 Jasmin K. Kroeger1,2
Jasmin K. Kroeger1,2 Karl Hassan3
Karl Hassan3 Aniko Vörös1
Aniko Vörös1 Roger Simm1†Massoud Saidijam4†
Roger Simm1†Massoud Saidijam4† Kim E. Bettaney4Andreas Bechthold2
Kim E. Bettaney4Andreas Bechthold2 Ian T. Paulsen3Peter J. F. Henderson4
Ian T. Paulsen3Peter J. F. Henderson4 Anne-Brit Kolstø1*
Anne-Brit Kolstø1*- 1Laboratory for Microbial Dynamics, Department of Pharmaceutical Biosciences, School of Pharmacy, University of Oslo, Oslo, Norway
- 2Institut für Pharmazeutische Biologie und Biotechnologie, Albert-Ludwigs Universität, Freiburg, Germany
- 3Department of Chemistry and Biomolecular Sciences, Macquarie University, Sydney, NSW, Australia
- 4School of BioMedical Sciences and Astbury Centre for Structural Molecular Biology, University of Leeds, Leeds, UK
Phylogenetic classification divides the major facilitator superfamily (MFS) into 82 families, including 25 families that are comprised of transporters with no characterized functions. This study describes functional data for BC3310 from Bacillus cereus ATCC 14579, a member of the “unknown major facilitator family-2” (UMF-2). BC3310 was shown to be a multidrug efflux pump conferring resistance to ethidium bromide, SDS and silver nitrate when heterologously expressed in Escherichia coli DH5α ΔacrAB. A conserved aspartate residue (D105) in putative transmembrane helix 4 was identified, which was essential for the energy dependent ethidium bromide efflux by BC3310. Transport proteins of the MFS comprise specific sequence motifs. Sequence analysis of UMF-2 proteins revealed that they carry a variant of the MFS motif A, which may be used as a marker to distinguish easily between this family and other MFS proteins. Genes orthologous to bc3310 are highly conserved within the B. cereus group of organisms and thus belong to the core genome, suggesting an important conserved functional role in the normal physiology of these bacteria.
Introduction
Bacillus cereus sensu stricto (B. cereus) is a Gram-positive, endospore forming organism known to cause foodborne illness in humans. It is a member of the B. cereus group of bacteria (Bacillus cereus sensu lato) that, in addition to B. cereus encompasses the species B. anthracis, B. thuringiensis, B. mycoides, B. pseudomycoides, B. weihenstephanensis, and B. cytotoxicus (Kolsto et al., 2009; Guinebretiere et al., 2013). The B. cereus group members are genetically closely related with high level of syntheny (conserved gene order). The high similarity results in an intertwinement of the B. cereus, B. thuringiensis, and B. weihenstephanensis branches in the phylogenetic tree (Ash et al., 1991). However, the B. cereus group organisms exhibit different phenotypes, inhabit diverse ecological niches and are pathogenic against different hosts. The three species B. mycoides, B. pseudomycoides, and B. weihenstephanansis are regarded as non-pathogenic. B. anthracis is the causative agent of anthrax in humans and animals (Mock and Fouet, 2001). B. thuringiensis is an insect pathogen that is commercially used as a biopesticide (Melo et al., 2014). B. cytotoxicus causes enteritis in humans and is thermotolerant and highly cytotoxic (Guinebretiere et al., 2013). In the natural environment B. cereus is found as a saprophyte in soil, associated with the rizosphere of plants and in the gut of invertebrates (Jensen et al., 2003; Berg et al., 2005). Even though B. cereus is most frequently associated with food-borne enteric infections in humans, it is able to cause other local or systemic infections such as endophthalmitis, cutaneous infections, endocarditis, central nervous system infection, or bacteremia (Steen et al., 1992; Callegan et al., 1999; Centers for Disease Control and Prevention, 2005; Callegan et al., 2006; Martinez et al., 2007; Kim et al., 2010; Sasahara et al., 2011; Stevens et al., 2012). Clinically serious infections of B. cereus are treated with antibiotics such as carbapenems, clindamycin, ciprofloxacin, and vancomycin (Kervick et al., 1990; Bottone, 2010; Uchino et al., 2012; Matsuda et al., 2014). However, resistance against carbapenem and clindamycin has been reported, which eventually led to failed treatments including cases with fatal outcomes (Kervick et al., 1990; Kiyomizu et al., 2008; Savini et al., 2009; Uchino et al., 2012).
According to the transportdb database, the B. cereus group strains constitute between 390 and 455 transporters per strain (Ren et al., 2007; Ren and Paulsen, 2007). The unusually high number of transporters per B. cereus group strain may reflect the different lifestyles of these bacteria. Importantly, each group member contains approximately 100 transporters, predicted to efflux drugs.
Drug efflux systems are part of the resistance machinery to counteract antibiotics (Sun et al., 2014). They are divided into six different transporter superfamilies: (i) MFS (major facilitator superfamily); (ii) ABC (ATP binding cassette) transporter superfamily; (iii) MATE (multidrug and toxic compound extrusion) family; (iv) RND (resistance nodulation division) family; (v) DMT (drug/metabolite transporter) superfamily, and (vi) PACE (proteobacterial antimicrobial compound efflux) (Poole, 2007; Hassan et al., 2015). Of these, MFS pumps constitute the majority of efflux transporters encoded in B. cereus group strains, typically more than 50 per strain. The MFS comprises secondary transporters that use the electrochemical gradient of protons or sodium ions across the cell membrane to energize substrate transport, including drug efflux (Pao et al., 1998; Saier et al., 1999; Reddy et al., 2012). The ‘transporter classification system’ (see http://www.tcdb.org/) classifies the MFS into 82 families. With respect to drug efflux pumps, the drug:H+ antiporter families (DHA)1 to 3 are the largest and best investigated drug exporter families in the MFS (Saier et al., 2014).
In this study, we characterize the phylogenetic and some functional properties of the putative multidrug transporter BC3310 from B. cereus ATCC 14579. BC3310 was classified by in silico analysis as a member of the major facilitator superfamily and the phylogenetic relationship within this group was determined. A deletion mutant of bc3310 was constructed and overexpression of BC3310 allowed for functional characterization in a heterogenous host as well as purification and partial biochemical characterization in vitro.
Materials and Methods
Bioinformatics Analyses
Bacterial sequence information was collected using the IMG homepage from the Joint Genome Institute (Markowitz et al., 2012). Sequence alignments were performed using MEGA MUSCLE alignment with default settings (Tamura et al., 2013) and the phylogenetic tree was constructed using MrBayes (Ronquist et al., 2012). Prediction of the transmembrane helices was done by submitting the primary protein sequence of BC3310 (UniProt Q81B77) to HMMTOP (Tusnady and Simon, 2001).
Construction of B. cereus bc3310 Deletion Mutant
A markerless mutant of bc3310 was constructed as described (Simm et al., 2012) in the B. cereus ATCC 14579 wild type according to the method of Janes and Stibitz (2006) and using the primers listed in Table 1. The B. cereus plasmid pBClin15 was lost during the process of making the markerless mutant and therefore a plasmid cured strain was used for phenotypic comparison as in previous investigations (Voros et al., 2013). The presence of the deletion was confirmed by sequencing. B. cereus was grown in LB medium at 30°C, unless otherwise stated.

TABLE 1. Primers used in this study.
Escherichia coli BC3310 Expression Constructs
The expression levels of genes cloned into pTTQ18-based plasmids are inducible by isopropyl β-D-thiogalactopyranoside (IPTG). Furthermore, the genes are fused with a sequence coding for a C-terminal (His)6 tag for identification and purification of the expressed protein. The plasmid construct pTTQ18-bc3310 (pbc3310) was made by general molecular biology techniques according to Sambrook and Russell (2001) by amplifying the gene bc3310 from genomic DNA of B. cereus ATCC 14579 using the primers listed in Table 1. The plasmids for expressing BC3310 D105 mutants pbc3310D105A, pbc3310D105N, and pbc3310D105E were made using sequence and ligation-independent cloning (Li and Elledge, 2007). The presence of each mutation was confirmed by sequencing. The E. coli strain DH5α ΔacrAB (Simm et al., 2012) carrying pTTQ18 empty vector or the overexpression plasmids was made for minimal inhibition concentration (MIC) testing. For protein purification the E. coli strain BL21 was transformed with pbc3310.
Escherichia coli strains harboring plasmids were grown in 50 or 250 ml LB medium with ampicillin (100 μg ml-1) at 37°C and 180 rpm in 250 ml or 1 l baffled flasks or on LB agar plates at 37°C, unless otherwise stated.
MIC Tests
Overnight cultures of B. cereus ATCC 14579 (without pBClin) and B. cereus Δ3310 or E. coli DH5α ΔacrAB (Simm et al., 2012) with relevant plasmid were inoculated 1:100 and grown to an OD600 between 0.8 and 1.0 at 37°C and 180 rpm. These pre-cultures were diluted to a final OD600 of 0.02. The test was performed at least three times in duplicate in microtiter plates and antibiotics were added in a 2-fold serial dilution. For susceptibility assay using E. coli strains 100 μg ml-1 ampicillin and 0.01 mM IPTG were added to all cultures. The cultures were incubated at 37°C for 20–24 h and visually inspected for growth. The lowest concentration, at which no growth was observed, was determined as the MIC.
Ethidium Bromide Accumulation Assay
Escherichia coli strains DH5α ΔacrAB with the plasmids pTTQ18 and pbc3310 were grown on LB agar plates supplemented with 100 μg ml-1 ampicillin and 0.01 mM IPTG at 37°C over night. Cells were collected with a loop and resuspended in PBS supplemented with 0.4% glucose (pH 7-7.4) to an OD600 of 1.000 (±0.005). These cells were applied on a microtiter plate and, where appropriate, carbonyl cyanide m-chlorophenylhydrazone (CCCP) was added to achieve an end concentration of 200 μM. Thereafter, ethidium bromide was added to an end concentration of 25 μM and the fluorescence change was measured over 60 min in a Safire spectrophotometer (Tecan, Crailsheim, Germany) with excitation and emission wavelength of 518 and 605 nm, respectively. Duplicate measurements were recorded on at least two cultures.
Heterologous Expression of BC3310 and Its Mutants with (His)6-tag and Western Blot
Overnight cultures of E. coli DH5α ΔacrAB carrying pbc3310, the empty vector (pTTQ18) or plasmids encoding the bc3310 mutants (pbc3310D105A, pbc3310D105N, or pbc3310D105E) were transferred to fresh LB (amp) medium and grown to an OD680 between 0.4 and 0.6. Expression was induced with 0.75 mM IPTG and the cultures were grown for another 3 h. For quantification of expression, Western blot assays were performed. One milliliter of the overexpression cultures was harvested by centrifugation at 15000 g, 4°C for 5 min. The pellet was washed (20 mM Tris-HCl pH 7.6, 100 mM NaCl, 5% glycerol, 1 mM phenylmethanesulfonylfluoride (PMSF)) and resuspended depending on cell mass in ice-cold lysis buffer (50 mM Tris-HCl pH 7.6, 100 mM NaCl, 5% glycerol, 5 mM β-mercaptoethanol, 1 mM PMSF, 1 μg ml-1 DNase). Cells were lysed by continuous sonication for 25 min in a cold water bath. SDS-PAGE and Western blots were performed as described in Sambrook and Russell (2001). (His)6-tag detection was done using a mouse anti-(His)6 antibody (Qiagen, Hilden, Germany) and a horse anti-mouse horseradish peroxidase-labeled secondary antibody (New England Biolabs. ECL advanced chemiluminescence detection reagent (Amersham Pharmacia Biotech, Pittsburgh, PA, USA) was used and chemiluminescence was measured by using the Analyzer Universal hood (Bio Rad, München) and the Quantity one 4.6.6 Software. Quantification was performed by pixel counting of five biological replicates on five different Western blots.
Purification of the BC3310 Protein by Affinity Chromatography
For protein expression and purification, the method described by Ward et al. (2000) was used. In short, E. coli strain BL21 pbc3310 was grown in 2TY medium (1.6% tryptone, 1% yeast extract, 0.5% sodium chloride, pH 7) and expression was induced at an OD680 between 0.4 and 0.6 with 0.75 mM IPTG. The culture was grown for another 3 h and cells were harvested. For inner membrane preparation, E. coli cells were resuspended in 20 mM Tris-HCl (pH 8.0), 0.5 mM EDTA and kept frozen at –80°C. After thawing, cells were disrupted with a Continuous Flow Disruptor (Constant Systems, UK) and inner membranes isolated by sucrose gradient centrifugation. Samples were kept at –80°C in Tris-HCl (pH 7.5) and EDTA.
Inner membranes were solubilized in 20 mM CAPSO (pH 10.0), 300 mM sodium chloride, 20% glycerol, 1% n-dodecyl β-D-maltoside (DDM), 20 mM imidazole (pH10.0). Immobilized metal affinity chromatography (IMAC) was performed using 20 mM CAPSO (pH 10.0), 10% glycerol, 0.05% DDM, 20 mM imidazole (pH 10.0) as wash buffer and 20 mM CAPSO (pH 10.0), 200 mM imidazole, 5% glycerol, and 0.05% DDM as elution buffer.
Circular Dichroism Measurement
Purified protein was washed using a spin concentrator with 20 mM CAPSO (pH 10.0), 5% glycerol and 0.05% DDM until imidazole-free. CD spectral analysis was performed from 270 to 195 nm in a 1 nm step resolution using a spectropolarimeter (Jasco J-715) with constant nitrogen flushing and a scan rate of 10 nm min-1. Response time was set at 1 s with a sensitivity of 100 mdeg and 10 nm bandwidth. The data comprised an accumulation of 20 scans, from which the buffer contribution was subtracted.
Results
BC3310 is Conserved in the B. cereus Group
To date, 228 strains of the B. cereus group of bacteria have been sequenced (Markowitz et al., 2012). A BLASTP search showed that the protein BC3310 is highly conserved within this group. In 225 strains BC3310 orthologs with >91% amino acid identity were identified. The predicted ortholog from the reduced genome sized B. cereus cytotoxis NVH 391-98 displayed 88% identity. The two strains (B. anthracis 3154 and B. anthracis A2012) in which no BC3310 ortholog was found are draft genomes which display a gap at the relevant genomic position (data not shown). Orthologs of the BC3310 protein are also found in other bacteria of the order Bacillales including B. subtilis (51% amino acid identity), Listeria innocua (47% amino acid identity), Geobacillus kaustophilus (47% amino acid identity), Lysinibacillus sphaericus (50% amino acid identity), Exiguobacterium sibiricum (39% amino acid identity), Anoxybacillus flavithermus (49% amino acid identity), Macrococcus caseolyticus (42% amino acid identity), Brevibacillus brevis (41% amino acid identity). The phylogenetic relationship of BC3310 to a selection of orthologs is depicted in a dendrogram (Figure 1). BC3310 clusters very closely with orthologous proteins from other B. cereus group members, thus forming a distinct cluster separate from the orthologs of other Bacillales species.
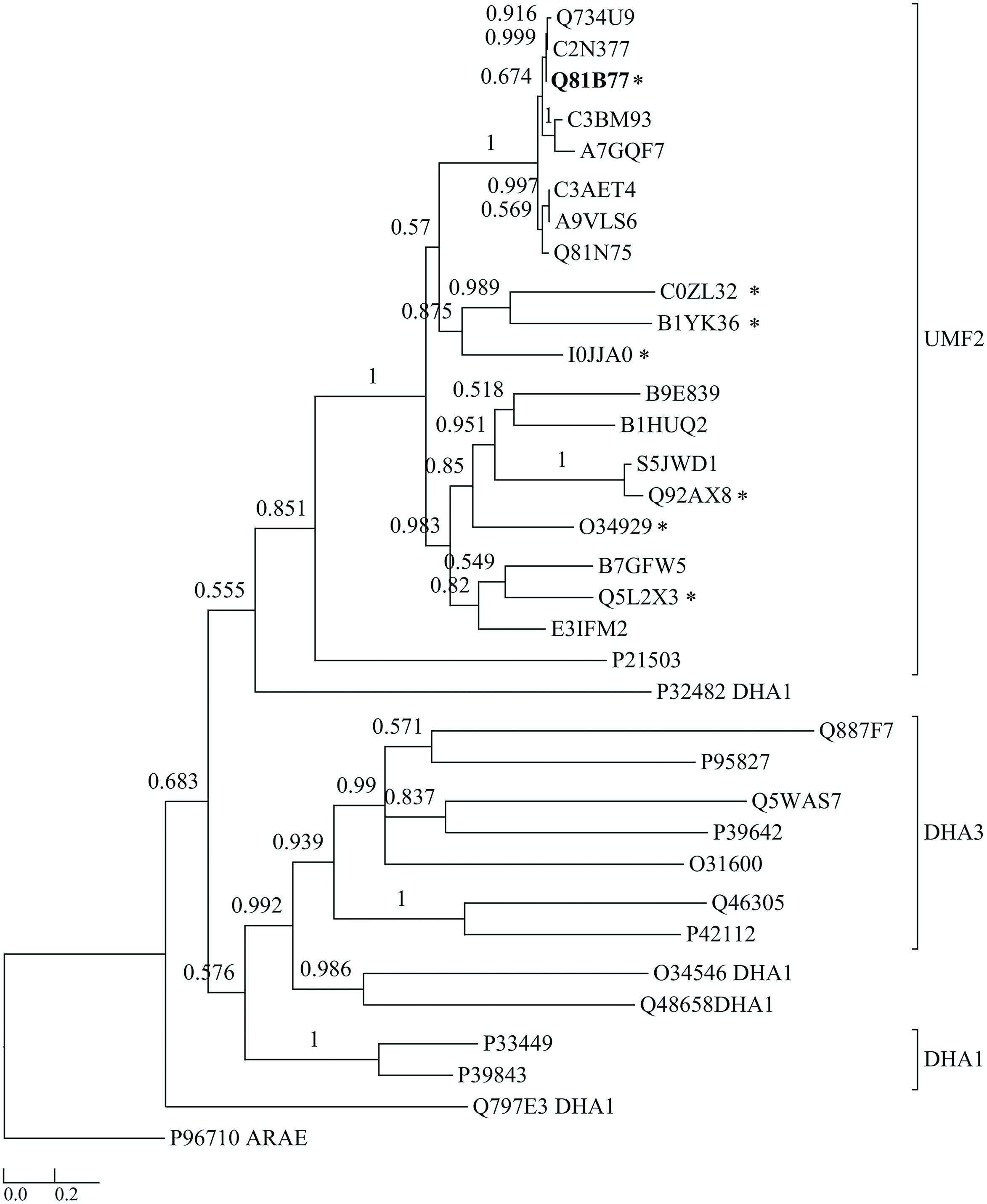
FIGURE 1. Dendrogram comparing BC3310 from Bacillus cereus ATCC 14579 with orthologous proteins and other multidrug transporters from the DHA1 and DHA3 families. BC3310 from B. cereus ATCC 14579 (UniProt accession number: Q81B77; bold font) and orthologous proteins from B. cereus ATCC 10987 (Q734U9), B. cereus ATCC 10876 (C2N377), B. anthracis str. Ames (Q81N75), B. cereus ssp. cytotoxis (A7GQF7), B. weihenstephanensis (A9VLS6), B. mycoides (C3AET4), B. pseudomycoides (C3BM93), Geobacillus sp. Y4.1MC1 (E3IFM2), Halobacillus halophilus (I0JJA0), B. subtilis (O34929), Listeria innocua (Q92AX8), Listeria monocytogenes (S5JWD1), Geobacillus kaustophilus (Q5L2X3), Lysinibacillus sphaericus (B1HUQ2), Exiguobacterium sibiricum (B1YK36), Anoxybacillus flavithermus (B7GFW5), M. caseolyticus (B9E839), Brevibacillus brevis (C0ZL32), Escherichia coli (P21503), and DHA1 proteins from Lactococcus lactis (Q48658), B. subtilis (Q797E3, O34546, P39843, P33449), Pseudomonas aeruginosa (P32482) and DHA3 proteins from Streptococcus pyrogenes (P95827), B. subtilis (P39642, O31600, P42112), B. clausii (Q5WAS7), Pseudomonas syringae (Q887F7), Clostridium perfringens (Q46305) and the sugar transporter AraE from B. subtilis (P96710) as an outgroup were used to build the tree. Posterior probability values are shown at each node and the bar represents the expected number of amino acid substitutions per site. The seven protein sequences marked with ∗ were aligned in Figure 5.
B. cereus Δbc3310 is More Susceptible to Ethidium Bromide Compared to the Wild Type
To examine the role of BC3310 in conferring drug tolerance in B. cereus ATCC 14579 a microbroth dilution test was conducted comparing the B. cereus wild type to its isogenic markerless knock-out mutant. Growth of the strains in twofold serial dilutions of ten compounds, including antibiotics from different classes, was tested. The susceptibility of the Δbc3310 mutant only differed from the susceptibility of the wild type strain for one of the 10 tested compounds. B. cereus Δbc3310 was two times more susceptible to ethidium bromide compared to the wild type (Table 2). It is possible that redundancy among efflux transporters masks the substrate range of the BC3310 transporter or that the transporter is not expressed under the conditions studied. Hence, a heterologous E. coli expression system with a hypersensitive E. coli strain and IPTG-inducible BC3310 expression was used to further investigate possible substrates.
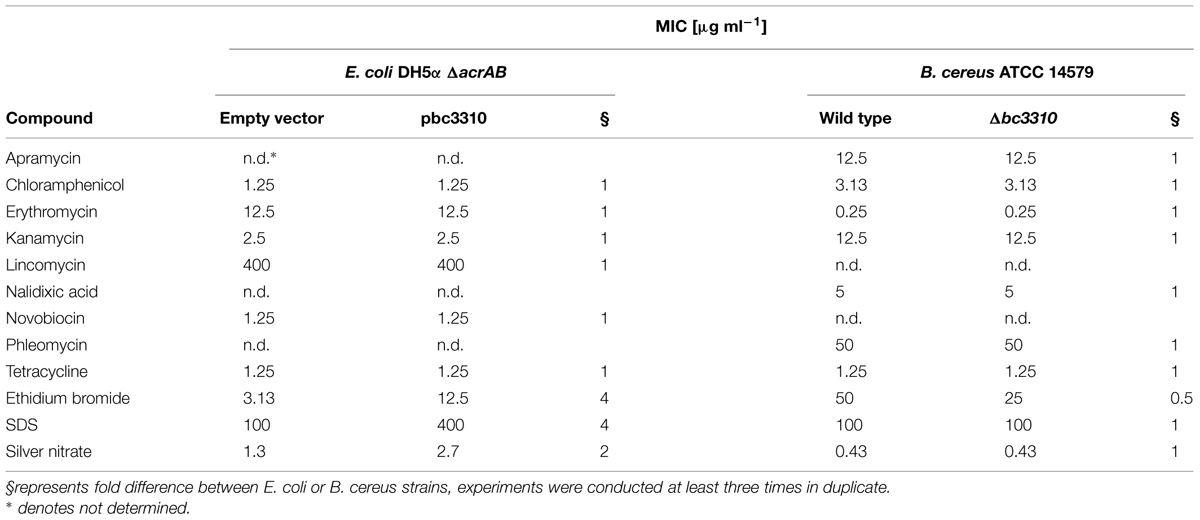
TABLE 2. Minimal inhibition concentration (MIC) of E. coli DH5α ΔacrAB expressing BC3310 (pbc3310) compared to empty vector control (pTTQ18) and Bacillus cereus ATCC 14579 Δbc3310 (Δbc3310) compared to B. cereus ATCC 14579 (wild type).
Expression of BC3310 Protein in E. coli
The ability of E. coli to heterologously express intact BC3310 protein was investigated. The bc3310 gene was cloned into the expression vector pTTQ18 as described (Saidijam et al., 2006, 2011; Szakonyi et al., 2007). BC3310 was expressed with a C-terminal RGSHis6 tag and detected by Western blotting using an antibody against the RGSHis6 tag (Figure 2). The protein was solubilized from the inner membrane fraction with DDMand purified by affinity chromatography (Figure 2). The major band on the Coomassie stained gel was subjected to Edman degradation and confirmed to contain the first eight predicted amino acids of BC3310. Topology analysis with HMMTOP predicted 12 transmembrane helices in the BC3310 transport protein. Circular dichroism measurements of the purified protein resulted in a spectrum with nodes at 210 and 222 nm (Figure 3), indicating a prevailing α-helical structure (Wallace et al., 2003) and thus confirming the integrity of the heterologously produced protein.
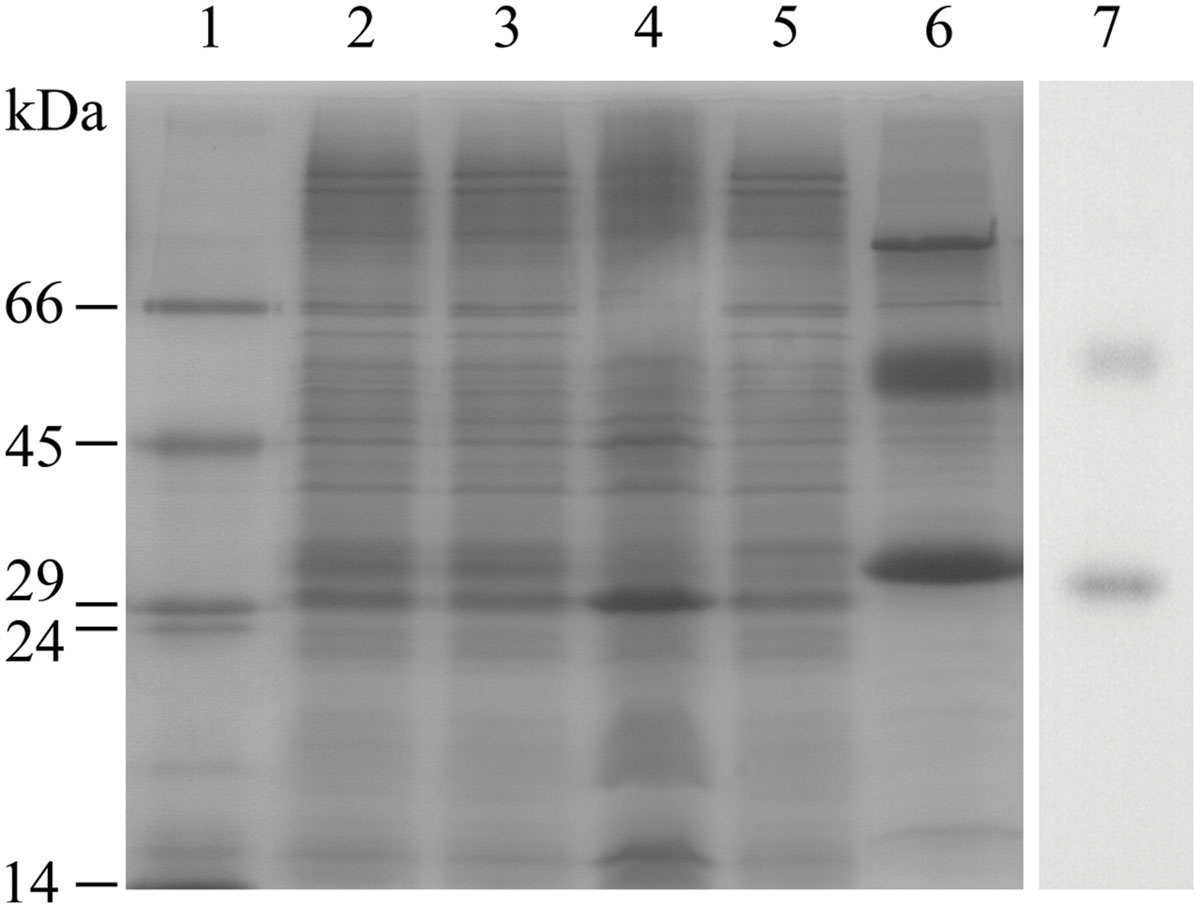
FIGURE 2. Purification of BC3310-RGSHis6. BC3310 was expressed with RGS(His)6-tag in E. coli and inner and outer membranes were separated. BC3310-RGSHis6 was solubilized with DDM and purified by immobilized metal affinity chromatography (IMAC). The SDS-PAGE was loaded as follows and stained with Coomassie Blue: (1) molecular weight marker; (2) inner membrane fraction; (3) solubilized protein; (4) unsolubilized protein; (5) flow-through after IMAC binding; (6) eluted protein. (7) Western blot detection of eluted BC3310 protein after affinity chromatography using an antibody to the RGSHis6 epitope.
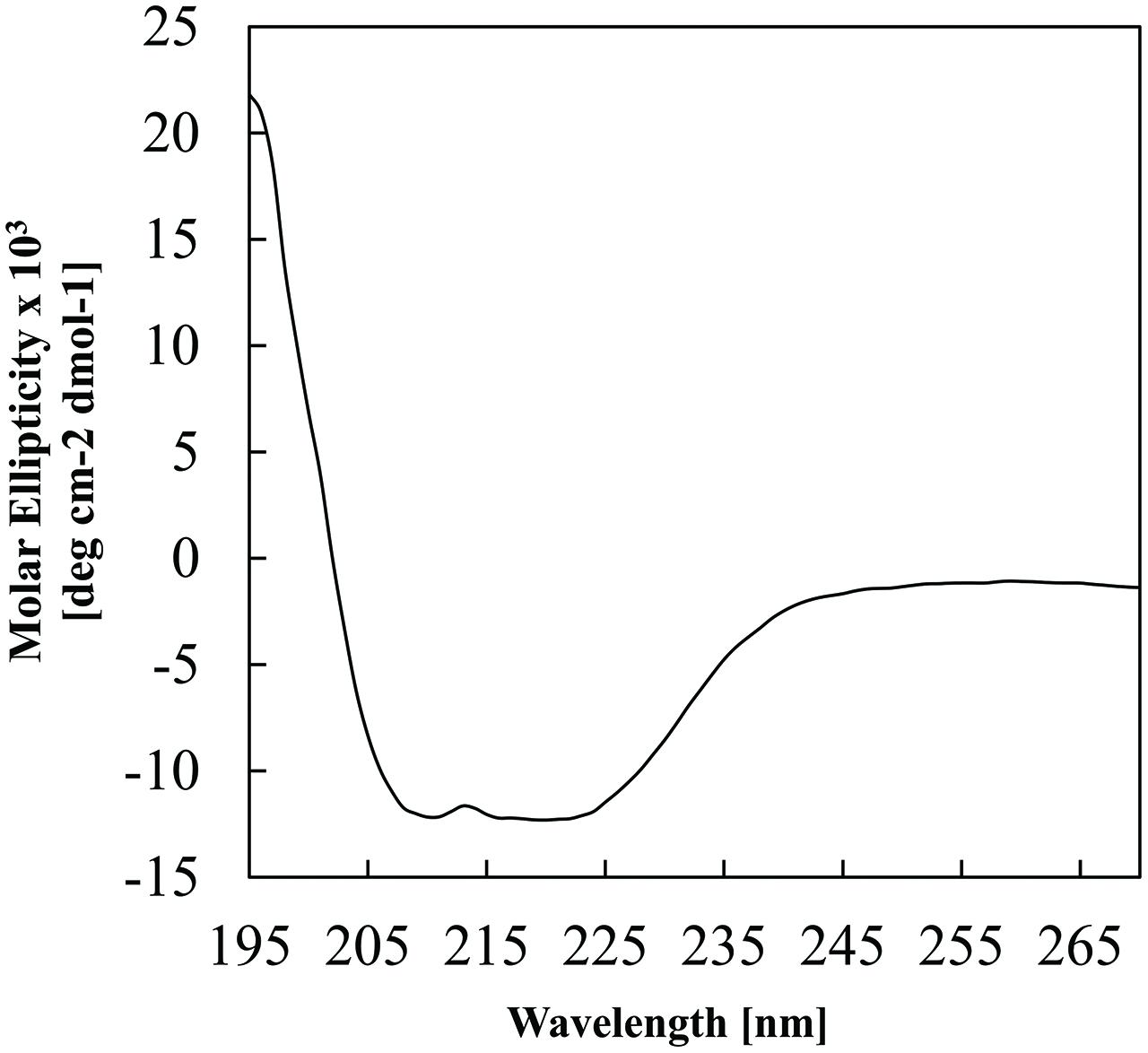
FIGURE 3. Circular dichroism analysis of purified BC3310-RGSHis6 protein. The analysis was performed at 1 nm intervals over 270–190 nm with a scan rate of 10 nm min-1. The spectrum represents an averaged accumulation of 20 scans, from which the buffer contribution was subtracted.
Thereafter the substrate range of heterologously expressed BC3310 was determined. A susceptibility assay was performed using E. coli DH5α ΔacrAB in which the major multidrug efflux complex was disrupted. The MICs of different compounds for the strain expressing BC3310 from pTTQ18 were compared to the MICs for the empty vector control. The E. coli strain expressing BC3310 showed a fourfold higher MIC for ethidium bromide and SDS and a twofold higher MIC for silver nitrate (Table 2).
Ethidium Bromide Efflux of BC3310 is Disrupted by CCCP
Major facilitator superfamily efflux proteins are secondary active transporters that utilize the electrochemical gradient across the cell membrane to extrude compounds. The BC3310 protein sequence displays motifs characteristic of an MFS transporter (see below) and so the ability of BC3310 to confer resistance to ethidium bromide by means of drug efflux was investigated further. A whole cell ethidium bromide accumulation assay with the E. coli DH5α ΔacrAB strain expressing BC3310 was performed. Ethidium bromide fluoresces upon binding to double-stranded DNA, and the fluorescence intensity correlates with the accumulation of ethidium bromide. The E. coli strain expressing bc3310 (pbc3310) showed less fluorescence compared to the empty vector control (pTTQ18), thereby implying that BC3310 exports ethidium bromide (Figure 4). Addition of the protonophore CCCP led to an increase in fluorescence intensity in the strain expressing bc3310 to approximately the control level (pTTQ18) (Figure 4, dark gray bars). This increase indicates the inability of BC3310 to export ethidium bromide due to the disruption of the electrochemical gradient.
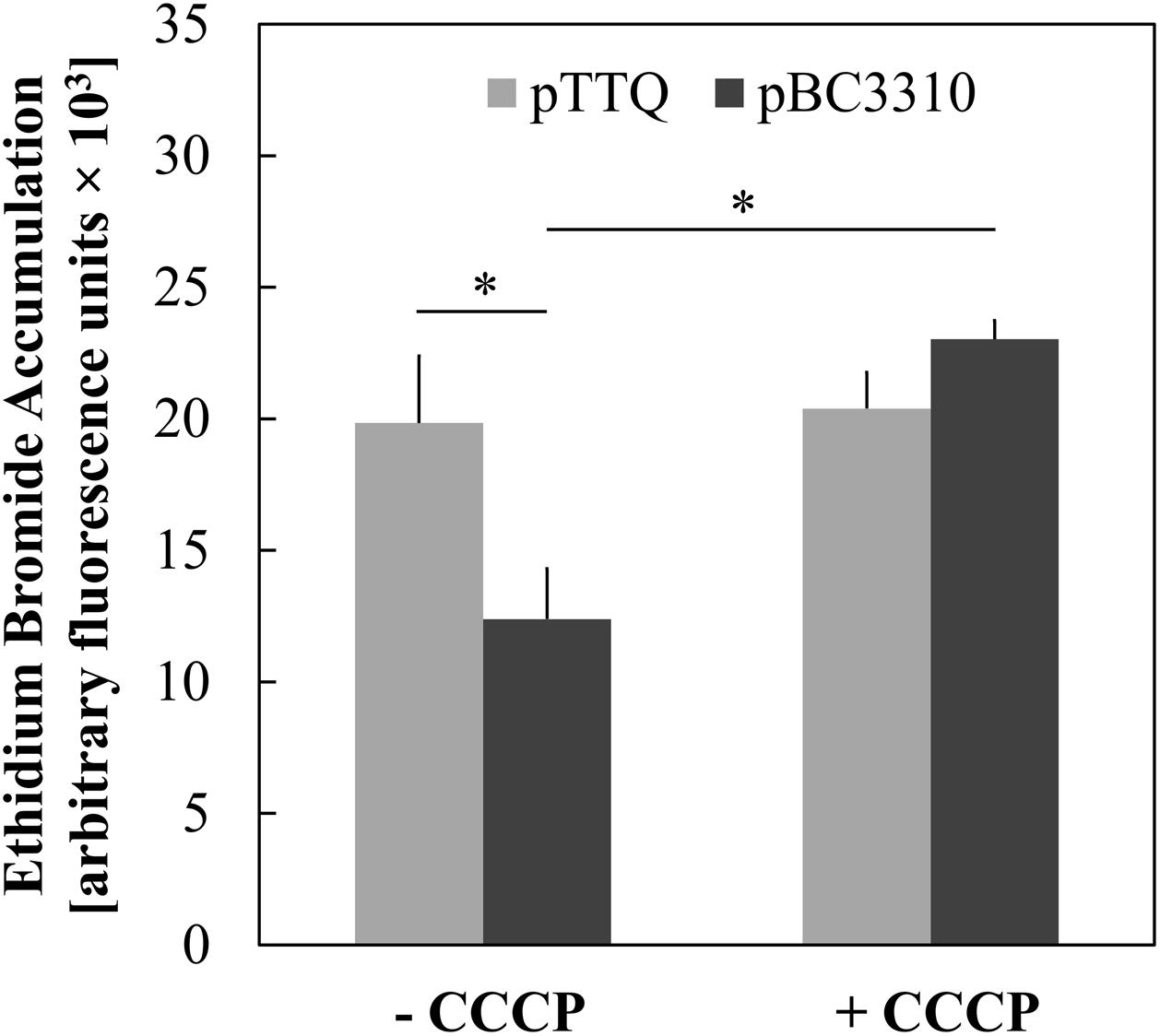
FIGURE 4. Uncoupler-sensitive efflux of ethidium associated with expression of BC3310. Accumulation of ethidium bromide after 30 min was measured in IPTG-induced E. coli DH5α ΔacrAB host cells either expressing bc3310 (pBC3310, dark gray) or not using an empty vector control (pTTQ18, light gray) without (-CCCP) and with addition of CCCP (+ CCCP). Values are means of four independent experiments and error bars indicate standard deviations, ∗p < 0.01, unpaired Student’s t-test.
Mutation of the Conserved Aspartic Acid Residue (D105) Abolishes Ethidium Bromide Efflux
Proton or substrate translocations by transport proteins often require acidic residues within transmembrane helices (Paulsen et al., 1996a; Edgar and Bibi, 1997; Sanderson et al., 1998; Dang et al., 2010). Sequence alignment of BC3310 with orthologous proteins revealed a conserved acidic residue in putative TMS 4 (Figure 5). In order to investigate the importance of this conserved aspartate residue (D105) for efflux activity, mutational analyses were conducted. Three constructs were made in which the aspartate residue was mutated to glutamate (D105E), asparagine (D105N), or alanine (D105A). The expression of the mutant proteins was detected and quantified by Western blot (Figure 6). This showed that the expression of all mutant proteins was three to four times higher compared to the expression of wild type protein. MIC determination of ethidium bromide and silver nitrate was performed to investigate the functionality of the mutant BC3310 proteins (Table 3). Even though more mutant protein was expressed, the susceptibility of strains expressing mutant BC3310 was reduced to levels approximating those of the empty vector control-strain. Thus, mutational change of the aspartate residue to another acidic or a structurally similar residue abolished the efflux ability of BC3310 for ethidium bromide and silver nitrate, indicating that both the size and charge of the side chain at position 105 are important for protein function.
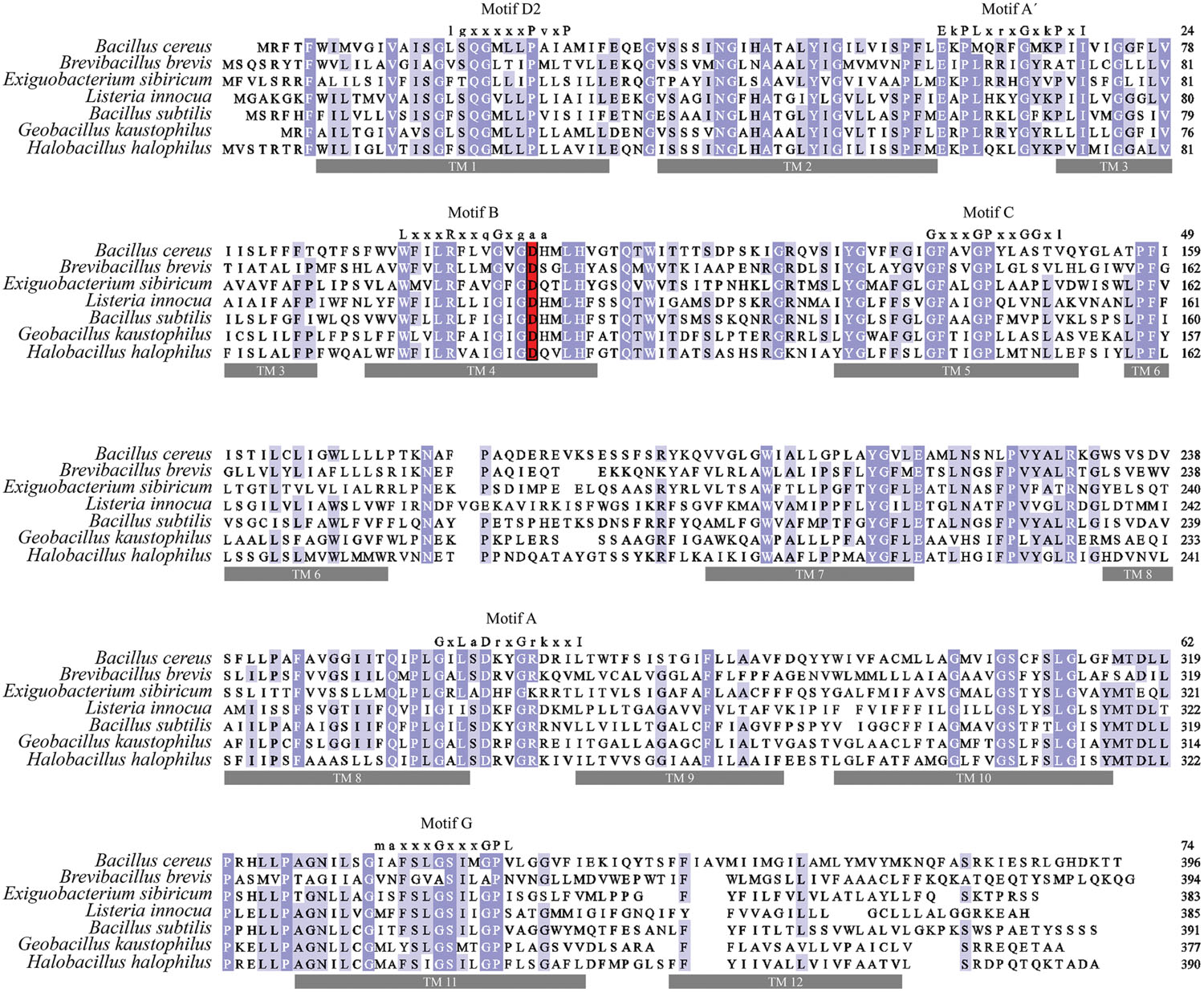
FIGURE 5. Multiple sequence alignment of BC3310 and its homologs. BC3310 from B. cereus ATCC 14579 (UniProt accession number: Q81B77) was aligned using MUSCLE with orthologs from Brevibacillus brevis (C0ZL32), Exiguobacterium sibiricum (B1YK36), Listeria innocua (Q92AX8), B. subtilis (O34929), Geobacillus kaustophilus (Q5L2X3) and Halobacillus halophilus (I0JJA0). Shading corresponds to >80% (dark blue), >60% (blue), >40% (light blue), and ≤40% (white) amino acid identity, respectively. The conserved acidic residue in transmembrane region 4 is displayed in red. Transmembrane regions have gray bars under and conserved MFS motifs are depicted above the sequence.
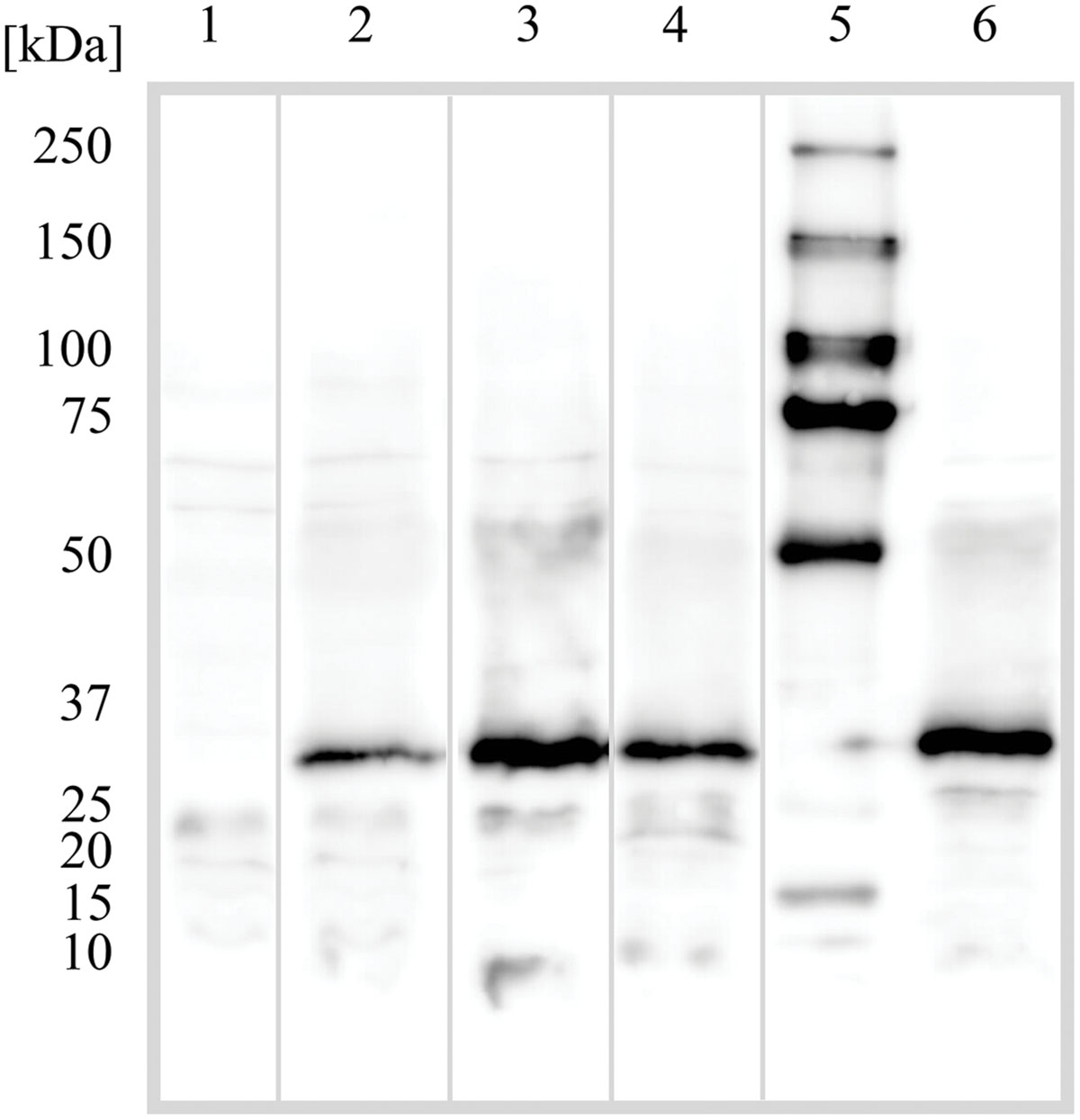
FIGURE 6. Western blot detection of the BC3310 protein and its mutants in Escherichia coli. E. coli DH5α ΔacrAB was transformed with empty pTTQ18 vector plasmid or pTTQ18 with the indicated insert; wild type BC3310 with his-tag and mutants D105N, D105A, and D105E under an IPTG inducible promoter. The cells were grown and prepared for SDS-PAGE as described in Section “Materials and Methods”. Numbers indicate: (1) empty pTTQ18 vector; (2) BC3310 wild type; (3) D105N mutant; (4) D105A mutant; (5) molecular weight marker; (6) D105E mutant. BC3310 and its mutants migrate at ∼30 kDa.

TABLE 3. Relative expression rate and relative MIC of E. coli strains producing no BC3310, BC3310 wild type, D105N, D105A, or D105E mutant protein.
BC3310 Belongs to the UMF-2 Family of the MFS
BC3310 showed prevailing α-helical structure in our CD analysis and is predicted to be a 12-TMS multidrug transporter belonging to the MFS. Most of the 12 TMS-containing MFS proteins that efflux several drugs are members of the drug:H+ antiporter families DHA1 and DHA3. To determine if BC3310 belongs to one of these families within the MFS, a multiple alignment of sequences orthologous to BC3310 and sequences from the well described DHA1 and DHA3 families was performed. From this alignment a dendrogram was built which showed clustering of BC3310 and orthologs in a distinct clade separate from the DHA1 and DHA3 family proteins included in the analysis (Figure 1). This analysis supported the transporter classification database (TCDB) division of YfkF, the BC3310 ortholog in B. subtilis, into a separate family, the unknown major facilitator family-2 (UMF-2) (Saier et al., 2014).
Transport Proteins within the UMF-2 Family Contain a Variant of the MFS Signature Motif A
Sequence alignment revealed that amino acid sequence motifs characteristic for MFS transporters, namely motif A, B, C, and G were conserved in BC3310 and orthologous proteins (Figure 5) (Henderson and Maiden, 1987; Griffith et al., 1992; Paulsen et al., 1996b). Motif A is conserved in the loop region between transmembrane segments (TMS) 2 and 3, and has been called the MFS signature motif due to its conservation across the superfamily. In the majority of MFS transporters, including the DHA1 family proteins, the motif A consensus sequence is G-x-L-a-D-r/k-x-G-r/k-r/k-x-x-I (x indicating any amino acid; capital and lower case letters representing amino acid frequency of >70% and 40–70%, respectively; Henderson and Maiden, 1987; Griffith et al., 1992; Paulsen et al., 1996b). However, a functional variant of this motif has been described in the Clostridium perfringens DHA3 family tetracycline efflux protein TetA(P): E-x-P-x-x-x-x-x-D-x-x-x-R-K (bold letters overlap with D, r/k,r/k of the canonical motif A) (Bannam et al., 2004). In BC3310 and its orthologs a modified motif A (motif A′) was identified, which represents a hybrid of the canonical motif A and the TetA(P) motif A (Table 4) (Paulsen et al., 1996b; Bannam et al., 2004). The N-terminal sequence of motif A′ in BC3310 orthologs resembles the TetA(P) (DHA3) motif A, with E and P conserved in both motifs, whereas the C-terminal sequence corresponds to the DHA1 motif A. This results in the BC3310 modified motif A′ sequence E-r/k-P-L-x-r/k-x-G-x-r/k-P-x-I (bold letters correspond to sequences of the previously described motif A sequences).
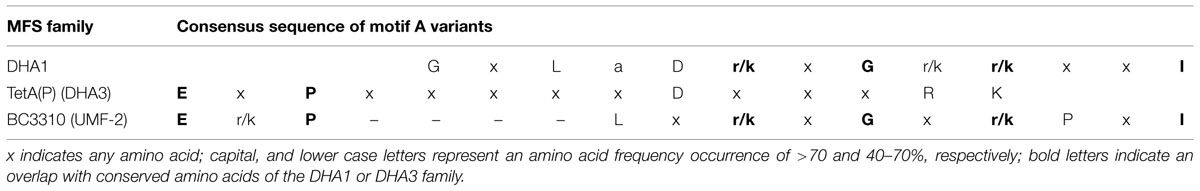
TABLE 4. Consensus sequences of motif A variants found in MFS drug export families.
As in other MFS transporters, a second motif A-like sequence is present between TMS 8 and TMS 9 in BC3310 (consensus sequence: G-x-L-S-D-r/k-x-G-R-r/k-x-x-i/l). This sequence coincides more with the signature motif A compared to the motif A′ sequence between TMS 2 and TMS 3 (Henderson and Maiden, 1987; Griffith et al., 1992).
Discussion
Heterologous expression of BC3310 in a drug hypersusceptible E. coli strain increased the tolerance of the bacteria to AgNO3, SDS, and ethidium bromide, indicating that it has a role in resistance to multiple drugs. Whole cell accumulation assays of ethidium bromide in E. coli expressing bc3310 demonstrated CCCP-sensitive efflux of ethidium in the drug hypersusceptible E. coli strain confirming a function as a drug efflux protein. Hence, BC3310 is an energy-dependent multidrug efflux pump. Inactivation of bc3310 in B. cereus ATCC 14579, also resulted in increased susceptibility to ethidium bromide, but not to SDS or AgNO3, suggesting, low basal expression of bc3310 under the conditions used in our experiments. It has, however, previously been reported that addition of 1 mM AgNO3 to exponentially growing cultures of B. cereus ATCC 14579 induced expression of bc3310 (Babu et al., 2011) and we detected AgNO3-induced temporal expression of bc3310 by qRT-PCR under our experimental settings (data not shown). Therefore, although BC3310 seems to have a role in transport of Ag+ and/or it is not essential in conferring AgNO3 resistance under the conditions tested, but may be important under specific circumstances. B. cereus ATCC 14579 contains 93 genes annotated as drug transporter which corresponds to 1.7% of the protein coding genes in the genome (Saidijam et al., 2006, 2011; Ren et al., 2007). In comparison, B. subtilis and E. coli display 32 and 37 genes encoding drug transport proteins, respectively, which correspond to 0.8 and 0.9% of the protein coding genes (Nishino and Yamaguchi, 2001; Ren et al., 2004). Considering the high number of annotated drug transporter genes in the genome of B. cereus, it is possible that one or more transporters compensate for the loss of BC3310, thereby concealing a potential effect of a gene disruption.
The efflux of ethidium bromide by BC3310 is dependent on a conserved aspartate residue, which could not be replaced by another acidic or hydrophobic amino acid. This indicates an important role of the aspartate residue at position 105 (D105) in the putative TMS 4. This residue is also conserved in BC3310 orthologs. Even though this aspartate residue is not reported to be one of the conserved residues, it falls into the boundaries of motif B. The motif B sequence of BC3310 and orthologs is W-x-x-L-R-x-x-x-G-x-G-D-x which overlaps to a large degree with canonical motif B L-x-x-x-R-x-x-q-G-x-g-a-a (bold letters indicate matching amino acids, underlined letter is D105 in BC3310). Motif B contains an absolutely conserved basic amino acid residue which is proposed to play a role in proton transfer (Paulsen and Skurray, 1993). This residue is also conserved in BC3310 (R98).
Sequence analyses classified BC3310 into the UMF-2 family of the MFS which is distinct from the well characterized drug efflux families DHA1 and DHA3 and consists of previously uncharacterized proteins. We have thus described the first functional data for a member of the UMF-2 family and showed that it includes multidrug efflux proteins. Previously transporters belonging to (at least) five of the 82 different families have been implicated in multidrug efflux. Besides the mentioned DHA1 and DHA3 families with 12 TMS-containing transporters, multidrug efflux proteins have been described for the Organic Cation Transporter family (2.A.1.19) (Koepsell, 2013). In addition, the DHA2 family is known to contain multidrug efflux proteins with 14 TMS (Paulsen et al., 1996b) and the gene encoding MdrA in Streptomyces coelicolor, classified into the Acriflavin Sensitivity family (2.A.1.36), is regulated by a TetR repressor that recognizes multiple drugs (Hayashi et al., 2013).
Interestingly, BC3310 and its orthologs contain an alternative motif A′ consensus sequence E-r/k-P-L-x-r/k-x-G-x-r/k-P-x-I between putative TMS 2 and 3. We propose that this consensus sequence can be used as a marker to distinguish the UMF-2 family from other MFS families. The presence of a second motif A in BC3310 is likely due to the duplication of 6 TMS during the evolution of the 12-TMS MFS transporters (Paulsen and Skurray, 1993). Similarly, motif G relates to a duplication of motif C (antiporter motif) (Paulsen et al., 1996b). Motif C is only conserved in exporters and not in importers (Paulsen and Skurray, 1993). This motif is also found with a high similarity (including the functionally important GP dipeptide; De Jesus et al., 2005) in BC3310 and orthologs which is in line with the efflux function of BC3310. Little similarity to MFS motif D2 is observed in the sequence alignment of BC3310 orthologs. As reported previously, motif D2 does not appear to be highly conserved in recently investigated 12-TMS MFS transporters and a function has not yet been assigned (Paulsen et al., 1996b; Kapoor et al., 2009).
The gene encoding the BC3310 transporter is highly conserved in the genomes of the B. cereus group members indicating that bc3310 belongs to the core genome of the B. cereus group. Comparison of the bc3310 genomic region of B. cereus ATCC 14579 with the equivalent regions of selected B. cereus group members, B. cereus ATCC 10987, B. cereus ATCC 10876, B. anthracis Ames Ancestor A2084, B. thuringiensis sv. kurstaki YBT-1520, and B. mycoides ATCC 6462 showed the same gene organization. The different species of the B. cereus group inhabit many different niches and display a high number of efflux transporter genes in the genome compared to other bacteria which could account for the different lifestyles (Saidijam et al., 2006, 2011). Thus, genes conserved in the genomes of the B. cereus group might play a role in the fundamental maintenance of physiological functions. Preliminary phenotypic microarray data using BIOLOG, however, did not reveal significant differences between B. cereus ATCC 14579 wild type and Δbc3310 mutant. Condition-dependent transcriptome analyses of the bc3310 ortholog, yfkF, in B. subtilis revealed relatively constant transcriptional activity across the conditions investigated (Nicolas et al., 2012). The highest level of gene expression was observed in cells within stationary (OD600 ∼2) or transition (OD600 ∼1.4) growth phases in LB medium or LB medium supplemented with glucose as well as on LB agar. Ethanol stress conditions revealed the lowest expression of this gene. Furthermore yfkF is predicted to be under the control of the housekeeping sigma factor SigA (Nicolas et al., 2012). Transcription of genes encoding multidrug transporters with a major role in protecting the cell against toxic compounds is generally activated by transcription factors that recognize toxic compounds or stress signals, such as AcrR, SoxS, MarR, and Rob in the case of AcrAB of E. coli (Ma et al., 1996; Sulavik et al., 2001; Randall and Woodward, 2002; Rosenberg et al., 2003). This fact and the minor intrinsic susceptibility against toxic compounds in the B. cereus Δbc3310 deletion mutant indicate that BC3310 is not a potent multidrug transporter with a main role in protecting the cell against toxic xenobiotics. It rather hints to an ancient and maybe general function in the normal physiology of the B. cereus group of bacteria. To further elucidate the role of this transporter the inactivation of other efflux proteins might be required.
Taken together, we have performed the first phylogenetic and functional characterization of a member of the UMF-2. The amino acid sequence of BC3310 comprises known motifs of the 12-TMS MFS transporters with a modified motif A′ between TMS 2 and TMS 3. BC3310 is a multidrug transporter with confirmed predominant α-helical structure. It confers resistance to ethidium bromide, SDS, and silver nitrate when expressed in E. coli. The export of ethidium bromide is energy dependent and requires a conserved aspartate residue in TMS 4. The deletion of bc3310 in B. cereus resulted in increased susceptibility to ethidium bromide under the conditions tested. The high conservation of bc3310 within the B. cereus group genomes indicates that it is part of the core genome. We hypothesize that the intrinsic role of BC3310 is not as a typical multidrug transporter, but rather as an important component in the normal physiology of the bacteria, under conditions that still remain to be identified.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was funded by the Norwegian Research Council (FUGE II Program, Project ID 183421). Collaborations between AK, JK, and AB are funded by the Deutscher Akademischer Austauschdienst Project ID 54564495 (DAAD) and the Norwegian Research Council. Financial support for this research in the PJFH laboratory was provided by grant agreement N° HEALTH-F4-2007-201924, European Drug Initiative for Channels and Transporters (EDICT) and by European Membrane Protein Consortium contract LSGH-CT-2004-504601 (EMeP). Consortium Collaborations between the three laboratories (AK, PH, IP) are funded by the EU BacMT, EDICT, and ATENS initiatives. We thank Ewa Jaroszewicz for valuable technical assistance, as well as Prof. Kern and Sabine Schuster for providing the opportunity and help during fluorescence measurements.
References
Ash, C., Farrow, J. A., Dorsch, M., Stackebrandt, E., and Collins, M. D. (1991). Comparative analysis of Bacillus anthracis, Bacillus cereus, and related species on the basis of reverse transcriptase sequencing of 16S rRNA. Int. J. Syst. Bacteriol. 41, 343–346.
Babu, M. M., Sridhar, J., and Gunasekaran, P. (2011). Global transcriptome analysis of Bacillus cereus ATCC 14579 in response to silver nitrate stress. J. Nanobiotechnol. 9, 49. doi: 10.1186/1477-3155-9-49
Bannam, T. L., Johanesen, P. A., Salvado, C. L., Pidot, S. J., Farrow, K. A., and Rood, J. I. (2004). The Clostridium perfringens TetA(P) efflux protein contains a functional variant of the Motif A region found in major facilitator superfamily transport proteins. Microbiology 150, 127–134. doi: 10.1099/mic.0.26614-0
Berg, G., Eberl, L., and Hartmann, A. (2005). The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 7, 1673–1685. doi: 10.1111/j.1462-2920.2005.00891.x
Bottone, E. J. (2010). Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 23, 382–398. doi: 10.1128/CMR.00073-09
Callegan, M. C., Booth, M. C., Jett, B. D., and Gilmore, M. S. (1999). Pathogenesis of gram-positive bacterial endophthalmitis. Infect. Immun. 67, 3348–3356.
Callegan, M. C., Cochran, D. C., Kane, S. T., Ramadan, R. T., Chodosh, J., Mclean, C., et al. (2006). Virulence factor profiles and antimicrobial susceptibilities of ocular Bacillus isolates. Curr. Eye Res. 31, 693–702. doi: 10.1080/02713680600850963
Centers for Disease Control and Prevention (2005) Outbreak of cutaneous Bacillus cereus infections among cadets in a university military program–Georgia, August 2004. MMWR Morb. Mortal. Wkly. Rep. 54, 1233–1235.
Dang, S., Sun, L., Huang, Y., Lu, F., Liu, Y., Gong, H., et al. (2010). Structure of a fucose transporter in an outward-open conformation. Nature 467, 734–738. doi: 10.1038/nature09406
De Jesus, M., Jin, J., Guffanti, A. A., and Krulwich, T. A. (2005). Importance of the GP dipeptide of the antiporter motif and other membrane-embedded proline and glycine residues in tetracycline efflux protein Tet(L). Biochemistry 44, 12896–12904. doi: 10.1021/bi050762c
Edgar, R., and Bibi, E. (1997). MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 179, 2274–2280.
Griffith, J. K., Baker, M. E., Rouch, D. A., Page, M. G., Skurray, R. A., Paulsen, I. T., et al. (1992). Membrane transport proteins: implications of sequence comparisons. Curr. Opin. Cell Biol. 4, 684–695. doi: 10.1016/0955-0674(92)90090-Y
Guinebretiere, M. H., Auger, S., Galleron, N., Contzen, M., De Sarrau, B., De Buyser, M. L., et al. (2013). Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of the Bacillus cereus Group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 63, 31–40. doi: 10.1099/ijs.0.030627-0
Hassan, K. A., Liu, Q., Henderson, P. J., and Paulsen, I. T. (2015). Homologs of the Acinetobacter baumannii AceI transporter represent a new family of bacterial multidrug efflux systems. MBio 6:e01982-14. doi: 10.1128/mBio.01982-14
Hayashi, T., Tanaka, Y., Sakai, N., Okada, U., Yao, M., Watanabe, N., et al. (2013). SCO4008, a putative TetR transcriptional repressor from Streptomyces coelicolor A3(2), regulates transcription of sco4007 by multidrug recognition. J. Mol. Biol. 425, 3289–3300. doi: 10.1016/j.jmb.2013.06.013
Henderson, P. J. F., and Maiden, M. C. J. (1987). Sugars, antibiotics, microbes and men. Trends Genet. 3, 62–64. doi: 10.1016/0168-9525(87)90174-0
Janes, B. K., and Stibitz, S. (2006). Routine markerless gene replacement in Bacillus anthracis. Infect. Immun. 74, 1949–1953. doi: 10.1128/IAI.74.3.1949-1953.2006
Jensen, G. B., Hansen, B. M., Eilenberg, J., and Mahillon, J. (2003). The hidden lifestyles of Bacillus cereus and relatives. Environ. Microbiol. 5, 631–640. doi: 10.1046/j.1462-2920.2003.00461.x
Kapoor, K., Rehan, M., Kaushiki, A., Pasrija, R., Lynn, A. M., and Prasad, R. (2009). Rational mutational analysis of a multidrug MFS transporter CaMdr1p of Candida albicans by employing a membrane environment based computational approach. PLoS Comput. Biol. 5:e1000624. doi: 10.1371/journal.pcbi.1000624
Kervick, G. N., Flynn, H. W. Jr., Alfonso, E., and Miller, D. (1990). Antibiotic therapy for Bacillus species infections. Am. J. Ophthalmol. 110, 683–687. doi: 10.1016/S0002-9394(14)77068-0
Kim, J. B., Jeong, H. R., Park, Y. B., Kim, J. M., and Oh, D. H. (2010). Food poisoning associated with emetic-type of Bacillus cereus in Korea. Foodborne Pathog. Dis. 7, 555–563. doi: 10.1089/fpd.2009.0443
Kiyomizu, K., Yagi, T., Yoshida, H., Minami, R., Tanimura, A., Karasuno, T., et al. (2008). Fulminant septicemia of Bacillus cereus resistant to carbapenem in a patient with biphenotypic acute leukemia. J. Infect. Chemother. 14, 361–367. doi: 10.1007/s10156-008-0627-y
Koepsell, H. (2013). The SLC22 family with transporters of organic cations, anions and zwitterions. Mol. Aspects Med. 34, 413–435. doi: 10.1016/j.mam.2012.10.010
Kolsto, A. B., Tourasse, N. J., and Okstad, O. A. (2009). What sets Bacillus anthracis apart from other Bacillus species? Annu. Rev. Microbiol. 63, 451–476. doi: 10.1146/annurev.micro.091208.073255
Li, M. Z., and Elledge, S. J. (2007). Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods 4, 251–256. doi: 10.1038/nmeth1010
Ma, D., Alberti, M., Lynch, C., Nikaido, H., and Hearst, J. E. (1996). The local repressor AcrR plays a modulating role in the regulation of acrAB genes of Escherichia coli by global stress signals. Mol. Microbiol. 19, 101–112. doi: 10.1046/j.1365-2958.1996.357881.x
Markowitz, V. M., Chen, I. M., Palaniappan, K., Chu, K., Szeto, E., Grechkin, Y., et al. (2012). IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res. 40, D115–D122. doi: 10.1093/nar/gkr1044
Martinez, M. F., Haines, T., Waller, M., Tingey, D., and Gomez, W. (2007). Probable occupational endophthalmitis from Bacillus cereus. Arch. Environ. Occup. Health 62, 157–160. doi: 10.3200/AEOH.62.3.157-160
Matsuda, S., Kirishima, T., Okamoto, N., Hisano, Y., Takai, K., Motoyoshi, T., et al. (2014). Bacillus cereus septicemia and necrotizing fasciitis in a patient with liver cirrhosis: a case report. Nihon Shokakibyo Gakkai Zasshi 111, 2013–2020.
Melo, A. L., Soccol, V. T., and Soccol, C. R. (2014). Bacillus thuringiensis: mechanism of action, resistance, and new applications: a review. Crit. Rev. Biotechnol. 1–10. doi: 10.3109/07388551.2014.960793 [Epub ahead of print].
Mock, M., and Fouet, A. (2001). Anthrax. Annu. Rev. Microbiol. 55, 647–671. doi: 10.1146/annurev.micro.55.1.647
Nicolas, P., Mader, U., Dervyn, E., Rochat, T., Leduc, A., Pigeonneau, N., et al. (2012). Condition-dependent transcriptome reveals high-level regulatory architecture in Bacillus subtilis. Science 335, 1103–1106. doi: 10.1126/science.1206848
Nishino, K., and Yamaguchi, A. (2001). Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol. 183, 5803–5812. doi: 10.1128/JB.183.20.5803-5812.2001
Pao, S. S., Paulsen, I. T., and Saier, M. H. Jr. (1998). Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 62, 1–34.
Paulsen, I. T., Brown, M. H., Littlejohn, T. G., Mitchell, B. A., and Skurray, R. A. (1996a). Multidrug resistance proteins QacA and QacB from Staphylococcus aureus: membrane topology and identification of residues involved in substrate specificity. Proc. Natl. Acad. Sci. U.S.A. 93, 3630–3635. doi: 10.1073/pnas.93.8.3630
Paulsen, I. T., Brown, M. H., and Skurray, R. A. (1996b). Proton-dependent multidrug efflux systems. Microbiol. Rev. 60, 575–608.
Paulsen, I. T., and Skurray, R. A. (1993). Topology, structure and evolution of two families of proteins involved in antibiotic and antiseptic resistance in eukaryotes and prokaryotes–an analysis. Gene 124, 1–11. doi: 10.1016/0378-1119(93)90755-R
Poole, K. (2007). Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 39, 162–176. doi: 10.1080/07853890701195262
Randall, L. P., and Woodward, M. J. (2002). The multiple antibiotic resistance (mar) locus and its significance. Res. Vet. Sci. 72, 87–93. doi: 10.1053/rvsc.2001.0537
Reddy, V. S., Shlykov, M. A., Castillo, R., Sun, E. I., and Saier, M. H. Jr. (2012). The major facilitator superfamily (MFS) revisited. FEBS J. 279, 2022–2035. doi: 10.1111/j.1742-4658.2012.08588.x
Ren, Q., Chen, K., and Paulsen, I. T. (2007). TransportDB: a comprehensive database resource for cytoplasmic membrane transport systems and outer membrane channels. Nucleic Acids Res. 35, D274–D279. doi: 10.1093/nar/gkl925
Ren, Q., Kang, K. H., and Paulsen, I. T. (2004). TransportDB: a relational database of cellular membrane transport systems. Nucleic Acids Res. 32, D284–D288. doi: 10.1093/nar/gkh016
Ren, Q., and Paulsen, I. T. (2007). Large-scale comparative genomic analyses of cytoplasmic membrane transport systems in prokaryotes. J. Mol. Microbiol. Biotechnol. 12, 165–179. doi: 10.1159/000099639
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Hohna, S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rosenberg, E. Y., Bertenthal, D., Nilles, M. L., Bertrand, K. P., and Nikaido, H. (2003). Bile salts and fatty acids induce the expression of Escherichia coli AcrAB multidrug efflux pump through their interaction with Rob regulatory protein. Mol. Microbiol. 48, 1609–1619. doi: 10.1046/j.1365-2958.2003.03531.x
Saidijam, M., Benedetti, G., Ren, Q., Xu, Z., Hoyle, C. J., Palmer, S. L., et al. (2006). Microbial drug efflux proteins of the major facilitator superfamily. Curr. Drug Targets 7, 793–811. doi: 10.2174/138945006777709575
Saidijam, M., Bettaney, K. E., Leng, D., Ma, P., Xu, Z., Keen, J. N., et al. (2011). The MFS efflux proteins of gram-positive and gram-negative bacteria. Adv. Enzymol. Relat. Areas Mol. Biol. 77, 147–166.
Saier, M. H. Jr., Beatty, J. T., Goffeau, A., Harley, K. T., Heijne, W. H., Huang, S. C., et al. (1999). The major facilitator superfamily. J. Mol. Microbiol. Biotechnol. 1, 257–279.
Saier, M. H. Jr., Reddy, V. S., Tamang, D. G., and Vastermark, A. (2014). The transporter classification database. Nucleic Acids Res. 42, D251–D258. doi: 10.1093/nar/gkt1097
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sanderson, N. M., Qi, D., Steel, A., and Henderson, P. J. (1998). Effect of the D32N and N300F mutations on the activity of the bacterial sugar transport protein, GalP. Biochem. Soc. Trans. 26, S306. doi: 10.1042/bst026s306
Sasahara, T., Hayashi, S., Morisawa, Y., Sakihama, T., Yoshimura, A., and Hirai, Y. (2011). Bacillus cereus bacteremia outbreak due to contaminated hospital linens. Eur. J. Clin. Microbiol. Infect. Dis. 30, 219–226. doi: 10.1007/s10096-010-1072-2
Savini, V., Favaro, M., Fontana, C., Catavitello, C., Balbinot, A., Talia, M., et al. (2009). Bacillus cereus heteroresistance to carbapenems in a cancer patient. J. Hosp. Infect. 71, 288–290. doi: 10.1016/j.jhin.2008.11.002
Simm, R., Voros, A., Ekman, J. V., Sodring, M., Nes, I., Kroeger, J. K., et al. (2012). BC4707 is a major facilitator superfamily multidrug resistance transport protein from Bacillus cereus implicated in fluoroquinolone tolerance. PLoS ONE 7:e36720. doi: 10.1371/journal.pone.0036720
Steen, M. K., Bruno-Murtha, L. A., Chaux, G., Lazar, H., Bernard, S., and Sulis, C. (1992). Bacillus cereus endocarditis: report of a case and review. Clin. Infect. Dis. 14, 945–946. doi: 10.1093/clinids/14.4.945
Stevens, M. P., Elam, K., and Bearman, G. (2012). Meningitis due to Bacillus cereus: a case report and review of the literature. Can. J. Infect. Dis. Med. Microbiol. 23, e16–e19.
Sulavik, M. C., Houseweart, C., Cramer, C., Jiwani, N., Murgolo, N., Greene, J., et al. (2001). Antibiotic susceptibility profiles of Escherichia coli strains lacking multidrug efflux pump genes. Antimicrob. Agents Chemother. 45, 1126–1136. doi: 10.1128/AAC.45.4.1126-1136.2001
Sun, J., Deng, Z., and Yan, A. (2014). Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 453, 254–267. doi: 10.1016/j.bbrc.2014.05.090
Szakonyi, G., Leng, D., Ma, P., Bettaney, K. E., Saidijam, M., Ward, A., et al. (2007). A genomic strategy for cloning, expressing and purifying efflux proteins of the major facilitator superfamily. J. Antimicrob. Chemother. 59, 1265–1270. doi: 10.1093/jac/dkm036
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Tusnady, G. E., and Simon, I. (2001). The HMMTOP transmembrane topology prediction server. Bioinformatics 17, 849–850. doi: 10.1093/bioinformatics/17.9.849
Uchino, Y., Iriyama, N., Matsumoto, K., Hirabayashi, Y., Miura, K., Kurita, D., et al. (2012). A case series of Bacillus cereus septicemia in patients with hematological disease. Intern. Med. 51, 2733–2738. doi: 10.2169/internalmedicine.51.7258
Voros, A., Simm, R., Kroeger, J. K., and Kolsto, A. B. (2013). Gene transcription from the linear plasmid pBClin15 leads to cell lysis and extracellular DNA-dependent aggregation of Bacillus cereus ATCC 14579 in response to quinolone-induced stress. Microbiology 159, 2283–2293. doi: 10.1099/mic.0.069674-0
Wallace, B. A., Lees, J. G., Orry, A. J., Lobley, A., and Janes, R. W. (2003). Analyses of circular dichroism spectra of membrane proteins. Protein Sci. 12, 875–884. doi: 10.1110/ps.0229603
Ward, A., Sanderson, N. M., O’reilly, J., Rutherford, N. G., Poolman, B., and Henderson, P. J. F. (2000). “The amplified expression, identification, purification, assay and properties of histidine-tagged bacterial membrane transport proteins,” in Membrane Transport – A Practical Approach, ed. S. A. Baldwin (Oxford: Blackwell Press), 141–166.
Keywords: MFS, drug resistance, efflux protein, Bacillus cereus, UMF-2
Citation: Kroeger JK, Hassan K, Vörös A, Simm R, Saidijam M, Bettaney KE, Bechthold A, Paulsen IT, Henderson PJF and Kolstø A-B (2015) Bacillus cereus efflux protein BC3310 – a multidrug transporter of the unknown major facilitator family, UMF-2. Front. Microbiol. 6:1063. doi: 10.3389/fmicb.2015.01063
Received: 17 August 2015; Accepted: 15 September 2015;
Published: 12 October 2015.
Edited by:
Miklos Fuzi, Semmelweis University, HungaryReviewed by:
Atte Von Wright, University of Eastern Finland, FinlandDmitri Debabov, NovaBay Pharmaceuticals, USA
Copyright © 2015 Kroeger, Hassan, Vörös, Simm, Saidijam, Bettaney, Bechthold, Paulsen, Henderson and Kolstø. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anne-Brit Kolstø, Laboratory for Microbial Dynamics, Department of Pharmaceutical Biosciences, School of Pharmacy, University of Oslo, Postbox 1068 Blindern, 0316 Oslo, Norway, a.b.kolsto@farmasi.uio.no
†Present address: Roger Simm, Norwegian Veterinary Institute, Oslo, Norway; Massoud Saidijam, Research Centre for Molecular Medicine, Department of Molecular Medicine and Genetics, School of Medicine, Hamadan University of Medical Sciences, Hamadan, Iran