 Elisa Flávia L. C. Bailão1†
Elisa Flávia L. C. Bailão1† Patrícia de Sousa Lima1
Patrícia de Sousa Lima1 Mirelle G. Silva-Bailão1†
Mirelle G. Silva-Bailão1† Alexandre M. Bailão1
Alexandre M. Bailão1 Gabriel da Rocha Fernandes2†
Gabriel da Rocha Fernandes2† Daniel J. Kosman3
Daniel J. Kosman3 Célia Maria de Almeida Soares1*
Célia Maria de Almeida Soares1*- 1Laboratório de Biologia Molecular, Instituto de Ciências Biológicas, Universidade Federal de Goiás, Goiânia, Brazil
- 2Laboratório de Bioinformática, Universidade Católica de Brasília, Brasília, Brazil
- 3Department of Biochemistry, School of Medicine and Biomedical Sciences, State University of New York at Buffalo, Buffalo, NY, USA
Iron is an essential micronutrient for almost all organisms, including fungi. Usually, fungi can uptake iron through receptor-mediated internalization of a siderophore or heme, and/or reductive iron assimilation (RIA). Traditionally, the RIA pathway consists of ferric reductases (Fres), ferroxidase (Fet3) and a high-affinity iron permease (Ftr1). Paracoccidioides spp. genomes do not present an Ftr1 homolog. However, this fungus expresses zinc regulated transporter homologs (Zrts), members of the ZIP family of membrane transporters that are able in some organisms to transport zinc and iron. A 2,3,5-triphenyltetrazolium chloride (TTC)-overlay assay indicates that both Pb01 and Pb18 express a ferric reductase activity; however, 59Fe uptake assays indicate that only in Pb18 is this activity coupled to a reductase-dependent iron uptake pathway. In addition, Zrts are up-regulated in iron deprivation, as indicated by RNAseq and qRT-PCR using Pb01 transcripts. RNAseq strategy also demonstrated that transcripts related to siderophore uptake and biosynthesis are up-regulated in iron-deprived condition. The data suggest that the fungus could use both a non-classical RIA, comprising ferric reductases and Fe/Zn permeases (Zrts), and siderophore uptake pathways under iron-limited conditions. The study of iron metabolism reveals novel surface molecules that could function as accessible targets for drugs to block iron uptake and, consequently, inhibit pathogen's proliferation.
Introduction
Iron is the most common cofactor in biology. This fact could be explained by the high abundance of the element in nature and by its chemical properties, mainly by its redox ability. Since iron participates in several metabolic pathways, either aerobes or anaerobes must obtain ferrous iron (Fe2+), since ferric iron (Fe3+) is insoluble. Typically, fungi accumulate iron through two different strategies: (1) receptor-mediated internalization of ferric-siderophore complexes and/or heme group; and (2) reductive iron assimilation (RIA), involving iron reduction, followed by a ferroxidase-permeation step (Kosman, 2013). The first strategy has been recently described in Paracoccidioides spp. It has been shown that this fungus is able to utilize siderophores and hemoglobin as iron sources through receptor-mediated pathways (Bailão et al., 2014; Silva-Bailão et al., 2014). However, the RIA in Paracoccidioides spp. remains elusive.
Ferric reductases have a central role in both strategies of iron capture (Baek et al., 2008). These enzymes, along with ferroxidases and permeases, promote high-affinity iron uptake (Kosman, 2003). Moreover, ferric reductases are important in removing iron from siderophores (Yun et al., 2001) or from host iron sources, such as transferrin and hemin (Knight et al., 2005; Saikia et al., 2014). In addition to the essential role in iron uptake, ferric reductases are also important for melanin production, resistance to azole drugs, virulence (Saikia et al., 2014) and host adaptation (Hu et al., 2014) in Cryptococcus neoformans. In C. albicans, the ferric reductases also participate in the oxidative stress response, filamentous development and virulence (Xu et al., 2014). Besides the transmembrane ferric reductase enzymes, a secreted γ-glutamyltransferase (Ggt1) activity has been described in Histoplasma capsulatum, Blastomyces dermatitidis, Paracoccidioides sp. and Sporothrix schenckii (Zarnowski and Woods, 2005).
Multicopper oxidases (MCOs) consist of four enzyme superfamilies: laccases, ascorbate oxidases, ferroxidases, and ceruloplasmin, that catalyze the four-electron reduction of molecular oxygen to two molecules of water (Sirim et al., 2011). In fungal ferroxidases, as noted for Saccharomyces cerevisiae Fet3, the specificity for Fe2+ as an electron donor derives from the presence of two carboxyl groups, E185 and D409, which are in H-bond contact with the T1 Cu ligands H489 and H413, respectively (Kosman, 2010). In most fungi, the Fe3+ product derived from a ferroxidase reaction is channeled to a high-affinity iron permease, Ftr1, which is associated to the ferroxidase in the fungal plasma membrane (Kwok et al., 2006; Ziegler et al., 2011).
The Paracoccidioides genus comprises thermodimorphic fungal pathogens causing paracoccidioidomycosis (PCM), which is a deep systemic mycosis with a high prevalence in Brazil, Colombia, Venezuela, and Argentina (San-Blas et al., 2002). Since the notification of PCM cases is not obligatory, prevalence of this fungal disease is likely significantly under-reported (Martinez, 2010). The treatment available for PCM is long and may include several side effects and pathogen resistance. Thus, there is a great demand for the development of safer alternative therapies that are able to overcome resistance (Bocca et al., 2013). In this context, blocking iron acquisition is a good strategy to prevent or treat fungal diseases since iron is an essential nutrient for pathogen proliferation in vertebrate hosts (Kronstad et al., 2013). In this way, the knowledge about Paracoccidioides spp. iron acquisition mechanisms can contribute to reveal new targets to antifungal therapy.
The iron uptake mechanisms in Paracoccidioides spp. have been investigated in part (Parente et al., 2011; Silva et al., 2011; Bailão et al., 2014; Silva-Bailão et al., 2014). It has been demonstrated by our group that Paracoccidioides spp. presents multiple ferric reductases and MCOs and a ggt1 homolog, but no ftr1 homolog (Silva et al., 2011). Moreover, it was observed that ferric reductase transcripts (Bailão et al., 2006, 2007, 2012a), zinc regulated transporters (Zrts) homologs (Bailão et al., 2006), hemoglobin receptor homologs and transcripts related to siderophore synthesis and uptake (Parente et al., 2011; Silva-Bailão et al., 2014) are induced during in vitro iron deprivation or when in vivo models of infection were used. Siderophore production and acquisition and host iron sources have been investigated in Paracoccidioides spp. In iron-depleted condition, siderophore secretion by this fungus increases. Moreover, Paracoccidioides spp. are able to use siderophores as iron source, increasing the fungus ability to survive inside macrophages, a poor-iron environment (Silva-Bailão et al., 2014). We also demonstrated that hemoglobin is the preferential host iron source for Paracoccidioides spp. To acquire hemoglobin, the fungus presents hemolytic activity and the ability to internalize the entire molecule instead of promoting the iron release extracellularly. A GPI-anchored hemoglobin receptor, Rbt5, is a virulence factor (Bailão et al., 2014). Since Paracoccidioides spp. do not express an Ftr1 homolog, Fre homologs could reduce the iron, which could be imported by iron/zinc permeases (Bailão et al., 2007; Silva et al., 2011), but this hypothesis remains elusive.
In this work, the RIA pathway was investigated. We have demonstrated that Paracoccidioides spp. is able to reduce iron, and in the case of Pb18, this reductase activity is linked to ferric iron uptake. In contrast, this reductase activity in Pb01 does not appear to be up-stream from this uptake. After reduction, the data suggest that Fe2+ is probably internalized through a Fe/Zn permease (Zrt). This suggestion is because Paracoccidioides spp. genomes do not present an ftr1 homolog and the zrt1 and zrt2 transcripts are up-regulated during iron deprivation. In addition, transcripts related to siderophore uptake and biosynthesis are up-regulated upon iron deprivation. The data suggest that the fungus could use both a non-classical RIA, comprising ferric reductases and Fe/Zn permeases, and siderophore uptake pathways under iron-limited conditions.
Materials and Methods
Strains and Growth Conditions
Paracoccidioides Pb01 (ATCC MYA-826; Paracoccidioides lutzii) (Teixeira et al., 2014) and Pb18 (ATCC 32069; Paracoccidioides brasiliensis, phylogenetic species S1) (Carrero et al., 2008) were used in this work. The fungus was maintained in brain heart infusion (BHI) medium supplemented with 4% (w/v) glucose at 36°C to cultivate the yeast form. Before each experiment, the cells were grown in liquid BHI supplemented with 4% (w/v) glucose for 72 h at 36°C under rotation.
In silico Sequences Analysis
Pb01 and Pb18 putative ferric reductase and ferroxidase amino acid sequences were obtained in the Paracoccidioides genome database (http://www.broadinstitute.org/annotation/genome/paracoccidioides_brasiliensis/MultiHome.html). The Paracoccidioides spp. sequences were compared using the ClustalX2 program (Larkin et al., 2007). Comparisons were performed with amino acid sequences from other fungi, as following: Aspergillus fumigatus (http://www.aspergillusgenome.org), Aspergillus nidulans (http://www.broadinstitute.org/annotation/fungi/aspergillus_nidulans_old), H. capsulatum (http://www.broadinstitute.org/annotation/genome/histoplasma_capsulatum/MultiHome.html). Coccidioides immitis, Coccidioides posadasii (http://www.broadinstitute.org/annotation/genome/coccidioides_group/MultiHome.html), B. dermatitidis (http://www.broadinstitute.org/annotation/genome/blastomyces_dermatitidis/MultiHome.html), Ustilago maydis (http://www.broadinstitute.org/annotation/genome/ustilago_maydis/Home.html), C. neoformans (http://www.broadinstitute.org/annotation/genome/cryptococcus_neoformans/MultiHome.html), S. pombe (http://www.pombase.org), C. albicans (http://www.candidagenome.org/), and S. cerevisiae (http://www.yeastgenome.org). ClustalX2 program (Larkin et al., 2007) and TreeView v.1.6.6 program (Page, 1996) were used for phylogenetic analysis and visualization, respectively, applying the neighbor-joining method and the tree architecture was inferred from 1000 bootstraps. Domains in amino acid sequences were localized using SMART online tool (http://smart.embl-heidelberg.de/).
A structural model of the putative ferroxidase, Pb01 PAAG_06004 was obtained using the Modeler program (Sali et al., 1995) at the Max-Planck Institute for Developmental Biology website (http://toolkit.tuebingen.mpg.de/modeller#). The input for the model used the structure of S. cerevisiae Fet3 as template (PDB 1ZPU). InsightII was used subsequently to further energy minimize the model using the cvff forcefield with 5000 iterations and a CG convergence of 1.0.
RNA Extraction and Quantitative Real Time PCR (qRT-PCR)
Pb01 and Pb18 yeast cells were incubated in Synthetic Complete medium (SC medium: 6.67 g/l yeast nitrogen base without amino acids, 2% glucose plus amino acids mixture) with no supplementation or with addition of 200 μM of bathophenanthroline disulfonic acid (BPS) or 10 μM FeCl3 at 36°C under rotation. After 2 h, the cells were harvested and total RNA was extracted using TRIzol (TRI Reagent, Sigma-Aldrich, St. Louis, MO, USA) and mechanical cell rupture (Mini-Beadbeater—Biospec Products Inc., Bartlesville, OK). The total RNA was treated with DNAse I (Promega Corporation, Madison, WI, USA) and used as template in in vitro reverse transcription (SuperScript III First-Strand Synthesis SuperMix; Invitrogen, Life Technologies). Then, the cDNAs were submitted to a qRT-PCR reaction, which was performed using SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA) in a StepOnePlus Real-Time PCR System (Applied Biosystems Inc.). The expression values were calculated using the transcript that encoded alpha tubulin (XM_002796593) as the endogenous control as previously reported (Bailão et al., 2012b). The annealing temperature for all primers was 62°C. The qRT-PCR reaction was performed in biological triplicate for each cDNA sample, and a melting curve analysis was performed to confirm single PCR products. The relative standard curve was generated using a pool of cDNAs from all the conditions that were used, which was serially diluted 1:5–1:625. Relative expression levels of transcripts of interest were calculated using the standard curve method for relative quantification (Bookout et al., 2006). Student's t-test was applied in the statistical analyses. For confirmation of Zrts induction in iron-deprived condition by qRT-PCR, all steps were performed as described, except for Pb01 incubation in chemically defined MMcM medium (Restrepo and Jiménez, 1980) supplemented with 3.5 μM inorganic iron [Fe(NH4)2(SO4)2] or with 50 μM of the iron chelator bathophenanthroline disulfonic acid (BPS: Sigma-Aldrich, Germany) for 3 or 24 h before cell harvesting.
TTC Indicator Plates
To detect Pb01 and Pb18 surface reductase activity, a plate assay was used as described previously (Ogur et al., 1957), except that the 2,3,5-triphenyltetrazolium chloride (TTC)-containing overlay was poured on fresh colonies grown on SC plates, supplemented or not with 50 μM BPS (no iron condition), 30 μM inorganic iron [Fe(NH4)2(SO4)2], 30 μM hemoglobin, 120 μM hemin, 30 μg/ml ferritin, or 30 μM transferrin, independently. All host iron sources were purchased from Sigma-Aldrich, St. Louis, MO, USA.
59Fe Uptake Assays
After 24 h of Pb01 and Pb18 incubation in SC medium with no supplementation or with addition of 200 μM BPS or 10 μM FeCl3 at 36°C under rotation, the cells were harvested and washed with 1 mM EDTA in citrate uptake buffer (2% analar glucose, 0.1 MES buffer, and 20 mM Na-citrate, pH 6.0). After, the cells were washed twice with citrate uptake buffer and incubated for 15 min at 36°C under rotation. At this time, an aliquot of cells was collected and counted in a hemocytometer. Then, 20 mM of ascorbic acid was added (reductive-independent 59Fe uptake assay) or not (reductive-dependent 59Fe uptake assay); Cl4K2Pt (PtII compound) was added or not; and 0.2 μM 59Fe solution (Perkin-Elmer, Waltham, MA) was added independent of the condition. A triplicate of point 0 and 60 min were collected in a filter membrane (type A/C 25 mm glass fiber filter, Sigma-Aldrich, St. Louis, MO, USA) and the cells were washed with ice-cold 1X Quench buffer (37.5 mM succinic acid, 62.5 mM Tris, and 12.8 mM EDTA, pH 6.0). Then filters containing the cells were placed on the bottom of the test tubes and internalized 59Fe was measured using a Wallac γ counter (LKB Wallac CompuGamma). 59Fe values were normalized with the number of cells.
High-throughput mRNA Sequencing (RNA-seq)
Pb01 yeast cells were incubated in chemically defined MMcM medium supplemented with 3.5 μM Fe(NH4)2(SO4)2 or with 50 μM BPS (Sigma-Aldrich, Germany) for 24 h at 36°C on a rotary shaker at 150 rpm. After that, the cells for both conditions, in biological triplicates, were treated with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) to obtain RNA molecules. The cDNAs libraries were prepared from poly(A)-fragment selected mRNA and processed on the Illumina HiSeq2000 Sequencing System (http://www.illumina.com/). As a result, approximately 40 million of reads of 100 bp paired-end sequencing were obtained for each sample. The sequencing reads were mapped to reference the Pb01 genome (http://www.broadinstitute.org/annotation/genome/paracoccidioides_brasiliensis/ MultiHome.html) using the Bowtie 2 tool (Langmead et al., 2009). Briefly, each read was allowed to alignment in just one site of the genome and the reads were counted. The default parameters were used to perform the alignment. The number of mismatches allowed in seed alignment (−N) is 0, and the length of each seed (−L) is 20. Reads mapped to a gene region were counted to infer expression measurement. Differentially expressed genes were identified using Fisher Exact test in DEGseq package (Wang et al., 2010). A p-value lower than 0.001 and minimum fold change of 1.5 were used to filter the most relevant candidates. Meaning that transcripts with log2 (fold change) higher than 0.58 or less than −0.58 were selected and classified as up- and down-regulated transcripts, respectively. Transcript's identifications and annotations were determined from the Paracoccidioides genome database. The biological processes were obtained using the Pedant on MIPS (http://pedant.helmholtzmuenchen.de/pedant3htmlview/pedant3view?Method=analysis&Db=p3_r48325_Par_brasi_Pb01) which provides a tool to browse and search the Functional Categories (FunCat) of proteins.
The predicted proteins encoded by the transcripts identified through RNAseq strategy were also classified as copper−, iron−, and zinc-binding proteins from Paracoccidioides genome as previously described (Tristão et al., 2015). Briefly, metalloproteins were identified by using the RDGB tool (Andreini et al., 2011) with default options. In the RDGB strategy, the protein domains defined in the Pfam library are used to identify putative homologs in any desired genome or list of genomes. Copper−, iron−, and zinc-binding Pfam domains were initially identified in the sequence of copper−, iron−, and zinc-binding proteins of known 3D structures, which are available from the Protein Data Bank (PDB). When a particular metal is present within the 3D structure of the protein, this information can be readily extracted from the PDB database along with the pattern of amino acids that are involved in the interaction of the protein with the metal. In addition, other specific domains were also identified to predicted encoded transcripts using the Paracoccidioides genome database (http://www.broadinstitute.org/annotation/genome/paracoccidioides_brasiliensis/MultiHome.html).
Results
Characterization of Paracoccidioides spp. Ferric Reductases
The Paracoccidioides spp. Genome Database searching for proteins possessing all the three domains: FRD (PF01794), FAD-binding domain (PF08022), and ferric reductase NAD-binding domain (PF08030) resulted in seven Pb01 and seven Pb18 putative ferric reductases. Moreover, amino acid sequences of Paracoccidioides spp. Ggt1 homologs were collected in the same database. These sequences were used to construct a phylogenetic tree comprising ferric reductase sequences of different fungi: A. nidulans, A. fumigatus, H. capsulatum, C. immitis, C. posadasii, B. dermatitidis, U. maydis, C. neoformans, S. pombe, S. cerevisiae, and C. albicans (Supplementary Figure 1). It could be observed that all the Paracoccidioides spp. sequences are grouped with H. capsulatum, C. immitis, C. posadasii, and B. dermatitidis sequences. Moreover, BLAST analyses revealed the presence of three groups of proteins in this phylogenetic three: (1) transmembrane ferric reductases; (2) NADPH oxidases; and (3) γ-glutamyltransferases. The sequences identified as NADPH oxidases were excluded from posterior analyses, since they are out of the manuscript's scope.
All the Pb01 and Pb18 sequences classified as transmembrane ferric reductases contained characteristic features of ferric reductases, such as the FRD, NAD, and FAD-binding domains, a bis-heme motif and at least five transmembrane domains (Figure 1). The sequences classified as γ-glutamyltransferases were aligned and presented similarity, mainly in some regions of γ-glutamyltranspeptidase domain (PF01019) (Supplementary Figure 2), indicating that other fungi, such as A. fumigatus, A. nidulans, C. albicans, C. neoformans, S. cerevisiae, S. pombe, U. maydis, C. immitis, and C. posadasii, not tested yet for extracellular glutathione-dependent ferric reductase (GSH-FeR) activity could utilize this alternative iron reduction route.
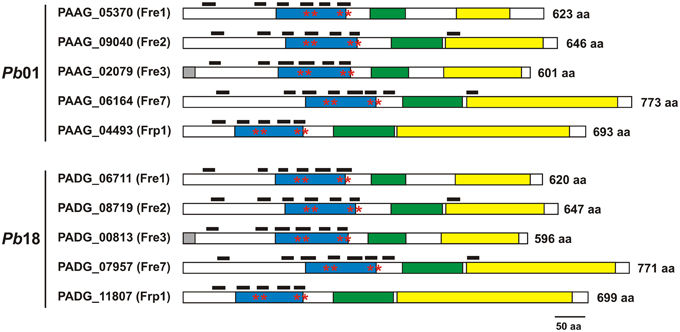
Figure 1. Paracoccidioides spp. ferric reductases present typical domains. Domains identified in Pb01 and Pb18 transmembrane ferric reductases were: ferric reductase domain (blue box), FAD-binding domain (green box), and NAD-binding domain (yellow box). Asterisks indicate the bis-hememotif comprising conserved histidine residues. Black boxes above each sequence indicate the lengh and position of transmembrane domains. The number of amino acids (aa) of each protein is indicated adjacent to the sequences.
Paracoccidioides spp. Ferric Reductase Regulation and Iron Uptake
In order to evaluate the Pb01 and Pb18 putative ferric reductase transcripts, the yeast cells were cultivated in presence of 10 μM inorganic iron or 200 μM BPS. After 2 h, it was observed no relevant regulation of all Pb01 transcripts analyzed in response to iron availability. Pb18 fre3, fre7, and ggt1 transcripts presented a slight regulation in response to iron availability. Under iron supplementation, Pb18 fre3 transcript level increased, whereas Pb18 fre7 and ggt1 transcripts level reduced, comparing to the SC medium without any supplementation. Under iron deprivation (presence of BPS), Pb18 fre7 transcript level reduced, whereas Pb18 ggt1 transcript level increased, comparing to the SC medium without any supplementation (Figure 2). Overall, there was no indication of a pattern of iron regulation of ferric reductase transcripts abundance in Paracoccidioides spp.
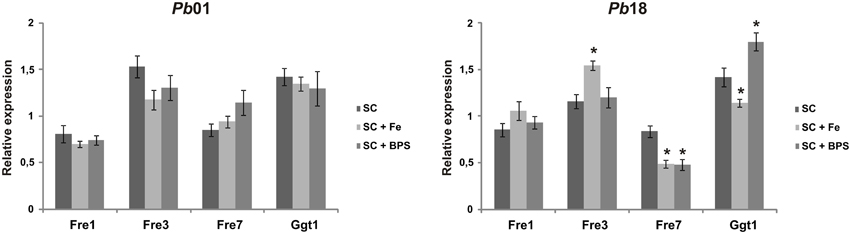
Figure 2. Expression profile of transcripts putatively encoding Paracoccidioides spp. ferric reductases. Pb01 and Pb18 yeast cells were recovered from SC medium, which was supplemented or not with 200 μM BPS or 10 μM FeCl3, after 2 h incubation. After RNA extraction and cDNA synthesis, Pb01 and Pb18 transcript levels were quantified using qRT-PCR. The expression values were calculated using alpha tubulin as the endogenous control. The data are expressed as the mean ± SD from triplicates. *Statistically significant data as determined by Student's t-test (p < 0.05).
To investigate if the RIA pathway could be functional when Paracoccidioides spp. grow in presence of different iron sources, inorganic iron, hemoglobin, hemin, ferritin, or transferrin were used. SC medium not supplemented with iron or chelated with BPS (no iron) were also used as controls. The Pb01 and Pb18 yeast cells growing in all conditions presented cell surface reductase activity, as evidenced by the red colony color in the presence of TTC (Figure 3). In addition, 59Fe uptake was quantified in presence or absence of the reducing agent ascorbic acid using Pb01 and Pb18 yeast cells grown in presence or absence of iron. The results showed that cells cultivated under iron deprivation (200 μM BPS) presented an increased 59Fe uptake rate, when compared to cells cultivated in the presence of 10 μM FeCl3 (Figure 4). When ascorbic acid was omitted, 59Fe uptake should be dependent on ferric reduction; only Pb18 cells cultivated under iron deprivation were able to internalize 59Fe added as ferric iron indicating that the reductase activity in this strain was coupled to iron uptake.
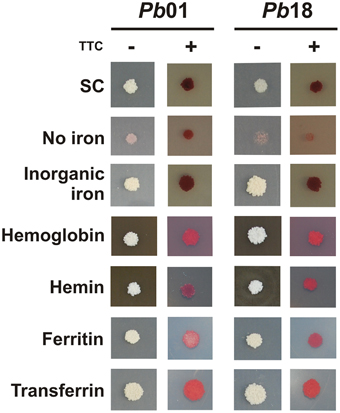
Figure 3. Activation of ferric reductase activity in presence of different iron sources by Paracoccidioides spp. Pb01 and Pb18 cell cultures were collected after 24 h in presence of 200 μM BPS, washed and 104 cells were spotted on SC medium plates, which were supplemented or not with 50 μM BPS, 30 μM inorganic iron, 30 μM hemoglobin, 120 μM hemin, 30 mg/ml ferritin, or 30 μM transferrin, independently. After 10 days of growth, TTC-containing agar solution was poured on the plates containing the colonies. The red colonies after TTC addition indicate a reduction of the colorless electron acceptor TTC to a red formazan precipitate.
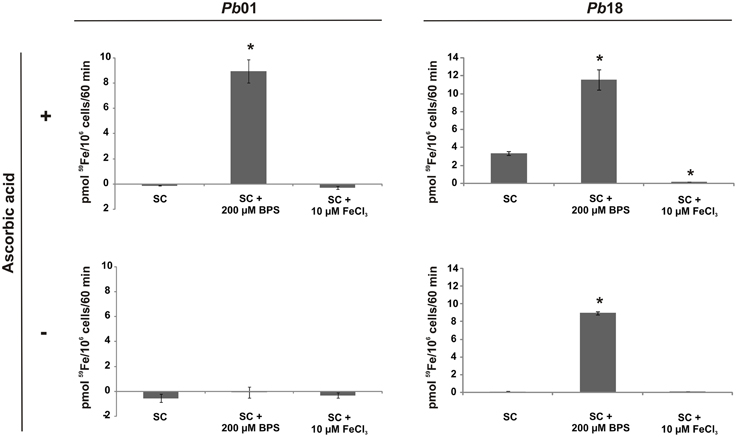
Figure 4. The iron uptake rate in Paracoccidioides spp. is dependent on the culture conditions and on the oxidation state of available iron. Pb01 and Pb18 yeast cells were cultivated on SC medium, which was supplemented or not with 200 μM BPS or 10 μM FeCl3, for 24 h. Then, the cells were collected, washed and incubated in presence of 59Fe with addition (+) or not (−) of ascorbic acid for 60 min. After this, the cells were collected and washed again and submitted to a γ counter for iron uptake rate establishment. The data is present as the mean ± SD from triplicates or quadruplicates. *Statistically significant data, as determined by Student's t-test (p < 0.05), relative to yeast cells cultivated on SC medium.
Since the ferric reductase activity can be subject to inhibition by Pt(II) (Eide et al., 1992), the Pb18 ferric reductase sensitivity to this element was tested using as an indicator the sensitivity of 59Fe uptake to this transition metal. Pb18 cells cultivated under iron deprivation exhibited a modest decrease in reductase-dependent 59Fe uptake in the presence of Pt(II) (Figure 5A); the limited uptake by Pb01 of 59Fe added as ferric iron was not strongly affected by this addition (Figure 5B).
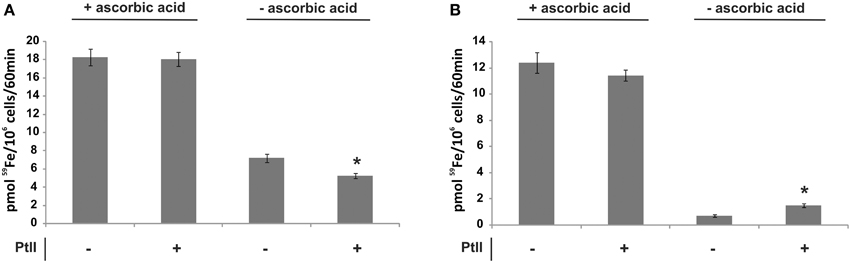
Figure 5. Effect of a platinum compound addition on the iron uptake rate of Paracoccidioides spp. Pb01 and Pb18 yeast cells were cultivated on SC medium supplemented or not with 200 μM BPS for 24 h. Then, the cells were collected, washed and incubated in presence of 59Fe with addition (+) or not (−) of ascorbic acid and/or a platinum compound (PtII) for 60 min. After this, the cells were collected and washed again and submitted to a γ counter for iron uptake rate establishment in Pb18 (A) and Pb01 (B). The data is present as the mean ± SD from triplicates or quadruplicates. *Statistically significant data, as determined by Student's t-test (p < 0.05), relative to yeast cells cultivated on abscence of a PtII compound.
The Multicopper Oxidases in Paracoccidioides spp.
To start an investigation about Paracoccidioides spp. ferroxidases, a searching for MCOs in Laccase Engineering Database (LccED: http://www.lcced.uni-stuttgart.de) was developed. It resulted in no Paracoccidioides spp. sequences classified in family E, corresponding to fungal ferroxidases. Paracoccidioides spp. MCOs are classified as basidiomycete laccases (family A), ascomycete laccases (family B), and fungal pigment MCOs (family D) (http://www.lcced.uni-stuttgart.de/cgi-bin/LccED1.2/index.pl?page=org&id=35). However, the sequences of all the Pb01 and Pb18 MCOs deposited in Paracoccidioides Genome Database were collected and aligned with S. cerevisiae Fet3 (Supplementary Figure 3) to identify both potential Cu-binding residues and the carboxylate side chains that support ferrous oxidase activity in fungal ferroxidases (Kosman, 2010). This examination indicated that only PAAG_00163 do not clearly displayed all four Cu-binding motifs and therefore cannot support the ferroxidase activity typical in the RIA pathway.
The ferroxidase activity in fungal MCOs is due to two acidic residues; in Fet3 these are E185 and D407. The global alignments (Supplementary Figure 3) provide insight into the presence of homologous residues in two Paracoccidioides spp. proteins, PAAG_06004 and PADG_05994, which are likely ferroxidases. A clear difference between Fet3 and Paracoccidioides spp. proteins is that the former presents a C-terminal transmembrane domain, whereas PAAG_06004 and PADG_05994 present N-terminal transmembrane domain (Supplementary Figure 3). Due to high similarity in ferroxidase motifs and in residues involved in Fe2+ binding between the PAAG_06004 and PADG_05994, a structural model of PAAG_06004 was constructed based on the structure of Fet3 (PBD 1ZPU). Despite the fact that the two proteins have only 22% identity, a satisfactory model was generated that did, however, contain a relatively large (unfavorable) Z-score for torsion angle energy. However, the model faithfully reproduced the conformation of the protein's four predicted Cu-coordination sites and indicated that in PAAG_06004 E280 and D536 could be part of a ferroxidase site in this protein (Figure 6). Isolation and characterization of this protein is necessary to test this possible ferroxidase activity.
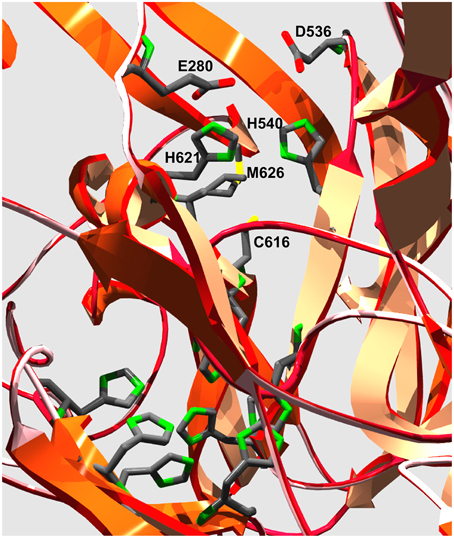
Figure 6. Model of the putative Cu-coordination sites in Pb01 PAAG_06004. The ribbon diagram illustrates the protein folds that contain the 11 side chains that make up the coordination sites for the 4 Cu-atoms found in fungal MCOs. The six His side chains in the lower left quadrant are ligands to the T2 and T3 Cu atoms that make up the trinuclear cluster to which O2 binds and is reduced to 2H2O. The grouping of H540, C616, H621, and M626 are ligands to the T1 Cu that is the electron acceptor from the reducing substrate. Ferroxidases contain two acidic side chains outer-sphere to this ligand grouping that are in H-bond contact with each of the His side chains. The unbiased computer model of PAAG_06004 has E280 and D536 in this conformation relative to the Fet3 T1 Cu His ligands H621 and H540, respectively, suggesting that this protein could exhibit ferroxidase activity.
The Iron Permeases in Paracoccidioides sp.
A whole transcriptome sequencing strategy was performed as previously described (Lima et al., 2014). The number of the reads counted for each transcript in iron replete and iron-deprived conditions was represented by scattered dots (Supplementary Figure 4). The transcripts were represented by dots, which could present a different number of reads in each condition (Supplementary Figure 4A). We also applied a statistical test to identify differentially expressed transcripts, represented by red dots (Supplementary Figure 4B). A total of 549 transcripts were statistically significant (Supplementary Figure 4B) but a cut-off of 1.5-fold change (Amich et al., 2013) generated 30 up- and 44 down-regulated transcripts (Supplementary Tables 1, 2).
Previous analysis has indicated that Paracoccidioides spp. does not express a Ftr1 homolog and thus the fungus could use zinc permeases to transport iron (Bailão et al., 2007; Silva et al., 2011). The RNAseq analysis revealed that zinc permeases transcripts (zrt1 and zrt2) are up-regulated under iron deprivation in Pb01 yeast cells, consistent with this hypothesis (Supplementary Table 1 and Figure 7). The results also revealed that a siderophore uptake encoding transcript (mirB) and transcripts encoding enzymes, such as carbapenemantibiotics biosynthesis protein (carD) and NADP-specific glutamate dehydrogenase (gdh), involved in ornithine biosynthesis, a hydroxamate precursor, are up-regulated under iron deprivation (Supplementary Table 1 and Figure 7). This fact points to an increase in siderophore synthesis and uptake during iron deprivation. Down-regulated transcripts are related to amino acid and lipid metabolism, which could generate acetyl-CoA, precursor for several metabolic pathways, and with heme biosynthesis, an iron-dependent molecule. The acetyl-CoA decreased production in association with the down-regulation of the electron transport chain could indicate an impaired aerobic metabolism (Figure 7). In fact, iron-sulfur proteins, such as succinate dehydrogenase iron-sulfur subunit (sdh) and cytosolic Fe-S cluster assembling factor NBP35, that also need iron as a cofactor, were down-regulated (Supplementary Table 2 and Figure 7), indicating that the fungus faces an iron-deprived condition.
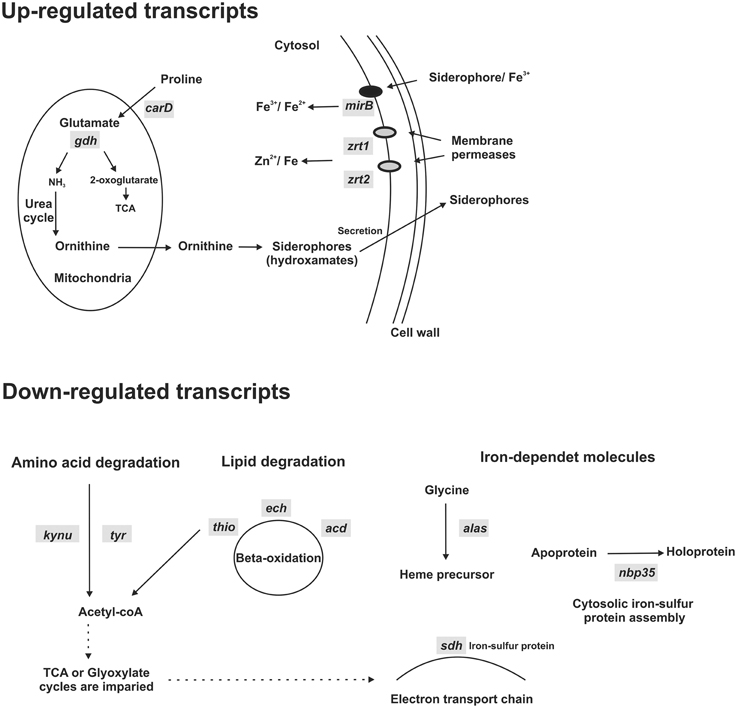
Figure 7. Schematic representation of up and down-regulated transcripts under iron deprivation. Pb01 yeast cells were cultivated on MMcM medium supplemented with 3.5 μM inorganic iron or with 50 μM BPS (iron deprivation) for 24 h. Then, a high-throughput mRNA sequencing (RNA-seq) strategy was used to compare transcripts expressed in iron deprivation and in presence of iron. Some transcripts obtained using this strategy was selected to construct this scheme. In iron deprivation condition, transcripts involved with siderophore synthesis (carD and gdh) and uptake (mirB) and with iron uptake using Fe/Zn permeases (zrt1 and zrt2) were up-regulated. On the other hand, transcripts involved with amino acid (kynu and tyr) and lipid degradation (thio, echand acd) and with synthesis of putative iron-dependent molecules (alas, sdh, and nbp35), such as heme and iron-sulfur proteins, were down-regulated. carD, carbapenemantibiotics biosynthesis protein; gdh, NADP-specific glutamate dehydrogenase; mirB, siderophore iron transporter; zrt1 and zrt2, zinc regulated transporters; kynu, kynureninase; tyr, tyrosinase central domain-containing protein; thio, 3-ketoacyl-CoA thiolase; ech, delta(3,5)-delta(2,4)-dienoyl-CoA isomerase; acd, acyl-CoA dehydrogenase family protein; alas, 5-aminolevulinate synthase; sdh, succinate dehydrogenase iron-sulfur subunit; nbp35, cytosolic Fe-S cluster assembling factor.
When Pb01 zrt1 and zrt2 transcripts were analyzed using qRT-PCR, similar results were obtained. After 24 h of iron deprivation, zrt1 and zrt2 expression increased (Figure 8), indicating that these genes could play a role in iron acquisition.
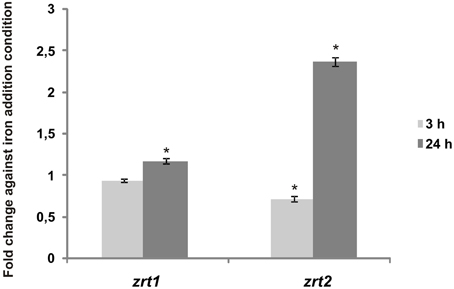
Figure 8. Transcripts putatively encoding Paracoccidioides sp. Fe/Zn permeases are iron-regulated. Pb01 yeast cells were recovered from MMcM medium, which was supplemented with 50 μM BPS or 3.5 μM inorganic iron for 3 or 24 h. After RNA extraction and cDNA synthesis, levels of Pb01 zrt1 and zrt2 were quantified by qRT-PCR. The expression values were calculated using alpha tubulin as the endogenous control. The values that were plotted on the bar graph were normalized against the expression data that were obtained from the iron addition condition (fold change). The data are expressed as the mean ± SD from triplicates. *Statistically significant data as determined by Student's t-test (p < 0.05).
The Iron Uptake Strategies in Paracoccidioides spp.
Based on previously published data (Bailão et al., 2007, 2014; Parente et al., 2011; Silva-Bailão et al., 2014) and those obtained in this study, an updated model regarding the iron uptake strategies used by Paracoccidioides spp. was proposed (Figure 9). Under iron deprivation, the fungus may use both ferric/ferrous iron uptake and siderophore uptake pathways, since an increase in zrts transcripts and in those involved with siderophore production and uptake was observed in this study and/or in previously published data (Parente et al., 2011; Silva-Bailão et al., 2014). Hemoglobin uptake through the hemoglobin receptors may be used preferentially in the presence of this host molecule, since Pb01 putative hemoglobin receptors present reduced transcription under iron depletion (Bailão et al., 2014). In the same way, transmembrane ferric reductases seem to be up-regulated in presence of host iron sources, such as transferrin and hemoglobin, since Pb01 fre3 transcript (named previously as fre2) presented an increased expression during Pb01 yeast cells incubation with human plasma (Bailão et al., 2007) and human blood (Bailão et al., 2006).
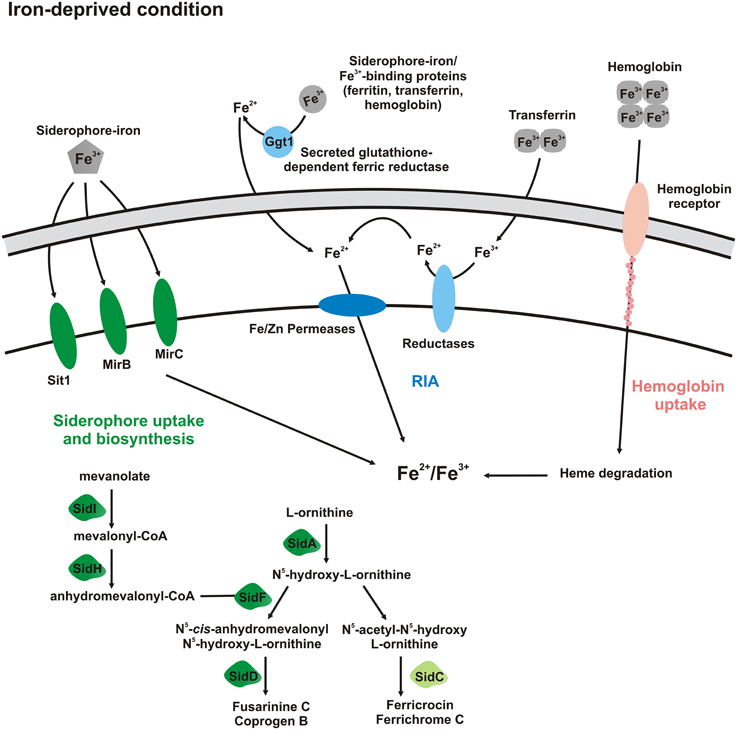
Figure 9. Schematic representation of Paracoccidioides spp. activated pathways in iron-deprived condition. Forms with dark colors indicate proteins whose transcription is up-regulated and that ones with light colors do not present transcription regulation. In that way, in iron-deprived condition, siderophore uptake, and biosynthesis of extracellular siderophores (coprogen B), as well as, Fe/Zn permeases (Zrts) are up-regulated. It seems that transmembrane ferric reductases and hemoglobin receptors are up-regulated mainly in presence of host iron sources, such as transferrin and hemoglobin, respectively.
Discussion
As described previously, Paracoccidioides spp. presents multiple ferric reductases (Silva et al., 2011). These proteins are very similar to homologs found in H. capsulatum, C. immitis, C. posadasii, and B. dermatitidis, as expected, since all the organisms are classified in the Onygenales order (Sharpton et al., 2009). There are five putative transmembrane ferric reductases either for Pb01 or for Pb18. Moreover these fungi present one glutathione-dependent ferric reductase homolog, whose activity has been described (Zarnowski and Woods, 2005). The Paracoccidioides spp. transmembrane ferric reductases present domains described for homologs from other pathogenic fungi, such as FRD, NAD and FAD-binding domains, bis-heme motif and transmembrane domains (Almeida et al., 2009; Saikia et al., 2014). The expression of these ferric reductases seems not to be iron-dependent, since the differences observed between the iron-replete and the iron-deprived conditions were subtle. Similar results were observed for C. neoformans, in which subtle differences in ferric reductases expression were observed in presence or absence of iron (Saikia et al., 2014). However, an increase in Paracoccidioides spp. ferric reductase expression has been observed in the presence of human blood and plasma (Bailão et al., 2006, 2007). This apparent inconsistency could be explained if the ferric reductase expression would be activated in the presence of human iron sources, such as transferrin, an iron source that could be used by Paracoccidioides spp. (Bailão et al., 2014).
59Fe uptake assays demonstrated that Pb18 reduce and uptake iron in scarcity of this metal. On the other hand, Pb01 express an apparently inefficient RIA system, since 59Fe uptake in the absence of ascorbic acid was low in any culture condition used in this work although TTC assay indicates this strain does exhibit cell surface ferric reductase activity. It has been proposed that A. nidulans also lack an efficient RIA pathway, because a siderophore-deficient strain is not able to utilize Fe3+ (Eisendle et al., 2003). Perhaps the main route to acquire iron in these fungi is the siderophore uptake, which has been demonstrated to be efficient either in A. nidulans (Eisendle et al., 2003) or in Paracoccidioides spp. (Silva-Bailão et al., 2014).
Paracoccidioides spp. MCOs (PAAG_06004 and PADG_05994), with a low Fet3 identity, were identified in Paracoccidioides spp. genome database. To confirm if these proteins could act as ferroxidases and if the MCO domain face or not the cytoplasm, the proteins should be isolated, characterized, and cellular localized, that is our focus in future studies. What we hypothesize at this moment is that an MCO does not play a significant role in RIA pathway in Paracoccidioides spp., since in previous work (Bailão et al., 2006, 2007, 2014; Costa et al., 2007; Parente et al., 2011), as well as in this work, ferroxidase regulation in low-iron conditions was not observed. Moreover, this pathway appears to be unusual, since it does not include an Ftr1 homolog, common to several fungi (Stearman et al., 1996; Ziegler et al., 2011). Similarly, a homology search in A. nidulans genome database failed to identify Fet3 and Ftr1 homologs (Eisendle et al., 2003).
We note, however, that Paracoccidioides spp. express Zrts homologs that are induced in iron- (this work) or zinc-deprivation (Parente et al., 2013). In plants and mammals, it has been demonstrated that ZIP (Zrt/Irt-like Proteins) family of membrane transporters is able to transport not only zinc, but also iron (Zhao et al., 2010; Milner et al., 2013). Additionally, it has been observed that Paracoccidioides spp. Zrts present more than one possible metal-binding domain (HXHXHXH) (Zhao and Eide, 1996) in the sequence (Silva et al., 2011), suggesting that these domains could be responsible for zinc and iron binding. These facts corroborate the hypothesis that Paracoccidioides Zrts are able to transport both iron and zinc. Moreover, it has been demonstrated that Zrt expression increases in yeast cells recovered from liver of infected mice and in presence of human plasma (Bailão et al., 2006, 2007). These data suggest that the permease is regulated in iron-limited conditions either in vitro or in vivo and in presence of human iron sources, such as transferrin. Maybe the iron-regulation in Paracoccidioides spp. impacts most strongly on the permeases and not on the ferric reductases.
A whole transcriptome sequencing strategy also revealed that both a siderophore transporter and the ornithine, a hydroxamate precursor (Eisendle et al., 2003), biosynthesis are up-regulated, indicating that Paracoccidioides spp. utilize the siderophore uptake pathway during iron scarcity, as previously demonstrated (Silva-Bailão et al., 2014). At the same time, expression of Zrts is also up-regulated, indicating that the fungus use more than one iron uptake strategy to acquire this metal when in iron scarcity. On the other hand, aerobic respiration and iron-dependent molecule synthesis pathways seems to be down-regulated, such as the synthesis of iron-sulfur proteins and heme precursors. It has been observed that tricarboxylic acid cycle and electron transport chain proteins decreased in abundance under iron limiting conditions (Parente et al., 2011). It could occur because iron is used as prosthetic group in a plenty of energy metabolism pathways (Kosman, 2013).
The RIA pathway in Paracoccidioides spp. appears constitutive in that ferric reductase expression is relatively insensitive to type or concentration of iron source. In contrast, in iron-deprived condition (Figure 9), transcripts related to siderophore uptake and biosynthesis of extracellular siderophores (coprogen B) are up-regulated as observed in this work and in others previously published (Parente et al., 2011; Silva-Bailão et al., 2014). Transcripts related to iron and zinc permeases are also up-regulated in Paracoccidioides spp. This indicates that the fungus can use both RIA and siderophore uptake pathways in iron limited condition. Paracoccidioides spp. also may use hemoglobin receptors to acquire this host iron source; those receptors are activated preferentially in presence of hemoglobin instead of iron deprivation (Bailão et al., 2014). In addition, in presence of transferrin (abundant in human plasma) and hemoglobin (abundant in human blood), the transcription of ferric reductases is up-regulated (Bailão et al., 2006, 2007). These data indicate that both RIA and siderophore uptake pathways could function under iron deprivation.
The study of iron metabolism is important to reveal novel molecules that contribute to iron uptake, an essential micronutrient for almost all organisms (Johnson, 2008). These proteins could function as accessible targets for drugs to block iron uptake, what could impair the pathogen's life. Moreover, iron permeases might also be exploited as vaccine targets or as route for antifungal internalization (Kronstad et al., 2013). Then, Paracoccidioides spp. proteins involved with iron uptake could be used as vaccine or drug targets or as drug internalization route to inhibit the proliferation of the pathogen.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the financial support from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Ensino Superior (CAPES), Financiadora de Estudos e Projetos (FINEP) and Fundação de Apoio à Pesquisa do Estado de Goiás (FAPEG). EB, PL, and MS received scholarships from CAPES.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.00821
References
Almeida, R. S., Wilson, D., and Hube, B. (2009). Candida albicans iron acquisition within the host. FEMS Yeast Res. 9, 1000–1012. doi: 10.1111/j.1567-1364.2009.00570.x
Amich, J., Schafferer, L., Haas, H., and Krappmann, S. (2013). Regulation of sulphur assimilation is essential for virulence and affects iron homeostasis of the human-pathogenic mould Aspergillus fumigatus. PLoS Pathog. 9:e1003573. doi: 10.1371/journal.ppat.1003573
Andreini, C., Bertini, I., Cavallaro, G., Decaria, L., and Rosato, A. (2011). A simple protocol for the comparative analysis of the structure and occurrence of biochemical pathways across superkingdoms. J. Chem. Inf. Model 51, 730–738. doi: 10.1021/ci100392q
Baek, Y. U., Li, M., and Davis, D. A. (2008). Candida albicans ferric reductases are differentially regulated in response to distinct forms of iron limitation by the Rim101 and CBF transcription factors. Eukaryot. Cell 7, 1168–1179. doi: 10.1128/EC.00108-08
Bailão, A. M., Nogueira, S. V., Rondon Caixeta Bonfim, S. M., de Castro, K. P., de Fátima da Silva, J., Mendes Giannini, M. J., et al. (2012b). Comparative transcriptome analysis of Paracoccidioides brasiliensis during in vitro adhesion to type I collagen and fibronectin: identification of potential adhesins. Res. Microbiol. 163, 182–191. doi: 10.1016/j.resmic.2012.01.004
Bailão, A. M., Schrank, A., Borges, C. L., Dutra, V., Molinari-Madlum, E. E. W. I., Felipe, M. S. S., et al. (2006). Differential gene expression by Paracoccidioides brasiliensis in host interaction conditions: representational difference analysis identifies candidate genes associated with fungal pathogenesis. Microbes. Infect. 8, 2686–2697. doi: 10.1016/j.micinf.2006.07.019
Bailão, A. M., Shrank, A., Borges, C. L., Parente, J. A., Dutra, V., Felipe, M. S., et al. (2007). The transcriptional profile of Paracoccidioides brasiliensis yeast cells is influenced by human plasma. FEMS Immunol. Med. Microbiol. 51, 43–57. doi: 10.1016/j.micinf.2006.07.019
Bailão, E. F. L. C., Parente, A. F., Parente, J. A., Silva-Bailão, M. G., Castro, K. P. D., Kmetzsch, L., et al. (2012a). Metal acquisition and homeostasis in fungi. Curr. Fungal. Infect. Rep. 6, 257–266. doi: 10.1007/s12281-012-0108-8
Bailão, E. F., Parente, J. A., Pigosso, L. L., de Castro, K. P., Fonseca, F. L., Silva-Bailão, M. G., et al. (2014). Hemoglobin uptake by Paracoccidioides spp. is receptor-mediated. PLoS Negl. Trop Dis. 8:e2856. doi: 10.1371/journal.pntd.0002856
Bocca, A. L., Amaral, A. C., Teixeira, M. M., Sato, P. K., Shikanai-Yasuda, M. A., and Soares Felipe, M. S. (2013). Paracoccidioidomycosis: eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 8, 1177–1191. doi: 10.2217/fmb.13.68
Bookout, A. L., Cummins, C. L., Mangelsdorf, D. J., Pesola, J. M., and Kramer, M. F. (2006). High-throughput real-time quantitative reverse transcription PCR. Curr. Protoc. Mol. Biol. Chapter 15, Unit 15 18. doi: 10.1002/0471142727.mb1508s73
Carrero, L. L., Niño-Vega, G., Teixeira, M. M., Carvalho, M. J., Soares, C. M., Pereira, M., et al. (2008). New Paracoccidioides brasiliensis isolate reveals unexpected genomic variability in this human pathogen. Fungal Genet. Biol. 45, 605–612. doi: 10.1016/j.fgb.2008.02.002
Costa, M., Borges, C. L., Bailão, A. M., Meirelles, G. V., Mendonca, Y. A., Dantas, S. F., et al. (2007). Transcriptome profiling of Paracoccidioides brasiliensis yeast-phase cells recovered from infected mice brings new insights into fungal response upon host interaction. Microbiology 153, 4194–4207. doi: 10.1099/mic.0.2007/009332-0
Eide, D., Davis-Kaplan, S., Jordan, I., Sipe, D., and Kaplan, J. (1992). Regulation of iron uptake in Saccharomyces cerevisiae. The ferrireductase and Fe(II) transporter are regulated independently. J. Biol. Chem. 267, 20774–20781.
Eisendle, M., Oberegger, H., Zadra, I., and Haas, H. (2003). The siderophore system is essential for viability of Aspergillus nidulans: functional analysis of two genes encoding l-ornithine N 5-monooxygenase (sidA) and a non-ribosomal peptide synthetase (sidC). Mol. Microbiol. 49, 359–375. doi: 10.1046/j.1365-2958.2003.03586.x
Hu, G., Chen, S. H., Qiu, J., Bennett, J. E., Myers, T. G., and Williamson, P. R. (2014). Microevolution during serial mouse passage demonstrates FRE3 as a virulence adaptation gene in Cryptococcus neoformans. MBio 5, e00941–e00914. doi: 10.1128/mBio.00941-14
Johnson, L. (2008). Iron and siderophores in fungal-host interactions. Mycol. Res. 112, 170–183. doi: 10.1016/j.mycres.2007.11.012
Knight, S. A., Vilaire, G., Lesuisse, E., and Dancis, A. (2005). Iron acquisition from transferrin by Candida albicans depends on the reductive pathway. Infect. Immun. 73, 5482–5492. doi: 10.1128/IAI.73.9.5482-5492.2005
Kosman, D. J. (2003). Molecular mechanisms of iron uptake in fungi. Mol. Microbiol. 47, 1185–1197. doi: 10.1046/j.1365-2958.2003.03368.x
Kosman, D. J. (2010). Multicopper oxidases: a workshop on copper coordination chemistry, electron transfer, and metallophysiology. J. Biol. Inorg. Chem. 15, 15–28. doi: 10.1007/s00775-009-0590-9
Kosman, D. J. (2013). Iron metabolism in aerobes: managing ferric iron hydrolysis and ferrous iron autoxidation. Coord. Chem. Rev. 257, 210–217. doi: 10.1016/j.ccr.2012.06.030
Kronstad, J. W., Cadieux, B., and Jung, W. H. (2013). Pathogenic yeasts deploy cell surface receptors to acquire iron in vertebrate hosts. PLoS Pathog. 9:e1003498. doi: 10.1371/journal.ppat.1003498
Kwok, E. Y., Severance, S., and Kosman, D. J. (2006). Evidence for iron channeling in the Fet3p-Ftr1p high-affinity iron uptake complex in the yeast plasma membrane. Biochemistry 45, 6317–6327. doi: 10.1021/bi052173c
Langmead, B., Trapnell, C., Pop, M., and Salzberg, S. L. (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10:R25. doi: 10.1186/gb-2009-10-3-r25
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam, H., et al. (2007). Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404
Lima, P. D. S., Casaletti, L., Bailão, A. M., de Vasconcelos, A. T., Fernandes Gda, R., and Soares, C. M. (2014). Transcriptional and proteomic responses to carbon starvation in Paracoccidioides. PLoS Negl. Trop Dis. 8:e2855. doi: 10.1371/journal.pntd.0002855
Martinez, R. (2010). Paracoccidioidomycosis: the dimension of the problem of a neglected disease. Rev. Soc. Bras. Med. Trop. 43, 480. doi: 10.1590/S0037-86822010000400034
Milner, M. J., Seamon, J., Craft, E., and Kochian, L. V. (2013). Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 64, 369–381. doi: 10.1093/jxb/ers315
Ogur, M., St. John, R., and Nagai, S. (1957). Tetrazolium overlay technique for population studies of respiration deficiency in yeast. Science 125, 928–929.
Page, R. D. (1996). TreeView: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 12, 357–358.
Parente, A. F., Bailão, A. M., Borges, C. L., Parente, J. A., Magalhães, A. D., Ricart, C. A., et al. (2011). Proteomic analysis reveals that iron availability alters the metabolic status of the pathogenic fungus Paracoccidioides brasiliensis. PLoS ONE 6:e22810. doi: 10.1371/journal.pone.0022810
Parente, A. F., de Rezende, T. C., de Castro, K. P., Bailão, A. M., Parente, J. A., Borges, C. L., et al. (2013). A proteomic view of the response of Paracoccidioides yeast cells to zinc deprivation. Fungal Biol. 117, 399–410. doi: 10.1016/j.funbio.2013.04.004
Restrepo, A., and Jiménez, B. E. (1980). Growth of Paracoccidioides brasiliensis yeast phase in a chemically defined culture medium. J. Clin. Microbiol. 12, 279–281.
Saikia, S., Oliveira, D., Hu, G., and Kronstad, J. (2014). Role of ferric reductases in iron acquisition and virulence in the fungal pathogen Cryptococcus neoformans. Infect. Immun. 82, 839–850. doi: 10.1128/IAI.01357-13
Sali, A., Potterton, L., Yuan, F., Van Vlijmen, H., and Karplus, M. (1995). Evaluation of comparative protein modeling by MODELLER. Proteins 23, 318–326. doi: 10.1002/prot.340230306
San-Blas, G., Niño-Vega, G., and Iturriaga, T. (2002). Paracoccidioides brasiliensis and paracoccidioidomycosis: molecular approaches to morphogenesis, diagnosis, epidemiology, taxonomy and genetics. Med. Mycol. 40, 225–242. doi: 10.1080/mmy.40.3.225.242
Sharpton, T. J., Stajich, J. E., Rounsley, S. D., Gardner, M. J., Wortman, J. R., Jordar, V. S., et al. (2009). Comparative genomic analyses of the human fungal pathogens Coccidioides and their relatives. Genome Res. 19, 1722–1731. doi: 10.1101/gr.087551.108
Silva, M. G., Schrank, A., Bailão, E. F. L. C., Bailão, A. M., Borges, C. L., Staats, C. C., et al. (2011). The homeostasis of iron, copper, and zinc in Paracoccidioides brasiliensis, Cryptococcus neoformans var. grubii, and Cryptococcus gattii: a comparative analysis. Front. Microbiol. 2:49. doi: 10.3389/fmicb.2011.00049
Silva-Bailão, M. G., Bailão, E. F., Lechner, B. E., Gauthier, G. M., Lindner, H., Bailão, A. M., et al. (2014). Hydroxamate production as a high affinity iron acquisition mechanism in Paracoccidioides spp. PLoS ONE 9:e105805. doi: 10.1371/journal.pone.0105805
Sirim, D., Wagner, F., Wang, L., Schmid, R. D., and Pleiss, J. (2011). The Laccase Engineering Database: a classification and analysis system for laccases and related multicopper oxidases. Database (Oxford) 2011:bar006. doi: 10.1093/database/bar006
Stearman, R., Yuan, D. S., Yamaguchi-Iwai, Y., Klausner, R. D., and Dancis, A. (1996). A permease-oxidase complex involved in high-affinity iron uptake in yeast. Science 271, 1552–1557.
Teixeira, M. M., Theodoro, R. C., Oliveira, F. F., Machado, G. C., Hahn, R. C., Bagagli, E., et al. (2014). Paracoccidioides lutzii sp. nov.: biological and clinical implications. Med. Mycol. 52, 19–28. doi: 10.3109/13693786.2013.794311
Tristão, G. B., Assunção Ldo, P., Dos Santos, L. P., Borges, C. L., Silva-Bailão, M. G., Soares, C. M., et al. (2015). Predicting copper-, iron-, and zinc-binding proteins in pathogenic species of the Paracoccidioides genus. Front. Microbiol. 5:761. doi: 10.3389/fmicb.2014.00761
Wang, L., Feng, Z., Wang, X., Wang, X., and Zhang, X. (2010). DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 26, 136–138. doi: 10.1093/bioinformatics/btp612
Xu, N., Qian, K., Dong, Y., Chen, Y., Yu, Q., Zhang, B., et al. (2014). Novel role of the Candida albicans ferric reductase gene CFL1 in iron acquisition, oxidative stress tolerance, morphogenesis and virulence. Res. Microbiol. 165, 252–261. doi: 10.1016/j.resmic.2014.03.001
Yun, C. W., Bauler, M., Moore, R. E., Klebba, P. E., and Philpott, C. C. (2001). The role of the FRE family of plasma membrane reductases in the uptake of siderophore-iron in Saccharomyces cerevisiae. J. Biol. Chem. 276, 10218–10223. doi: 10.1074/jbc.M010065200
Zarnowski, R., and Woods, J. P. (2005). Glutathione-dependent extracellular ferric reductase activities in dimorphic zoopathogenic fungi. Microbiology 151, 2233–2240. doi: 10.1099/mic.0.27918-0
Zhao, H., and Eide, D. (1996). The yeast ZRT1 gene encodes the zinc transporter protein of a high-affinity uptake system induced by zinc limitation. Proc. Natl. Acad. Sci. U.S.A. 93, 2454–2458.
Zhao, N., Gao, J., Enns, C. A., and Knutson, M. D. (2010). ZRT/IRT-like protein 14 (ZIP14) promotes the cellular assimilation of iron from transferrin. J. Biol. Chem. 285, 32141–32150. doi: 10.1074/jbc.M110.143248
Keywords: iron reduction, iron uptake, ferric reductase, zinc-regulated transporter, multicopper oxidases, paracoccidioidomycosis
Citation: Bailão EFLC, Lima PdS, Silva-Bailão MG, Bailão AM, Fernandes GdR, Kosman DJ and Soares CMdA (2015) Paracoccidioides spp. ferrous and ferric iron assimilation pathways. Front. Microbiol. 6:821. doi: 10.3389/fmicb.2015.00821
Received: 24 April 2015; Accepted: 27 July 2015;
Published: 12 August 2015.
Edited by:
Helio K. Takahashi, Universidade Federal de São Paulo, BrazilReviewed by:
Marcelo Tolmasky, California State University Fullerton, USAMarcos Sergio Toledo, Universidade Federal de São Paulo, Brazil
Copyright © 2015 Bailão, Lima, Silva-Bailão, Bailão, Fernandes, Kosman and Soares. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Célia Maria de Almeida Soares, Laboratório de Biologia Molecular, Instituto de Ciências Biológicas II, Universidade Federal de Goiás, Prédio ICB II, Sala 206, Câmpus Samambaia, 74690-900 Goiânia, Brazil, cmasoares@gmail.com
†Present Address: Elisa Flávia L. C. Bailão, Universidade Estadual de Goiás, Anápolis, Brazil; Mirelle G. Silva-Bailão, Universidade Federal de Goiás, Unidade Acadêmica Ciências da Saúde, Jataí, Brazil; Gabriel da Rocha Fernandes, Centro de Pesquisas Rene Rachou - Fiocruz, Grupo de Genômica e Biologia Computacional, Belo Horizonte, Brazil