 George E. Anasontzis1†
George E. Anasontzis1† Elisavet Kourtoglou2
Elisavet Kourtoglou2 Silas G. Villas-Boâs3
Silas G. Villas-Boâs3 Dimitris G. Hatzinikolaou1
Dimitris G. Hatzinikolaou1 Paul Christakopoulos4*
Paul Christakopoulos4*- 1Microbial Biotechnology Unit, Sector of Botany, Department of Biology, National and Kapodistrian University of Athens, Zografou, Greece
- 2BIOtechMASS Unit, Biotechnology Laboratory, School of Chemical Engineering, National Technical University of Athens, Zografou, Greece
- 3Centre for Microbial Innovation, School of Biological Sciences, University of Auckland, Auckland, New Zealand
- 4Biochemical and Chemical Process Engineering, Division of Sustainable Process Engineering, Department of Civil, Environmental and Natural Resources Engineering, Luleå University of Technology, Luleå, Sweden
Fusarium oxysporum is one of the few filamentous fungi capable of fermenting ethanol directly from plant cell wall biomass. It has the enzymatic toolbox necessary to break down biomass to its monosaccharides and, under anaerobic and microaerobic conditions, ferments them to ethanol. Although these traits could enable its use in consolidated processes and thus bypass some of the bottlenecks encountered in ethanol production from lignocellulosic material when Saccharomyces cerevisiae is used—namely its inability to degrade lignocellulose and to consume pentoses—two major disadvantages of F. oxysporum compared to the yeast—its low growth rate and low ethanol productivity—hinder the further development of this process. We had previously identified phosphoglucomutase and transaldolase, two major enzymes of glucose catabolism and the pentose phosphate pathway, as possible bottlenecks in the metabolism of the fungus and we had reported the effect of their constitutive production on the growth characteristics of the fungus. In this study, we investigated the effect of their constitutive production on ethanol productivity under anaerobic conditions. We report an increase in ethanol yield and a concomitant decrease in acetic acid production. Metabolomics analysis revealed that the genetic modifications applied did not simply accelerate the metabolic rate of the microorganism; they also affected the relative concentrations of the various metabolites suggesting an increased channeling toward the chorismate pathway, an activation of the γ-aminobutyric acid shunt, and an excess in NADPH regeneration.
Introduction
Lignocellulose is the complex matrix of cellulose, hemicellulose, and lignin that constitutes the plant cell wall. Being a waste product from agricultural activities, forestry, and the pulp and paper industry, lignocellulose can be a sustainable source of carbohydrates for biotechnological applications (Fujita et al., 2002; Malherbe and Cloete, 2002).
The commonly perceived process for lignocellulose use in a biorefinery for ethanol production comprises of two main steps; hydrolysis and fermentation. Hydrolysis refers to the breakdown of polysaccharides present in biomass to monosaccharides and fermentation refers to the use of these monosaccharides for the synthesis of the end-product. Hydrolysis and fermentation can be performed either sequentially or in parallel (Ask et al., 2012). Selection of the set-up depends on the raw material. However, since each set-up has different requirements in terms of physical and chemical conditions, different biomass-degrading enzyme mixtures are required for the efficient release of the available sugars. For example, when hydrolysis and fermentation are performed simultaneously, the temperature is maintained at 30°C, which is not necessarily the optimum temperature for the commercial enzyme mixtures. When hydrolysis is performed separately from fermentation, the temperature can be closer to the optimum for the enzyme mixture, but other factors—such as product inhibition—can affect the efficiency of the process (Ask et al., 2012).
As far as the fermentation part is concerned Saccharomyces cerevisiae is undoubtedly the most important microorganism, due to its high productivity (reported as high as 3.3 gethanol L−1 h−1; Cavka et al., 2010) and generally high metabolic rate, which make it a suitable microorganism for industrial use (Kuyper et al., 2004). This microorganism is capable of fermenting hexoses, such as glucose and mannose, to ethanol. Glucose is present in various concentrations and structures in plant biomass, but predominantly in the form of cellulose; mannose is abundant as a hemicellulose constituent from gymnosperms. The capacity of yeast to efficiently produce ethanol has prompted its use for the production of biofuels, and as a production host for the biosynthesis of other bulk and fine biochemicals (Nevoigt, 2008).
Despite its obvious advantages, S. cerevisiae has several drawbacks that hinder its use in lignocellulose-based processes; these include its dependence on previous chemical or enzymatic steps in order to break down the plant cell wall biomass to monosaccharides, and also its inability to metabolize pentoses such as xylose (la Grange et al., 2010). Despite the development of S. cerevisiae strains capable of fermenting xylose (Bettiga et al., 2009), which has allowed extensive work on increasing tolerance to inhibitors present in lignocellulose (Koppram et al., 2012) and on optimizing the fermentation process from lignocellulosic materials (Koppram et al., 2013), these bottlenecks have also encouraged the development of microorganisms that can use plant biomass, saccharify the polymers, and ferment the product of interest in a consolidated bioprocess (Lynd et al., 1996). This can be based either on the engineering of microorganisms that are naturally capable of breaking down biomass in order to improve the efficiency of the production step, or on the engineering of non-cellulolytic microorganisms, but with a high yield for the product of interest, in order to acquire the ability to extract the sugars from lignocellulosic biomass (la Grange et al., 2010). Despite the progress achieved in both of these strategies, outlined in recent reviews (la Grange et al., 2010; den Haan et al., 2015), there are no such strains available today that could make the consolidated bioprocessing system advantageous on an industrial scale.
Fusarium oxysporum is a filamentous fungus. It is one of the few fungal species reported to ferment plant carbohydrate polymers to ethanol in a one-step process (Christakopoulos et al., 1989; Singh and Kumar, 1991; Anasontzis et al., 2011). It can produce a range of plant cell wall degrading enzymes (Christakopoulos et al., 1996a,c, 1997) and ferment D-glucose, D-xylose, cellulose, and hemicellulose to ethanol. Compared to yeasts, its main disadvantage is the low conversion rate (Singh and Kumar, 1991).
The strain F3 of F. oxysporum was selected for its high cellulase production, and it has been extensively studied (Christakopoulos et al., 1989; Panagiotou et al., 2005a,b,c, 2008). The accumulation by F. oxysporum F3 of high levels of glucose-1,6-phosphate at the end of both the aerobic and the anaerobic phases, when glucose was used as carbon source, was attributed to what was believed to be a low phosphoglucomutase activity, which was consequently restricting the metabolism of glucose toward cell wall biosynthesis (Panagiotou et al., 2005c), glycolysis, and ethanol production. This enabled us to study this enzyme further and to show that it follows the ping-pong mechanism for the conversion of glucose-1-P to glucose-6-P and vice versa (Kourtoglou et al., 2011). Also, the accumulation of sedoheptulose-7-P when the microorganism was grown in glucose-xylose mixtures under oxygen-limiting conditions, and of erythrose-4-P during xylose consumption (Panagiotou et al., 2005b), indicated a second bottleneck in the F. oxysporum metabolism. Fan et al. reported the effect on F. oxysporum of the overexpression of transaldolase genes from Pichia stipitis and S. cerevisiae, which increased both xylose consumption and ethanol production when either glucose or xylose was used as carbon source (Fan et al., 2011a,b) in shake flask batch cultures.
We had previously investigated the effect of overproduction of these two enzymes on the metabolism of F. oxysporum under aerobic conditions. Through a double transformation using customized vectors providing hygromycin resistance, we had generated a number of strains that constitutively overexpressed the phosphoglucomutase and/or the transaldolase gene under the regulation of the gpdA promoter of Aspergillus nidulans. Among these strains, FF11 produced the highest amounts of both enzymes. Thus, we compared FF11 and the parental strain F3 in order to evaluate the effect of the modification on the growth characteristics of the microorganism. We found that the applied modification increased the growth rate and the fungal biomass yield when glucose was used as carbon source. We also detected changes in the metabolome that indicated a facilitation of the channeling of the sugars through the glycolysis and pentose phosphate metabolism, such as the accumulation of glutamate as a possible response to the high NADPH availability from higher flux through the pentose phosphate pathway (Anasontzis et al., 2014a).
In this study, we evaluated the effect of the constitutive production of phosphoglucomutase and tranlsaldolase on ethanol productivity when glucose was used as carbon source. Thus, we compared the parental strain F3 with the genetically modified FF11 under anaerobic conditions. We showed that the latter outperformed F3 in terms of ethanol production, glucose consumption and by-product generation. We also used metabolomics tools, in an attempt to understand if the metabolome profile indicates a continuously high NADPH and NADH availability which resulted in improved fermentation characteristics.
Materials and Methods
Microorganisms and Media
Fusarium oxysporum wild type F3 (Christakopoulos et al., 1989) was maintained on PDA plates (Potato Dextrose Agar; Applichem, Germany) and recombinant FF11 (Anasontzis et al., 2014a) was maintained on PDA plates supplemented with 50 mg L−1 hygromycin at 4°C, following growth for 4 days at 25°C.
The basal medium for growth in bioreactors and for metabolome analysis had the following composition (in g.L−1): NaH2PO4.2H2O, 7; Na2HPO4.2H2O, 9.5; KH2PO4, 1; (NH4)2HPO4, 10; MgSO4.7H2O, 0.3; CaCl2.2H2O, 0.3 (Christakopoulos et al., 1996b).
Precultures were grown for 24 h in 500-mL Erlenmeyer flasks (100 mL working volume) using the basal medium supplemented with glucose at 20 g.L−1 as carbon source. The whole preculture was used to inoculate a 2-L stirred tank bioreactor (BioFlo310, Benchtop Fermenter/Bioreactor; New Brunswick Scientific GmbH, Germany) with a working volume of 1.5 L and at a final concentration of 7% (v/v), under aerobic conditions as previously described (Anasontzis et al., 2014a). After the end of the aerobic phase, which corresponded to glucose depletion, the air supply was stopped and 60 g of sterilized glucose was added.
Genetic Modification
Genetic modification of F. oxysporum F3 was performed as previously described (Kourtoglou et al., 2011; Anasontzis et al., 2014a). In brief, the genes encoding for translaldolase and phosphoglucomutase—tal and pgm— were isolated from the genomic DNA of F. oxysporum with PCR and appropriately designed primers. The overexpression vectors were constructed by substituting the xyl2 gene from pBlXyl (Anasontzis et al., 2011) with the PCR fragments of the two genes. pBLXyl had the gpdA promoter upstream and the trpC terminator of A. nidulans downstream the xyl2. The hygromycin resistance gene was isolated from the plasmid pCSN44 and ligated to the plasmids, after digestion with XhoI. The resulting plasmids pBlTAL-hyg (Genebank accession JF756587) and pBLPGM-hyg (Anasontzis et al., 2011; Kourtoglou et al., 2011) were then used for the co-transformation of F. oxysporum F3. Protoplast preparation and transformation were performed as previously described (Kourtoglou et al., 2011). Integration of the genes in the genome was confirmed with PCR and Southern analysis, while gene expression was evaluated with northern analysis. The successful overproduction of the two enzymes was verified with enzymatic assays. The transformant FF11, which had incorporated both inserts and overproduced both enzymes, was selected for this study.
Cell Density and Glucose Concentration
Cell density was calculated based on dry cell weight (DCW). To determine the glucose concentration in the culture medium, samples were first centrifuged at 4°C and 10,000 × g. The supernatant was then analyzed by HPLC, on a Jasco (PU-987) HPLC system equipped with an ion-moderated partition chromatography column (Aminex HPX-87H; Bio-Rad), and a Waters 410 refractive index detector. The mobile phase (5 mM H2SO4) was 0.6 mL.min−1 and the temperature 45°C. All measurements are the result of technical duplicates from two identical fermentations (biological duplicates).
Quenching and Sample Metabolite Analysis
Sample quenching and derivatization, and the subsequent analyses of amino and non-amino organic acids were performed as previously described (Smart et al., 2010; Anasontzis et al., 2014a). In brief, triplicate 50-mL samples from the cultures were obtained at the last sampling point of the anaerobic phase (139.5 h for FF11 and 124 h for F3), either just before depletion of the carbon source or after the culture had shifted to the stationary phase.
The samples were filtered under vacuum to separate the fungal biomass from the culture medium. The mycelium was then washed with saline solution at 4°C and quenched with cold methanol-water (1:1 v/v) at −30°C. From each filtrate, 1 mL was sampled and supplemented with 0.2 μmol 2,3,3,3-d4-alanine as internal standard, and stored at −80°C for the extracellular metabolite analysis. From the quenched mycelium, cold methanol-water freeze and thaw cycles quenched the intracellular metabolites. Alanine was also added as internal standard before the extraction. Both quenched culture medium and the extracted intracellular metabolites from the mycelium were freeze-dried using a BenchTop K manifold freeze dryer (VirTis) before chemical derivatization. The derivatization of the freeze-dried samples was performed with the methyl chloroformate method (MCF), and the derivatives were analyzed in an Agilent GC7890 system coupled to a MSD5975 mass selective detector and a ZB-1701 GC capillary column (30 m × 250 μm internal diameter × 0.15 μm with 5-m guard column; Phenomenex) as previously described (Smart et al., 2010). Metabolomic data analysis was performed as previously described (Smart et al., 2010; Anasontzis et al., 2014a). The deconvolution of GC-MS chromatograms and the identification of metabolites was performed with the software AMDIS using an in-house MCF mass spectra library. Both MS spectrum and its respective retention time were used for metabolite identification. The relative abundance of identified metabolites was determined with ChemStation (Agilent) by using the GC base-peak value of a selected reference ion, normalized by the initial biomass of each sample as well by the internal standard concentration (2,3,3,3-d4-alanine). Samples were prepared in triplicates from two identical bioreactors for each strain.
Results
Based on the hypothesis that the overproduction of phosphoglucomutase and transaldolase could help bypass the assumed bottleneck (Panagiotou et al., 2005c) in the metabolism of F. oxysporum, we previously described the expression of the pgm and tal genes under the regulation of the constitutive promoter of gpdA of Aspergillus nidulans. Strain FF11, which constitutively produced both enzymes, was selected for further evaluation. We reported how the overproduction of these two enzymes affected the metabolic channeling and the growth of the microorganism under aerobic conditions. A higher specific growth rate and a higher abundance for the majority of the detected amino and non-amino organic acids was observed at the end of the aerobic phase—when glucose was fully consumed or growth reached a plateau (Anasontzis et al., 2014b).
Considering the perspective of using F. oxysporum for ethanol production from lignocellulose in a consolidated bioprocess, we evaluated the effect of constitutive expression of the respective genes on ethanol productivity using glucose as the sole carbon source.
The recombinant strain, FF11, showed significantly higher ethanol yield than the wild type, and in parallel, a lower yield of acetic acid (Table 1). FF11 produced more than 20 g.L−1 of ethanol after 139 h of anaerobic fermentation, while the wild type seemed to decrease its glucose consumption rate at around 120 h, with an ethanol concentration of less than 10 g L−1. Another interesting point was the lower production of acetate by the recombinant strain compared to the wild type one; acetate is a common by-product of fermentation, and was also detected as a major by-product in the case of the wild type (Figure 1). Ethanol productivity was also higher for FF11 compared to F3 (0.146 and 0.080 g L−1 h−1, respectively), as was the glucose consumption rate (0.290 and 0.223 g L−1 h−1).
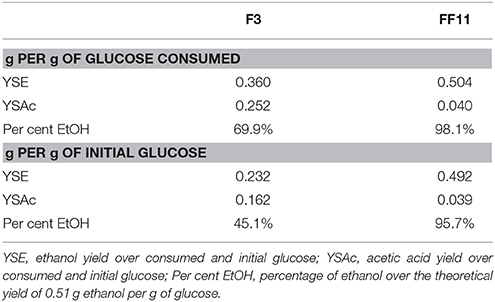
Table 1. Yield of ethanol and acetic acid.
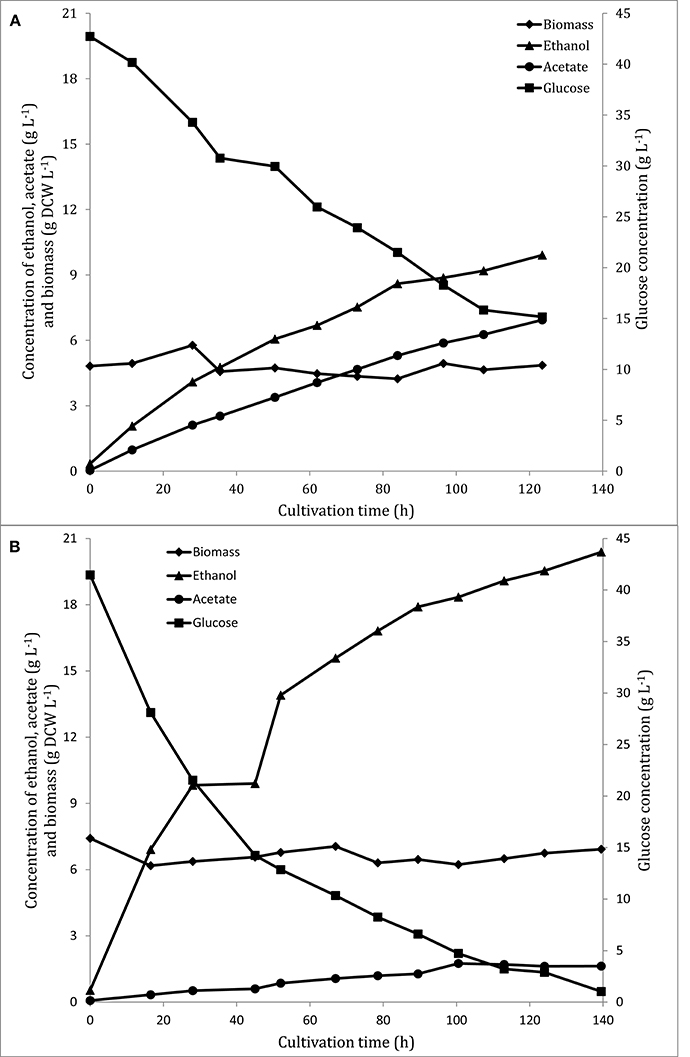
Figure 1. Glucose (■), ethanol (▴), acetic acid (•), and biomass (♦) during the anaerobic phase of F. oxysporum F3 (A) and FF11 (B) fermentation. These are results of duplicate measurements (technical duplicates) from two different fermenters (biological duplicates). Standard deviation was lower than 3%.
In order to further investigate the metabolic state of the two strains during the anaerobic stage of the culture, to identify possible bottlenecks, and to understand if the more efficient redox balance achieved by the recombinant strain during the aerobic phase was retained also under anaerobic conditions, we performed metabolomics analysis of amino and non-amino acid metabolites at the final sampling point of each fermentation—when glucose consumption reached a plateau for strain F3 and when glucose was practically depleted for strain FF11. The sampling point was selected in order to obtain comparable results with our previous work, when metabolomics analysis was performed at the end of the aerobic phase, and to allow accumulation of metabolites in detectable and significant levels (Anasontzis et al., 2014a). The results of the comparison between the two strains under anaerobic conditions are presented in Figure 2. It is evident that the citric acid cycle was more active in the recombinant strain, at least after the γ-aminobutyric acid (GABA) shunt. Higher concentrations of lactic acid, nicotinic acid, and citramalate were also observed. Itaconic acid levels were higher for the recombinant strain only in the extracellular samples, although still low as absolute concentrations. Several amino acids had lower concentrations in FF11 compared to the wild type strain. Table 2 shows the average values of the intracellular and extracellular metabolites under anaerobic conditions for both strains.
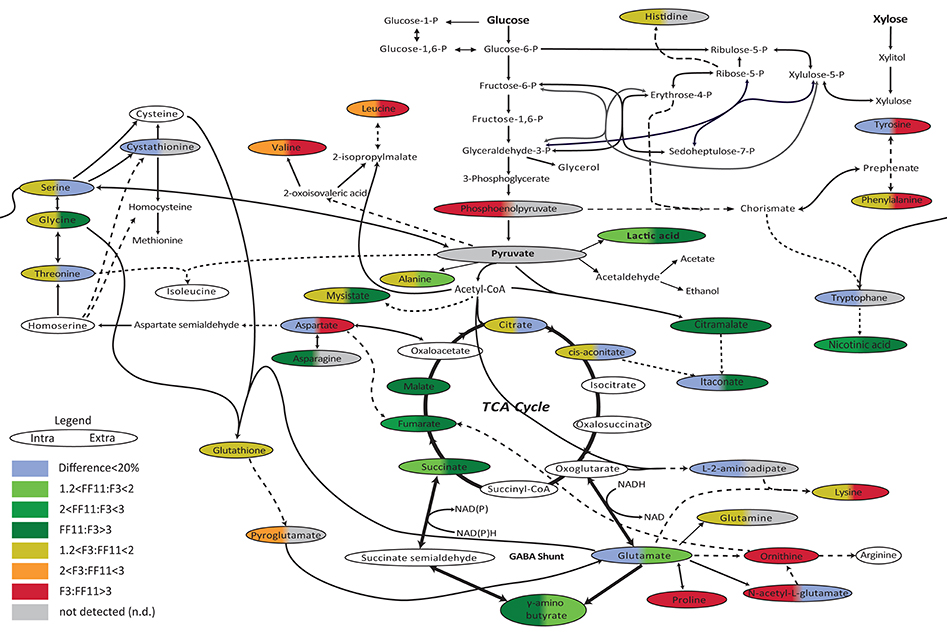
Figure 2. Schematic demonstration of the relative intracellular concentrations of amino and non-amino organic acids during anaerobic growth of F. oxysporum F3 and FF11 strains on glucose. Single lines: One enzyme reaction; dashed lines: several reactions; Colors on the left and right correspond to the difference in intracellular and extracellular metabolites, respectively. Red: F3:FF11 > 3; Orange: 2 < F3:FF11 < 3; green/yellow: 1.2 < F3:FF11 < 2: Dark green: FF11:F3 > 3; Green: 2 < FF11:F3 < 3; Light green: 1.2 < FF11:F3 < 2: blue: difference smaller than 20%; gray: not detected (adapted from Anasontzis et al., 2014a).
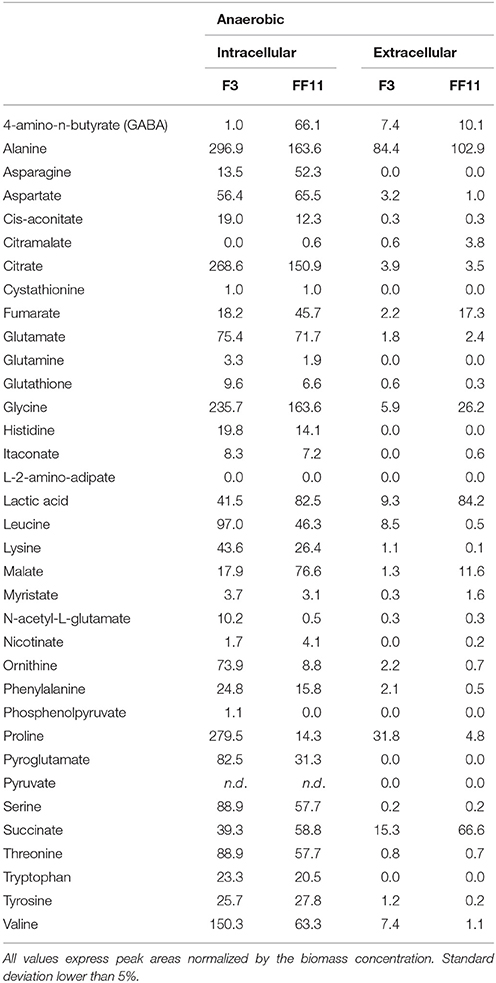
Table 2. Amino acid and non-amino acid intracellular and extracellular metabolites detected at the end of the anaerobic phase, for the two strains.
Discussion
Ethanol Productivity
Our results show an increase in ethanol productivity in the recombinant strain. This indicates that the genetic modification performed was in the right direction, as it has improved the yield of the main product while reducing that of acetic acid, the main by-product. Phosphoglucomutase overexpression would be expected to increase the channeling of glucose to pyruvate and then to ethanol and acetic acid. However, it is not clear why acetaldehyde catabolism shifts toward ethanol instead of acetic acid. An explanation could be that due to the constitutive production of transaldolase and the more active PPP, the NADPH recycling is more efficient and therefore less acetic acid production is required to regenerate NADPH through the NADP-dependent acetaldehyde dehydrogenase (E.C. 1.2.1.3). The higher concentration of lactic acid in the recombinant strain confirms this, as lactic acid production regenerates NAD+ through the action of lactic acid dehydrogenase.
Despite the very high ethanol yield and the improvement in the ethanol productivity (Table 1), F. oxysporum still cannot compete with other microorganisms in terms of productivity and final ethanol concentration. As Table 3 shows, both these values from strains F3 and FF11 are still much lower in comparison to values in the literature. For example, Rhizopus oryzae has been reported to have an ethanol productivity of 1.87 g L−1 h−1 (Millati et al., 2005). The maximum ethanol concentration was also very low compared to values achieved routinely with S. cerevisiae in pure or mixed substrates (Saitoh et al., 2010). Nevertheless, FF11 gave among the highest yields of glucose to ethanol, higher than 98% of the theoretical value. In a similar study, Fan et al. demonstrated the effect of the expression of a transaldolase gene from P. stipitis in ethanol production in flask cultures (Fan et al., 2011a). Although the experimental set-up was different, and parameters such as productivity and yield are not readily comparable, extrapolation of their results indicates an improvement in ethanol yields. We observed even greater improvement, which could be attributed to simultaneous overproduction of both transaldolase and phosphoglucomutase.
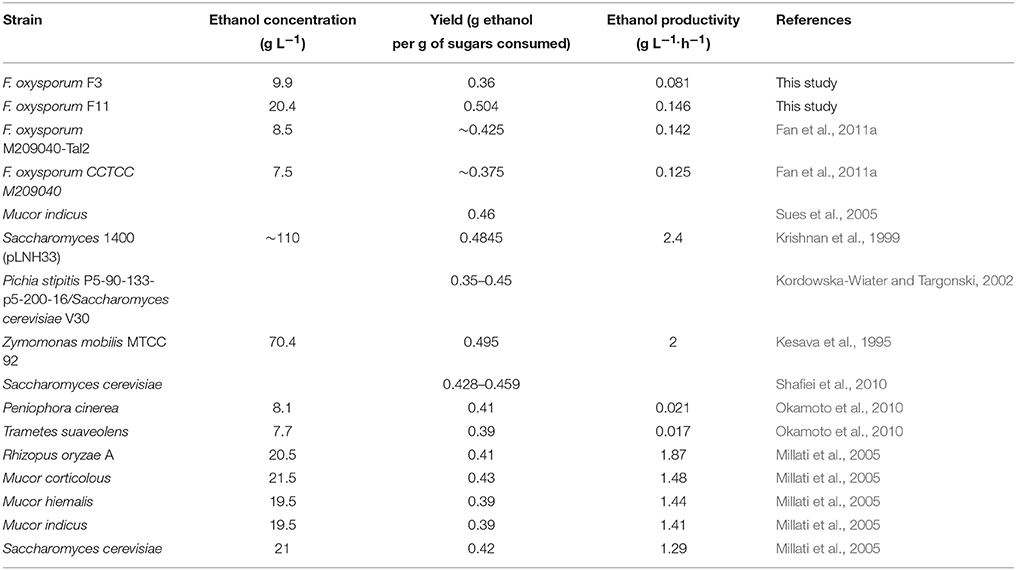
Table 3. Ethanol productivity of different strains reported in the literature, using glucose as the carbon source.
Metabolite Concentrations
The relative levels of tryptophan, tyrosine, phenylalanine, and nicotinic acid for FF11, between the aerobic phase (Anasontzis et al., 2014a) and the anaerobic phase, showed no significant variation. For the wild type, though, all three of these amino acids showed higher levels in the anaerobic phase than in the aerobic one. It seems that in the wild type, their production is induced under anaerobic conditions. However, in the recombinant strain, there appears to be a constant need for their synthesis, indicating that the modifications have increased the metabolic flow toward their production in both culture regimes. The lack of detection of phosphoenolpuryvate under anaerobic conditions supports this idea, and it suggests increased channeling toward either the TCA cycle or the chorismate pathway. Although nicotinic acid levels are constant in the recombinant strain, they are lower in the wild type under anaerobic conditions.
For S. cerevisiae, it has been reported that under aerobic conditions nicotinic acid is synthesized from tryptophan, while under anaerobic conditions it appears to be produced from aspartate and glutamate (Ahmad and Moat, 1966). Strain F3 concentrations of nicotinic acid, glutamate, and aspartate drop during the anaerobic phase, an observation that indicates a similar mechanism of regulation. However, for the recombinant strain, although aspartate and glutamate also decrease under anaerobic conditions, nicotinic acid is only slightly affected. It is possible that the constitutive expression of the two genes has led to the partial retainment of the aerobic nicotinic acid production pathway (the chorismate pathway). Nicotinic acid is a precursor of NAD synthesis; however, it is not clear if the higher concentrations of nicotinic acid in the recombinant strain are related to the availability of the different forms of NAD.
The levels of all following amino acids—serine, glycine, alanine, leucine, threonine, lysine, glutamine, and proline—increased in the recombinant strain under aerobic conditions (Anasontzis et al., 2014a), but were relatively lower under anaerobic conditions. This could be loosely related to the protein production levels under the latter conditions. If the excessive production of fumarate during both phases in the recombinant strain acts as the electron acceptor during protein production, as has been previously suggested for S. cerevisiae (Liu et al., 2015), the intracellular concentrations of at least several of the protein building blocks could be expected to be lower, as they are more readily consumed. Nevertheless, under aerobic conditions the recombinant strain has higher intracellular levels of these amino acids, but whether this is a result of an increase in their production rate or a decrease in their consumption rate remains unclear with the current experimental set-up. The different relative levels of other amino acids such as valine, aspartate, asparagine, histidine, and glutamate further complicate the picture.
Apart from fumarate, other metabolites of the TCA cycle that are linked to the GABA shunt showed higher concentrations in the recombinant strain under both aerobic and anaerobic conditions. It seems that this effect is restricted after the GABA shunt and it is not reflected on the glutamate—the precursor of GABA—, citrate, and cis-aconitate levels. On the other hand, the GABA shunt of the wild type appears to be completely deactivated under anaerobic conditions, while the TCA cycle remains active, albeit with reduced metabolite concentrations. It seems that under anaerobic conditions, there is a reduced need in the wild type for GABA production, a compound that plays an important role in response to environmental stresses and especially to reactive oxygen stress (Mead et al., 2013), while the genetic modifications keep the GABA shunt active. The low production of acetic acid, which indicates no need for NADPH regeneration, may be associated with the active GABA shunt—as such pathways become extremely important as sinks for excess NAD(P)H which occurs when the TCA cycle is inhibited (Streeter and Salminen, 1990).
An active TCA cycle under anaerobic conditions has also been reported in other microorganisms. In Rhizopus arrhizus, it has been suggested that both TCA and pyruvate carboxylation are active but in the opposite direction (Kenealy et al., 1986; Roa Engel et al., 2008). The higher concentrations of fumarate, succinate, malate and GABA in the recombinant strain FF11 propose a similar equilibrium, leading to the accumulation of GABA. The higher succinate production could contribute to the NAD+ recycling, as it has been reported for Bifidobacterium sp. (Van der Meulen et al., 2006).
In order for the cell to regenerate its reductive potential, it is necessary to recycle the cofactors and maintain the balance of NAD+/NADH and NADP+/NADPH. This balance allows energy production and use of the substrate for compound synthesis (Roger et al., 2004). Both glycolysis and pentose phosphate pathway are two key metabolic pathways both present in even between distantly related microorganisms such as F. oxysporum (Panagiotou et al., 2005a) and lactic acid bacteria (Roger et al., 2004). Modification of the metabolic fluxes to increase or reduce the accumulation of a specific metabolite is often the target, but selection of the modification that will give the expected result is not always possible, as the effect can be unpredictable (Roger et al., 2004). The repression of an enzyme, such as the lactic acid dehydrogenase which also regenerates NAD+ (Monnet et al., 2000) or the acetaldehyde dehydrogenase which regenerates NAD(P)H (Panagiotou et al., 2005b), could drive the metabolism to alternative pathways and equilibria with the same or unexpected results.
The overproduction of phosphoglucomutase and transaldolase was not aimed at disrupting the redox equilibrium of F. oxysporum, but instead to accelerate the metabolism at specific points, and eventually increase its ethanol productivity. Nevertheless, it seems that the redox equilibrium has indeed been affected in a beneficial way, with several metabolites supporting it under both aerobic (Anasontzis et al., 2014a) and anaerobic conditions.
Despite the positive effect of the constitutive production of phosphoglucomutase and transaldolase directly on ethanol productivity and indirectly on redox equilibrium, further improvements would be required to allow the large scale perspectives of F. oxysporum in biofuel production from lignocellulose. Other aspects, such as ethanol tolerance, also need to be addressed (Paschos et al., 2015). Screening for the selection of wild type strains that are more efficient ethanol producers and better understanding of the gene regulation and bottlenecks under ethanol producing conditions (Ali et al., 2012, 2013), further genetic modifications on selected pathways, evolutionary engineering, and novel bioprocessing set-ups, are necessary in order to achieve the production of ethanol above the limit of 5% (w/v) from lignocellulose (Anasontzis and Christakopoulos, 2014). Under the scope of this work, investigation and increase of the efficiency of alcohol dehydrogenase would help determine other major bottlenecks in ethanol production from F. oxysporum, while studies of the performance of the selected strain on lignocellulose material would also shed light on the limitations that might have been removed or added through this approach. Constitutive production of these enzymes might not be the most optimal modification; thus fine-tuning using different promoters could improve the growth and ethanol characteristics of the modified strains.
Author Contributions
GA contributed to the design of the work, performed the fermentations and the sampling for metabolome analysis, interpreted the data, and drafted the manuscript. EK performed the fermentations, sampling, and HPLC analyses, and also drafted the results. SV designed and performed the metabolomics analysis and revised the manuscript. DH contributed to the design of the study, interpretation of the data, and revision of the manuscript. PC conceived the study, contributed to the design and interpretation of the results, and revised the manuscript. All the authors have approved the final version to be published and have agreed to be accountable for all aspects of the work.
Funding
This work has been co-funded by the project PENED 2003 and the European Community's Seventh Framework Program (FP7/2007-2013) under grant agreement 213139 (the HYPE project). PENED is co-financed 80% from public expenditure through the EC-European Social Fund, 20% from public expenditure through the Greek Ministry of Development—General Secretariat of Research and Technology, and through private sector, under measure 8.3 of the OPERATIONAL PROGRAMME “COMPETITIVENESS” in the Third Community Support Program.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ahmad, F., and Moat, A. G. (1966). Nicotinic acid biosynthesis in prototrophs and tryptophan auxotrophs of Saccharomyces cerevisiae. J. Biol. Chem. 241, 775–780.
Ali, S. S., Khan, M., Fagan, B., Mullins, E., and Doohan, F. M. (2012). Exploiting the inter-strain divergence of Fusarium oxysporum for microbial bioprocessing of lignocellulose to bioethanol. AMB Express 2:16. doi: 10.1186/2191-0855-2-16
Ali, S. S., Nugent, B., Mullins, E., and Doohan, F. M. (2013). Insights from the fungus Fusarium oxysporum point to high affinity glucose transporters as targets for enhancing ethanol production from lignocellulose. PLoS ONE 8:e54701. doi: 10.1371/journal.pone.0054701
Anasontzis, G., and Christakopoulos, P. (2014). Challenges in ethanol production with Fusarium oxysporum through consolidated bioprocessing. Bioengineered 5, 393–395. doi: 10.4161/bioe.36328
Anasontzis, G. E., Kourtoglou, E., Mamma, D., Villas-Boas, S. G., Hatzinikolaou, D. G., and Christakopoulos, P. (2014a). Constitutive homologous expression of phosphoglucomutase and transaldolase increases the metabolic flux of Fusarium oxysporum. Microb Cell Fact 13:43. doi: 10.1186/1475-2859-13-43
Anasontzis, G. E., Kourtoglou, E., Mamma, D., Villas-Boâs, S. G., Hatzinikolaou, D. G., and Christakopoulos, P. (2014b). Constitutive homologous expression of phosphoglucomutase and transaldolase increases the metabolic flux of Fusarium oxysporum. Microb. Cell Fact. 13, 1–13. doi: 10.1186/1475-2859-13-43
Anasontzis, G. E., Zerva, A., Stathopoulou, P. M., Haralampidis, K., Diallinas, G., Karagouni, A. D., et al. (2011). Homologous overexpression of xylanase in Fusarium oxysporum increases ethanol productivity during consolidated bioprocessing (CBP) of lignocellulosics. J. Biotechnol. 152, 16–23. doi: 10.1016/j.jbiotec.2011.01.002
Ask, M., Olofsson, K., Di Felice, T., Ruohonen, L., Penttilä, M., Lidén, G., et al. (2012). Challenges in enzymatic hydrolysis and fermentation of pretreated Arundo donax revealed by a comparison between SHF and SSF. Process Biochem. 47, 1452–1459. doi: 10.1016/j.procbio.2012.05.016
Bettiga, M., Bengtsson, O., Hahn-Hagerdal, B., and Gorwa-Grauslund, M. F. (2009). Arabinose and xylose fermentation by recombinant Saccharomyces cerevisiae expressing a fungal pentose utilization pathway. Microb. Cell Fact. 8:40. doi: 10.1186/1475-2859-8-40
Cavka, A., Alriksson, B., Rose, S. H., Zyl, W. H., and Jönsson, L. J. (2010). Biorefining of wood: combined production of ethanol and xylanase from waste fiber sludge. J. Indust. Microbiol. Biotechnol. 38, 891–899. doi: 10.1007/s10295-010-0856-9
Christakopoulos, P., Kekos, D., Macris, B. J., Claeyssens, M., and Bhat, M. K. (1996a). Purification and characterisation of a major xylanase with cellulase and transferase activities from Fusarium oxysporum. Carbohydr. Res. 289, 91–104.
Christakopoulos, P., Macris, B. J., and Kekos, D. (1989). Direct fermentation of cellulose to ethanol by Fusarium oxysporum. Enzyme Microb. Technol. 11, 236–239. doi: 10.1016/0141-0229(89)90098-7
Christakopoulos, P., Mamma, D., Nerinckx, W., Kekos, D., Macris, B., and Claeyssens, M. (1996b). Production and partial characterization of xylanase from Fusarium oxysporum. Bioresour. Technol. 58, 115–119.
Christakopoulos, P., Nerinckx, W., Kekos, D., Macris, B., and Claeyssens, M. (1996c). Purification and characterization of two low molecular mass alkaline xylanases from Fusarium oxysporum F3. J. Biotechnol. 51, 181–189.
Christakopoulos, P., Nerinckx, W., Kekos, D., Macris, B., and Claeyssens, M. (1997). The alkaline xylanase III from Fusarium oxysporum F3 belongs to family F/10. Carbohydr. Res. 302, 191–195. doi: 10.1016/S0008-6215(97)00075-X
den Haan, R., van Rensburg, E., Rose, S. H., Gorgens, J. F., and van Zyl, W. H. (2015). Progress and challenges in the engineering of non-cellulolytic microorganisms for consolidated bioprocessing. Curr. Opin Biotechnol. 33, 32–38. doi: 10.1016/j.copbio.2014.10.003
Fan, J.-X., Yang, X.-X., Song, J.-Z., Huang, X.-M., Cheng, Z.-X., Yao, L., et al. (2011b). Heterologous expression of transaldolase gene tal from saccharomyces cerevisiae in Fusarium oxysporum for enhanced bioethanol production. Appl. Biochem. Biotechnol. 164, 1023–1036. doi: 10.1007/s12010-011-9191-5
Fan, J.-X., Yang, Q., Liu, Z.-H., Huang, X.-M., Song, J.-Z., Chen, Z.-X., et al. (2011a). The characterization of transaldolase gene tal from Pichia stipitis and its heterologous expression in Fusarium oxysporum. Mol. Biol. Rep. 38, 1831–1840. doi: 10.1007/s11033-010-0299-4
Fujita, Y., Takahashi, S., Ueda, M., Tanaka, A., Okada, H., Morikawa, Y., et al. (2002). Direct and efficient production of ethanol from cellulosic material with a yeast strain displaying cellulolytic enzymes. Appl. Environ. Microbiol. 68, 5136–5141. doi: 10.1128/AEM.68.10.5136-5141.2002
Kenealy, W., Zaady, E. D. U., Preez, J. C., Stieglitz, B., and Goldberg, I. (1986). Biochemical aspects of fumaric acid accumulation by Rhizopus arrhizus. Appl. Envir. Microbiol. 52, 128–133.
Kesava, S. S., Rakshit, S. K., and Panda, T. (1995). Production of ethanol by zymomonas-mobilis - the effect of batch step-feeding of glucose and relevant growth-factors. Process Biochem. 30, 41–47. doi: 10.1016/0032-9592(95)95703-L
Koppram, R., Albers, E., and Olsson, L. (2012). Evolutionary engineering strategies to enhance tolerance of xylose utilizing recombinant yeast to inhibitors derived from spruce biomass. Biotechnol. Biofuels 5:32. doi: 10.1186/1754-6834-5-32
Koppram, R., Nielsen, F., Albers, E., Lambert, A., Wannstrom, S., Welin, L., et al. (2013). Simultaneous saccharification and co-fermentation for bioethanol production using corncobs at lab, PDU and demo scales. Biotechnol. Biofuels 6:2. doi: 10.1186/1754-6834-6-2
Kordowska-Wiater, M., and Targonski, Z. (2002). Ethanol fermentation on glucose/xylose mixture by co-cultivation of restricted glucose catabolite repressed mutants of Pichia stipitis with respiratory deficient mutants of Saccharomyces cerevisiae. Acta Microbiol. Pol. 51, 345–352.
Kourtoglou, E., Anasontzis, G. E., Mamma, D., Topakas, E., Hatzinikolaou, D. G., and Christakopoulos, P. (2011). Constitutive expression, purification and characterization of a phosphoglucomutase from Fusarium oxysporum. Enzyme Microb. Technol. 48, 217–224. doi: 10.1016/j.enzmictec.2010.10.007
Krishnan, M., Ho, N., and Tsao, G. (1999). Fermentation kinetics of ethanol production from glucose and xylose by recombinant Saccharomyces 1400(pLNH33). Appl. Biochem. Biotechnol. 77–79, 373–388. doi: 10.1385/ABAB:78:1-3:373
Kuyper, M., A, A., van Dijken, J. P., and Pronk, J. T. (2004). Minimal metabolic engineering of Saccharomyces cerevisiae for efficient anaerobic xylose fermentation: a proof of principle. FEMS Yeast Res. 4, 655–664. doi: 10.1016/j.femsyr.2004.01.003
la Grange, D. C., den Haan, R., and van Zyl, W. H. (2010). Engineering cellulolytic ability into bioprocessing organisms. Appl. Microbiol. Biotechnol. 87, 1195–1208. doi: 10.1007/s00253-010-2660-x
Liu, L., Zhang, Y., Liu, Z., Petranovic, D., and Nielsen, J. (2015). Improving heterologous protein secretion at aerobic conditions by activating hypoxia-induced genes in Saccharomyces cerevisiae. FEMS Yeast Res. 15:fov070. doi: 10.1093/femsyr/fov070
Lynd, L., Elamder, R., and Wyman, C. (1996). Likely features and costs of mature biomass ethanol technology. Appl. Biochem. Biotechnol. 57–58, 741–761. doi: 10.1007/BF02941755
Malherbe, S., and Cloete, T. E. (2002). Lignocellulose biodegradation: fundamentals and applications. Rev. Environ. Sci. Technol. 1, 105–114. doi: 10.1023/a:1020858910646
Mead, O., Thynne, E., Winterberg, B., and Solomon, P. S. (2013). Characterising the role of GABA and its metabolism in the wheat pathogen Stagonospora nodorum. PLoS ONE 8:e78368. doi: 10.1371/journal.pone.0078368
Millati, R., Edebo, L., and Taherzadeh, M. J. (2005). Performance of Rhizopus, Rhizomucor, and Mucor in ethanol production from glucose, xylose, and wood hydrolyzates. Enzyme Microb. Technol. 36, 294–300. doi: 10.1016/j.enzmictec.2004.09.007
Monnet, C., Aymes, F., and Corrieu, G. (2000). Diacetyl and alpha -acetolactate overproduction by Lactococcus lactis subsp. lactis biovar diacetylactis mutants that are deficient in alpha -acetolactate decarboxylase and have a low lactate dehydrogenase activity. Appl. Environ. Microbiol. 66, 5518–5520. doi: 10.1128/AEM.66.12.5518-5520.2000
Nevoigt, E. (2008). Progress in metabolic engineering of Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 72, 379–412. doi: 10.1128/MMBR.00025-07
Okamoto, K., Imashiro, K., Akizawa, Y., Onimura, A., Yoneda, M., Nitta, Y., et al. (2010). Production of ethanol by the white-rot basidiomycetes Peniophora cinerea and Trametes suaveolens. Biotechnol. Lett. 32, 909–913. doi: 10.1007/s10529-010-0243-7
Panagiotou, G., Christakopoulos, P., and Olsson, L. (2005a). The influence of different cultivation conditions on the metabolome of Fusarium oxysporum. J. Biotechnol. 118, 304–315. doi: 10.1016/j.jbiotec.2005.05.004
Panagiotou, G., Christakopoulos, P., Villas-Boas, S. G., and Olsson, L. (2005b). Fermentation performance and intracellular metabolite profiling of Fusarium oxysporum cultivated on a glucose-xylose mixture. Enzyme Microb. Technol. 36, 100–106. doi: 10.1016/j.enzmictec.2004.07.009
Panagiotou, G., Pachidou, F., Petroutsos, D., Olsson, L., and Christakopoulos, P. (2008). Fermentation characteristics of Fusarium oxysporum grown on acetate. Bioresour. Technol. 99, 7397–7401. doi: 10.1016/j.biortech.2008.01.017
Panagiotou, G., Villas-Boas, S. G., Christakopoulos, P., Nielsen, J., and Olsson, L. (2005c). Intracellular metabolite profiling of Fusarium oxysporum converting glucose to ethanol. J. Biotechnol. 115, 425–434. doi: 10.1016/j.jbiotec.2004.09.011
Paschos, T., Xiros, C., and Christakopoulos, P. (2015). Ethanol effect on metabolic activity of the ethalogenic fungus Fusarium oxysporum. BMC Biotechnol. 15:15. doi: 10.1186/s12896-015-0130-3
Roa Engel, C., Straathof, A., Zijlmans, T., van Gulik, W., and van der Wielen, L. (2008). Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 78, 379–389. doi: 10.1007/s00253-007-1341-x
Roger, O., Lavigne, R., Mahmoud, M., Buisson, C., Onno, B., Zhang, B.-L., et al. (2004). Quantitative 2H NMR at natural abundance can distinguish the pathway used for glucose fermentation by lactic acid bacteria. J. Biol. Chem. 279, 24923–24928. doi: 10.1074/jbc.M401232200
Saitoh, S., Hasunuma, T., Tanaka, T., and Kondo, A. (2010). Co-fermentation of cellobiose and xylose using beta-glucosidase displaying diploid industrial yeast strain OC-2. Appl. Microbiol. Biotechnol. 87, 1975–1982. doi: 10.1007/s00253-010-2714-0
Shafiei, M., Karimi, K., and Taherzadeh, M. J. (2010). Pretreatment of spruce and oak by N-methylmorpholine-N-oxide (NMMO) for efficient conversion of their cellulose to ethanol. Bioresour. Technol. 101, 4914–4918. doi: 10.1016/j.biortech.2009.08.100
Singh, A., and Kumar, P. K. (1991). Fusarium oxysporum: status in bioethanol production. Crit. Rev. Biotechnol. 11, 129–147. doi: 10.3109/07388559109040619
Smart, K. F., Aggio, R. B. M., Van Houtte, J. R., and Villas-Boas, S. G. (2010). Analytical platform for metabolome analysis of microbial cells using methyl chloroformate derivatization followed by gas chromatography-mass spectrometry. Nat. Protocols 5, 1709–1729. doi: 10.1038/nprot.2010.108
Streeter, J., and Salminen, S. (1990). Periplasmic metabolism of glutamate and aspartate by intact Bradyrhizobium japonicum bacteroids. Biochim. Biophys. Acta 1035, 257–265. doi: 10.1016/0304-4165(90)90087-D
Sues, A., Millati, R., Edebo, L., and Taherzadeh, M. J. (2005). Ethanol production from hexoses, pentoses, and dilute-acid hydrolyzate by Mucor indicus. FEMS Yeast Res. 5, 669–676. doi: 10.1016/j.femsyr.2004.10.013
Keywords: Fusarium oxysporum, ethanol, consolidated bioprocessing, filamentous fungi, biofuels, fermentation, γ-aminobutyric acid shunt, NADPH regeneration
Citation: Anasontzis GE, Kourtoglou E, Villas-Boâs SG, Hatzinikolaou DG and Christakopoulos P (2016) Metabolic Engineering of Fusarium oxysporum to Improve Its Ethanol-Producing Capability. Front. Microbiol. 7:632. doi: 10.3389/fmicb.2016.00632
Received: 22 February 2016; Accepted: 18 April 2016;
Published: 04 May 2016.
Edited by:
Vijai Kumar Gupta, National University of Ireland, Galway, IrelandReviewed by:
Michael Sauer, University of Natural Resources and Life Sciences, AustriaAntonella Amore, National Renewable Energy Laboratory, USA
Copyright © 2016 Anasontzis, Kourtoglou, Villas-Boâs, Hatzinikolaou and Christakopoulos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul Christakopoulos, paul.christakopoulos@ltu.se
†Present Address: George E. Anasontzis, Biodiversité et Biotechnologie Fongiques, INRA - Aix Marseille Université, Marseille, France