 Carmen B. Lupiañez1,2
Carmen B. Lupiañez1,2 María T. Villaescusa3,4
María T. Villaescusa3,4 Agostinho Carvalho5,6
Agostinho Carvalho5,6 Jan Springer7Michaela Lackner8José M. Sánchez-Maldonado1Luz M. Canet1Cristina Cunha5,6Juana Segura-Catena1,2
Jan Springer7Michaela Lackner8José M. Sánchez-Maldonado1Luz M. Canet1Cristina Cunha5,6Juana Segura-Catena1,2 Laura Alcazar-Fuoli9Carlos Solano10Luana Fianchi11
Laura Alcazar-Fuoli9Carlos Solano10Luana Fianchi11 Livio Pagano11Leonardo Potenza12José M. Aguado13Mario Luppi12Manuel Cuenca-Estrella9Cornelia Lass-Flörl8Hermann Einsele7Lourdes Vázquez3 PCRAGA Study Group14Rafael Ríos-Tamayo1,2Jurgen Loeffler7Manuel Jurado1,2
Livio Pagano11Leonardo Potenza12José M. Aguado13Mario Luppi12Manuel Cuenca-Estrella9Cornelia Lass-Flörl8Hermann Einsele7Lourdes Vázquez3 PCRAGA Study Group14Rafael Ríos-Tamayo1,2Jurgen Loeffler7Manuel Jurado1,2 Juan Sainz1,2*
Juan Sainz1,2*- 1Genomic Oncology Area, GENYO, Center for Genomics and Oncological Research, Pfizer/University of Granada/Andalusian Regional Government, PTS Granada, Granada, Spain
- 2Hematology Department, Virgen de las Nieves University Hospital, Granada, Spain
- 3Hematology Department, University Hospital of Salamanca, Salamanca, Spain
- 4Hematology Department, Jiménez Díaz Foundation, Madrid, Spain
- 5Life and Health Sciences Research Institute (ICVS), School of Health Sciences, University of Minho, Braga, Portugal
- 6ICVS/3B's - PT Government Associate Laboratory, Braga, Portugal
- 7Universitätsklinikum Würzburg, Medizinische Klinik II, Würzburg, Germany
- 8Division of Hygiene and Medical Microbiology, Medical University of Innsbruck, Innsbruck, Austria
- 9Mycology Reference Laboratory, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Madrid, Spain
- 10Hematology Department, Clinic University Hospital of Valencia, Valencia, Spain
- 11Istituto di Ematologia, Università Cattolica del S. Cuore, Rome, Italy
- 12Department of Medical and Surgical Sciences, University of Modena and Reggio Emilia, AOU Policlinico, Modena, Italy
- 13Unit of Infectious Diseases, University Hospital 12 de Octubre, Research Institute of Hospital 12 de Octubre (i+12), Madrid, Spain
- 14PCRAGA Study Group, Salamanca, Spain
Invasive Aspergillosis (IA) is an opportunistic infection caused by Aspergillus, a ubiquitously present airborne pathogenic mold. A growing number of studies suggest a major host genetic component in disease susceptibility. Here, we evaluated whether 14 single-nucleotide polymorphisms within NFκB1, NFκB2, RelA, RelB, Rel, and IRF4 genes influence the risk of IA in a population of 834 high-risk patients (157 IA and 677 non-IA) recruited through a collaborative effort involving the aspBIOmics consortium and four European clinical institutions. No significant overall associations between selected SNPs and the risk of IA were found in this large cohort. Although a hematopoietic stem cell transplantation (HSCT)-stratified analysis revealed that carriers of the IRF4rs12203592T/T genotype had a six-fold increased risk of developing the infection when compared with those carrying the C allele (ORREC = 6.24, 95%CI 1.25–31.2, P = 0.026), the association of this variant with IA risk did not reach significance at experiment-wide significant threshold. In addition, we found an association of the IRF4AATC and IRF4GGTC haplotypes (not including the IRF4rs12203592T risk allele) with a decreased risk of IA but the magnitude of the association was similar to the one observed in the single-SNP analysis, which indicated that the haplotypic effect on IA risk was likely due to the IRF4rs12203592 SNP. Finally, no evidence of significant interactions among the genetic markers tested and the risk of IA was found. These results suggest that the SNPs on the studied genes do not have a clinically relevant impact on the risk of developing IA.
Introduction
Invasive Aspergillosis (IA) is an opportunistic infection often caused by species of Aspergillus, a common saprophytic filamentous fungus that is ubiquitously present in the environment. The LIFE initiative has estimated that around 30 million patients worldwide are at high risk of developing IA every year and over 200,000 patients develop the infection annually. Despite the substantial improvement in efficacy of newly developed anti-fungal drugs (mainly azoles alone or in combination with other antifungal drugs), IA continues to be a life-threatening infection in an increasing proportion of immunocompromised or critically ill subjects (Neofytos et al., 2009; Steinbach et al., 2012). IA is frequently encountered in patients of intensive care (Meersseman et al., 2007) and solid organ transplantation units (Singh et al., 2013) but also among those patients who undergo allogeneic hematopoietic stem cell transplantation (allo-HSCT) or are diagnosed either with acute myeloid leukemia (AML) or acute lymphoid leukemia (ALL) and receive high-dose chemotherapy regimens (Steinbach et al., 2012; Neofytos et al., 2013a,b).
Although major clinical risk factors for IA have been identified (Kousha et al., 2011) and the management of high-risk patients has been improved through the optimization of prevention strategies and early initiation of anti-fungal prophylaxis, mortality rates for IA remain still today unacceptably high (up to 30–60%; Neofytos et al., 2009; Steinbach et al., 2012; Karthaus and Buchheidt, 2013). These observational findings suggest that additional factors may contribute to the risk of developing IA. In this regard, a growing number of studies have suggested that host genetic polymorphisms within or near immune-related genes may contribute to determine the risk of developing the infection (Kesh et al., 2005; Sainz et al., 2007a,b, 2008a,b, 2010, 2012; Bochud et al., 2008; Mezger et al., 2008; Ramaprakash et al., 2009; Cunha et al., 2010, 2011, 2014; Chai et al., 2011; Grube et al., 2013; Stappers et al., 2014; Wojtowicz et al., 2015a,b). Interestingly, a substantial proportion of the susceptibility markers identified to date for IA are located in genes directly or indirectly implicated in the activation of the nuclear factor-kappa B (NFκB) signaling pathway, which suggests a relevant role of this biological route in determining the risk of developing this fungal infection. In particular, it has been reported that single nucleotide polymorphisms (SNPs) within toll-like receptors (TLRs; Mambula et al., 2002; Kesh et al., 2005; Bochud et al., 2008; Pamer, 2008; Ramaprakash et al., 2009; Carvalho et al., 2012; Grube et al., 2013), C-type lectins (Cunha et al., 2010; Chai et al., 2011; Sainz et al., 2012), PTX3 (Cunha et al., 2014), and tumor necrosis factor receptors (TNFRs; Sainz et al., 2007b, 2010), which are pathogen recognition receptors (PRRs) that often culminate in the activation of NFκB pathway, may render patients more susceptible to develop IA.
Based on these findings but also those that have demonstrated that NFκB1 (p105/p50), NFκB2 (p100/p52), Rel A (p65), RelB, c-Rel genes may form homo- and hetero-dimers to regulate the activation of the canonical and non-canonical NFκB pathways (Moynagh, 2005; Gilmore, 2006; Hoffmann et al., 2006; Schlitzer et al., 2013; Bajaña et al., 2016) but also IRF4-dependent immune responses (Boddicker et al., 2015), we hypothesized that the presence of common genetic polymorphisms within NFκB1, NFκB2, RelA, RelB, Rel, and IRF4 genes might influence the risk of developing IA in high-risk patients. Thus, the aim of the present study was to investigate the relationship between 14 single nucleotide polymorphisms within these genes and the risk of IA but also to determine whether these variants could interact with each other to modify the risk of developing the infection.
Material and Methods
Study Population
Eight hundred and thirty-four high-risk European Caucasian patients undergoing allo-HSCT or being diagnosed with acute myeloid leukemia (AML) or acute lymphoid leukemia (ALL) and receiving intensive remission-induction chemotherapy were recruited in this case-control population-based study (Lupiañez et al., 2015). Three hundred and thirty-five patients were ascertained from the aspBIOmics consortium (http://www.aspbiomics.eu) whereas 341 patients were collected from two Spanish medical institutions (University Hospital of Salamanca and Clinic University Hospital of Valencia) and through a Spanish multicenter clinical trial (PCRAGA, EU clinical trial number: 2010-019406-17; Aguado et al., 2015). In addition, 148 patients were recruited from two Italian medical institutions (Università Cattolica del S. Cuore, Rome; and University of Modena and Reggio Emilia, AOU Policlinico, Modena). Of those 834 patients, a total of 157 patients were diagnosed with proven or probable IA according to the updated EORTC/MSG criteria (De Pauw et al., 2008) whereas the remaining 677 patients did not show any sign of fungal infection.
SNP Selection and Genotyping
Fourteen polymorphisms within the NFκB1, NFκB2, RelA, RelB, Rel, and IRF4 genes were selected to be genotyped in the whole population (Table 1). SNP selection was based on previously reported associations with cancer (Curtin et al., 2010; Do et al., 2010; Han et al., 2011; Slattery et al., 2011; Seufert et al., 2013; Wang et al., 2014) and immune-related diseases including infections (Chen et al., 2006, 2016; Gregersen et al., 2009; Trynka et al., 2009; Eyre et al., 2010; Varadé et al., 2011; Bowes et al., 2012; Ellinghaus et al., 2012; Ali et al., 2013; Leung et al., 2014; Pan et al., 2015) but also because their potential functionality according to the Haploreg (http://www.broadinstitute.org/mammals/haploreg/haploreg.php) and ENCODE annotation data (https://genome.ucsc.edu/ENCODE/). The genotyping of the selected polymorphisms was carried out at GENYO (Center for Genomics and Oncological Research: Pfizer/University of Granada/Andalusian Regional Government, Granada, Spain) using KASPar® assays (LGC Genomics, Hoddesdon, UK) according to manufacturer's instructions. For internal quality control, 5% of samples were randomly selected and included as duplicates. Concordance between the original and the duplicate samples for the 14 SNPs was ≥99.0%. Call rates for all SNPs were ≥90.0% with the exception of the IRF4rs872071 SNP that was excluded from further analysis.
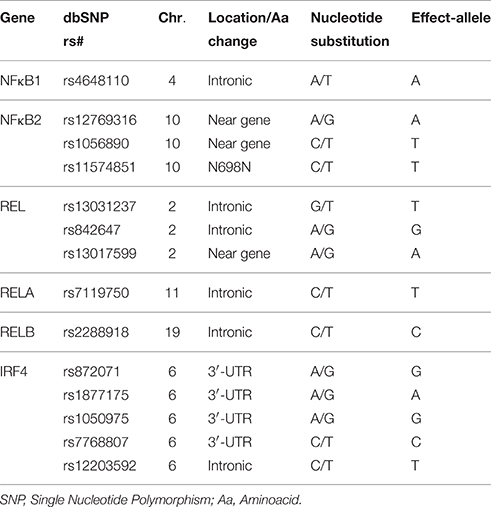
Table 1. NFκB-related polymorphisms.
Statistical Analysis
The Hardy-Weinberg Equilibrium (HWE) test was performed in the control group (non-IA patients) by a standard observed-expected chi-square (χ2). Logistic regression analysis adjusted for age, sex, country of origin, allo-HSCT, underlying disease and prophylaxis status was used to assess the main effects of the selected SNPs on IA risk. We also performed gene-HSCT interaction analyses to determine whether the association between SNPs and IA was of similar magnitude in HSCT (at highest risk) and non-HSCT patients. Although the selection of variables for adjustment was based on well-established risk factors for IA, the partial availability of data regarding prophylaxis status, type of chemotherapy, immunosuppressive drugs (HSCT), HLA mismatch (HSCT), or CMV status did not allow us to assess the impact of these factors on the genetic associations tested. Statistical power of the overall and HSCT-stratified analyses was estimated using Quanto software (http://hydra.usc.edu/gxe/). All tests were conducted using the statistical software SPSS (v.20) and STATA (v.12) for MAC.
In order to account for multiple comparisons, we calculated an adjusted significance level using the Meff method (Nyholt, 2004) but also considering the number of inheritance models tested (codominant, dominant, recessive, and log-additive). Thus, the significant threshold used for the main effect analysis was 0.001 ([0.05]/13 independent genetic markers]/4 inheritance models).
Linkage Disequilibrium (LD) and Haplotype Analysis
Haplotype blocks were constructed from the genotyping data of the non-IA control group using the SNPtool (http://www.dkfz.de/de/molgen_epidemiology/tools/SNPtool.html; Chen et al., 2009) and the Haploview software (v.4.2). Selected polymorphisms within the same locus were not in linkage disequilibrium ensuring that these variants represented independent variability signals (Supplementary Figure 1). In addition to the analysis based on a single SNP, we performed haplotype frequency estimation and haplotype association analysis using SNPStats (http://bioinfo.iconcologia.net/SNPstats). Haplotype frequencies were determined using the Expectation-maximization (EM) algorithm and the minimum haplotype frequency was set at 0.01, therefore assessing association for common haplotypes.
eQTL Analysis
We also assessed whether selected polymorphisms correlated with mRNA expression in the publicly available eQTL IGV browser for primary cells (http://www.gtexportal.org/home/) or using the Haploreg data.
SNP-SNP Interaction Analysis
We also were interested in testing whether NFκB-related SNPs could interact with each other to modify the risk of IA using the multifactor dimensionality reduction (MDR) software. A detailed description of the MDR method has been reported elsewhere (Ritchie et al., 2003; Moore, 2004). A 10-fold cross-validation analysis (exhaustive search) and permutation testing were used to confirm the best interaction models. MDR models were considered statistically significant at P < 0.05 (P sign). Statistical significance of each particular model was then re-evaluated using a 1.000-fold permutation test to compare observed testing balanced accuracies with those expected under the null hypothesis of no association (using the MDR permutation testing module 0.4.9 alpha). Interactions were visualized by constructing an interaction dendrogram according to the method described by Moore et al. (Moore, 2004). MDR software and MDR permutation testing module are open-source and freely available from http://www.epistasis.org.
Results
In this population-based case-control study a total of 834 hematological patients were recruited. Demographic and clinical characteristics of these patients are summarized in Table 2. IA and non-IA patients had similar age but male patients were more prone to develop IA than females (male/female ratio = 1.86 vs. 1.18, P = 0.015). HSCT was common and equally distributed between IA and non-IA groups (45.85 vs. 45.49%) and the underlying disease (hematological disorder) was also uniform between both groups (Table 2). As expected, patients without prophylaxis were more prone to develop IA compared to those patients who take antifungal drugs (23.28 vs. 14.51%; Table 2).
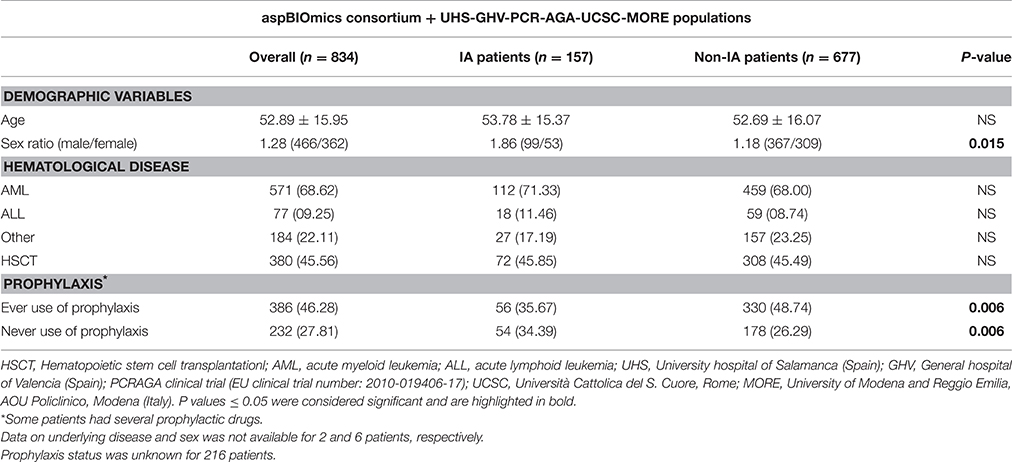
Table 2. Demographic and clinical characteristic of IA and non-IA patients.
All SNPs were in Hardy-Weinberg in the control group with the exception of the RELrs13031237 that was excluded from further analyses (non-IA; P < 0.001). Allele and genotype frequencies of selected SNPs were in line with those reported in Hapmap. Logistic regression analysis adjusted for age, sex, country of origin, allo-HSCT, and underlying disease showed that none of the selected SNPs was significantly associated with the risk of IA according to codominant, dominant, recessive and log-additive models of inheritance (Table 3). In addition, although prophylaxis status was only partially available in our population (n = 618), no significant changes in association estimates were observed when this clinical variable was included as covariate for adjustment (data not shown). No significant associations between the NFκB1, NFκB2, cRel, RelB, and IRF4 polymorphisms and IA risk were also found in an allo-HSCT-restricted analysis considering donor genotypes and IA episodes occurred after transplantation (Table 4). Although our statistical power was limited (Supplementary Table 1), we found a fairly weak association of the IRF4rs12203592T/T genotype with the risk of developing the infection at the nominal significance level of P < 0.05. Carriers of the IRF4rs12203592T/T genotype showed a six-fold increased risk of developing IA when compared with patients carrying the wild type C-allele (ORREC = 6.24, 95%CI 1.25–31.2, P = 0.026). Interestingly, we found that the presence of the minor allele of the rs12203592 (T) strongly correlated with IRF4 mRNA expression levels in whole blood samples and Epstein-Barr virus (EBV)-transformed lymphocytes (P = 6.0 • 10−7 and P = 3.0 • 10−7, respectively), which suggested a possible functional role of this polymorphism. Based on these interesting results, we decided to explore the potential functional impact of this polymorphism using Haploreg and ENCODE annotation data. This analysis revealed that the IRF4rs12203592 SNP resides on a strong enhancer and near of an active promoter that might constitute a regulatory element for IRF4. In addition, this analysis showed that this intronic polymorphism was predicted to change binding motifs for NFκB, HDAC2, and HMG-IY, which are proteins implicated in the transcriptional regulation of multiple innate and adaptive immune-related genes (Liu et al., 2001; Hayden et al., 2006; Falkenberg and Johnstone, 2014). In line with the findings suggesting a functional role of this variant, we also found that the rs12203592 SNP mapped among promoter and enhancer histone marks in different primary T-cell subtypes (TCD8+, TCD4+, Treg, and Th17) of different origin (peripheral blood, hematopoietic stem cells, cord blood, etc.) but also in primary B-cells, natural killer cells and monocytes. Although at this point it was tempting to speculate that the IRF4rs12203592 SNP might play a role in modulating the risk of developing IA likely through the regulation of IRF4 mRNA expression levels, the association of this SNP with IA risk did not remained significant after correction for multiple testing (P = 0.001; Table 4) and, therefore, requires further confirmation. In agreement with the single-SNP analysis, haplotype association analysis showed fairly weak associations of the IRF4AATC and IRF4GGTC haplotypes (not including the IRF4rs12203592T risk allele) with a decreased risk of IA. However, these associations were of similar magnitude to the one observed for the IRF4rs12203592 SNP in the single-SNP analysis (OR = 0.28, 95%CI 0.08–0.95, P = 0.042 and OR = 0.04, 95%CI 0.00–0.71; Table 5) suggesting that the haplotypic effect was due to this intronic variant.
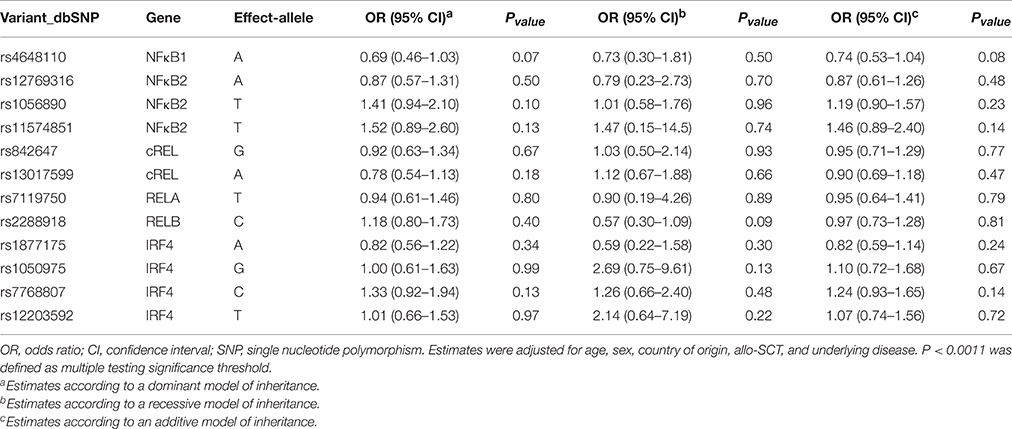
Table 3. Association estimates for NFκB-related polymorphisms and IA.
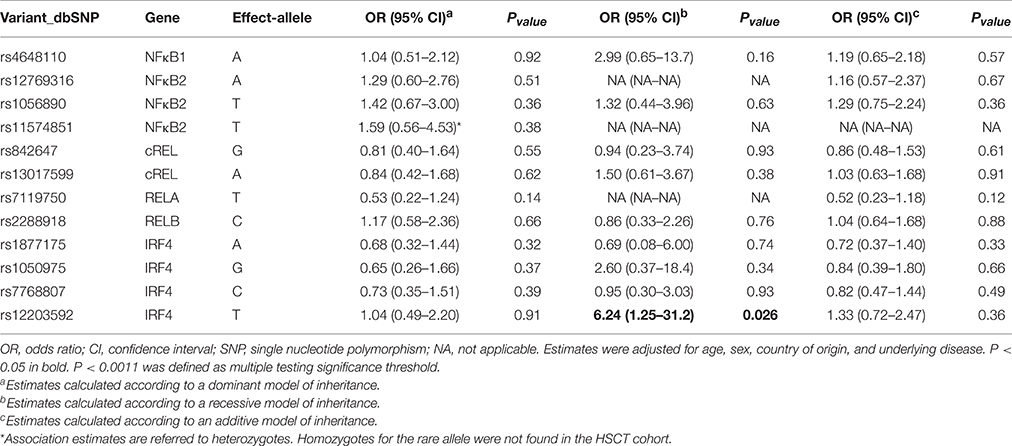
Table 4. Association estimates for NFκB-related polymorphisms and IA in HSCT patients (n = 239).
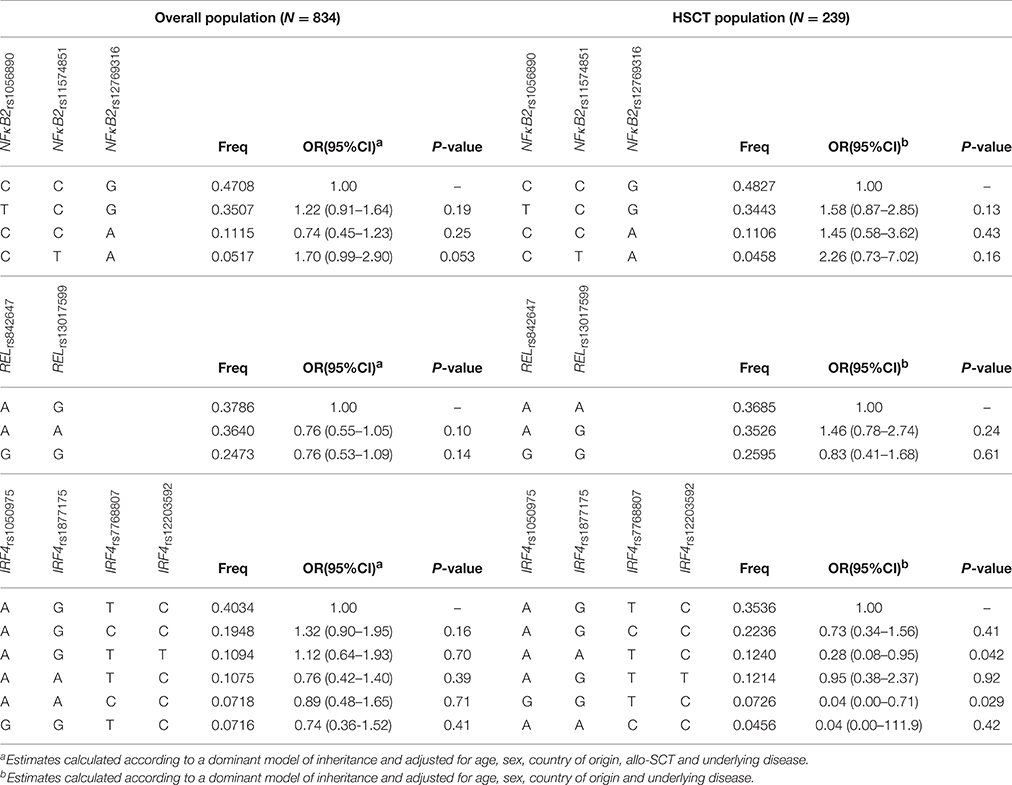
Table 5. Haplotype association analysis and risk of IA.
Finally, given the involvement of physiological complexes (p50/p65, p52/RelB, and p65-c-Rel) in the activation of the NFκB signaling pathway and the role of the p52/RelB complex in the transcriptional activation of the IRF4 (Boddicker et al., 2015), we also decided to investigate whether NFκB-related SNPs might interact to each other to modify the risk of IA. Results of the MDR analysis evaluating all possible combinations among the NFκB-related SNPs are shown in Table 6 and Figure 1. The best model suggested an interaction between the RELrs842647, NFκB2rs1056890, IRF4rs7768807, and RELrs13017599 SNPs to synergistically increase the risk of developing IA (TA = 0.5488, PSign = 0.0010). Although this model was the best to predict IA and presented a high cross validation consistency (CVC) (9/10), it did not remain statistically significant following 1000-fold permutation test (P = 0.31). In addition, we found a significant 2-locus interaction model including the NFκB2rs1056890 and IRF4rs7768807 SNPs to increase the risk of getting the infection (TA = 0.5353, PSign = 0.0010) but this interaction model also failed to retain statistical significance after 1000-fold permutation test (P = 0.47; Table 6). None of the best models included the IRF4rs12203592 SNP.
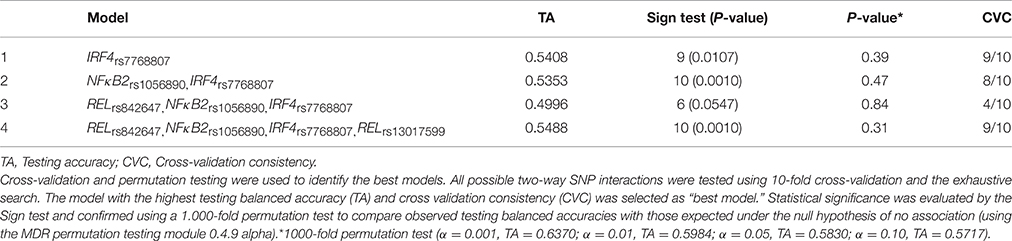
Table 6. Multifactor dimensionality reduction analysis summary.
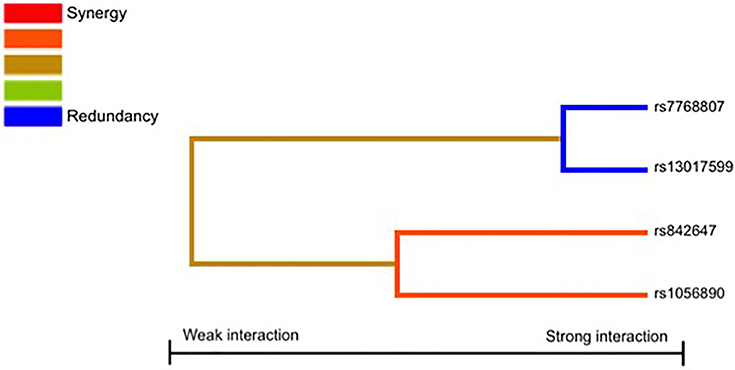
Figure 1. Interaction dendrogram. The interaction dendrogram reveals no significant interactions among the studied SNPs to modulate the risk of IA.
Discussion
The NFκB pathway is implicated in fostering a wide variety of physiological processes such as immune cell turnover (Hayden et al., 2006), inflammation (Karin and Greten, 2005), T-cell differentiation (Th1, Th2, Th17, and Treg subsets; Oh and Ghosh, 2013), DC maturation (Burkly et al., 1995; Abe et al., 2003), cell apoptosis (Wang et al., 1998; Karin and Greten, 2005), cell adhesion (Chen et al., 1995; Lockyer et al., 1998; Chiang et al., 2008), and angiogenesis (Beinke and Ley, 2004). In the presence of fungal pathogens such as Aspergillus fumigatus, the canonical NFκB pathway involving NEMO-IKKα-IKKβ and NFκB-p50/p65 complexes can be activated by membrane pattern recognition receptors (PRRs) such as toll-like receptors (TLRs; Mambula et al., 2002; Bellocchio et al., 2004; Roeder et al., 2004), C-type lectin receptors (CLRs; Serrano-Gómez et al., 2004; LeibundGut-Landmann et al., 2007; Gringhuis et al., 2009; Rivera et al., 2011; Saijo and Iwakura, 2011), cytokine receptors (IL1R, TNFRs) (Bozza et al., 2014), or soluble PRRs like pentraxins (PTX3; Garlanda et al., 2002) that leads to the immediate secretion of pro-inflammatory cytokines (TNF, IL1α, IL1β, IL6, IL12; Nicholson et al., 1996; Roeder et al., 2004), chemokines (CCL2, CCL3, CXCL2 and CXCL10; Shahan et al., 1998; Phadke and Mehrad, 2005; Reid et al., 2009), and reactive oxygen species (ROS; superoxide, hydroxyl radical, nitric oxide; Philippe et al., 2003; Chiang et al., 2008) but also to the expression of specific immune receptors and cell surface adhesion molecules (VCAM-1, E-Selectin; Akira et al., 2001; Chiang et al., 2008; Gringhuis et al., 2009). In addition, binding of fungal compounds to C-type lectins (Dectin-1 and Dectin-2) and the activation of certain TNF family cytokines (CD40L, BAFF, or LT-β) observed during fungal infections leads to the activation of the non-canonical NFκB pathway (p52/RelB), which is implicated in B-cell and DC maturation (Hu et al., 2011; Sun, 2012) and γδTh17 cell development (Powolny-Budnicka et al., 2011) but also in the control of the expression of a wide range of immune-related genes (Geijtenbeek and Gringhuis, 2009; Vallabhapurapu and Karin, 2009; Plato et al., 2013). On the other hand, it has recently been suggested the existence of a CD30-p52/Relb-IRF4 loop to regulate the NFκB pathway and cell proliferation (Boddicker et al., 2015) and a role of IRF4 in modulating the differentiation of different DC (Bajaña et al., 2016) and Th17-mediated immune responses against A. fumigatus (Schlitzer et al., 2013).
Considering the central role of the PRRs-NFκB pathways and IRF4 in the immune responses against A. fumigatus (Oh and Ghosh, 2013; Schlitzer et al., 2013; Williams et al., 2013) and those studies suggesting that genetic host factors may account for differences in susceptibility to IA (Ok et al., 2011; van der Velden et al., 2011), we decided to investigate the link between genetic polymorphisms within NFκB1, NFκB2, RelA, RelB, Rel, and IRF4 genes and the risk of IA. In spite of the growing number of studies assessing the role of genetic polymorphisms within PRRs (TLR2, TLR4, TLR9, Dectin-1, Dectin-2, DC-SIGN, PTX3; Kesh et al., 2005; Bochud et al., 2008; Pamer, 2008; Cunha et al., 2010, 2011, 2014; Chai et al., 2011; Sainz et al., 2012; Grube et al., 2013; Wojtowicz et al., 2015b), cytokines (IL1, IL10, IFNG; Sainz et al., 2007a, 2008a; Lupiañez et al., 2015), and their receptors (IL4R, TNFR1, and TNFR2; Sainz et al., 2007b, 2010; Lupiañez et al., 2015) in determining the susceptibility to invasive fungal infections, this is the first study that attempts to evaluate the impact of polymorphisms within NFκB1, NFκB2, RelA, RelB, REL, and IRF4 genes on the risk of developing IA. Our data showed no significant overall associations between selected SNPs and IA infection. The best result was the association of the IRF4rs12203592 SNP with an increased risk of IA in HSCT patients that suggested a weak effect of this locus on the risk of IA that might become evident only in those patients with a more profound degree of immunosuppression. However, despite the potential interest of these results and the evidences supporting a functional role of the IRF4rs12203592 variant in regulating IRF4 mRNA expression and thereby modulating the NFκB pathway (Boddicker et al., 2015) and IRF4-dependent immune responses (Schlitzer et al., 2013; Bajaña et al., 2016), the association of this variant did not remain significant at the experiment-wide significance threshold, suggesting that the association of this polymorphism with IA could be due to chance. Haplotype analysis showed a small effect of common haplotypes on IA risk but the magnitude of these effects suggested that the observed association was likely due to the IRF4rs12203592 SNP rather than the combined effect of the IRF4 SNPs. In addition, when we evaluated whether there was any significant interaction among the SNPs analyzed, we did not observe any consistent interaction model that could affect the risk of developing IA.
In conclusion, this case-control study does not provide strong evidence of a relationship between polymorphisms within NFκB1, NFκB2, cRel, RelB, and IRF4 genes and IA risk. Nonetheless, given the limited statistical power of the HSCT-stratified analysis (80% to detect odds ratio of 2.1 at α = 0.001 for a SNP with a frequency of 0.25, dominant model) and the evidences suggesting a functional role of the IRF4rs12203592 SNP, we cannot dismiss the possibility of a small but still real effect of this variant or its haplotypes on the risk of IA in HSCT patients. Future case-control population-based studies conducted in larger HSCT populations are now warranted to further evaluate whether the IRF4 locus may have a role in determining the susceptibility to IA.
Ethics Statement
The study protocol was approved by the local ethics review boards of all participating centers and written informed consent was obtained from each patient before inclusion in accordance with the Declaration of Helsinki. Ethical approval for this study was provided by the Comité de Ética e Investigación Clinica (CEIC) of the Virgen de las Nieves Hospital (Granada), University Hospital of Salamanca (Salamanca), Clinic University Hospital of Valencia (Valencia), and the Centro Nacional de Microbiologia (27_2012). The PCRAGA trial is registered with ClinicalTrials.gov (NCT01742026) and EudraCT (2010-019406-17). Ethical approval was also provided by the Subcomissao de Etica para as Ciencias da Vida e Saude (SECVS), University of Minho (approval SECVS 125/2014), (23533/16), the Comitato Etico Provinciale di Modena, the Ethics Committee of the Medical Faculty of the University of Wuerzburg (42/06) and the Ethic votum of the Medical University Innsbruck (UN4529).
Author Contributions
MJ and JS conceived the study and participated in its design and coordination. CBL performed the genetic analyses. TV, AC, JSp, ML, JMS-M, LMC, CC, JS-C, LA-F, CS, LF, LP, LPo, JMA, MLu, MC-E, CL-F, HE, LV, RR-T, JL, MJ and PCRAGA Study Group coordinated patient's recruitment and provided the clinical data. JS analysed the data. MJ and JS drafted the manuscript. All authors read and approved the final version of the manuscript.
Funding
This study was supported by grants PI12/02688 from Fondo de Investigaciones Sanitarias (Instituto de Salud Carlos III, Madrid, Spain), the ERA-NET PathoGenoMics (03159000A; Ministerio de Ciencia e Innovación PIM2010EPA-00756, Madrid, Spain), the Collaborative Research Center / Transregio 124 FungiNet, the Austrian Science Fundation (FWF I-656-B09), the Fundação para a Ciência e Tecnologia (FCT), cofunded by Programa Operacional Regional do Norte (ON.2—O Novo Norte), the Quadro de Referência Estratégico Nacional (QREN) through the Fundo Europeu de Desenvolvimento Regional (FEDER) and the Projeto Estratégico – LA 26 – 2013–2014 (PEst-C/SAU/LA0026/2013). Agostinho Carvalho and Cristina Cunha were supported by the Fundação para a Ciência e Tecnologia (FCT), Portugal (IF/00735/2014 and SFRH/BPD/96176/2013, respectively). The PCRAGA trial was supported by an unrestricted grant from Pfizer, which had no involvement or control over the collection, analysis, and interpretation of data; the writing of the report; or the decision to submit the paper for publication. This study was also supported by Astellas Pharma Inc. and a donation from Consuelo González Moreno, an acute myeloid leukemia survivor.
Conflict of Interest Statement
Dr. MC has been an advisor/consultant to the Panamerican Health Organization, Astellas Pharma, Gilead Sciences, Merck Sharp & Dohme, Pfizer, and Schering Plough. He has been paid for talks on behalf of Gilead Sciences, Merck Sharp & Dohme, Pfizer, Astellas Pharma, and Schering Plough.
The other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01243
Supplementary Table 1. Statistical power calculations for NFκB-related polymorphisms. SNP, Single Nucleotide Polymorphism; MAF, Aa, Aminoacid; ND, not determined; NP, no statistical power. Assumptions: IA incidence of 3.5% (Morgan et al. Med Mycol 2005; 43:S49-S58) and an α = 0.001 (study-wide significance). *The statistical power was calculated based on genotype frequencies and assuming a dominant model of inheritance. *The statistical power was calculated based on genotype frequencies and assuming a recessive model of inheritance.
Supplementary Figure 1. Linkage disequilibrium (LD) blocks in the NFκB-related genes calculated in our population. Numbers into squares indicate r2 values.
References
Abe, K., Yarovinsky, F. O., Murakami, T., Shakhov, A. N., Tumanov, A. V., Ito, D., et al. (2003). Distinct contributions of TNF and LT cytokines to the development of dendritic cells in vitro and their recruitment in vivo. Blood 101, 1477–1483. doi: 10.1182/blood.V101.4.1477
Aguado, J. M., Vázquez, L., Fernandez-Ruiz, M., Villaescusa, T., Ruiz-Camps, I., Barba, P., et al. (2015). Serum galactomannan versus a combination of galactomannan and polymerase chain reaction-based Aspergillus DNA detection for early therapy of invasive aspergillosis in high-risk hematological patients: a randomized controlled trial. Clin. Infect. Dis. 60, 405–414. doi: 10.1093/cid/ciu833
Akira, S., Takeda, K., and Kaisho, T. (2001). Toll-like receptors: critical proteins linking innate and acquired immunity. Nat. Immunol. 2, 675–680. doi: 10.1038/90609
Ali, F. R., Barton, A., Smith, R. L., Bowes, J., Flynn, E., Mangino, M., et al. (2013). An investigation of rheumatoid arthritis loci in patients with early-onset psoriasis validates association of the REL gene. Br. J. Dermatol. 168, 864–866. doi: 10.1111/bjd.12106
Bajaña, S., Turner, S., Paul, J., Ainsua-Enrich, E., and Kovats, S. (2016). IRF4 and IRF8 Act in CD11c+ cells to regulate terminal differentiation of lung tissue dendritic cells. J. Immunol. 196, 1666–1677. doi: 10.4049/jimmunol.1501870
Beinke, S., and Ley, S. C. (2004). Functions of NF-κB1 and NF-κB2 in immune cell biology. Biochem. J. 382(Pt 2), 393–409. doi: 10.1042/BJ20040544
Bellocchio, S., Moretti, S., Perruccio, K., Fallarino, F., Bozza, S., Montagnoli, C., et al. (2004). TLRs govern neutrophil activity in aspergillosis. J. Immunol. 173, 7406–7415. doi: 10.4049/jimmunol.173.12.7406
Bochud, P. Y., Chien, J. W., Marr, K. A., Leisenring, W. M., Upton, A., Janer, M., et al. (2008). Toll-like receptor 4 polymorphisms and aspergillosis in stem-cell transplantation. N. Engl. J. Med. 359, 1766–1777. doi: 10.1056/NEJMoa0802629
Boddicker, R. L., Kip, N. S., Xing, X., Zeng, Y., Yang, Z. Z., Lee, J. H., et al. (2015). The oncogenic transcription factor IRF4 is regulated by a novel CD30/NF-κB positive feedback loop in peripheral T-cell lymphoma. Blood 125, 3118–3127. doi: 10.1182/blood-2014-05-578575
Bowes, J., Ho, P., Flynn, E., Ali, F., Marzo-Ortega, H., Coates, L. C., et al. (2012). Comprehensive assessment of rheumatoid arthritis susceptibility loci in a large psoriatic arthritis cohort. Ann. Rheum. Dis. 71, 1350–1354. doi: 10.1136/annrheumdis-2011-200802
Bozza, S., Campo, S., Arseni, B., Inforzato, A., Ragnar, L., Bottazzi, B., et al. (2014). PTX3 binds MD-2 and promotes TRIF-dependent immune protection in aspergillosis. J. Immunol. 193, 2340–2348. doi: 10.4049/jimmunol.1400814
Burkly, L., Hession, C., Ogata, L., Reilly, C., Marconi, L. A., Olson, D., et al. (1995). Expression of relB is required for the development of thymic medulla and dendritic cells. Nature 373, 531–536. doi: 10.1038/373531a0
Carvalho, A., De Luca, A., Bozza, S., Cunha, C., D'Angelo, C., Moretti, S., et al. (2012). TLR3 essentially promotes protective class I-restricted memory CD8(+) T-cell responses to Aspergillus fumigatus in hematopoietic transplanted patients. Blood 119, 967–977. doi: 10.1182/blood-2011-06-362582
Chai, L. Y., de Boer, M. G., van der Velden, W. J., Plantinga, T. S., van Spriel, A. B., Jacobs, C., et al. (2011). The Y238X stop codon polymorphism in the human β-glucan receptor dectin-1 and susceptibility to invasive aspergillosis. J. Infect. Dis. 203, 736–743. doi: 10.1093/infdis/jiq102
Chen, B., Wilkening, S., Drechsel, M., and Hemminki, K. (2009). SNP_tools: A compact tool package for analysis and conversion of genotype data for MS-Excel. BMC Res. Notes 2, 214. doi: 10.1186/1756-0500-2-214
Chen, C. C., Rosenbloom, C. L., Anderson, D. C., and Manning, A. M. (1995). Selective inhibition of E-selectin, vascular cell adhesion molecule-1, and intercellular adhesion molecule-1 expression by inhibitors of I κ B-α phosphorylation. J. Immunol. 155, 3538–3545.
Chen, F., Xu, L., Zhao, T., Xiao, X., Pan, Y., and Hou, S. (2016). Genetic variation in the REL gene increases risk of Behcet's Disease in a Chinese han population but that of PRKCQ does not. PLoS ONE 11:e0147350. doi: 10.1371/journal.pone.0147350
Chen, W. J., Yang, J. Y., Lin, J. H., Fann, C. S., Osyetrov, V., King, C. C., et al. (2006). Nasopharyngeal shedding of severe acute respiratory syndrome-associated coronavirus is associated with genetic polymorphisms. Clin. Infect. Dis. 42, 1561–1569. doi: 10.1086/503843
Chiang, L. Y., Sheppard, D. C., Gravelat, F. N., Patterson, T. F., and Filler, S. G. (2008). Aspergillus fumigatus stimulates leukocyte adhesion molecules and cytokine production by endothelial cells in vitro and during invasive pulmonary disease. Infect. Immun. 76, 3429–3438. doi: 10.1128/IAI.01510-07
Cunha, C., Aversa, F., Lacerda, J. F., Busca, A., Kurzai, O., Grube, M., et al. (2014). Genetic PTX3 deficiency and aspergillosis in stem-cell transplantation. N. Engl. J. Med. 370, 421–432. doi: 10.1056/NEJMoa1211161
Cunha, C., Di Ianni, M., Bozza, S., Giovannini, G., Zagarella, S., Zelante, T., et al. (2010). Dectin-1 Y238X polymorphism associates with susceptibility to invasive aspergillosis in hematopoietic transplantation through impairment of both recipient- and donor-dependent mechanisms of antifungal immunity. Blood 116, 5394–5402. doi: 10.1182/blood-2010-04-279307
Cunha, C., Giovannini, G., Pierini, A., Bell, A. S., Sorci, G., Riuzzi, F., et al. (2011). Genetically-determined hyperfunction of the S100B/RAGE axis is a risk factor for aspergillosis in stem cell transplant recipients. PLoS ONE 6:e27962. doi: 10.1371/journal.pone.0027962
Curtin, K., Wolff, R. K., Herrick, J. S., Abo, R., and Slattery, M. L. (2010). Exploring multilocus associations of inflammation genes and colorectal cancer risk using hapConstructor. BMC Med. Genet. 11:170. doi: 10.1186/1471-2350-11-170
De Pauw, B., Walsh, T. J., Donnelly, J. P., Stevens, D. A., Edwards, J. E., Calandra, T., et al. (2008). Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) Consensus Group. Clin. Infect. Dis. 46, 1813–1821. doi: 10.1086/588660
Do, T. N., Ucisik-Akkaya, E., Davis, C. F., Morrison, B. A., and Dorak, M. T. (2010). An intronic polymorphism of IRF4 gene influences gene transcription in vitro and shows a risk association with childhood acute lymphoblastic leukemia in males. Biochim. Biophys. Acta 1802, 292–300. doi: 10.1016/j.bbadis.2009.10.015
Ellinghaus, E., Stuart, P. E., Ellinghaus, D., Nair, R. P., Debrus, S., Raelson, J. V., et al. (2012). Genome-wide meta-analysis of psoriatic arthritis identifies susceptibility locus at REL. J. Invest. Dermatol. 132, 1133–1140. doi: 10.1038/jid.2011.415
Eyre, S., Hinks, A., Flynn, E., Martin, P., Wilson, A. G., Maxwell, J. R., et al. (2010). Confirmation of association of the REL locus with rheumatoid arthritis susceptibility in the UK population. Ann. Rheum. Dis. 69, 1572–1573. doi: 10.1136/ard.2009.122887
Falkenberg, K. J., and Johnstone, R. W. (2014). Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat. Rev. Drug Discov. 13, 673–691. doi: 10.1038/nrd4360
Garlanda, C., Hirsch, E., Bozza, S., Salustri, A., De Acetis, M., Nota, R., et al. (2002). Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 420, 182–186. doi: 10.1038/nature01195
Geijtenbeek, T. B., and Gringhuis, S. I. (2009). Signalling through C-type lectin receptors: shaping immune responses. Nat. Rev. Immunol. 9, 465–479. doi: 10.1038/nri2569
Gilmore, T. D. (2006). Introduction to NF-κB: players, pathways, perspectives. Oncogene 25, 6680–6684. doi: 10.1038/sj.onc.1209954
Gregersen, P. K., Amos, C. I., Lee, A. T., Lu, Y., Remmers, E. F., Kastner, D. L., et al. (2009). REL, encoding a member of the NF-κB family of transcription factors, is a newly defined risk locus for rheumatoid arthritis. Nat. Genet. 41, 820–823. doi: 10.1038/ng.395
Gringhuis, S. I., den Dunnen, J., Litjens, M., van der Vlist, M., Wevers, B., Bruijns, S. C., et al. (2009). Dectin-1 directs T helper cell differentiation by controlling noncanonical NF-κB activation through Raf-1 and Syk. Nat. Immunol. 10, 203–213. doi: 10.1038/ni.1692
Grube, M., Loeffler, J., Mezger, M., Krüger, B., Echtenacher, B., Hoffmann, P., et al. (2013). TLR5 stop codon polymorphism is associated with invasive aspergillosis after allogeneic stem cell transplantation. Med. Mycol. 51, 818–825. doi: 10.3109/13693786.2013.809630
Han, J., Qureshi, A. A., Nan, H., Zhang, J., Song, Y., Guo, Q., et al. (2011). A germline variant in the interferon regulatory factor 4 gene as a novel skin cancer risk locus. Cancer Res. 71, 1533–1539. doi: 10.1158/0008-5472.CAN-10-1818
Hayden, M. S., West, A. P., and Ghosh, S. (2006). NF-κB and the immune response. Oncogene 25, 6758–6780. doi: 10.1038/sj.onc.1209943
Hoffmann, A., Natoli, G., and Ghosh, G. (2006). Transcriptional regulation via the NF-κB signaling module. Oncogene 25, 6706–6716. doi: 10.1038/sj.onc.1209933
Hu, H., Wu, X., Jin, W., Chang, M., Cheng, X., and Sun, S. C. (2011). Noncanonical NF-κB regulates inducible costimulator (ICOS) ligand expression and T follicular helper cell development. Proc. Natl. Acad. Sci. U.S.A. 108, 12827–12832. doi: 10.1073/pnas.1105774108
Karin, M., and Greten, F. R. (2005). NF-κB: linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 5, 749–759. doi: 10.1038/nri1703
Karthaus, M., and Buchheidt, D. (2013). Invasive aspergillosis: new insights into disease, diagnostic and treatment. Curr. Pharm. Des. 19, 3569–3594. doi: 10.2174/13816128113199990330
Kesh, S., Mensah, N. Y., Peterlongo, P., Jaffe, D., Hsu, K., Van den Brink, M., et al. (2005). TLR1 and TLR6 polymorphisms are associated with susceptibility to invasive aspergillosis after allogeneic stem cell transplantation. Ann. N. Y. Acad. Sci. 1062, 95–103. doi: 10.1196/annals.1358.012
Kousha, M., Tadi, R., and Soubani, A. O. (2011). Pulmonary aspergillosis: a clinical review. Eur. Respir. Rev. 20, 156–174. doi: 10.1183/09059180.00001011
LeibundGut-Landmann, S., Gross, O., Robinson, M. J., Osorio, F., Slack, E. C., Tsoni, S. V., et al. (2007). Syk- and CARD9-dependent coupling of innate immunity to the induction of T helper cells that produce interleukin 17. Nat. Immunol. 8, 630–638. doi: 10.1038/ni1460
Leung, G., Baggott, C., West, C., Elboim, C., Paul, S. M., Cooper, B. A., et al. (2014). Cytokine candidate genes predict the development of secondary lymphedema following breast cancer surgery. Lymphat. Res. Biol. 12, 10–22. doi: 10.1089/lrb.2013.0024
Liu, F., Chau, K. Y., Arlotta, P., and Ono, S. J. (2001). The HMG I proteins: dynamic roles in gene activation, development, and tumorigenesis. Immunol. Res. 24, 13–29. doi: 10.1385/IR:24:1:13
Lockyer, J. M., Colladay, J. S., Alperin-Lea, W. L., Hammond, T., and Buda, A. J. (1998). Inhibition of nuclear factor-κB-mediated adhesion molecule expression in human endothelial cells. Circ. Res. 82, 314–320. doi: 10.1161/01.RES.82.3.314
Lupiañez, C. B., Canet, L. M., Carvalho, A., Alcazar-Fuoli, L., Springer, J., Lackner, M., et al. (2015). Polymorphisms in host immunity-modulating genes and risk of invasive aspergillosis: results from the AspBIOmics Consortium. Infect. Immun. 84, 643–657. doi: 10.1128/IAI.01359-15
Mambula, S. S., Sau, K., Henneke, P., Golenbock, D. T., and Levitz, S. M. (2002). Toll-like receptor (TLR) signaling in response to Aspergillus fumigatus. J. Biol. Chem. 277, 39320–39326. doi: 10.1074/jbc.M201683200
Meersseman, W., Lagrou, K., Maertens, J., and Van Wijngaerden, E. (2007). Invasive aspergillosis in the intensive care unit. Clin. Infect. Dis. 45, 205–216. doi: 10.1086/518852
Mezger, M., Steffens, M., Beyer, M., Manger, C., Eberle, J., Toliat, M. R., et al. (2008). Polymorphisms in the chemokine (C-X-C motif) ligand 10 are associated with invasive aspergillosis after allogeneic stem-cell transplantation and influence CXCL10 expression in monocyte-derived dendritic cells. Blood 111, 534–536. doi: 10.1182/blood-2007-05-090928
Moore, J. H. (2004). Computational analysis of gene-gene interactions using multifactor dimensionality reduction. Expert Rev. Mol. Diagn. 4, 795–803. doi: 10.1586/14737159.4.6.795
Moynagh, P. N. (2005). The NF-κB pathway. J. Cell Sci. 118(Pt 20), 4589–4592. doi: 10.1242/jcs.02579
Neofytos, D., Horn, D., Anaissie, E., Steinbach, W., Olyaei, A., Fishman, J., et al. (2009). Epidemiology and outcome of invasive fungal infection in adult hematopoietic stem cell transplant recipients: analysis of Multicenter Prospective Antifungal Therapy (PATH) Alliance registry. Clin. Infect. Dis. 48, 265–273. doi: 10.1086/595846
Neofytos, D., Lu, K., Hatfield-Seung, A., Blackford, A., Marr, K. A., Treadway, S., et al. (2013a). Epidemiology, outcomes, and risk factors of invasive fungal infections in adult patients with acute myelogenous leukemia after induction chemotherapy. Diagn. Microbiol. Infect. Dis. 75, 144–149. doi: 10.1016/j.diagmicrobio.2012.10.001
Neofytos, D., Treadway, S., Ostrander, D., Alonso, C. D., Dierberg, K. L., Nussenblatt, V., et al. (2013b). Epidemiology, outcomes, and mortality predictors of invasive mold infections among transplant recipients: a 10-year, single-center experience. Transpl. Infect. Dis. 15, 233–242. doi: 10.1111/tid.12060
Nicholson, W. J., Slight, J., and Donaldson, K. (1996). Inhibition of the transcription factors NF-κB and AP-1 underlies loss of cytokine gene expression in rat alveolar macrophages treated with a diffusible product from the spores of Aspergillus fumigatus. Am. J. Respir. Cell Mol. Biol. 15, 88–96. doi: 10.1165/ajrcmb.15.1.8679226
Nyholt, D. R. (2004). A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am. J. Hum. Genet. 74, 765–769. doi: 10.1086/383251
Oh, H., and Ghosh, S. (2013). NF-κB: roles and regulation in different CD4(+) T-cell subsets. Immunol. Rev. 252, 41–51. doi: 10.1111/imr.12033
Ok, M., Einsele, H., and Loeffler, J. (2011). Genetic susceptibility to Aspergillus fumigatus infections. Int. J. Med. Microbiol. 301, 445–452. doi: 10.1016/j.ijmm.2011.04.013
Pamer, E. G. (2008). TLR polymorphisms and the risk of invasive fungal infections. N. Engl. J. Med. 359, 1836–1838. doi: 10.1056/NEJMe0806412
Pan, W., Zhang, A. Q., Gu, W., Gao, J. W., Du, D. Y., Zhang, L. Y., et al. (2015). Identification of haplotype tag single nucleotide polymorphisms within the nuclear factor-κB family genes and their clinical relevance in patients with major trauma. Crit. Care 19, 95. doi: 10.1186/s13054-015-0836-6
Phadke, A. P., and Mehrad, B. (2005). Cytokines in host defense against Aspergillus: recent advances. Med. Mycol. 43 (Suppl. 1), S173–S176. doi: 10.1080/13693780500052099
Philippe, B., Ibrahim-Granet, O., Prévost, M. C., Gougerot-Pocidalo, M. A., Sanchez Perez, M., Van der Meeren, A., et al. (2003). Killing of Aspergillus fumigatus by alveolar macrophages is mediated by reactive oxidant intermediates. Infect. Immun. 71, 3034–3042. doi: 10.1128/IAI.71.6.3034-3042.2003
Plato, A., Willment, J. A., and Brown, G. D. (2013). C-type lectin-like receptors of the dectin-1 cluster: ligands and signaling pathways. Int. Rev. Immunol. 32, 134–156. doi: 10.3109/08830185.2013.777065
Powolny-Budnicka, I., Riemann, M., Tanzer, S., Schmid, R. M., Hehlgans, T., and Weih, F. (2011). RelA and RelB transcription factors in distinct thymocyte populations control lymphotoxin-dependent interleukin-17 production in gammadelta T cells. Immunity 34, 364–374. doi: 10.1016/j.immuni.2011.02.019
Ramaprakash, H., Ito, T., Standiford, T. J., Kunkel, S. L., and Hogaboam, C. M. (2009). Toll-like receptor 9 modulates immune responses to Aspergillus fumigatus conidia in immunodeficient and allergic mice. Infect. Immun. 77, 108–119. doi: 10.1128/IAI.00998-08
Reid, D. M., Gow, N. A., and Brown, G. D. (2009). Pattern recognition: recent insights from Dectin-1. Curr. Opin. Immunol. 21, 30–37. doi: 10.1016/j.coi.2009.01.003
Ritchie, M. D., Hahn, L. W., and Moore, J. H. (2003). Power of multifactor dimensionality reduction for detecting gene-gene interactions in the presence of genotyping error, missing data, phenocopy, and genetic heterogeneity. Genet. Epidemiol. 24, 150–157. doi: 10.1002/gepi.10218
Rivera, A., Hohl, T. M., Collins, N., Leiner, I., Gallegos, A., Saijo, S., et al. (2011). Dectin-1 diversifies Aspergillus fumigatus-specific T cell responses by inhibiting T helper type 1 CD4 T cell differentiation. J. Exp. Med. 208, 369–381. doi: 10.1084/jem.20100906
Roeder, A., Kirschning, C. J., Rupec, R. A., Schaller, M., Weindl, G., and Korting, H. C. (2004). Toll-like receptors as key mediators in innate antifungal immunity. Med. Mycol. 42, 485–498. doi: 10.1080/13693780400011112
Saijo, S., and Iwakura, Y. (2011). Dectin-1 and Dectin-2 in innate immunity against fungi. Int. Immunol. 23, 467–472. doi: 10.1093/intimm/dxr046
Sainz, J., Hassan, L., Perez, E., Romero, A., Moratalla, A., López-Fernández, E., et al. (2007a). Interleukin-10 promoter polymorphism as risk factor to develop invasive pulmonary aspergillosis. Immunol. Lett. 109, 76–82. doi: 10.1016/j.imlet.2007.01.005
Sainz, J., Lupiáñez, C. B., Segura-Catena, J., Vazquez, L., Ríos, R., Oyonarte, S., et al. (2012). Dectin-1 and DC-SIGN polymorphisms associated with invasive pulmonary Aspergillosis infection. PLoS ONE 7:e32273. doi: 10.1371/journal.pone.0032273
Sainz, J., Pérez, E., Gómez-Lopera, S., and Jurado, M. (2008a). IL1 gene cluster polymorphisms and its haplotypes may predict the risk to develop invasive pulmonary aspergillosis and modulate C-reactive protein level. J. Clin. Immunol. 28, 473–485. doi: 10.1007/s10875-008-9197-0
Sainz, J., Perez, E., Gomez-Lopera, S., Lopez-Fernandez, E., Moratalla, L., Oyonarte, S., et al. (2008b). Genetic variants of IL6 gene promoter influence on C-reactive protein levels but are not associated with susceptibility to invasive pulmonary aspergillosis in haematological patients. Cytokine 41, 268–278. doi: 10.1016/j.cyto.2007.11.018
Sainz, J., Pérez, E., Hassan, L., Moratalla, A., Romero, A., Collado, M. D., et al. (2007b). Variable number of tandem repeats of TNF receptor type 2 promoter as genetic biomarker of susceptibility to develop invasive pulmonary aspergillosis. Hum. Immunol. 68, 41–50. doi: 10.1016/j.humimm.2006.10.011
Sainz, J., Salas-Alvarado, I., López-Fernández, E., Olmedo, C., Comino, A., García, F., et al. (2010). TNFR1 mRNA expression level and TNFR1 gene polymorphisms are predictive markers for susceptibility to develop invasive pulmonary aspergillosis. Int. J. Immunopathol. Pharmacol. 23, 423–436. doi: 10.1177/039463201002300205
Schlitzer, A., McGovern, N., Teo, P., Zelante, T., Atarashi, K., Low, D., et al. (2013). IRF4 transcription factor-dependent CD11b+ dendritic cells in human and mouse control mucosal IL-17 cytokine responses. Immunity 38, 970–983. doi: 10.1016/j.immuni.2013.04.011
Serrano-Gómez, D., Dominguez-Soto, A., Ancochea, J., Jimenez-Heffernan, J. A., Leal, J. A., and Corbi, A. L. (2004). Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin mediates binding and internalization of Aspergillus fumigatus conidia by dendritic cells and macrophages. J. Immunol. 173, 5635–5643. doi: 10.4049/jimmunol.173.9.5635
Seufert, B. L., Poole, E. M., Whitton, J., Xiao, L., Makar, K. W., Campbell, P. T., et al. (2013). IkappaBKbeta and NFκB1, NSAID use and risk of colorectal cancer in the Colon Cancer Family Registry. Carcinogenesis 34, 79–85. doi: 10.1093/carcin/bgs296
Shahan, T. A., Sorenson, W. G., Paulauskis, J. D., Morey, R., and Lewis, D. M. (1998). Concentration- and time-dependent upregulation and release of the cytokines MIP-2, KC, TNF, and MIP-1alpha in rat alveolar macrophages by fungal spores implicated in airway inflammation. Am. J. Respir. Cell Mol. Biol. 18, 435–440. doi: 10.1165/ajrcmb.18.3.2856
Singh, N., Husain, S., and AST Infectious Diseases Community of Practice. (2013). Aspergillosis in solid organ transplantation. Am. J. Transplant. 13 (Suppl 4), 228–241. doi: 10.1111/ajt.12115
Slattery, M. L., Lundgreen, A., Bondurant, K. L., and Wolff, R. K. (2011). Interferon-signaling pathway: associations with colon and rectal cancer risk and subsequent survival. Carcinogenesis 32, 1660–1667. doi: 10.1093/carcin/bgr189
Stappers, M. H., Thys, Y., Oosting, M., Plantinga, T. S., Ioana, M., Reimnitz, P., et al. (2014). Polymorphisms in cytokine genes IL6, TNF, IL10, IL17A and IFNG influence susceptibility to complicated skin and skin structure infections. Eur. J. Clin. Microbiol. Infect. Dis. 33, 2267–2274. doi: 10.1007/s10096-014-2201-0
Steinbach, W. J., Marr, K. A., Anaissie, E. J., Azie, N., Quan, S. P., Meier-Kriesche, H. U., et al. (2012). Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J. Infect. 65, 453–464. doi: 10.1016/j.jinf.2012.08.003
Sun, S. C. (2012). The noncanonical NF-κB pathway. Immunol. Rev. 246, 125–140. doi: 10.1111/j.1600-065X.2011.01088.x
Trynka, G., Zhernakova, A., Romanos, J., Franke, L., Hunt, K. A., Turner, G., et al. (2009). Coeliac disease-associated risk variants in TNFAIP3 and REL implicate altered NF-κB signalling. Gut 58, 1078–1083. doi: 10.1136/gut.2008.169052
Vallabhapurapu, S., and Karin, M. (2009). Regulation and function of NF-κB transcription factors in the immune system. Annu. Rev. Immunol. 27, 693–733. doi: 10.1146/annurev.immunol.021908.132641
van der Velden, W. J., Blijlevens, N. M., and Donnelly, J. P. (2011). Genetic variants and the risk for invasive mould disease in immunocompromised hematology patients. Curr. Opin. Infect. Dis. 24, 554–563. doi: 10.1097/QCO.0b013e32834ab1f4
Varadé, J., Palomino-Morales, R., Ortego-Centeno, N., Díaz-Rubio, M., Fernandez-Gutierrez, B., Gonzalez-Gay, M. A., et al. (2011). Analysis of the REL polymorphism rs13031237 in autoimmune diseases. Ann. Rheum. Dis. 70, 711–712. doi: 10.1136/ard.2010.134593
Wang, C. Y., Mayo, M. W., Korneluk, R. G., Goeddel, D. V., and Baldwin, A. S. Jr. (1998). NF-κB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science 281, 1680–1683. doi: 10.1126/science.281.5383.1680
Wang, S., Yan, Q., Chen, P., Zhao, P., and Gu, A. (2014). Association of interferon regulatory factor 4 gene polymorphisms rs12203592 and rs872071 with skin cancer and haematological malignancies susceptibility: a meta-analysis of 19 case-control studies. BMC Cancer 14:410. doi: 10.1186/1471-2407-14-410
Williams, J. W., Tjota, M. Y., Clay, B. S., Vander Lugt, B., Bandukwala, H. S., Hrusch, C. L., et al. (2013). Transcription factor IRF4 drives dendritic cells to promote Th2 differentiation. Nat. Commun. 4:2990. doi: 10.1038/ncomms3990
Wójtowicz, A., Gresnigt, M. S., Lecompte, T., Bibert, S., Manuel, O., Joosten, L. A., et al. (2015a). IL1B and DEFB1 polymorphisms increase susceptibility to invasive mold infection after solid-organ transplantation. J. Infect. Dis. 211, 1646–1657. doi: 10.1093/infdis/jiu636
Keywords: Invasive Aspergillosis, genetic polymorphisms, susceptibility, NFκB-related genes, interaction
Citation: Lupiañez CB, Villaescusa MT, Carvalho A, Springer J, Lackner M, Sánchez-Maldonado JM, Canet LM, Cunha C, Segura-Catena J, Alcazar-Fuoli L, Solano C, Fianchi L, Pagano L, Potenza L, Aguado JM, Luppi M, Cuenca-Estrella M, Lass-Flörl C, Einsele H, Vázquez L, PCRAGA Study Group, Ríos-Tamayo R, Loeffler J, Jurado M and Sainz J (2016) Common Genetic Polymorphisms within NFκB-Related Genes and the Risk of Developing Invasive Aspergillosis. Front. Microbiol. 7:1243. doi: 10.3389/fmicb.2016.01243
Received: 14 April 2016; Accepted: 26 July 2016;
Published: 12 August 2016.
Edited by:
Frederic Lamoth, Centre Hospitalier Universitaire Vaudois, SwitzerlandReviewed by:
William J. Steinbach, Duke University, USAPierre-Yves Bochud, Centre Hospitalier Universitaire Vaudois, Switzerland
Copyright © 2016 Lupiañez, Villaescusa, Carvalho, Springer, Lackner, Sánchez-Maldonado, Canet, Cunha, Segura-Catena, Alcazar-Fuoli, Solano, Fianchi, Pagano, Potenza, Aguado, Luppi, Cuenca-Estrella, Lass-Flörl, Einsele, Vázquez, PCRAGA Study Group, Ríos-Tamayo, Loeffler, Jurado and Sainz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan Sainz, juan.sainz@genyo.es