 Christina Westermann1†
Christina Westermann1† Sinéad C. Corr
Sinéad C. Corr Christian U. Riedel
Christian U. Riedel- 1Institute of Microbiology and Biotechnology, University of Ulm, Ulm, Germany
- 2Department of Microbiology, Moyne Institute of Preventative Medicine, School of Genetics and Microbiology, Trinity College Dublin, Dublin, Ireland
Bifidobacteria are common inhabitants of the human gastrointestinal tract that, despite a long history of research, have not shown any pathogenic potential whatsoever. By contrast, some bifidobacteria are associated with a number of health-related benefits for the host. The reported beneficial effects of bifidobacteria include competitive exclusion of pathogens, alleviation of symptoms of irritable bowel syndrome and inflammatory bowel disease, and modulation of intestinal and systemic immune responses. Based on these effects, bifidobacteria are widely used as probiotics by pharmaceutical and dairy industries. In order to exert a beneficial effect bifidobacteria have to, at least transiently, colonize the host in a sufficient population size. Besides other criteria such as resistance to manufacturing processes and intestinal transit, potential probiotic bacteria are tested for adhesion to the host structures including intestinal epithelial cells, mucus, and extracellular matrix components. In the present review article, we summarize the current knowledge on bifidobacterial structures that mediate adhesion to host tissue and compare these to similar structures of pathogenic bacteria. This reveals that most of the adhesive structures and mechanisms involved in adhesion of bifidobacteria to host tissue are similar or even identical to those employed by pathogens to cause disease. It is thus reasonable to assume that these structures and mechanisms are equally important for commensal or probiotic bacteria and play a similar role in the beneficial effects exerted by bifidobacteria.
Introduction
The mammalian GIT is home to an extremely complex and diverse microbial ecosystem consisting primarily of prokaryotes. This microbial community is collectively referred to as the gut microbiota and exerts a number of profound effects on host health (Marchesi et al., 2016). During the first period of life when newborns are exclusively breast-fed, members of the genus Bifidobacterium are one of the predominant bacterial groups of the microbiota in the lower GIT (Yatsunenko et al., 2012; Bäckhed et al., 2015; Walker et al., 2015). The major source for bifidobacteria is the intestinal microbiota of the mother to which the newborn is exposed during (vaginal) delivery (Grönlund et al., 2011; Matamoros et al., 2013). More recently, breast milk has also been shown to contain viable bifidobacteria (Fernández et al., 2013; Jost et al., 2015). Although their relative proportion decreases over time, bifidobacteria are still a subdominant group amongst intestinal bacteria of adult humans (Arumugam et al., 2011).
With the exception of Bifidobacterium dentium, which has been associated with dental caries (Ventura et al., 2009), bifidobacteria have to date not shown any pathogenic potential. By contrast, a number of health promoting effects have been attributed to the presence of bifidobacteria in the GIT including improvement of symptoms of irritable bowel syndrome, inflammatory bowel disease and infectious diarrhea, modulation of intestinal and systemic immune responses, and resistance against colonization by pathogens (Gareau et al., 2010; Buffie and Pamer, 2013). Of note, a very recent study links bifidobacteria in the gut microbiota to enhanced anti-tumor immune responses and support of checkpoint-inhibition cancer therapy using a monoclonal antibody (Sivan et al., 2015). Based on these findings bifidobacteria are widely used as probiotics, i.e., live microorganisms which when administered in adequate amounts confer a health benefit to the host (Holmes et al., 2012; Foligné et al., 2013).
Besides the health promoting effects, several criteria are applied during selection of a suitable probiotic candidate strain including stability during manufacturing processes, viability during gastrointestinal transit and functionality at the desired target site (Foligné et al., 2013). One of the classical selection criteria for potential probiotic bacteria is adhesion to mucus and/or IECs (Klaenhammer and Kullen, 1999; Tuomola et al., 2001; Papadimitriou et al., 2015).
It may be argued that adhesion is not important for probiotic functionality since probiotic bacteria do not have access to host tissue due to the thick mucus layer covering the (healthy) gut epithelium. However, a number of bifidobacteria were shown to adhere to mucus (He et al., 2001; Izquierdo et al., 2008) and utilize host-derived mucins as a substrate for growth (Tailford et al., 2015). Also, bifidobacteria are discussed as potential treatment options for conditions with an impaired mucus layer (Whelan and Quigley, 2013; Johansson, 2014) facilitating direct access of (bifido)bacteria to the epithelium. Moreover, various bacterial pathogens must overcome the mucosal barriers and gain access to the epithelial layer to cause disease. For example, pathogenic Escherichia coli strains and related organisms use pili, fimbriae, and/or intimin with its translocated intimin receptor for adhesion to epithelial cells (Niemann et al., 2004). Another example is the interaction of InlA of Listeria monocytogenes with E-cadherin on host epithelial cells which is crucially required for infection (Stavru et al., 2011). Only once adhesion of these pathogens to the epithelium has been achieved despite the presence of an intact mucus layer, progression to later stages of infection and disease are possible (Bhavsar et al., 2007).
On the other hand, a number of probiotic traits may be directly linked to adhesion to host structures. One of the proposed health benefits of bifidobacteria is resistance against colonization or infection by pathogens. This may involve a variety of adhesion-independent mechanisms such as competition for nutrients or production of antimicrobial compounds (Buffie and Pamer, 2013; Lawley and Walker, 2013). Nevertheless, adhesion to IECs, mucus and ECM components by commensal and probiotic bacteria may also directly block access of pathogens to these structures (Bernet et al., 1993; Collado et al., 2005; Candela et al., 2008; Serafini et al., 2013) either by competition for attachment sites or steric hinderance. Also, there are numerous reports of immunomodulatory effects of bifidobacteria in vitro and in animal models (Bermudez-Brito et al., 2012). All these effects crucially depend on interaction with (and thus adhesion to) epithelial cells, dendritic cells, monocytes, macrophages and or other immune cells. Finally, even if not directly implicated mechanistically, adhesion might contribute to beneficial effects by allowing initial colonization or prolonging persistence of (probiotic) bifidobacteria in the GIT.
Factors for Adhesion of Bifidobacteria to Host Structures
A number of factors and structures involved in adhesion to IECs, ECM components, and mucus have been identified in bifidobacteria (Table 1). These studies have been performed almost exclusively in in vitro model systems.
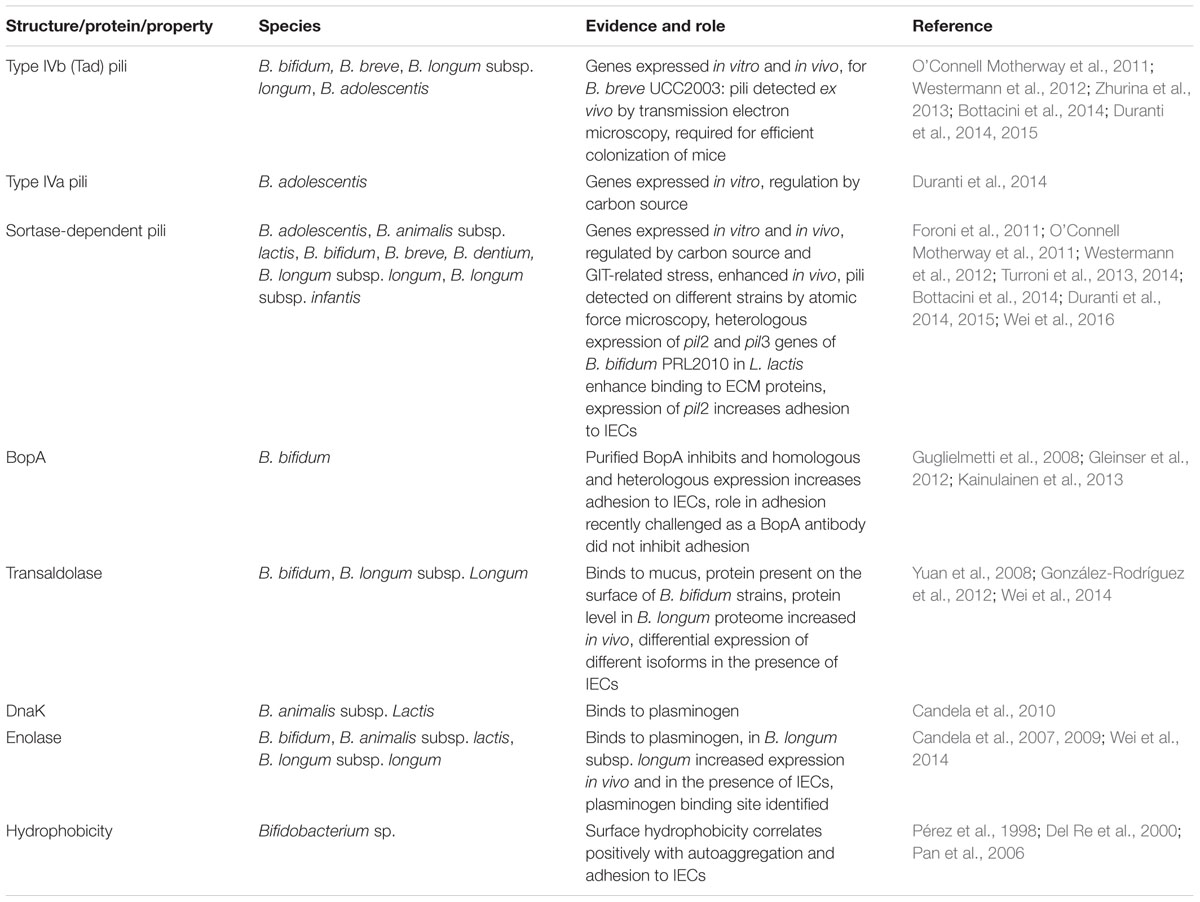
TABLE 1. Adhesive structures identified in bifidobacteria.
Adhesion to mucus is mostly analyzed using microtiter plate assays with immobilized mucus with quantification of adherent bacteria after metabolic labeling using radioisotopes or fluorescent dyes (He et al., 2001; Izquierdo et al., 2008; González-Rodríguez et al., 2012; Kainulainen et al., 2013). Similar assays are performed to analyze adhesion to immobilized ECM proteins (Kainulainen et al., 2013) or detection of ECM proteins bound to bacterial cells or protein extracts by specific antibodies (Candela et al., 2007, 2009, 2010).
The methods and cell lines used to determine adhesion to IECs differ largely between studies and groups. The most widely used cell lines are Caco-2, HT-29, and T84 (Guglielmetti et al., 2009; Preising et al., 2010; Gleinser et al., 2012; González-Rodríguez et al., 2012; Kainulainen et al., 2013; Grimm et al., 2014). In studies that employ more than one cell line, absolute adhesion of different strains may vary between cell lines but relative differences between strains are usually conserved (Riedel et al., 2006; Preising et al., 2010; Gleinser et al., 2012). One observation is that, although there is considerable strain-to-strain variability, strains of the species B. bifidum generally tend to adhere better to IECs than strains of other species (Guglielmetti et al., 2008; Gleinser et al., 2012). Detection of adherent bacteria is performed by metabolic labeling using radionucleotides (Riedel et al., 2006; Kainulainen et al., 2013), enumeration of colony forming units of adherent bacteria (Gleinser et al., 2012; González-Rodríguez et al., 2012), microscopic imaging and calculation of adhesion indices, i.e., the ratio of adherent bacteria and cells (Guglielmetti et al., 2008, 2009, 2010), or expression of fluorescent proteins (Grimm et al., 2014). However, the method of quantification does not seem to impact on adhesion itself as comparable results are obtained using radioactive or fluorescent labeling and plate counting (Riedel et al., 2006; Gleinser et al., 2012; Grimm et al., 2014).
In the following sections, the current knowledge on bifidobacterial adhesion to host structures will be summarized and the involved factors will be compared to adhesion factors of pathogens.
Pili
A wide range of Gram-positive and -negative bacteria possess proteinaceous surface appendages termed fimbriae or pili (Proft and Baker, 2009). In general, pili are adhesive structures that are involved in biofilm formation, conjugation, motility, and adhesion to biotic and abiotic surfaces (Maier and Wong, 2015). These hair-like structures extend to some distance (up to 3 μm) from the bacterial cell surface (Proft and Baker, 2009). It is hypothesized that they are able to bridge the repulsive forces between microbial cells and biological substrates, which under physiological conditions are both negatively charged (Proft and Baker, 2009). Pili are well-known for their role as virulence factors of Gram-positive and -negative pathogens and are important for initial attachment to host tissues (Telford et al., 2006; Proft and Baker, 2009).
There is increasing evidence that bifidobacteria also encode and express pilus-like structures on their cell surface. The first report of pili in bifidobacteria was the presence of genes encoding type IVb tight adherence (Tad) pili in B. breve UCC2003 (O’Connell Motherway et al., 2011). Since then, genes for Tad, Type VIa, and/or sortase-dependent pili were found in basically all sequenced genomes of bifidobacteria (Table 1). Interestingly, in most cases bifidobacteria posses more than one pilus-coding locus and B. dentium harbours as much as seven gene clusters for sortase-dependent pili (Foroni et al., 2011).
Transcriptional analysis revealed that at least some of the genes are expressed under in vitro conditions and are regulated in response to substrate, presence of other bacteria, growth phase, or stress conditions related to the GIT (Foroni et al., 2011; Westermann et al., 2012; Duranti et al., 2014; Turroni et al., 2014). Moreover, pilin proteins are present in the in vitro proteome of B. bifidum S17 (Wei et al., 2016) and pilus-like structures were observed on several bifidobacteria by electron and atomic force microscopy (Foroni et al., 2011; O’Connell Motherway et al., 2011; Duranti et al., 2014). Collectively, this suggests that bifidobacteria possess functional pili. There is also evidence that bifidobacterial pili have a role in colonization of the host and attachment to epithelial cells. In B. breve UCC2003, expression of the tad locus was up-regulated in the GIT of mice and is required for efficient colonization in the presence of a competing microbiota (O’Connell Motherway et al., 2011). In a B. adolescentis strain, expression of pilus gene clusterss and presence of pili were enhanced when bacteria were isolated from the murine GIT or grown on starch, cellobiose or maltodextrin, i.e., substrates abundantly present in the GIT (Duranti et al., 2014). Similarly, expression of two of the three sortase-dependent pili clusters of B. bifidum PRL2010 is enhanced in the murine GIT and in the presence of human IECs in vitro (Turroni et al., 2013). Although a direct role for colonization and adhesion by inactivation of the corresponding genes is missing (probably due to the lack of appropriate genetic tools for the species B. bifidum), heterologous expression of the two gene clusters in Lactococcus lactis led to presence of pilus-like structures. Moreover, the recombinant L. lactis strains displayed increased adhesion to cultured IECs (pil2 cluster) and ECM proteins laminin, fibronectin, fibrinogen, and plasminogen (pil2 and pil3 clusters; Turroni et al., 2013). Adhesion to fibronectin seems to be mediated by sugar-binding domains of the pili since enzymatic deglycosylation of fibronectin markedly reduced adhesion of the recombinant L. lactis strains expressing the pil2 and pil3 gene clusters of B. bifidum PRL2010 (Turroni et al., 2013).
Moonlighting Proteins
A rather obscure group of proteins involved in adhesion of bacteria to host tissues are so-called moonlighting proteins (Huberts and van der Klei, 2010). These proteins are multifunctional and usually have an enzymatic role in bacterial metabolism or other cellular processes but at the same time are involved in totally unrelated biological functions (Huberts and van der Klei, 2010). In more than 90 pathogenic bacteria, proteins with a moonlighting function in virulence have been identified (Henderson, 2014). Interestingly, a large number of moonlighting proteins are cytoplasmic enzymes of the central metabolism that lack secretion signals raising the question if these proteins are actively exported to mediate virulence related functions. The best characterized examples are adhesins of pathogenic bacteria that are involved in primary attachment to host tissue and are important for later stages of infection (Henderson and Martin, 2011).
Enzymes of glycolysis with a moonlighting function in adhesion of pathogens include aldolase (or transaldolase), enolase, and glyceraldehyde-3-phosphate dehydrogenase (Henderson and Martin, 2011). These proteins were detected in proteomes of different bifidobacteria (Yuan et al., 2006, 2008; Ruiz et al., 2009; Gilad et al., 2011; Liu et al., 2011; Wei et al., 2016; Zhu et al., 2016). Transaldolase, a cytoplasmatic key enzyme of the bifidus shunt, was found to be present on the surface of several B. bifidum strains (González-Rodríguez et al., 2012). Using an in vitro binding assay the transaldolase could be identified as a mucin-binding protein and the specificity of this interaction was confirmed by increased mucus binding of recombinant L. lactis strains expressing transaldolase (González-Rodríguez et al., 2012). Enolase of different B. longum, B. bifidum, B. animalis subsp. lactis and B. breve strains was shown to interact with plasminogen (Candela et al., 2007, 2009; Wei et al., 2014). Moreover, the plasminogen binding site in the B. lactis enolase was shown to be homologous to that of Streptococcus pneumoniae and specific amino acid residues crucial for plasminogen binding have been identified (Candela et al., 2009). Another moonlighting protein that serves as an adhesin for bifidobacteria is DnaK, which has a primary function as a chaperone (Henderson and Martin, 2011). For B. animalis subsp. lactis BI07, DnaK was shown to bind plasminogen (Candela et al., 2007, 2010). Further potential moonlighting proteins of B. animalis subsp. lactis BI07 with plasminogen-binding activity are glutamine synthetase, bilesalt hydrolase, and phosphoglycerate mutase (Candela et al., 2007).
For B. longum NCC2705, transaldolase was detected at higher levels incubated in vivo in a rabbit intestinal loop compared to in vitro growth (Yuan et al., 2008) and enolase and transaldolase were more abundant in the proteome following co-cultivation with IECs (Wei et al., 2014). Also, expression of DnaK and enolase is upregulated in several bifidobacteria in response to bile (Savijoki et al., 2005; Candela et al., 2010). This indicates that bifidobacteria might be able to sense the conditions of the intestinal environment and presence of IECs (or receptors on IECs) and respond by enhancing expression of adhesive molecules.
Other Adhesion Factors
A rather general, and non-specific property of bacteria that has been associated occasionally with adhesion of pathogens to host tissue is surface hydrophobicity (Hirt et al., 2000; Kouidhi et al., 2010). Several studies have tested different strains and species of bifidobacteria for hydrophobicity, autoaggregation and adhesion to IECs (Pérez et al., 1998; Del Re et al., 2000; Pan et al., 2006). Overall, the results suggest that (i) strains with higher surface hydrophobicity show higher autoaggregation and adhesion to IECs and (ii) B. bifidum strains tend to be more hydrophobic than strains of other Bifidobacterium sp. This is in line with other studies showing that B. bifidum strains adhere better to IECs than strains of other species (Preising et al., 2010; Gleinser et al., 2012).
Other non-proteinaceous component of the bacterial envelope that have been associated with adhesion to host tissue of Gram positive pathogens are glycoconjugates including exopolysaccharides, lipoteichoic, and wall teichoic acids (Weidenmaier and Peschel, 2008; Tytgat and Lebeer, 2014; Tan et al., 2015). Despite the presence of genes (potentially) involved in biosynthesis of exopolysaccharides and teichoic acids in most of the sequenced genomes of bifidobacteria (Hidalgo-Cantabrana et al., 2014; Colagiorgi et al., 2015), a contribution to adhesion have not been demonstrated conclusively so far. However, one study links exopolysaccharide production of bifidobacteria with adhesion to mucus by showing that purified exopolysaccharides of two bifidobacteria reduced adhesion of intact bacterial cells of these strains (Ruas-Madiedo et al., 2006).
Besides the abovementioned pili and moonlighting proteins no specific adhesins such as intimin, internalins, lectins, fibronectin-binding proteins as described for a number of pathogens (Niemann et al., 2004; Kline et al., 2009) have been characterized for bifidobacteria. A bioinformatic screen of the genome of B. bifidum S17 yielded a number of proteins with domains such as fibronectin type III domain, concanavalin A-like lectin, and collagen triple helix repeat domains, suggesting that bifidobacteria might have similar adhesins (Westermann et al., 2012). A definite role of the corresponding proteins in adhesion to host structures has yet to be demonstrated.
One specific protein that has been suspected to play a role in adhesion of bifidobacteria to IECs is BopA, lipoprotein of the cell envelope specifically found in B. bifidum strains (Guglielmetti et al., 2008; Gleinser et al., 2012). However, BopA contains the characteristic domains of a solute-binding protein and is part of an operon that encodes a putative oligopeptide ABC-transporter (Gleinser et al., 2012). Moreover, a recent study has challenged the idea that BopA serves a function in adhesion by showing that blocking BopA using a specific antibody does not affect adhesion of B. bifidum MIMBb75 to IECs (Kainulainen et al., 2013). Thus, BopA might be another example for a moonlighting protein but whether it has a role in adhesion of B. bifidum strains to intestinal tissue in humans needs to be elucidated in further studies.
Conclusion
A large number of Bifidobacterium sp. strains were shown to adhere to IECs, mucus, and/or ECM proteins. For some bifidobacteria, adhesive structures have been characterized and include pili and different moonlighting proteins. Lactobacilli, another group of potential probiotic, Gram-positive microorganisms use exactly the same structures to adhere to the same target sites on host tissues (Vélez et al., 2007; van Tassell and Miller, 2011). Pathogenic microorganisms employ similar or even identical structures to adhere to host structures. The genus Bacteroides contains highly abundant commensal species as well as opportunistic pathogens that even may cause cancer (Wexler, 2007; Sears et al., 2014). Both commensal and pathogenic strains were shown to adhere to IECs, ECM, or mucus (Brook and Myhal, 1991; Ferreira et al., 2002; Macfarlane et al., 2005; de O Ferreira et al., 2006; Huang et al., 2011; Ferreira Ede et al., 2013) and pili, specific ECM-binding proteins, EPS etc. (Brook and Myhal, 1991; de O Ferreira et al., 2006; Pumbwe et al., 2006; Ferreira Ede et al., 2013) are involved in the process. Collectively, this illustrates that both pathogenic and commensal, in some cases even beneficial, bacteria employ the same strategies to attach to host structures. There is no doubt that adhesion of pathogens to host tissue is required or helps to promote infection. Bifidobacteria are generally regarded as safe microorganisms, which despite intensive studies of the past decades have not shown any pathogenic potential whatsoever. Instead, there are a number of health-related benefits associated with bifidobacteria. Although definitive proof is missing in most cases, it is reasonable to assume that adhesion to host tissue by beneficial bacteria are also required for or support their health-promoting effects. Moreover, the impressive number of different adhesion factors encoded by individual strains of bifidobacteria suggests that adhesion to host tissue is important for bifidobacteria to colonize and strive in the highly competitive ecosystem of the GIT.
Author Contributions
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This study was partially supported by the German Academic Exchange Service/Federal Ministry of Education and Research to CUR (Grant D/09/04778). CW was supported by a by Ph.D. fellowship of the “Landesgraduiertenförderung Baden-Württemberg.” The funders had no role in the decision to publish or the content of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
ECM, extracellular matrix; GIT, gastrointestinal tract; IECs, intestinal epithelial cells.
References
Arumugam, M., Raes, J., Pelletier, E., Le Paslier, D., Yamada, T., Mende, D. R., et al. (2011). Enterotypes of the human gut microbiome. Nature 473, 174–180. doi: 10.1038/nature09944
Bäckhed, F., Roswall, J., Peng, Y., Feng, Q., Jia, H., Kovatcheva-Datchary, P., et al. (2015). Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 17, 690–703. doi: 10.1016/j.chom.2015.04.004
Bermudez-Brito, M., Plaza-Díaz, J., Muñoz-Quezada, S., Gómez-Llorente, C., and Gil, A. (2012). Probiotic mechanisms of action. Ann. Nutr. Metab. 61, 160–174. doi: 10.1159/000342079
Bernet, M. F., Brassart, D., Neeser, J. R., and Servin, A. L. (1993). Adhesion of human bifidobacterial strains to cultured human intestinal epithelial cells and inhibition of enteropathogen-cell interactions. Appl. Environ. Microbiol. 59, 4121–4128.
Bhavsar, A. P., Guttman, J. A., and Finlay, B. B. (2007). Manipulation of host-cell pathways by bacterial pathogens. Nature 449, 827–834. doi: 10.1038/nature06247
Bottacini, F., O Connell Motherway, M., Kuczynski, J., O Connell, K. J., Serafini, F., Duranti, S., et al. (2014). Comparative genomics of the Bifidobacterium breve taxon. BMC Genomics 15:170. doi: 10.1186/1471-2164-15-170
Brook, I., and Myhal, M. L. (1991). Adherence of Bacteroides fragilis group species. Infect. Immun. 59, 742–744.
Buffie, C. G., and Pamer, E. G. (2013). Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 13, 790–801. doi: 10.1038/nri3535
Candela, M., Bergmann, S., Vici, M., Vitali, B., Turroni, S., Eikmanns, B. J., et al. (2007). Binding of human plasminogen to Bifidobacterium. J. Bacteriol. 189, 5929–5936. doi: 10.1128/JB.00159-07
Candela, M., Biagi, E., Centanni, M., Turroni, S., Vici, M., Musiani, F., et al. (2009). Bifidobacterial enolase, a cell surface receptor for human plasminogen involved in the interaction with the host. Microbiology 155, 3294–3303. doi: 10.1099/mic.0.028795-0
Candela, M., Centanni, M., Fiori, J., Biagi, E., Turroni, S., Orrico, C., et al. (2010). DnaK from Bifidobacterium animalis subsp. lactis is a surface-exposed human plasminogen receptor upregulated in response to bile salts. Microbiology 156, 1609–1618. doi: 10.1099/mic.0.038307-0
Candela, M., Perna, F., Carnevali, P., Vitali, B., Ciati, R., Gionchetti, P., et al. (2008). Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol. 125, 286–292. doi: 10.1016/j.ijfoodmicro.2008.04.012
Colagiorgi, A., Turroni, F., Mancabelli, L., Serafini, F., Secchi, A., van Sinderen, D., et al. (2015). Insights into teichoic acid biosynthesis by Bifidobacterium bifidum PRL2010. FEMS Microbiol. Lett. 362:fnv141. doi: 10.1093/femsle/fnv141
Collado, M. C., Gueimonde, M., Hernández, M., Sanz, Y., and Salminen, S. (2005). Adhesion of selected Bifidobacterium strains to human intestinal mucus and the role of adhesion in enteropathogen exclusion. J. Food Prot. 68, 2672–2678.
de O Ferreira, E., Araújo Lobo, L., Barreiros Petrópolis, D., dos S Avelar, K. E., Ferreira, M. C., e Silva Filho, F. C., et al. (2006). A Bacteroides fragilis surface glycoprotein mediates the interaction between the bacterium and the extracellular matrix component laminin-1. Res. Microbiol. 157, 960–966. doi: 10.1016/j.resmic.2006.09.005
Del Re, B., Sgorbati, B., Miglioli, M., and Palenzona, D. (2000). Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 31, 438–442. doi: 10.1046/j.1365-2672.2000.00845.x
Duranti, S., Milani, C., Lugli, G. A., Turroni, F., Mancabelli, L., Sanchez, B., et al. (2015). Insights from genomes of representatives of the human gut commensal Bifidobacterium bifidum. Environ. Microbiol. 17, 2515–2531. doi: 10.1111/1462-2920.12743
Duranti, S., Turroni, F., Lugli, G. A., Milani, C., Viappiani, A., Mangifesta, M., et al. (2014). Genomic characterization and transcriptional studies of the starch-utilizing Bifidobacterium adolescentis 22L. Appl. Environ. Microbiol. 80, 6080–6090. doi: 10.1128/AEM.01993-14
Fernández, L., Langa, S., Martín, V., Maldonado, A., Jiménez, E., Martín, R., et al. (2013). The human milk microbiota: origin and potential roles in health and disease. Pharmacol. Res. 69, 1–10. doi: 10.1016/j.phrs.2012.09.001
Ferreira, E. O., Falcão, L. S., Vallim, D. C., Santos, F. J., Andrade, J. R. C., Andrade, A. F. B., et al. (2002). Bacteroides fragilis adherence to Caco-2 cells. Anaerobe 8, 307–314. doi: 10.1016/S1075-9964(03)00008-8
Ferreira Ede, O., Teixeira, F. L., Cordeiro, F., Araujo Lobo, L., Rocha, E. R., Smith, J. C., et al. (2013). The Bfp60 surface adhesin is an extracellular matrix and plasminogen protein interacting in Bacteroides fragilis. Int. J. Med. Microbiol. 303, 492–497. doi: 10.1016/j.ijmm.2013.06.007
Foligné, B., Daniel, C., and Pot, B. (2013). Probiotics from research to market: the possibilities, risks and challenges. Curr. Opin. Microbiol. 16, 284–292. doi: 10.1016/j.mib.2013.06.008
Foroni, E., Serafini, F., Amidani, D., Turroni, F., He, F., Bottacini, F., et al. (2011). Genetic analysis and morphological identification of pilus-like structures in members of the genus Bifidobacterium. Microb. Cell Fact. 10(Suppl. 1), S16. doi: 10.1186/1475-2859-10-S1-S16
Gareau, M. G., Sherman, P. M., and Walker, W. A. (2010). Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 7, 503–514. doi: 10.1038/nrgastro.2010.117
Gilad, O., Svensson, B., Viborg, A. H., Stuer-Lauridsen, B., and Jacobsen, S. (2011). The extracellular proteome of Bifidobacterium animalis subsp. lactis BB-12 reveals proteins with putative roles in probiotic effects. Proteomics 11, 2503–2514. doi: 10.1002/pmic.201000716
Gleinser, M., Grimm, V., Zhurina, D., Yuan, J., and Riedel, C. U. (2012). Improved adhesive properties of recombinant bifidobacteria expressing the Bifidobacterium bifidum-specific lipoprotein BopA. Microb. Cell Fact. 11, 80. doi: 10.1186/1475-2859-11-80
González-Rodríguez, I., Sánchez, B., Ruiz, L., Turroni, F., Ventura, M., Ruas-Madiedo, P., et al. (2012). Role of extracellular transaldolase from Bifidobacterium bifidum in mucin adhesion and aggregation. Appl. Environ. Microbiol. 78, 3992–3998. doi: 10.1128/AEM.08024-11
Grimm, V., Gleinser, M., Neu, C., Zhurina, D., and Riedel, C. U. (2014). Expression of fluorescent proteins in bifidobacteria for analysis of host-microbe interactions. Appl. Environ. Microbiol. 80, 2842–2850. doi: 10.1128/AEM.04261-13
Grönlund, M.-M., Grześkowiak, L., Isolauri, E., and Salminen, S. (2011). Influence of mother’s intestinal microbiota on gut colonization in the infant. Gut Microbes 2, 227–233. doi: 10.4161/gmic.2.4.16799
Guglielmetti, S., Tamagnini, I., Minuzzo, M., Arioli, S., Parini, C., Comelli, E., et al. (2009). Study of the adhesion of Bifidobacterium bifidum MIMBb75 to human intestinal cell lines. Curr. Microbiol. 59, 167–172. doi: 10.1007/s00284-009-9415-x
Guglielmetti, S., Tamagnini, I., Mora, D., Minuzzo, M., Scarafoni, A., Arioli, S., et al. (2008). Implication of an outer surface lipoprotein in adhesion of Bifidobacterium bifidum to Caco-2 cells. Appl. Environ. Microbiol. 74, 4695–4702. doi: 10.1128/AEM.00124-08
Guglielmetti, S., Taverniti, V., Minuzzo, M., Arioli, S., Zanoni, I., Stuknyte, M., et al. (2010). A dairy bacterium displays in vitro probiotic properties for the pharyngeal mucosa by antagonizing group A streptococci and modulating the immune response. Infect. Immun. 78, 4734–4743. doi: 10.1128/IAI.00559-10
He, F., Ouwehan, A. C., Hashimoto, H., Isolauri, E., Benno, Y., and Salminen, S. (2001). Adhesion of Bifidobacterium spp. to human intestinal mucus. Microbiol. Immunol. 45, 259–262. doi: 10.1111/j.1348-0421.2001.tb02615.x
Henderson, B. (2014). An overview of protein moonlighting in bacterial infection. Biochem. Soc. Trans. 42, 1720–1727. doi: 10.1042/BST20140236
Henderson, B., and Martin, A. (2011). Bacterial virulence in the moonlight: multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 79, 3476–3491. doi: 10.1128/IAI.00179-11
Hidalgo-Cantabrana, C., Sánchez, B., Milani, C., Ventura, M., Margolles, A., and Ruas-Madiedo, P. (2014). Genomic overview and biological functions of exopolysaccharide biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 80, 9–18. doi: 10.1128/AEM.02977-13
Hirt, H., Erlandsen, S. L., and Dunny, G. M. (2000). Heterologous inducible expression of Enterococcus faecalis pCF10 aggregation substance asc10 in Lactococcus lactis and Streptococcus gordonii contributes to cell hydrophobicity and adhesion to fibrin. J. Bacteriol. 182, 2299–2306. doi: 10.1128/JB.182.8.2299-2306.2000
Holmes, E., Kinross, J., Gibson, G. R., Burcelin, R., Jia, W., Pettersson, S., et al. (2012). Therapeutic modulation of microbiota-host metabolic interactions. Sci. Transl. Med. 4:137rv6. doi: 10.1126/scitranslmed.3004244
Huang, J. Y., Lee, S. M., and Mazmanian, S. K. (2011). The human commensal Bacteroides fragilis binds intestinal mucin. Anaerobe 17, 137–141. doi: 10.1016/j.anaerobe.2011.05.017
Huberts, D. H. E. W., and van der Klei, I. J. (2010). Moonlighting proteins: an intriguing mode of multitasking. Biochim. Biophys. Acta 1803, 520–525. doi: 10.1016/j.bbamcr.2010.01.022
Izquierdo, E., Medina, M., Ennahar, S., Marchioni, E., and Sanz, Y. (2008). Resistance to simulated gastrointestinal conditions and adhesion to mucus as probiotic criteria for Bifidobacterium longum strains. Curr. Microbiol. 56, 613–618. doi: 10.1007/s00284-008-9135-7
Johansson, M. E. V. (2014). Mucus layers in inflammatory bowel disease. Inflamm. Bowel Dis. 20, 2124–2131. doi: 10.1097/MIB.0000000000000117
Jost, T., Lacroix, C., Braegger, C., and Chassard, C. (2015). Impact of human milk bacteria and oligosaccharides on neonatal gut microbiota establishment and gut health. Nutr. Rev. 73, 426–437. doi: 10.1093/nutrit/nuu016
Kainulainen, V., Reunanen, J., Hiippala, K., Guglielmetti, S., Vesterlund, S., Palva, A., et al. (2013). BopA has no major role in the adhesion of Bifidobacterium bifidum to intestinal epithelial cells, extracellular matrix proteins and mucus. Appl. Environ. Microbiol. 79, 6989–6997. doi: 10.1128/AEM.01993-13
Klaenhammer, T. R., and Kullen, M. J. (1999). Selection and design of probiotics. Int. J. Food Microbiol. 50, 45–57. doi: 10.1016/S0168-1605(99)00076-8
Kline, K. A., Fälker, S., Dahlberg, S., Normark, S., and Henriques-Normark, B. (2009). Bacterial adhesins in host-microbe interactions. Cell Host Microbe 5, 580–592. doi: 10.1016/j.chom.2009.05.011
Kouidhi, B., Zmantar, T., Hentati, H., and Bakhrouf, A. (2010). Cell surface hydrophobicity, biofilm formation, adhesives properties and molecular detection of adhesins genes in Staphylococcus aureus associated to dental caries. Microb. Pathog. 49, 14–22. doi: 10.1016/j.micpath.2010.03.007
Lawley, T. D., and Walker, A. W. (2013). Intestinal colonization resistance. Immunology 138, 1–11. doi: 10.1111/j.1365-2567.2012.03616.x
Liu, D., Wang, S., Xu, B., Guo, Y., Zhao, J., Liu, W., et al. (2011). Proteomics analysis of Bifidobacterium longum NCC2705 growing on glucose, fructose, mannose, xylose, ribose, and galactose. Proteomics 11, 2628–2638. doi: 10.1002/pmic.201100035
Macfarlane, S., Woodmansey, E. J., and Macfarlane, G. T. (2005). Colonization of mucin by human intestinal bacteria and establishment of biofilm communities in a two-stage continuous culture system. Appl. Environ. Microbiol. 71, 7483–7492. doi: 10.1128/AEM.71.11.7483-7492.2005
Maier, B., and Wong, G. C. L. (2015). How bacteria use type IV pili machinery on surfaces. Trends Microbiol. 23, 775–788. doi: 10.1016/j.tim.2015.09.002
Marchesi, J. R., Adams, D. H., Fava, F., Hermes, G. D. A., Hirschfield, G. M., Hold, G., et al. (2016). The gut microbiota and host health: a new clinical frontier. Gut 65, 330–339. doi: 10.1136/gutjnl-2015-309990
Matamoros, S., Gras-Leguen, C., Le Vacon, F., Potel, G., and de La Cochetiere, M.-F. (2013). Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 21, 167–173. doi: 10.1016/j.tim.2012.12.001
Niemann, H. H., Schubert, W. D., and Heinz, D. W. (2004). Adhesins and invasins of pathogenic bacteria: a structural view. Microbes Infect. 6, 101–112. doi: 10.1016/j.micinf.2003.11.001
O’Connell Motherway, M., Zomer, A., Leahy, S. C., Reunanen, J., Bottacini, F., Claesson, M. J., et al. (2011). Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. U.S.A. 108, 11217–11222. doi: 10.1073/pnas.1105380108
Pan, W.-H., Li, P.-L., and Liu, Z. (2006). The correlation between surface hydrophobicity and adherence of Bifidobacterium strains from centenarians’ faeces. Anaerobe 12, 148–152. doi: 10.1016/j.anaerobe.2006.03.001
Papadimitriou, K., Zoumpopoulou, G., Foligné, B., Alexandraki, V., Kazou, M., Pot, B., et al. (2015). Discovering probiotic microorganisms: in vitro, in vivo, genetic and omics approaches. Front. Microbiol. 6:58. doi: 10.3389/fmicb.2015.00058
Pérez, P. F., Minnaard, Y., Disalvo, E. A., and De Antoni, G. L. (1998). Surface properties of bifidobacterial strains of human origin. Appl. Environ. Microbiol. 64, 21–26.
Preising, J., Philippe, D., Gleinser, M., Wei, H., Blum, S., Eikmanns, B. J., et al. (2010). Selection of bifidobacteria based on adhesion and anti-inflammatory capacity in vitro for amelioration of murine colitis. Appl. Environ. Microbiol. 76, 3048–3051. doi: 10.1128/AEM.03127-09
Proft, T., and Baker, E. N. (2009). Pili in Gram-negative and Gram-positive bacteria - structure, assembly and their role in disease. Cell. Mol. Life Sci. 66, 613–635. doi: 10.1007/s00018-008-8477-4
Pumbwe, L., Skilbeck, C. A., and Wexler, H. M. (2006). The Bacteroides fragilis cell envelope: quarterback, linebacker, coach—or all three? Anaerobe 12, 211–220. doi: 10.1016/j.anaerobe.2006.09.004
Riedel, C. U., Foata, F., Goldstein, D. R., Blum, S., and Eikmanns, B. J. (2006). Interaction of bifidobacteria with Caco-2 cells-adhesion and impact on expression profiles. Int. J. Food Microbiol. 110, 62–68. doi: 10.1016/j.ijfoodmicro.2006.01.040
Ruas-Madiedo, P., Gueimonde, M., Margolles, A., de los Reyes-Gavilán, C. G., and Salminen, S. (2006). Exopolysaccharides produced by probiotic strains modify the adhesion of probiotics and enteropathogens to human intestinal mucus. J. Food Prot. 69, 2011–2015.
Ruiz, L., Couté, Y., Sánchez, B., de los Reyes-Gavilán, C. G., Sanchez, J.-C., and Margolles, A. (2009). The cell-envelope proteome of Bifidobacterium longum in an in vitro bile environment. Microbiology 155, 957–967. doi: 10.1099/mic.0.024273-0
Savijoki, K., Suokko, A., Palva, A., Valmu, L., Kalkkinen, N., and Varmanen, P. (2005). Effect of heat-shock and bile salts on protein synthesis of Bifidobacterium longum revealed by [35S]methionine labelling and two-dimensional gel electrophoresis. FEMS Microbiol. Lett. 248, 207–215. doi: 10.1016/j.femsle.2005.05.032
Sears, C. L., Geis, A. L., and Housseau, F. (2014). Bacteroides fragilis subverts mucosal biology: from symbiont to colon carcinogenesis. J. Clin. Invest. 124, 4166–4172. doi: 10.1172/JCI72334
Serafini, F., Strati, F., Ruas-Madiedo, P., Turroni, F., Foroni, E., Duranti, S., et al. (2013). Evaluation of adhesion properties and antibacterial activities of the infant gut commensal Bifidobacterium bifidum PRL2010. Anaerobe 21, 9–17. doi: 10.1016/j.anaerobe.2013.03.003
Sivan, A., Corrales, L., Hubert, N., Williams, J. B., Aquino-Michaels, K., Earley, Z. M., et al. (2015). Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 350, 1084–1089. doi: 10.1126/science.aac4255
Stavru, F., Archambaud, C., and Cossart, P. (2011). Cell biology and immunology of Listeria monocytogenes infections: novel insights. Immunol. Rev. 240, 160–184. doi: 10.1111/j.1600-065X.2010.00993.x
Tailford, L. E., Crost, E. H., Kavanaugh, D., and Juge, N. (2015). Mucin glycan foraging in the human gut microbiome. Front. Genet. 6:81. doi: 10.3389/fgene.2015.00081
Tan, F. Y. Y., Tang, C. M., and Exley, R. M. (2015). Sugar coating: bacterial protein glycosylation and host–microbe interactions. Trends Biochem. Sci. 40, 342–350. doi: 10.1016/j.tibs.2015.03.016
Telford, J. L., Barocchi, M. A., Margarit, I., Rappuoli, R., and Grandi, G. (2006). Pili in gram-positive pathogens. Nat. Rev. Microbiol. 4, 509–519. doi: 10.1038/nrmicro1443
Tuomola, E., Crittenden, R., Playne, M., Isolauri, E., and Salminen, S. (2001). Quality assurance criteria for probiotic bacteria. Am. J. Clin. Nutr. 73, 393S–398S.
Turroni, F., Serafini, F., Foroni, E., Duranti, S., O’Connell Motherway, M., Taverniti, V., et al. (2013). Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions. Proc. Natl. Acad. Sci. U.S.A. 110, 11151–11156. doi: 10.1073/pnas.1303897110
Turroni, F., Serafini, F., Mangifesta, M., Arioli, S., Mora, D., van Sinderen, D., et al. (2014). Expression of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in response to environmental gut conditions. FEMS Microbiol. Lett. 357, 23–33. doi: 10.1111/1574-6968.12509
Tytgat, H. L. P., and Lebeer, S. (2014). The sweet tooth of bacteria: common themes in bacterial glycoconjugates. Microbiol. Mol. Biol. Rev. 78, 372–417. doi: 10.1128/MMBR.00007-14
van Tassell, M. L., and Miller, M. J. (2011). Lactobacillus adhesion to mucus. Nutrients 3, 613–636. doi: 10.3390/nu3050613
Vélez, M. P., De Keersmaecker, S. C. J., and Vanderleyden, J. (2007). Adherence factors of Lactobacillus in the human gastrointestinal tract. FEMS Microbiol. Lett. 276, 140–148. doi: 10.1111/j.1574-6968.2007.00908.x
Ventura, M., Turroni, F., Zomer, A., Foroni, E., Giubellini, V., Bottacini, F., et al. (2009). The Bifidobacterium dentium Bd1 genome sequence reflects its genetic adaptation to the human oral cavity. PLoS Genet. 5:e1000785. doi: 10.1371/journal.pgen.1000785
Walker, A. W., Martin, J. C., Scott, P., Parkhill, J., Flint, H. J., and Scott, K. P. (2015). 16S rRNA gene-based profiling of the human infant gut microbiota is strongly influenced by sample processing and PCR primer choice. Microbiome 3, 26. doi: 10.1186/s40168-015-0087-4
Wei, X., Wang, S., Zhao, X., Wang, X., Li, H., Lin, W., et al. (2016). Proteomic profiling of Bifidobacterium bifidum S17 cultivated under in vitro conditions. Front. Microbiol. 7:97. doi: 10.3389/fmicb.2016.00097
Wei, X., Yan, X., Chen, X., Yang, Z., Li, H., Zou, D., et al. (2014). Proteomic analysis of the interaction of Bifidobacterium longum NCC2705 with the intestine cells Caco-2 and identification of plasminogen receptors. J. Proteomics 108, 89–98. doi: 10.1016/j.jprot.2014.04.038
Weidenmaier, C., and Peschel, A. (2008). Teichoic acids and related cell-wall glycopolymers in Gram-positive physiology and host interactions. Nat. Rev. Microbiol. 6, 276–287. doi: 10.1038/nrmicro1861
Westermann, C., Zhurina, D. S., Baur, A., Shang, W., Yuan, J., and Riedel, C. U. (2012). Exploring the genome sequence of Bifidobacterium bifidum S17 for potential players in host-microbe interactions. Symbiosis 58, 191–200. doi: 10.1007/s13199-012-0205-z
Wexler, H. M. (2007). Bacteroides: the good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 20, 593–621. doi: 10.1128/CMR.00008-07
Whelan, K., and Quigley, E. M. M. (2013). Probiotics in the management of irritable bowel syndrome and inflammatory bowel disease. Curr. Opin. Gastroenterol. 29, 184–189. doi: 10.1097/MOG.0b013e32835d7bba
Yatsunenko, T., Rey, F. E., Manary, M. J., Trehan, I., Dominguez-Bello, M. G., Contreras, M., et al. (2012). Human gut microbiome viewed across age and geography. Nature 486, 222–227. doi: 10.1038/nature11053
Yuan, J., Wang, B., Sun, Z., Bo, X., Yuan, X., He, X., et al. (2008). Analysis of host-inducing proteome changes in bifidobacterium longum NCC2705 grown in Vivo. J. Proteome Res. 7, 375–385. doi: 10.1021/pr0704940
Yuan, J., Zhu, L., Liu, X., Li, T., Zhang, Y., Ying, T., et al. (2006). A proteome reference map and proteomic analysis of Bifidobacterium longum NCC2705. Mol. Cell. Proteomics 5, 1105–1118. doi: 10.1074/mcp.M500410-MCP200
Zhu, D., Sun, Y., Liu, F., Li, A., Yang, L., and Meng, X.-C. (2016). Identification of surface-associated proteins of Bifidobacterium animalis ssp. lactis KLDS 2.0603 by enzymatic shaving. J. Dairy Sci. 99, 5155–5172. doi: 10.3168/jds.2015-10581
Keywords: Bifidobacterium, probiotic, adhesion, host interaction
Citation: Westermann C, Gleinser M, Corr SC and Riedel CU (2016) A Critical Evaluation of Bifidobacterial Adhesion to the Host Tissue. Front. Microbiol. 7:1220. doi: 10.3389/fmicb.2016.01220
Received: 27 May 2016; Accepted: 22 July 2016;
Published: 05 August 2016.
Edited by:
Francesca Turroni, University College Cork, IrelandReviewed by:
Marco Ventura, University of Parma, ItalySimone Rampelli, University of Bologna, Italy
Copyright © 2016 Westermann, Gleinser, Corr and Riedel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian U. Riedel, christian.riedel@uni-ulm.de
†These authors have contributed equally to this work.