deresinski, S. C., and Stevens, D. A. (2003). Caspofungin. Clin. Infect. Dis. 36, 1445–1457. doi: 10.1086/375080
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
De Souza, C. P., Hashmi, S. B., Osmani, A. H., Andrews, P., Ringelberg, C. S., Dunlap, J. C., et al. (2013). Functional analysis of the Aspergillus nidulans kinome. PLoS ONE 8:e58008. doi: 10.1371/journal.pone.0058008
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dichtl, K., Ebel, F., Dirr, F., Routier, F. H., Heesemann, J., and Wagener, J. (2010). Farnesol misplaces tip-localized Rho proteins and inhibits cell wall integrity signalling in Aspergillus fumigatus. Mol. Microbiol. 76, 1191–1204. doi: 10.1111/j.1365-2958.2010.07170.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dichtl, K., Helmschrott, C., Dirr, F., and Wagener, J. (2012). Deciphering cell wall integrity signalling in Aspergillus fumigatus: identification and functional characterization of cell wall stress sensors and relevant Rho GTPases. Mol. Microbiol. 83, 506–519. doi: 10.1111/j.1365-2958.2011.07946.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dichtl, K., Samantaray, S., Aimanianda, V., Zhu, Z., Prevost, M. C., Latge, J. P., et al. (2015). Aspergillus fumigatus devoid of cell wall beta-1,3-glucan is viable, massively sheds galactomannan and is killed by septum formation inhibitors. Mol. Microbiol. 95, 458–471. doi: 10.1111/mmi.12877
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dinamarco, T. M., Freitas, F. Z., Almeida, R. S., Brown, N. A., Dos Reis, T. F., Ramalho, L. N., et al. (2012). Functional characterization of an Aspergillus fumigatus calcium transporter (PmcA) that is essential for fungal infection. PLoS ONE 7:e37591. doi: 10.1371/journal.pone.0037591
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dirr, F., Echtenacher, B., Heesemann, J., Hoffmann, P., Ebel, F., and Wagener, J. (2010). AfMkk2 is required for cell wall integrity signaling, adhesion, and full virulence of the human pathogen Aspergillus fumigatus. Int. J. Med. Microbiol. 300, 496–502. doi: 10.1016/j.ijmm.2010.03.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dupres, V., Alsteens, D., Wilk, S., Hansen, B., Heinisch, J. J., and Dufrene, Y. F. (2009). The yeast Wsc1 cell surface sensor behaves like a nanospring in vivo. Nat. Chem. Biol. 5, 857–862. doi: 10.1038/nchembio.220
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Eddouzi, J., Parker, J. E., Vale-Silva, L. A., Coste, A., Ischer, F., Kelly, S., et al. (2013). Molecular mechanisms of drug resistance in clinical Candida species isolated from Tunisian hospitals. Antimicrob. Agents Chemother. 57, 3182–3193. doi: 10.1128/AAC.00555-13
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ejzykowicz, D. E., Cunha, M. M., Rozental, S., Solis, N. V., Gravelat, F. N., Sheppard, D. C., et al. (2009). The Aspergillus fumigatus transcription factor Ace2 governs pigment production, conidiation and virulence. Mol. Microbiol. 72, 155–169. doi: 10.1111/j.1365-2958.2009.06631.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ejzykowicz, D. E., Solis, N. V., Gravelat, F. N., Chabot, J., Li, X., Sheppard, D. C., et al. (2010). Role of Aspergillus fumigatus DvrA in host cell interactions and virulence. Eukaryot. Cell 9, 1432–1440. doi: 10.1128/EC.00055-10
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fang, Y., Hu, L., Zhou, X., Jaiseng, W., Zhang, B., Takami, T., et al. (2012). A genomewide screen in Schizosaccharomyces pombe for genes affecting the sensitivity of antifungal drugs that target ergosterol biosynthesis. Antimicrob. Agents Chemother. 56, 1949–1959. doi: 10.1128/AAC.05126-11
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Firon, A., Beauvais, A., Latge, J. P., Couve, E., Grosjean-Cournoyer, M. C., and D’enfert, C. (2002). Characterization of essential genes by parasexual genetics in the human fungal pathogen Aspergillus fumigatus: impact of genomic rearrangements associated with electroporation of DNA. Genetics 161, 1077–1087.
Fortwendel, J. R., Juvvadi, P. R., Perfect, B. Z., Rogg, L. E., Perfect, J. R., and Steinbach, W. J. (2010). Transcriptional regulation of chitin synthases by calcineurin controls paradoxical growth of Aspergillus fumigatus in response to caspofungin. Antimicrob. Agents Chemother. 54, 1555–1563. doi: 10.1128/AAC.00854-09
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fortwendel, J. R., Juvvadi, P. R., Rogg, L. E., Asfaw, Y. G., Burns, K. A., Randell, S. H., et al. (2012). Plasma membrane localization is required for RasA-mediated polarized morphogenesis and virulence of Aspergillus fumigatus. Eukaryot. Cell 11, 966–977. doi: 10.1128/EC.00091-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fortwendel, J. R., Zhao, W., Bhabhra, R., Park, S., Perlin, D. S., Askew, D. S., et al. (2005). A fungus-specific ras homolog contributes to the hyphal growth and virulence of Aspergillus fumigatus. Eukaryot. Cell 4, 1982–1989. doi: 10.1128/EC.4.12.1982-1989.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fuchs, B. B., and Mylonakis, E. (2009). Our paths might cross: the role of the fungal cell wall integrity pathway in stress response and cross talk with other stress response pathways. Eukaryot. Cell 8, 1616–1625. doi: 10.1128/EC.00193-09
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fuller, K. K., Richie, D. L., Feng, X., Krishnan, K., Stephens, T. J., Wikenheiser-Brokamp, K. A., et al. (2011). Divergent Protein Kinase A isoforms co-ordinately regulate conidial germination, carbohydrate metabolism and virulence in Aspergillus fumigatus. Mol. Microbiol. 79, 1045–1062. doi: 10.1111/j.1365-2958.2010.07509.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Garcia, R., Rodriguez-Pena, J. M., Bermejo, C., Nombela, C., and Arroyo, J. (2009). The high osmotic response and cell wall integrity pathways cooperate to regulate transcriptional responses to zymolyase-induced cell wall stress in Saccharomyces cerevisiae. J. Biol. Chem. 284, 10901–10911. doi: 10.1074/jbc.M808693200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gastebois, A., Mouyna, I., Simenel, C., Clavaud, C., Coddeville, B., Delepierre, M., et al. (2010). Characterization of a new beta(1-3)-glucan branching activity of Aspergillus fumigatus. J. Biol. Chem. 285, 2386–2396. doi: 10.1074/jbc.M109.077545
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gautam, P., Shankar, J., Madan, T., Sirdeshmukh, R., Sundaram, C. S., Gade, W. N., et al. (2008). Proteomic and transcriptomic analysis of Aspergillus fumigatus on exposure to amphotericin B. Antimicrob. Agents Chemother. 52, 4220–4227. doi: 10.1128/AAC.01431-07
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gehrke, A., Heinekamp, T., Jacobsen, I. D., and Brakhage, A. A. (2010). Heptahelical receptors GprC and GprD of Aspergillus fumigatus Are essential regulators of colony growth, hyphal morphogenesis, and virulence. Appl. Environ. Microbiol. 76, 3989–3998. doi: 10.1128/AEM.00052-10
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gray, K. C., Palacios, D. S., Dailey, I., Endo, M. M., Uno, B. E., Wilcock, B. C., et al. (2012). Amphotericin primarily kills yeast by simply binding ergosterol. Proc. Natl. Acad. Sci. U.S.A. 109, 2234–2239. doi: 10.1073/pnas.1117280109
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grice, C. M., Bertuzzi, M., and Bignell, E. M. (2013). Receptor-mediated signaling in Aspergillus fumigatus. Front. Microbiol. 4:26. doi: 10.3389/fmicb.2013.00026
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grosse, C., Heinekamp, T., Kniemeyer, O., Gehrke, A., and Brakhage, A. A. (2008). Protein kinase A regulates growth, sporulation, and pigment formation in Aspergillus fumigatus. Appl. Environ. Microbiol. 74, 4923–4933. doi: 10.1128/AEM.00470-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gsaller, F., Hortschansky, P., Beattie, S. R., Klammer, V., Tuppatsch, K., Lechner, B. E., et al. (2014). The Janus transcription factor HapX controls fungal adaptation to both iron starvation and iron excess. EMBO J. 33, 2261–2276. doi: 10.15252/embj.201489468
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Haas, H. (2014). Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 31, 1266–1276. doi: 10.1039/C4NP00071D
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heinekamp, T., Thywissen, A., Macheleidt, J., Keller, S., Valiante, V., and Brakhage, A. A. (2012). Aspergillus fumigatus melanins: interference with the host endocytosis pathway and impact on virulence. Front. Microbiol. 3:440. doi: 10.3389/fmicb.2012.00440
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Henry, C., Latge, J. P., and Beauvais, A. (2012). alpha1,3 glucans are dispensable in Aspergillus fumigatus. Eukaryot. Cell 11, 26–29. doi: 10.1128/EC.05270-11
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Herrmann, M., Sprote, P., and Brakhage, A. A. (2006). Protein kinase C (PkcA) of Aspergillus nidulans is involved in penicillin production. Appl. Environ. Microbiol. 72, 2957–2970. doi: 10.1128/AEM.72.4.2957-2970.2006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Horn, F., Heinekamp, T., Kniemeyer, O., Pollmacher, J., Valiante, V., and Brakhage, A. A. (2012). Systems biology of fungal infection. Front. Microbiol. 3:108. doi: 10.3389/fmicb.2012.00108
Hortschansky, P., Eisendle, M., Al-Abdallah, Q., Schmidt, A. D., Bergmann, S., Thon, M., et al. (2007). Interaction of HapX with the CCAAT-binding complex–a novel mechanism of gene regulation by iron. EMBO J. 26, 3157–3168. doi: 10.1038/sj.emboj.7601752
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hu, W., Sillaots, S., Lemieux, S., Davison, J., Kauffman, S., Breton, A., et al. (2007). Essential gene identification and drug target prioritization in Aspergillus fumigatus. PLoS Pathog 3:e24. doi: 10.1371/journal.ppat.0030024
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jahn, B., Koch, A., Schmidt, A., Wanner, G., Gehringer, H., Bhakdi, S., et al. (1997). Isolation and characterization of a pigmentless-conidium mutant of Aspergillus fumigatus with altered conidial surface and reduced virulence. Infect. Immun. 65, 5110–5117.
Jain, R., Valiante, V., Remme, N., Docimo, T., Heinekamp, T., Hertweck, C., et al. (2011). The MAP kinase MpkA controls cell wall integrity, oxidative stress response, gliotoxin production and iron adaptation in Aspergillus fumigatus. Mol. Microbiol. 82, 39–53. doi: 10.1111/j.1365-2958.2011.07778.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jimenez-Ortigosa, C., Aimanianda, V., Muszkieta, L., Mouyna, I., Alsteens, D., Pire, S., et al. (2012). Chitin synthases with a myosin motor-like domain control the resistance of Aspergillus fumigatus to echinocandins. Antimicrob. Agents Chemother. 56, 6121–6131. doi: 10.1128/AAC.00752-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Karthaus, M. (2011). Prophylaxis and treatment of invasive aspergillosis with voriconazole, posaconazole and caspofungin: review of the literature. Eur. J. Med. Res. 16, 145–152. doi: 10.1186/2047-783X-16-4-145
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Khot, P. D., Suci, P. A., Miller, R. L., Nelson, R. D., and Tyler, B. J. (2006). A small subpopulation of blastospores in candida albicans biofilms exhibit resistance to amphotericin B associated with differential regulation of ergosterol and beta-1,6-glucan pathway genes. Antimicrob. Agents Chemother. 50, 3708–3716. doi: 10.1128/AAC.00997-06
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kontoyiannis, D. P., Lewis, R. E., Osherov, N., Albert, N. D., and May, G. S. (2003). Combination of caspofungin with inhibitors of the calcineurin pathway attenuates growth in vitro in Aspergillus species. J. Antimicrob. Chemother. 51, 313–316. doi: 10.1093/jac/dkg090
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kraus, P. R., Fox, D. S., Cox, G. M., and Heitman, J. (2003). The Cryptococcus neoformans MAP kinase Mpk1 regulates cell integrity in response to antifungal drugs and loss of calcineurin function. Mol. Microbiol. 48, 1377–1387. doi: 10.1046/j.1365-2958.2003.03508.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lamoth, F., Juvvadi, P. R., Fortwendel, J. R., and Steinbach, W. J. (2012). Heat shock protein 90 is required for conidiation and cell wall integrity in Aspergillus fumigatus. Eukaryot. Cell 11, 1324–1332. doi: 10.1128/EC.00032-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Langfelder, K., Streibel, M., Jahn, B., Haase, G., and Brakhage, A. A. (2003). Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal. Genet. Biol. 38, 143–158. doi: 10.1016/S1087-1845(02)00526-1
Latge, J. P., and Beauvais, A. (2014). Functional duality of the cell wall. Curr. Opin. Microbiol. 20, 111–117. doi: 10.1016/j.mib.2014.05.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lelievre, L., Groh, M., Angebault, C., Maherault, A. C., Didier, E., and Bougnoux, M. E. (2013). Azole resistant Aspergillus fumigatus: an emerging problem. Med. Mal. Infect. 43, 139–145. doi: 10.1016/j.medmal.2013.02.010
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Levin, D. E. (2011). Regulation of cell wall biogenesis in Saccharomyces cerevisiae: the cell wall integrity signaling pathway. Genetics 189, 1145–1175. doi: 10.1534/genetics.111.128264
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lew, R. R. (2011). How does a hypha grow? The biophysics of pressurized growth in fungi. Nat. Rev. Microbiol. 9, 509–518. doi: 10.1038/nrmicro2591
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, H., Zhou, H., Luo, Y., Ouyang, H., Hu, H., and Jin, C. (2007). Glycosylphosphatidylinositol (GPI) anchor is required in Aspergillus fumigatus for morphogenesis and virulence. Mol. Microbiol. 64, 1014–1027. doi: 10.1111/j.1365-2958.2007.05709.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liu, W., Li, L., Sun, Y., Chen, W., Wan, Z., Li, R., et al. (2012). Interaction of the echinocandin caspofungin with amphotericin B or voriconazole against Aspergillus biofilms in vitro. Antimicrob. Agents Chemother. 56, 6414–6416. doi: 10.1128/AAC.00687-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Macheleidt, J., Scherlach, K., Neuwirth, T., Schmidt-Heck, W., Straßburger, M., Spraker, J., et al. (2015). Transcriptome analysis of cAMP-dependent protein kinase A regulated genes reveals the production of the novel natural compound fumipyrrole by Aspergillus fumigatus. Mol. Microbiol. 96, 148–162. doi: 10.1111/mmi.12926
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Malavazi, I., Da Silva Ferreira, M. E., Soriani, F. M., Dinamarco, T. M., Savoldi, M., Uyemura, S. A., et al. (2009). Phenotypic analysis of genes whose mRNA accumulation is dependent on calcineurin in Aspergillus fumigatus. Fungal Genet. Biol. 46, 791–802. doi: 10.1016/j.fgb.2009.06.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maubon, D., Park, S., Tanguy, M., Huerre, M., Schmitt, C., Prevost, M. C., et al. (2006). AGS3, an alpha(1-3)glucan synthase gene family member of Aspergillus fumigatus, modulates mycelium growth in the lung of experimentally infected mice. Fungal Genet. Biol. 43, 366–375. doi: 10.1016/j.fgb.2006.01.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mellado, E., Diaz-Guerra, T. M., Cuenca-Estrella, M., and Rodriguez-Tudela, J. L. (2001). Identification of two different 14-alpha sterol demethylase-related genes (cyp51A and cyp51B) in Aspergillus fumigatus and other Aspergillus species. J. Clin. Microbiol. 39, 2431–2438. doi: 10.1128/JCM.39.7.2431-2438.2001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mellado, E., Garcia-Effron, G., Buitrago, M. J., Alcazar-Fuoli, L., Cuenca-Estrella, M., and Rodriguez-Tudela, J. L. (2005). Targeted gene disruption of the 14-alpha sterol demethylase (cyp51A) in Aspergillus fumigatus and its role in azole drug susceptibility. Antimicrob. Agents Chemother. 49, 2536–2538. doi: 10.1128/AAC.49.6.2536-2538.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mesa-Arango, A. C., Trevijano-Contador, N., Roman, E., Sanchez-Fresneda, R., Casas, C., Herrero, E., et al. (2014). The production of reactive oxygen species is a universal action mechanism of Amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drug. Antimicrob. Agents Chemother. 58, 6627–6638. doi: 10.1128/AAC.03570-14
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Minguet, E. G., Vera-Sirera, F., Marina, A., Carbonell, J., and Blazquez, M. A. (2008). Evolutionary diversification in polyamine biosynthesis. Mol. Biol. Evol. 25, 2119–2128. doi: 10.1093/molbev/msn161
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mouyna, I., Hartl, L., and Latge, J. P. (2013). beta-1,3-glucan modifying enzymes in Aspergillus fumigatus. Front. Microbiol. 4:81. doi: 10.3389/fmicb.2013.00081
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mouyna, I., Morelle, W., Vai, M., Monod, M., Lechenne, B., Fontaine, T., et al. (2005). Deletion of GEL2 encoding for a beta(1-3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus. Mol. Microbiol. 56, 1675–1688. doi: 10.1111/j.1365-2958.2005.04654.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Müller, S., Baldin, C., Groth, M., Guthke, R., Kniemeyer, O., Brakhage, A. A., et al. (2012). Comparison of transcriptome technologies in the pathogenic fungus Aspergillus fumigatus reveals novel insights into the genome and MpkA dependent gene expression. BMC Genomics 13:519. doi: 10.1186/1471-2164-13-519
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Munro, C. A., Selvaggini, S., De Bruijn, I., Walker, L., Lenardon, M. D., Gerssen, B., et al. (2007). The PKC, HOG and Ca2+ signalling pathways co-ordinately regulate chitin synthesis in Candida albicans. Mol. Microbiol. 63, 1399–1413. doi: 10.1111/j.1365-2958.2007.05588.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Muszkieta, L., Aimanianda, V., Mellado, E., Gribaldo, S., Alcazar-Fuoli, L., Szewczyk, E., et al. (2014). Deciphering the role of the chitin synthase families 1 and 2 in the in vivo and in vitro growth of Aspergillus fumigatus by multiple gene targeting deletion. Cell Microbiol. 16, 1784–1805. doi: 10.1111/cmi.12326
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nanduri, J., and Tartakoff, A. M. (2001). The arrest of secretion response in yeast: signaling from the secretory path to the nucleus via Wsc proteins and Pkc1p. Mol. Cell. 8, 281–289. doi: 10.1016/S1097-2765(01)00312-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nivoix, Y., Zamfir, A., Lutun, P., Kara, F., Remy, V., Lioure, B., et al. (2006). Combination of caspofungin and an azole or an amphotericin B formulation in invasive fungal infections. J. Infect. 52, 67–74. doi: 10.1016/j.jinf.2005.01.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Perez, P., Garcia-Acha, I., and Duran, A. (1983). Effect of papulacandin B on the cell wall and growth of Geotrichum lactis. J. Gen. Microbiol. 129, 245–250. doi: 10.1099/00221287-129-1-245
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Puttikamonkul, S., Willger, S. D., Grahl, N., Perfect, J. R., Movahed, N., Bothner, B., et al. (2010). Trehalose 6-phosphate phosphatase is required for cell wall integrity and fungal virulence but not trehalose biosynthesis in the human fungal pathogen Aspergillus fumigatus. Mol. Microbiol. 77, 891–911. doi: 10.1111/j.1365-2958.2010.07254.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rispail, N., Soanes, D. M., Ant, C., Czajkowski, R., Grunler, A., Huguet, R., et al. (2009). Comparative genomics of MAP kinase and calcium-calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet. Biol. 46, 287–298. doi: 10.1016/j.fgb.2009.01.002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rocha, E. M., Garcia-Effron, G., Park, S., and Perlin, D. S. (2007). A Ser678Pro substitution in Fks1p confers resistance to echinocandin drugs in Aspergillus fumigatus. Antimicrob. Agents Chemother. 51, 4174–4176. doi: 10.1128/AAC.00917-07
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rogg, L. E., Fortwendel, J. R., Juvvadi, P. R., Lilley, A., and Steinbach, W. J. (2011). The chitin synthase genes chsA and chsC are not required for cell wall stress responses in the human pathogen Aspergillus fumigatus. Biochem. Biophys. Res. Commun. 411, 549–554. doi: 10.1016/j.bbrc.2011.06.180
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Romano, J., Nimrod, G., Ben-Tal, N., Shadkchan, Y., Baruch, K., Sharon, H., et al. (2006). Disruption of the Aspergillus fumigatus ECM33 homologue results in rapid conidial germination, antifungal resistance and hypervirulence. Microbiology 152, 1919–1928. doi: 10.1099/mic.0.28936-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Samantaray, S., Neubauer, M., Helmschrott, C., and Wagener, J. (2013). Role of the guanine nucleotide exchange factor Rom2 in cell wall integrity maintenance of Aspergillus fumigatus. Eukaryot. Cell 12, 288–298. doi: 10.1128/EC.00246-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sawistowska-Schroder, E. T., Kerridge, D., and Perry, H. (1984). Echinocandin inhibition of 1,3-beta-D-glucan synthase from Candida albicans. FEBS Lett. 173, 134–138. doi: 10.1016/0014-5793(84)81032-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schmaler-Ripcke, J., Sugareva, V., Gebhardt, P., Winkler, R., Kniemeyer, O., Heinekamp, T., et al. (2009). Production of pyomelanin, a second type of melanin, via the tyrosine degradation pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 75, 493–503. doi: 10.1128/AEM.02077-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schmalhorst, P. S., Krappmann, S., Vervecken, W., Rohde, M., Müller, M., Braus, G. H., et al. (2008). Contribution of galactofuranose to the virulence of the opportunistic pathogen Aspergillus fumigatus. Eukaryot. Cell 7, 1268–1277. doi: 10.1128/EC.00109-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Seo, K., Akiyoshi, H., and Ohnishi, Y. (1999). Alteration of cell wall composition leads to amphotericin B resistance in Aspergillus flavus. Microbiol. Immunol. 43, 1017–1025. doi: 10.1111/j.1348-0421.1999.tb01231.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Singh, R., Shivaprakash, M. R., and Chakrabarti, A. (2011). Biofilm formation by zygomycetes: quantification, structure and matrix composition. Microbiology 157, 2611–2618. doi: 10.1099/mic.0.048504-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Steinbach, W. J., Cramer, R. A. Jr., Perfect, B. Z., Asfaw, Y. G., Sauer, T. C., Najvar, L. K., et al. (2006). Calcineurin controls growth, morphology, and pathogenicity in Aspergillus fumigatus. Eukaryot. Cell 5, 1091–1103. doi: 10.1128/EC.00139-06
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ullmann, A. J., and Cornely, O. A. (2006). Antifungal prophylaxis for invasive mycoses in high risk patients. Curr. Opin. Infect. Dis. 19, 571–576. doi: 10.1097/QCO.0b013e3280108e45
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Valiante, V., Heinekamp, T., Jain, R., Hartl, A., and Brakhage, A. A. (2008). The mitogen-activated protein kinase MpkA of Aspergillus fumigatus regulates cell wall signaling and oxidative stress response. Fungal Genet. Biol. 45, 618–627. doi: 10.1016/j.fgb.2007.09.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Valiante, V., Jain, R., Heinekamp, T., and Brakhage, A. A. (2009). The MpkA MAP kinase module regulates cell wall integrity signaling and pyomelanin formation in Aspergillus fumigatus. Fungal Genet. Biol. 46, 909–918. doi: 10.1016/j.fgb.2009.08.005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vandeputte, P., Tronchin, G., Larcher, G., Ernoult, E., Berges, T., Chabasse, D., et al. (2008). A nonsense mutation in the ERG6 gene leads to reduced susceptibility to polyenes in a clinical isolate of Candida glabrata. Antimicrob. Agents Chemother. 52, 3701–3709. doi: 10.1128/AAC.00423-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Verwer, P. E., Van Duijn, M. L., Tavakol, M., Bakker-Woudenberg, I. A., and Van De Sande, W. W. (2012). Reshuffling of Aspergillus fumigatus cell wall components chitin and beta-glucan under the influence of caspofungin or nikkomycin Z alone or in combination. Antimicrob. Agents Chemother. 56, 1595–1598. doi: 10.1128/AAC.05323-11
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wagener, J., Echtenacher, B., Rohde, M., Kotz, A., Krappmann, S., Heesemann, J., et al. (2008). The putative alpha-1,2-mannosyltransferase AfMnt1 of the opportunistic fungal pathogen Aspergillus fumigatus is required for cell wall stability and full virulence. Eukaryot. Cell 7, 1661–1673. doi: 10.1128/EC.00221-08
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Walker, L. A., Munro, C. A., De Bruijn, I., Lenardon, M. D., Mckinnon, A., and Gow, N. A. (2008). Stimulation of chitin synthesis rescues Candida albicans from echinocandins. PLoS Pathog 4:e1000040. doi: 10.1371/journal.ppat.1000040
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wiederhold, N. P. (2007). Attenuation of echinocandin activity at elevated concentrations: a review of the paradoxical effect. Curr. Opin. Infect. Dis. 20, 574–578. doi: 10.1097/QCO.0b013e3282f1be7f
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wiederhold, N. P. (2009). Paradoxical echinocandin activity: a limited in vitro phenomenon? Med. Mycol. 47(Suppl. 1), S369–S375. doi: 10.1080/13693780802428542
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wiederhold, N. P., Kontoyiannis, D. P., Prince, R. A., and Lewis, R. E. (2005). Attenuation of the activity of caspofungin at high concentrations against Candida albicans: possible role of cell wall integrity and calcineurin pathways. Antimicrob. Agents Chemother. 49, 5146–5148. doi: 10.1128/AAC.49.12.5146-5148.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Winkelstroter, L. K., Bom, V. L., De Castro, P. A., Ramalho, L. N., Goldman, M. H., Brown, N. A., et al. (2015). High Osmolarity Glycerol response (HOG) PtcB phosphatase is important for Aspergillus fumigatus virulence. Mol. Microbiol. 96, 42–54. doi: 10.1111/mmi.12919
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Young, L. Y., Hull, C. M., and Heitman, J. (2003). Disruption of ergosterol biosynthesis confers resistance to amphotericin B in Candida lusitaniae. Antimicrob. Agents Chemother. 47, 2717–2724. doi: 10.1128/AAC.47.9.2717-2724.2003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhao, W., Li, C., Liang, J., and Sun, S. (2014). The Aspergillus fumigatus beta-1,3-glucanosyltransferase Gel7 plays a compensatory role in maintaining cell wall integrity under stress conditions. Glycobiology 24, 418–427. doi: 10.1093/glycob/cwu003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: Aspergillus fumigatus, cell wall integrity, signaling pathways, virulence, mitogen activated protein kinases (MAPKs)
Citation: Valiante V, Macheleidt J, Föge M and Brakhage AA (2015) The Aspergillus fumigatus cell wall integrity signaling pathway: drug target, compensatory pathways, and virulence. Front. Microbiol. 6:325. doi: 10.3389/fmicb.2015.00325
Received: 10 February 2015; Accepted: 31 March 2015;
Published online: 16 April 2015
Edited by:
Praveen Rao Juvvadi, Duke University Medical Center, USAReviewed by:
Vishukumar Aimanianda, Pasteur Institute, FranceJohannes Wagener, Ludwig Maximilian University Munich, Germany
Copyright © 2015 Valiante, Macheleidt, Föge and Brakhage. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vito Valiante, Molecular Biotechnology of Natural Products, Department of Molecular and Applied Microbiology, Leibniz Institute for Natural Product Research and Infection Biology – Hans Knöll Institute, Beutenbergstrasse 11a, 07745 Jena, Germany vito.valiante@hki-jena.de