 Lin Zhu1,2†
Lin Zhu1,2† Anouk Zancarini2†
Anouk Zancarini2† Imen Louati2†
Imen Louati2† Silvia De Cesare2
Silvia De Cesare2 Charlotte Duval3
Charlotte Duval3 Kevin Tambosco2
Kevin Tambosco2 Cécile Bernard3
Cécile Bernard3 Didier Debroas4
Didier Debroas4 Lirong Song1
Lirong Song1 Julie Leloup2
Julie Leloup2 Jean-François Humbert2*
Jean-François Humbert2*- 1State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China
- 2UPMC-INRA, iEES-Paris UMR 7618, Paris, France
- 3CNRS, MCAM, Muséum National d’Histoire Naturelle, UMR 7245, Sorbonne Universités, Paris, France
- 4Laboratoire Microorganismes: Génome et Environnement, Université de Clermont-Ferrand, Clermont-Ferrand, France
To overcome the limitations associated with studying the interactions between bacterial communities (BCs) and cyanobacteria in natural environments, we compared the structural and functional diversities of the BCs associated with 15 non-axenic cyanobacterial strains in culture and two natural BCs sampled during cyanobacterial blooms. No significant differences in richness and diversity were found between the natural and cultivated BCs, although some of the cyanobacterial strains had been isolated 11 years earlier. Moreover, these BCs shared some similar characteristics, such as a very low abundance of Actinobacteria, but they display significant differences at the operational taxonomic unit (OTU) level. Overall, our findings suggest that BCs associated with cyanobacteria in culture are good models to better understand the interactions between heterotrophic bacteria and cyanobacteria. Additionally, BCs associated with heterocystous cyanobacterial strains cultivated in Z8X culture medium without nitrate (Aphanizomenon–Dolichospermum) demonstrated significant differences compared to BCs associated with non-heterocystous strains cultivated in Z8 culture medium (Planktothrix–Microcystis) in terms of their composition and their ability to utilize different carbon sources, suggesting the potential influence of cyanobacterial metabolism and/or culture media on associated BCs. Finally, half of the dominant OTUs in these BCs were specifically associated with cyanobacteria or other phytoplankton, whereas the remaining OTUs were generally associated with ecosystems containing high organic matter content, such as sludge or intestines.
Introduction
In the last 10 years, a growing number of published studies have investigated the bacterial communities (BCs) associated with phytoplankton. In marine ecosystems, a limited number of heterotrophic bacterial lineages displaying distinct metabolic strategies dominate these BCs (see review of Buchan et al., 2014). In freshwater ecosystems, the majority of studies have focused on cyanobacterial blooms due to their potential impact on human health (e.g., Dodds et al., 2009). For example, it has been reported that epiphytic bacteria and archaea are directly associated with cyanobacteria (e.g., Stevenson and Waterbury, 2006; Grossart et al., 2011; Woodhouse et al., 2013), and that several bacterial species degrade cyanotoxins to use them as carbon or nitrogen sources (e.g., Maruyama et al., 2004; Zhu et al., 2014). Several publications have also evaluated the impact of cyanobacterial blooms on the structure and composition of BCs within the same ecosystems (e.g., Eiler and Bertilsson, 2004; Berg et al., 2009; Louati et al., 2015; Woodhouse et al., 2016). Microcystis sp., a toxic bloom-forming cyanobacteria, has been a specific focus given its high prevalence in many ecosystems around the world as well as its colonial organization, which offers a potential niche and physical support for direct interactions with heterotrophic prokaryotes (e.g., Shen et al., 2011; Li et al., 2012; Shi et al., 2012; Shao et al., 2014).
Most studies are performed on natural samples and have produced interesting findings. For example, BCs associated with cyanobacteria are distinct from those that are free-living, implying direct interactions between cyanobacteria and bacteria (Cai et al., 2013; Parveen et al., 2013). However, the impact of numerous environmental factors and processes on BCs in natural environments (see Dziallas and Grossart, 2011) makes it difficult to separate the influence of cyanobacteria from that of the environment. To overcome these constraints, we compared the functional and structural diversities of BCs associated with 15 non-axenic cyanobacterial strains belonging to four genera: Microcystis, Planktothrix, Aphanizomenon, and Dolichospermum.
We addressed two primary goals to our study. The first goal was to estimate the potential utility of cultivated cyanobacterial strains to study the relationships between cyanobacteria and bacteria by comparing (i) the richness and diversity within the BCs associated with cyanobacteria in culture to estimate any decrease in these two parameters in BCs associated with cyanobacterial strains with respect to the isolation date of these strains, and (ii) the structure and composition of BCs associated with two strains (one Microcystis and one Dolichospermum) in culture compared to natural BCs sampled from the pond from which these two strains were isolated to determine whether culture conditions had a deep impact on the BCs associated with cyanobacteria. The second goal of this study was to identify the major factors influencing the structural and functional diversity of BCs associated with the 15 cultured cyanobacterial strains based on different strain characteristics such as their taxonomic affiliation, culture time, N2-fixing capacity (heterocystous versus non-heterocystous strains) and media composition used for the culture of these cyanobacteria.
Materials and Methods
Experimental Design
The 15 selected non-axenic cultures were obtained from the Paris Museum Collection (PMC) of cyanobacteria. A description of the different genera and geographical origins is provided in Table 1. All cyanobacterial strains were cultured under the same conditions as the PMC collection of cyanobacteria (25°C and illumination of 30 μmol quanta/m2/s with a photoperiod of 16/8 h of day/night). Cultures were maintained in 250-mL Erlenmeyer flasks containing 100 mL of Z8 medium (Kotai, 1972) for Microcystis and Planktothrix strains and Z8X medium (without nitrate but with ammonia in trace element solution) for N2-fixing Dolichospermum and Aphanizomenon. We chose to cultivate the N2-fixing (N2+) and non-N2-fixing (N2-) cyanobacterial strains in two different media to maintain continuity, as they had been cultured in these media since their isolation, and we did not want to disturb the cyanobacteria and their associated BCs by changing the culture conditions. All experiments were performed in triplicate batch cultures. The cultures were sampled after 27 days, during the logarithmic growth phase.
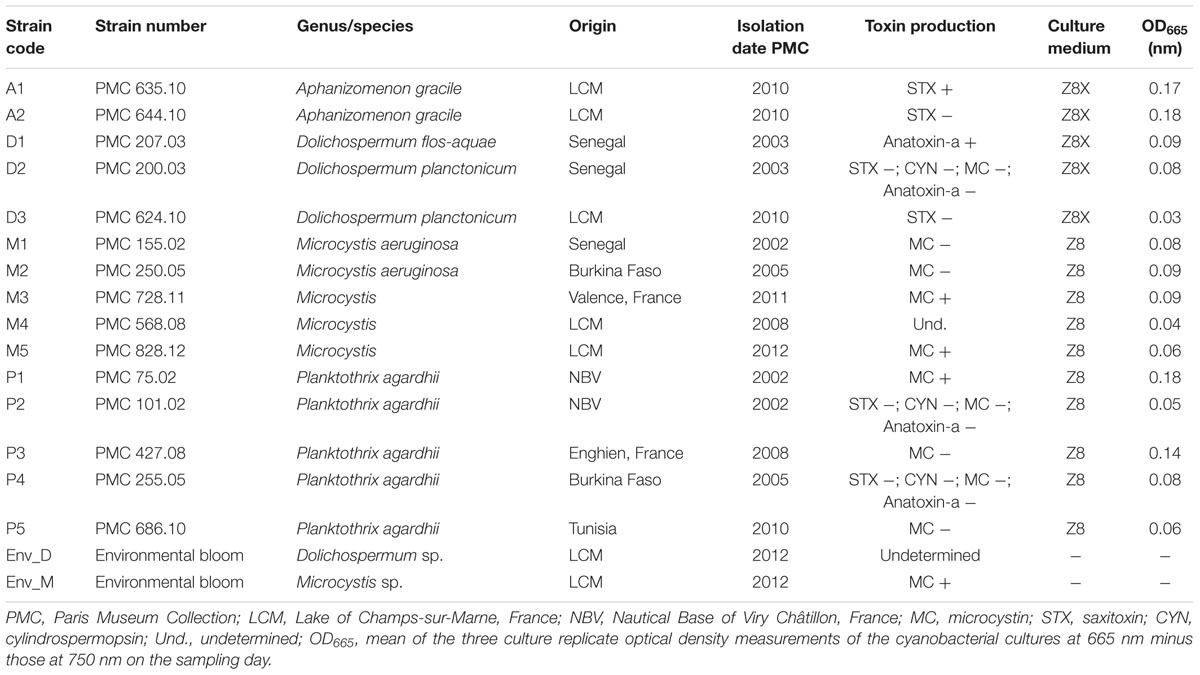
TABLE 1. Characteristics of the cyanobacterial strains and environmental samples used in this study.
For each culture, a double filtration was performed. One hundred milliliters of each culture sample was first filtered through a 1.2-μm pore-size filter membrane (Isopore Membrane Filters, Millipore) to remove cyanobacterial cells; then, 20 mL of this filtrate was utilized for a Biolog EcoPlate experiment. Finally, 80 mL of filtrate was aseptically filtered through a 0.2-μm pore-size filter (Track-Etched Membranes, Whatman®NucleporeTM). These filters were stored at -20°C for further molecular analyses. This removal step by filtration was required to access to the major BCs associated to cyanobacteria in cultures, but in return, bacteria attached to organic particles or to cyanobacteria might have also been removed. However, considering that these attached bacteria have a free-living phase and that the carbon sources available for heterotrophic bacteria were only provided by cyanobacteria, we have considered that the sampled BCs were directly influenced by cyanobacteria in our cultures.
Environmental cyanobacteria were collected from a recreational lake located near the city of Champs-sur-Marne (France, 48°51′47.0 N, 02°35′53.9 E) in 2012. Two cyanobacterial genera bloomed during the summer season, Dolichospermum in July and Microcystis in September, and they were sampled as previously described (Pascault et al., 2015). The cultivated Microcystis strain M5 (Table 1) used in this study was also isolated at the same time, while the Dolichospermum strain D3 (Table 1) was isolated from this lake 2 years prior. Natural bloom samples were counted by microscope, and 95% of cells belonged to Dolichospermum during the bloom of this genus, while the Microcystis bloom sample consisted of 84% Microcystis and 15% Pseudoanabaena mucicola.
Carbon Source Utilization
The functional diversity of BCs associated with cyanobacterial strains in culture was assessed by implementing a Biolog EcoPlate approach (EcoPlates; Biolog, Hayward, CA, USA). The Biolog EcoPlate experiment used 96-well microtiter plates containing a total of 31 carbon sources and a blank containing only water, all in triplicate. In addition to the carbon source, each well-contained the redox agent tetrazolium violet as an indicator of substrate oxidation. Positive wells developed a purple color that was quantified spectrophotometrically. Each well was inoculated with 150 μL of sample, and the plates were then incubated at 25°C for 1 week. During the incubation, the absorbance of the plates was measured every day at a wavelength of 590 nm using a spectrophotometric microplate reader (Molecular Devices, USA).
The optical density in each well was averaged among technical replicates and among culture triplicates. The carbon metabolic activity of the BCs was analyzed for all 31 carbon sources using principal component analysis (PCA). For each BC associated with the different cyanobacterial strains, microbial activity was expressed as the average well-color development (AWCD), determined as follows: AWCD = ΣODi/31, where ODi is the mean optical density value from each well. Carbon metabolic richness was calculated as the number of oxidized carbon substrates using an OD of 0.25 as the threshold for a positive response (Garland, 1996). The nitrogen use index (NUI) was calculated and expressed as the ratio of total substrate utilization to substrates containing nitrogen (Sala et al., 2006).
Molecular Approaches
The structural diversity of BCs associated with cyanobacterial strains in culture and with natural blooms of cyanobacteria was assessed using a 16S rRNA gene fragment pyrosequencing approach. The DNA extraction procedure involved mechanical and chemical extraction and was adapted from the procedure described by Massana et al. (1997). Each 0.2-μm filter was resuspended in 1.1 mL of lysis buffer (40 mM EDTA, 50 mM Tris-HCl, 0.75 M sucrose) and crushed using Lysing Matrix E tubes in a Fast Prep (MP Biomedical, Fast Prep Instrument®-24). In addition, enzymatic lysis was performed by adding lysozyme (0.5 mg/mL). After 45 min at 37°C, deproteinization was carried out by bringing the mixture to a final concentration of 10% sodium dodecyl sulfate (SDS) and adding proteinase K (0.2 mg/mL, Thermo Scientific) for 1.5 h at 55°C. Then, samples were centrifuged, and the supernatants were collected and purified with phenol-chloroform-isoamyl alcohol followed by chloroform. After precipitating with a 0.1 sample volume of sodium acetate and a 0.6 sample volume of isopropanol, the nucleic acids were washed with 80% ethanol and re-suspended in 100 μL of ultrapure water. The samples were stored at -20°C.
A region of the 16S rRNA gene including the variable region V4–V5 was selected for tag pyrosequencing. This region was amplified using the bacterial forward primer 563F (Claesson et al., 2010), which also includes the Roche 454 pyrosequencing adapter FLX A and a unique 10-bp barcode, and the bacterial reverse primer 907rm (Schauer et al., 2003), which includes the Roche 454 pyrosequencing adapter FLX B. Polymerase chain reaction (PCR) was performed under the following conditions: 98°C for 2 min; 30 cycles of 98°C for 10 s, 52°C for 30 s and 72°C for 1 min; 72°C for 10 min. Each DNA sample was amplified in two replicate PCR reactions, which were then pooled. These PCR products were then purified using the MinElute Gel Extraction Kit (Qiagen, Venlo, Pays-Bas). The amount of DNA in each sample was quantified using the Qubit dsDNA HS assay (Invitrogen, Carlsbad, CA, USA). Finally, the PCR products were combined in equimolar amounts and sequenced using a Genome Sequencer GS FLX Titanium 454 of GATC Biotech (Roche Company, Branford, CT, USA).
Bioinformatics Analysis
The 454 pyrosequencing of the 16S rRNA gene produced 514,905 raw sequences. All sequences were cleaned by applying PANGEA trimming (Giongo et al., 2010) with a quality threshold (>23) and a minimum sequence length of 270 bp and by removing sequences with errors in the forward primer. The remaining sequences were clustered using USEARCH (Edgar, 2010) at a 97% similarity threshold and were then grouped according to different taxonomic levels using the SILVA base (Pruesse et al., 2007). The LCA (Lowest Common Ancestor) assignment method was employed. The process was automated by PANAM pipeline1, which constructs phylogenetic trees for taxonomic annotation (Taib et al., 2013). After the removal of low-quality sequences, a range of 2,127 to 17,266 sequences was obtained for each BC associated with different cyanobacterial strain triplicates. After removing the cyanobacterial sequences (representing approximately 25% (± 22%) of the total bacterial sequences among the different samples) and singletons (every sequence found only once in the entire data set), 1,005 to 16,383 sequences were retrieved per sample. For further analyses, sequence data sets were normalized to 1,004 sequences. Within the 50,200 sequences analyzed (1,004 sequences per sample), a total of 1,125 operational taxonomic units (OTUs) were obtained from cyanobacteria-associated BCs in culture, and 1,458 OTUs were obtained from combined analyses of cyanobacteria-associated BCs in culture and those from the two natural bloom samples. OTU thresholds were defined by an abundance of reads >1% within a pooled sample corresponding to an abundant OTU, whereas a proportion <0.01% represented a rare OTU, as described in Pedrós-Alió (2006) and Galand et al. (2009).
Statistical Analysis
All statistical analyses were performed using the statistical software package R 2.15.1 (R Development Core Team, Vienna, Austria). Only differences at p < 0.05 were considered significant.
To test the potential effects of the different characteristics of the cyanobacterial strains (strain, genus, sampling date, putative toxicity, N2 fixation, and culture medium) on the different variables studied (AWCD, carbon metabolic richness, NUI, the three first PCA axes of the two PCAs based on EcoPlate and 454 pyrosequencing data), we first chose the best model according to the Akaike Information Criterion (AIC) in a Stepwise Algorithm using the “step” function in R, and we then calculated type II analysis-of-variance (ANOVA) tables. We took into account certain factors that were not independent, such as the cyanobacterial strain and genus or N2 fixation and culture medium. Then, differences in carbon substrate use, the NUI, the AWCD and the carbon metabolic richness according to the cyanobacterial genera were tested by ANOVA and Tukey’s tests.
Principal component analysis and co-inertia analyses were performed using Ade4TkGUI software (Thioulouse and Dray, 2007). A Hellinger transformation had previously been carried out on the 454 pyrosequencing data using the vegan package (Dixon, 2003). Permutational multivariate ANOVA (PERMANOVA) and Mantel tests were performed using the vegan package to test for differences between BCs associated with N2+ cyanobacteria and those associated with N2- cyanobacteria. Diversity indices were calculated using the vegan package with the normalized data set (Dixon, 2003).
The effects of N2 fixation/culture medium on the distribution of the major bacterial orders and classes and on the dominant bacterial OTUs (sequence numbers > 1% of the total sequences) associated with the different cyanobacterial strains were analyzed by performing ANOVA followed by Tukey’s tests.
A network approach was implemented to visualize OTU distribution among the different cyanobacterial strains. The network was constructed based on the presence/absence of data for each OTU in all cyanobacterial cultures after pooling the replicates. Cyanobacterial strains were used as source nodes, and OTUs were used as target nodes, with edges (that is, lines connecting nodes) corresponding to positive associations between OTUs and cyanobacterial strains. Then, the network was visualized using the edge-weighted spring-embedded layout algorithm in Cytoscape Software (Shannon et al., 2003).
Results
Richness and Diversity in BC Associated with Cyanobacteria in Culture and in Natural Blooms of Cyanobacteria
As shown in Table 2, the richness and diversity values of BCs associated with the two cultured strains Dolichospermum D3 and Microcystis M5 were substantially lower than those of BCs associated with natural blooms of these two genera in the lake from which these two strains were isolated. However, when all values obtained for the 15 strains in culture and for the two natural blooms were compared, no significant differences in the diversity indices of these BC types were noted (Kruskal–Wallis tests; p = 0.06, 0.06, and 0.19 for richness, Chao1 and Shannon, respectively; Table 2). Furthermore, there were no significant differences in the richness and diversity of BCs associated with cyanobacterial strains isolated before or after 2008 (Kruskal–Wallis tests; p = 0.94, 0.80, and 0.54 for richness, Chao1 and Shannon, respectively).
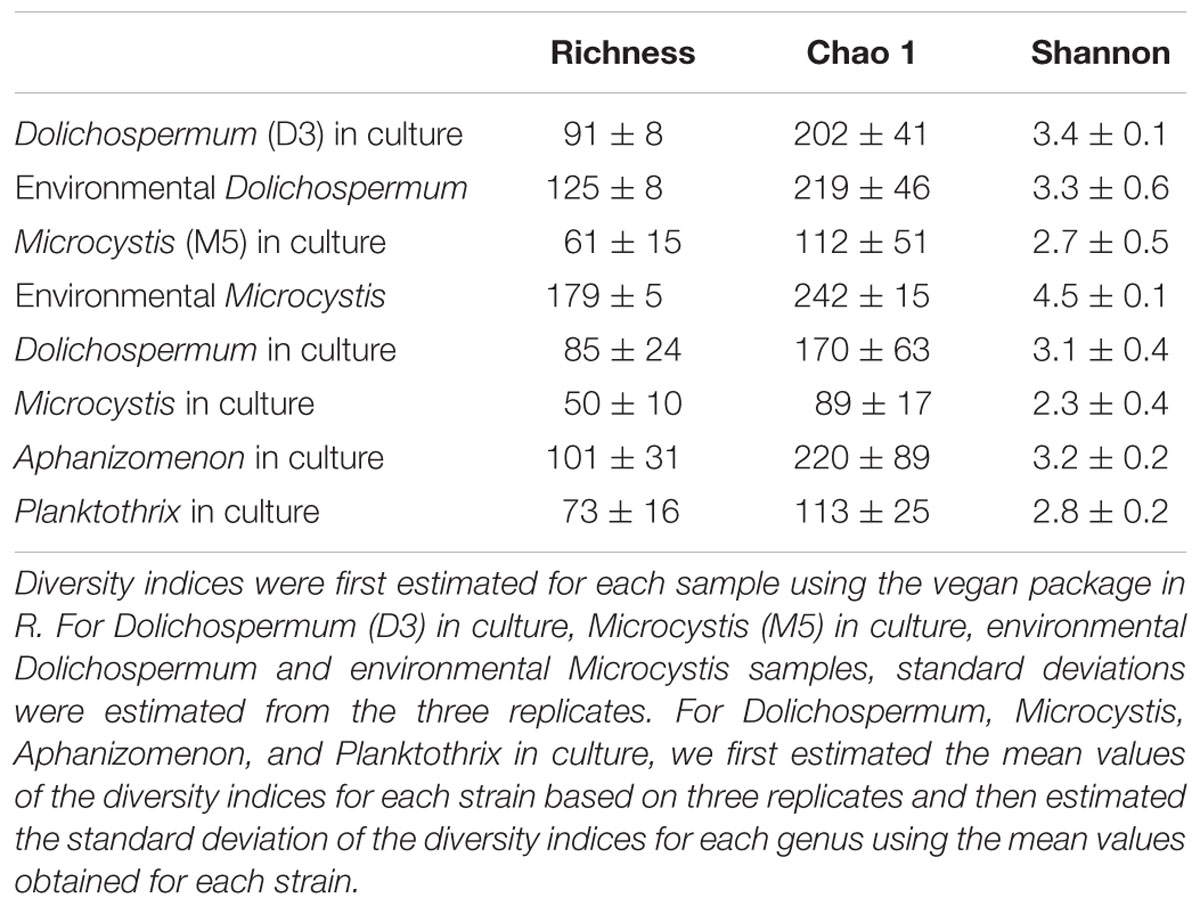
TABLE 2. Richness, Chao1 and diversity values estimated for the bacterial communities associated with cyanobacteria in culture and with the two environmental samples.
Comparison of the global composition and structure of the BCs associated with cyanobacteria in culture and those associated with natural blooms of Dolichospermum and Microcystis revealed a shared dominance by Proteobacteria, particularly Alphaproteobacteria, and Bacteroidetes in these communities. These BCs primarily differed in terms of the relative abundances of Actinobacteria and Betaproteobacteria and, to a lesser extent, Verrucomicrobia (Figure 1); these phyla were more abundant in BCs associated with cyanobacteria in natural blooms than those in culture. At the OTU level, significant differences were found between natural and cultivated BCs, as these BCs shared only 5.6% of their OTUs (Figure 2). However, these shared OTUs contained more than 26.2% of all sequences; thus, the majority of these shared OTUs were likely dominant in all BCs (Figure 2). Interestingly, PCA performed on all data revealed the distribution of BCs in three different groups: BCs from environmental samples, BCs from N2+ cyanobacteria in culture and BCs from N2- cyanobacteria in culture (Supplementary Figure S1).
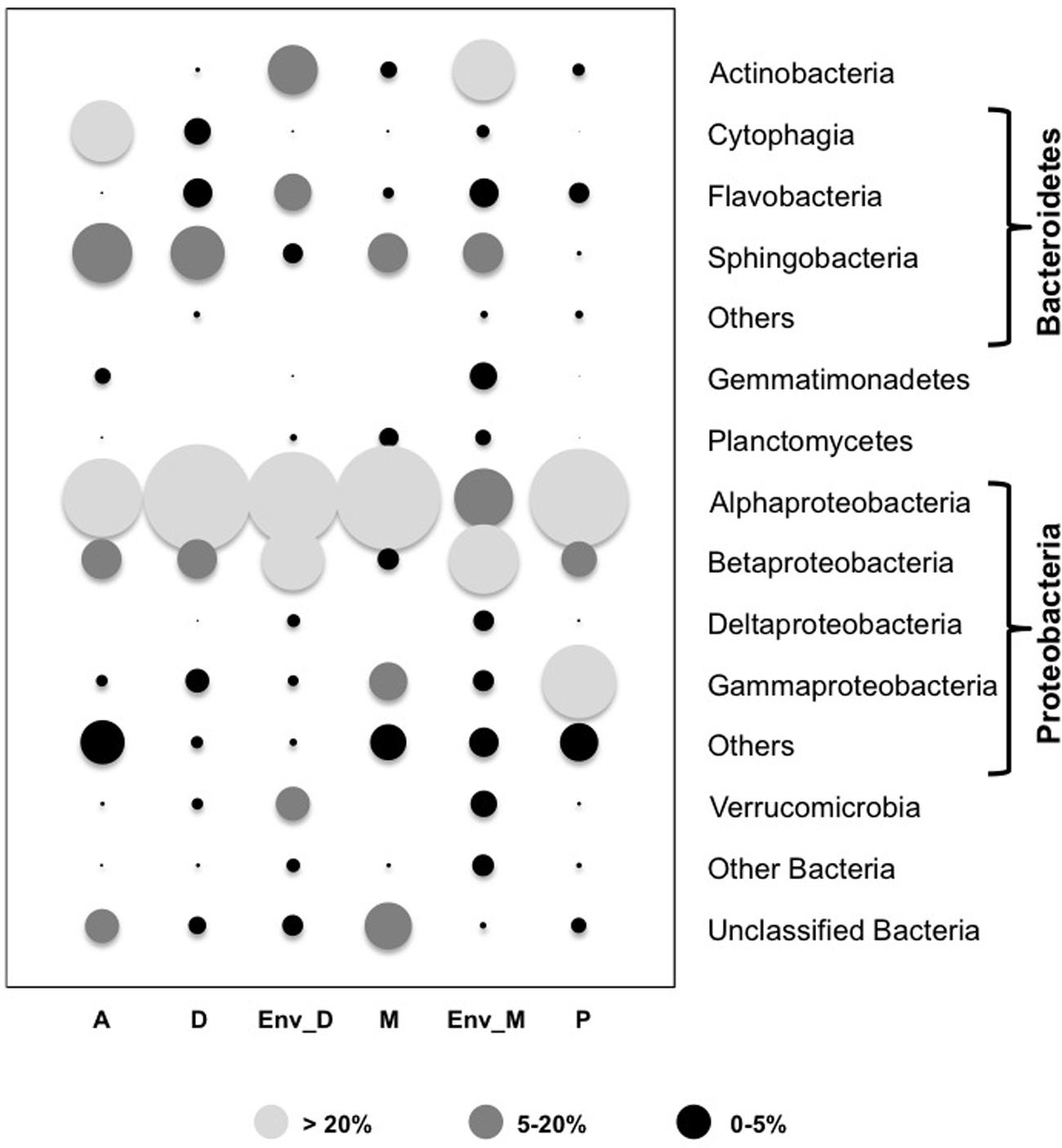
FIGURE 1. Relative abundance of main groups found in the bacterial communities (BCs) associated with the four cyanobacterial genera in culture and with the two natural blooms of cyanobacteria after normalization to the smallest sample (n = 1004). The size of the circles was estimated after averaging in a first time, the number of reads found in the three replicates for each strain and in a second time the mean number of reads found in each strain of a given genus. A: Aphanizomenon strains; D: Dolichospermum strains; M: Microcystis strains; P: Planktothrix strains; Env_D: Environmental sample of an Dolichospermum bloom; Env_M: Environmental sample of a Microcystis bloom; Others (within the Bacteroidetes and the Proteobacteria sections): all the sequences of minority classes (<1% for all the Cyanobacterial genera) and also included for the Proteobacteria section, sequences only identified as Proteobacteria using the SILVA base and the assignment LCA (Lowest Common Ancestor); Other Bacteria: all the sequences of non-dominant phyla (<1% for all the Cyanobacterial genera); Unclassified Bacteria: sequences only identified as Bacteria using the SILVA by the LCA method.
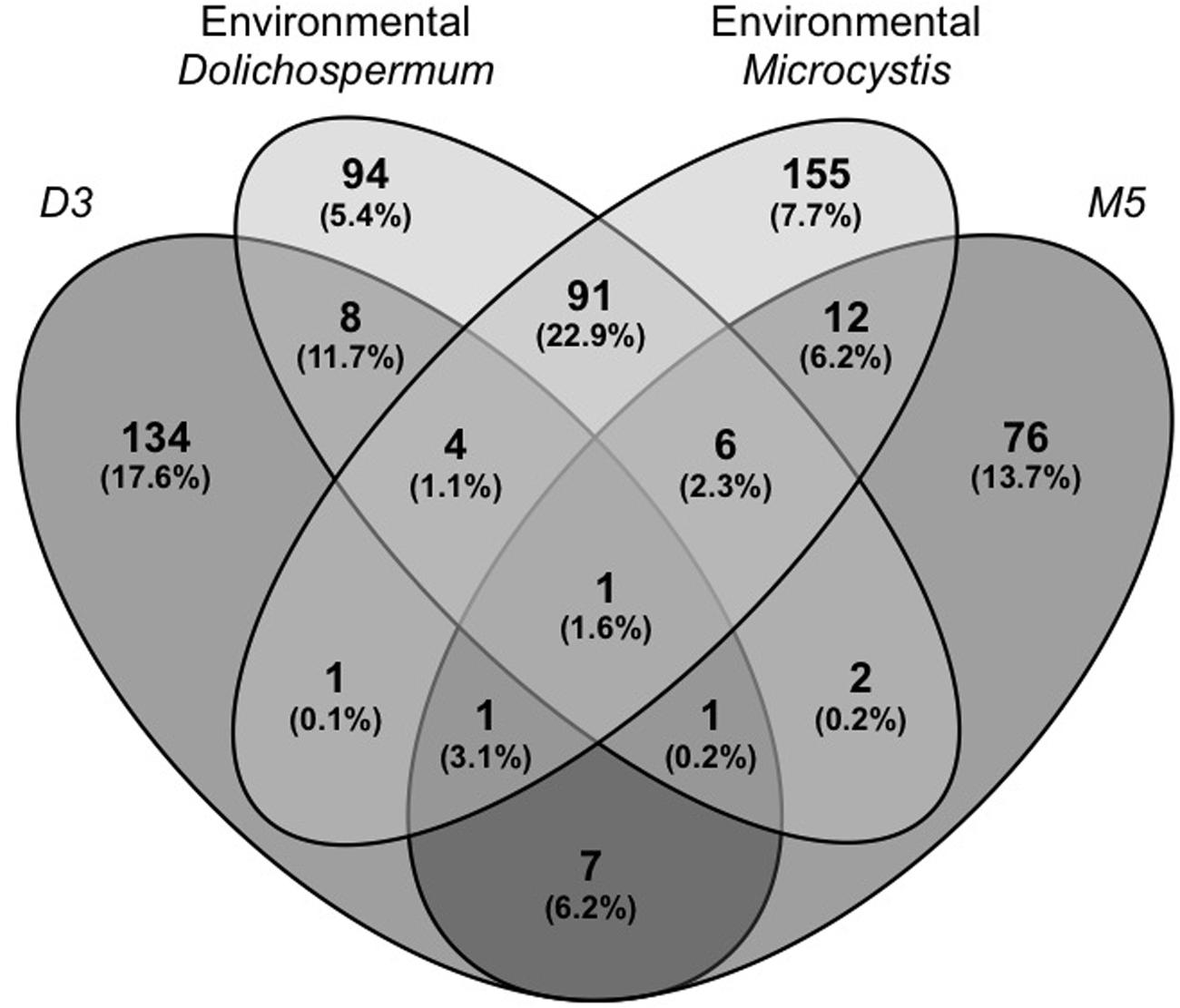
FIGURE 2. Venn diagram showing the distribution and abundance of the bacterial OTUs found in the BCs sampled during the two natural blooms of Microcystis and Dolichospermum and in the BCs associated with the Microcystis (M5) and Dolichospermum (D3) strains in culture. The abundances (total read percentages) of these OTUs are represented in brackets. The number of reads was first added for the three replicates, and their abundances were then calculated for the different OTU groups based on total OTUs for these four cyanobacteria, both for natural blooms and individual strains (n = 12,048 reads). D3: BC associated with Dolichospermum strain 3 (isolated from Champs/Marne); M5: BC associated with Microcystis strain 5 (isolated from Champs/Marne); Environmental Dolichospermum/Microcystis: BC associated with the two bloom samples of Dolichospermum and Microcystis from Champs/Marne.
Comparison of the Carbon and Nitrogen Metabolic Activities of BCs Associated with Cyanobacterial Cultures
To determine whether BCs associated with different cyanobacterial strains in culture exhibited divergent catabolic profiles, we implemented a Biolog EcoPlate approach. The number of carbon substrates and the AWCD were significantly higher (ANOVA analysis: df = 1, F-value = 126.1, p = 2.4 10-10 and df = 1, F-value = 198.8, p = 3.5 10-12, respectively) for BCs associated with N2- cyanobacteria (Microcystis and Planktothrix strains) cultivated in Z8 medium than for those associated with N2+ cyanobacteria (Dolichospermum and Aphanizomenon strains) cultivated in Z8X medium. More specifically, after grouping every carbon source by chemical category, amino acids, carbohydrates and phenolic substances were highly consumed by BCs associated with N2- cyanobacteria (Figure 3A). Similarly, the NUI (Urakawa et al., 2013) was significantly higher for BCs associated with N2- cyanobacteria than for those associated with N2+ cyanobacteria (ANOVA analysis: df = 1, F-value = 289.5, p = 9.3 10-14) (Figure 3B). Overall, among the different characteristics of the cyanobacterial strains (N2 fixation, culture medium, date of isolation, genus and putative toxicity), only N2 fixation/culture medium and the interactions among cyanobacterial strain, genus and N2 fixation/culture medium had significant effects on carbon and nitrogen metabolic activities [ANOVA analyses using best models given by AIC analyses (∼Fixation + Fixation:Strain:Genus), df = 1 and F-value > 126.1 for N2 fixation; df = 13 and F-value < 18.1 for Fixation:Strain:Genus, p < 0.001].
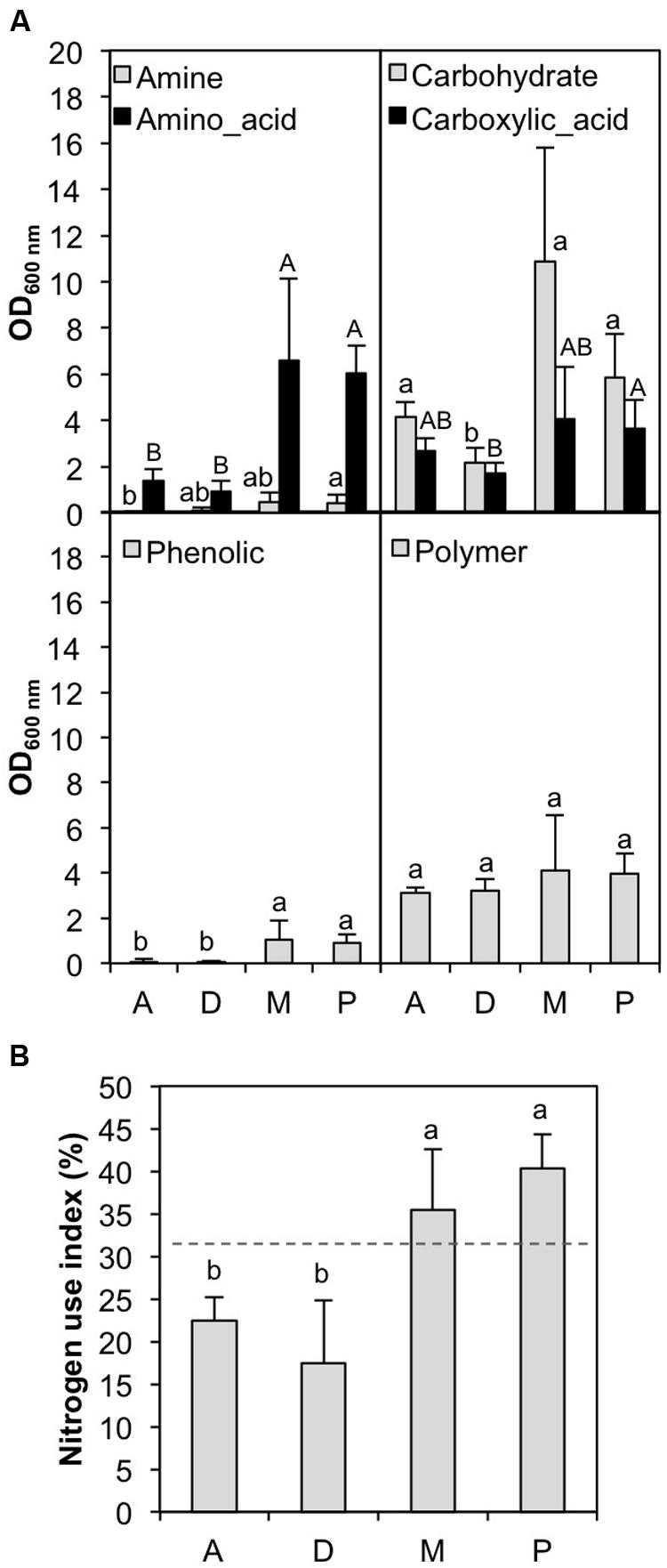
FIGURE 3. Comparative use of (A) different families of carbon substrates available in the Biolog EcoPlates and (B) nitrogen use index by the BCs associated with cyanobacterial strains in culture. D: Dolichospermum strains; A: Aphanizomenon strains; M: Microcystis strains; P: Planktothrix strains. Letters with different labels indicate significant differences (p < 0.05).
Finally, PCA revealed a clear distinction between the BCs associated with Dolichospermum and Aphanizomenon strains that grouped together and those associated with Planktothrix and Microcystis strains (PERMANOVA analysis, df = 1, R2 = 0.35, p = 0.0001; Figure 4A). Furthermore, the BCs associated with different Microcystis strains and, to a lesser extent, the Planktothrix strains demonstrated higher carbon source variability than those associated with the Dolichospermum and Aphanizomenon strains.
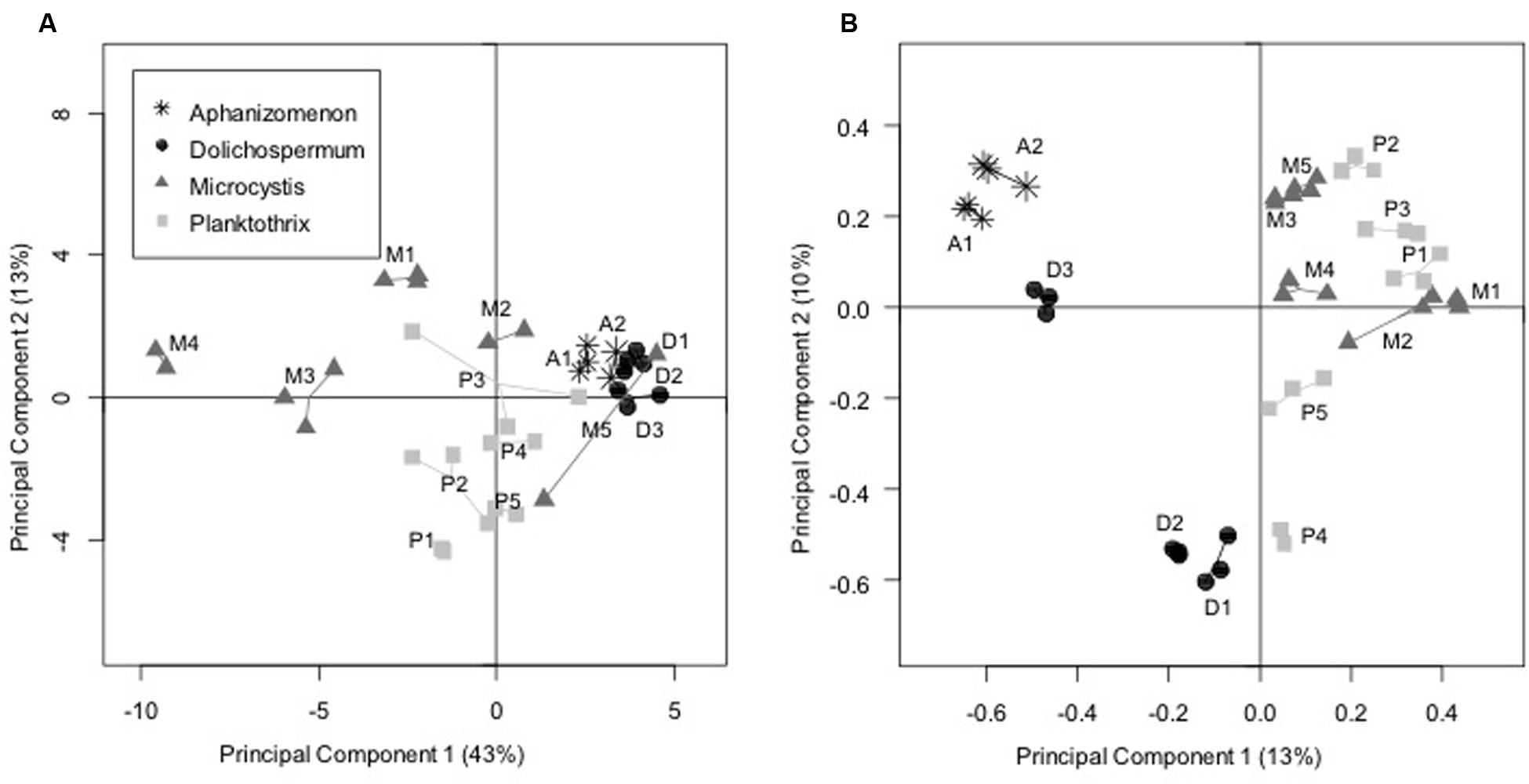
FIGURE 4. Principal component analyses performed (A) on the use of the 31 carbon sources (EcoPlate approach) and (B) on the OTU composition (pyrosequencing approach) of the BCs associated with the cyanobacterial strains in culture. For each strain, the replicates were connected together in order to facilitate the reading. A: Aphanizomenon strains; D: Dolichospermum strains; M: Microcystis strains; P: Planktothrix strains.
Comparison of the Structure and Composition of BCs Associated with Cyanobacterial Cultures
Based on overall BC structure, Proteobacteria (particularly Alphaproteobacteria) and Bacteroidetes were the two most abundant phyla in all culture samples (Figure 1), while the read numbers of other phyla did not exceed 2% (0.7–2.0%). Even if the global structures of BC communities were very similar, regardless of the cyanobacterial genera, some groups were preferentially detected in only one cyanobacterial genus (Figure 1). For example, Planctomycetes were primarily found in BCs associated with Microcystis strains, whereas Cytophaga (Bacteroidetes) were primarily found in heterocystous cyanobacterial cultures.
When the BCs associated with the 15 cyanobacterial strains at the OTU level were compared in our network analysis (Figure 5), almost all abundant OTUs were shared by several BCs (OTUs located at the center of Figure 5), while numerous OTUs with low abundance were generally found in only one BC (OTUs located at the periphery of the Figure 5).
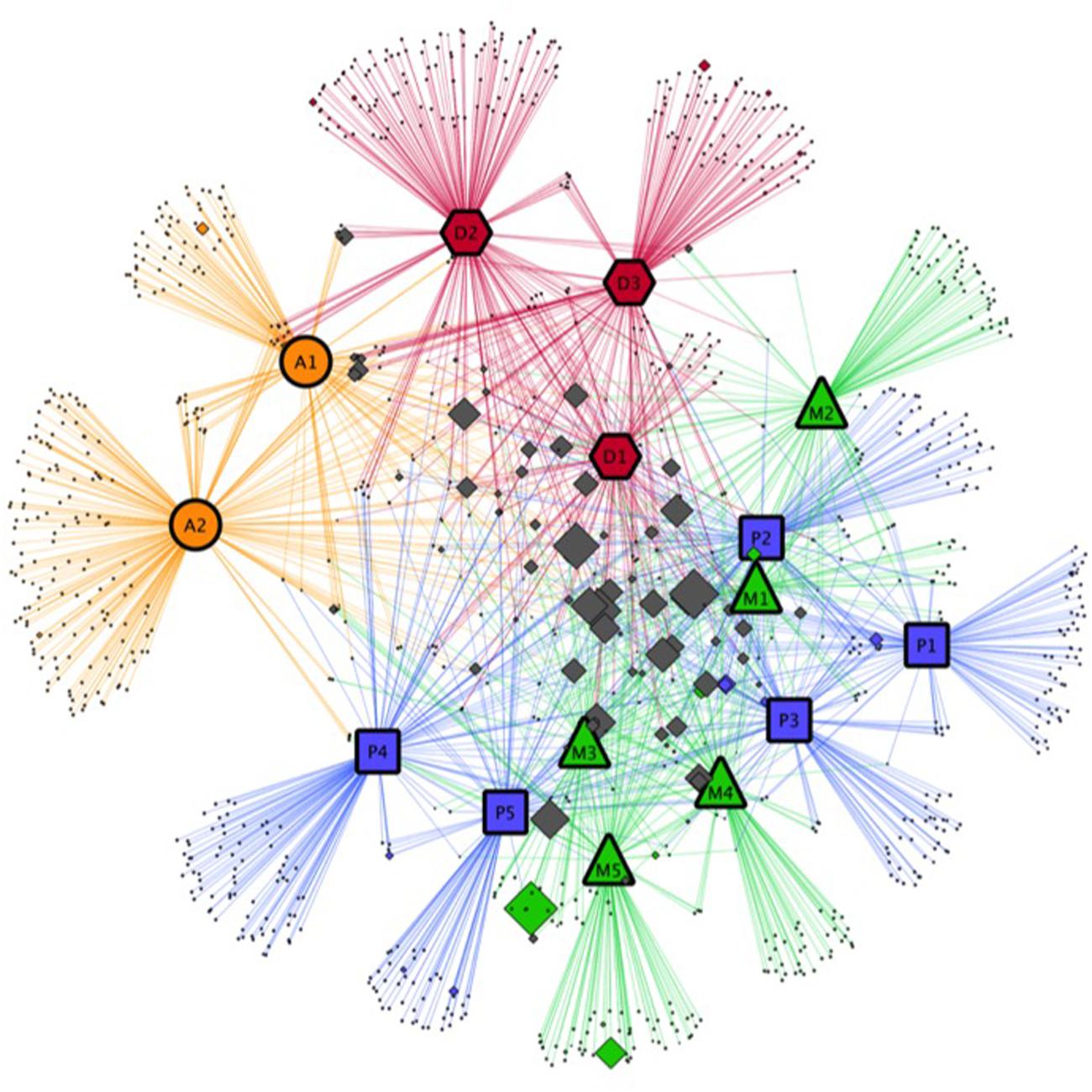
FIGURE 5. Network analysis on the relationships between cyanobacterial strains in culture and the bacterial OTU identified in our study. Colored symbols with large black border represent the different cyanobacterial strains: orange circles, red hexagons, green triangles and blue rectangles represent respectively Aphanizomenon strains (A), Dolichospermum strains (D), Microcystis strains (M), and Planktothrix strains (P). Diamonds represent the bacterial OTU found in associated with cyanobacterial strains. Colored diamonds are used for bacterial OTU associated with only one cyanobacterial strain (for example, green diamonds correspond to bacterial OTU only associated with one Dolichospermum strain). Gray diamonds represent OTU associated with several cyanobacterial strains. The size of the diamonds is proportional to the relative abundance of the OTU.
Special attention was focused on the distribution of the 100 most abundant OTUs. Forty seven of these OTUs demonstrated a significant difference (ANOVA analysis, p < 0.05) in their relative abundance when comparing BCs associated with N2+ cyanobacteria cultivated in Z8X medium and those associated with N2- strains cultivated in Z8 medium. Second, BLAST analysis was performed with these 100 OTUs in the GenBankTM (GB) database to determine how many sequences exhibited 98% sequence identity across more than 98% of their sequence length to our 100 OTUs. The taxonomic assignments of these OTUs obtained via BLAST analysis were congruent with those obtained from the SILVA database (see Materials and Methods). Interestingly, for approximately 50% of our OTUs, only a limited number of matches were found in GB (less than 50 sequences for each OTU), whereas for the rest of the OTUs, numerous matches were found in GB (at least 100 sequences but frequently several 100). In instances where few sequences were retrieved from GB, almost all were associated with freshwater ecosystems, and the majority of these OTUs were associated with cyanobacterial blooms (Supplementary Table S1). Conversely, when more than 100 sequences were retrieved from GB, these sequences originated from various ecosystems that were generally characterized by a high level of organic matter (OM), including freshwater ecosystems but also wastewater sludge, sediment, human waste and skin, and soils (Supplementary Table S1). In addition to their affinity for OM, we paid special attention to the potential ability of these dominant bacterial OTUs associated with cyanobacteria in culture to fix free nitrogen. Within the dominant OTUs, we identified OTUs belonging to three genera (Rhizobium, Azospirillum, and Pseudomonas) known to contain potentially N2-fixing species. Their abundance in each cyanobacterial genus varied widely according to the strain and replicate (from 0 to 23.5% in Dolichospermum, 0 to 7.2% in Aphanizomenon, 0 to 37.6% in Microcystis, and 0 to 43.3% in Planktothrix). Rhizobium and Pseudomonas were primarily associated with N2- cyanobacteria while Azospirillum was primarily associated with N2+ cyanobacteria.
Principal component analysis performed on the distribution of all bacterial OTUs in BCs associated with the 15 cyanobacterial strains (in triplicate) revealed a significant difference (PERMANOVA analysis: df = 1, R2 = 0.14, p = 0.0001) along the first axis of the analysis (Figure 4B) between BCs associated with N2+ cyanobacteria cultivated in Z8X medium and those associated with N2- cyanobacteria cultivated in Z8 medium. When testing the potential effects of certain cyanobacterial strain characteristics (N2 fixation, culture medium, strain, genus, sampling date, and putative toxicity), only N2 fixation/culture medium and the interactions among cyanobacterial strain, genus, and N2 fixation/culture medium had significant effects on the BC distribution along the first two PCA axes (ANOVA analyses using best models given by AIC analyses (∼Fixation + Fixation:Strain:Genus), df = 1, R2 = 0.14, p = 0.0001 for N2 fixation and df = 13, R2 = 0.74, p = 0.0001 for interactions among cyanobacterial strain, genus, and N2 fixation/culture medium). Finally, a co-inertia analysis revealed a significant correlation between the two PCA analyses performed on the 454 pyrosequencing and on the EcoPlate data (Mantel test: R2 = 0.45, p = 0.0001).
Discussion
With regard to the first aim of our study, which investigated the potential utility of cultivated cyanobacterial strains to study the relationships between cyanobacteria and bacteria, our data indicate a high level of richness and diversity maintained in BCs associated with cultured cyanobacteria, even in strains isolated a long time ago. Two major processes may be involved in the conservation of BC diversity. First, changes in the quality and the quantity of OM produced by cyanobacteria during growth (Leloup et al., 2013) may cause temporal successions in BCs (Bagatini et al., 2014; Brauer et al., 2015), maintaining a large degree of richness and diversity. Second, the influence of bacteriophages on the dynamics and diversity of BCs may also sustain a high level of richness in our cultured communities (see reviews of Wommack and Colwell, 2000; Sime-Ngando, 2014).
In addition to these findings, the comparison of BC communities associated with cyanobacterial cultures and those living in freshwater ecosystems hosting cyanobacterial blooms revealed several shared similarities at the phylum level, specifically a low proportion of Actinobacteria and a relatively high abundance of Gammaproteobacteria in addition to the dominance of Alpha- and Betaproteobacteria. In freshwater ecosystems, Actinobacteria are very abundant, while Gammaproteobacteria represent a tiny minority when there are no cyanobacterial blooms (e.g., Humbert et al., 2009). In addition, the abundance of these Actinobacteria decreases while the abundance of Gammaproteobacteria increases during cyanobacterial blooms (Parveen et al., 2013; Ghai et al., 2014), which we observed in both cultivated and natural BCs. At the OTU level, there were large differences between cultivated and natural communities, but a small number of OTUs were shared by natural and cultured BCs. These OTUs contained a high number of reads, indicating their presence among the core species of these BCs. BCs contain an assemblage of core and satellite species as defined by Gibson et al. (1999); the former are abundant and widely distributed, while the abundances of the latter are much more variable, and their distribution is limited to a restricted number of sites (Pedrós-Alió, 2006; Humbert et al., 2009). The presence of these core species in cultivated BCs supports their ability to survive under culture conditions and to play an important role in community functions.
Finally, when considering our data in terms of the potential utility of these cultivated communities to study the interactions between heterotrophic bacteria and cyanobacteria, the complexity of the cultivated communities and the presence of OTUs belonging to the core species of natural communities suggest that these cultivated strains are good models for further studies.
The second aim of this study was to compare the BCs associated with cyanobacterial strains belonging to four genera. Interestingly, these BCs displayed differences in their structural and functional diversity. In particular, BCs associated with N2+ genera cultivated in Z8X medium (Aphanizomenon and Dolichospermum) were clearly distinguishable from those associated with N2- genera cultivated in Z8 medium (Microcystis and Planktothrix). Within these two groups, separation between BCs according to genus was less evident. Significant congruence of the results obtained from the EcoPlate and pyrosequencing approaches was noted; thus, differences in the OTU composition of BCs associated with cyanobacteria in cultures likely correspond to differences in their ability to use different carbon sources.
One of the major questions raised by these findings concerns the relative importance of the cyanobacteria versus that of the culture media in terms of the differences observed in BCs. In natural aquatic ecosystems (Sarmento and Gasol, 2012; Landa et al., 2014) and in culture (Bagatini et al., 2014; Landa et al., 2014), the composition of microbial communities is primarily influenced by the quality and quantity of OM available in these systems. Furthermore, N2+ cyanobacteria release significant amounts of NH4+ (Mulholland, 2007; Ploug et al., 2011; Adam et al., 2016), which is used by the surrounding BC and consequently prevents putative BC limitation by nitrogen in Z8X media. In this study, the EcoPlate approach revealed a greater NUI for BCs associated with N2- cyanobacteria cultivated in Z8 medium than for BCs associated with N2+ cyanobacteria cultivated in Z8X medium without nitrogen; this difference may be a consequence of potential nitrogen limitation in BCs associated with N2- cyanobacteria. However, Brauer et al. (2015) recently demonstrated competition and facilitation between a marine N2-fixing cyanobacterium and its associated BC and found the influence of nitrogen and carbon availability on the dynamics and composition of BCs associated with the cyanobacterium to be highly complex and dependent on carbon availability and physical parameters such as temperature. It is very difficult to evaluate the relative importance of the culture medium and of nitrogen fixation by cyanobacteria on the observed differences between BCs associated with N2+ and N2- cyanobacteria; thus, new studies focusing specifically on the nitrogen cycle will facilitate a better answer to this question.
The second interesting finding obtained by comparing the structure and composition of BCs associated with cyanobacteria in culture was the observation of two major categories of dominant OTUs in these BCs: OTUs specifically associated with cyanobacteria or other phytoplankton species (phyto-OM specialized bacteria) and OTUs found in environments characterized by high quantities of OM, such as sludge, human waste, and the rhizosphere (generalist-OM bacteria). Interestingly, several OTUs that were widely distributed among both our cultivated BCs and in natural environments are known to potentially fix N2. The majority of these were closely related (i) to the genus Rhizobium, known to engage in symbiotic relationships with plant roots and to fix free nitrogen, even when free-living (Dreyfus et al., 1983; Ludwig, 1984; Alazard, 1990); (ii) to the genus Azospirillum, known to contain endophytic diazotrophic rhizobacteria (Santi et al., 2013); and (iii) to the genus Pseudomonas, which contains certain species that were recently described as potential N2 fixers in marine environments (Dang et al., 2013). Further studies on the diversity and the expression of functional genes such as nifH will permit to estimate the contribution of these potentially diazotrophic bacteria in the nitrogen cycle during cyanobacterial cultures. For example, in marine environments it has been shown that some Gammaproteobacterial diazotrophs could have a significant contribution to N budget in these ecosystems (Moisander et al., 2014). Moreover, it would be interesting to distinguish BCs that are attached to cyanobacteria and organic aggregates, from the free living BCs because it has been shown that suboxic conditions in aggregates may promote the N2 fixation activity of heterotrophic diazotrophs (see review of Dang and Lovell, 2016).
Conclusion
Our study revealed the complex structures of BCs associated with cultivated cyanobacterial strains, which contained several abundant OTUs that were also highly abundant in BCs associated with natural cyanobacterial blooms. These results highlighted the relevance of the culture models employed to study interactions between bacteria and cyanobacteria. An additional, original finding of this study is the observation that BCs associated with different cyanobacterial genera exhibited functional and structural differences related to the phylogenetic relationships among these cyanobacterial genera and, potentially, to the quality of the OM produced. Finally, BCs associated with cyanobacteria were composed of a variety of OM-generalist and specialist species, which will be explored in our future work.
Author Contributions
LZ, AZ, IL, SD, CB, LS, JL, and J-FH have worked on the design of the experiments. LZ, AZ, IL, SD, CD, and KT have performed the experiments. LZ, AZ, IL, DD, JL, and J-FH have performed the data analysis. CD and KT help to the data analysis. LZ, AZ, IL, CD, CB, DD, LS, JL, and J-FH have written the paper. J-FH co-led the project with JL, CB, and LS.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was funded by the PHYCOCYANO project in the Jeunes Chercheurs-Jeunes Chercheuses program of the French ANR (Agence Nationale de la Recherche; ANR-11-JSV7-014-01). The English text has been checked by a Language Editing Society.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01662
FIGURE S1 | Principal component analyses performed on the OTU composition (pyrosequencing approach) of the BCs associated with the two natural blooms of Microcystis and Dolichospermum and the 15 cyanobacterial strains in culture. For each strain, the replicates are connected to facilitate reading. Env. Dolichospermum and Env. Microcystis: environmental samples from a Dolichospermum bloom and a Microcystis bloom, respectively.
TABLE S1 | Description of the results obtained from the BLAST analysis performed on the 100 most abundant OTUs found in BCs associated with cyanobacteria in culture.
Footnotes
References
Adam, B., Klawonn, I., Svedén, J. B., Bergkvist, J., Nahar, N., Walve, J., et al. (2016). N2-fixation, ammonium release and N-transfer to the microbial and classical food web within a plankton community. ISME J. 10, 450–459. doi: 10.1038/ismej.2015.126
Alazard, D. (1990). Nitrogen fixation in pure culture by rhizobia isolated from stem nodules of tropical Aeschynomene species. FEMS Microbiol. Lett. 68, 177–182. doi: 10.1111/j.1574-6968.1990.tb04145.x
Bagatini, I. L., Eiler, A., Bertilsson, S., Klaveness, D., Tessarolli, L. P., and Vieira, A. A. H. (2014). Host-specificity and dynamics in bacterial communities associated with bloom-forming freshwater phytoplankton. PLoS ONE 9:e85950. doi: 10.1371/journal.pone.0085950
Berg, K. A., Lyra, C., Sivonen, K., Paulin, L., Suomalainen, S., Tuomi, P., et al. (2009). High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 3, 314–325. doi: 10.1038/ismej.2008.110
Brauer, V. S., Stompe, M., Bouvier, T., Fouilland, E., Leboulanger, C., Confurius-Guns, V., et al. (2015). Competition and facilitation between the marine nitrogen-fixing cyanobacterium Cyanothece and its associated bacterial community. Front. Microbiol. 5:795. doi: 10.3389/fmicb.2014.00795
Buchan, A., LeCleir, G. R., Gulvik, C. A., and González, J. M. (2014). Master recyclers: features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 12, 686–698. doi: 10.1038/nrmicro3326
Cai, H. Y., Yan, Z. S., Wang, A. J., Krumholz, L. R., and Jiang, H. L. (2013). Analysis of the attached microbial community on mucilaginous cyanobacterial aggregates in the eutrophic Lake Taihu reveals the importance of Planctomycetes. Microb. Ecol. 66, 73–83. doi: 10.1007/s00248-013-0224-1
Claesson, M. J., Wang, Q., O’Sullivan, O., Greene-Diniz, R., Cole, J. R., Ross, R. P., et al. (2010). Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res. 38:e200. doi: 10.1093/nar/gkq873
Dang, H., and Lovell, C. R. (2016). Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 80, 91–138. doi: 10.1128/MMBR.00037-15
Dang, H., Yang, J., Li, J., Luan, X., Zhang, Y., Gu, G., et al. (2013). Environment-dependent distribution of the sediment nifH-harboring microbiota in the northern south China sea. Appl. Environ. Microbiol. 79, 121–132. doi: 10.1128/AEM.01889-12
Dixon, P. (2003). VEGAN, a package of R functions for community ecology. J. Veg. Sci. 14, 927–930. doi: 10.1111/j.1654-1103.2003.tb02228.x
Dodds, W. K., Bouska, W. W., Eitzmann, J. L., Pilger, T. J., Pitts, K. L., Riley, A. J., et al. (2009). Eutrophication of US freshwaters: analysis of potential economic damages. Env. Sci. Tech. 43, 12–19. doi: 10.1021/es801217q
Dreyfus, B., Elmerich, C., and Dommergues, Y. (1983). Free-living Rhizobium strain able to grow on N2 as the sole nitrogen source. Appl. Environ. Microbiol. 45, 711–713.
Dziallas, C., and Grossart, H. P. (2011). Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp. Environ. Microbiol. 13, 1632–1641. doi: 10.1111/j.1462-2920.2011.02479.x
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Eiler, A., and Bertilsson, S. (2004). Composition of freshwater bacterial communities associated with cyanobacterial blooms in four Swedish lakes. Environ. Microbiol. 6, 1228–1243. doi: 10.1111/j.1462-2920.2004.00657.x
Galand, P. E., Casamayor, E. O., Kirchman, D. L., and Lovejoy, C. (2009). Ecology of the rare microbial biosphere of the Arctic Ocean. Proc. Nat. Acad. Sci. U.S.A. 106, 22427–22432. doi: 10.1073/pnas.0908284106
Garland, J. L. (1996). Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biol. Biochem. 28, 213–221. doi: 10.1016/0038-0717(95)00112-3
Ghai, R., Megumi Mizuno, C., Picazo, A., Camacho, A., and Roriguez-Valera, F. (2014). Key roles for freshwater Actinobacteria revealed by deep metagenomic sequencing. Mol. Ecol. 23, 6073–6090. doi: 10.1111/mec.12985
Gibson, D. J., Ely, J. S., and Collins, S. L. (1999). The core-satellite species hypothesis provides a theoretical basis for Grime’s classification of dominant, subordinate, and transient species. J. Ecol. 87, 1064–1067. doi: 10.1046/j.1365-2745.1999.00424.x
Giongo, A., Crabb, D. B., Davis-Richardson, A. G., Chauliac, D., Mobberley, J. M., Gano, K. A., et al. (2010). PANGEA: pipeline for analysis of next generation amplicons. ISME J. 4, 852–861. doi: 10.1038/ismej.2010.16
Grossart, H. P., Frindte, K., Dziallas, C., Eckert, W., and Tang, K. W. (2011). Microbial methane production in oxygenated water column of an oligotrophic lake. Proc. Nat. Acad. Sci. U.S.A. 108, 19657–19661. doi: 10.1073/pnas.1110716108
Humbert, J. F., Dorigo, U., Cecchi, P., Le Berre, B., Debroas, D., and Bouvy, M. (2009). Comparison of the structure and composition of bacterial communities from temperate and tropical freshwater ecosystems. Environ. Microbiol. 11, 2339–2350. doi: 10.1111/j.1462-2920.2009.01960.x
Kotai, J. (1972). Instructions for preparation of modified nutrient solution Z8 for algae. Norw. Inst. Water Res. Oslo 11, 5.
Landa, M., Cottrell, M., Kirchman, D., Kaiser, K., Medeiros, P., Tremblay, L., et al. (2014). Phylogenetic and structural response of heterotrophic bacteria to dissolved organic matter of different chemical composition in a continuous culture study. Environ. Microbiol. 16, 1668–1681. doi: 10.1111/1462-2920.12242
Leloup, M., Nicolau, R., Pallier, V., Yéprémian, C., and Feuillade-Cathalifaud, G. (2013). Organic matter produced by algae and cyanobacteria: quantitative and qualitative characterization. J. Enviorn. Sci. 25, 1089–1097. doi: 10.1016/S1001-0742(12)60208-3
Li, H., Xing, P., and Wu, Q. L. (2012). The high resilience of the bacterioplankton community in the face of a catastrophic disturbance by a heavy Microcystis bloom. FEMS Microbiol. Ecol. 82, 192–201. doi: 10.1111/j.1574-6941.2012.01417.x
Louati, I., Pascault, N., Debroas, D., Bernard, C., Humbert, J. F., and Leloup, J. (2015). Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differ according to the cyanobacterial genus. PLoS ONE 10:e0140614. doi: 10.1371/journal.pone.0140614
Ludwig, R. A. (1984). Rhizobium free-living nitrogen fixation occurs in specialized nongrowing cells. Proc. Nat. Acad. Sci. U.S.A. 81, 1566–1569. doi: 10.1073/pnas.81.5.1566
Maruyama, T., Kato, K., and Park, H. D. (2004). Population dynamics of free-living bacteria related to the microcystin-degrading strain Y2 in Lake Suwa and in microcystin amended enrichments. Microbes Environ. 19, 137–146. doi: 10.1264/jsme2.19.137
Massana, R., Murray, A. E., Preston, C. M., and DeLong, E. F. (1997). Vertical distribution and phylogenetic characterization of marine planktonic Archaea in the santa barbara channel. Appl. Environ. Microbiol. 63, 50–56.
Moisander, P. H., Serros, T., Paerl, R. W., Beinart, R. A., and Zehr, J. P. (2014). Gammaproetobacterial diazotrophs and nifH gene expression in surface waters of the South Pacific Ocean. ISME J. 8, 1962–1973. doi: 10.1038/ismej.2014.49
Mulholland, M. R. (2007). The fate of nitrogen fixed by diazotrophs in the ocean. Biogeosciences 4, 37–51. doi: 10.5194/bg-4-37-2007
Parveen, B., Ravet, V., Djediat, C., Mary, I., Quiblier, C., Debroas, D., et al. (2013). Bacterial communities associated with Microcystis colonies differ from free–living communities living in the same ecosystem. Environ. Microbiol. Rep. 5, 716–724. doi: 10.1111/1758-2229.12071
Pascault, N., Loux, V., Derozier, S., Martin, V., Debroas, D., Maloufi, S., et al. (2015). Technical challenges in metatranscriptomic studies applied to the bacterial communities of freshwater ecosystems. Genetica 143, 157–167. doi: 10.1007/s10709-014-9783-4
Pedrós-Alió, C. (2006). Marine microbial diversity: can it be determined? Trends Microbiol. 14, 257–263. doi: 10.1016/j.tim.2006.04.007
Ploug, H., Adam, B., Musat, N., Kalvelage, T., Lavik, G., Wolf-Gladrow, D., et al. (2011). Carbon, nitrogen and O2 fluxes associated with the cyanobacterium Nodularia spumigena in the Baltic Sea. ISME J. 5, 1549–1558. doi: 10.1038/ismej.2011.20
Pruesse, E., Quast, C., Knittel, K., Fuchs, B. M., Ludwig, W., Peplies, J., et al. (2007). SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 35, 7188–7196. doi: 10.1093/nar/gkm864
Sala, M. M., Estrada, M., and Gasol, J. M. (2006). Seasonal changes in the functional diversity of bacterioplankton in contrasting coastal environments of the NW Mediterranean. Aquat. Microb. Ecol. 44, 1–9. doi: 10.3354/ame042001
Santi, C., Bogusz, D., and Franche, C. (2013). Biological nitrogen fixation in non-legume plants. Ann. Bot. 111, 743–767. doi: 10.1093/aob/mct048
Sarmento, H., and Gasol, J. M. (2012). Use of phytoplankton–derived dissolved organic carbon by different types of bacterioplankton. Environ. Microbiol. 14, 2348–2360. doi: 10.1111/j.1462-2920.2012.02787.x
Schauer, M., Balagué, V., Pedrós-Alió, C., and Massana, R. (2003). Seasonal changes in the taxonomic composition of bacterioplankton in a coastal oligotrophic system. Aquat. Microb. Ecol. 31, 163–174. doi: 10.3354/ame031163
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504.
Shao, K., Zhang, L., Wang, Y., Yao, X., Tang, X., Qin, B., et al. (2014). The responses of the taxa composition of particle-attached bacterial community to the decomposition of Microcystis blooms. Sci. Total Environ. 488, 236–242. doi: 10.1016/j.scitotenv.2014.04.101
Shen, H., Niu, Y., Xie, P., Tao, M., and Yang, X. (2011). Morphological and physiological changes in Microcystis aeruginosa as a result of interactions with heterotrophic bacteria. Freshw. Biol. 56, 1065–1080. doi: 10.1111/j.1365-2427.2010.02551.x
Shi, L., Cai, Y., Kong, F., and Yu, Y. (2012). Specific association between bacteria and buoyant Microcystis colonies compared with other bulk bacterial communities in the eutrophic Lake Taihu, China. Enviorn. Microbiol. Rep. 4, 669–678. doi: 10.1111/1758-2229.12001
Sime-Ngando, T. (2014). Environmental bacteriophages: viruses of microbes in aquatic ecosystems. Front. Microbiol. 5:355. doi: 10.3389/fmicb.2014.00355
Stevenson, B. S., and Waterbury, J. B. (2006). Isolation and identification of an epibiotic bacterium associated with heterocystous Anabaena cells. Biol. Bull. 210, 73–77. doi: 10.2307/4134596
Taib, N., Mangot, J. F., Domaizon, I., Bronner, G., and Debroas, D. (2013). Phylogenetic affiliation of SSU rRNA genes generated by massively parallel sequencing: new insights into the freshwater protist diversity. PLoS ONE 8:e58950. doi: 10.1371/journal.pone.0058950
Thioulouse, J., and Dray, S. (2007). Interactive multivariate data analysis in R with the ade4 and ade4TkGUI Packages. J. Stat. Softw. 22, 1–14. doi: 10.18637/jss.v022.i05
Urakawa, H., Ali, J., Ketover, R. D., Talmage, S. D., Garcia, J. C., Campbell, I. S., et al. (2013). Shifts of bacterioplankton metabolic profiles along the salinity gradient in a subtropical estuary. ISRN Oceanogr. 2013:410814. doi: 10.5402/2013/410814
Wommack, K. E., and Colwell, R. R. (2000). Virioplankton: viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 64, 69–114. doi: 10.1128/MMBR.64.1.69-114.2000
Woodhouse, J. N., Kinsela, A. S., Collins, R. N., Bowling, L. C., Honeyman, G. L., Holliday, J. K., et al. (2016). Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lakes. ISME J. 10, 1337–1351. doi: 10.1038/ismej.2015.218
Woodhouse, J. N., Ongley, S. E., Brown, M. V., and Neilan, B. A. (2013). Microbial diversity and diazotrophy associated with the freshwater non-heterocyst forming cyanobacterium Lyngbya robusta. J. Appl. Phycol. 25, 1039–1045. doi: 10.1007/s10811-012-9909-y
Keywords: bacterial communities, cyanobacteria, cultures, natural samples, functional and structural diversity, pyrosequencing, EcoPlates
Citation: Zhu L, Zancarini A, Louati I, De Cesare S, Duval C, Tambosco K, Bernard C, Debroas D, Song L, Leloup J and Humbert J -F (2016) Bacterial Communities Associated with Four Cyanobacterial Genera Display Structural and Functional Differences: Evidence from an Experimental Approach. Front. Microbiol. 7:1662. doi: 10.3389/fmicb.2016.01662
Received: 22 April 2016; Accepted: 05 October 2016;
Published: 24 October 2016.
Edited by:
Hongyue Dang, Xiamen University, ChinaReviewed by:
Hans-Peter Grossart, Leibniz Institute of Freshwater Ecology and Inland Fisheries (IGB), GermanyGuang Gao, Nanjing Institute of Geography and Limnology (CAS), China
Copyright © 2016 Zhu, Zancarini, Louati, De Cesare, Duval, Tambosco, Bernard, Debroas, Song, Leloup and Humbert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-François Humbert, jean-francois.humbert@upmc.fr
†These authors have contributed equally to this work.