 Warren Albertin1,2*
Warren Albertin1,2* Mathabatha E. Setati3
Mathabatha E. Setati3 Cécile Miot-Sertier1,4
Cécile Miot-Sertier1,4 Talitha T. Mostert3
Talitha T. Mostert3 Benoit Colonna-Ceccaldi5
Benoit Colonna-Ceccaldi5 Joana Coulon6Patrick Girard5Virginie Moine6Myriam Pillet7Franck Salin7
Joana Coulon6Patrick Girard5Virginie Moine6Myriam Pillet7Franck Salin7 Marina Bely1
Marina Bely1 Benoit Divol3
Benoit Divol3 Isabelle Masneuf-Pomarede1,8
Isabelle Masneuf-Pomarede1,8- 1Unité de recherche Œnologie, Institut de la Science de la Vigne et du Vin, University Bordeaux, Villenave d'Ornon, France
- 2ENSCBP, Bordeaux INP, Pessac, France
- 3Department of Viticulture and Oenology, Institute for Wine Biotechnology, Stellenbosch University, Matieland, South Africa
- 4Institut National de la Recherche Agronomique, Institut de la Science de la Vigne et du Vin, USC 1366 Institut National de la Recherche Agronomique, Villenave d'Ornon, France
- 5Centre de Recherche Pernod Ricard, Créteil, France
- 6Biolaffort, Bordeaux, France
- 7Institut National de la Recherche Agronomique, UMR Biodiversité Gènes et Ecosystèmes, PlateForme Génomique, Cestas, France
- 8Bordeaux Sciences Agro, Gradignan, France
Hanseniaspora uvarum is one of the most abundant yeast species found on grapes and in grape must, at least before the onset of alcoholic fermentation (AF) which is usually performed by Saccharomyces species. The aim of this study was to characterize the genetic and phenotypic variability within the H. uvarum species. One hundred and fifteen strains isolated from winemaking environments in different geographical origins were analyzed using 11 microsatellite markers and a subset of 47 strains were analyzed by AFLP. H. uvarum isolates clustered mainly on the basis of their geographical localization as revealed by microsatellites. In addition, a strong clustering based on year of isolation was evidenced, indicating that the genetic diversity of H. uvarum isolates was related to both spatial and temporal variations. Conversely, clustering analysis based on AFLP data provided a different picture with groups showing no particular characteristics, but provided higher strain discrimination. This result indicated that AFLP approaches are inadequate to establish the genetic relationship between individuals, but allowed good strain discrimination. At the phenotypic level, several extracellular enzymatic activities of enological relevance (pectinase, chitinase, protease, β-glucosidase) were measured but showed low diversity. The impact of environmental factors of enological interest (temperature, anaerobia, and copper addition) on growth was also assessed and showed poor variation. Altogether, this work provided both new analytical tool (microsatellites) and new insights into the genetic and phenotypic diversity of H. uvarum, a yeast species that has previously been identified as a potential candidate for co-inoculation in grape must, but whose intraspecific variability had never been fully assessed.
Introduction
Hanseniaspora uvarum (anamorph Kloeckera apiculata) is an apiculate yeast species frequently found on mature fruits (Spencer et al., 1992; Morais et al., 1995) and particularly on grapes where it forms part of the grape and fermentation microbiome.Its association with grapes and the first stages of alcoholic fermentation (AF) has been reported repeatedly during the last century (Castelli, 1955; Schütz and Gafner, 1993; Hierro et al., 2006) and for most—if not all—vineyard regions worldwide (Heard and Fleet, 1985; Holloway et al., 1990; Mateo et al., 1991; Comi et al., 2001; Beltran et al., 2002; Jolly et al., 2003; Combina et al., 2005; Li et al., 2010; Zott et al., 2010; Kachalkin et al., 2015). H. uvarum is also frequently isolated from other fermented beverages such as cider (Lachance, 1995; Cabranes et al., 1997; Valles et al., 2007; Pando Bedrinana et al., 2012), palm wine and cashew juice (Owuama and Saunders, 1990), tequila (Bilbao et al., 1997), sugar-cane aguardente (Morais et al., 1997), etc. It is part of the natural microbiome of many fermented food processes, including coffee (Masoud et al., 2004) and cocoa (Batista et al., 2015) fermentations. In some biotechnological processes such as yogurt (Kosse et al., 1997), orange juice (Renard et al., 2008), beer (Wiles, 1950), and honey (Pulvirenti et al., 2009) production, H. uvarum is considered as a spoilage species. H. uvarum also displays industrially relevant antagonistic properties against the development of molds responsible for fruit spoilage. The species is thus extensively assessed as a biocontrol agent against Botrytis cinerea (gray mold) on grapes and strawberries (Long et al., 2005; Liu et al., 2010a,b; Cai et al., 2015), Penicilium spp. (fruit rot) on citrus (Long et al., 2005), Colletotrichum capsici (fruit rot) on chili (Basha and Ramanujam, 2014), etc., while the underlying mechanisms of action are actively studied (Liu et al., 2014; Pu et al., 2014). The ecological extent of H. uvarum is large: it has been collected from soils (Capriotti, 1955), plants (Sláviková et al., 2009), insects (Nguyen et al., 2007), birds (Kocan and Hasenclever, 1972), molluscs (de Araujo et al., 1995), and shrimps (Pagnocca et al., 1989), while its occurrence as clinical isolate on humans is rare and considered as opportunistic (Emmanouil-Nikoloussi et al., 1994; Garcia-Martos et al., 1999).
In winemaking, the presence of indigenous apiculate yeasts has long been viewed as undesirable (Velázquez et al., 1991; Ciani, 1998; Comitini and Ciani, 2010), and methods or factors to limit their proliferation during AF have been described (Farías and Manca de Nadra, 2003; Sosa et al., 2008; Comitini and Ciani, 2010). However, the renewed interest in non-conventional yeasts in the wine industry has led to the reassessment of the species suitable—and beneficial—for winemaking purpose. Several studies report on the characterization of the outcome of AF by H. uvarum in mixed or sequential inoculation with Saccharomyces cerevisiae in grape must, as H. uvarum alone is not able to complete AF (i.e., to consume all the sugar contained in grape musts). Wines resulting from mixed or sequential inoculation of H. uvarum and S. cerevisiae were shown to differ from pure cultures (S. cerevisiae) in their chemical composition. Indeed, the concentrations of some organic acids, aldehydes and minor alcohols (Hong and Park, 2013), higher alcohols and volatile metabolites (Zironi et al., 1993; Zohre and Erten, 2002; Moreira et al., 2011), isoamyl acetate (Moreira et al., 2008), butanediol and acetoin (Romano et al., 1993, 2000), and a few other compounds were reported as significantly different. Some of these alterations could be associated with the secretion of extracellular enzymes. Indeed, several enzymatic activities of technological interest have been characterized, such as β-glucosidase, xylosidase, protease, and lipase activities (Charoenchai et al., 1997; Manzanares et al., 1999; Capece et al., 2005). Moreover, some strains of the H. uvarum species were shown to be low producers of ochratoxin A (OTA), the main mycotoxin found in wine (Angioni et al., 2007). For all these reasons, the ability of H. uvarum to be preserved by lyophilization and cryopreservation was assessed recently, and was found satisfactory enough to maintain its fermentation ability (de Arruda Moura Pietrowski et al., 2015).
The abiotic and biotic factors to which H. uvarum is exposed in grape must have also been investigated. The data showed that the growth of H. uvarum was significantly affected by temperature, pH, sulfite, and ethanol concentrations (Gao and Fleet, 1988; Heard and Fleet, 1988; Albertin et al., 2014b), with some of these factors having synergistic or buffering effects. Several authors reported the existence of interactions between H. uvarum and S. cerevisiae during AF (Mendoza et al., 2007; Wang et al., 2014), associated with various underlying mechanisms including production of killer toxin (Radler et al., 1985, 1990; Schmitt and Neuhausen, 1994), and release of yet unidentified metabolites (Wang et al., 2015).
However, most of these studies evaluated single strains of H. uvarum. Only a few authors considered several strains to account for potential diversity within the species (Comi et al., 2001; Capece et al., 2005), but even then, the genetic relationships between the different strains remained obscure due to the lack of dedicated tools. Indeed, the molecular approaches available to date allowed intraspecific discrimination, but not establishment of genetic distance: RAPD (randomly amplified polymorphic DNA, Capece et al., 2005) or restriction endonuclease analysis associated with pulse-field gel electrophoresis (REA-PFGE, Versavaud and Hallet, 1995) were described to discriminate H. uvarum strains. By contrast, PCR fingerprinting was not able to discriminate H. uvarum strains in Aglianico wines (Caruso et al., 2002). More recently, FT-IR (Fourier transform infrared spectroscopy) was successfully applied to the intraspecific discrimination of H. uvarum from grape berries and the winery environment (Grangeteau et al., 2015). However, none of these approaches allows the establishment of genetic relationships between the different isolates. Consequently, the extent of the diversity within the H. uvarum species remains uncharacterized.
In this study, 115 strains of H. uvarum were isolated from winemaking environments in France and South Africa. Their genetic variability was analyzed using two different approaches: microsatellite markers and AFLP (amplified fragment-length polymorphism). Their phenotypic diversity regarding enzymatic activities and response to environmental factors was also investigated.
Material and Methods
Yeast Strains
One hundred and eleven strains of Hanseniaspora spp., including mainly H. uvarum and a few Hanseniaspora guillermondii, were isolated from French and South African winemaking areas between 2003 and 2014 (Table 1). These strains were identified using molecular techniques like rDNA ITS analysis (Granchi et al., 1999), and sequencing of the D1/D2 domain of 26S rDNA (O'Donnell, 1993; Kurtzman and Robnett, 1998) or the ITS sequence (White et al., 1990; Esteve-Zarzoso et al., 1999). D1/D2 and ITS sequences were then blasted again either NCBI database or YeastIP, a curated yeast database (Weiss et al., 2013).
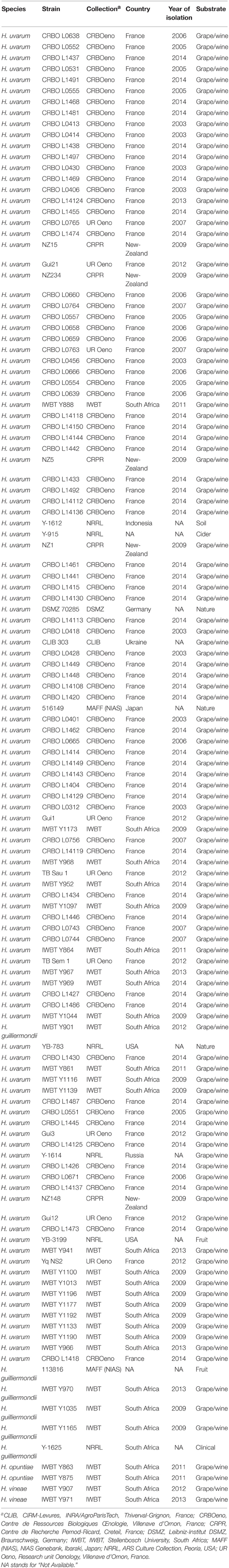
Table 1. Hanseniaspora sp. strains used in this study.
Fifteen strains from other geographical and substrate (nature, cider, etc.) origins were included (Table 1). For phenotypic characterization, several control strains were used: S. cerevisiae VIN13 (Mocke, 2005) was used as positive control for killer activity, S. cerevisiae ZIM 1859 S6 (Zagorc et al., 2001) was used as killer sensitive yeasts. Metschnikowia pulcherrima IWBT Y1123, Schwanniomyces polymorphus var. africanus CBS 8047, Saccharomyces paradoxus RO88 (Redzepovic et al., 2003), Metschnikowia chrysoperlae IWBT Y955 were used as positive controls for the acid protease, β-glucosidase, pectinase, and chitinase tests, respectively.
All strains were grown at 24°C in traditional YPD medium containing 1% yeast extract, 1% peptone, and 2% glucose (w/v), supplemented or not with 2% agar (w/v).
Microsatellite analysis was applied to all Hanseniapora spp. strains available, while only a subset of strains was used for both AFLP and phenotyping assays, more time-consuming and less reproducible over large number of experiments. For AFLP approach, 47 strains were selected, and for phenotyping data we used a subset of 30 strains (all included in the AFLP panel) as well as 10 other Hanseniaspora spp.
Genome Sequencing, Microsatellite Loci Identification, and Primers Design
A draft genomic sequence was produced using Ion Torrent technology. Briefly, a genomic library of strain CRBO L0551 was produced using the Ion Xpress Plus Fragment Library Kit (Life Technologies, Carlsbad, USA), with an enzymatic shearing of 10 min at 37°C. DNA was sequenced on an Ion Torrent PGM (Life Technologies, Carlsbad, CA). After trimming on quality threshold (Phred-type quality score of Q20, QPhred = 20) and length threshold (50 bp) using CLC GenomicsWorkbench 7.0.3 (CLC bio, Boston, MA), Newbler software (version 2.7, 454 Life Sciences) was used to produce a de novo assembly of 1665 contigs of more than 1000 bp. This draft assembly forms a 7.68 Mb sequence for an estimated genome size of 8–9 Mb (Esteve-Zarzoso et al., 2001).
Microsatellites (di- to tetranucleotide repeats) were searched within the de novo genome assembly as described previously (Albertin et al., 2014a), and primers were designed using the Design primers' tool on the SGD website (http://www.yeastgenome.org/cgi-bin/web-primer) by applying Schuelke's method (Schuelke, 2000) to reduce costs. Amplified fragment sizes varied from 120 to 466 bp, allowing subsequent multiplexing of the amplicons (Table 2).
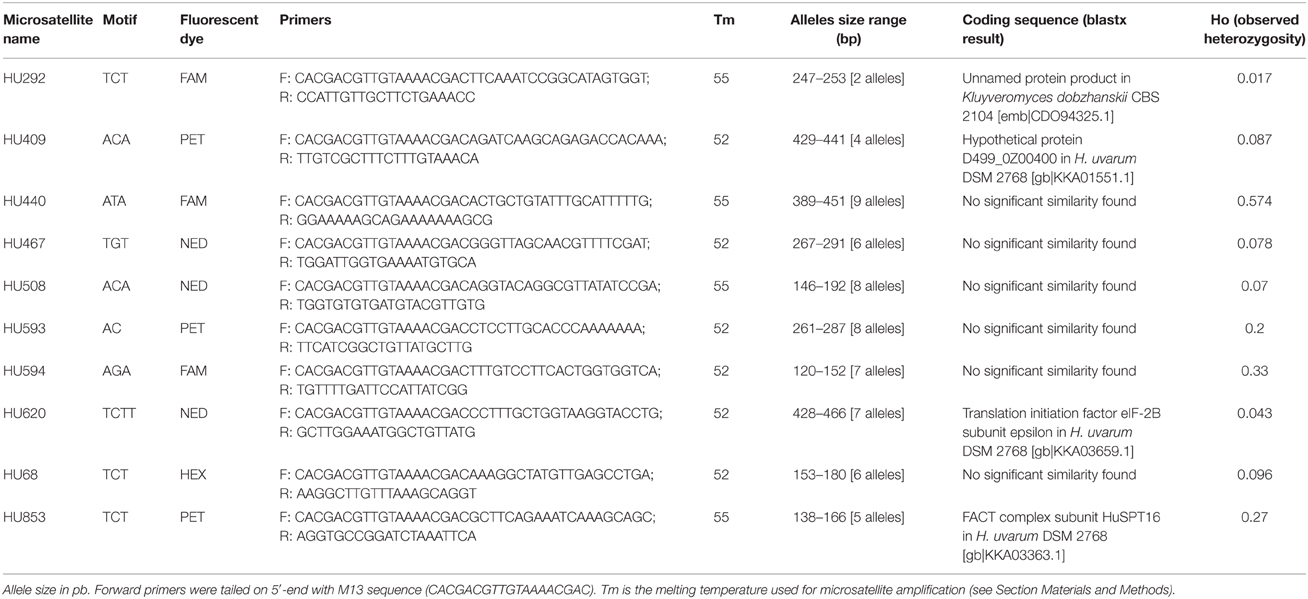
Table 2. Microsatellite loci for Hanseniaspora uvarum genotyping.
Microsatellites Amplification
DNA was prepared as followed: yeast cells were diluted in 20 mM NaOH (concentration of 1.108 cells/mL), then heated 10 min at 94°C. This solution was used as DNA templates for further PCR reactions.
PCR were performed in a final volume of 15 μL containing 1 μL of DNA template, 0.05 μM of forward primer, 0.5 μM of reverse primer and labeled primer, 1X Taq-&GO (MP Biomedicals, Illkirch, France). Universal M13 primers were labeled with either FAM-, HEX-, PET-, or NED-fluorescent dyes (Eurofins MWG Operon, Les Ulis, France).
Touch-down PCR were carried out using iCycler (Biorad, Hercules, CA) thermal cycler. The program encompassed an initial denaturation step of 1 min at 94°C followed by 10 cycles of 30 s at 94°C, 30 s at Tm + 10°C (followed by a 1°C decrease per cycle until Tm is reached) and 30 s at 72°C, then 20 cycles of 30 s at 94°C, 30 s at Tm and 30 s at 72°C, and a final extension step of 2 min at 72°C.
Amplicons were initially analyzed by a microchip electrophoresis system (MultiNA, Shimadzu) and the optimal conditions for PCR amplifications were assessed. Then, the sizes of the amplified fragments were measured on an ABI3730 DNA analyzer (Applied Biosystems). For that purpose, PCR amplicons were diluted (1800-fold for FAM, 600-fold for HEX, 1200-fold for PET, and 1800-fold for NED-labeled amplicons respectively) and multiplexed in formamide. LIZ 600 molecular marker (ABI GeneScan 600 LIZ Size Standard, Applied Biosystem) was 100-fold diluted and added for each multiplex. Before loading, diluted amplicons were heated 4 min at 94°C. Allele size was recorded using GeneMarker Demo software V2.4.0 (SoftGenetics).
Microsatellite Analysis
Microsatellite analysis, based on allele size, was used to investigate the genetic relationships between isolates. A dendrogram was built using Bruvo's distance (Bruvo et al., 2004) and Ward's clustering, by means of R (R Development Core Team, 2010). Bruvo's distance is particularly well adapted in the case of multiple and/or unknown ploidy levels, which is the case for H. uvarum species. Since classical bootstrap resampling is poorly reliable with microsatellite data, we assessed the robustness of the tree nodes using multiscale bootstrap resampling of the loci associated with an approximately unbiased test (Shimodaira, 2002) by means of R and the pvclust package v1.2-2 (Suzuki and Shimodaira, 2006; R Development Core Team, 2010).
Analysis of molecular variance (AMOVA) was performed by means of the pegas package (Paradis, 2010) with n = 1000 permutations. We tested whether the genetic distance was significantly explained by geographical localization (i.e., the country of isolation was used as grouping factor) or year of isolation (from 2003 to 2014).
Amplified Fragment Length Polymorphism
For all yeast species and isolates, genomic DNA was extracted using mechanical cell breakage with glass beads (Hoffman, 2001). DNA concentrations were determined using the NanoDrop™ 1000 spectrophotometer. The AFLP reactions were performed according to Esteve-Zarzoso et al. (2010). Briefly, 1.5 μg DNA was digested for 4 h with EcoRI and MseI at 37°C followed by ligation of the EcoRI (5′-CTCGTAGACTGCGTACC-3′ and 5′-AATTGGTACGCAGTC-3′) and MseI (5′-GACGATGAGTCCTGAG-3′ and 5′-TACTCAGGACTCAT-3′) adaptors. The primer pair EcoRI-0 (5′-GACTGCGTACCAATTC-3′) and MseI-C (5′-GATGAGTCCTGAGTAAC-3′) was used for the non-selective PCR of a 5 μL aliquot of the ligation mix diluted 10 × with TE buffer, while the selective primer was performed using EcoRI-C (5′-GACTGCGTACCAATTCC-3′) and MseI-AC (5′-GATGAGTCCTGAGTAAAC-3′) primer pair. The bands were resolved on a 2% (w/v) agarose gel with 1 × TBE buffer at 80 V. The gel was stained with GelRed and visualized under UV. The presence/absence of AFLP markers was scored against a GeneRuler™ 100 bp Plus DNA ladder (Fermentas Life Sciences, Finland) using GeneTools version 4.01 (SynGene, Synoptics Ltd., Cambridge, England). AFLP fragment sizes were rounded to the closest integer and a binary matrix (presence/absence) of 263 AFLP bands, ranging from 94 to 1865 pb was created. A dendrogram was subsequently built using Euclidean's distance, Ward's clustering and multiscale bootstrap resampling.
Screening for Extracellular Enzyme Activities of Enological Relevance
All yeast species were grown overnight in YPD broth (Biolab-Merck, Wadeville, South Africa) at 30°C on a rotary wheel. In order to standardize the number of cells spotted, the cultures were diluted to an optical density of 0.1 at a wavelength of 600 nm. On each plate, 10 μL of the diluted culture was spotted and incubated for 3 days at 30°C. The following activities were screened on solid agar media as previously reported in literature.
β-Glucosidase Activity
Extracellular β-glucosidase activity was tested on arbutin substrate [1% (w/v) yeast extract, 2% (w/v) peptone, 0.5% (w/v) arbutin, 20 mL 1% ammonium ferric] at pH 3.5 according to the method described by Strauss et al. (2001). S. polymorphus var. africanus (previously Debaryomyces polymorphus var. africanus) CBS 8047 was used as a positive control.
Acid Protease Activity
This assay was performed according to the method described by Bilinski et al. (1987). Sixty milliliters of phosphate-sodium buffer (24 g/L KH2PO4 + 35 g/L Na2HPO4-7H2O) was microwaved with 70 mL skim milk solution (100 g/L skim milk in 0.05 M citrate phosphate buffer) for ~45 s or until it starts simmering. Four hundred and eighty milliliters agar (20 g/L, pH adjusted to 3.5) was then added and the plates poured. M. pulcherrima IWBT Y1123 (Reid et al., 2012) was used as a positive control for protease activity.
Polygalacturonase Activity
The assay was carried out following the method described by van Wyk et al. (van Wyk and Divol, 2010). Polygalacturonic acid [1.25% (w/v)] was dissolved in 0.68% (w/v) potassium phosphate (pH 3.5), together with 0.67% (w/v) YNB, 1% (w/v) glucose, 2% (w/v) agar. Positive activity was measured against the control yeast S. paradoxus RO88 (Mocke, 2005).
Chitinase Activity
Colloidal chitin [0.45% (w/v)] was used as substrate to test for chitinase activity according to the method described by Agrawal and Kotasthane (2012). The pH of the medium was adjusted to 4.7. M. chrysoperlae IWBT Y955 was used as a positive control (Ghosh, 2015).
Screening for Killer Activity
Hanseniaspora spp. isolates were tested for their potential killer activity against S. cerevisiae ZIM1859 S6 previously reported as killer sensitive strains. The so-called “spot-on-the-lawn” technique was used, as described by Mehlomakulu et al. (2014). Briefly, all the strains were cultivated overnight in 5 mL YPD broth on a rotary wheel. The cells were harvested by centrifugation and re-suspended in saline [0.9% (w/v) NaCl] to an OD600nm of 0.1 (~3 × 107 cells/mL). To prepare the seeded cultures, 1 mL of the sensitive cells was mixed with 4 mL of a 4% (w/v) pre-autoclaved agar solution and 5 mL of a filter sterilized commercial preservative-free white table grape juice supplemented with 0.5% (w/v) yeast extract, adjusted to pH 4.5. The solution was poured into Petri dishes and allowed to set. Thereafter, 10 μL of overnight cultures of the potential killer strains in saline were spotted on the surface. The plates were incubated at 20°C until a lawn of seeded yeasts was visible and a zone of inhibition around the killer positive strain S. cerevisiae VIN13 was observed.
Sporulation
Sporulation ability of Hanseniapora spp. isolates was assessed on three different media: McClary's acetate agar (10% glucose, 1.8 g/L potassium chloride, 8.2 g/L sodium acetate trihydrate, 2.5 g/L yeast extract, 15 g/L agar), malt extract agar (5% malt extract, 2% agar) as described by Kurtzman et al. (2011), and potassium acetate agar (10 g/L Potassium Acetate, 15 g/L agar) by streaking colonies on these media. The cells were then stained according to the method described by Merritt and Hurley (1972).
Growth Assays under Various Environmental Conditions
Strain ability to metabolize glycerol as sole carbon source was tested as followed: strains were plated on 2% glycerol agar plates (1% yeast extract, 2% peptone, 2% glycerol, 1.5% agar) and incubated at 25°C for up to 7 days.
In order to test the impact of low temperature (12°C), anaerobia and the addition of copper solution, yeast strains were grown for 24 h in YNB (BD Difco) pH 3.5 at 25°C with constant agitation then serially 10-fold diluted and spotted on YNB agar plates (pH 3.5). Ten microliters of serial cellular concentrations were tested (103 cells/ml, 104 cells/ml, 105 cells/ml,) and gave similar results. Cellular suspensions were spotted using a Steers multipoint inoculator. Anaerobic conditions were created in sachet by AnaeroGen sachet AN0025 (Oxoid). Actual anaerobia was checked using GasPak™ Dry Anaerobic Indicator Strips (BD). The presence or absence of growth was recorded after 48 h incubation (12°C or 25°C, aerobia or anaerobia).
Susceptibility to copper was estimated by plating the yeast strains on YNB pH 3.5 containing either CuSO4 (copper sulfate, the molecule usually contained in Bordeaux mixture) or Cu(OH)2 (copper hydroxide, as contained in ChampFlo, Nufarm) at concentrations varying from 0.03 to 32 μg/mL of CuSO4 or Cu(OH)2 respectively. After 48H incubation at 25°C, the minimum inhibitory concentration (MIC) was determined.
Results
Development of Microsatellite Markers for Hanseniaspora Uvarum
Next generation sequencing was used to produce a de novo assembly of the genome sequence of CRBO L0551, a strain isolated from grape must in Bordeaux region in 2005. Although this de novo assembly displayed an important number of contigs (1665 contigs of more than 1000 bp), it was sufficient to locate repeated sequence. Microsatellite loci (dinucleotide to tetranucleotide) were selected on the basis of their location: on different contigs and not within the 5′-end and 3′-end of the contigs (3 kb exclusion in order to exclude possible telomeric or subtelomeric positions). Primers were designed to amplify 11 microsatellite loci, four of them being located within putative coding sequence (Table 2). The amplicons were separated using a microchip electrophoresis system (MultiNA), and the optimal conditions for microsatellites amplifications were assessed on a subpanel of five strains of H. uvarum (data not shown). After optimization, the microsatellites markers were tested on other species of the Hanseniaspora genus: H. guillermondii Y-1625T, 113816, IWBT Y1035, IWBT Y1165, IWBT Y901, IWBT Y970; H. opuntiae IWBT Y863 and IWBT Y875; H. vineae IWBT Y907, and IWBT Y971. No amplification was observed for these non-uvarum strains (data not shown), except for strain IWBT Y901. Strain IWBT Y901 was identified as H. guillermondii by sequencing both ITS and LSU D1/D2 rRNA regions, yet allowed the normal amplification of all 11 microsatellites markers.
The 11 microsatellites markers were then used to genotype 115 strains, including 101 H. uvarum strains isolated from various wineries in France near Bordeaux and in South Africa near Stellenbosch (Table 1). A few other isolates from winemaking environments were added: the type strain Y-1614 from Russia, five strains from New-Zealand (NZ1, NZ5, NZ15, NZ148, and NZ234) and CLIB 303 from Ukraine. Six strains from non-enological environments were also genotyped: Y-1612 (soil, Indonesia), Y-915 (cider), DSMZ 70285 (soil), 516149 (maize, Japan), YB-783 (tree, USA), and YB-3199 (fruit, USA). Strain IWBT Y901, identified as H. guillermondii but able to amplify all microsatellites, was also added. All microsatellites were polymorphic on this panel of 115 strains, with only two alleles for HU292 and up to 9 alleles for HU440 (Table 2). Although the polymorphism of the microsatellite loci was limited compared to other species (Legras et al., 2005; Albertin et al., 2014a,c; Masneuf-Pomarede et al., 2015), altogether they were discriminant enough to detect 86 different genotypes over the 115 tested. Twenty strains displayed only one allele per locus, while 95 showed heterozygosity for at least 1 upon 10 loci. Heterozygosity was detected for all loci, with observed heterozygosity ranging from 0.017 for the less polymorphic locus HU292 to 0.574 for HU440, the more polymorphic locus.
Exploring the Genetic Relationships Between H. uvarum Isolates Using Microsatellites
The genetic relationships between the 115 isolates of H. uvarum were studied using Bruvo's distance (Bruvo et al., 2004) and Ward's clustering. The resulting dendrogram (Figure 1) shows three main clusters: one cluster (group C) contained almost all strains from South African winemaking environments (19 of the 21), and was highly supported (bootstrap value of 96). The two other groups contained mostly wine strains from France, but interestingly, these groups clustered on the basis of the year of isolation: most strains collected before 2009 clustered in group A (19 strains upon 29), with high bootstrap value (91). Group B contained 40 strains, most of them (25) being isolated after 2009 from winemaking environments in France (boostrap value of 91).
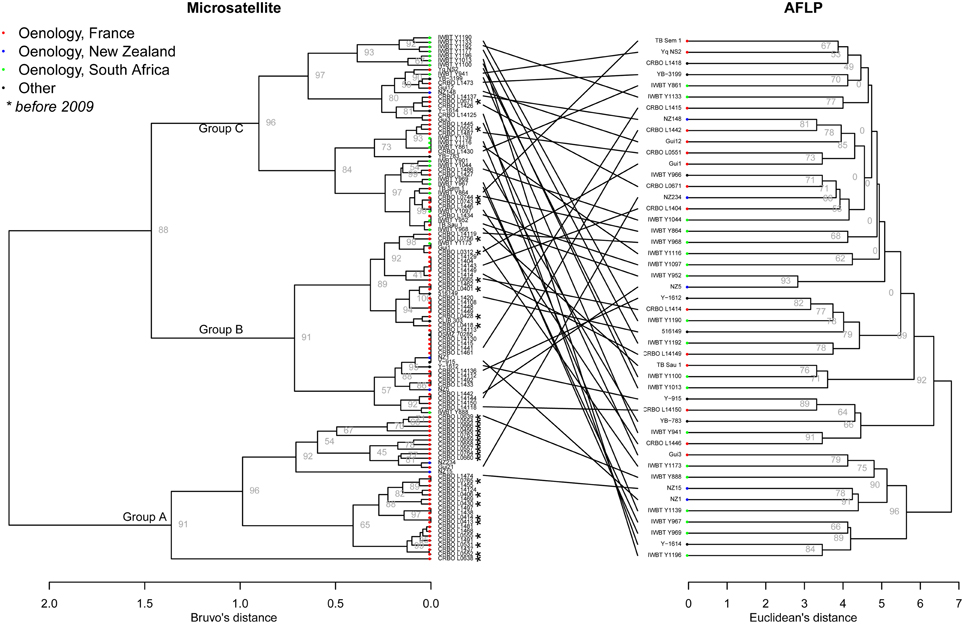
Figure 1. Dendrogram trees of Hanseniaspora uvarum from microsatellite dataset (115 strains) and AFLP dataset (47 strains). Bruvo'as and Euclidean distance were used for microsatellite and AFLP data, respectively. Ward's clustering and multiscale bootstrap resampling were used in both cases. For visibility, only the boostraps of the higher nodes were shown for microsatellite data.
To confirm the genetic clustering based on both geographical distance and year of isolation, we performed AMOVA. When using the country of origin as grouping factor, AMOVA was significant (p = 0.00099), and the country explained 8.54% of the total variation of the microsatellite dataset (Table 3). The year of isolation was also used as a grouping factor, and explained much more variation (20.62%, p < 10−6). These results confirmed that year of isolation as well as geographical origin significantly shaped the diversity of H. uvarum populations from winemaking environments.
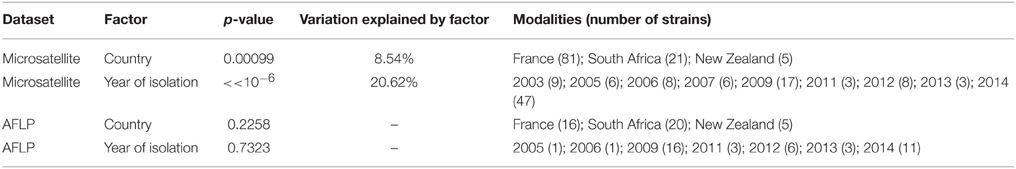
Table 3. AMOVA results using microsatellite or AFLP dataset, for country and year of isolation as grouping factors.
Genetic Diversity of H. uvarum Populations in Winemaking Environments
The wine strains used in this study were isolated from several wineries, sometimes from different samples over several years. This is the case of winery G, for which 19 strains were isolated from grape must between 2003 and 2014 (Table 4). These 19 strains displayed 17 different genotypes distributed throughout the dendrogram, indicating that no clone was a specific signature of this winery. The same pattern was observed for all wineries: in most instances, several genotypes from different genetic groups were identified within the same winery, suggesting that the absence of genetic signature at the winery level was common for H. uvarum population.
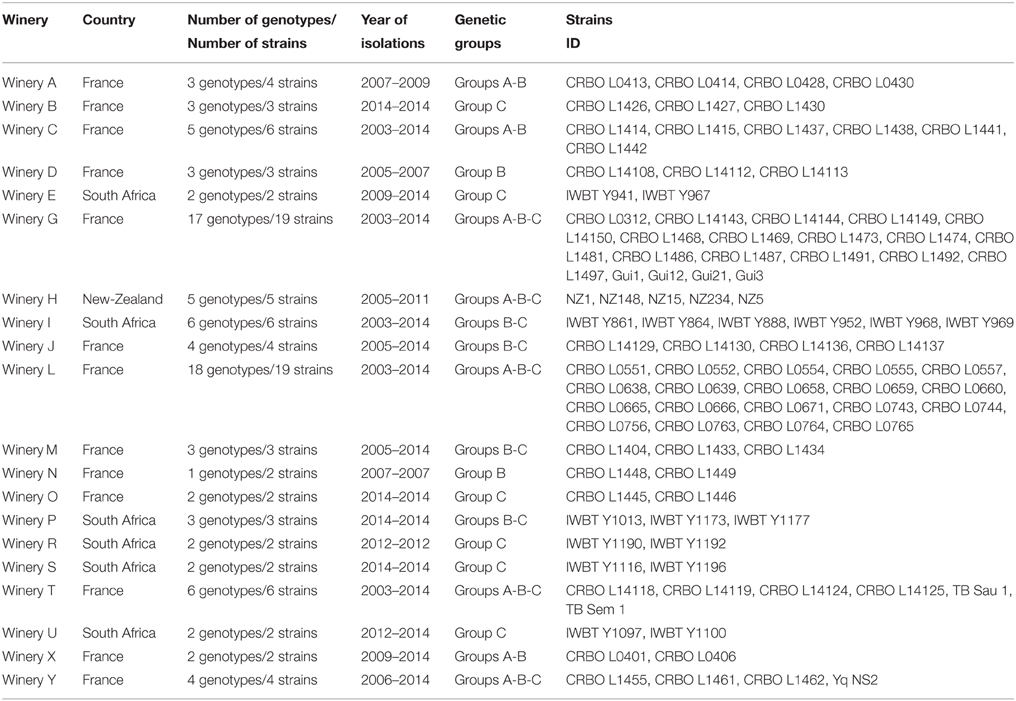
Table 4. Diversity of Hanseniaspora uvarum populations for 20 wineries as detected by microsatellite genotyping.
We also studied the genetic diversity at sample level. For example, five strains from grape must were isolated and genotyped from the same sample in winery. The five strains (CRBO L0743, CRBO L0756, CRBO L0763, CRBO L0764, and CRBO L0765) exhibited five different genotypes distributed on the tree. In addition, in 2005, for winery L, some strains were isolated on days 1, 3, 4, and 5 during the pre-fermentative stage of the same tank. The four corresponding strains (CRBO L0552, CRBO L0554, CRBO L0555, and CRBO L0557) clustered in group A on the dendrogram, but they all displayed different genotypes. Altogether, these results suggest that the diversity of the population of H. uvarum is high in winemaking environments and that no specific genetic signature exists in a given winery.
Comparing Microsatellite and AFLP Typing
AFLP techniques are viewed as moderately repeatable over a high number of experiments, so that AFLP analyses are usually applied to a limited number of strains in order to be reliable. Here, we chose to apply AFLP analysis to a subpanel of 47 strains, in order to compare the results obtained from microsatellite data and AFLP data. The AFLP dendrogram was produced using Euclidean distance and Ward's clustering (Figure 1). Comparison between microsatellite and AFLP dendrograms revealed important differences with no obvious clustering for AFLP data for country origin or vintage, even when comparing exactly the same subset of strains (Supplementary Figure 1). We subsequently performed AMOVA analysis using the distance matrix produced from AFLP data (Table 3). When using the country or the year of isolation as grouping factor, AMOVA was not significant (p > 0.05) using AFLP data, indicating that AFLP clustering was not able to detect the genetic structure depending on geographical origin nor year of isolation. However, it has to be noted that AFLP tool was able to discriminate 47 strains upon 47 on the basis of their AFLP patterns. By contrast, on the same subset, the microsatellite tool identified 37 different genotypes. This indicated that although AFLP tool lacked robustness to assess the genetic relationship of individuals, it was more discriminant than the microsatellite tool.
Phenotyping Hanseniaspora sp. Isolates
As phenotyping assays are time-consuming, a subpanel of 30 strains of H. uvarum and 10 other Hanseniaspora spp. were selected and subjected to various plate assays to assess whether they possessed any extracellular enzyme activity that could be of interest in enology (Table 5). Their killer activity against two strains of S. cerevisiae that are sensitive to S. cerevisiae's killer toxins was also investigated. Finally, their ability to grow when exposed to various environmental factors of scientific or enological interest (low temperature, anaerobia, copper presence, glycerol as the only carbon source) was recorded.
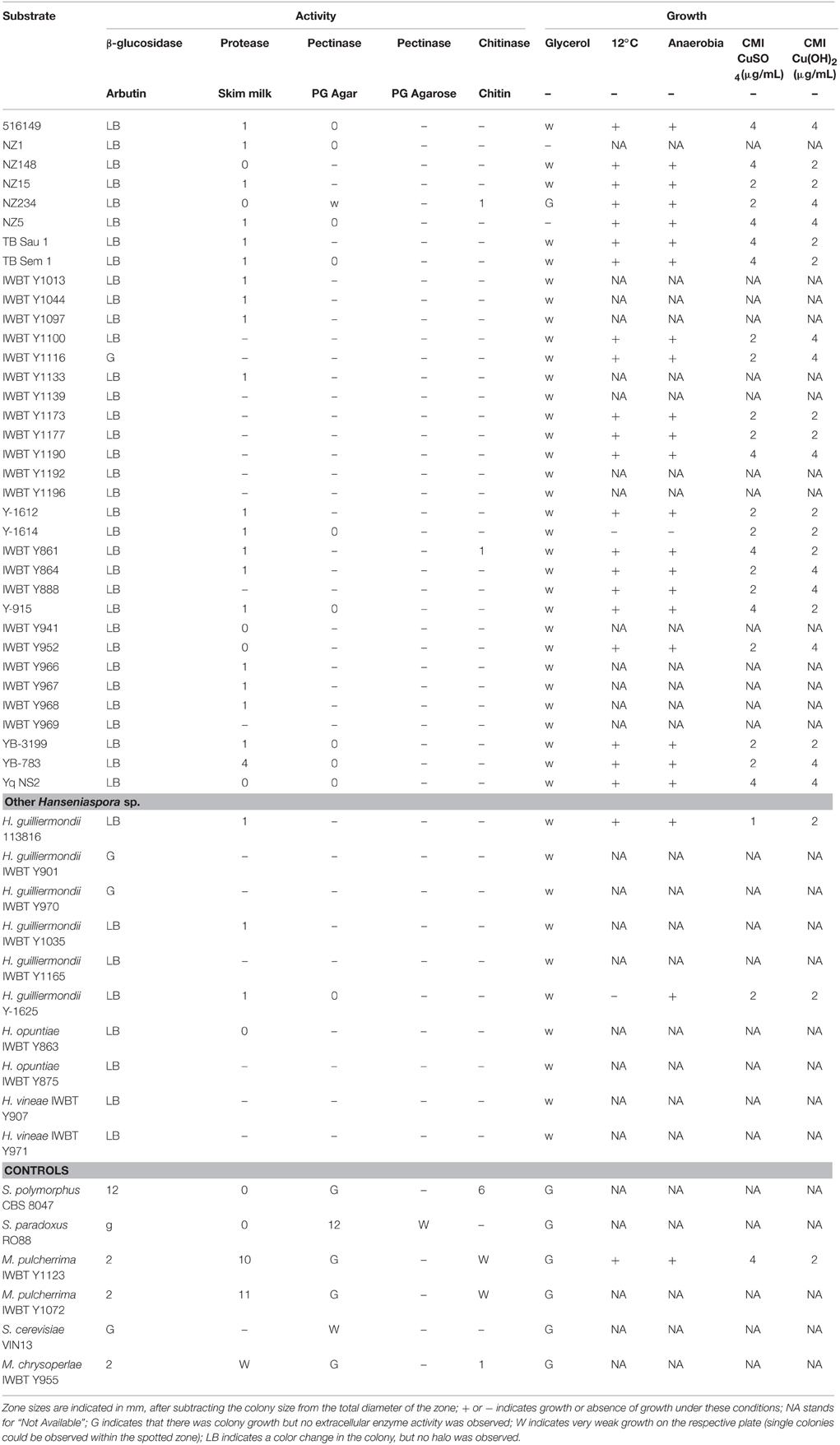
Table 5. Phenotyping 45 Hanseniaspora sp. strains for enzymatic activities and growth ability.
With regards to extracellular enzyme activity, all strains showed growth on the arbutin plates, although no distinct halo could be observed. Most Hanseniaspora strains showed a slight browning of the colony which might be due to weak activity or even intracellular β-glucosidase activity. Weak acid protease activity was observed for most strains, with YB-783 showing the largest halo (4 mm) and strains Gui21 and CRBO L0551 showing halos of 2 mm after 72-h incubation. No polygalacturonase activity was observed. Finally, weak chitinase activity could be visualized in isolates NZ234 and Y-861 (1-mm halo) while the other isolates showed no growth on the chitin media.
None of the strains was able to sporulate. Regarding their growth ability, most strains were unable to grow on 2% glycerol agar plates after incubation at 30°C for up to 7 days. Their growth ability under various environmental conditions was tested: ability to grow at low temperature (12 and 30°C), under oxic or anoxic, in presence of various copper concentrations. The MIC for copper sulfate and copper hydroxide was either 2 or 4 mg/L with no specific correlation between these 2 factors. All strains tested showed similar ability to grow under these conditions of enological interest, and limited phenotypic variations were recorded. Indeed, only strain Y-1614 did not show any growth at 12°C and anaerobiosis. Finally, no killer activity was observed against S. cerevisiae.
Discussion
Comparing Microsatellite and AFLP Genotyping
In this paper, we compared the intraspecific clustering using two different techniques: AFLP and microsatellites. Both approaches allowed discrimination at the strain level: 47 different patterns were scored for AFLP (for 47 strains), while 86 genotypes were evidenced for 115 strains with microsatellite data. Indeed, both methods proved to be discriminant as previously reported (Mariette et al., 2001; Gaudeul et al., 2004), with AFLP having a higher discriminant power in our case. However, it has to be noted that using AFLP, the amplification of multiple bands in a single run may lead to competition between amplicons and therefore to differences of band intensity that complicate data analysis. In addition, AFLP techniques are usually viewed as moderately repeatable thereby making the technique usually poorly reliable, while the repeatability of microsatellites markers is usually higher (Jones et al., 1997) and can be thus applied to a larger number of individuals.
Moreover, AFLP markers are non-codominant markers, so that homozygosity or heterozygosity is difficult to assess (Gaudeul et al., 2004). By contrast, microsatellites are codominant markers, allowing assessing heterozygosity status. Here, we found that 95 out of 115 strains showed heterozygosity, allowing an unprecedented insight into the genetics of the species. Microsatellites are widely used to estimate relatedness among individuals or differentiation among groups. By contrast, AFLP should be taken with caution due to the lack of complete genotypic information caused by dominance (Parker et al., 1998). Indeed, the dendrogram obtained by both approaches are not comparable and the genetic structure based on year of isolation and geographical origin evidenced using microsatellite was completely missed by AFLP analysis.
As expected, microsatellite genotyping proved to be a better tool for establishing genetic relation between strains and getting new insights within species at genetic level (Ross et al., 1999). By contrast, AFLP is interesting to perform assays where genetic relatedness is not needed, which is usually the case for several biotechnological applications in enology: assessing the global population diversity, testing for the prevalence/implantation of a specific (known) strain, searching for contamination evidence, etc. In these latter instances, the technical simplicity and rapidity of AFLP, associated with low cost, is definitively advantageous compared to microsatellite genotyping.
New Insights into the Genetic Structure of Hanseniaspora uvarum from Winemaking Environments
Like many non-conventional yeasts of enological interest, the genetic structure of H. uvarum from winemaking environments remained elusive. Here, using microsatellite data, we show that an important number of H. uvarum strains (95/115) are heterozygous. This result could be congruent with the hypothesis of a diploid species, although the possibility of aneuploidy could not be excluded. Additional work should be performed to confirm its diploid status, but will be complicated by the absence of sporulation on classical medium. The absence of sporulation could be explained, at least in part, by the weak ability of the species to metabolize glycerol, suggesting poor respiration ability (sporulation being strongly linked to respiration ability in S. cerevisiae, Codon et al., 1995). Indeed, all H. uvarum strains (except Y1614) showed unperturbed growth under anaerobic conditions, indicating that respiratory metabolism is not necessary for their normal growth.
Interestingly, strain IWBT Y901 –showing 456/456 identities for D1/D2 sequence with Hanseniaspora guilliermondii CBS 465T- was clustered among H. uvarum strains in group C. While some microsatellite markers can cross the species, most microsatellites are intraspecific. The fact that IWBT Y901 is the sole H. guillermondii strain to be amplified by all 11 markers is clearly unusual. The possibility of a contamination of this strain with an actual H. uvarum strain can be eliminated as IWBT Y901 differed from all other strains we genotyped: its closest relative IWBT Y1044 differs from two alleles at two different loci. One possible explanation for these unexpected results is that IWBT Y901 could derived from an interspecific hybrid between H. uvarum and H. guillermondii, a hypothesis that remains to be demonstrated formally.
Microsatellite analysis also reveals a genetic structure related to geographical localization. Since we genotyped mostly strains from France and South Africa, it could be interesting to extend our analysis to strains from other countries in order to assess the extent of relationship between genetic structure and geographical origin. A few wine yeasts were shown to be genetically structured, at least partially, by geography, as it is the case of Saccharomyces uvarum, Candida zemplinina, or Torulaspora delbrueckii (Albertin et al., 2014a; Almeida et al., 2014; Masneuf-Pomarede et al., 2015). More surprisingly, our data show a strong relationship between the year of isolation and the genetic structure. This result indicates that H. uvarum populations isolated from winemaking environments show a temporal clustering in addition to a spatial one. More data are required to determine whether this temporal variation exists only for French and South African strains over the period we tested (2003–2014), or if this trend is also detected for other vineyards and/or larger periods of time. In addition, further investigation is required to determine which factor(s) could be related to this temporal evolution. Factors to be tested include environmental factors such as temperature, pH, sugar concentrations, ability to survive from season to season, etc. Viticultural and enological practices should also be considered, including phytosanitary treatments, sulfite addition, cold pre-fermentation stage, turbidity, or starter culture addition that were shown to impact Hanseniaspora populations during the early stage of AF (Albertin et al., 2014b).
Finally, our data failed to identify any specific genetic signature associated with wineries and/or samples. Moreover, we globally identified high level of genetic diversity within all wineries/samples tested, with no evidence for clonal dominance. Such high genetic diversity was previously shown for Hanseniaspora populations in grape must and other environments of the winery using FT-IR (Grangeteau et al., 2015). High diversity was also detected for the wine yeast C. zemplinina (Masneuf-Pomarede et al., 2015), while other wine species like the spoilage yeast Brettanomyces bruxellensis showed clonal populations and maintenance over decades in winemaking environments (Albertin et al., 2014c). H. uvarum is known to be insect associated and therefore we can speculate that its diversity may depend on the diversity and frequency of insect occurrence during ripening (Lam and Howell, 2015). Such hypothesis could be tested by investigating the insect-associated yeast diversity and comparing it with the grape/winery diversity.
H. uvarum Displays Low Phenotypic Variability for the Traits Investigated in this Study
In order to investigate whether the genetic clustering evidenced above reflects a certain phenotypic diversity, a number of phenotypes of enological relevance were tested: secretion of typical enzymes, as well as ability to grow at cold temperature (similar to that occurring at the beginning of the winemaking process and possibly during the fermentation of white wine), in anaerobiosis (typically occurring during wine fermentation) and ability to tolerate copper, a typical anti-fungal treatment used in the vineyard. The ability to grow on glycerol as sole source of carbon was also tested, following a preliminary observation that H. uvarum could not utilize glycerol (not shown). Although both microsatellites and AFLP revealed large genetic variation, the phenotypic variability was very low for the factors investigated (Table 4). Indeed, with a few exceptions, most strains exhibited similar extracellular enzyme activity, tolerance to copper and ability to grow at low temperature or poor ability to use glycerol as sole source of carbon. No clear connection could be observed between these phenotypes and the genetic clustering reported above. H. uvarum is usually categorized as a good producer of extracellular enzymes (Dizy and Bisson, 2000) and it is typically reported to exhibit all the enzymatic activities investigated in this study, although this seems to be strain dependent. However, most authors did not adjust the pH of their screening media to wine pH. There seems to be a general consensus between our data and previous studies, that when pH is adjusted to 3.5, most strains of H. uvarum display β-glucosidase and protease activities (Lagace and Bisson, 1990; Charoenchai et al., 1997), but not polygalacturonase activity (Charoenchai et al., 1997). No study however investigated the actual impact of these extracellular enzymes on wine composition. All the strains investigated in our study except one could grow at 12°C. This is in agreement with literature. Indeed, it has been reported that low temperatures favor biomass production in H. uvarum (Ciani et al., 2006; Mendoza et al., 2009). Surprisingly, H. uvarum seems to be poorly able to consume glycerol, even in the presence of oxygen. None of the strains investigated in this study were found able to sporulate. They should therefore all be classified as H. uvarum (teleomorph) and not K. apiculata (anamorph). As reviewed by Jolly et al. (2006), the region of isolation seems to play a role in the distribution of H. uvarum and K. apiculata. In temperate regions, an equal mixture of teleomorph/anamorph is found, while in warmer climates, only the teleomorph H. uvarum is detected. Yet our strains were isolated from both temperate and warm climate regions (France and South Africa, respectively) and none of the strains studied here were found able to sporulate. Another explanation could be the amount of time that these strains spent as freeze cultures. Indeed, some authors have reported that the time between isolation and analysis plays a role in the ability of H. uvarum to sporulate (see review by Jolly et al., 2006).
Overall, with regard to the traits investigated in this study, H. uvarum seems to display very little phenotypic variability. In literature, a greater diversity seems to occur in terms of intracellular metabolism. Indeed, several studies report on the influence of H. uvarum inoculated in pure or mixed culture with S. cerevisiae and describe its production of esters, higher alcohols, and fatty acids (Moreira et al., 2008; Suzzi et al., 2012). These studies are not always in full agreement, as mentioned by the latter authors, and these would point out toward some intraspecific diversity at this level. Nevertheless, our results show that upon inoculation in grape juice, H. uvarum could survive the typical cold temperatures applied early in the winemaking process and during the fermentation of white wine as well as the anaerobic conditions also occurring during fermentation. Furthermore, it could potentially release glycosylated compounds and break down proteins through the activity of its extracellular enzymes, both properties being of strong enological interest. Since H. uvarum displayed no killer activity against the strains of S. cerevisiae tested, these two yeast species could be co-inoculated without threatening the overall proceedings of AF.
In conclusion, we describe in this paper a new analytical tool (microsatellite markers) that allowed estimating the genetic diversity and the genetic relationship between H. uvarum from winemaking environments. Our results indicate that H. uvarum populations are structured by both geographical origin and the year of isolation from a genetic viewpoint. By contrast, the phenotypic variability was more limited regarding extracellular enzymatic activities and response to environmental factors. Subsequent analysis of a larger number of isolates will help determine the extent of such results in winemaking environments.
Author Contributions
WA and MS designed and performed most of the experiments. CM and TM performed the screening experiments. BC, JC, PG, and VM performed microsatellite development. MP and FS performed genome sequencing and microsatellite analysis. WA, MME, MB, BD and IMF conceived the project, wrote and edited the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01569
Supplementary Figure 1. Comparaison of dendrogram trees of Hanseniaspora uvarum obtained from microsatellite and AFLP approaches on the same subset of strains. Bruvo'as and Euclidean distance were used for microsatellite and AFLP data, respectively.
References
Agrawal, T., and Kotasthane, A. S. (2012). Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springerplus 1:73. doi: 10.1186/2193-1801-1-73
Albertin, W., Chasseriaud, L., Comte, G., Panfili, A., Delcamp, A., Salin, F., et al. (2014a). Winemaking and bioprocesses strongly shaped the genetic diversity of the ubiquitous yeast Torulaspora delbrueckii. PLoS ONE 9:e94246. doi: 10.1371/journal.pone.0094246
Albertin, W., Miot-Sertier, C., Bely, M., Marullo, P., Coulon, J., Moine, V., et al. (2014b). Oenological prefermentation practices strongly impact yeast population dynamics and alcoholic fermentation kinetics in Chardonnay grape must. Int. J. Food Microbiol. 178, 87–97. doi: 10.1016/j.ijfoodmicro.2014.03.009
Albertin, W., Panfili, A., Miot-Sertier, C., Goulielmakis, A., Delcamp, A., Salin, F., et al. (2014c). Development of microsatellite markers for the rapid and reliable genotyping of Brettanomyces bruxellensis at strain level. Food Microbiol. 42, 188–195. doi: 10.1016/j.fm.2014.03.012
Almeida, P., Gonçalves, C., Teixeira, S., Libkind, D., Bontrager, M., Masneuf-Pomarède, I., et al. (2014). A Gondwanan imprint on global diversity and domestication of wine and cider yeast Saccharomyces uvarum. Nat. Commun. 5:4044. doi: 10.1038/ncomms5044
Angioni, A., Caboni, P., Garau, A., Farris, A., Orro, D., Budroni, M., et al. (2007). In vitro interaction between ochratoxin A and different strains of Saccharomyces cerevisiae and Kloeckera apiculata. J. Agric. Food Chem. 55, 2043–2048. doi: 10.1021/jf062768u
Basha, H., and Ramanujam, B. (2014). Growth promotion effect of Pichia guilliermondii in chilli and biocontrol potential of Hanseniaspora uvarum against Colletotrichum capsici causing fruit rot. Biocont. Sci. Technol. 25, 185–206. doi: 10.1080/09583157.2014.968092
Batista, N. N., Ramos, C. L., Ribeiro, D. D., Pinheiro, A. C. M., and Schwan, R. F. (2015). Dynamic behavior of Saccharomyces cerevisiae, Pichia kluyveri and Hanseniaspora uvarum during spontaneous and inoculated cocoa fermentations and their effect on sensory characteristics of chocolate. LWT Food Sci. Technol. 63, 221–227. doi: 10.1016/j.lwt.2015.03.051
Beltran, G., Torija, M. J., Novo, M., Ferrer, N., Poblet, M., Guillamón, J. M., et al. (2002). Analysis of yeast populations during alcoholic fermentation: a six year follow-up study. Syst. Appl. Microbiol. 25, 287–293. doi: 10.1078/0723-2020-00097
Bilbao, A., Irastorza, A., Dueñas, M., and Fernandez, K. (1997). The effect of temperature on the growth of strains of Kloeckera apiculata and Saccharomyces cerevisiae in apple juice fermentation. Lett. Appl. Microbiol. 24, 37–39. doi: 10.1046/j.1472-765X.1997.00340.x
Bilinski, C. A., Russell, I., and Stewart, G. G. (1987). Applicability of yeast extracellular proteinases in brewing: physiological and biochemical aspects. Appl. Environ. Microbiol. 53, 495–499.
Bruvo, R., Michiels, N. K., D'souza, T. G., and Schulenburg, H. (2004). A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 13, 2101–2106. doi: 10.1111/j.1365-294X.2004.02209.x
Cabranes, C., Mangas, J. J., and Blanco, D. (1997). Selection and biochemical characterisation of Saccharomyces cerevisiae and Kloeckera apiculata strains isolated from spanish cider. J. Inst. Brew. 103, 165–169. doi: 10.1002/j.2050-0416.1997.tb00945.x
Cai, Z., Yang, R., Xiao, H., Qin, X., and Si, L. (2015). Effect of preharvest application of Hanseniaspora uvarum on postharvest diseases in strawberries. Postharvest Biol. Technol. 100, 52–58. doi: 10.1016/j.postharvbio.2014.09.004
Capece, A., Fiore, C., Maraz, A., and Romano, P. (2005). Molecular and technological approaches to evaluate strain biodiversity in Hanseniaspora uvarum of wine origin. J. Appl. Microbiol. 98, 136–144. doi: 10.1111/j.1365-2672.2004.02434.x
Capriotti, A. (1955). Yeasts in some Netherlands soils. Antonie Van Leeuwenhoek 21, 145–156. doi: 10.1007/BF02543809
Caruso, M., Capece, A., Salzano, G., and Romano, P. (2002). Typing of Saccharomyces cerevisiae and Kloeckera apiculata strains from Aglianico wine. Lett. Appl. Microbiol. 34, 323–328. doi: 10.1046/j.1472-765X.2002.01090.x
Castelli, T. (1955). Yeasts of wine fermentations from various regions of Italy. Am. J. Enol. Vitic. 6, 18–19.
Charoenchai, C., Fleet, G. H., Henschke, P. A., and Todd, B. E. N. T. (1997). Screening of non-Saccharomyces wine yeasts for the presence of extracellular hydrolytic enzymes. Aust. J. Grape Wine Res. 3, 2–8. doi: 10.1111/j.1755-0238.1997.tb00109.x
Ciani, M. (1998). Wine vinegar production using base wines made with different yeast species. J. Sci. Food Agric. 78, 290–294.
Ciani, M., Beco, L., and Comitini, F. (2006). Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 108, 239–245. doi: 10.1016/j.ijfoodmicro.2005.11.012
Codon, A. C., Gasentramirez, J. M., and Benitez, T. (1995). Factors which affect the frequency of sporulation and tetrad formation in Saccharomyces cerevisiae bakers yeasts. Appl. Environ. Microbiol. 61, 1677–1677.
Combina, M., Elía, A., Mercado, L., Catania, C., Ganga, A., and Martinez, C. (2005). Dynamics of indigenous yeast populations during spontaneous fermentation of wines from Mendoza, Argentina. Int. J. Food Microbiol. 99, 237–243. doi: 10.1016/j.ijfoodmicro.2004.08.017
Comi, G., Romano, P., Cocolin, L., and Fiore, C. (2001). Characterization of Kloeckera apiculata strains from the Friuli region in Northern Italy. World J. Microbiol. Biotechnol. 17, 391–394. doi: 10.1023/A:1016731420425
Comitini, F., and Ciani, M. (2010). The zymocidial activity of Tetrapisispora phaffii in the control of Hanseniaspora uvarum during the early stages of winemaking. Lett. Appl. Microbiol. 50, 50–56. doi: 10.1111/j.1472-765X.2009.02754.x
de Araujo, F. V., Soares, C. A. G., Hagler, A. N., and Mendonça-Hagler, L. C. (1995). Ascomycetous yeast communities of marine invertebrates in a Southeast Brazilian mangrove ecosystem. Antonie Van Leeuwenhoek 68, 91–99. doi: 10.1007/BF00873096
de Arruda Moura Pietrowski, G., Grochoski, M., Sartori, G. F., Gomes, T. A., Wosiacki, G., and Nogueira, A. (2015). Viability of Hanseniaspora uvarum yeast preserved by lyophilization and cryopreservation. Yeast 32, 559–565. doi: 10.1002/yea.3079
Dizy, M., and Bisson, L. F. (2000). Proteolytic activity of yeast strains during grape juice fermentation. Am. J. Enol. Vitic. 51, 155–167.
Emmanouil-Nikoloussi, E., Kanellaki-Kyparissi, M., Papavassiliou, P., Koliakos, K., Dermentzopoulou, M., and Foroglou, C. (1994). “Hanseniaspora uvarum” the ultrastructural morphology of a rare ascomycete, isolated from oral thrush. Bull. Group. Int. Rech. Sci. Stomatol. Odontol. 37, 13–17.
Esteve-Zarzoso, B., Belloch, C., Uruburu, F., and Querol, A. (1999). Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 49, 329–337. doi: 10.1099/00207713-49-1-329
Esteve-Zarzoso, B., Hierro, N., Mas, A., and Guillamon, J. M. (2010). A new simplified AFLP method for wine yeast strain typing. LWT Food Sci. Technol. 43, 1480–1484. doi: 10.1016/j.lwt.2010.05.016
Esteve-Zarzoso, B., Peris-Torán, M., Ramón, D., and Querol, A. (2001). Molecular characterisation of Hanseniaspora species. Antonie Van Leeuwenhoek 80, 85–92. doi: 10.1023/A:1012268931569
Farías, M. E., and Manca de Nadra, M. C. (2003). Flocculation and cell surface characterization of Kloeckera apiculata from wine. J. Appl. Microbiol. 95, 457–462. doi: 10.1046/j.1365-2672.2003.01994.x
Gao, C., and Fleet, G. H. (1988). The effects of temperature and pH on the ethanol tolerance of the wine yeasts, Saccharomyces cerevisiae, Candida stellata and Kloeckera apiculata. J. Appl. Bacteriol. 65, 405–409. doi: 10.1111/j.1365-2672.1988.tb01909.x
Garcia-Martos, P., Hernandez-Molina, J. M., Galan, F., Ruiz-Henestrosa, J. R., Garcia-Agudo, R., J., P. M., and Mira, J. (1999). Isolation of Hanseniaspora uvarum (Kloeckera apiculata) in humans. Mycopathologia 144, 73–75. doi: 10.1023/A:1006900909455
Gaudeul, M., Till-Bottraud, I., Barjon, F., and Manel, S. (2004). Genetic diversity and differentiation in Eryngium alpinum L. (Apiaceae): comparison of AFLP and microsatellite markers. Heredity 92, 508–518. doi: 10.1038/sj.hdy.6800443
Ghosh, S. (2015). Metagenomic Screening of Cell Wall Hydrolases, Their Anti-Fungal Activities and Potential Role in Wine Fermentation. Ph.D. thesis, Stellenbosch University.
Granchi, L., Bosco, M., Messini, A., and Vincenzini, M. (1999). Rapid detection and quantification of yeast species during spontaneous wine fermentation by PCR-RFLP analysis of the rDNA ITS region. J. Appl. Microbiol. 87, 949–956. doi: 10.1046/j.1365-2672.1999.00600.x
Grangeteau, C., Gerhards, D., Rousseaux, S., von Wallbrunn, C., Alexandre, H., and Guilloux-Benatier, M. (2015). Diversity of yeast strains of the genus Hanseniaspora in the winery environment: what is their involvement in grape must fermentation? Food Microbiol. 50, 70–77. doi: 10.1016/j.fm.2015.03.009
Heard, G. M., and Fleet, G. H. (1985). Growth of natural yeast flora during the fermentation of inoculated wines. Appl. Environ. Microbiol. 50, 727–728.
Heard, G. M., and Fleet, G. H. (1988). The effects of temperature and pH on the growth of yeast species during the fermentation of grape juice. J. Appl. Bacteriol. 65, 23–28. doi: 10.1111/j.1365-2672.1988.tb04312.x
Hierro, N., González, Á., Mas, A., and Guillamón, J. M. (2006). Diversity and evolution of non-Saccharomyces yeast populations during wine fermentation: effect of grape ripeness and cold maceration. FEMS yeast Res. 6, 102–111. doi: 10.1111/j.1567-1364.2005.00014.x
Hoffman, C. S. (2001). “Preparation of Yeast DNA,” in Current Protocols in Molecular Biology, eds F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (John Wiley & Sons Inc), 13.11.11–13.11.14.
Holloway, P., Subden, R. E., and Lachance, M. A. (1990). The yeasts in a riesling must from the Niagara grape-growing region of Ontario. Can. Inst. Food Sci. Technol. J. 23, 212–216. doi: 10.1016/S0315-5463(90)70245-3
Hong, Y.-A., and Park, H.-D. (2013). Role of non-Saccharomyces yeasts in Korean wines produced from Campbell early grapes: potential use of Hanseniaspora uvarum as a starter culture. Food Microbiol. 34, 207–214. doi: 10.1016/j.fm.2012.12.011
Jolly, N. P., Augustyn, O. P. H., and Pretorius, I. S. (2003). The occurrence of non-Saccharomyces cerevisiae yeast species over three vintages in four vineyards and grape musts from four production regions of the Western Cape. S. Afr. J. Enol. Vitic. 24, 35–42.
Jolly, N. P., Augustyn, O. P. H., and Pretorius, I. S. (2006). The role and use of non-Saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 27, 15–39.
Jones, C. J., Edwards, K. J., Castaglione, S., Winfield, M. O., Sala, F., van de Wiel, C., et al. (1997). Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Mol. Breed. 3, 381–390. doi: 10.1023/A:1009612517139
Kachalkin, A. V., Abdullabekova, D. A., Magomedova, E. S., Magomedov, G. G., and Chernov, I. Y. (2015). Yeasts of the vineyards in Dagestan and other regions. Microbiology 84, 425–432. doi: 10.1134/S002626171503008X
Kocan, R. M., and Hasenclever, H. F. (1972). Normal yeast flora of the upper digestive tract of some wild columbids. J. Wildl. Dis. 8, 365–368. doi: 10.7589/0090-3558-8.4.365
Kosse, D., Seiler, H., Amann, R., Ludwig, W., and Scherer, S. (1997). Identification of Yoghurt-spoiling Yeasts with 18S rRNA-targeted Oligonucleotide Probes. Syst. Appl. Microbiol. 20, 468–480. doi: 10.1016/S0723-2020(97)80016-1
Kurtzman, C. P., Fell, J. W., and Boekhout, T. (2011). The Yeasts: A Taxonomic Study. Amsterdam: Elsevier.
Kurtzman, C. P., and Robnett, C. J. (1998). Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 73, 331–371. doi: 10.1023/A:1001761008817
Lachance, M.-A. (1995). Yeast communities in a natural tequila fermentation. Antonie Van Leeuwenhoek 68, 151–160. doi: 10.1007/BF00873100
Lagace, L. S., and Bisson, L. F. (1990). Survey of yeast acid proteases for effectiveness of wine haze reduction. Am. J. Enol. Vitic. 41, 147–155.
Lam, S. S., and Howell, K. S. (2015). Drosophila-associated yeast species in vineyard ecosystems. FEMS Microbiol Lett 362. doi: 10.1093/femsle/fnv170
Legras, J. L., Ruh, O., Merdinoglu, D., and Karst, F. (2005). Selection of hypervariable microsatellite loci for the characterization of Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 102, 73–83. doi: 10.1016/j.ijfoodmicro.2004.12.007
Li, S.-S., Cheng, C., Li, Z., Chen, J.-Y., Yan, B., Han, B.-Z., et al. (2010). Yeast species associated with wine grapes in China. Int. J. Food Microbiol. 138, 85–90. doi: 10.1016/j.ijfoodmicro.2010.01.009
Liu, H., Guo, J., Cheng, Y., Luo, L., Liu, P., Wang, B., et al. (2010a). Control of gray mold of grape by Hanseniaspora uvarum and its effects on postharvest quality parameters. Ann. Microbiol. 60, 31–35. doi: 10.1007/s13213-010-0018-3
Liu, H. M., Guo, J. H., Luo, L., Liu, P., Wang, B. Q., Cheng, Y. J., et al. (2010b). Improvement of Hanseniaspora uvarum biocontrol activity against gray mold by the addition of ammonium molybdate and the possible mechanisms involved. Crop Prot. 29, 277–282. doi: 10.1016/j.cropro.2009.10.020
Liu, P., Cheng, Y., Yang, M., Liu, Y., Chen, K., Long, C. A., et al. (2014). Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 14:242. doi: 10.1186/s12866-014-0242-2
Long, C.-A., Wu, Z., and Deng, B.-X. (2005). Biological control of Penicillium italicum of Citrus and Botrytis cinerea of grape by strain 34–9 of Kloeckera apiculata. Eur. Food Res. Technol. 221, 197–201. doi: 10.1007/s00217-005-1199-z
Manzanares, P., Ramón, D., and Querol, A. (1999). Screening of non-Saccharomyces wine yeasts for the production of β-D-xylosidase activity. Int. J. Food Microbiol. 46, 105–112. doi: 10.1016/S0168-1605(98)00186-X
Mariette, S., Chagné, D., Lézier, C., Pastuszka, P., Raffin, A., Plomion, C., et al. (2001). Genetic diversity within and among Pinus pinaster populations: comparison between AFLP and microsatellite markers. Heredity 86, 469–479. doi: 10.1046/j.1365-2540.2001.00852.x
Masneuf-Pomarede, I., Juquin, E., Miot-Sertier, C., Renault, P., Laizet, Y. H., Salin, F., et al. (2015). The yeast Starmerella bacillaris (synonym Candida zemplinina) shows high genetic diversity in winemaking environments. FEMS Yeast Res. 15:fov045. doi: 10.1093/femsyr/fov045
Masoud, W., Bjørg Cesar, L., Jespersen, L., and Jakobsen, M. (2004). Yeast involved in fermentation of Coffea arabica in East Africa determined by genotyping and by direct denaturating gradient gel electrophoresis. Yeast 21, 549–556. doi: 10.1002/yea.1124
Mateo, J. J., Jimenez, M., Huerta, T., and Pastor, A. (1991). Contribution of different yeasts isolated from musts of monastrell grapes to the aroma of wine. Int. J. Food Microbiol. 14, 153–160. doi: 10.1016/0168-1605(91)90102-U
Mehlomakulu, N. N., Setati, M. E., and Divol, B. (2014). Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int. J. Food Microbiol. 188, 83–91. doi: 10.1016/j.ijfoodmicro.2014.07.015
Mendoza, L. M., Manca de Nadra, M. C., and Farías, M. E. (2007). Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 29, 1057–1063. doi: 10.1007/s10529-007-9355-0
Mendoza, L., Nadra, M. M., Bru, E., and Farías, M. (2009). Influence of wine-related physicochemical factors on the growth and metabolism of non-Saccharomyces and Saccharomyces yeasts in mixed culture. J. Ind. Microbiol. Biotechnol. 36, 229–237. doi: 10.1007/s10295-008-0489-4
Merritt, A. E., and Hurley, R. (1972). Evaluation of sporulation media for yeasts obtained from pathological material. J. Med. Microbiol. 5, 21–30. doi: 10.1099/00222615-5-1-21
Mocke, B. A. (2005). The Breeding of Yeast Strains for Novel Oenological Outcomes. Ph.D. thesis, Stellenbosch University.
Morais, P. B., Martins, M. B., Klaczko, L. B., Mendonça-Hagler, L. C., and Hagler, A. N. (1995). Yeast succession in the Amazon fruit Parahancornia amapa as resource partitioning among Drosophila spp. Appl. Environ. Microbiol. 61, 4251–4257.
Morais, P. B., Rosa, C. A., Linardi, V. R., Pataro, C., and Maia, A. B. R. A. (1997). Short communication: characterization and succession of yeast populations associated with spontaneous fermentations during the production of Brazilian sugar-cane aguardente. World J. Microbiol. Biotechnol. 13, 241–243. doi: 10.1023/A:1018558302062
Moreira, N., Mendes, F., Guedes de Pinho, P., Hogg, T., and Vasconcelos, I. (2008). Heavy sulphur compounds, higher alcohols and esters production profile of Hanseniaspora uvarum and Hanseniaspora guilliermondii grown as pure and mixed cultures in grape must. Int. J. Food Microbiol. 124, 231–238. doi: 10.1016/j.ijfoodmicro.2008.03.025
Moreira, N., Pina, C., Mendes, F., Couto, J. A., Hogg, T., and Vasconcelos, I. (2011). Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Cont. 22, 662–667. doi: 10.1016/j.foodcont.2010.07.025
Nguyen, N. H., Suh, S. O., and Blackwell, M. (2007). Five novel Candida species in insect-associated yeast clades isolated from Neuroptera and other insects. Mycologia 99, 842–858. doi: 10.3852/mycologia.99.6.842
O'Donnell, K. (1993). “Fusarium and its near relatives,” in The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics. Wallingford, UK: CAB International.
Owuama, C. I., and Saunders, J. R. (1990). Physiological variants of Saccharomyces cerevisiae and Kloeckera apiculata from palm wine and cashew juice. J. Appl. Bacteriol. 68, 491–494. doi: 10.1111/j.1365-2672.1990.tb02901.x
Pagnocca, F. G., Mendonca-Hagler, L. C., and Hagler, A. N. (1989). Yeasts associated with the white shrimp Penaeus schmitti, sediment, and water of Sepetiba Bay, Rio de Janeiro, Brasil. Yeast 5 Spec No, S479–S483.
Pando Bedrinana, R., Lastra Queipo, A., and Suarez Valles, B. (2012). Screening of enymatic activities in non-Saccharomyces cider yeasts. J. Food Biochem. 36, 683–689. doi: 10.1111/j.1745-4514.2011.00583.x
Paradis, E. (2010). Pegas: an R package for population genetics with an integrated-modular approach. Bioinformatics 26, 419–420. doi: 10.1093/bioinformatics/btp696
Parker, P. G., Snow, A. A., Schug, M. D., Booton, G. C., and Fuerst, P. A. (1998). What molecules can tell us about populations: choosing and using a molecular marker. Ecology 79, 361–382. doi: 10.2307/176939
Pu, L., Jingfan, F., Kai, C., Chao-an, L., and Yunjiang, C. (2014). Phenylethanol promotes adhesion and biofilm formation of the antagonistic yeast Kloeckera apiculata for the control of blue mold on citrus. FEMS Yeast Res. 14, 536–546. doi: 10.1111/1567-1364.12139
Pulvirenti, A., Giudici, P., zucchi, P., Desalvo, F., Marcazzan, G., and Sabatini, A. G. (2009). Qualitative aspects of spoiled honey. Ingredienti Alimentari 8, 6–14.
Radler, F., Pfeiffer, P., and Dennert, M. (1985). Killer toxins in new isolates of the yeasts Hanseniaspora uvarum and Pichia kluyveri. FEMS Microbiol. Lett. 29, 269–272. doi: 10.1111/j.1574-6968.1985.tb00874.x
Radler, F., Schmitt, M. J., and Meyer, B. (1990). Killer toxin of Hanseniaspora uvarum. Arch. Microbiol. 154, 175–178. doi: 10.1007/BF00423329
R Development Core Team (2010). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Redzepovic, S., Orlic, S., Majdak, A., Kozina, B., Volschenk, H., and Viljoen-Bloom, M. (2003). Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int. J. Food Microbiol. 83, 49–61. doi: 10.1016/S0168-1605(02)00320-3
Reid, V. J., Theron, L. W., du Toit, M., and Divol, B. (2012). Identification and partial characterization of extracellular aspartic protease genes from Metschnikowia pulcherrima IWBT Y1123 and Candida apicola IWBT Y1384. Appl. Environ. Microbiol. 78, 6838–6849. doi: 10.1128/AEM.00505-12
Renard, A., Gómez di Marco, P., Egea-Cortines, M., and Weiss, J. (2008). Application of whole genome amplification and quantitative PCR for detection and quantification of spoilage yeasts in orange juice. Int. J. Food Microbiol. 126, 195–201. doi: 10.1016/j.ijfoodmicro.2008.05.021
Romano, P., Palla, G., Caligiani, A., Brandolini, V., Maietti, A., and Salzano, G. (2000). Evaluation of stereoisomers of 2,3-butanediol and acetoin to differentiate Saccharomyces cerevisiae and Kloeckera apiculata wine strains. Biotechnol. Lett. 22, 1947–1951. doi: 10.1023/A:1026741625019
Romano, P., Suzzi, G., Zironi, R., and Comi, G. (1993). Biometric study of acetoin production in Hanseniaspora guilliermondii and Kloeckera apiculata. Appl. Environ. Microbiol. 59, 1838–1841.
Ross, K. G., Shoemaker, D. D., Krieger, M. J., DeHeer, C. J., and Keller, L. (1999). Assessing genetic structure with multiple classes of molecular markers: a case study involving the introduced fire ant Solenopsis invicta. Mol. Biol. Evol. 16, 525–543. doi: 10.1093/oxfordjournals.molbev.a026134
Schmitt, M. J., and Neuhausen, F. (1994). Killer toxin-secreting double-stranded RNA mycoviruses in the yeasts Hanseniaspora uvarum and Zygosaccharomyces bailii. J. Virol. 68, 1765–1772.
Schuelke, M. (2000). An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 18, 233–234. doi: 10.1038/72708
Schütz, M., and Gafner, J. (1993). Analysis of yeast diversity during spontaneous and induced alcoholic fermentations. J. Appl. Bacteriol. 75, 551–558. doi: 10.1111/j.1365-2672.1993.tb01594.x
Shimodaira, H. (2002). An approximately unbiased test of phylogenetic tree selection. Syst. Biol. 51, 492–508. doi: 10.1080/10635150290069913
Sláviková, E., Vadkertiová, R., and Vránová, D. (2009). Yeasts colonizing the leaves of fruit trees. Ann. Microbiol. 59, 419–424. doi: 10.1007/BF03175125
Sosa, O., de Nadra, M. M., and Farías, M. (2008). Modification by glucose of the flocculent phenotype of a Kloeckera apiculata wine strain. J. Ind. Microbiol. Biotechnol. 35, 851–857. doi: 10.1007/s10295-008-0357-2
Spencer, D. M., Spencer, J. F. T., De Figueroa, L., and Heluane, H. (1992). Yeasts associated with rotting citrus fruits in Tucumán, Argentina. Mycol. Res. 96, 891–892. doi: 10.1016/S0953-7562(09)81038-1
Strauss, M. L., Jolly, N. P., Lambrechts, M. G., and van Rensburg, P. (2001). Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 91, 182–190. doi: 10.1046/j.1365-2672.2001.01379.x
Suzuki, R., and Shimodaira, H. (2006). Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 22, 1540–1542. doi: 10.1093/bioinformatics/btl117
Suzzi, G., Schirone, M., Sergi, M., Marianella, R. M., Fasoli, G., Aguzzi, I., et al. (2012). Multistarter from organic viticulture for red wine Montepulciano d'Abruzzo production. Front. Microbiol. 3:135. doi: 10.3389/fmicb.2012.00135
Valles, B. S., Bedriñana, R. P., Tascón, N. F., Simón, A. Q., and Madrera, R. R. (2007). Yeast species associated with the spontaneous fermentation of cider. Food Microbiol. 24, 25–31. doi: 10.1016/j.fm.2006.04.001
van Wyk, H., and Divol, B. (2010). Recovery of endo-polygalacturonase activity in wine yeast and its effect on wine aroma. FEMS Yeast Res. 10, 58–71. doi: 10.1111/j.1567-1364.2009.00588.x
Velázquez, J. B., Longo, E., Sieiro, C., Cansado, J., Calo, P., and Villa, T. G. (1991). Improvement of the alcoholic fermentation of grape juice with mixed cultures of Saccharomyces cerevisiae wild strains. Negative effect of Kloeckera apiculata. World J. Microbiol. Biotechnol. 7, 485–489. doi: 10.1007/BF00303374
Versavaud, A., and Hallet, J.-N. (1995). Pulsed-field gel electrophoresis combined with rare-cutting endonucleases for strain differentiation of Candida famata, Kloeckera apiculata and Schizosaccharomyces pombe with dhromosome number and size estimation of the two former. Syst. Appl. Microbiol. 18, 303–309. doi: 10.1016/S0723-2020(11)80401-7
Wang, C., Esteve-Zarzoso, B., and Mas, A. (2014). Monitoring of Saccharomyces cerevisiae, Hanseniaspora uvarum, and Starmerella bacillaris (synonym Candida zemplinina) populations during alcoholic fermentation by fluorescence in situ hybridization. Int. J. Food Microbiol. 191, 1–9. doi: 10.1016/j.ijfoodmicro.2014.08.014
Wang, C., Mas, A., and Esteve-Zarzoso, B. (2015). Interaction between Hanseniaspora uvarum and Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 206, 67–74. doi: 10.1016/j.ijfoodmicro.2015.04.022
Weiss, S., Samson, F., Navarro, D., and Casaregola, S. (2013). YeastIP: a database for identification and phylogeny of Saccharomycotina yeasts. FEMS Yeast Res 13, 117–125. doi: 10.1111/1567-1364.12017
White, T. J., Bruns, T. D., Lee, S. B., and Taylor, J. W. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR Protocols: A Guide to Methods and Applications, eds M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White (London: Academic Press), 315–322.
Wiles, A. E. (1950). Studies of some yeasts causing spoilage of draught beers. J. Inst. Brew. 56, 183–193. doi: 10.1002/j.2050-0416.1950.tb01531.x
Zagorc, T., Maráz, A., Cadez, N., Jemec, K. P., Péter, G., Resnik, M., et al. (2001). Indigenous wine killer yeasts and their application as a starter culture in wine fermentation. Food Microbiol. 18, 441–451. doi: 10.1006/fmic.2001.0422
Zironi, R., Romano, P., Suzzi, G., Battistutta, F., and Comi, G. (1993). Volatile metabolites produced in wine by mixed and sequential cultures of Hanseniaspora guilliermondii or Kloeckera apiculata and Saccharomyces cerevisiae. Biotechnol. Lett. 15, 235–238. doi: 10.1007/BF00128311
Zohre, D. E., and Erten, H. (2002). The influence of Kloeckera apiculata and Candida pulcherrima yeasts on wine fermentation. Proc. Biochem. 38, 319–324. doi: 10.1016/S0032-9592(02)00086-9
Keywords: Hanseniaspora uvarum, wine, intraspecific diversity, microsatellites, phenotypic screening
Citation: Albertin W, Setati ME, Miot-Sertier C, Mostert TT, Colonna-Ceccaldi B, Coulon J, Girard P, Moine V, Pillet M, Salin F, Bely M, Divol B and Masneuf-Pomarede I (2016) Hanseniaspora uvarum from Winemaking Environments Show Spatial and Temporal Genetic Clustering. Front. Microbiol. 6:1569. doi: 10.3389/fmicb.2015.01569
Received: 05 November 2015; Accepted: 27 December 2015;
Published: 20 January 2016.
Edited by:
Alberto Mas, University Rovira i Virgili, SpainReviewed by:
Maurizio Ciani, Università Politecnica delle Marche, ItalyMatthias Sipiczki, University of Debrecen, Hungary
Copyright © 2016 Albertin, Setati, Miot-Sertier, Mostert, Colonna-Ceccaldi, Coulon, Girard, Moine, Pillet, Salin, Bely, Divol and Masneuf-Pomarede. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Warren Albertin, warren.albertin@u-bordeaux.fr