 Kin-Kwan Lai
Kin-Kwan Lai Austin G. Davis-Richardson
Austin G. Davis-Richardson Raquel Dias
Raquel Dias Eric W. Triplett
Eric W. Triplett- Microbiology and Cell Science Department, Institute of Food and Agricultural Sciences, University of Florida, Gainesville, FL, USA
Here Tn5 random transposon mutagenesis was used to identify the essential elements for culturing Liberibacter crescens BT-1 that can serve as antimicrobial targets for the closely related pathogens of citrus, Candidatus Liberibacter asiaticus (Las) and tomato and potato, Candidatus Liberibacter solanacearum (Lso). In order to gain insight on the virulence, metabolism, and culturability of the pathogens within the genus Liberibacter, a mini-Tn5 transposon derivative system consisting of a gene specifying resistance to kanamycin, flanked by a 19-base-pair terminal repeat sequence of Tn5, was used for the genome-wide mutagenesis of L. crescens BT-1 and created an insertion mutant library. By analyzing the location of insertions using Sanger and Illumina Mi-Seq sequencing, 314 genes are proposed as essential for the culture of L. crescens BT-1 on BM-7 medium. Of those genes, 76 are not present in the uncultured Liberibacter pathogens and, as a result, suggest molecules necessary for the culturing these pathogens. Those molecules include the aromatic amino acids, several vitamins, histidine, cysteine, lipopolysaccharides, and fatty acids. In addition, the 238 essential genes of L. crescens in common with L. asiaticus are potential targets for the development of therapeutics against the disease.
Introduction
Citrus greening disease, also known as Huanglongbing disease (HLB), is a devastating disease of citrus worldwide. A phloem-restricted α-proteobacterium, namely Candidatus Liberibacter asiaticus (Las), is the casual agent of the disease (Bové, 2006; Tyler et al., 2009). Las is found in North America and Asia while two other species, Candidatus Liberibacter africanus (Laf) and Candidatus Liberibacter americanus (Lar), are found in Africa and Brazil respectively. HLB is naturally transmitted by the psyllid Diaphorina citri and is capable of infecting all known commercial varieties of citrus. The highest titer of Las is found in the peduncle and is unequally distributed in the phloem of infected plants (Tatineni et al., 2008). Some of the HLB symptoms include yellow shoots, asymmetric blotchy leaf mottle, thicken leaves, veins enlargement, twig dieback, small off-taste fruits, and premature fruit drops. Starch accumulation occurs in the phloem, which damages the phloem and prevents nutrients transport. The infected trees eventually die when the conditions are too severe (Wang and Trivedi, 2013). Currently there are no cures for HLB. Thermotherapy and chemotherapy are recently proposed but still under evaluation (Pagliai et al., 2015; Canales et al., 2016; Fan et al., 2016). Methods on controlling the HLB infection are based on chemical treatments on the psyllid vectors and complete removal of the infected trees (Gottwald et al., 2007). However, these preventative methods are not as efficient as the spreading of the disease.
During the past few years, genomes of several Liberibacter species were sequenced (Duan et al., 2009; Lin et al., 2011, 2015; Wulff et al., 2014). Genes involved in transcriptional regulation, major metabolic pathways, secretion/transportation system, motility and signal transduction were predicted as putative virulence factors (Cong et al., 2012; Yan et al., 2013). However, the inability to culture any of the Liberibacter pathogens limits our understanding of the mechanism of infection and delays the development of treatments for citrus greening disease. Liberibacter crescens BT-1 is a Gram-negative, rod-shaped, α-proteobacterium isolated from mountain papaya (Leonard et al., 2012; Fagen et al., 2014a). It is the closest cultured relative of the causal agents of citrus greening disease (Las, Laf, and Lar), and Ca. L. solanacearum (Lso), the causal agent of zebra chip on potato and tomato yellows (Fagen et al., 2014a,b). To date, a plant host for L. crescens has not been identified. In addition, L. crescens has not experienced far less genome reduction than the Liberibacter pathogens. These two observations suggest that L. crescens may not be a plant pathogen. L. crescens can serve as a model to understand the mechanism of infection and culturing of Liberibacter pathogens.
Genome-wide saturation mutagenesis has been used in several studies to identify the essential genes and pathways critical for the survival or pathogenesis of bacteria in culture or in association with a host (Akerley et al., 2002; Forsyth et al., 2002; Langridge et al., 2009; Moule et al., 2014). An essential gene is defined as one whose loss is lethal to an organism under the growth conditions of interest. Identifying the essential gene set allows us to not only understand the nutritional requirement for growth but also suggests potential targets for antimicrobial drug development (Lee et al., 2015).
Transposon mutagenesis is a powerful tool for producing randomized gene mutation in bacterial genomes. It has been utilized for the study of bacterial pathogenesis extensively and has been used to identify virulence factors in different bacteria (Mei et al., 1997; Hava and Camilli, 2002). Bacterial clones containing transposon insertions that significantly reduce fitness for in vitro growth will not survive. Using the genomic DNA from the mutant pools, Illumina nucleotide sequencing can be used to amplify from the transposon and sequence into the adjacent target DNA to identify the insertion sites in the genome. The distribution of transposon insertion sites thus provides a map of the genes and other elements essential for in vitro culturing, which can also serve as antimicrobial targets for the closely related pathogens (Miesel et al., 2003).
Here, the Tn5 transposon was used to mutagenize L. crescens BT-1 in order to predict gene essentiality. We believe that the predicted essential genes will be extremely useful for expanding the capacity of L. crescens BT-1 as a model organism to develop treatments for citrus greening disease.
Materials and Methods
Bacterial Strains and Culture Conditions
Escherichia coli donor strain SM17-1λpir carrying pUT-miniTn5Km1 plasmid (de Lorenzo et al., 1990) was cultured on Luria-Bertani (LB) liquid medium supplemented with 25 μg mL−1 kanamycin at 37°C with agitation at 250 rpm. L. crescens BT-1 was cultured on liquid BM7 medium (Fagen et al., 2014a) at 27°C with moderate agitation (125 rpm).
Preparation of L. crescens BT-1 Electrocompetent Cell
A single colony of L. crescens BT-1 was picked from BM7 agar plate and grown in 2 mL of BM7 medium until OD600 reached 0.3–0.4. It was used to inoculate 30 mL of BM7 medium. Cells were incubated on ice for 15 min and harvested by centrifugation (1400 x g, 15 min, 4°C) when OD600 reached 0.3–0.4. Cells were washed 2 times with 30 mL of ice-cold 10% glycerol. The competent cells were resuspended in 300 μL of ice-cold 10% glycerol and stored at −80°C in small aliquots.
Electro-Transformation of L. crescens BT-1 using EZ-Tn5 Transponsome
EZ-Tn5 < KAN-2>Tnp Transposome (20 ng, Epicentre, USA) was mixed with 50 μL of L. crescens BT-1 competent cells and transferred to a pre-chilled 0.1 cm gene pulser cuvette (Bio-Rad, USA). Electro-transformation was carried out using Bio-Rad Gene Pulser Xcell™ Electroporation System set to 1.8 kV, 25 μF, and 200 ω. Cells were immediately resuspended into 2 mL of BM7 medium and grown for 24 h. Cells were harvested by centrifugation (1400 x g, 5 min) and spread on BM7 agar plate supplemented with 5 μg mL−1 kanamycin. Cells were incubated at 27°C until colonies formed. Individual colonies were picked and inoculated into 2 mL of BM7 medium. Cells were harvested when OD600 reached 0.3–0.4 and suspended in 25% glycerol for storage and subsequent DNA extraction.
Transposon Mutagenesis of L. crescens BT-1 using pUTminiTn5km1
The pUTminiTn5Km1 transposon was delivered into L. crescens BT-1 by bi-parental mating with the E. coli donor strain SM17-1λpir carrying pUTminiTn5Km1 plasmid. Ten milliliter of L. crescens BT-1 culture (OD600 ~ 0.3) and 1 mL of E. coli donor culture were harvested by centrifugation. Cells were washed 2 times with 1 mL of 10 mM MgSO4 and were resuspended in the same volume of 10 mM MgSO4. One microliter of the washed E. coli donor cells was added to the 1 mL of the washed L. crescens BT-1 cells. The mixture was filtered through a sterile 25 mm-diameter Millipore type HA 0.45 μm filter (EMD Millipore, USA). The filter was transferred to a BM7 agar plate and incubated for 18 h at 27°C. The filter was transferred to 5 mL of BM7 supplemented with 10 mM MgSO4 to release cells from the filter and into the medium. Cells were harvested from 200 μL of the cell suspension by centrifugation and spread on BM7 agar plate supplemented with antibiotics (15 μg mL−1 nalidixic acid and 5 μg mL−1 kanamycin) to select for L. crescens BT-1 containing transposon insertions. Transformed cells were then incubated at 27°C until cell layer formed. Cells were scraped off the plate and grown in 5 mL of BM7 medium supplemented with antibiotics (15 μg mL−1 nalidixic acid and 5 μg mL−1 kanamycin). Next, cells were harvested at OD600 0.3–0.4 and then stored at −80°C in 25% glycerol.
Genomic DNA Extraction of L. crescens BT-1 Tn5 Mutants
Genomic DNA of Tn5 mutants were extracted using hexadecyltrimethylammonium bromide (CTAB) phenol chloroform extraction method. Cell pellets were resuspended in 567 μL TE buffer (10 mM Tris-HCl pH 8, 1 mM EDTA), 30 μL 10% SDS, and 3 μL proteinase K (20 mg mL−1 in TE buffer) and incubated for 1 h at 37°C. Cells were mixed thoroughly after addition of 100 μL 5 M NaCl and 80 μL of CTAB-NaCl (10% CTAB, 0.7 M NaCl). After the mixture was incubated at 65°C for 10 min, 780 μL of chloroform/isoamyl alcohol (24:1, v/v) was added, gently mixed, and centrifuged at 15,000 x g for 5 min. The supernatant was transferred to a new tube followed by addition of an equal volume of phenol/chloroform/isoamyl alcohol (25:24:1, v/v/v) which was then, gently mixed, and centrifuged at 15000 x g for 5 min. To the supernatant, 0.6 volume of isopropanol was added, gently mixed, and stored at −20°C overnight. The DNA pellet was harvested by centrifugation at 15,000 x g for 5 min. The pellet was washed with 700 μL of 70% ethanol and centrifuged. The ethanol was discarded. The pellet was allowed to air dry for 10 min and rehydrated in 100 μL of water. The concentration of DNA was determined with a qubit fluorometer (Life Technologies, USA).
Determination of EZ-Tn5 L. crescens BT-1 Insertion Site
Single primer polymerase chain reaction (sp-PCR) was used to identify the EZ-Tn5 insertion site (Karlyshev et al., 2000). Primer inv-1 (ATGGCTCATAACACCCCTTGTATTA) or inv-2 (GAACTTTTGCTGAGTTGAAGGATCA) was used in the PCR with GoTaq® G2 DNA Polymerase (Promega, USA). DNA was amplified with 5 min at 95°C followed by 30 cycles of 30 s at 95°C, 30 s at 56°C, and 30 s at 72°C; 30 cycles of 30 s at 95°C, 30 s at 30°C, and 30 s at 72°C; 30 cycles of 30 s at 95°C, 30 s at 56°C, and 2 min at 72°C; and 10 min at 72°C. The PCR products were purified using Qiagen QIAquick PCR purification kit. Sanger sequencing was performed using primer FP1 (ACCTACAACAAAGCTCTCATCAACC, for inv-2 PCR product) or RP1 (GCAATGTAACATCAGAGATTTTGAG, for inv-1 PCR product). The insertion sites were determined by homology to the L. crescens genome using NCBI Blastn. A total of 2070 high quality sequences of the EZ-Tn5 insertion sites are provided as Table S1.
Illumina Sequencing of pUTminiTn5km1 L. crescens BT-1
Equal amounts of genomic DNA from each batch of pUTminiTn5Km1 L. crescens BT-1 mutants were pooled together. Pooled DNA (5 μg) was fragmented to an average size of 300 bp by ultrasonication. The fragments were further size selected to 250–350 bp by gel excision. The fragmented DNA was purified, end repaired, and A-tailed according to Illumina library preparation protocol. Two single-stranded primers Ind_Ad_T (ACACTCTTTCCCTACACGACGCTCTTCCGATC*T) and Ind_Ad_B (pGATCGGAAGAGCGGTTCAGCAGGAATGCCGAGACCGATCTC) were annealed together to form the adapter. The adapter was ligated to the fragmented DNA. The DNA was quantified with an Agilent NAD1000 chip and qubit fluorometer. PCR was preformed to enrich the fragments containing the transposon insertion sites using primer RInV3.3 (CAAGCAGAAGACGGCATACGAGATCGGTACACTCTTTCCCTACACGACGCTCTTCCGATCT) and primer MiniTn5_P5_3pr_3 (AATGATACGGCGACCACCGAGATCTACACCTAGGCtGCGGCtGCACTTGTG) with Phusion® High-Fidelity DNA Polymerase (NEB, USA). PCR amplification conditions included 2 min at 98°C; 22 cycles of 30 s at 98°C, 30 s at 65°C, 30 s at 72°C; and 10 min at 72°C. Amplified library was cleaned using Qiagen QIAquick PCR purification kit. DNA fragment library was sequenced using 1 × 50 cycles Illumina flow cell on an Illumina MiSeq sequencer with a custom Tn5 sequencing primer Tn5_ill_seq (CCTAGGCtGCGGCtGCACTTGTG), which is designed such that the first 12 bp of each read is the end 12 bp of the transposon sequence.
Analysis of the pUTminiTn5km1 Sequence Data
The 12-bp transposon tag (TATAAGAGTCAG) was trimmed from the raw Illumina sequence data. Reads were mapped to the L. crescens BT-1 chromosome (GI: 430799321) using CLC Workbench version 6.5 (CLC bio, USA). A custom script was used to count the number of insertions by gene and is available online (https://github.com/triplett/lai-2015). The total number of high quality sequences obtained from these insertion events was 16,185,251. Raw sequence data can be accessed at NCBI sequence read archive with study accession number SRP065838.
Statistical Analysis
The probability of having 1 insertion in each of the genes was calculated using the poisson distribution equation (Watabe et al., 2014) as follow:
N is the number of insertions needed; P is the probability of having 1 insertion in each of the genes; f is the frequency of insertion, which is the average gene size divided by the genome size. The probability of finding 1 insertion within a particular gene was calculated using neutral base-pair model (Laia et al., 2009):
n is the number of insertions; P is the probability of finding 1 insertion within a particular gene; X is the length of the gene; G is the length of the genome.
Gene Comparison and Functional Categorization of Essential Genes
Gene annotations were acquired from NCBI. Essential gene homologs were identified by manual NCBI BLAST searches. Genes were classified into functional groups/subsystems using Rapid Annotation Subsystem Technology (RAST; Overbeek et al., 2014).
Results
Tn5 Transposon Mutagenesis of L. crescens BT-1
L. crescens BT-1 genome has 1379 open reading frames and 54 RNA-coding genes (Leonard et al., 2012). It has a genome size of 1504659 bp and an average gene length of 895.3 bp. Poisson distribution equation was used to estimate the number of insertions or mutants required to saturate the genome to have at least 1 insertion per gene as seen in Table 1. Two methods were used for the Tn5 transposon mutagenesis of L. crescens BT-1: electro-transformation with the commercial EZ-Tn5 Transposome (EZ-Tn5) and bi-parental mating with the E. coli donor strain SM17-1λpir harboring pUTminiTn5Km1 plasmid (pUT-Tn5). Approximately 50–100 colonies (2.5 × 103–5 × 103 transformants/μg of transposon DNA) were formed in each successful electro-transformation with the EZ-Tn5 method. A total of 2205 EZ-Tn5 colonies were isolated. Tn5 insertion sites were identified in 2070 (93.9%) EZ-Tn5 isolated mutants using sp-PCR and Sanger sequencing. Of those, 188 unique insertions were mapped to intergenic regions and 1355 unique insertions were mapped to 547 ORFs and 5 RNA-coding genes of the L. crescens BT-1 genome (Table S2).
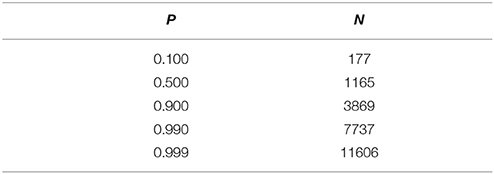
Table 1. Probability estimation of having at least 1 insertion per gene in L. crescens mutagenesis with poisson distribution equation.
In order to achieve genome-wide saturation mutagenesis of L. crescens BT-1, a more robust mutation method was required to generate a larger number of mutants. A thin cell layer was formed on the BM7 agar plate supplemented with nalidixic acid and kanamycin in each successful bi-parental mating with the pUT-Tn5 method. Each batch resulted in 500–1000 colonies as determined by the standard serial dilution plate count method (data not shown). A total of 40 batches of pUT-Tn5 mutants were made independently. Illumina Mi-Seq sequencing analysis identified 4250 unique insertions. Of those 3591 were mapped to 835 ORFs and 11 RNA-coding genes respectively of the L. crescens BT-1 genome (Table S3). The distribution of mapped sequence reads across the whole genome is shown in Figure 1.
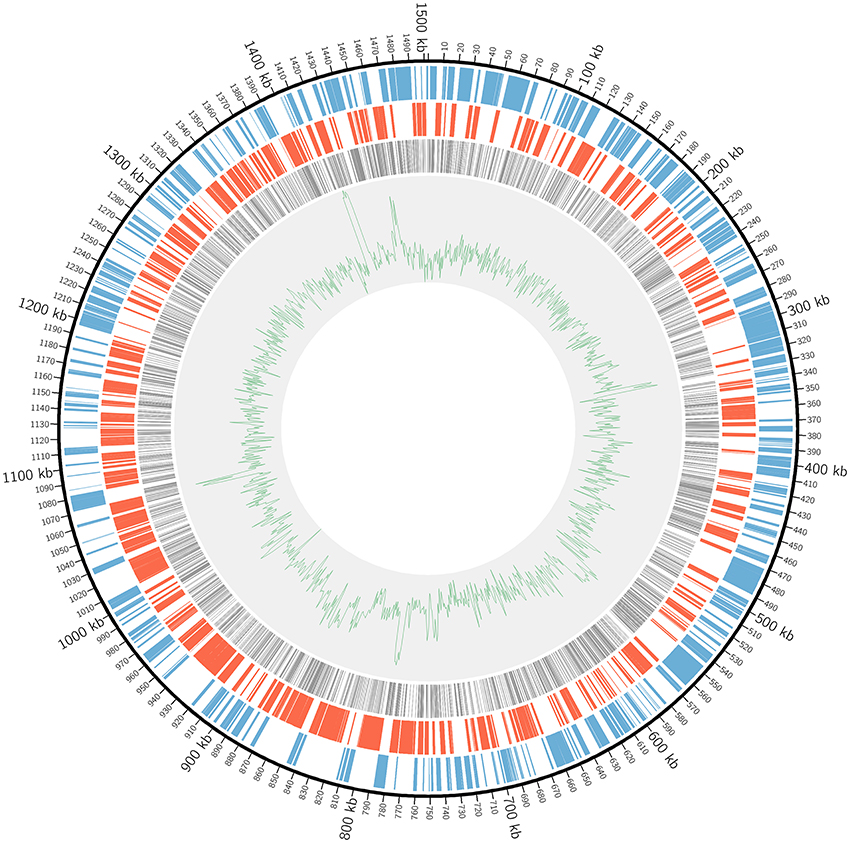
Figure 1. The distribution of Tn5 insertions in L. crescens BT-1 genome. The black circular line represents the genome sequence with the origin of replication at coordinate zero. The blue bars represent genes transcribed clockwise. The red bars represent genes transcribed counterclockwise. The gray bars represent Tn5 insertions within ORFs. The green line represents the GC plot.
Identification of Candidate Essential Genes
Combining both mutation protocols, Tn5 insertions were detected in 994 protein-coding genes and 13 RNA-coding genes from the L. crescens genome. Tn5 insertions were not detected in 385 protein-coding and 41 RNA-coding genes, suggesting these genes are essential for the growth of L. crescens on BM7 medium (Table S4). A fair number of these essential genes code for small hypothetical proteins. We questioned whether these are truly essential as small gene is less likely to have a mutation than a large gene. Neutral base-pair model allows us to estimate how many genes in this set are truly essential and how many were not hit because of chance alone. It assumes that every base pair in the genome has the same probability of containing an insertion. The gene is less likely to be essential when P < 0.5. The neutral base-pair model estimates that 71 protein-coding genes and all 41 RNA-coding genes in the essential gene set were not disrupted because of chance (Table S4), which reduces the essential protein-coding genes set to 314. With 4938 unique insertions within ORFs and 1433 protein- and RNA-coding genes in the genome, there was an average of 3.45 unique insertions per gene. Excluding the 314 protein-coding genes with no insertions, there was an average of 4.41 unique insertions per putatively non-essential gene.
Functional Classification of Candidate Essential Genes
Genes essential for growth make excellent drug target since inhibiting their function will likely lead to lethality. Here, a total of 314 protein-coding genes are proposed as the essential gene set for L. crescens. Of the essential protein-coding genes, 74 code for hypothetical genes with no known functions. Most of the annotated essential genes are involved in diverse core cellular processes such as DNA and protein metabolism, transporters, cell division, regulation, and cell wall/membrane biogenesis (Table S4).
The essential gene set was further classified using RAST. Unlike the Genbank annotation pipeline which provides a specific annotation for each gene, the RAST pipeline classifies proteins into functional groups called subsystems. RAST was used previously in the comparative genomics of Liberibacter species (Fagen et al., 2014a). A functional comparison was done between the essential gene set and the L. crescens genome (Table S5). In that analysis, a far great proportion of unclassified genes is seen in the essential gene set (43%) than in the whole genome (28.1%). Also in the essential gene set a higher proportion of genes involved in fatty acid, lipid, and isoprenoid metabolism and a much lower proportion of genes with roles in amino acid, RNA, and carbohydrate metabolism as well as cell wall synthesis, motility, chemotaxis, and stress response.
Of the 314 essential protein-coding genes, 238 have homologs in Las (CEGs-Las+). These 238 genes are potentially excellent candidates for drug targeting against Las (Table S4). As expected, only 28 of these 238 genes (12%) are hypothetical as most of these would be expected to play a role in core metabolism or essential transport functions.
The remaining 76 protein-coding essential genes of L. crescens have no homologs in Las (CEGs-Las−) and these genes should help us understand why L. crescens is cultured while Las has not yet been cultured (Table S6). Of those, 46 genes (61%) are hypothetical which is expected given that we have not yet solved the riddle of culturing Las.
As expected, homologs of both Lso and Las are highly conserved in the L. crescens essential protein-coding genes set with a small different. Of the 314 essential protein-coding genes, 242 have homologs and 72 have no homologs in Lso (Tables S7, S8).
The subsystem categories of CEGs-Las+ and CEGs-Las− protein sets are divided into functional categories including amino acid synthesis and uptake, cofactor and vitamin synthesis and uptake prosthetic group synthesis, pigment formation, gene regulation (Figure 2). These results suggest that Las may be more reliant on the uptake of available resources from the environment than L. crescens. The fact that most of the genes are categorized as unclassified suggests that the functions of these genes are still poorly understood.
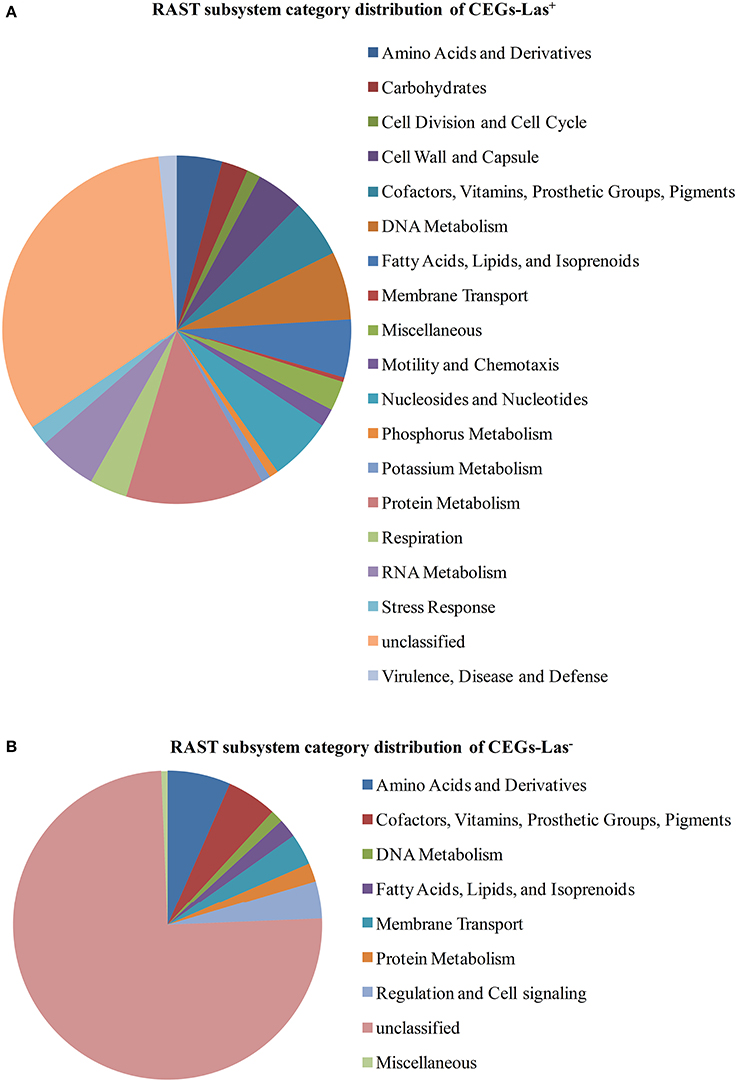
Figure 2. RAST subsystem category distribution of CEGs. CEGs were classified into different category using RAST. Unclassified group contains proteins with no known metabolic involvement and putative/hypothetical proteins. (A) CEGs-Las+. (B) CEGs-Las−.
Essential Gene Set and Las Gene Expression Profile Comparison
Genes involved in bacterial adaptation to the host environment during infection in plant pathogenic bacteria are known to be up-regulated (Chowdhury et al., 1996). A study by Yan showed the relative expression of 381 Las genes in planta versus in psyllid (Yan et al., 2013). Several of these up-regulated Las genes are homologs of L. crescens essential genes as shown in Table 2.
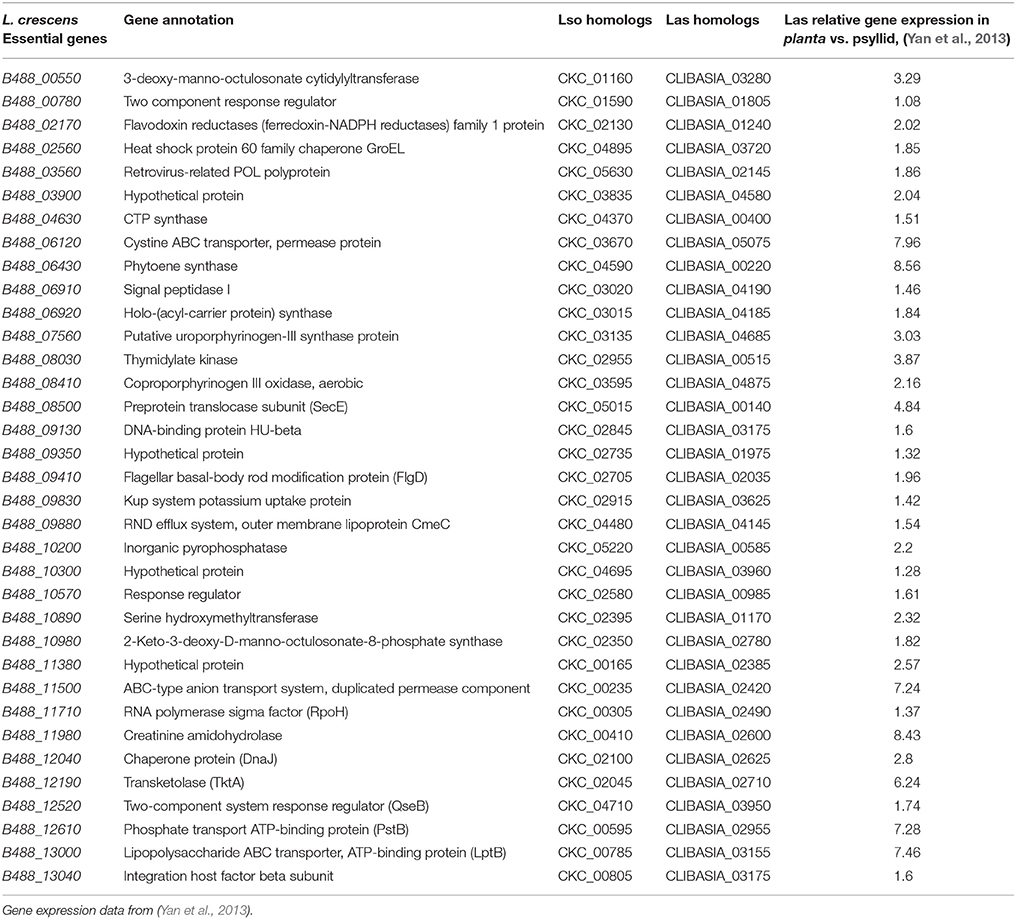
Table 2. Essential genes of L. crescens with homologs in Las and Lso that are up-regulated in Las in citrus.
Discussion
Functional Comparison between the Essential Gene Set and the Entire L. crescens Genome
The much higher proportion of unclassified genes in the essential gene set compared to the whole genome was a surprise. The vast majority of the essential genes were expected to be core metabolism genes that are already well understood but a large number are of unknown function with many of those also with homologs in the two uncultured Ca. Liberibacter plant pathogens. Clearly, much of the metabolism of fastidious bacteria that interact with plants and insects is largely unknown. Not surprisingly, the rich medium used for the culture of L. crescens in this work, precludes the requirement for many of the genes involved in amino acid, carbohydrate, and RNA metabolism.
Implications of the L. crescens Essential Gene Set for Las Culturing
Despite vigorous efforts to culture Las, there is still no reproducible, robust culturing method available. This greatly hinders research on understanding the pathogenesis of this organism as well as limit progress in treating infected citrus trees. L. crescens is the sole culturable member of the Liberibacter genus (Fagen et al., 2014a). The stream lined genome of Las possesses 356 fewer genes than does that of L. crescens genome (Fagen et al., 2014b). The majority of the L. crescens essential genes that are unique to L. crescens encode hypothetical proteins.
Among the essential genes in L. crescens that are absent in Las are genes that code for proteins involved in stress responses. Our hypothesis here is the plant and insect habitats of Las do not expose this pathogen to some of the stresses that L. crescens must survive in culture. For example, B488_09700 encodes protein-L-isoaspartate O-methyltransferase (PIMT). One of the several processes that can damage proteins and affect the cellular functioning is the formation of L-isoaspartyl residues on proteins. PIMT acts as a protein repair enzyme, converting the abnormal L-isoaspartyl residues back to the normal L-aspartyl residues (Clarke, 2003). Oxidative damage is another process that inactive proteins. Reactive oxygen species (ROS) and reactive nitrogen intermediates (RNI) are generated as natural byproducts during cell metabolism and stress responses. These byproducts can damage proteins by oxidizing the methionine residues, forming methionine sulfoxide. B488_10230 encodes peptide methionine sulfoxide reductase (MsrA), which catalyzes the reduction of methionine sulfoxide residues in proteins to methionine during oxidative damage (Weissbach et al., 2002). Low dosage of anti-oxidants such as ascorbic acid and glutathione can be added to the culture medium to protect the cells (La Scola et al., 2014). Both processes prevent the accumulation of potentially dysfunctional proteins and avoid the unnecessary protein degradation and resynthesis, indicating L. crescens has a better mechanism on protein repairing than in L. asiaticus.
The functions of another set of genes missing in Las but required for the growth of L. crescens in BM7 medium can be replaced by simple additions to the medium such as high levels of folate and aromatic amino acids. Folate biosynthesis is an essential pathway to synthesize folate (an essential cofactor for cell metabolism) in most of the bacteria due to the fact that folate-dependent formylation of the initiator tRNA is a hallmark of bacterial translation (de Crécy-Lagard et al., 2007). However, genes code for 2-amino-4-hydroxy-6-hydroxymethyldihydropteridine pyrophosphokinase (B488_08090), a putative dihydroneopterin aldolase (B488_08100), and dihydropteroate synthase (B488_08110) are missing in Las but present in L. crescens and Lso. For the synthesis of aromatic amino acids, the L. crescens genome codes for indole-3-glycerol phosphate synthase (B488_03020), 2-keto-3-deoxy-D-arabino-heptulosonate-7-phosphate synthase II (B488_07360), cyclohexadienyl dehydrogenase (B488_11240), and 3-dehydroquinate synthase (B488_11320), which are involved in phenylalanine, tyrosine, and tryptophan biosynthesis/shikimate pathway. All of these are essential proteins for the growth of L. crescens on BM7 medium but absent in Las.
One of the L. crescens essential proteins is nicotinamidase (PncA, B488_07080). The function of PncA is to hydrolyze nicotinamide to nicotinic acid for the production of NAD+, maintaining NAD+ homeostasis (Gazzaniga et al., 2009). Although L. crescens has a niacin/nicotinic acid transporter NiaP (B488_08370), it was determined to be non-essential in this study. The result is in agreement to the fact that the composition of BM7 medium does not include nicotinic acid. NAD is an important cofactor that contributes to important biological processes in both prokaryote and eukaryote (Vrablik et al., 2009; Evans et al., 2010). It acts as electron carrier during redox reaction and even involves in posttranslational modifications. Las does not appear to have PncA nor NiaP; indicating nicotinic acid is not required for the growth of the bacterium.
However, the key to culturing Las may still lie within the majority of the genes encoding hypothetical proteins that are essential in L. crescens but also found in Las. Improved bacterial genome annotation is required to unravel unknown metabolic pathways that may be required for Las in culture.
Implications of the L. crescens Essential Gene Set for Las Inhibition
Inhibition of an essential function for growth is a common means to develop antibiotics against a pathogen of interest (Clatworthy et al., 2007). Since L. crescens BT-1 and Las are close relatives, it is not surprising that 238 essential genes of L. crescens have homologs in Las. The availability of a list of essential genes provides a significant advantage because it allows us to choose specific targets, such as enzymes and regulators, to introduce a lethal effect on Las. Antibiotics can be designed to target more than one essential gene and thus minimize the development of resistance. Genes involved in cell wall or membrane biosynthesis are important targets for inhibition of the growth of Las. The incomplete synthesis of cell walls or membranes may prevent the bacteria from resisting high osmotic pressure and high sucrose concentrations present within phloem. Several L. crescens essential proteins such as UDP-N-acetylglucosamine O-acyltransferase (B488_00500), tetraacyldisaccharide 4′-kinase (B488_02350), and 2-keto-3-deoxy-D-manno-octulosonate-8-phosphate synthase (B488_10980) are involved in lipopolysaccharide biosynthesis. Homolog of B488_10980 in Las (CLIBASIA_02780) was also up-regulated during infection in planta, indicating CLIBASIA_02780 is a promising target for growth inhibition. Effort has been shown to discover inhibitors for the biosynthetic pathway (Shapiro et al., 2013).
Other essential genes with homologs in Las [B488_01060 and B488_02950: enoyl-(acyl-carrier-protein) reductase; B488_01080: 3-hydroxydecanoyl-(acyl-carrier-protein) dehydratase; B488_03630: phosphate:acyl-ACP acyltransferase PlsX; B488_10110: acyl carrier protein AcpP; B488_10130: malonyl CoA-acyl carrier protein transacylase; B488_11350: acetyl-coenzyme A carboxyl transferase alpha chain] are involved in fatty acid synthesis. In E. coli, genes in the fatty acid biosynthetic pathway are essential (Egan and Russell, 1973; Zhang and Cronan, 1998). This pathway is a common target for novel antibacterials triclosan, isoniazid, thiolactomycin, and cerulenin (Heath and Rock, 2004).
Differences on Essential Gene Set Comparison between Las and Lso
Only five of the L. crescens essential gene homologs that are present in Las are absent in Lso while nine of the L. crescens essential genes found in Lso are absent in Las. All 14 of these genes are highly conserved. The essential genes uniquely missing in Las are involved in folate-dependent formylation of tRNA. The few differences observed in the comparison of the L. crescens essential gene set with both Las and Lso suggests that the inability to culture both pathogens may be very similar. Thus, we predict that a medium that can successfully culture Las will culture Lso.
Increased in planta Expression of L. crescens Essential Gene Homologs in Las Using Gene Expression Data from Yan et al. (2013)
In this study, 314 of L. crescens essential genes were suggested. A study of the relative expression of 381 Las genes in planta versus in psyllid is publicly available. We hypothesized that Las genes that are up-regulated in planta and are common in the L. crescens essential set are the most promising candidates for drug targeting. Within the 314 L. crescens essential genes, 35 Las homologs were up-regulated during the infection in planta (Table 2). Genes related to ABC transporter (CLIBASIA_05075, CLIBASIA_02420, CLIBASIA_02955, CLIBASIA_03155) were especially up-regulated, up to 7.96-fold. Transporters are known virulence factors in pathogenic bacteria, aiding in uptake of nutrients and metal ions (Li et al., 2012; Yan et al., 2013). Gene encoding lipopolysaccharide ABC transporter (CLIBASIA_03155), together with other lipopolysaccharide biosynthesis genes (CLIBASIA_03280, CLIBASIA_02780) were also up-regulated. Lipopolysaccharide is an essential component for cell wall synthesis, protecting the cells from the harsh environment. Antimicrobial compounds can be designed to target these genes, controlling HLB disease (Akula et al., 2012). The potential role in pathogenesis of the other genes in Table 2 is unknown. In addition, the up regulation of some of these genes in the citrus may be less important than their down regulation for survival of Las in the psyllid.
Summary
The essential gene set of L. crescens described here provides suggestions for both culturing (essential genes that are unique to L. crescens) and inhibition of Las (essential genes that are common in Liberibacter pathogens). A general workflow of this study is summarized in Figure 3. However, there are limitations to this approach. First, it only describes those genes that are essential for growth on BM7 medium. Second, some of the putative essential genes may simply have escaped mutagenesis by chance, which would suggest that our mutagenesis did not saturate the genome. Third, some of the insertions may not have interrupted gene function. Nevertheless, the number of genes in our 314 essential gene set is larger than those observed in E. coli, with 295 essential genes (Juhas et al., 2014), and Bacillus subtilis, with 261 essential genes (Commichau et al., 2013).
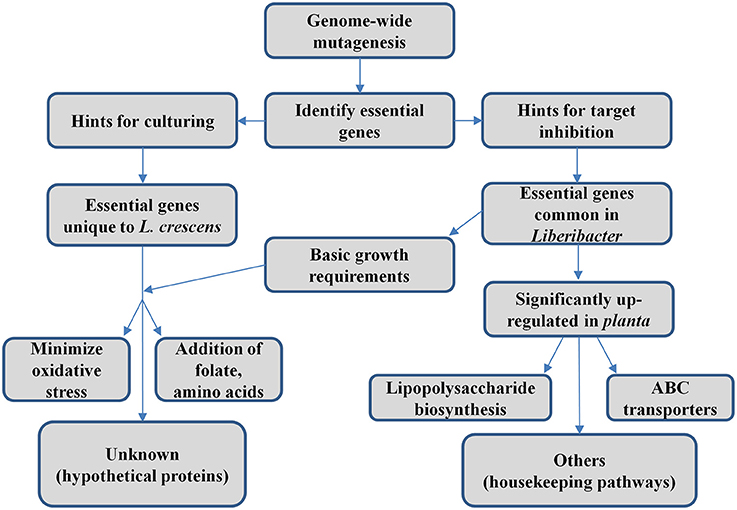
Figure 3. Conceptual diagram of work summary.
Given that the study of the genetics and biochemistry of L. crescens is in its infancy, it is not surprising that the current list of essential genes in L. crescens is larger compared to those of the model systems of E. coli and B. subtilis. The essential gene sets of both model systems have reduced in size as our knowledge grows (Juhas et al., 2014). However, the growth conditions and physiology of the three bacteria are very different so that each bacterium uses its own specific set of genes to survive in a particular growth medium. Indeed, some of the essentials genes in E. coli are not present in the L. crescens genome at all. Of the 295 protein-coding genes in the E. coli essential gene set, 39 have no homologs in L. crescens.
The gene essentiality is specific to the experimental growth condition and likely differs from those genes required in the natural habitat of L. crescens. The results described here provide an approximation of the essential elements for L. crescens BT-1 growth in vitro.
In summary, beside the nutrients in BM7 that support the growth of L. crescens, the study herein suggests folic acid and amino acids such as phenylalanine, tyrosine, and tryptophan should be supplemented when formulating a growth medium for Las. Our results are consistent with the finding as in (Fagen et al., 2014b). The majority of hypothetical proteins in the essential gene set still hinder the culturing of Las. In order to cure HLB, ABC transporter, and lipopolysaccharide biosynthesis genes should be used as the primary targets for developing antimicrobials. Triclosan, isoniazid, thiolactomycin, and cerulenin are novel antimicrobials that can inhibit the fatty acid biosynthetic pathway. More insights are needed from worldwide citrus researchers to fully solve the riddle of HLB.
Author Contributions
All authors listed, have made substantial, direct, and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Ronald Canepa for editing Figure 1. This research was supported by the Citrus Research and Development Foundation projects 767 and 769 and the University of Florida's Institute of Food and Agricultural Sciences.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00547
References
Akerley, B. J., Rubin, E. J., Novick, V. L., Amaya, K., Judson, N., and Mekalanos, J. J. (2002). A genome-scale analysis for identification of genes required for growth or survival of Haemophilus influenzae. Proc. Natl. Acad. Sci. U.S.A. 99, 966–971. doi: 10.1073/pnas.012602299
Akula, N., Trivedi, P., Han, F. Q., and Wang, N. (2012). Identification of small molecule inhibitors against SecA of Candidatus Liberibacter asiaticus by structure based design. Eur. J. Med. Chem. 54, 919–924. doi: 10.1016/j.ejmech.2012.05.035
Bové, J. M. (2006). Huanglongbing: a destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 88, 7–37. doi: 10.4454/jpp.v88i1.828
Canales, E., Coll, Y., Hernández, I., Portieles, R., Rodríguez García, M., López, Y., et al. (2016). ‘Candidatus Liberibacter asiaticus,’ causal agent of citrus huanglongbing, is reduced by treatment with brassinosteroids. PLoS ONE 11:e0146223. doi: 10.1371/journal.pone.0146223
Chowdhury, R., Sahu, G. K., and Das, J. (1996). Stress response in pathogenic bacteria. J. Biosci. 21, 149–160. doi: 10.1007/BF02703105
Clarke, S. (2003). Aging as war between chemical and biochemical processes: protein methylation and the recognition of age-damaged proteins for repair. Ageing Res. Rev. 2, 263–285. doi: 10.1016/S1568-1637(03)00011-4
Clatworthy, A. E., Pierson, E., and Hung, D. T. (2007). Targeting virulence: a new paradigm for antimicrobial therapy. Nat. Chem. Biol. 3, 541–548. doi: 10.1038/nchembio.2007.24
Commichau, F. M., Pietack, N., and Stülke, J. (2013). Essential genes in Bacillus subtilis: a re-evaluation after ten years. Mol. Biosyst. 9, 1068–1075. doi: 10.1039/c3mb25595f
Cong, Q., Kinch, L. N., Kim, B. H., and Grishin, N. V. (2012). Predictive sequence analysis of the Candidatus Liberibacter asiaticus proteome. PLoS ONE 7:e41071. doi: 10.1371/journal.pone.0041071
de Crécy-Lagard, V., El Yacoubi, B., de la Garza, R. D., Noiriel, A., and Hanson, A. D. (2007). Comparative genomics of bacterial and plant folate synthesis and salvage: predictions and validations. BMC Genomics 8:245. doi: 10.1186/1471-2164-8-245
de Lorenzo, V., Herrero, M., Jakubzik, U., and Timmis, K. N. (1990). Mini-Tn5 transposon derivatives for insertion mutagenesis, promoter probing, and chromosomal insertion of cloned DNA in gram-negative eubacteria. J. Bacteriol. 172, 6568–6572.
Duan, Y., Zhou, L., Hall, D. G., Li, W., Doddapaneni, H., Lin, H., et al. (2009). Complete genome sequence of citrus huanglongbing bacterium, ‘Candidatus Liberibacter asiaticus’ obtained through metagenomics. Mol. Plant Microbe Interact. 22, 1011–1020. doi: 10.1094/MPMI-22-8-1011
Egan, A. F., and Russell, R. R. (1973). Conditional mutations affecting the cell envelope of Escherichia coli K-12. Genet. Res. 21, 139–152. doi: 10.1017/S001667230001332X
Evans, C., Bogan, K. L., Song, P., Burant, C. F., Kennedy, R. T., and Brenner, C. (2010). NAD+ metabolite levels as a function of vitamins and calorie restriction: evidence for different mechanisms of longevity. BMC Chem. Biol. 10:2. doi: 10.1186/1472-6769-10-2
Fagen, J. R., Leonard, M. T., Coyle, J. F., McCullough, C. M., Davis-Richardson, A. G., Davis, M. J., et al. (2014a). Liberibacter crescens gen. nov., sp. nov., the first cultured member of the genus Liberibacter. Int. J. Syst. Evol. Microbiol. 64, 2461–2466. doi: 10.1099/ijs.0.063255-0
Fagen, J. R., Leonard, M. T., McCullough, C. M., Edirisinghe, J. N., Henry, C. S., Davis, M. J., et al. (2014b). Comparative genomics of cultured and uncultured strains suggests genes essential for free-living growth of Liberibacter. PLoS ONE 9:e84469. doi: 10.1371/journal.pone.0084469
Fan, G.-C., Xia, Y.-L., Lin, X.-J., Hu, H.-Q., Wang, X.-D., Ruan, C.-Q., et al. (2016). Evaluation of thermotherapy against Huanglongbing (citrus greening) in the greenhouse. J. Integrat. Agric. 15, 111–119. doi: 10.1016/S2095-3119(15)61085-1
Forsyth, R. A., Haselbeck, R. J., Ohlsen, K. L., Yamamoto, R. T., Xu, H., Trawick, J. D., et al. (2002). A genome-wide strategy for the identification of essential genes in Staphylococcus aureus. Mol. Microbiol. 43, 1387–1400. doi: 10.1046/j.1365-2958.2002.02832.x
Gazzaniga, F., Stebbins, R., Chang, S. Z., McPeek, M. A., and Brenner, C. (2009). Microbial NAD metabolism: lessons from comparative genomics. Microbiol. Molec. Biol. Rev. 73, 529–541. doi: 10.1128/mmbr.00042-08
Gottwald, T. R., Da Graça, J. V., and Bassanezi, R. B. (2007). Citrus huanglongbing: the pathogen and its impact. Plant Health Progress 6. doi: 10.1094/PHP-2007-0906-01-RV
Hava, D. L., and Camilli, A. (2002). Large-scale identification of serotype 4 Streptococcus pneumoniae virulence factors. Molec. Microbiol. 45, 1389–1406. doi: 10.1046/j.13652958.2002.03106.x
Heath, R. J., and Rock, C. O. (2004). Fatty acid biosynthesis as a target for novel antibacterials. Curr. Opin. Investig. Drugs 5, 146–153.
Juhas, M., Reuß, D. R., Zhu, B., and Commichau, F. M. (2014). Bacillus subtilis and Escherichia coli essential genes and minimal cell factories after one decade of genome engineering. Microbiology 160, 2341–2351. doi: 10.1099/mic.0.079376-0
Karlyshev, A. V., Pallen, M. J., and Wren, B. W. (2000). Single-primer PCR procedure for rapid identification of transposon insertion sites. Biotechniques 28, 1078, 1080, 1082.
Laia, M. L., Moreira, L. M., Dezajacomo, J., Brigati, J. B., Ferreira, C. B., Ferro, M. I., et al. (2009). New genes of Xanthomonas citri subsp. citri involved in pathogenesis and adaptation revealed by a transposon-based mutant library. BMC Microbiol. 9:12. doi: 10.1186/1471-2180-9-12
Langridge, G. C., Phan, M. D., Turner, D. J., Perkins, T. T., Parts, L., Haase, J., et al. (2009). Simultaneous assay of every Salmonella Typhi gene using one million transposon mutants. Genome Res. 19, 2308–2316. doi: 10.1101/gr.097097.109
La Scola, B., Khelaifia, S., Lagier, J. C., and Raoult, D. (2014). Aerobic culture of anaerobic bacteria using antioxidants: a preliminary report. Eur. J. Clin. Microbiol. Infect. Dis. 33, 1781–1783. doi: 10.1007/s10096-014-2137-4
Lee, S. A., Gallagher, L. A., Thongdee, M., Staudinger, B. J., Lippman, S., Singh, P. K., et al. (2015). General and condition-specific essential functions of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 112, 5189–5194. doi: 10.1073/pnas.1422186112
Leonard, M. T., Fagen, J. R., Davis-Richardson, A. G., Davis, M. J., and Triplett, E. W. (2012). Complete genome sequence of Liberibacter crescens BT-1. Stand. Genomic Sci. 7, 271–283. doi: 10.4056/sigs.3326772
Li, W., Cong, Q., Pei, J., Kinch, L. N., and Grishin, N. V. (2012). The ABC transporters in Candidatus Liberibacter asiaticus. Proteins 80, 2614–2628. doi: 10.1002/prot.24147
Lin, H., Lou, B., Glynn, J. M., Doddapaneni, H., Civerolo, E. L., Chen, C., et al. (2011). The complete genome sequence of ‘Candidatus Liberibacter solanacearum,’ the bacterium associated with potato zebra chip disease. PLoS ONE 6:e19135. doi: 10.1371/journal.pone.0019135
Lin, H., Pietersen, G., Han, C., Read, D. A., Lou, B., Gupta, G., et al. (2015). Complete genome sequence of “Candidatus Liberibacter africanus,” a bacterium associated with citrus huanglongbing. Genome Announc. 3, e00733–e00715. doi: 10.1128/genomeA.00733-15
Mei, J. M., Nourbakhsh, F., Ford, C. W., and Holden, D. W. (1997). Identification of Staphylococcus aureus virulence genes in a murine model of bacteraemia using signature-tagged mutagenesis. Molec. Microbiol. 26, 399–407. doi: 10.1046/j.1365-2958.1997.5911966.x
Miesel, L., Greene, J., and Black, T. A. (2003). Genetic strategies for antibacterial drug discovery. Nat. Rev. Genet. 4, 442–456. doi: 10.1038/nrg1086
Moule, M. G., Hemsley, C. M., Seet, Q., Guerra-Assunção, J. A., Lim, J., Sarkar-Tyson, M., et al. (2014). Genome-wide saturation mutagenesis of Burkholderia pseudomallei K96243 predicts essential genes and novel targets for antimicrobial development. MBio 5, e00926–e00913. doi: 10.1128/mBio.00926-13
Overbeek, R., Olson, R., Pusch, G. D., Olsen, G. J., Davis, J. J., Disz, T., et al. (2014). The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 42, D206–D214. doi: 10.1093/nar/gkt1226
Pagliai, F. A., Gonzalez, C. F., and Lorca, G. L. (2015). Identification of a Ligand Binding Pocket in LdtR from Liberibacter asiaticus. Front. Microbiol. 6:1314. doi: 10.3389/fmicb.2015.01314
Shapiro, A. B., Ross, P. L., Gao, N., Livchak, S., Kern, G., Yang, W., et al. (2013). A high-throughput-compatible fluorescence anisotropy-based assay for competitive inhibitors of Escherichia coli UDP-N-acetylglucosamine acyltransferase (LpxA). J. Biomol. Screen. 18, 341–347. doi: 10.1177/1087057112462062
Tatineni, S., Sagaram, U. S., Gowda, S., Robertson, C. J., Dawson, W. O., Iwanami, T., et al. (2008). In planta distribution of ‘Candidatus Liberibacter asiaticus’ as revealed by polymerase chain reaction (PCR) and real-time PCR. Phytopathology 98, 592–599. doi: 10.1094/PHYTO-98-5-0592
Tyler, H. L., Roesch, L. F., Gowda, S., Dawson, W. O., and Triplett, E. W. (2009). Confirmation of the sequence of ‘Candidatus Liberibacter asiaticus’ and assessment of microbial diversity in Huanglongbing-infected citrus phloem using a metagenomic approach. Mol. Plant Microbe Interact. 22, 1624–1634. doi: 10.1094/MPMI-22-12-1624
Vrablik, T. L., Huang, L., Lange, S. E., and Hanna-Rose, W. (2009). Nicotinamidase modulation of NAD+ biosynthesis and nicotinamide levels separately affect reproductive development and cell survival in C. elegans. Development 136, 3637–3646. doi: 10.1242/dev.028431
Wang, N., and Trivedi, P. (2013). Citrus huanglongbing: a newly relevant disease presents unprecedented challenges. Phytopathology 103, 652–665. doi: 10.1094/PHYTO-12-12-0331-RVW
Watabe, K., Mimuro, M., and Tsuchiya, T. (2014). Development of a high-frequency in vivo transposon mutagenesis system for Synechocystis sp. PCC 6803 and Synechococcus elongatus PCC 7942. Plant Cell Physiol. 55, 2017–2026. doi: 10.1093/pcp/pcu128
Weissbach, H., Etienne, F., Hoshi, T., Heinemann, S. H., Lowther, W. T., Matthews, B., et al. (2002). Peptide methionine sulfoxide reductase: structure, mechanism of action, and biological function. Arch. Biochem. Biophys. 397, 172–178. doi: 10.1006/abbi.2001.2664
Wulff, N. A., Zhang, S., Setubal, J. C., Almeida, N. F., Martins, E. C., Harakava, R., et al. (2014). The complete genome sequence of ‘Candidatus Liberibacter americanus,’ associated with Citrus huanglongbing. Mol. Plant Microbe Interact. 27, 163–176. doi: 10.1094/MPMI-09-13-0292-R
Yan, Q., Sreedharan, A., Wei, S., Wang, J., Pelz-Stelinski, K., Folimonova, S., et al. (2013). Global gene expression changes in Candidatus Liberibacter asiaticus during the transmission in distinct hosts between plant and insect. Mol. Plant Pathol. 14, 391–404. doi: 10.1111/mpp.12015
Keywords: Liberibacter, transposon mutagenesis, essential genes, citrus greening disease, Huanglongbing, HLB, citrus
Citation: Lai K-K, Davis-Richardson AG, Dias R and Triplett EW (2016) Identification of the Genes Required for the Culture of Liberibacter crescens, the Closest Cultured Relative of the Liberibacter Plant Pathogens. Front. Microbiol. 7:547. doi: 10.3389/fmicb.2016.00547
Received: 30 June 2015; Accepted: 04 April 2016;
Published: 20 April 2016.
Edited by:
John R. Battista, Louisiana State University and Agricultural and Mechanical College, USAReviewed by:
Pankaj Trivedi, Univesity of Western Sydney, AustraliaElena Gonella, Università degli Studi di Torino, Italy
Copyright © 2016 Lai, Davis-Richardson, Dias and Triplett. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric W. Triplett, ewt@ufl.edu