 Zhilin Wu1,2
Zhilin Wu1,2 Xuebin Yin2,3,4
Xuebin Yin2,3,4 Gary S. Bañuelos5
Gary S. Bañuelos5 Zhi-Qing Lin6Zhu Zhu7Ying Liu2,3,4Linxi Yuan2,3,4*
Zhi-Qing Lin6Zhu Zhu7Ying Liu2,3,4Linxi Yuan2,3,4* Miao Li1,4*
Miao Li1,4*- 1Key Laboratory of Agri-Food Safety of Anhui Province and Laboratory of Quality and Safty Risk Assessment for Agricultural Products on Storage and Preservation of the Ministry of Agriculture (Hefei), School of Plant Protection – School of Resources and Environment, Anhui Agricultural University, Hefei, China
- 2School of Earth and Space Sciences, University of Science and Technology of China, Hefei, China
- 3Jiangsu Bio-Engineering Research Centre of Selenium, Suzhou, China
- 4Institute of Advanced Technology, University of Science and Technology of China, Hefei, China
- 5Water Management Research Unit, United States Department of Agriculture – Agricultural Research Service, Parlier, CA, USA
- 6Environmental Sciences Program and Department of Biological Sciences, Southern Illinois University Edwardsville, Edwardsville, IL, USA
- 7School of Chemistry and Biological Engineering, University of Technology and Science Beijing, Beijing, China
Selenium (Se) has important benefits for crop growth and stress tolerance at low concentrations. However, there is very little information on antimicrobial effect of Se against the economically important fungus Botrytis cinerea. In the present study, using sodium selenite as Se source, we investigated the effect of Se salts on spore germination and mycelial growth of the fungal pathogen in vitro and gray mold control in harvested tomato fruit. Se treatment at 24 mg/L significantly inhibited spore germination of the fungal pathogen and effectively controlled gray mold in harvested tomato fruit. Se treatment at 24 mg/L seems to induce the generation of intracellular reactive oxygen species in the fungal spores. The membrane integrity damage was observed with fluorescence microscopy following staining with propidium iodide after treatment of the spores with Se. These results suggest that Se has the potential for controlling gray mold rot of tomato fruits and might be useful in integrated control against gray mold disease of postharvest fruits and vegetables caused by B. cinerea. The mechanisms by which Se decreased gray mold decay of tomato fruit may be directly related to the severe damage to the conidia plasma membrane and loss of cytoplasmic materials from the hyphae.
Introduction
Mineral nutrients has important benefits for the growth and development of many organisms, and are essential factors in biotic plant interactions while influencing plant health (Ahn et al., 2005; Datnoff et al., 2007; Cabot et al., 2013; Crane et al., 2014). The most recent information regarding the effect of nutrients on disease resistance and tolerance to pathogens has been reported by others (Boyd, 2007; Fones et al., 2010; Hörger et al., 2013). Specific micronutrients (Cu, Fe, Mn, Zn, and B) reportedly affect many plant diseases (Dordas, 2008; Yao et al., 2012; Fones and Preston, 2013). In addition, some relatively non-toxic organic and inorganic salts such as potassium silicate and calcium chloride have been investigated for inhibition of fungal pathogens on fruits, vegetables, field crops, and ornamentals (Hervieux et al., 2002; Shi et al., 2011; Yaganza et al., 2014). Though selenium (Se) has been found to be beneficial to plants, its effect on plant–disease interaction has little been evaluated, and there are few investigations on Se salt treatment for control of plant diseases (Hanson et al., 2003; Trumble and Sorensen, 2008; Companioni et al., 2012).
Selenium is an essential trace element for humans and animals but at high concentrations, Se becomes toxic to organisms due to Se replacing sulfur in proteins (Zhao and McGrath, 2009; Zhu et al., 2009; Hasanuzzaman et al., 2010, 2014; Wu et al., 2015). The range between beneficial and harmful Se concentrations is relatively narrow for humans and animals. The minimal Se concentration in livestock feed is 0.05–0.10 mg/kg dry forage, while the toxic Se concentration in animal feed is 2–5 mg/kg dry forage. In humans, the World Health Organization (WHO) and USDA recommended the required human dietary intake of Se to be 55–200 μg/day for adults (Wu et al., 2015). Se is of great interest in many facets of biomedicine, biochemistry, and environmental science (Tapiero et al., 2003; Bañuelos, 2006; Doucha et al., 2009; Winkel et al., 2011; Yuan et al., 2013). Moreover, Se is used for obtaining more bioactivity and safety for chemical pesticide (Ma et al., 2006; Singh, 2012). Selenium sulfide and sodium selenite have been tested for inhibition of some pathogens (Brotherton, 1967; Wu et al., 2014). A mixture fungicide of Dithane and sodium selenite was more effective against Aspergillus funiculosus and Alternaria tenuis (Razak et al., 1991). The application of Se at low concentrations as possible alternatives to synthetic fungicides for the control of plant diseases, may reduce the potential hazardous effect on the environment and human health (Hasanuzzaman et al., 2010; Wu et al., 2014).
Gray mold decay, caused by species of Botrytis cinerea, is one of the most important postharvest diseases of fresh fruit and vegetables worldwide (Qin et al., 2010; Youssef and Roberto, 2014). Infections by B. cinerea that could lead to severe economic losses both at pre- and post-harvest stages (Soylu et al., 2010; Cabot et al., 2013). Currently, the use of synthetic chemical fungicides is the primary means to control postharvest diseases (Zhu et al., 2010). However, growing concern for the potential impact of fungicide residues on the environment and human health risks, the development of fungicide-resistant strains of pathogen, and the lack of the most effective fungicides have created an interest in exploring for alternative approaches for the disease management (Janisiewicz and Korsten, 2002). Among alternative control methods, the application of antagonistic microorganisms, and the use of bicarbonates, chitosan, hot water, essential oils, either alone or in combination with other treatments as part of the integrated management of postharvest diseases strategies, have been investigated for the control of postharvest decay of fruits and vegetables with some success (Karabulut et al., 2004, 2005; Gabler et al., 2005; Droby et al., 2009; Soylu et al., 2010; Zhang et al., 2014; de Souza et al., 2015). However, there is a need for developing alternative methods to control postharvest gray mold of fruits and vegetables that are safe, effective, economical, and compatible with value-added practices such as producing Se-biofortified fruits and vegetables.
Our previous studies showed that sodium selenite could control postharvest disease of fruits and vegetables caused by Penicillium expansum (Wu et al., 2014). However, to our knowledge, little information is available concerning the antimicrobial activity of sodium selenite against postharvest pathogen B. cinerea and that the mechanism of action still remains essentially unexplored. Therefore, we aimed to explore the potential of using sodium selenite as an antifungal agent for the control of B. cinerea that causes gray mold disease on postharvest fruits and vegetables. In the present study, the experiments in vitro and in vivo were designed with sodium selenite treatments to directly inhibit B. cinerea, and to investigate the control of gray mold disease on postharvest tomato fruit. The underlying mechanisms of action by which sodium selenite caused fungal death, and affected intracellular reactive oxygen species (ROS) and the integrity of the plasma membrane of B. cinerea were also assessed.
Materials and Methods
Pathogen and Chemicals
The postharvest fungal pathogen B. cinerea used in this study was obtained from Key Laboratory of Plant Resources, Institute of Botany, Chinese Academy of Sciences. The fungus was inoculated and re-isolated from tomato fruit to maintain pathogenicity. The isolates were routinely grown on potato dextrose agar (PDA) plates for 2 weeks at 23°C in the darkness. The spores were obtained from the surface of the agar and suspended in 5 mL of sterile distilled water containing 0.1% v/v Tween 20. Spore suspensions were filtered through four layers of sterile cheesecloth to remove mycelia fragments. A hemocytometer was used to calculate the number of spores and the spore concentration was adjusted to 5.0 × 105 spores/mL before conducting subsequent experiments.
Sodium selenite used as Se source in this study and was purchased from Sigma–Aldrich (St. Louis, MO, USA). Propidium Iodide (PI), 2,7-dichlorodihydrofluorescein diacetate (DCHF-DA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals used in this study were of high analytical grade.
Inhibitory Effect of Se on Spore Germination and Germ Tube Elongation of B. cinerea
The effect of sodium selenite on spore germination and germ tube elongation of B. cinerea was assayed in potato dextrose broth (PDB) medium following the method described by Qin et al. (2010). In brief, aliquots of a spore suspension of B. cinerea were added to wells of a 24-well microtitration plate containing PDB medium to obtain a final concentration of 5 × 105 spores/ml. The culture medium was supplemented with different concentrations of sodium selenite at 0, 6, 12, 18, 24 mg Se/L at pH of 7.0, which are referred to Razak et al. (1991) and Wu et al. (2014, 2015). The microtitration plate was incubated at 23°C on a rotary shaker at 200 rpm. About 200 spores were examined microscopically for germination level and germ tube length after 8 h of incubation. Spores were considered germinated if the germ tube was equal to or greater than the diameter of the spore. Germ tube length was determined with an ocular micrometer and germinated spores were expressed as percentage of the total number of evaluated spores. There were three replicates for each treatment, and the experiment was repeated twice.
Effect of Se on Mycelial Growth of B. cinerea
The effect of sodium selenite on mycelial growth of B. cinerea was assayed in PDA following the method described by Qin et al. (2010). A 5-mm diameter plug of mycelial agar was obtained from the growing edge of 7-day-old cultures of the fungi and placed in the center of a 9-cm-diameter petri dish containing PDA medium with different concentrations (0, 6, 12, 18, 24 mg Se/L) as sodium selenite solutions. Sodium selenite solutions were filtered through a 0.45 μm Millipore filter before they were, respectively, added to the autoclaved PDA medium that had cooled to 60°C. Results are reported as the diameter of the fungal colony (mycelium) in the petri dish minus the diameter of the agar plugs (5 mm). Radial growth of B. cinerea was observed after incubation at 23°C for 3, 4, 5, 6, and 7 days, respectively. Radial growth of B. cinerea was calculated as the average of the orthogonal diameter, the results were expressed as minimum inhibitory concentration (MIC) according to Arslan et al. (2009). Each treatment contained three replicates and the entire experiment was repeated twice.
Reactive Oxygen Species (ROS) Assay
The oxidant-sensitive probe DCHF-DA was used to assess the intracellular ROS levels in B. cinerea according to Shi et al. (2012) with some modifications. Spores of B. cinerea were cultured in PDB medium supplemented with 0 or 24 mg/L sodium selenite and incubated for 1 and 3 h. The spores were washed with 10 mM potassium phosphate buffer (pH 7.0) and incubated for 1 h in the same buffer containing 10 μM DCHF-DA (dissolved in dimethyl sulfoxide). After washing twice with potassium phosphate buffer, spores were examined under a AMG microscope (AMG, EVOS-S1, USA) using a fluorescein 2,7-dichlorodihydro-specific filter. At least 100 spores were examined for each treatment with three replications.
Membrane Integrity Assay and Microscopy
The fluorescence microscopy following staining with PI was used to assess the membrane integrity of B. cinerea conidia according to the methods of Wu et al. (2014) with some modifications. Spores of B. cinerea were cultured in PDB medium supplemented with 0 and 24 mg/L sodium selenite and incubated for 1, 3, and 5 h. Then, the spores were collected and stained with 10 μg/ml PI for 5 min at 30°C. Because PI is membrane impermeant, only cells that have lost membrane integrity will show red staining. The spores were centrifuged and washed twice with 10 mM potassium phosphate buffer (pH 7.0) to remove residual dye. The spores were then observed on a AMG microscope (AMG, EVOS-S1, USA) equipped with individual fluorescein rhodamine filter set (excitation BP 546/12 nm, emission LP 590 nm), and the images were collected. At least 100 spores were examined for each treatment with three replications.
Cellular Leakage Assay
The leakage of cytoplasmic contents from mycelium of B. cinerea was determined according to the methods of Lai et al. (2011) with some modifications. B. cinerea was grown in conical flasks (250 mL) containing 100 mL PDB medium at 23°C on a rotary shaker at 200 rpm and mycelia harvested after 3 days of incubation. After pooled and washed with sterile distilled water, the mycelia were re-suspended in 100 mL sterile distilled water containing 0–24 mg Se/L as sodium selenite, and incubated on a rotary shaker at 23°C for 1, 2, 3, and 4 h. Mycelia were then filtered from the solutions through a 0.2 μm pore size membrane and the aqueous solutions were used for determination of soluble proteins and carbohydrates. The Bradford assay was performed on the filtrates to quantify proteins by the various treatments (Bradford, 1976). Soluble carbohydrate was determined with anthrone reagent using glucose as the standard (Xu et al., 2006).
Measurement of Cellular Enzymatic Activity
Botrytis cinerea was grown in conical flasks (50 mL) containing 20 mL PDB medium with 0–24 mg Se/L as sodium selenite at 23°C on a rotary shaker at 200 rpm and mycelia were harvested after 1, 2, and 3 days of incubation. The leakage of cytoplasmic contents from mycelium of B. cinerea was determined according to the method of Lai et al. (2011) with some modifications. The 0.5 g mycelia were filtered from the solutions and ground into homogenate with phosphate buffer (pH 6.8) and a small quantity of quartz sand in liquid nitrogen. The homogenate was centrifuged at 12000 rpm for 20 min and the supernatant was used for determination of enzymatic activity. The Bradford assay was performed on the supernatant to quantify release of proteins by the various treatments according to the method of Bradford (1976) with some modifications. The content of glutathione (GSH), methane dicarboxylic aldehyde (MDA), and superoxide dismutase (SOD) was determined with the previously reported method (Liu et al., 2005). Hydrogen peroxide (H2O2) and superoxide anion (O2-) production rate were determined with the methods of Xu et al. (2006).
Effect of Se on Gray Mold Disease on Tomato Fruit
Seeds of tomato (Lycopersicon esculentum Mill. cv. Zhenbao) were obtained from the Academy of Agricultural Science in Anhui, China. During all experiments, tomato plants were greenhouse grown at Suzhou Key Advanced Laboratory for Selenium and Human Health, Suzhou Institute for Advanced Study, University of Science and Technology of China (Suzhou, China) with a mean temperature cycle of 31°C day/24°C night, relative humidity of 43%/85%, and a light period of 16 h, at 450–500 μmol/m2/s maximum photon flux densities that were measured at plant canopy level (IRGA, model LCA-4, Hoddesdon, UK). The fruits were harvested at commercial maturity (surface color of the fruit was red), and sorted based on size and the absence of physical injuries or disease infection. Before treatments, fruits were surfaced-disinfected with 2% sodium hypochlorite for 3 min, then rinsed with sterile water, and air-dried. The surface of the fruit was prepared for inoculation by inflicting a single 1-mm deep wound in the middle of each fruit with a sterile needle. Each wound was then inoculated with the pathogen B. cinerea by placing 10 μL of spore suspension (1 × 104 spores/L) on the wound. The inoculated fruit were incubated overnight in a sterile box at 23°C before dipping it in the sodium selenite solution at concentration of 24 mg/L for 30 min. Fruit dipped in sterile distilled water served as the control. The treated fruit were incubated in a moist plastic box at 23°C for 6 days and disease development was assessed by measuring the lesion diameter and decay incidence of the gray mold on tomato fruit. Each treatment consisted of 50 fruits with three replications and the experiment was repeated twice.
Statistical Analysis
All statistical analyses were performed using the SPSS software version 13.0 (SPSS Inc., Chicago, IL, USA) and analyzed by one-way analysis of variance (ANOVA). Mean separations were performed by Duncan’s multiple range tests. Differences at p < 0.05 were considered significant.
Results
Effects of Se on Spore Germination and Germ Tube Elongation of B. cinerea
As shown in Table 1, selenite was effective in inhibiting spore germination and germ tube elongation of B. cinerea following a concentration-dependent trend (p < 0.05). After incubation for 8 h at 23°C, germination of B. cinerea spores was strongly inhibited by selenite at the concentration of 24 mg/L.

TABLE 1. Effects of Se on spore germination and germ tube elongation of Botrytis cinerea after 8 h incubation at 23°C.
Inhibitory Effect of Se on Mycelial Growth of B. cinerea
As shown in Table 2, sodium selenite was effective in inhibiting the mycelial growth of B. cinerea on PDA. After incubation for 3–7 days at 23°C, the growth of B. cinerea was significantly inhibited when the treatment concentration of sodium selenite was 24 mg/L (Table 2). Moreover, there was a significantly positive correlation between sodium selenite concentration and the effect on mycelial growth (Table 2).
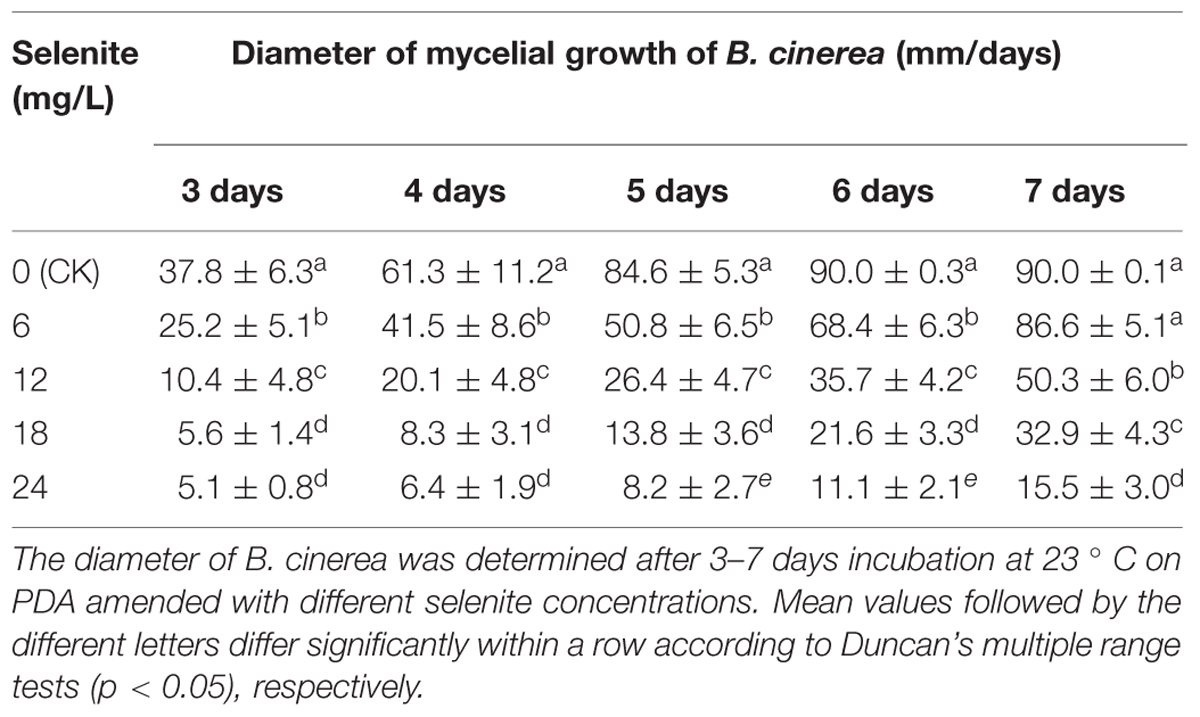
TABLE 2. Radial growth inhibition of Botrytis cinerea by Se salts in vitro.
Generation of Reactive Oxygen Species
Reactive oxygen species generation was monitored when B. cinerea was exposed to 24 mg/L selenite for 1, 2, and 3 h at 23°C (Figures 1A,B). After 1 h of incubation, a majority of spores in the control treatment were not stained by DCHF-DA, implying poor ROS production at the time. However, in the selenite treatment about 32% of spores were stained by DCHF-DA (Figure 1B). With an increased incubation period, even higher levels of ROS were detected in Se-treated B. cinerea spores compared with the control. The highest percentage of stained spores (57%) was observed following 3 h of incubation with 24 mg Se/L, indicating that an increasing amount of oxidizing molecules was produced in cells exposed to selenite (Figures 1A,B).
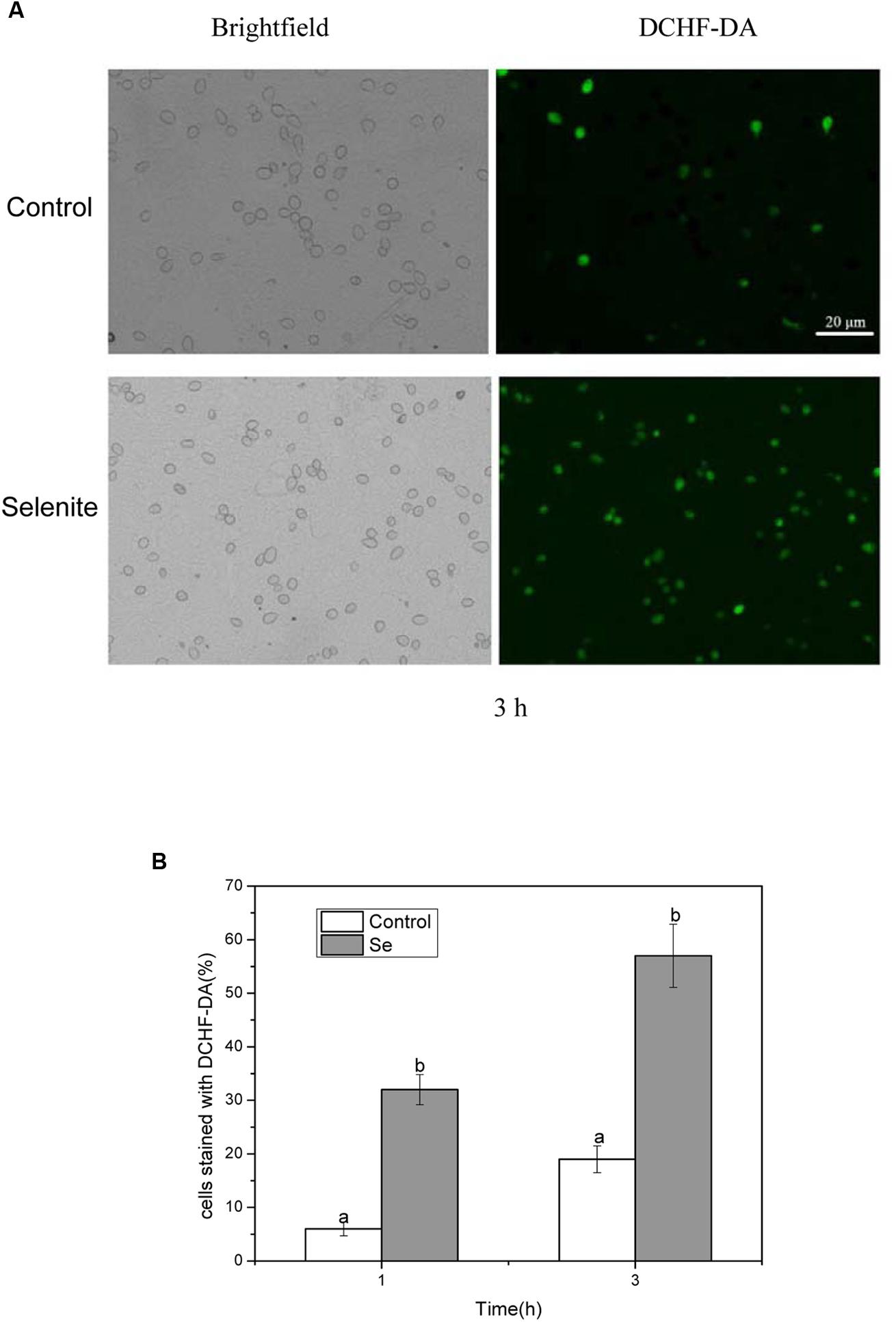
FIGURE 1. (A) Effect of selenite at 24 mg/L on production of reactive oxygen species in spores of Botrytis cinerea. (B) The percentage of spores stained with DCHF-DA. Spores incubated for 1 and 3 h. Vertical bars indicate standard deviations. Columns followed by different letters are statistically different according to the Duncan’s multiple range test (p < 0.05).
Assessment of Plasma Membrane Integrity by PI Staining
The plasma membranes integrity was assessed by PI staining when B. cinerea was exposed to 24 mg/L selenite for 1, 3, and 5 h at 23°C (Figures 2A,B). After 1 h of incubation, the plasma membranes of B. cinerea were damaged under selenite stress (p < 0.05; Figures 2A,B). Membrane integrity of B. cinerea spores markedly declined with increasing incubation period in PDB containing selenite compared with the control (Figures 2A,B). Most (≥90%) untreated spores appeared to have intact plasma membranes, whereas the membrane integrity of spores decreased from 63% to 42% following 3 to 5 h of incubation with 24 mg Se/L (Figures 2A,B).
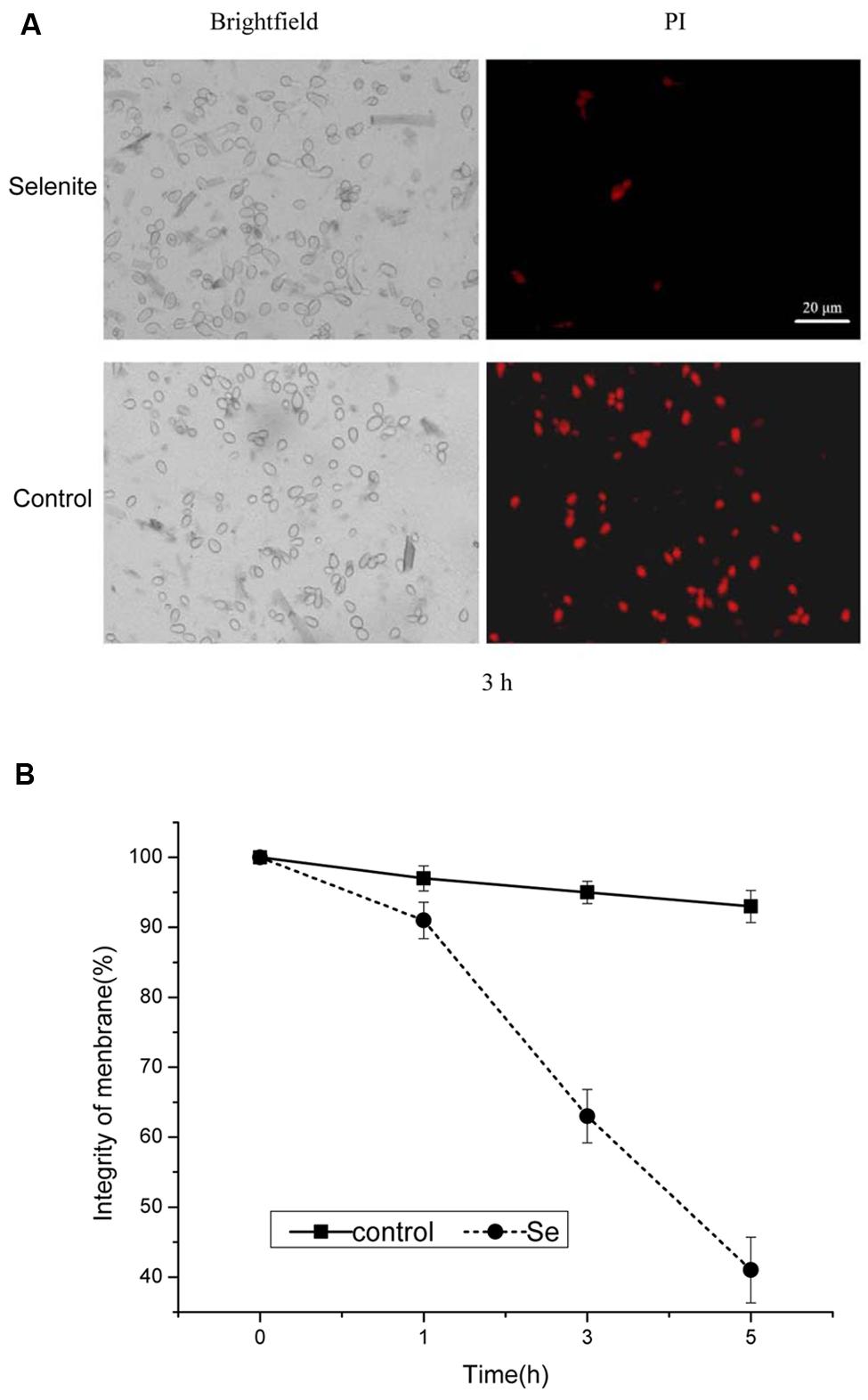
FIGURE 2. (A) Effect of selenite at 24 mg/L on plasma membrane integrity of B. cinerea conidia. (B) Percentage of plasma membrane integrity of B. cinerea spores. Spores incubated for 1, 3, and 5 h. Vertical bars indicate standard deviations. Columns followed by different letters are statistically different according to the Duncan’s multiple range test (p < 0.05).
Evaluation of Cellular Leakage
The leakage of proteins and carbohydrates were significantly increased under selenite at 24 mg/L stress compared with the control (Figures 3A,B). These results suggests that loss of membrane integrity would lead to the release of cytoplasmic contents from the cells. The production of soluble protein and soluble carbohydrates were increased significantly when the damage of cell membrane integrity caused by the leakage of cellular constituents.
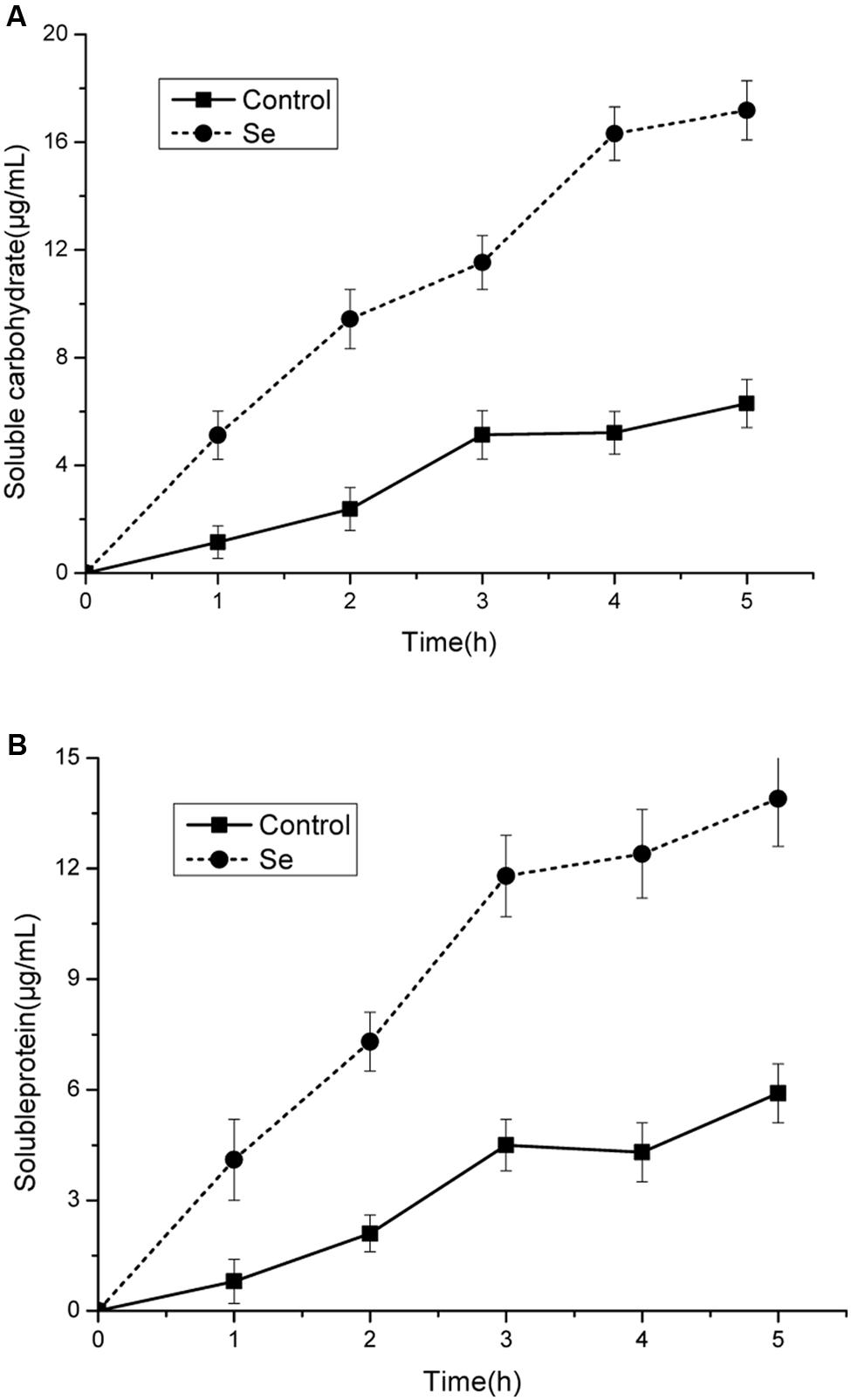
FIGURE 3. Effect of selenite at 24 mg/L on leakage of carbohydrates (A) and protein (B) of B. cinerea mycelia cultured in sterile double-distilled water at 23°C. Values for both soluble proteins and carbohydrates were expressed as μg/mL of aqueous solutions. Vertical bars indicate standard deviations.
Changes of SOD Activities, H2O2, MDA, GSH content, and O2- Production Rate
As shown in Figures 4A,B, both SOD activities and GSH content presented the similar decreasing trend from 1 to 3 days under selenite at 24 mg/L stress compared with the control. Moreover, upon exposure to selenite at 24 mg/L, the values of SOD and GSH were significantly lower than those of the control at each time point, and the decrease of SOD activity and GSH content was more fast than the control. Minimum values for SOD activity were 3.8 U/mg protein and 4.6 μg/g GSH content.
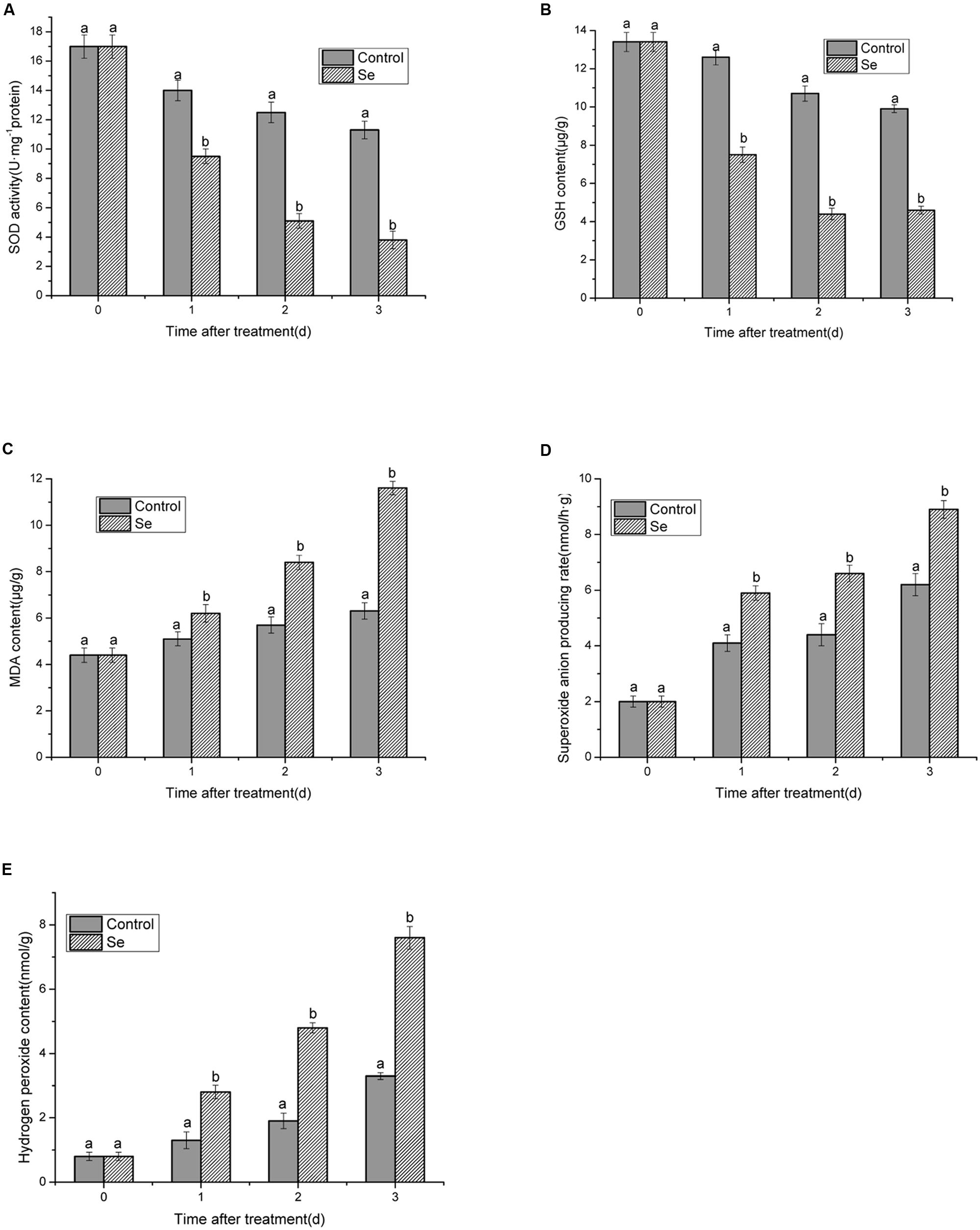
FIGURE 4. Effects of selenite at 24 mg/L on activities of SOD (A), GSH, and MDA content (B,C), O2- and H2O2 production rate (D,E) of B. cinerea spores after 1, 2, and 3 days of incubation at 23°C. Vertical bars indicate standard deviations. Columns followed by different letters are statistically different according to the Duncan’s multiple range test (p < 0.05).
The MDA content significantly increased under selenite at 24 mg/L stress compared with the control, especially after 3 days with a maximum of 11.6 μg/g (Figure 4C). H2O2 content, and O2- production rate of the control still increased, however, the production rate of them was clearly lower than that observed with selenite at 24 mg/L treatment. Rates increased to 8.9 nmol/g and 7.6 nmol/g for H2O2 and O2-, respectively (Figures 4D,E).
Effect of Se on Gray Mold Disease on Tomato Fruit
Selenite at 24 mg/L was effective in inhibiting the lesion diameter of postharvest gray mold of tomato fruit for 6 days storage at 23°C. (p < 0.05). After inoculation of tomato fruit for 2 days, the control fruits started to decay, while the Se-treatment group did not show any disease symptoms. Compared with 100% disease incidence in the control, the disease incidence in the Se-treatment fruits was less than 80% (Figure 5A). After inoculation for 3 and 6 days, the lesion diameters were only 8 and 24 mm in the Se-treatment group, while the control group reached 14 and 34 mm, respectively (Figures 5B,C). These results indicated that sodium selenite was effective for postharvest gray mold control in tomato fruit (at the concentration of 24 mg Se/L).
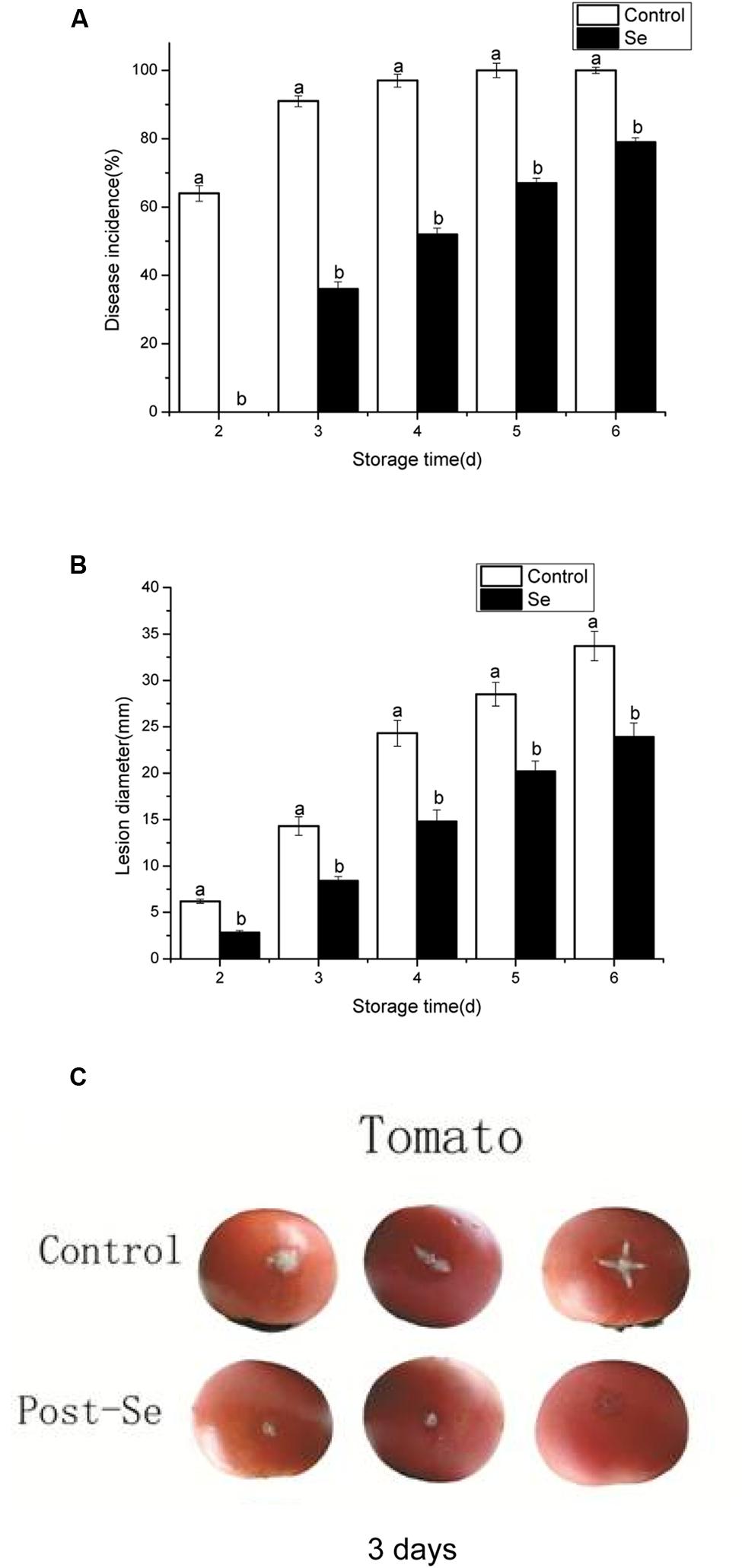
FIGURE 5. Effect of selenite at 24 mg/L on disease incidence (A) and lesion diameter (B) of gray mold on tomato fruit cause by B. cinerea after 1–6 days incubation at 23°C. Symptoms in inoculated tomato fruit stored in 23°C for 3 days (C). Vertical bars indicate standard deviations. Mean values present in each bar followed by different letters are statistically different according to the Duncan’s multiple range test (p < 0.05).
Comparison of the Inhibition Effects of Se with Several Inorganic Salts on B. cinerea
As shown in Table 3, Se could inhibit B. cinerea at minimum concentrations (e.g., Se at 0.012 g/L), as reported several inorganic salts such as silicate and borate, some of which have antimicrobial properties and could be useful as postharvest or preharvest treatments for gray mold disease control in the literature (Franck et al., 2005; Huang et al., 2008; Qin et al., 2010; Cabot et al., 2013; Harren and Tudzynski, 2013). This finding suggests that sodium selenite might serve as a potential alternative to synthetic fungicides for the reduction of the postharvest disease of fruit and vegetables caused by B. cinerea.

TABLE 3. Comparison of inhibitory effects of several similar inorganic salts on Botrytis cinerea.
Discussion
Mineral nutrients increase plant growth and development while influencing plant health. Specific elements such as Si and B have reportedly affected many plant diseases (Fauteux et al., 2005; Datnoff et al., 2007; Qin et al., 2010). Moreover, some inorganic and organic salts could be useful as postharvest or/and pre-harvest treatments for postharvest diseases control (Hervieux et al., 2002; Shi et al., 2011; Yaganza et al., 2014). Among of these salts, silicon (Si), and Se are widely studied trace elements and have been found to be protective for plants under abiotic and biotic stress conditions (Fauteux et al., 2005; Ma and Yamaji, 2006; Liang et al., 2007; Hasanuzzaman et al., 2014; Rahman et al., 2015). Both Si and Se were reported to play roles in conferring oxidative stress tolerance by enhancement of the antioxidant defense system in plants. Si is the second most abundant element and its presence in the form of silicic acid allows its uptake by plants, however, those plants not supplied with sufficient natural sources of Si may benefit from its exogenous application (Fauteux et al., 2005; Ma and Yamaji, 2006; Liang et al., 2007; Hasanuzzaman et al., 2014; Rahman et al., 2015). Se, an essential element for animals and humans, has also been found to be beneficial to plants, which also plays a protective role in conferring tolerance to certain abiotic and biotic stresses when applied at lower concentrations, while higher concentrations show phytotoxicity (Zhao and McGrath, 2009; Zhu et al., 2009; Hasanuzzaman et al., 2010; Wu et al., 2014).
Recently, some studies demonstrate that Se has important physiological functions for plant growth and quality improvement in fruit at low concentrations (Zhao and McGrath, 2009; Zhu et al., 2009; Hasanuzzaman et al., 2010; Wu et al., 2015). In recent years, based on the elemental defense hypothesis in relation to pathogens, Se has been shown to exert a positive effect on stress tolerance and used to control plant diseases in some crops (Hanson et al., 2003; Quinn et al., 2010; Companioni et al., 2012; Hladun et al., 2013). Hanson et al. (2003) showed Se accumulation could protect plants against invertebrate herbivory damage and fungal infection. Se containing substances in leaves and stems of alfalfa and cruciferous crops act as repellents, so that aphids, caterpillars, and spider mites avoid these plants (Galeas et al., 2008; Quinn et al., 2010). Even in small amounts these substances are also toxic for these pests (Freeman et al., 2007, 2009, 2012; Galeas et al., 2008; Hladun et al., 2013). Moreover, Leaves of plant supplied with Se were less susceptible to both Alternaria brassicicola and a Fusarium sp. (Hanson et al., 2003). From experiments with Indian mustard (Brassica juncea) it appears that application of Se can enhance resistance to fungal diseases (Hanson et al., 2003). Se is not commonly added to nutrient solutions in tomato, but in a Mexican study (Companioni et al., 2012) application of sodium selenite reduced damage inflicted by Fusarium wilt. Additionally, total protein content and antioxidant activity increased in both susceptible and resistant cultivars of tomato.
In previous studies, we found that germination and germ tube elongation of P. expansum were significantly inhibited by the addition of 20 mg/L selenite to the PDB. In addition, we found that the mechanisms involved damage of Se to cellular oxygen-eliminating system (Wu et al., 2014). In the current study, we found that selenite significantly inhibited spore germination and germ tube elongation of B. cinerea (Table 1) and reduced the lesion diameter of gray mold disease in harvested tomato fruit after artificially inoculating pathogen spores (Figure 5). From these results we could conclude that selenite has a potential as a promising antifungal agent and also as a potential alternatives to synthetic fungicides in the protection of tomato fruit against B. cinerea, and selenite has a broadly inhibiting effect on postharvest fungal pathogens. The mechanism of action appeared to be closely related to the generation of active oxygen. ROS may cause oxidative damage to cellular compounds and lead to cellular dysfunction or cell death (Shi et al., 2012). In this study, we used an oxidant-sensitive probe DCHF-DA to investigate the ROS accumulation in B. cinerea spores, and observed that Se-treated spores showed a higher ratio of stained cells than that in control (Figures 1 and 2), indicating that Se induced ROS accumulation in B. cinerea spores. ROS accumulation can cause oxidative damage of the fungal spore and result in lower spore germination.
In addition, understanding the mode of action by which sodium selenite inhibits B. cinerea will help in the application of sodium selenite for management of pre- or/and postharvest gray mold disease of fruits and vegetables. Therefore, further studies are needed on the molecular mechanism of sodium selenite against B. cinerea by the “Omics approaches” (e.g., proteomics, metabolomics, genomics, transcriptomics), the effect of Se on nutrition and quality of tomato fruit, using confocal microscopy and TEM to confirm if selenite treatment could lead to mitochondrial damage, and the development of unique valued-added Se-enriched nutraceutical agricultural products from Se-biofortified fruits and vegetables, etc. Specifically, only sodium selenite and B. cinerea was tested in the study. Much more plant pathogenic fungal and bacterial species need to be tested to investigate if the antimicrobial effect of Se compounds such as sodium selenite and/or sodium selenate at low concentration against fungal and/or bacterial species is specific. Moreover, Se is one of essential micronutrients for human and animals health but at high concentrations, Se becomes toxic to vertebrates, especially birds, such that widespread use of this antimicrobial agent needs to be evaluated in the context of potential ecological impacts.
Conclusion
In this study, sodium selenite at a concentration of 24 mg Se/L was effective in controlling gray mold in tomato fruits caused by B. cinerea. Moreover, we conclude that selenite treatment can stimulate ROS accumulation in fungal spores, resulting in oxidative damage, which may act as the antifungal mechanism of selenite inhibiting spore germination of B. cinerea and controlling gray mold in tomato fruits. In summary, we have shown that selenite could directly inhibit the growth of B. cinerea in vitro. Moreover, Se resulted in oxidative damage, which then played an important role in the inhibitory effect on B. cinerea. Additionally, application of selenite is promising as an alternative to synthetic fungicides for postharvest gray mold disease control in tomato fruit.
Author Contributions
ZW and ML wrote the main manuscript text, and ZW and LY prepared all figures and tables. GB and Z-qL revised the manuscript. XY, YL, and ZZ have provided input and assistance to the submission of the final manuscript. All authors reviewed the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant 31400091, 31401545), the Natural Science Foundation of Anhui province (Grant 1408085MC68), Technology Innovation Fund by Institute of Advanced Technology, USTC (Grant 2013-1-16) the Natural Science Foundation of Jiangsu Province (Grant BK2012195 and BK2012202), and China Postdoctoral Science Foundation (Grant 20100470108 and 2013M540518).
References
Ahn, I. P., Kim, S., and Lee, Y. H. (2005). Vitamin B1 functions as an activator of plant disease resistance. Plant Physiol. 138, 1505–1515. doi: 10.1104/pp.104.058693
Arslan, U., Ilhan, K., Vardar, C., and Karabulut, O. A. (2009). Evaluation of antifungal activity of food additives against soilborne phytopathogenic fungi. World J. Microbiol Biotechnol. 25, 537–543. doi: 10.1007/s11274-008-9921-1
Bañuelos, G. S. (2006). Phyto-products may be essential for sustainability and implementation of phytoremediation. Environ. Pollu. 144, 19–23. doi: 10.1016/j.envpol.2006.01.015
Boyd, R. S. (2007). The defense hypothesis of elemental hyperaccumulation: status, challenges and new directions. Plant Soil. 293, 153–176. doi: 10.1007/s11104-007-9240-6
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Brotherton, J. (1967). The sulphur metabolism of Pityrosporum ovale and its inhibition by selenium compounds. Microbiology 49, 393–400.
Cabot, C., Gallego, B., Martos, S., Barceló, J., and Poschenrieder, C. (2013). Signal cross talk in Arabidopsis exposed to cadmium, silicon, and Botrytis cinerea. Planta 237, 337–349. doi: 10.1007/s00425-012-1779-7
Companioni, B., Medrano, J., Torres, J. A., Flores, A., Rodríguez, E., and Benavides, A. (2012). “Protective action of sodium selenite against Fusarium wilt in tomato: total protein contents, levels of phenolic compounds and changes in antioxidant potential,” in Proceedings of the II International Symposium on Soilless Culture and Hydroponics, Acta Horticulturae 947, eds L. I. Trejo-Téllez, F. C. Gómez-Merino and M. N. Rodriguez-Mendoza, (Mexico city: International Society for Horticultural Science), 321–328.
Crane, J. M., Frodyma, M. E., and Bergstrom, G. C. (2014). Nutrient-induced spore germination of a Bacillus amyloliquefaciens biocontrol agent on wheat spikes. J. Appl. Microbiol. 116, 1572–1583. doi: 10.1111/jam.12480
Datnoff, L. E., Elmer, W. H., and Huber, D. M. (eds). (2007). Mineral Nutrition and Plant Disease. (St. Paul, MN: APS Press), 1–25.
de Souza, E. L., Sales, C. V., de Oliveira, C. E., Lopes, L. A., da Conceição, M. L., Berger, L. R., et al. (2015). Efficacy of a coating composed of chitosan from Mucor circinelloides and carvacrol to control Aspergillus flavus and the quality of cherry tomato fruits. Front. Microbiol. 6:732. doi: 10.3389/fmicb.2015.00732
Dordas, C. (2008). Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron Sustain Dev. 28, 33–46. doi: 10.1051/agro:2007051
Doucha, J., Lívanský, K., Kotrbácek, V., and Zachleder, V. (2009). Production of Chlorella biomass enriched by selenium and its use in animal nutrition: a review. Appl. Microbiol. Biotechnol. 83, 1001–1008. doi: 10.1007/s00253-009-2058-9
Droby, S., Wisniewskib, M., Macarisinb, D., and Wilsonc, C. (2009). Twenty years of postharvest biocontrol research: is it time for a new paradigm? Postharvest Biol. Technol. 52, 137–145. doi: 10.1016/j.postharvbio.2008.11.009
Fauteux, F., Rémus-Borel, W., Menzies, J. G., and Bélanger, R. R. (2005). Silicon and plant disease resistance against pathogenic fungi. FEMS Microbiol. Lett. 249, 1–6. doi: 10.1016/j.femsle.2005.06.034
Fones, H., Davis, C. A., Rico, A., Fang, F., Smith, J. A., and Preston, G. M. (2010). Metal hyperaccumulation armors plants against disease. PLoS Pathog. 6:e1001093. doi: 10.1371/journal.ppat.1001093
Fones, H., and Preston, G. M. (2013). The impact of transition metals on bacterial plant disease. FEMS Microbiol. Rev. 37, 495–519. doi: 10.1111/1574-6976.12004
Franck, J., Latorre, B. A., Torres, R., and Zoffoli, J. P. (2005). The effect of preharvest fungicide and postharvest sulfur dioxide use on postharvest decay of table grapes caused by Penicillium expansum. Postharvest Biol. Technol. 37, 20–30. doi: 10.1016/j.postharvbio.2005.02.011
Freeman, J. L., Lindblom, S. D., Quinn, C. F., Fakra, S., Marcus, M. A., and Pilon-Smits, E. A. (2007). Selenium accumulation protects plants from herbivory by Orthoptera via toxicity and deterrence. New Phytol. 175, 490–500. doi: 10.1111/j.1469-8137.2007.02119.x
Freeman, J. L., Marcus, M. A., Fakra, S. C., Devonshire, J., McGrath, S. P., Quinn, C. F., et al. (2012). Selenium hyperaccumulator plants Stanleya pinnata and Astragalus bisulcatus are colonized by Se-resistant, Se-excluding wasp and beetle seed herbivores. PLoS ONE 7:e50516. doi: 10.1371/journal.pone.0050516
Freeman, J. L., Quinn, C. F., Lindblom, S. D., Klamper, E. M., and Pilon-Smits, E. A. (2009). Selenium protects the hyperaccumulator Stanleya pinnata against black-tailed prairie dog herbivory in native seleniferous habitats. Am. J. Bot. 96, 1075–1085. doi: 10.3732/ajb.0800287
Gabler, F. M., Smilanick, J. L., Ghosoph, J. M., and Margosan, D. A. (2005). Impact of postharvest hot water or ethanol treatment of table grapes on gray mold incidence, quality, and ethanol content. Plant Dis. 89, 309–316. doi: 10.1094/PD-89-0309
Galeas, M. L., Klamper, E. M., Bennett, L. E., Freeman, J. L., Kondratieff, B. C., Quinn, C. F., et al. (2008). Selenium hyperaccumulation reduces plant arthropod loads in the field. New Phytol. 177, 715–724. doi: 10.1111/j.1469-8137.2007.02285.x
Hanson, B., Garifullina, G. F., Lindblom, S. D., Wangeline, A., Ackley, A., Kramer, K., et al. (2003). Selenium accumulation protects Brassica juncea from invertebrate herbivory and fungal infection. New Phytol. 159, 461–469. doi: 10.1046/j.1469-8137.2003.00786.x
Harren, K., and Tudzynski, B. (2013). Cch1 and Mid1 are functionally required for vegetative growth under low-calcium conditions in the phytopathogenic ascomycete Botrytis cinerea. Eukaryot. Cell 12, 712–724. doi: 10.1128/EC.00338-12
Hasanuzzaman, M., Hossain, M. A., and Fujita, M. (2010). Selenium in higher plants: physiological role, antioxidant metabolism and abiotic stress tolerance. J. Plant Sci. 5, 354–375. doi: 10.3923/jps.2010.354.375
Hasanuzzaman, M., Nahar, K., and Fujita, M. (2014). Silicon and Selenium: two vital trace elements that confer abiotic stress tolerance to plants. emerging technologies and management of crop stress tolerance, Chapter 16. Biol. Techniq. 1, 377–422.
Hervieux, V., Yaganza, E. S., Arul, J., and Tweddell, R. J. (2002). Effect of organic and inorganic salts on the development of Helminthosporium solani, the causal agent of potato silver scurf. Plant Dis. 86, 1014–1018. doi: 10.1094/PDIS.2002.86.9.1014
Hladun, K. R., Kaftanoglu, O., Parker, D. R., Tran, K. D., and Trumble, J. T. (2013). Effects of selenium on development, survival, and accumulation in the honeybee (Apis mellifera L.). Environ. Toxicol. Chem. 32, 2584–2592. doi: 10.1002/etc.2357
Hörger, A. C., Fones, H. N., and Preston, G. M. (2013). The current status of the elemental defense hypothesis in relation to pathogens. Front. Plant Sci. 4:395. doi: 10.3389/fpls.2013.00395
Huang, F., Wang, J. M., and Xu, Y. M. (2008). Primary study on mechanism of borate inhibiting spore germination of Botrytis cinerea. Acta Phytopathol. Sin. 38, 370–376.
Janisiewicz, W. J., and Korsten, L. (2002). Biological control of postharvest diseases of fruits. Annu. Rev. Phytopathol. 40, 411–441. doi: 10.1146/annurev.phyto.40.120401.130158
Karabulut, O. A., Arslan, U., Ilhan, K., and Kuruoglu, G. (2005). Integrated control of postharvest diseases of sweet cherry with yeast antagonists and sodium bicarbonate applications within a hydrocooler. Postharvest. Biol. Technol. 37, 135–141. doi: 10.1016/j.postharvbio.2005.03.003
Karabulut, O. A., Gabler, F. M., Mansour, M., and Smilanick, J. L. (2004). Postharvest ethanol and hot water treatments of table grapes to control gray mold. Postharvest. Biol Technol. 34, 169–177.
Lai, T. F., Li, B. Q., Qin, G. Z., and Tian, S. P. (2011). Oxidative damage involves in the inhibitory effect of nitric oxide on spore germination of Penicillium expansum. Curr. Microbiol. 62, 229–234. doi: 10.1007/s00284-010-9695-1
Liang, Y., Sun, W., Zhu, Y. G., and Christie, P. (2007). Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: a review. Environ. Pollut. 147, 422–428. doi: 10.1016/j.envpol.2006.06.008
Liu, H., Jiang, W. B., Bi, Y., and Luo, Y. B. (2005). Postharvest BTH treatment induces resistance of peach (Prunus persica L. cv. Jiubao) fruit to infection by Penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest. Biol. Technol. 35, 263–269. doi: 10.1016/j.postharvbio.2004.08.006
Ma, J. F., and Yamaji, N. (2006). Silicon uptake and accumulation in higher plants. Trends Plant Sci. 11, 392–397. doi: 10.1016/j.tplants.2006.06.007
Ma, Y., Liu, R. H., Gong, X. Y., Li, Z., Huang, Q. H., Wang, H. S., et al. (2006). Synthesis and herbicidal activity of, N., N-diethyl-3-(arylselenonyl)-1 H-1, 2, 4-triazole-1-carboxamide. J. Agricul. Food Chem. 54, 7724–7728. doi: 10.1021/jf0609328
Qin, G. Z., Zong, Y. Y., Chen, Q. L., Hua, D. L., and Tian, S. P. (2010). Inhibitory effect of boron against Botrytis cinerea on table grapes and its possible mechanisms of action. Int. J. Food Microbiol. 138, 145–150. doi: 10.1016/j.ijfoodmicro.2009.12.018
Quinn, C. F., Freeman, J. L., Reynolds, R. J., Cappa, J. J., Fakra, S. C., Marcus, M. A., et al. (2010). Selenium hyperaccumulation offers protection from cell disruptor herbivores. BMC Ecol. 10:19. doi: 10.1186/1472-6785-10-19
Rahman, A., Wallis, C. M., and Uddin, W. (2015). Silicon-induced systemic defense responses in perennial ryegrass against infection by Magnaporthe oryzae. Phytopathology 105, 748–757. doi: 10.1094/PHYTO-12-14-0378-R
Razak, A. A., El-Tantawy, H., El-Sheikh, H. H., and Gharieb, M. M. (1991). Influence of selenium on the efficiency of fungicide action against certain fungi. Biol. Trace Element Res. 28, 47–56. doi: 10.1007/BF02990462
Shi, X. Q., Li, B. Q., Qin, G. Z., and Tian, S. P. (2011). Antifungal activity and possible mode of action of borate against Colletotrichum gloeosporioides on Mango. Plant Dis. 95, 63–69. doi: 10.1094/PDIS-06-10-0437
Shi, X. Q., Li, B. Q., Qin, G. Z., and Tian, S. P. (2012). Mechanism of antifungal action of borate against Colletotrichum gloeosporioides related to mitochondrial degradation in spores. Postharvest. Biol. Technol. 67, 138–143. doi: 10.1016/j.postharvbio.2012.01.003
Singh, P. K. (2012). Synthesis and fungicidal activity of novel 3-(substituted/unsubstituted phenylselenonyl)-1-ribosyl/deoxyribosyl-1 H-1, 2, 4-triazole. J. Agricul. Food Chem. 60, 5813–5818. doi: 10.1021/jf300730f
Soylu, E. M., Kurt, S., and Soylu, S. (2010). In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 143, 183–189. doi: 10.1016/j.ijfoodmicro.2010.08.015
Tapiero, H., Townsend, D., and Tew, K. (2003). The antioxidant role of selenium and seleno-compounds. Biomed. Pharmacother. 57, 134–144. doi: 10.1016/S0753-3322(03)00035-0
Trumble, J., and Sorensen, M. (2008). Selenium and the elemental defense hypothesis. New Phytol. 177, 569–572. doi: 10.1111/j.1469-8137.2008.02353.x
Winkel, L. H. E., Johnson, C. A., Lenz, M., Grundl, T., Leupin, O. X., Amini, M., et al. (2011). Environmental selenium research: from microscopic processes to global understanding. Environ. Sci. Technol. 46, 571–579. doi: 10.1021/es203434d
Wu, Z., Bañuelos, G. S., Lin, Z. Q., Liu, Y., Yuan, L., Yin, X., et al. (2015). Biofortification and phytoremediation of selenium in China. Front. Plant Sci. 6:136. doi: 10.3389/fpls.2015.00136
Wu, Z. L., Yin, X. B., Lin, Z. Q., Bañuelos, G. S., Yuan, L. X., Liu, Y., et al. (2014). Inhibitory effect of selenium against Penicillium expansum and its possible mechanisms of action. Curr. Microbiol. 69, 192–201. doi: 10.1007/s00284-014-0573-0
Xu, S., Li, J. L., Zhang, X. Q., Wei, H., and Cui, L. J. (2006). Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot. 56, 274–285. doi: 10.1016/j.envexpbot.2005.03.002
Yaganza, E. S., Tweddell, R. J., and Arul, J. (2014). Postharvest application of organic and inorganic salts to control potato (Solanum tuberosum L.) storage soft rot: plant tissue–salt physicochemical interactions. J. Agric. Food Chem. 62, 9223–9231. doi: 10.1021/jf5017863
Yao, Y. A., Wang, J., Ma, X., Lutts, S., Sun, C., Ma, J., et al. (2012). Proteomic analysis of Mn-induced resistance to powdery mildew in grapevine. J. Exp. Bot. 63, 5155–5170. doi: 10.1093/jxb/ers175
Youssef, K., and Roberto, S. R. (2014). Salt strategies to control Botrytis mold of ‘Benitaka’ table grapes and to maintain fruit quality during storage. Postharvest. Biol. Technol. 95, 95–102. doi: 10.1016/j.postharvbio.2013.08.011
Yuan, L., Zhu, Y., Lin, Z. Q., Banuelos, G., Li, W., and Yin, X. (2013). A novel selenocystine-accumulating plant in selenium-mine drainage area in Enshi, China. PLoS ONE 8:e65615. doi: 10.1371/journal.pone.0065615
Zhang, H., Ge, L., Chen, K., Zhao, L., and Zhang, X. (2014). Enhanced biocontrol activity of Rhodotorula mucilaginosa cultured in media containing chitosan against postharvest diseases in strawberries: possible mechanisms underlying the effect. J. Agric. Food Chem. 62, 4214–4224. doi: 10.1021/jf500065n
Zhao, F. J., and McGrath, S. P. (2009). Biofortification and phytoremediation. Curr. Opin. Plant Biol. 12, 373–380. doi: 10.1016/j.pbi.2009.04.005
Zhu, Y. G., Pilon-Smits, E. A., Zhao, F. J., Williams, P. N., and Meharg, A. A. (2009). Selenium in higher plants: understanding mechanisms for biofortification and phytoremediation. Trends Plant Sci. 14, 436–442. doi: 10.1016/j.tplants.2009.06.006
Keywords: Botrytis cinerea, selenium, postharvest treatment, reactive oxygen species, membrane integrity, cellular leakage
Citation: Wu Z, Yin X, Bañuelos GS, Lin Z-Q, Zhu Z, Liu Y, Yuan L and Li M (2016) Effect of Selenium on Control of Postharvest Gray Mold of Tomato Fruit and the Possible Mechanisms Involved. Front. Microbiol. 6:1441. doi: 10.3389/fmicb.2015.01441
Received: 25 July 2015; Accepted: 03 December 2015;
Published: 06 January 2016.
Edited by:
Sahdeo Prasad, The University of Texas MD Anderson Cancer Center, USAReviewed by:
Maria Gougouli, Aristotle University of Thessaloniki, GreeceParag A. Parekh, MD Anderson Cancer Center, USA
Shivangi Agarwal, Northwestern University, USA
Copyright © 2016 Wu, Yin, Bañuelos, Lin, Zhu, Liu, Yuan and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Linxi Yuan, yuanli@ustc.edu.cn; Miao Li, miaoli@ustc.edu.cn