 Dorota M. Krzyżanowska1
Dorota M. Krzyżanowska1 Adam Ossowicki1Magdalena Rajewska1
Adam Ossowicki1Magdalena Rajewska1 Tomasz Maciąg1Magdalena Jabłońska1Michał Obuchowski2
Tomasz Maciąg1Magdalena Jabłońska1Michał Obuchowski2 Stephan Heeb3
Stephan Heeb3 Sylwia Jafra1*
Sylwia Jafra1*- 1Laboratory of Biological Plant Protection, Department of Biotechnology, Intercollegiate Faculty of Biotechnology of University of Gdansk and Medical University of Gdansk, Gdansk, Poland
- 2Laboratory of Molecular Bacteriology, Department of Medical Biotechnology, Intercollegiate Faculty of Biotechnology University of Gdansk and Medical University of Gdansk, Medical University of Gdansk, Gdansk, Poland
- 3School of Life Sciences, Faculty of Medicine and Health Sciences, University of Nottingham, Nottingham, UK
Dickeya solani and Pectobacterium carotovorum subsp. brasiliense are recently established species of bacterial plant pathogens causing black leg and soft rot of many vegetables and ornamental plants. Pseudomonas sp. strain P482 inhibits the growth of these pathogens, a desired trait considering the limited measures to combat these diseases. In this study, we determined the genetic background of the antibacterial activity of P482, and established the phylogenetic position of this strain. Pseudomonas sp. P482 was classified as Pseudomonas donghuensis. Genome mining revealed that the P482 genome does not contain genes determining the synthesis of known antimicrobials. However, the ClusterFinder algorithm, designed to detect atypical or novel classes of secondary metabolite gene clusters, predicted 18 such clusters in the genome. Screening of a Tn5 mutant library yielded an antimicrobial negative transposon mutant. The transposon insertion was located in a gene encoding an HpcH/HpaI aldolase/citrate lyase family protein. This gene is located in a hypothetical cluster predicted by the ClusterFinder, together with the downstream homologs of four nfs genes, that confer production of a non-fluorescent siderophore by P. donghuensis HYST. Site-directed inactivation of the HpcH/HpaI aldolase gene, the adjacent short chain dehydrogenase gene, as well as a homolog of an essential nfs cluster gene, all abolished the antimicrobial activity of the P482, suggesting their involvement in a common biosynthesis pathway. However, none of the mutants showed a decreased siderophore yield, neither was the antimicrobial activity of the wild type P482 compromised by high iron bioavailability. A genomic region comprising the nfs cluster and three upstream genes is involved in the antibacterial activity of P. donghuensis P482 against D. solani and P. carotovorum subsp. brasiliense. The genes studied are unique to the two known P. donghuensis strains. This study illustrates that mining of microbial genomes is a powerful approach for predictingthe presence of novel secondary-metabolite encoding genes especially when coupled with transposon mutagenesis.
Introduction
Pseudomonas spp. constitute a highly diverse group of γ-proteobacteria, inhabiting various ecological niches (Palleroni, 2005) Over 200 species have been reported in the literature1, with at least 144 validated (Gomila et al., 2015). Representatives of the genus include pathogens affecting humans and animals (Pseudomonas aeruginosa; Sadikot et al., 2005), insects (Pseudomonas entomophila; Vodovar et al., 2005) and plants (mainly Pseudomonas syringae; Young, 2010; Baltrus et al., 2011). However, most pseudomonads are harmless commensals, some of which show plant-beneficial or biodegradation properties valuable for biotechnological applications (Adesemoye and Kloepper, 2009; Mercado-Blanco, 2015; Novik et al., 2015). Pseudomonas spp. produce a diverse array of biologically active metabolites, including antibiotics, bacteriocins, biosurfactants, toxins, volatiles, and iron-scavenging siderophores (Gross and Loper, 2009; Silby et al., 2011; Schulz-Bohm et al., 2015). Many of these metabolites increase the competitive potential of Pseudomonas spp., either by facilitating the colonization of a given niche and nutrient uptake, or through direct influence on the coexisting (micro)organisms (Pliego et al., 2011; Mercado-Blanco, 2015). This competitive edge is of particular importance in environments such as soil and the rhizosphere, where local “hot spots” of microbial activity are formed due to limited and uneven distribution of nutrients.
Numerous Pseudomonas-derived metabolites have been studied, many for their antimicrobial activity toward fungi and oomycetes and, to a considerably lesser extent, toward bacteria (Haas and Defago, 2005; Weller, 2007; Gross and Loper, 2009; Pierson and Pierson, 2010; Raaijmakers et al., 2010). Among the antimicrobials, the majority are polyketides (PK), (cyclic)non-ribosomal (lipo)peptides (NRPs or CLPs), or hybrid compounds (PK-NRP; Raaijmakers et al., 2006; Gross and Loper, 2009). Some strains, such as Pseudomonas protegens Pf-5 and CHA0T produce several antimicrobial compounds of different chemical classes (Loper et al., 2008; Ramette et al., 2011). The onset of the genomic era has created opportunities for discovering new antimicrobials especially given that over 1800 assemblies of Pseudomonas spp. genome sequences are currently available in GenBank (February 2016). The sizes of these genomes range from 4.17 Mbp for P. stutzeri JM300 (Busquets et al., 2012) to 7.7 Mbp for P. protegens Pf-5 (Paulsen et al., 2005). However, the majority of Pseudomonas spp. genomes can be considered large (≈6 Mbp), thereby reflecting the pool of genes and regulatory elements necessary to thrive in complex and dynamic environments (Raes et al., 2007; Goldfarb et al., 2011; Silby et al., 2011; Wu et al., 2011; Loper et al., 2012). In addition, the core genome of the genus is relatively small (approximately 25–35%). Taken together, this creates a considerable pool of strain-specific genes, some of which are involved in secondary metabolism conferring unique properties (Loper et al., 2012).
Pseudomonas sp. P482 is a tomato rhizosphere isolate, able to inhibit the growth of several plant pathogens, including the stone fruit pathogen P. syringae (Golanowska et al., 2012) and various strains of Dickeya and Pectobacterium (formerly Erwinia) genera (Krzyzanowska et al., 2012). The latter are plant pathogens that cause black leg and soft rot diseases of many vegetables and ornamental plants resulting in serious economic losses (Ma et al., 2007). As shown in this study, the spectrum of antibacterial activity of the P482 includes Dickeya solani and Pectobacterium carotovorum subsp. brasiliense, the recently established, highly virulent species of soft rot Enterobacteriaceae (SRE; Nabhan et al., 2012; van der Wolf et al., 2014). In the light of limited measures available to protect plants from soft rot (Czajkowski et al., 2011), as well the long history of Pseudomonas spp. strains as effective biological control agents against fungal diseases in agriculture and horticulture (Mercado-Blanco, 2015), the discovery of P482 with its novel antibacterial activity is potentially significant. Currently, the only chemically defined Pseudomonas-derived compound with antibacterial activity against soft rot bacteria is 2,4-diacetylphloroglucinol (2,4-DAPG). This polyketide antibiotic, although studied mainly for its antifungal properties (i.a., Harrison et al., 1993; Raaijmakers and Weller, 1998) also accounts for the in vitro antagonism of P. fluorescens F113 toward Erwinia carotovora subsp. atroseptica (currently Pectobacterium atrosepticum; Cronin et al., 1997). Despite other reports on Pseudomonas strains inhibiting the growth of soft rot bacteria, they all lack information on the mechanism of these antagonistic interactions (Krzyzanowska et al., 2012; Cigna et al., 2015; Raoul des Essarts et al., 2016).
Here, we elucidated the genetic background of the antibacterial activity of Pseudomonas sp. P482 toward SRE, with the focus on D. solani and P. carotovorum subsp. brasiliense. Recent publication of the draft genome of strain P482 (Krzyzanowska et al., 2014), as well as the ongoing development of bioinformatics tools, enabled us to employ genome mining data to identify novel secondary-metabolite gene clusters. Genomic data also enable us to establish the phylogenetic position of P482.
Materials and Methods
Bacterial Strains, Culture Conditions, and Growth Rate
Bacterial strains used in this study are listed in Table 1. All strains were cultured in Miller's Lysogeny Broth (LB) or on LB solidified with 1.5% agar (Novagen, Germany). The Pseudomonas spp. were grown at 28°C and the Escherichia coli ST18 was grown at 37°C. For the growth of the auxotrophic strain E. coli ST18, the medium was supplemented with 50 μg·ml−1 of 5-aminolevulonic acid (5-ALA; Sigma-Aldrich, USA). When necessary the medium was supplemented with kanamycin (30 μg·ml−1). For determination of bacterial growth rate the cells were cultured in 96-well plates and the OD595 measurements were performed hourly using an EnVision Multilabel Reader (PerkinElmer, USA).
Table 1. Bacterial strains used in this study.
Phylogenetic Analysis Based on the 16S rRNA Gene, MLSA, and Whole-Genome ANI
The 16S rRNA gene analysis was performed for partial (1384 nucl.) rss gene sequences of 52 Pseudomonas species (Table S1). The Multilocus Sequence Analysis (MLSA), performed according to Ramette et al. (2011), involved a concatenated set of partial sequences of three housekeeping genes: gyrB, rpoB, and rpoD. The total length of the concatenated set was 8328 nucleotides (2415, 4073, and 1840, respectively). All sequence alignments were performed using Clustal Omega (Sievers et al., 2011). Phylogenetic trees were constructed using MEGA 6.06 software (Tamura et al., 2013), Maximum Likelihood method, Kimura two-parameter model with bootstrap support 1000 replicates. Whole-genome average nucleotide identity (ANI) based on BLAST (version 2.2.18) was computed for pairwise alignment of stretches of genomes of using the JSpecies software with default settings2 (Richter and Rosselló-Móra, 2009). Apart from the type strains of respective Pseudomonas species, the well-studied strains Pseudomonas putida KT2440 and P. protegens Pf-5 were also subjected to ANI calculations. All nucleotide sequences used for the phylogenetic study were obtained from GenBank3, with the exception of the draft genome of Pseudomonas asplenii (unpublished data). The accession numbers of the analyzed genes and genomes are provided in Tables S1, S2, respectively.
Antibacterial Activity Assay
All bacterial strains for the assay were cultured overnight in LB medium at 28°C. Bacterial cells were harvested by centrifugation and re-suspended in sterile saline (0.9% NaCl). For the soft rot pathogens, the turbidity of each bacterial suspension was adjusted to 1 McFarland unit (DENSILAMETER II, Erba Lachema) and a sterile swab, soaked in the suspension, was used to inoculate the surface of an LB agar plate. For the potential antagonists, the turbidity of bacterial suspensions was adjusted to 4 McFarland units. Two microliter aliquots of each antagonist suspension were spotted on the surface of media pre-inoculated with the pathogens. The samples were incubated at 28°C for 16 h. The diameter of each pathogen inhibition zone was measured and the value obtained was normalized to the diameter of the bacterial antagonist colony forming the zone. Each experiment was performed in triplicate (n = 3), unless otherwise stated. To assess the role of iron availability in the antibiosis between P482 and the SRE, the LB agar plates were supplemented with filter-sterilized solution of FeSO4 to a final concentration of 15 μM.
Genome Mining for Secondary Metabolite-Encoding Genes
The annotated genome of strain P482 (JHTS00000000.1; Krzyzanowska et al., 2014) was searched for the presence of genes involved in the production of 26 metabolites (16 antimicrobials, six siderophores, two biosurfactants, and two compounds with unknown function) reported in literature to be produced by Pseudomonas spp. (Table S3). The search was performed at the protein sequence level using the local blastp tool (Altschul, 1997; Altschul et al., 2005) incorporated into Manatee4. Query coverage and identity values ≥50% were considered the cutoff values for a positive hit.
Additionally, the genome of the P482 strain was analyzed with an automatic pipeline called the “antibiotics and secondary metabolite analysis shell” (antiSMASH), version 2.05 (Medema et al., 2011; Blin et al., 2013). Both default settings (searched against the software's manually curated database) and settings involving the application of ClusterFinder algorithm (search based on Pfam domain probabilities; Cimermancic et al., 2014) have been used. More detailed information on the particular genes/gene products were obtained from the GenBank6 and the KEGG7 databases (Kanehisa and Goto, 2000).
Core and Variable Genome Analysis
The core genome and the pool of variable genes were determined using the EDGAR8 tool (Blom et al., 2009). The analyzed group comprised of Pseudomonas sp. P482 and three other, P482-related strains: P. donghuensis HYST (AJJP00000000), P. entomophila L48T (CT573326), and P. putida KT2440 (AE015451). For the purpose of the analysis, the genomes of HYST, L48T and KT2440 were re-annotated with the IGS Annotation pipeline, to match the annotation previously obtained for P482 (Krzyzanowska et al., 2014).
Site-Directed Mutagenesis
Fragments (316–453 bp) of genes to be inactivated were PCR-amplified using the Hot Start II Phusion DNA polymerase (Thermo Scientific). Details of the primer pairs used, annealing temperatures and the expected amplicons lengths are given in Tables S4, S5. The PCR products obtained were each cloned between the XbaI/XhoI restriction sites of the pKNOCK-Km suicide vector (Alexeyev, 1999). The resulting constructs, designated pKN1009, pKN3755, pKN4705, pKN4706, pKN4707, and pKN4709 (Table 2), were introduced into the E. coli ST18 donor strain (Thoma and Schobert, 2009) and subsequently transferred to Pseudomonas sp. P482 by biparental mating. In brief, cells from 1.5 ml of overnight LB cultures of both the donor and the recipient were washed twice with fresh LB medium and re-suspended in 0.5 ml of LB. The two suspensions were pooled (1:1) and the cells harvested by centrifugation. Bacterial pellets were re-suspended in a droplet (20–30 μl) of LB and spotted onto an LB agar plate. The sample was incubated for 16 h at 37°C. The macro-colony obtained was scratched from the medium and suspended in 1 ml of LB. One hundred microliter aliquots of the suspension and serial dilutions (10−1, 10−2, 10−3) were plated on LB agar supplemented with kanamycin (30 μg·ml−1) but lacking 5-ALA, thus preventing the growth of the E. coli ST18. The P482 transconjugants obtained were screened for the presence of the pKNOCK-Km insertion with primers F_pKNOCK_backbone and R_pKNOCK_backbone. To confirm that the suicide vector had incorporated into the target loci, genomic DNA of each mutant was used as template in a sequencing reaction with primer F_outof_pKNOCK. The results obtained enabled mapping of the pKNOCK-Km insertion site to the genome of the P482 strain. The sequencing was performed at Oligo.pl (Warsaw, Poland).

Table 2. Vectors used in this study.
Transposon Mutagenesis
Plasmid pRL27 bearing the mini-Tn5 transposon (Larsen et al., 2002) was transferred into E. coli ST18. The resulting strain E. coli ST18 [pRL27] was used to deliver pRL27 into P482 by biparental mating, using the same protocol as described above for the site-directed mutagenesis. The P482 mutants obtained were screened for the loss of antimicrobial activity against soft rot bacteria using D. solani IFB0102. In the case of mutants showing decreased antimicrobial activity, the integration site of the mini-Tn5 transposon was mapped. For this purpose, the genomic DNA of each mutant was used as a template in sequencing reactions with primers tpnRL13–2_F_LONG and tpnRL17–1_R_LONG (Table S4). Both primers anneal near the ends of the transposon and their 3′ ends face outwards. The resulting sequences, obtained at Oligo.pl (Warsaw, Poland), were used as query in a local blastn search against the genome of the P482 strain.
Detection of Siderophore Production on CAS Agar
Total siderophore production by the Pseudomonas spp. strains studied was assessed on CAS blue agar (Schwyn and Neilands, 1987). Bacterial cells were cultured overnight in LB medium, harvested by centrifugation, and re-suspended in sterile saline (0.9% NaCl). The turbidity of the suspensions was adjusted to 4 McFarland units. Two microliters of each suspension were spotted on CAS agar plates, in two technical replicates. The plates were incubated at 28°C for 24 h, and then another 96 h at room temperature (22°C) for the development of orange halos. Following incubation, the diameter of each halo was measured and the value obtained was normalized to the diameter of the bacterial colony. The experiment was performed twice (n = 2), with two technical replicates.
Pyoverdine Production in CAA and MKB Media
Two types of iron-poor media: CAA (5 g casamino acids, 1.18 g K2HPO4·3H2O, 0.25 g MgSO4·7 H2O, per liter; Kümmerli and Brown, 2010) and MKB (5 g casamino acids, 2.5 g K2HPO4, 15 ml glycerol, 2.5 g MgSO4·7 H2O, per liter, pH 7.2; Yu et al., 2014), were inoculated (1:1000) with overnight cultures of Pseudomonas sp. P482, P. donghuensis HYST and the P482 mutants: KN4705, KN4706, KN4707, KN4709, KN1009, KN3755. Strain Pseudomonas vranovensis DSM 16006T, a non-fluorescent pseudomonad unable to produce pyoverdine, was used as a reference. Two hundred microliter aliquots of the inoculated media were transferred to the wells of a 96-well plate, in four replicates for each strain and medium. The plates were incubated for 48 h at 28°C, without shaking. Following incubation, the pyoverdine fluorescence level (excitation λ = 400 nm, emission λ = 460 nm) was measured, together with the optical density of the cultures (λ = 600 nm; EnVision Multilabel Plate Reader, Perkin Elmer). The level of pyoverdine production per cell is presented as relative fluorescence units (RFU) = fluorescence (400/460)/OD600.
In silico Screening for Promoter and Terminator Regions
The sequence of interest (contig JHTS01000055.1, range 27,755–36,623) was analyzed for the presence of sigma housekeeping promoter sequence by three different programs: PromoterHunter9 (Klucar et al., 2010), Promoter prediction10 (Reese, 2001) and BPROM 11 (Solovyev and Salamov, 2011). As the PromoterHunter requires a weight matrix for the −10 and −35 sequences, such a matrix was created based on the P. aeruginosa promoters (Potvin et al., 2008), available at phiSITE12. The ARNold13 program was employed for the identification of rho-independent terminators (Lambert et al., 2004).
Identification of Prophages and Genomic Islands
The PHAST14 software (Zhou et al., 2011; was applied to identify prophage sequences within the draft genome sequence of P482. IslandPick 3.015 (Dhillon et al., 2015) was used to screen the genome for the presence of genomic islands. As part of the IslandPick analysis, the contigs comprising the draft genome were aligned using the genome of P. entomophila L48T as a reference. L48T was chosen as the most closely related species for which a complete genomic sequence is available.
Results
Pseudomonas sp. P482 Is a New Representative of P. donghuensis
Phylogenetic studies based on the analysis of 16S rRNA gene sequences (Figure 1A) and MLSA (Figure 1B) revealed that the closest relatives of strain P482 are P. donghuensis HYST, an isolate from lake water in China studied for high siderophore yield (Gao et al., 2015), P. vranovensis DSM 16006T, a non-fluorescent pseudomonad obtained from soil next to a highway in Czech Republic (Tvrzová et al., 2006), and a well-studied insect pathogen, P. entomophila L48T (Mulet et al., 2012). The availability of genomic data for P482 and the three related strains enabled us to investigate whether P482 belongs to one of these species by calculation of ANI-values for pairwise alignment of genomes (Konstantinidis and Tiedje, 2005; Goris et al., 2007; Richter and Rosselló-Móra, 2009). The highest ANI-value of 99.24% was obtained for the comparison of genomes of strains P482 and HYST (Table 3). ANI-values calculated for the comparison of genome of P482 with the genomes of P. vranovensis DSM 16006T and P. entomophila L48T were 85.34 and 81.48%. respectively, thus much below the single species value. Considering that the postulated single species threshold is ANI ~ 95–96%, the Pseudomonas sp. P482 should be classified as the same species as P. donghuensis. This species has recently been established by Gao et al. (2012), with strain HYST as a type strain and the only known representative. High similarity between P482 and HYST was also confirmed at the biochemical level using API 20NE and API 50CH (bioMérieux, France; Table S6). A comparative genome analysis using EDGAR showed that the number of unique ORFs found for the P482 and HYST is 222 (4.3%) and 345 (6.5%), respectively. Some of the differences may have a reason since both genomes are in a draft format. The core genome calculated for P482 and three other related Pseudomonas species (P. vranovensis DSM 16006T, P. entomophila L48T, P. putida KT 2440) represents 60–64% of each genome (Figure 2).
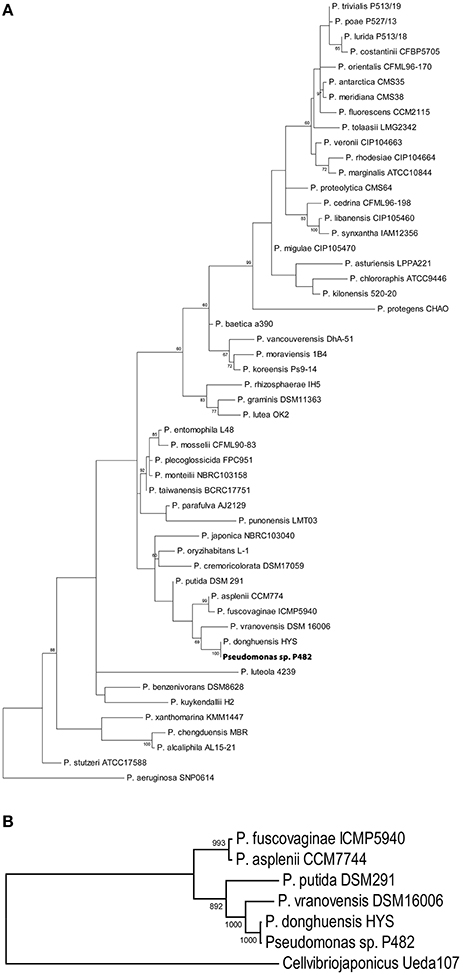
Figure 1. Phylograms depicting the results of 16S rRNA analysis (A) and MLSA (B) for P482 and related Pseudomonas spp. type strains. The phylogram based on 16S rRNA gene analysis was constructed using maximum likelihood method with Kimura 2-parameter using MEGA 6 software. Bootstrap values are shown at the nodes if the value is >60%. Except for Pseudomonas sp. P482, all strains used in the analysis are type strains. Pseudomonas aeruginosa SNP 0614 was used as the outgroup. Accession numbers of all the gene sequences included are listed in Table S1. The MLSA was performed for a set of partial nucleotide sequences of three genes: gyrB, rpoB, and rpoD (8328 nucleotides). Pseudomonas sp. P482 and five other Pseudomonas spp. strains were included because of the short genetic distance between them (see panel A). The MLSA-based phylogram was constructed using maximum likelihood method with GTR + I + G model estimated by jModelTest2 software. Bootstrap values are shown at the nodes. Cellvibrio japonicas Ueda107 was used as the outgroup.
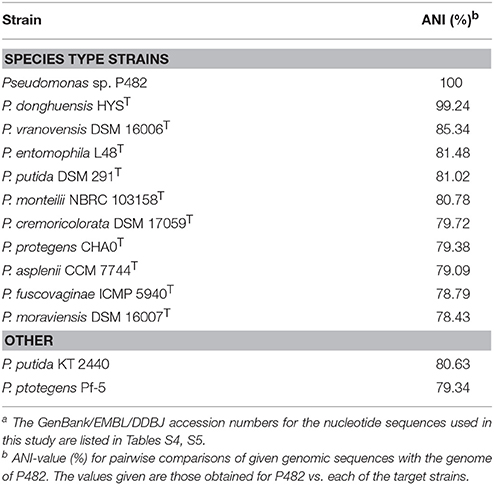
Table 3. ANI-values for pairwise alignment of genomes, calculated for the P482 and the type strains of closely related Pseudomonas speciesa.
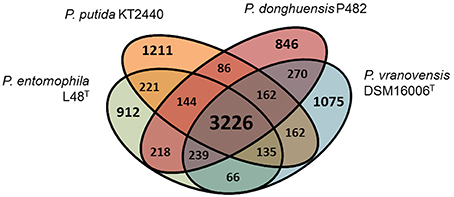
Figure 2. Venn diagram for the comparison of four Pseudomonas spp. genomes, including P. donghuensis P482 and three closely related strains. The calculated core genome (3226 ORFs, shown in large font) represents 60–64% of each genome. The results for P. donghuensis HYST are not shown for clarity. The number of unique ORFs found for the P482 and HYST was 222 (4.3%) and 345 (6.5%), respectively.
Comparison of the Antibacterial Activity of P482 with Other Related Pseudomonas spp.
Having established the phylogenetic position of P482, we investigated whether the ability to inhibit D. solani and P. carotovorum subsp. brasiliense growth is a unique property of P482, or rather that this trait is more widespread among P482-related species. P482 and 10 Pseudomonas spp. strains, closely related to the studied strain based on ANI calculations, were tested for their ability to inhibit the growth of D. solani and P. carotovorum subsp. brasiliense. Two strains from each pathogenic species were included in the assay—the type strain (IPO 2222T and LMG21371T) and one recently obtained environmental isolate (IFB0102 and JJ 56). This experiment revealed that only P. donghuensis HYST possessed similar antibacterial properties to P482, given that it inhibited growth of all soft rot strains tested (Figure 3). Among other Pseudomonas spp. strains, only P. entomophila L48T and the two 2,4-DAPG producing P. protegens strains, CHA0T and Pf-5, showed measurable antibiosis toward the soft rot pathogens. L48T inhibited the growth of D. solani (approximately 75% activity of that of P482) but was inactive against P. carotovorum subsp. brasiliense. In contrast the P. protegens strains inhibited the growth of all four pathogens. However, their activity with respect to P482 was relatively high against P. carotovorum subsp. brasiliense (57–100%), but low against D. solani (8–44%). This shows that the antimicrobial activity pattern of P482 and HYST is unique to the two P. donghuensis strains.
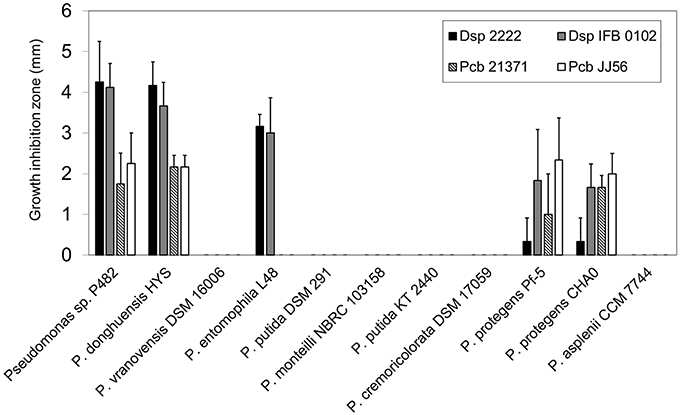
Figure 3. Antibacterial activity of Pseudomonas sp. P482 and the type strains of P482-related species against D. solani and P. carotovorum subsp. brasiliense. Four soft rot bacterial strains were tested: D. solani type strain IPO 2222T, D. solani IFB 0102, P. carotovorum subsp. brasiliense type strain LMG21371T, and P. carotovorum subsp. brasiliense JJ 56. The order of the Pseudomonas strains from left to right reflects the their degree of their relatedness to P482, as estimated from ANI calculations. The histogram shows the mean of three independent experiments, and error bars show standard deviations.
Mining of the P482 Genome for Genes Conferring the Production of Biologically Active Metabolites
Manual Search Focusing on Pseudomonas-Derived Compounds
The genome of P482 was manually searched for the presence of genetic elements essential for the synthesis of biologically active metabolites, previously described for Pseudomonas spp. The analysis included 16 metabolites with reported antifungal and/or antibacterial activity (2,4-DAPG, 5-dialkylresorcinols, quinolones, hydrogen cyanide, phenazines, xantholysin, massetolides, mupirocin, orfamides, pyoluteorin, pyrrolnitrin, rhizoxins, syringopeptin, syringomycin and viscosin), six siderophores (achromobactin, quinolobactin/thioquinolobactin, non-fluorescent siderophore, pyoverdin, pseudomonin, pyochelin), two compounds described as biosurfactants (arthrofactin, putisolvin), and two compounds of unknown function (paerucumarin, pseudoverdin). The analysis performed showed that P482 contains genes enabling the production of only three of the compounds investigated: toxic volatile hydrogen cyanide, the siderophore pyoverdine and a chemically undefined, recently described non-fluorescent siderophore of P. donghuensis HYST (Yu et al., 2014; Table S3).
Automated Search Using the antiSMASH 2.0 Pipeline
The genome of P482 was subjected to an automated search using the “antibiotics and secondary metabolite analysis shell” (antiSMASH 2.0). For detailed results of the antiSMASH analysis see Table 4. When the default settings were applied to the genome of P. donghuensis P482, five gene clusters were identified: two non-ribosomal peptide synthases (“NRPS”), one designated as “Bacteriocin,” and two classified more generally as “Other.” The two NRPS-type clusters identified consisted of genes potentially involved in the synthesis of pyoverdine (Table 4).
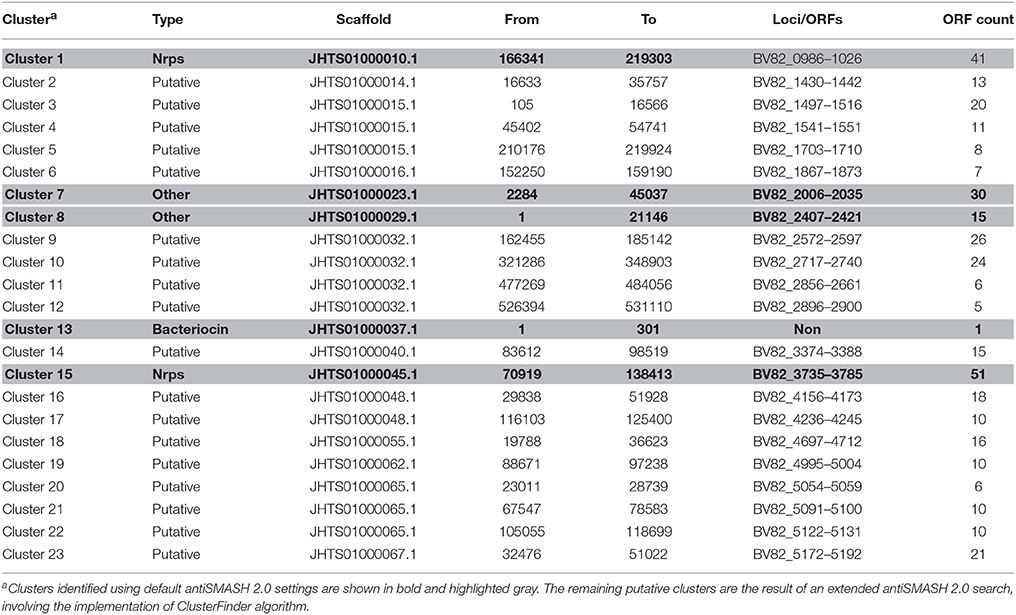
Table 4. Gene clusters potentially involved in the synthesis of secondary metabolites and antibiotics by P482, identified using the antiSMASH 2.0.
An “extended” search of the P482 genome, involving the implementation of ClusterFinder, led to the detection of 18 additional gene clusters with the status “Hypothetical” (Table 4). For “Hypothetical” clusters, no suggestions regarding their products and functions are provided. One of the “Hypothetical” clusters, no. 18, incorporated among other, loci BV82_4708–4711 (Figure 4), encoding the homologs of four genes essential for the production of the non-fluorescent siderophore of P. donghuensis HYST (Yu et al., 2014).
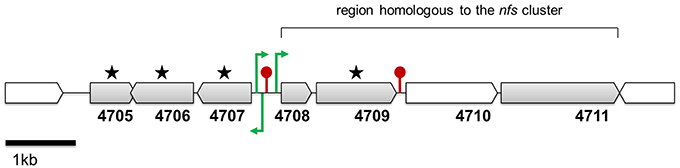
Figure 4. Genomic region conferring the antibacterial activity of strain Pseudomonas sp. P482 toward soft rot bacteria. Genes marked with stars were inactivated by mutagenesis and the corresponding P482 mutants were impaired in antibacterial activity. ORFs shown in gray encode proteins that have none or few homologs in other Pseudomonas spp. The locations of the promoters (green arrows) and the terminators (red pins) are not drawn to scale. Their precise locations in contig JHTS01000055.1 are provided in Supplementary Materials (Tables S9, S10). Annotations of the depicted genes: 4705—bacterial regulatory, tetR family protein; 4706—HpcH/HpaI aldolase/citrate lyase family protein; 4707—short chain dehydrogenase family protein; 4708—thioesterase superfamily protein; 4709—acyl-CoA dehydrogenase, C-terminal domain protein; 4710—phenylacetate-CoA ligase; 4711—thiamine pyrophosphate enzyme.
Overall this in silico search using antiSMASH led to identification of 23 gene clusters, potentially involved in the synthesis of secondary metabolites. None of the clusters was involved in the synthesis of previously described antibiotics of microbial origin. Twenty clusters, comprising 281 genes in total, had no predicted function.
Identification of Genes Involved in the Antibacterial Activity of P482
In order to identify gene(s) involved in the antibacterial activity of P482, we performed random mutagenesis using the mini-Tn5 transposon. Screening of over 5000 mutants allowed us to select S0405—a P482 transposon mutant unable to inhibit the growth of D. solani (IPO 2222T, IFB0102) and P. carotovorum subsp. brasiliense (LMG 21371T, JJ 56; Figure 5). The transposon insertion in mutant S0405 was located in locus BV82_4706. The product of this gene is annotated as HpcH/HpaI aldolase/citrate lyase family protein, and harbors a domain typical for CitE—the β subunit of the citrate lyase (EC 4.1.3.6). The holoenzyme is involved in the fermentation of citrate by a few bacterial species, and is composed of three protein subunits: α, β and γ, encoded by the citF, cite, and citD genes, respectively (Bott and Dimroth, 1994; Meyer et al., 1997). The putative CitE from P482 shares only 30% identity to the CitE protein of Klebsiella pneumoniae (CAA56216) but is not accompanied by the neighboring citF and citD genes. In line with this observation, blastp search of the P482 protein dataset using the CitF (CAA56217.1) and CitD (CAA56215.1) sequences as queries gave no relevant matches.
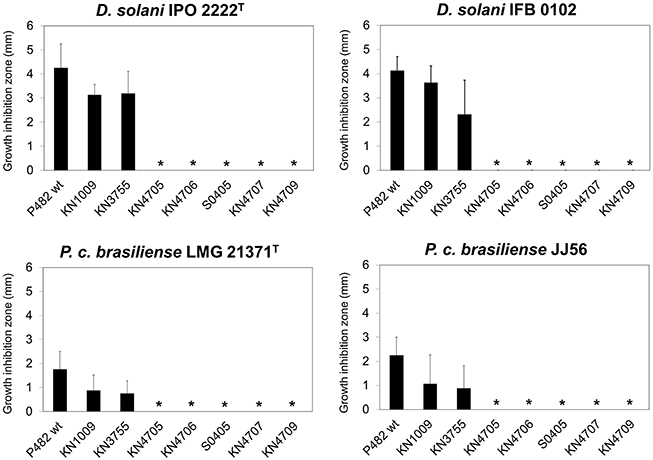
Figure 5. Antibacterial activity of Pseudomonas sp. P482 and its mutants toward D. solani and P. carotovorum subsp. brasiliense strains. The histogram shows the mean of four independent experiments (n = 4), and error bars show standard deviations. Results statistically different from those obtained for the reference strain (P482 wt) in two-tailed Student's t-test assuming equal variances (α = 0.05) are marked with stars.
The BV82_4706 gene is located upstream of the homologs of the four genes of the nfs cluster from P. donghuensis HYST (loci BV82_4708–4711) but oriented in the opposite direction. Moreover, each of the genes described above occurs in one of the antiSMASH-predicted “Hypothetical” clusters, here assigned number 18. At the nucleotide level, the whole of cluster 18 (8522 bp, loci BV82_4697-BV82_4712) shares 99% identity with the region 6854–15376 of Scaffold 1 (AJJP01000005.1) of the draft genome of P. donghuensis HYST.
Analysis of cluster 18 in more detail revealed that it consist of 16 open reading frames (loci BV82_4697-BV82_4712; Table 5). Compared with genomic regions found in other Pseudomonas spp., cluster 18 can be divided into two parts. The first part (loci BV82_4697-BV82_4704, with the exception of the internally located BV82_4702) encodes proteins having multiple homologs of Pseudomonas origin in the NCBI non-redundant protein database (over 100 hits with query coverage >90% and identity >70% each; Table 5). Among these loci, four (BV82_4697-BV82_4700) are putatively involved in the synthesis of alginate—an exopolysaccharide produced by bacteria for protection against environment threats and to enhance adhesion to solid surfaces (Boyd and Chakrabarty, 1995). In contrast, there were few blastp hits of Pseudomonas origin for the loci comprising the second part of the cluster (BV82_4705-BV82_4712, with the exception of BV82_4710; 3–20 hits per locus, including the P482 and HYST; Table 5). These results are in line with the comparative genome analysis performed for P482, HYST and three other related Pseudomonas species (P. vranovensis DSM 16006T, P. entomophila L48T, P. putida KT 2440; Figure 2). This analysis showed that the loci BV82_4705–4711 are among the reading frames that are unique to the P. donghuensis strains P482 and HYST (Figure 4).
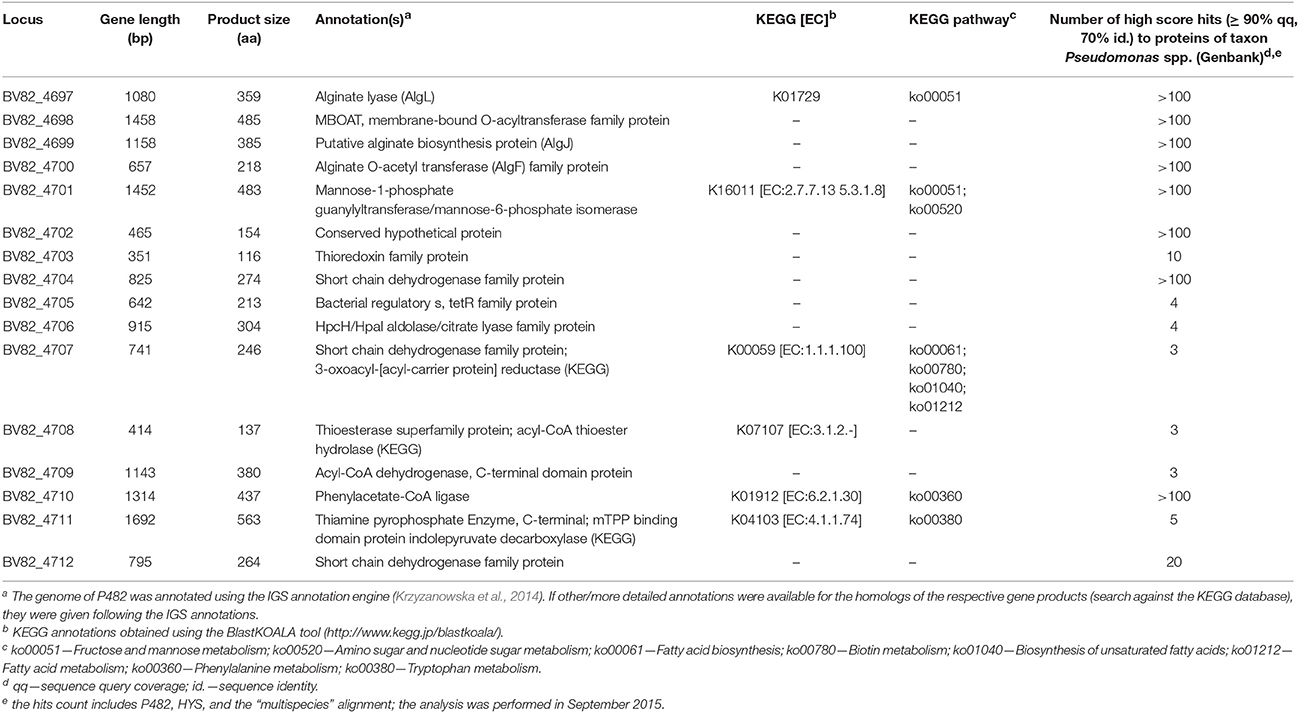
Table 5. Annotation and features of the genes comprising cluster 18—one of the antiSMASH-predicted gene clusters, putatively responsible for the synthesis of an unknown secondary metabolite(s) by Pseudomonas sp. P482.
Based on the results obtained in silico, we performed site-directed mutagenesis of four genes, all located in the “unique region” of cluster 18. The loci inactivated were (predicted products in brackets): BV82_4705 (regulatory protein of the TetR family), BV82_4706 (HpcH/HpaI aldolase/citrate lyase family protein), BV82_4707 (short chain acyl dehydrogenase family protein) and BV82_4709 (acyl-CoA dehydrogenase), the latter being an ortholog of the nfs cluster essential for non-fluorescent siderophore production (Yu et al., 2014; Figure 4, Table 5). Additionally, we carried out site-directed mutagenesis of loci BV82_1009 (psvA/pvdL) and BV82_3755 (in part similar to pvdD of P. aeruginosa), the major genes of the two antiSMASH-predicted “NRPS” clusters, potentially involved in the synthesis of a pyoverdine-like siderophore. Antibacterial activity assays have shown that mutants KN4705, KN4706, KN4707, and KN4709 had completely lost their in vitro antagonism toward the soft rot pathogens tested, as observed earlier for the mini-Tn5 mutant S0405 (Figure 5). In contrast, for mutants with disrupted pyoverdine production (KN1009 and KN3755), no statistically significant decrease in the diameter of growth inhibition zones could be observed, with respect to the parental strain P482 (Figure 5). None of the mutants constructed in this study had impaired growth rate in LB medium as used in the antibacterial activity assays (data not shown).
Thus, disruption of the nfs cluster, as well as three genes located upstream, affects the production of a compound playing a major role in the in vitro antagonism of P482 toward the soft rot bacteria.
Prediction of Promoters and Terminators in the Genomic Region BV82_4705–4712
The cluster 18 region comprising genes BV82_4705–4712 was analyzed for the presence of putative promoters and transcriptional terminators. Three out of several hypothetical promoters were unanimously predicted by all programs used, all located in the spacer region between genes BV82_4707 and BV82_4708 (Figure 4; Table S9). Two of these may drive transcription toward BV82_4708—the first of the four-gene ortholog nfs cluster. The third promoter is located in the opposite direction, and may drive transcription toward the BV82_4707 gene. Next, rho-independent terminators were searched using ARNold. Two possible terminator sequences were identified (Table S10). The first terminator is located between loci BV82_4707 and BV82_4708, thus separating the nfs cluster homologs from the BV82_4706 and BV82_4707 genes, encoding the HpcH/HpaI aldolase/citrate lyase and the short chain alkyl dehydrogenase, respectively. The second terminator sequence was found between open reading frames BV82_4709 and BV82_4710. The promoters and terminators identified are also present in the HYST strain. This in silico study requires experimental verification. However, both the relative position of the genes and the position of the putative promoters and terminators suggest that the insertional mutagenesis of the BV82_4705, BV82_4706, and BV82_4707 by the pKNOCK system should not affect on the downstream nfs loci. Nevertheless, some concerns arise because inactivation of the BV82_4705 gene may influence expression of the partially overlapping BV82_4706.
Total Siderophore Production
Total siderophore production on CAS agar was assessed for the P482 wt strain and its mutant derivatives: KN4705, KN4706, S0405, KN4707, KN4709, KN1009, and KN3755. In the case of KN1009 and KN3755, that both contain inactivated pyoverdine synthesis pathway genes, a significant (≈80%) decrease in siderophore production was observed (Figure 6, Figure S1A). The results obtained show that, under these assay conditions, pyoverdine plays the major iron scavenging role, despite the presence of a second siderophore biosynthetic locus. In contrast, the inactivation of the BV82_4709 (nfs gene homolog), as well as the three upstream genes (BV82_4705, BV82_4706, and BV82_4707), did not significantly affect total siderophore production (Figure 6).
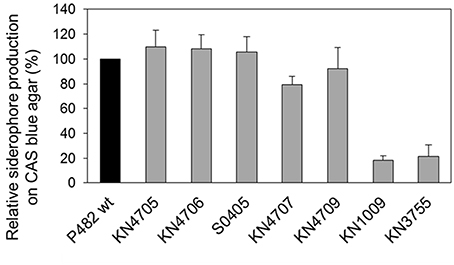
Figure 6. Total siderophore production on CAS blue agar. The histogram shows the relative siderophore production (%) by the P482 mutants (gray bars) with respect to the wild type strain (black bar). Error bars indicate standard deviations resulting from two independent experiments.
Additionally, total siderophore production exhibited by P482 was compared with that of HYST and four other well-studied Pseudomonas spp.: P. aeruginosa PAO1, P. protegens strains CHA0T and Pf-5, and P. putida DSM 291T. Under the experimental conditions used, strains P482 and HYST produced comparable levels of siderophores, however, these were markedly lower than CHA0T, Pf-5, DSM 291T, and PA01 (Figure S1B).
Production of Pyoverdine
The P. donghuensis strains P482 and HYST showed comparable levels of pyoverdine in CAA and MKB media (Figure 7). Four P482 mutants KN4705, KN4706, KN4707, and KN4709, showed no significant differences in pyoverdine levels with respect to the wild-type P482. In contrast, the P482 mutants KN1009 and KN3755 in common with P. vranovensis DSM 16006T, do not demonstrate any fluorescent siderophore activity suggesting that they do not produce pyoverdine (Tvrzová et al., 2006). This is consistent with the data mining results which indicated high similarity of the BV82_1009 and BV82_3755 gene products to sequences of proteins involved in pyoverdine synthesis.
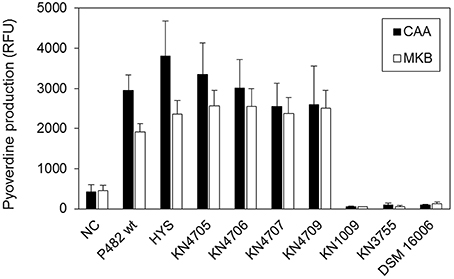
Figure 7. Pyoverdine production in CAA and MKB media. The level of pyoverdine production by Pseudomonas sp. P482 mutants and the related Pseudomonas spp. strains (P. donghuensis HYST and P. vranovensis DSM 16006T), was measured in two different iron-poor media: CAA (black bars) and MKB (white bars). Results are presented in relative fluorescence units (RFU), calculated as the ratio of fluorescence level (excitation 400 nm, emission 460 nm) to the optical density of the culture at 600 nm. Error bars on the histogram show standard deviations between eight technical replicates from a single experiment.
Role of Iron Availability in the In vitro Antagonism between P482 and Soft Rot Bacteria
To assess the influence of high iron availability on the antibacterial activity of P482 toward soft rot bacteria, the growth inhibition assay was performed on LB agar supplemented with 15 μM FeSO4. No reduction in growth inhibition zone was observed on this medium for any of the soft rot pathogens (Figure 8). This shows that antagonism between P482 and the soft rot bacteria does not depend on competition for iron alone. Moreover, the antibacterial compound is produced by P482 grown under conditions of high iron availability.
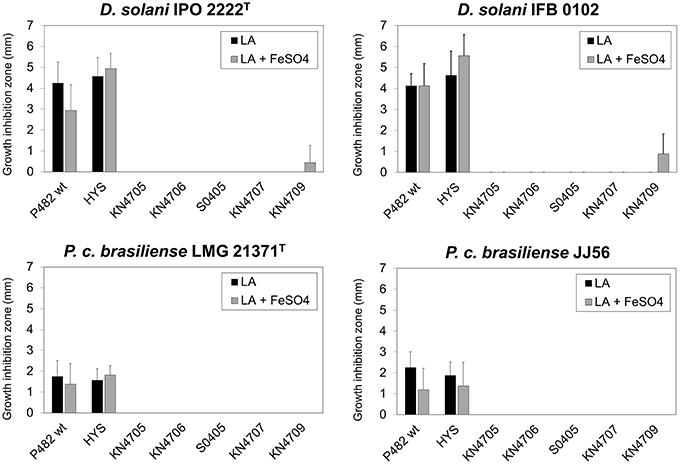
Figure 8. Influence of iron availability on the antimicrobial activity of Pseudomonas sp. P482 and its mutants. The histogram shows the mean of four independent experiments, and error bars show standard deviations. Black bars represent the diameter of the respective pathogen growth inhibition zone on LB agar medium. The gray bars are the results of the assay performed on LB agar supplemented with 15 μM FeSO4.
Investigation of the Origin of the Genes Involved in the Antibacterial Activity
The P482 draft genome harbors four prophage-like regions, including two prophages scored as “intact” (Table S7), and 11 putative genomic islands (Table S8). Two of the identified genomic islands, consisting of loci BV82_0239—BV82_0241 and BV82_3041—BV82_3048, are directly adjacent to two of the prophage-encoding regions. However, none of the regions showing evidence for horizontal acquisition overlap with the genes of cluster 18 that are involved in the antibacterial activity of P482.
Discussion
Pseudomonas spp. are well-known for their antagonistic properties toward different fungal pathogens as a consequence of the production of antimicrobials and other secondary metabolites (Haas and Defago, 2005; Raaijmakers and Mazzola, 2012). Some of these compounds are also active against bacteria (i.a., Keel et al., 1992), although the mode of antibacterial action of these Pseudomonas-derived compounds remains as yet, poorly understood. In this study, we determined the genetic background of the antibacterial activity of strain P482, a fluorescent pseudomonad obtained from the rhizosphere of tomato (Golanowska et al., 2012; Krzyzanowska et al., 2012), toward the pectolytic bacteria D. solani (van der Wolf et al., 2014) and P. carotovorum subsp. brasiliense (Nabhan et al., 2012). Over the last decade, these two microorganisms have been reported as highly aggressive plant pathogens causing major economic losses in potato production (Duarte et al., 2004; Sławiak et al., 2009; Tsror et al., 2009; van der Merwe et al., 2009; Laurila et al., 2010; Toth et al., 2011; Leite et al., 2014). Their worldwide dissemination requires the development of novel control strategies, preferably environmentally friendly (i.e., based on natural compounds and microorganisms), as encouraged by European Union regulations (2009/128/EC). Apart from establishing genetic determinants of antibacterial properties of P482, we sought to establish the taxonomic position of this strain.
Pseudomonas sp. P482 Is a Novel Representative of P. donghuensis
Our study revealed that P482 should be classified as Pseudomonas donghuensis—a species recently established with HYST as its type strain and sole representative (Gao et al., 2015). The genomes of the P482 and HYST are estimated to be of comparable size, respectively, 5.639 and 5.648 Mbp (Krzyzanowska et al., 2014; Gao et al., 2015) with an ANI-value for their comparison (the mean identity for the alignments of all homologous genes) exceeding 99%. A comparative genome analysis performed for this pair of strains using EDGAR revealed that the fraction of non-homologous genes, known as the accessory (unique) genome, is only 6.5% (HYST) to 4.3% (P482). Another potential sequence-based relationship between the strains could be the synteny of their genomes, thus the physical co-localization of genetic loci. This, however, cannot be compared for genomes that are still in their draft form, as is the case for both P482 and HYST. Nevertheless, the two strains, P482 and HYST, are highly similar at the genomic level and also phenotypically comparable.
P482 strain was obtained from the rhizosphere of tomato in Gdynia (Poland) and HYST originates from the waters of Donghu lake (China; Gao et al., 2015). The level of similarity between these microorganisms, isolated at distant locations, suggests a relatively recent common origin. This is likely to be a consequence of spreading via the international exchange of plant materials (i.e., Sławiak et al., 2009). However, P482 and HYST are not the only examples reported of close phylogenetic relations between non-pathogenic Pseudomonas spp., isolated from different geographical regions. For example, ANI-values above 98% were calculated for the alignment of the genome of Pseudomonas sp. Cab57, obtained from the rhizosphere of a shepherd's purse in Japan, with the genomes of both P. protegens CHA0T (98.47%) and P. protegens Pf-5 (98.22%; Takeuchi et al., 2014). The latter two were isolated from the rhizosphere of tobacco in Switzerland (Stutz, 1986) and the rhizosphere of cotton in USA (Howell and Stipanovic, 1979).
Growth-Inhibitory Properties of a Novel Antimicrobial Produced by P482
Multiple secondary metabolites of bacterial origin have been reported, and the genes essential for their synthesis identified (Newman and Cragg, 2012). For our study of Pseudomonas sp. P482, we assumed that the growth-inhibitory activity of this strain toward the soft rot pathogens was likely to depend on one or more previously identified compounds. However, mining of the P482 genome failed to reveal any genes involved in the synthesis of known antibiotics. In this respect, P482 was only found to harbor genes enabling the synthesis of hydrogen cyanide and two siderophores: pyoverdine (Cornelis and Matthijs, 2002; Visca et al., 2007) and a non-fluorescent siderophore, previously reported for P. donghuensis HYST (Yu et al., 2014). Hydrogen cyanide, a volatile produced by several bacterial species, is a potent inhibitor of cytochrome c oxidase and several other metalloenzymes, and hence highly toxic for eukaryotic cells (Blumer and Haas, 2000). The headspace volatiles of P482 grown on LB agar did not inhibit the growth of D. solani in an in vitro assay, thereby excluding contribution of HCN to antibiosis (Ossowicki, unpublished data). The non-ribosomal peptides from the pyoverdine family are high-affinity iron scavengers of the so-called “fluorescent pseudomonads” (Cornelis and Matthijs, 2002; Visca et al., 2007). To our knowledge, these compounds have not been reported as the primary cause of in vitro antibiosis between pseudomonads and other bacterial species. In line with this assumption, the pyoverdine-deficient P482 mutants, KN1009 and KN3755, retained their ability to inhibit the growth of the soft rot pathogens.
Identification of Genes Involved in the In vitro Antagonism of P482 toward D. solani and P. carotovorum subsp. brasiliense
Strain S0405, a P482 transposon mutant lacking antibacterial activity against soft rot bacteria, was found to carry the insertion in locus BV82_4706. This gene was annotated as encoding a HpcH/HpaI aldolase family/citrate lyase family protein. Although, the protein contains a domain known from the β subunit (CitE) of the citrate lyase (source: NCBI's CDD, Marchler-Bauer et al., 2014), its similarity to CitE from K. pneumoniae is low (Bott and Dimroth, 1994; Meyer et al., 1997). Moreover, P482 lacks genes encoding the α and γ subunits of the CitE holoenzyme, suggesting that the product of BV82_4706 plays a different role in P482 than it does in other citrate-fermenting bacteria (Bekal et al., 1998; Martín et al., 2004). Here, we have also established that the inactivation of the neighboring loci: BV82_4705, BV82_4707, as well as the nfs gene homolog BV82_4709, leads to the loss of antibiosis between P482 and the soft rot pathogens.
Interrelationship between Iron Availability, Siderophore Production, and the Antimicrobial Activity of P482
In P. donghuensis HYST, the homolog of the BV82_4709 gene was reported to be essential for the synthesis of an, as yet, chemically-undefined non-fluorescent siderophore, responsible for the potent iron-scavenging properties of this strain (Yu et al., 2014). HYST also produces pyoverdine, however its loss through inactivation of pvdA did not reduce the total siderophore yield of HYST, highlighting the compensatory role of the non-fluorescent siderophore. Assuming that these genes play an analogous role in both P482 and HYST, there is overlap between the P482 antibacterial activity, and the siderophore production, reported for HYST. In P482, contrary to the results obtained for HYST, the inactivation of BV82_4709, which corresponds to the nfs2 ORF of HYST, did not significantly influence total siderophore production by P482. Moreover, two pyoverdine-deficient mutants of the P482, KN1009 (psvA−), and KN3755 (pvdD−), showed a profound decrease (approximately 80%) in siderophore production with respect to the wild type strain. Thus, pyoverdine is the major iron chelator employed by P482 in the assay conditions used, even though it is not the only iron-scavenging compound produced by this strain.
Considering the differences between P482 and HYST, it is worth acknowledging that although both groups used CAS agar to detect siderophores, there were technical differences in the experimental setups used. For example, different incubation temperatures were used and it is known that the temperature alters siderophore production levels (Meyer and Stintzi, 1998).
We also investigated the interrelationship between iron availability and the antimicrobial activity of P482. The medium that we routinely use for growth-inhibition assays (LB agar) is relatively iron-rich (~10–17 μM; Goldberg et al., 1990; Abdul-Tehrani et al., 1999). However, additional supplementation with FeSO4 can be performed for studies requiring unlimited iron growth conditions (Massé et al., 2005; Ouyang and Isaacson, 2006). Our results showed that the wild type P. donghuensis P482 inhibits the growth of tested strains of SRE on an iron-rich medium (LB agar supplemented with additional 15 μM of FeSO4) in a similar manner to that on unsupplemented LB agar. This implies that the observed in vitro antagonism is not based on the competition for iron, and, by extension, on the activity of siderophores. Moreover, the iron-rich conditions do not repress production of antibacterial compound(s) by P482. This is interesting as a FeSO4 concentration of 10 μM repressed the production of the non-fluorescent siderophore by P. donghuensis HYST (Yu et al., 2014).
Preliminary experiments aiming isolation and purification of the biologically active compound(s) produced by P498, with use the chromatographic and spectrometric techniques failed. Thus, further investigation is required to elucidate the chemical nature of the antibacterial activity of P482 because this cannot be deduced by genomic analysis alone. Similar problem with determination of the chemical problem of novel antimicrobial from P. putida W15Oct28 was also described by Ye et al. (2014).
Antibacterial Activity of P. donghuensis against Soft Rot Pathogens: Unique Mechanism of a Not So Uncommon Trait?
The two P. donghuensis strains, P482 and HYST, exhibit a comparable level of antibiosis toward the Dickeya and Pectobacterium spp. Among nine related Pseudomonas spp. tested, three other strains also hindered the growth of the SRE: the P. protegens strains CHA0T and Pf-5, and the P. entomophila L48T. The CHA0T and Pf-5 produce, among others, 2,4-DAPG, a polyketide that inhibits the growth of Pectobacterium atrosepticum (Cronin et al., 1997), Bacillus cereus, Bacillus thuringensis, and Pseudomonas syringae (MIC-value, 5 μg·ml−1), and, to lesser extent, P. carotovorum subsp. carotovorum (MIC-value, 250 μg·ml−1; Keel et al., 1992). To our knowledge, P. entomophila L48T has not been previously tested for antibiosis toward the soft rot bacteria. This strain produces many secondary metabolites, including the cyclic lipopeptide xantholysin, shown to have antibacterial activity against xanthomonads and several Gram-positive bacterial species (Li et al., 2013; Molina-Santiago et al., 2015). For CHA0T, PF-5 and L48T, the spectrum of inhibited soft rot pathogens and/or the size of the growth inhibition zones observed differed from those of P482 and HYST. In general, the antibacterial activity of P. donghuensis was more potent than the other strains.
We established that the genes crucial for the production of antimicrobial(s) by P482 are unique to the two known representatives of P. donghuensis. These genes are absent from the genomes of P. putida PA14H7 and P. fluorescens PA3G8 and PA4C2, three strains previously reported to inhibit the growth of soft rot pathogens (Cigna et al., 2015; Raoul des Essarts et al., 2016). In the latter, in vitro screening of over 10000 potato-associated bacterial isolates yielded ~2.4% of strains with antibacterial activity toward at least one soft rot pathogens. Most of the selected antagonistic isolates were classified as Pseudomonas or Bacillus spp. In earlier work on soft rot antagonists, where we obtained comparable results for rhizobacteria of different herbs and vegetables, approximately 1% of these growth-inhibitors were pseudomonads and bacilli (Krzyzanowska et al., 2012) This provides a rough estimation of how many easily-cultivable bacteria from the sampled environments synthesize, in vitro, compounds affecting the growth of soft rot pathogens. This highlights that among Pseudomonas spp., this trait is not restricted to a specific strain, species, nor compound. However, for P482 and HYST genome analysis suggests that they share unique genes, which confer the production of novel antimicrobials.
Genomic regions exclusive to certain Pseudomonas spp. strains are often associated with horizontal gene transfer, intragenomic rearrangements, or the activity of mobile genetic elements (Loper et al., 2012). These events are generally recognized as important driving forces of genomic diversity (Darmon and Leach, 2014). However, here we have shown that the BV82_4705–47011 gene cluster of P482, as well as the adjacent genomic regions, do not appear to carry the marks of recent gene transfer (Langille et al., 2008; Zhou et al., 2011).
Applicability of Genome Mining for the Discovery of Novel Compounds—The P482 Perspective
The development of high-throughput sequencing technologies, as well as user-friendly bioinformatic tools, considerably facilitates the exploration of microbial secondary metabolomes (Donadio et al., 2010; Loman et al., 2012; Blin et al., 2013). Gene/protein-based searches have been effective in linking previously-studied compounds to new isolates, as well as in the discovery of novel compounds synthesized by well-conserved machineries, such as the NRPS, PKS, or terpene synthases (Weber and Marahiel, 2001; Gross et al., 2007; Loper et al., 2008; Rokni-Zadeh et al., 2011; Cane and Ikeda, 2012; Ye et al., 2014; Aleti et al., 2015; Horn et al., 2015; Song et al., 2015). An exciting development is that genome mining tools are now not only able to identify well-known pathways (compounds), but also to provide probability-based guesses of new chemical entities. An automated search of the P482 genome with ClusterFinder, an antiSMASH-integrated algorithm designed to detect atypical or even novel classes of secondary metabolite gene clusters (Cimermancic et al., 2014), led to the prediction of 18 such hypothetical clusters in P482. In total, the number of genes potentially involved in the synthesis of unknown metabolites was estimated to be 281, suggesting a considerable potential for the discovery of diverse novel antimicrobial(s). The value of these predictions is that they enabled discovery of genes essential for the antimicrobial activity of P482. However, this finding could not have been made without combining with transposon mutagenesis, as the high number of hypothetical candidate genes precluded the use of low throughput experimental methods, such as site-directed mutagenesis. Currently, this may be a general limiting factor for the application of genome mining for the discovery of novel secondary metabolites. Overcoming this limitation will be important for the future structural elucidation of compounds encoded by cryptic clusters not active in vitro and harbored by both culturable and non-culturable microbes.
Conclusions
Pseudomonas sp. P482 is a novel representative of Pseudomonas donghuensis species. The nfs cluster and the three upstream genes play a major role in the antibacterial activity of P482 toward the plant pathogens D. solani and P. carotovorum subsp. brasiliense. Moreover, the observed antibiosis does not depend on competition for bioavailable iron. The results obtained so far suggest that the gene cluster identified is unique to the two P. donghuensis strains P482 and HYST.
Mining of microbial genomes for genes encoding secondary metabolites is a powerful approach. However, when novel compounds are involved, major limitations are apparent because of the shortage of high-throughput experimental tools to verify the accuracy of in silico predictions.
Author Contributions
DK undertook the preliminary research, participated in the design of all experiments, performed genome-mining analyses and ANI calculations, constructed two of the P482 mutants, and wrote the first draft of the manuscript. AO performed the phylogenetic studies, including API biochemical assays, and obtained the P482 transposon mutant. MR performed the siderophore and pyoverdine production assays, contributed to the construction of P482 mutants, prepared growth curves, and critically revised the manuscript. TM constructed three of the mutants reported. MJ performed the antibacterial activity assays. MO performed the searches for promoter and terminator regions and helped to interpret the results. SH contributed to data analysis and revised the manuscript. SJ conceived and co-ordinated the study, helped to plan the experiments and completed the manuscript. All authors approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was funded by the Polish National Science Centre research grant no. DEC-2012/07/B/NZ9/01623. DK was supported by the National Science Centre personal scholarship ETIUDA1 2013/08/T/NZ9/01049 and MR by the MOBI4 Health program funded by the European Union's Seventh Framework Programme for research, technological development, and demonstration under agreement no. 316094. The authors thank Prof. Paul Williams (Nottingham University, UK) for a helpful discussion and English language correction, Dr. J. van der Waals (University of Pretoria, South Africa) for providing the P. carotovorum subsp. brasiliense JJ 56 strain, the IGS Analysis Engine team (University of Maryland School of Medicine, USA) for the opportunity to use Manatee, and Dr. J. Blom (Justus-Liebig-Universität Gießen, Germany) for access to EDGAR.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00782
Footnotes
2. ^http://www.imedea.uib.es/jspecies.
3. ^https://www.ncbi.nlm.nih.gov/genbank/.
4. ^http://manatee.sourceforge.net.
5. ^http://www.secondarymetabolites.org/.
6. ^http://www.ncbi.nlm.nih.gov/.
7. ^http://www.genome.jp/kegg/.
8. ^http://edgar.computational.bio.uni-giessen.de.
9. ^http://www.phisite.org/main/index.php?nav=toolsandnav_sel=hunter.
10. ^http://www.fruitfly.org/seq_tools/promoter.html.
11. ^http://linux1.softberry.com/berry.phtml?topic=bpromandgroup=programsandsubgroup=gfindb.
12. ^http://www.phisite.org/main/index.php?nav=browse.
13. ^http://rna.igmors.u-psud.fr/toolbox/arnold/.
14. ^http://phast.wishartlab.com/index.html.
15. ^http://www.pathogenomics.sfu.ca/islandviewer/.
References
Abdul-Tehrani, H., Hudson, A. J., Chang, Y. S., Timms, A. R., Hawkins, C., Williams, J. M., et al. (1999). Ferritin mutants of Escherichia coli are iron deficient and growth impaired, and fur mutants are iron deficient. J. Bacteriol. 181, 1415–1428.
Adesemoye, A. O., and Kloepper, J. W. (2009). Plant-microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 85, 1–12. doi: 10.1007/s00253-009-2196-0
Aleti, G., Sessitsch, A., and Brader, G. (2015). Genome mining: prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Comput. Struct. Biotechnol. J. 13, 192–203. doi: 10.1016/j.csbj.2015.03.003
Alexeyev, M. F. (1999). The pknock series of broad-host-range mobilizable suicide vectors for gene knockout and targeted DNA insertion into the chromosome of gram-negative bacteria. BioTechniques 26, 824–827.
Altschul, S. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Altschul, S. F., Wootton, J. C., Gertz, E. M., Agarwala, R., Morgulis, A., Schäffer, A. A., et al. (2005). Protein database searches using compositionally adjusted substitution matrices. FEBS J. 272, 5101–5109. doi: 10.1111/j.1742-4658.2005.04945.x
Ark, P. A., and Tompkins, C. M. (1946). Bacterial leaf blight of bird's-nest fern. Phytopathology 36, 758–761.
Bagdasarian, M., Lurz, R., Rückert, B., Franklin, F. C., Bagdasarian, M. M., Frey, J., et al. (1981). Specific-purpose plasmid cloning vectors. II. Broad host range, high copy number, RFS 1010-derived vectors, and a host-vector system for gene cloning in Pseudomonas. Gene 16, 237–247. doi: 10.1016/0378-1119(81)90080-9
Baltrus, D. A., Nishimura, M. T., Romanchuk, A., Chang, J. H., Mukhtar, M. S., Cherkis, K., et al. (2011). Dynamic evolution of pathogenicity revealed by sequencing and comparative genomics of 19 Pseudomonas syringae isolates. PLoS Pathog. 7:e1002132. doi: 10.1371/journal.ppat.1002132
Bekal, S., van Beeumen, J., Samyn, B., Garmyn, D., Henini, S., Diviès, C., et al. (1998). Purification of Leuconostoc mesenteroides citrate lyase and cloning and characterization of the citCDEFG gene cluster. J. Bacteriol. 180, 647–654.
Blin, K., Medema, M. H., Kazempour, D., Fischbach, M. A., Breitling, R., Takano, E., et al. (2013). Antismash 2.0–A versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 41, W204–W212. doi: 10.1093/nar/gkt449
Blom, J., Albaum, S. P., Doppmeier, D., Pühler, A., Vorhölter, F.-J., Zakrzewski, M., et al. (2009). Edgar: a software framework for the comparative analysis of prokaryotic genomes. BMC Bioinformatics 10, 154–154. doi: 10.1186/1471-2105-10-154
Blumer, C., and Haas, D. (2000). Mechanism, regulation, and ecological role of bacterial cyanide biosynthesis. Arch. Microbiol. 173, 170–177. doi: 10.1007/s002039900127
Bott, M., and Dimroth, P. (1994). Klebsiella pneumoniae genes for citrate lyase and citrate lyase ligase: localization, sequencing, and expression. Mol. Microbiol. 14, 347–356. doi: 10.1111/j.1365-2958.1994.tb01295.x
Boyd, A., and Chakrabarty, A. M. (1995). Pseudomonas aeruginosa biofilms: role of the alginate exopolysaccharide. J. Ind. Microbiol. 15, 162–168. doi: 10.1007/BF01569821
Busquets, A., Peña, A., Gomila, M., Bosch, R., Nogales, B., García-Valdés, E., et al. (2012). Genome sequence of Pseudomonas stutzeri strain JM300 (DSM 10701), a soil isolate and model organism for natural transformation. J. Bacteriol. 194, 5477–5478. doi: 10.1128/JB.01257-12
Cane, D. E., and Ikeda, H. (2012). Exploration and mining of the bacterial terpenome. Accounts Chem. Res. 45, 463–472. doi: 10.1021/ar200198d
Cigna, J., Raoul Des Essarts, Y., Mondy, S., Hélias, V., Beury-Cirou, A., and Faure, D. (2015). Draft genome sequences of Pseudomonas fluorescens strains PA4C2 and PA3G8 and Pseudomonas putida PA147, three biocontrol bacteria against Dickeya phytopathogens. Genome Announc. 3, e01503-14. doi: 10.1128/genomeA.01503-14
Cimermancic, P., Medema, M. H., Claesen, J., Kurita, K., Wieland Brown, L. C., Mavrommatis, K., et al. (2014). Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 158, 412–421. doi: 10.1016/j.cell.2014.06.034
Cornelis, P., and Matthijs, S. (2002). Diversity of siderophore-mediated iron uptake systems in fluorescent pseudomonads: not only pyoverdines. Environ. Microbiol. 4, 787–798. doi: 10.1046/j.1462-2920.2002.00369.x
Cronin, D., Moënne-Loccoz, Y., Fenton, A., Dunne, C., Dowling, D. N., and O'gara, F. (1997). Ecological interaction of a biocontrol Pseudomonas fluorescens strain producing 2,4-diacetylphloroglucinol with the soft rot potato pathogen Erwinia carotovora subsp. atroseptica. FEMS Microbiol. Ecol. 23, 95–106. doi: 10.1111/j.1574-6941.1997.tb00394.x
Czajkowski, R., Perombelon, M. C. M., van Veen, J. A., and van der Wolf, J. M. (2011). Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: a review. Plant Pathol. 60, 999–1013. doi: 10.1111/j.1365-3059.2011.02470.x
Darmon, E., and Leach, D. R. F. (2014). Bacterial genome instability. Microbiol. Mol. Biol. Rev. 78, 1–39. doi: 10.1128/MMBR.00035-13
Dhillon, B. K., Laird, M. R., Shay, J. A., Winsor, G. L., Lo, R., Nizam, F., et al. (2015). Islandviewer 3: more flexible, interactive genomic island discovery, visualization and analysis. Nucleic Acids Res. 43, W104–W108. doi: 10.1093/nar/gkv401
Donadio, S., Maffioli, S., Monciardini, P., Sosio, M., and Jabes, D. (2010). Antibiotic discovery in the twenty-first century: current trends and future perspectives. J. Antibiot. 63, 423–430. doi: 10.1038/ja.2010.62
Duarte, V., de Boer, S. H., Ward, L. J., and De Oliveira, A. M. R. (2004). Characterization of atypical Erwinia carotovora strains causing blackleg of potato in brazil. J. Appl. Microbiol. 96, 535–545. doi: 10.1111/j.1365-2672.2004.02173.x
Elomari, M., Coroler, L., Verhille, S., Izard, D., and Leclerc, H. (1997). Pseudomonas monteilii sp. nov., isolated from clinical specimens. Int. J. Syst. Bacteriol. 47, 846–852. doi: 10.1099/00207713-47-3-846
Gao, J., Xie, G., Peng, F., and Xie, Z. (2015). Pseudomonas donghuensis sp. nov., exhibiting high-yields of siderophore. Antonie Van Leeuwenhoek 107, 83–94. doi: 10.1007/s10482-014-0306-1
Gao, J., Yu, X., and Xie, Z. (2012). Draft genome sequence of high-siderophore-yielding Pseudomonas sp. strain HYS. J. Bacteriol. 194, 4121–4121. doi: 10.1128/jb.00688-12
Golanowska, M., Ankiewicz, H., Taraszkiewicz, A., Kamysz, W., Czajkowski, R., Krolicka, A., et al. (2012). Combined effect of the antagonistic potential of selected Pseudomonas spp. strains and the synthetic peptide “CAMEL” on Pseudomonas syringae pv. syringae and P. syringae pv. morsprunorum. J. Plant. Pathol. 94, S1.69–S61.73. doi: 10.4454/jpp.v94i1sup.012
Goldberg, M. B., Dirita, V. J., and Calderwood, S. B. (1990). Identification of an iron-regulated virulence determinant in Vibrio cholerae, using tnphoa mutagenesis. Infect. Immun. 58, 55–60.
Goldfarb, K. C., Karaoz, U., Hanson, C. A., Santee, C. A., Bradford, M. A., Treseder, K. K., et al. (2011). Differential growth responses of soil bacterial taxa to carbon substrates of varying chemical recalcitrance. Front. Microbiol. 2:94. doi: 10.3389/fmicb.2011.00094
Gomila, M., Peña, A., Mulet, M., Lalucat, J., and García-Valdés, E. (2015). Phylogenomics and systematics in Pseudomonas. Front. Microbiol. 6:214. doi: 10.3389/fmicb.2015.00214
Goris, J., Konstantinidis, K. T., Klappenbach, J. A., Coenye, T., Vandamme, P., and Tiedje, J. M. (2007). DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 57, 81–91. doi: 10.1099/ijs.0.64483-0
Gross, H., and Loper, J. E. (2009). Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 26, 1408–1446. doi: 10.1039/b817075b
Gross, H., Stockwell, V. O., Henkels, M. D., Nowak-Thompson, B., Loper, J. E., and Gerwick, W. H. (2007). The genomisotopic approach: a systematic method to isolate products of orphan biosynthetic gene clusters. Chem. Biol. 14, 53–63. doi: 10.1016/j.chembiol.2006.11.007
Haas, D., and Defago, G. (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3, 307–319. doi: 10.1038/nrmicro1129
Harrison, L. A., Letendre, L., Kovacevich, P., Pierson, E., and Weller, D. (1993). Purification of an antibiotic effective against Gaeumannomyces graminis var. tritici produced by a biocontrol agent, Pseudomonas aureofaciens. Soil Biol. Biochem. 25, 215–221. doi: 10.1016/0038-0717(93)90029-B
Holloway, B. W. (1955). Genetic recombination in Pseudomonas aeruginosa. J. Gen. Microbiol. 13, 572–581. doi: 10.1099/00221287-13-3-572
Horn, H., Hentschel, U., and Abdelmohsen, U. R. (2015). Mining genomes of three marine sponge-associated actinobacterial isolates for secondary metabolism. Genome Announc. 3:e01106-15. doi: 10.1128/genomeA.01106-15
Howell, C. R., and Stipanovic, R. D. (1979). Control of Rhizoctonia solani on cotton seedlings with Pseudomonas fluorescens and with an antibiotic produced by the bacterium. Phytopathology 69, 480–482. doi: 10.1094/Phyto-69-480
Kanehisa, M., and Goto, S. (2000). Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. doi: 10.1093/nar/28.1.27
Keel, C., Schnider, U., Maurhofer, M., Voisard, C., Laville, J., Burger, U., et al. (1992). Suppression of root diseases by Pseudomonas fluorescens CHA0: importance of the bacterial secondary metabolite 2,4-diacetylphloroglucinol. Mol. Plant Microbe Interact. 5, 4–13. doi: 10.1094/MPMI-5-004
Klucar, L., Stano, M., and Hajduk, M. (2010). phiSITE: database of gene regulation in bacteriophages. Nucleic Acids Res. 38, D366–D370. doi: 10.1093/nar/gkp911
Konstantinidis, K. T., and Tiedje, J. M. (2005). Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. U.S.A. 102, 2567–2572. doi: 10.1073/pnas.0409727102
Krzyzanowska, D. M., Ossowicki, A., and Jafra, S. (2014). Genome sequence of Pseudomonas sp. strain P482, a tomato rhizosphere isolate with broad-spectrum antimicrobial activity. Genome Announc. 2:e00394-14. doi: 10.1128/genomea.00394-14
Krzyzanowska, D., Potrykus, M., Golanowska, M., Polonis, K., Gwizdek-Wisniewska, A., Lojkowska, E., et al. (2012). Rhizosphere bacteria as potential biocontrol agents against soft rot caused by various Pectobacterium and Dickeya spp. strains. J. Plant Pathol. 94, 367–378. doi: 10.4454/JPP.FA.2012.042
Kümmerli, R., and Brown, S. P. (2010). Molecular and regulatory properties of a public good shape the evolution of cooperation. Proc. Natl. Acad. Sci. U.S.A. 107, 18921–18926. doi: 10.1073/pnas.1011154107
Lambert, A., Fontaine, J.-F., Legendre, M., Leclerc, F., Permal, E., Major, F., et al. (2004). The ERPIN server: an interface to profile-based rna motif identification. Nucleic Acids Res. 32, W160–W165. doi: 10.1093/nar/gkh418
Langille, M. G. I., Hsiao, W. W. L., and Brinkman, F. S. L. (2008). Evaluation of genomic island predictors using a comparative genomics approach. BMC Bioinformatics 9, 329–329. doi: 10.1186/1471-2105-9-329
Larsen, R. A., Wilson, M. M., Guss, A. M., and Metcalf, W. W. (2002). Genetic analysis of pigment biosynthesis in Xanthobacter autotrophicus Py2 using a new, highly efficient transposon mutagenesis system that is functional in a wide variety of bacteria. Arch. Microbiol. 178, 193–201. doi: 10.1007/s00203-002-0442-2
Laurila, J., Hannukkala, A., Nykyri, J., Pasanen, M., Hélias, V., Garlant, L., et al. (2010). Symptoms and yield reduction caused by Dickeya spp. strains isolated from potato and river water in Finland. Eur. J. Plant. Pathol. 126, 249–262. doi: 10.1007/s10658-009-9537-9
Leite, L. N., De Haan, E., Krijger, M., Kastelein, P., van Der Zouwen, P., van Den Bovenkamp, G., et al. (2014). First report of potato blackleg caused by Pectobacterium carotovorum subsp. brasiliensis in the Netherlands. New Dis. Rep. 29:24. doi: 10.5197/j.2044-0588.2014.029.024
Li, W., Rokni-Zadeh, H., De Vleeschouwer, M., Ghequire, M. G. K., Sinnaeve, D., Xie, G.-L., et al. (2013). The antimicrobial compound xantholysin defines a new group of Pseudomonas cyclic lipopeptides. PLoS ONE 8:e62946. doi: 10.1371/journal.pone.0062946
Loman, N. J., Constantinidou, C., Chan, J. Z. M., Halachev, M., Sergeant, M., Penn, C. W., et al. (2012). High-throughput bacterial genome sequencing: an embarrassment of choice, a world of opportunity. Nat. Rev. Microbiol. 10, 599–606. doi: 10.1038/nrmicro2850
Loper, J. E., Hassan, K. A., Mavrodi, D. V., Davis, E. W., Lim, C. K., Shaffer, B. T., et al. (2012). Comparative genomics of plant-associated Pseudomonas spp.: insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet. 8:e1002784. doi: 10.1371/journal.pgen.1002784
Loper, J. E., Henkels, M. D., Shaffer, B. T., Valeriote, F. A., and Gross, H. (2008). Isolation and identification of rhizoxin analogs from Pseudomonas fluorescens Pf-5 by using a genomic mining strategy. Appl. Environ. Microbiol. 74, 3085–3093. doi: 10.1128/AEM.02848-07
Ma, B., Hibbing, M. E., Kim, H. S., Reedy, R. M., Yedidia, I., Breuer, J., et al. (2007). Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya. Phytopathology 97, 1150–1163. doi: 10.1094/PHYTO-97-9-1150
Marchler-Bauer, A., Derbyshire, M. K., Gonzales, N. R., Lu, S., Chitsaz, F., Geer, L. Y., et al. (2014). Cdd: NCBI's conserved domain database. Nucleic Acids Res. 43, D222–D226. doi: 10.1093/nar/gku1221
Martín, M. G., Sender, P. D., Peirú, S., De Mendoza, D., and Magni, C. (2004). Acid-inducible transcription of the operon encoding the citrate lyase complex of Lactococcus lactis biovar diacetylactis CRL264. J. Bacteriol. 186, 5649–5660. doi: 10.1128/JB.186.17.5649-5660.2004
Massé, E., vanderpool, C. K., and Gottesman, S. (2005). Effect of RyhB small RNA on global iron use in Escherichia coli. J. Bacteriol. 187, 6962–6971. doi: 10.1128/JB.187.20.6962-6971.2005
Medema, M. H., Blin, K., Cimermancic, P., De Jager, V., Zakrzewski, P., Fischbach, M. A., et al. (2011). AntiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 39, W339–W346. doi: 10.1093/nar/gkr466
Mercado-Blanco, J. (2015). “Pseudomonas strains that exert biocontrol of plant pathogens,” in Pseudomonas: Volume 7: New Aspects of Pseudomonas Biology, ed J.-L. Ramos (Dordrecht: Springer Netherlands), 121–172. doi: 10.1007/978-94-017-9555-5_6
Meyer, J.-M., and Stintzi, A. (1998). “Iron metabolism and siderophores in Pseudomonas and related species,” in Pseudomonas, ed T. C. Montie (New York, NY: Springer US), 201–243. doi: 10.1007/978-1-4899-0120-0_7
Meyer, M., Dimroth, P., and Bott, M. (1997). In vitro binding of the response regulator citb and of its carboxy-terminal domain to A + T-rich DNA target sequences in the control region of the divergent citc and cits operons of Klebsiella pneumoniae. J. Mol. Biol. 269, 719–731. doi: 10.1006/jmbi.1997.1076
Molina-Santiago, C., Udaondo, Z., Daddaoua, A., Roca, A., Martín, J., Pérez-Victoria, I., et al. (2015). Efflux pump-deficient mutants as a platform to search for microbes that produce antibiotics. Microb. Biotechnol. 8, 716–725. doi: 10.1111/1751-7915.12295
Mulet, M., Gomila, M., Lemaitre, B., Lalucat, J., and García-Valdés, E. (2012). Taxonomic characterisation of Pseudomonas strain L48 and formal proposal of Pseudomonas entomophila sp. nov. Syst. Appl. Microbiol. 35, 145–149. doi: 10.1016/j.syapm.2011.12.003
Nabhan, S., De Boer, S. H., Maiss, E., and Wydra, K. (2012). Taxonomic relatedness between Pectobacterium carotovorum subsp. carotovorum, Pectobacterium carotovorum subsp. odoriferum and Pectobacterium carotovorum subsp. brasiliense subsp. nov. J. Appl. Microbiol. 113, 904–913. doi: 10.1111/j.1365-2672.2012.05383.x
Newman, D. J., and Cragg, G. M. (2012). Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 75, 311–335. doi: 10.1021/np200906s
Novik, G., Savich, V., and Kiseleva, E. (2015). “An insight into beneficial Pseudomonas bacteria,” in Microbiology in Agriculture and Human Health, ed M. M. Shah (InTech), 73–105.
Ouyang, Z., and Isaacson, R. (2006). Identification and characterization of a novel ABC iron transport system, Fit, in Escherichia coli. Infect. Immun. 74, 6949–6956. doi: 10.1128/IAI.00866-06
Palleroni, N. J. (2005). “Geuns I. Pseudomonas migula 1894, 237AL (Nom. Cos,. Opin. 5 of the jud. Comm. 1952, 121),” in Bergey's Manual of Systematic Bacteriology, vol. 2B, 2nd Edn., eds D. J. Brenner, R. W. Castenholz, G. M. Garrity, N. R. Krieg and J. T. Staley (New York, NY: Springer), 323–379.
Paulsen, I. T., Press, C. M., Ravel, J., Kobayashi, D. Y., Myers, G. S. A., Mavrodi, D. V., et al. (2005). Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf-5. Nat. Biotechnol. 23, 873–878. doi: 10.1038/nbt1110
Pierson, L. S., and Pierson, E. A. (2010). Metabolism and function of phenazines in bacteria: impacts on the behavior of bacteria in the environment and biotechnological processes. Appl. Microbiol. Biotechnol. 86, 1659–1670. doi: 10.1007/s00253-010-2509-3
Pliego, C., Kamilova, F., and Lugtenberg, B. (2011). “Plant growth-promoting bacteria: fundamentals and exploitation,” in Bacteria in Agrobiology: Crop Ecosystems, ed D. K. Maheshwari (Berlin; Heidelberg: Springer-Verlag), 61–96. doi: 10.1007/978-3-642-18357-7_11
Potvin, E., Sanschagrin, F., and Levesque, R. C. (2008). Sigma factors in Pseudomonas aeruginosa. FEMS Microbiol. Rev. 32, 38–55. doi: 10.1111/j.1574-6976.2007.00092.x
Raaijmakers, J. M., De Bruijn, I., and de Kock, M. J. D. (2006). Cyclic lipopeptide production by plant-associated Pseudomonas spp.: diversity, activity, biosynthesis, and regulation. Mol. Plant Microbe Interact. 19, 699–710. doi: 10.1094/MPMI-19-0699
Raaijmakers, J. M., de Bruijn, I., Nybroe, O., and Ongena, M. (2010). Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol. Rev. 34, 1037–1062. doi: 10.1111/j.1574-6976.2010.00221.x
Raaijmakers, J. M., and Mazzola, M. (2012). Diversity and natural functions of antibiotics produced by beneficial and plant pathogenic bacteria. Annu. Rev. Phytopathol. 50, 403–424. doi: 10.1146/annurev-phyto-081211-172908
Raaijmakers, J. M., and Weller, D. M. (1998). Natural plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. In take-all decline soils. Mol. Plant Microbe Interact. 11, 144–152. doi: 10.1094/MPMI.1998.11.2.144
Raes, J., Korbel, J. O., Lercher, M. J., von Mering, C., and Bork, P. (2007). Prediction of effective genome size in metagenomic samples. Genome Biol. 8:R10. doi: 10.1186/gb-2007-8-1-r10
Ramette, A., Frapolli, M., Fischer-Le Saux, M., Gruffaz, C., Meyer, J.-M., Défago, G., et al. (2011). Pseudomonas protegens sp. nov., widespread plant-protecting bacteria producing the biocontrol compounds 2,4-diacetylphloroglucinol and pyoluteorin. Syst. Appl. Microbiol. 34, 180–188. doi: 10.1016/j.syapm.2010.10.005
Raoul des Essarts, Y., Cigna, J., Quêtu-Laurent, A., Caron, A., Munier, E., Beury-Cirou, A., et al. (2016). Biocontrol of the potato blackleg and soft-rot diseases caused by Dickeya dianthicola. Appl. Environ. Microbiol. 82, 268–278. doi: 10.1128/AEM.02525-15
Reese, M. G. (2001). Application of a time-delay neural network to promoter annotation in the Drosophila melanogaster genome. Comput. Chem. 26, 51–56. doi: 10.1016/S0097-8485(01)00099-7
Richter, M., and Rosselló-Móra, R. (2009). Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. U.S.A. 106, 19126–19131. doi: 10.1073/pnas.0906412106
Rokni-Zadeh, H., Mangas-Losada, A., and de Mot, R. (2011). PCR detection of novel non-ribosomal peptide synthetase genes in lipopeptide-producing Pseudomonas. Microb. Ecol. 62, 941–947. doi: 10.1007/s00248-011-9885-9
Sadikot, R. T., Blackwell, T. S., Christman, J. W., and Prince, A. S. (2005). Pathogen-host interactions in Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 171, 1209–1223. doi: 10.1164/rccm.200408-1044SO
Schulz-Bohm, K., Zweers, H., de Boer, W., and Garbeva, P. (2015). A fragrant neighborhood: volatile mediated bacterial interactions in soil. Front. Microbiol. 6:1212. doi: 10.3389/fmicb.2015.01212
Schwyn, B., and Neilands, J. B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 46–56. doi: 10.1016/0003-2697(87)90612-9
Sievers, F., Wilm, A., Dineen, D., Gibson, T. J., Karplus, K., Li, W., et al. (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using clustal omega. Mol. Syst. Biol. 7, 539–539. doi: 10.1038/msb.2011.75
Silby, M. W., Winstanley, C., Godfrey, S. A., Levy, S. B., and Jackson, R. W. (2011). Pseudomonas genomes: diverse and adaptable. FEMS Microbiol. Rev. 35, 652–680. doi: 10.1111/j.1574-6976.2011.00269.x
Sławiak, M., Łojkowska, E., and van der Wolf, J. M. (2009). First report of bacterial soft rot on potato caused by Dickeya sp. (syn. Erwinia chrysanthemi) in Poland. Plant Pathol. 58, 794–794. doi: 10.1111/j.1365-3059.2009.02028.x
Solovyev, V., and Salamov, A. (2011). “Automatic annotation of microbial genomes and metagenomic sequences,” in Metagenomics and its Applications in Agriculture, Biomedicine and Environmental Studies, ed R. W. Li (Nova Science Publishers), 61–78.
Song, C., Schmidt, R., De Jager, V., Krzyzanowska, D., Jongedijk, E., Cankar, K., et al. (2015). Exploring the genomic traits of fungus-feeding bacterial genus Collimonas. BMC Genomics 16:1103. doi: 10.1186/s12864-015-2289-3
Stutz, E. W. (1986). Naturally occurring fluorescent pseudomonads involved in suppression of black root rot of tobacco. Phytopathology 76, 181–181. doi: 10.1094/Phyto-76-181
Takeuchi, K., Noda, N., and Someya, N. (2014). Complete genome sequence of the biocontrol strain Pseudomonas protegens Cab57 discovered in japan reveals strain-specific diversity of this species. PLoS ONE 9, e93683. doi: 10.1371/journal.pone.0093683
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). Mega6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Thoma, S., and Schobert, M. (2009). An improved Escherichia coli donor strain for diparental mating. FEMS Microbiol. Lett. 294, 127–132. doi: 10.1111/j.1574-6968.2009.01556.x
Toth, I. K., van der Wolf, J. M., Saddler, G., Lojkowska, E., Hélias, V., Pirhonen, M., et al. (2011). Dickeya species: an emerging problem for potato production in Europe. Plant Pathol. 60, 385–399. doi: 10.1111/j.1365-3059.2011.02427.x
Tsror, L., Erlich, O., Lebiush, S., Hazanovsky, M., Zig, U., Sławiak, M., et al. (2009). Assessment of recent outbreaks of Dickeya sp. (syn. Erwinia chrysanthemi) slow wilt in potato crops in Israel. Eur. J. Plant Pathol. 123, 311–320. doi: 10.1007/s10658-008-9368-0
Tvrzová, L., Schumann, P., Spröer, C., Sedlácek, I., Pácová, Z., Sedo, O., et al. (2006). Pseudomonas moraviensis sp. nov. and Pseudomonas vranovensis sp. nov., soil bacteria isolated on nitroaromatic compounds, and emended description of Pseudomonas asplenii. Int. J. Syst. Evol. Microbiol. 56, 2657–2663. doi: 10.1099/ijs.0.63988-0
Uchino, M., Shida, O., Uchimura, T., and Komagata, K. (2001). Recharacterization of Pseudomonas fulva Iizuka and Komagata 1963, and proposals of Pseudomonas parafulva sp. nov. and Pseudomonas cremoricolorata sp. nov. J. Gen. Appl. Microbiol. 47, 247–261. doi: 10.2323/jgam.47.247
van der Merwe, J. J., Coutinho, T. A., Korsten, L., and van der Waals, J. E. (2009). Pectobacterium carotovorum subsp. brasiliensis causing blackleg on potatoes in South Africa. Eur. J. Plant. Pathol. 126, 175–185. doi: 10.1007/s10658-009-9531-2
van der Wolf, J. M., Nijhuis, E. H., Kowalewska, M. J., Saddler, G. S., Parkinson, N., Elphinstone, J. G., et al. (2014). Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 64, 768–774. doi: 10.1099/ijs.0.052944-0
Visca, P., Imperi, F., and Lamont, I. L. (2007). Pyoverdine siderophores: from biogenesis to biosignificance. Trends Microbiol. 15, 22–30. doi: 10.1016/j.tim.2006.11.004
Vodovar, N., Vinals, M., Liehl, P., Basset, A., Degrouard, J., Spellman, P., et al. (2005). Drosophila host defense after oral infection by an entomopathogenic Pseudomonas species. Proc. Natl. Acad. Sci. U.S.A. 102, 11414–11419. doi: 10.1073/pnas.0502240102
Weber, T., and Marahiel, M. A. (2001). Exploring the domain structure of modular nonribosomal peptide synthetases. Structure 9, R3–R9. doi: 10.1016/S0969-2126(00)00560-8
Weller, D. M. (2007). Pseudomonas biocontrol agents of soilborne pathogens: looking back over 30 years. Phytopathology 97, 250–256. doi: 10.1094/PHYTO-97-2-0250
Wu, X., Monchy, S., Taghavi, S., Zhu, W., Ramos, J., and van der Lelie, D. (2011). Comparative genomics and functional analysis of niche-specific adaptation in Pseudomonas putida. FEMS Microbiol. Rev. 35, 299–323. doi: 10.1111/j.1574-6976.2010.00249.x
Ye, L., Hildebrand, F., Dingemans, J., Ballet, S., Laus, G., Matthijs, S., et al. (2014). Draft genome sequence analysis of a Pseudomonas putida W15Oct28 strain with antagonistic activity to Gram-positive and Pseudomonas sp. pathogens. PLoS ONE 9:e110038. doi: 10.1371/journal.pone.0110038
Young, J. M. (2010). Taxonomy of Pseudomonas syringae. J. Plant. Pathol. 92, S95–S14. doi: 10.4454/jpp.v92i1sup.2501
Yu, X., Chen, M., Jiang, Z., Hu, Y., and Xie, Z. (2014). The two-component regulators GacS and GacA positively regulate a nonfluorescent siderophore through the gac/rsm signaling cascade in high-siderophore-yielding Pseudomonas sp. strain HYS. J. Bacteriol. 196, 3259–3270. doi: 10.1128/jb.01756-14
Keywords: Dickeya, Pectobacterium, secondary metabolites, genome mining, antiSMASH, nfs
Citation: Krzyżanowska DM, Ossowicki A, Rajewska M, Maciąg T, Jabłońska M, Obuchowski M, Heeb S and Jafra S (2016) When Genome-Based Approach Meets the “Old but Good”: Revealing Genes Involved in the Antibacterial Activity of Pseudomonas sp. P482 against Soft Rot Pathogens. Front. Microbiol. 7:782. doi: 10.3389/fmicb.2016.00782
Received: 02 March 2016; Accepted: 09 May 2016;
Published: 26 May 2016.
Edited by:
Eric Altermann, AgResearch Ltd., New ZealandReviewed by:
Pierre Cornelis, Vrije Universiteit Brussel, BelgiumManuel Espinosa-Urgel, Estación Experimental del Zaidín, Consejo Superior de Investigaciones Científicas, Spain
Copyright © 2016 Krzyżanowska, Ossowicki, Rajewska, Maciąg, Jabłońska, Obuchowski, Heeb and Jafra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sylwia Jafra, sylwia.jafra@biotech.ug.edu.pl