Noncoding RNA Regulation of Dopamine Signaling in Diseases of the Central Nervous System
 William T. Carrick
William T. Carrick Brandi Burks1
Brandi Burks1  Murray J. Cairns
Murray J. Cairns Jannet Kocerha
Jannet Kocerha- 1Department of Chemistry, Georgia Southern University, Statesboro, GA, USA
- 2School of Biomedical Sciences and Pharmacy and the Hunter Medical Research Institute, University of Newcastle, Callaghan, NSW, Australia
- 3Schizophrenia Research Institute, Sydney, NSW, Australia
Dopaminergic neurotransmission mediates a majority of the vital central nervous system functions. Disruption of these synaptic events provokes a multitude of neurological pathologies, including Parkinson's, schizophrenia, depression, and addiction. Growing evidence supports a key role for noncoding RNA (ncRNA) regulation in the synapse. This review will discuss the role of both short and long ncRNAs in dopamine signaling, including bioinformatic examination of the pathways they target. Specifically, we focus on the contribution of ncRNAs to dopaminergic dysfunction in neurodegenerative as well as psychiatric disease.
Introduction
Dopamine is a potent neurotransmitter in the central nervous system that governs a diverse panel of neuronal functions. Accumulating studies indicate that noncoding RNAs (ncRNAs) modulate almost all aspects of dopamine signaling, including its receptors, and transporter, as well as differentiation and viability of dopaminergic neurons. The processing and transcription of those implicated ncRNAs are also subject to control. Moreover, there are emerging roles for both short and long ncRNAs (lncRNAs) in synaptic transmission, including miRNAs, and antisense transcripts. This review discusses ncRNA-mediated mechanisms which influence dopaminergic signaling under control and pathogenic conditions.
The diverse classes of ncRNAs target all stages in the lineage from stem cells to mature dopaminoceptive neurons (DN) which express dopamine receptors. One of the first ncRNAs reported to modulate midbrain DN (mDN) differentiation and viability was miR-133b (Kim et al., 2007). A subset of additional miRNAs are also now known to direct DN fate, including miR-132, miR-135a2, and miR-218 (Yang et al., 2012; Anderegg et al., 2013; Baek et al., 2014). Moreover, studies indicate that selected miRNAs, such as miR-132, influence DN maturation, and survival by binding to key neuronal transcription factors, including Nurr1 (Yang et al., 2012). Reports also suggest that miRNA processor genes, such as Dicer, are required for terminal differentiation of embryonic stem cells (ESC) into DN (Kim et al., 2007). Dicer cleaves the mature miRNA from its precursor transcript. Mouse studies revealed that Dicer depletion in dopaminoceptive neurons alters brain morphology and neurobehavioral patterns (Cuellar et al., 2008). Further, ablation of Dicer in mouse mDN led to nearly 90% cell death by 8 weeks of age; however, that level of degeneration was not seen when Dicer was depleted in neurons receiving input from dopamine-transporter expressing cells (Kim et al., 2007). These data suggest that miRNA processing deficits may impact specific classes of mature neurons more potently.
It is now evident that ncRNAs target expression of the receptors which bind dopamine. There are at least 5 reported subtypes of dopamine receptors (D1R-D5R), however, D1R, and D2R are the most abundant (Greengard, 2001; Girault and Greengard, 2004). Protein levels of D1R and D2R are well known to fluctuate during the normal aging process as well as in numerous neuronal pathologies (Hurley et al., 2001, 2003; Kaasinen and Rinne, 2002; Rangel-Barajas et al., 2015). In vitro analysis of varying deletion constructs for the D1R gene identified regions of putative miRNA binding sites within the D1R 3′UTR. It was discovered that a short 94 bp segment of the 3′UTR was adequate for post-transcriptional regulation and contains a miR-142-3p binding site (Tobón et al., 2012). Correspondingly, similar studies have also investigated ncRNA control of D2R. Antisense-knockdown of neuronally-enriched miR-9 and miR-326 provoked upregulated expression of D2R 3′UTR in luciferase based assays (Shi et al., 2014). It is important to note, however, that D1R and D2R pathways are also intricately connected and their receptors can even form heteromers (Hasbi et al., 2009; Perreault et al., 2011a,b, 2014a,b). Thus, a subset of ncRNAs may influence signaling of multiple receptor types.
It is well established that dopamine receptor signaling is synchronized with excitatory and inhibitory responses triggered by other neurotransmitters. The role of ncRNAs in mediating this cross-talk, however, is undefined. Saba et al suggests miR-181a as a candidate with a pleiotropic role in neurotransmission, as its expression is modulated by dopamine while it also targets the glutamate receptor subunit GluA2 (Saba et al., 2012). Notably, glutamatergic transmission emerges through AMPA, Kainate, NMDA, and metabotropic receptor classes, underscoring the complexity of synaptic physiology (Planells-Cases et al., 2006). Orchestration of this extensive neurotransmission network by ncRNAs may reveal enduring mechanistic questions that have stymied drug development for many neurological pathologies.
Dopamine signaling is ubiquitous throughout the brain, however, this pathway also functions in select tissues outside of the central nervous system, including the cardiovascular system and kidney (Lokhandwala and Barrett, 1982; Lokhandwala and Amenta, 1991). Impaired dopamine-signaling in the kidney is a primary contributor to hypertension (Banday and Lokhandwala, 2008; Chugh et al., 2012, 2013). Correspondingly, hypertension is repeatedly linked to neuronal dysfunction, including cognitive deficits in patients with Alzheimer's Disease. Investigating ncRNA regulation of dopamine-mediated hypertension in the kidney could reveal novel origins of neurological disease. A recent study identified five dysregulated miRNAs in renal cells from patients carrying hypertension-associated polymorphisms (SNP) in the D2R gene (Han et al., 2015). Of those 5 misregulated miRNAs in the SNP-DR2 cells, miR-217 expression was directly connected to D2R expression through a TGF β1-Wnt5a-Ror2 feedback loop. In a separate study, Zhang et found that let-7d silenced dopamine-receptor 3 (D3R) in renal cells at the chromatin level through histone-3 –lysine-9 dimethylation (Zhang et al., 2016). However, these studies did not explore whether specific disruption of these miRNAs induce hypertension and associated neuronal damage.
This review focuses on neuronal ncRNA mediators of dopamine signaling, although, it is evident this epigenetic pathway also exerts significant influence in other cell types. We outline a subset of these ncRNAs which are implicated in neurodegenerative and neuropsychiatric pathologies Table 1). Additionally, we bioinformatically examined the pathways targeted by the reported miRNAs. Using Ingenuity Pathway Analysis (IPA), several key dopamine-associated pathways were identified, including Dopamine-DARPP32 in cAMP Signaling, Dopamine Receptor Signaling, Synaptic LTP/LTD, CREB Signaling in Neurons, and Axon Guidance Signaling (Figure 1). Overall, the published findings suggest the entire neurotransmission cascade activated by dopamine, as well as its misregulation, is guided by ncRNAs.
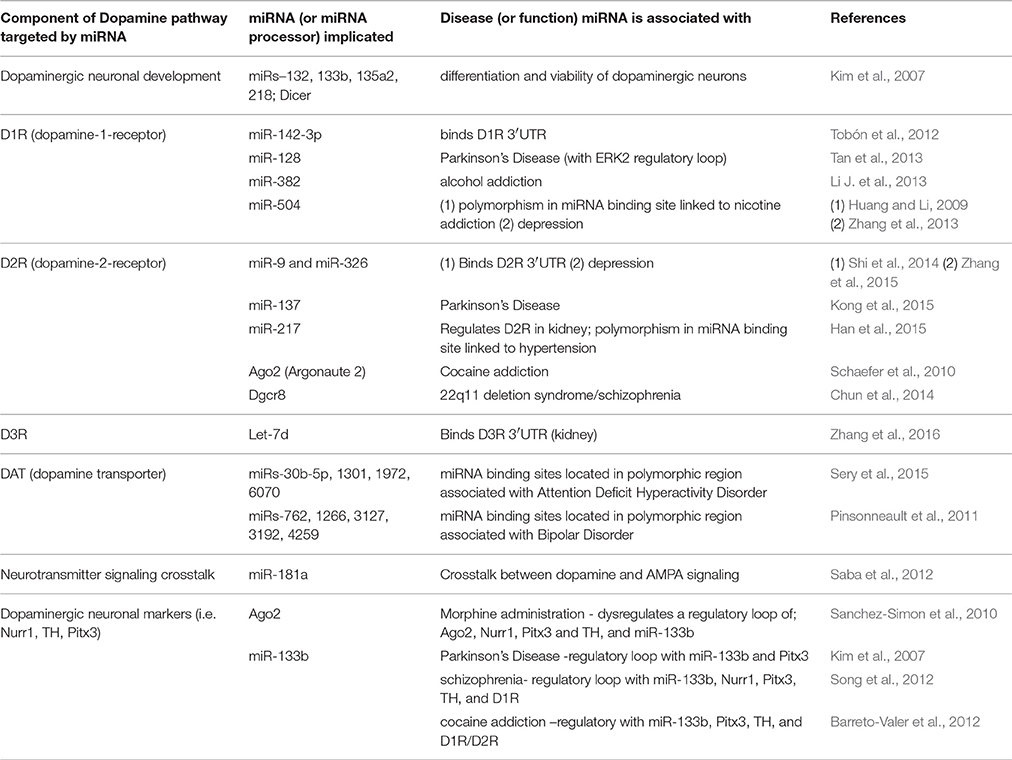
Table 1. Dopamine-responsive miRNAs and their association with neurological functions/disease.
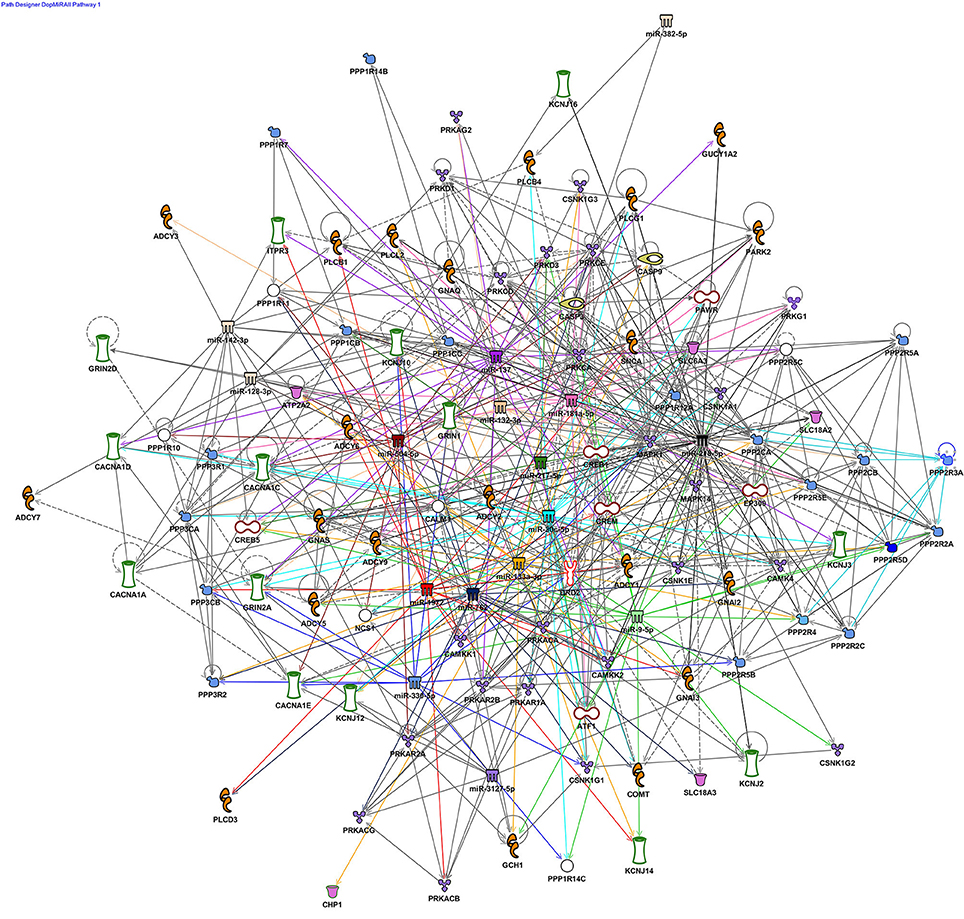
Figure 1. Dopaminergic miRNA nodes in a rich network of connections in synaptic plasticity pathways. Ingenuity pathway analysis (IPA) was used to explore the molecular connectivity predicted between miRNA implicated in dopaminergic development, function and disease (references for all implicated miRNAs examined in this figure are listed in Table 1). miRNA targets, both experimentally validated and predicted by TargetScan were filtered with respect to dopamine-associated canonical pathways and used to generate a network graph. Core miRNA nodes near the center of the graph are color-coded and have colored edges to denote their connections. The largest network defined by the filter was Dopamine-DARPP32 in cAMP Signaling (85 genes) followed by Dopamine Receptor Signaling (40). These networks were also enriched with synaptic plasticity pathways, Synaptic LTP/LTD (43/29 genes), CREB Signaling in Neurons (43 genes), and the Axon Guidance Signaling (27 genes).
miRNA-Mediated Regulation of Dopamine Signaling in Neurological Disease
Dopamine Receptors and miRNAs in Neurological Disease
All five classes of dopamine receptors are putative miRNA targets in neurological pathologies, although D1R and D2R are the most interrogated to date. In neurodegenerative Parkinson's Disease (PD), these patients classically display upregulated expression of D1R and D2R with hypersensitive response to dopamine (Seeman and Niznik, 1990; Kaasinen et al., 2000; Hurley et al., 2001; Gerfen et al., 2002; Zhu et al., 2002; Chu et al., 2004; Hisahara and Shimohama, 2011). Studies by Tan et al revealed that genetic knockout of miR-128 in D1-neurons of wild-type (WT) mice enhances D1R-activation by a dopamine agonist and provokes increased ERK2 expression (Tan et al., 2013). Parallel responses of D1R-hyperexcitablity/ERK2 in miR-128 deficient mice and PD patients suggests this miRNA is an active mechanism underlying PD pathology. In a separate study, a drosophila model which recapitulates PD through common genetic variants of alpha-synuclein uncovered a dysfunctional D2R-miR-137 pathway (Kong et al., 2015). Expression of miR-137 was elevated in the PD drosophila and anti-correlated with D2R protein levels, consistent with miRNA target regulation. Subsequently, miR-137 binding sites in the D2R gene were validated through in vitro analyses.
Targeting of dopamine receptor signaling by physiological stressors that trigger depression and related neuropsychiatric phenotypes is guided by select miRNAs (Zhang et al., 2013, 2015). Early life stress (ELS) can broadly impact normal development of the dopamine system, leading to inadequate coping methods for chronic-unpredictable stress (CUS) and depression in adults. Expression profiles of D1R, miR-504 (a D1R target), and D2R were explored in rats with ELS modeled through maternal deprivation (MD) coupled with exposure to CUS at mature age (MD/CUS) (Zhang et al., 2013). The MD/CUS rats indeed showed significant correlation between D1R, D2R, miR-504 as well as correlation of D2R and miR-504 with depression-related behaviors. In a separate study also conducted by Zhang et al. they subsequently identified several miRNAs (miR-9, miR-200a, miR-141, and miR-326) that target the 3′-UTR of the D2R (Zhang et al., 2015). In vitro analyses confirmed that miR-9 binds the D2R transcript (Zhang et al., 2015). They also found that miR-9, miR-326, and D2R correlated with depressive-like phenotypes in mature MD/CUS rats. In addition, aberrant expression of miR-326 in the MD/CUS rats was normalized with the anti-depressant escitalopram (Zhang et al., 2015). These results not only implicate dopamine-regulated miRNAs in depression, but also suggests these ncRNAs may mediate treatment response to anti-depressants.
Targeting of dopamine receptors by miRNAs was also uncovered in a model for addiction (Li J. et al., 2013). Post-administration of alcohol to rats, the NAc in the basal forebrain displayed repressed levels of miR-382 levels while its bioinformatic target, D1R, was upregulated. Cell-based studies as well as site-specific knockdown and overexpression of miR-382 in rat NAc confirmed it targets the D1R gene (Li J. et al., 2013). Moreover, miR-382 overexpression altered the neuronal excitability responses of D1R and diminished desire to consume alcohol. This report makes the direct connection between miRNA regulation of D1R and neuropsychiatric phenotypes in an animal model.
Disease-associated polymorphic regions in the dopamine receptor genes can impact miRNA binding to their target sites. One such reported example is the D1R rs686 polymorphism linked to nicotine dependence (Huang and Li, 2009). Two miRNA sites were bioinformatically identified within the rs686 region, miR-504 and miR-296. Further, miR-504 regulation of D1R was confirmed in vitro. Notably, miR-504 appears to exhibit stronger binding potential to the rs686 allele compared to the wild-type (WT) allele, which could alter the allelic representation of DR1.
Dopamine Transporter (DAT) and miRNAs in Neurological Disease
The dopamine transporter gene (DAT/SLC6A3) is critical for removal of dopamine in the synapse to halt neurotransmission (German et al., 2015; McHugh and Buckley, 2015). Correspondingly, the disruption of this gene reportedly provokes numerous distinct neuronal pathologies (German et al., 2015; McHugh and Buckley, 2015). Two separate studies investigated miRNA binding in polymorphic regions of the DAT gene. One study examined a 40-base pair 3′UTR region within the DAT transcript linked to Attention Deficit Hyperactivity Disorder (ADHD) (Sery et al., 2015). Bioinformatic analysis of this 40-base pair segment identified putative sites for miRs-1972, 30b-5p, 1301, and 6070 (Sery et al., 2015). A second study focused on a DAT polymorphism linked to bipolar disease, rs27072 (Pinsonneault et al., 2011). The rs27072 analysis revealed binding sequences for miRs-762, 4259, 3192, 3127, and 1266 (Pinsonneault et al., 2011). Collectively, although these findings are only predictive at this point, they provide a framework for future investigation.
In addition to dopamine, DAT can also transport the neurotoxin 6-hydroxydopamine (6-OHDA), eliciting phenotypes which recapitulate PD (Westerlund et al., 2010). 6-OHDA kills dopaminergic neurons, providing a platform to identify and study dopamine-regulated ncRNAs. A subset of miRNAs are reported to mediate 6-OHDA neuronal damage, including miRs-124, 126, 668-3p, let-7d-3p, 3077-3p, 665-5p, 99b-3p, 323-3p, 875, 207, 425-5p, 19b-3p, and 338-3p (Li L. et al., 2013; Kim et al., 2014; Saraiva et al., 2016).
Dopamine Signaling and miRNA Processing Machinery in Neurological Disease
The processing of miRNAs involves several stages, and their disruption is pathogenic (Kocerha et al., 2009, 2015; Chan and Kocerha, 2012; Chun et al., 2014). The primary-miRNA is cleaved by Dgcr8 and Drosha, followed by second cleavage with Dicer into the mature transcript (Bartel, 2004, 2009; Macias et al., 2013). The mature miRNA gets assembled, along with argonaute proteins, into RISC for target gene regulation (Bartel, 2004, 2009). Studies by Schaefer et al found that deficiency of the RISC-associated Argonaute 2 (Ago2) protein in D2R-neurons significantly reduced self-administration of cocaine in mice (Schaefer et al., 2010). This Ago2 mouse model was then used to compile a list of D2R-localized miRNAs which putatively control cocaine addiction.
Ago2 expression is also reportedly regulated by the pain medication morphine (García-Pérez et al., 2013). Acute administration of morphine in rats led to increased Ago2 levels as well as the disruption of a panel of dopaminergic markers such as Nurr1, Pitx3, and TH activity. Pitx3 and TH are mentioned below as part of a conserved miRNA mechanism with miR-133b in neurological disease. Notably, pain relief is mediated by dopamine release in the midbrain, a region where miR-133b functions are reported (Kim et al., 2007; Navratilova et al., 2015). Indeed, a separate study revealed morphine alters miR133b-Pitx3 expression and differentiation of dopaminergic neurons in a zebrafish model (Sanchez-Simon et al., 2010). These results suggest ncRNAs which are regulated by morphine influence neurotransmission.
The gene for the core miRNA processor, Dgcr8, is located within the 22ql1 chromosomal locus, a region persistently associated with schizophrenia and other neurological anomalies (Merico et al., 2015; Zhao et al., 2015). Studies by Chun et al showed that a mouse model for the 22q11 deletion impedes Dgcr8 function, leading to enhanced expression of D2R, and disruption of corticothalamic synaptic transmission (Chun et al., 2014). Moreover, the aberrant D2R signaling provokes a hypersensitive response to antipsychotics used for schizophrenia treatment. Identifying the specific miRNAs impacted by haploinsufficient Dgcr8 processing in the 22q11 mice would facilitate future studies. Importantly, these data also suggest ncRNA mechanisms can influence drug efficacy and medical outcomes.
Conserved Dopamine-Mediated miRNA Mechanisms in Disease; the miR-133b-Pitx3 Example
A shared characteristic of many neurological diagnoses is impaired synaptic transmission; thus, it is pertinent to uncover conserved dopamine-linked ncRNA mechanisms in brain pathologies (Henstridge et al., 2016). One conserved ncRNA pathway was identified through miRNA expression profiles of midbrain dopamine neurons (mDNs) in PD patients (Kim et al., 2007; Briggs et al., 2015). One of the dysregulated miRNAs, miR-133b, controls mDN differentiation through a feedback loop with Pitx3 (Kim et al., 2007). Pitx3 is a transcription factor essential for DN survival (Bergman et al., 2010). A separate study, however, reported that miR-133b expression is not impacted within the substantia nigra region of the midbrain (Schlaudraff et al., 2014). These results suggest the presence of site-specific miRNA mechanisms, providing an additional layer of dopaminergic influence.
The Pitx3-miR-133b pathway associated with PD is also implicated in the neuropsychiatric disorder schizophrenia. Song et al reported that Pitx3-miR-133b is dysregulated in mice with schizophrenia phenotypes provoked through overexpression of the stress protein heme-oxygenase-1 (HO-1) (Song et al., 2012). Moreover, these mice exhibit disrupted profiles of dopaminergic markers D1R, Nurr1 and tyrosine hydroxylase (TH), as well as neurotransmitters serotonin and dopamine. Parallel changes of dopamine and serotonin in this model is consistent with extensive cross-talk reported in synaptic signaling (Henstridge et al., 2016). Further, an independent study found this miRNA pathway is altered in another neuropsychiatric disorder, cocaine addiction (Barreto-Valer et al., 2012). Zebrafish embryos were exposed to cocaine followed by gene expression analysis at various developmental timepoints. In addition to dysregulated levels of miR-133b, Pitx3, D1R, TH in these embryos, there were also significant changes in D2R (Barreto-Valer et al., 2012). Collectively, these data suggest subsets of epigenetic mechanisms are conserved across neurological diseases and, further, may contribute to a general trend of synaptic dysfunction in these pathologies.
lncRNA Regulation of Dopamine Signaling in the Brain
Although miRNAs are the most studied to date, the detection of neuronal lncRNAs is expanding. One of the first annotated lncRNAs in the central nervous system, BC1, mediates D2R expression in vivo and related electrophysiological responses (Wang et al., 2002; Centonze et al., 2007; Zhong et al., 2009; Maccarrone et al., 2010). BC1 knockout mice displayed significant upregulation of D2R protein in the striatum and potentiated responses to D2R agonists (Centonze et al., 2007). Moreover, although BC1 was reported to act as a translational repressor, that is not the mechanism in which it regulates D2R (Wang et al., 2002; Centonze et al., 2007). This study suggests BC1 may have additional functions that have yet to be investigated.
Recently, Carrieri et al showed that a lncRNA antisense to the Uchl1 gene (AS-Uchl1) is significantly repressed in dopaminergic neurons of PD models (Carrieri et al., 2015). The AS-Uchl1 gene is regulated by Nurr1, a core transcription factor involved in the maturation and viability of dopamine neurons (Carrieri et al., 2015). Furthermore, the AS-Uchl1/PARK5 locus is linked to certain populations of PD patients, suggesting an intricate regulatory loop between dopamine-responsive lncRNAs and disease susceptibility genes (Belin and Westerlund, 2008).
Neurodegeneration may also be controlled by lncRNAs encoded within viral transcripts. Kuan et al showed that p137, a ncRNA transcribed from the human cytomegaloviral β2.7 gene, is neuroprotective against OHDA-induced cell damage (Kuan et al., 2012). Compellingly, p137 prevented progression of PD phenotypes within dopaminergic neurons for both in vitro and in vivo OHDA models. In addition to the miRNAs discussed previously, these data suggest that a diverse range of ncRNA classes influence neurodegenerative pathways. It is also likely these ncRNAs may regulate each other through feedback loops.
Investigation of midbrain dopamine neurons (mDN), which are primary responders to drugs of abuse, revealed lncRNA modulators of addiction phenotypes (Bannon et al., 2015). Bannon et al identified a subset of dysregulated lncRNAs in mDN from post-mortem tissue of cocaine addicts (Bannon et al., 2015). One of those misregulated lncRNAs is an antisense transcript to the gene for tumor necrosis factor receptor-associated factor 3-interacting protein 2 (TRAF3IP2). The antisense RNA was localized to the nucleus of the neurons compared to the global cellular distribution of TRAF3IP2, suggestive of epigenetic control at the chromatin level.
A recent report now also suggests Gomafu, a schizophrenia associated lncRNA, modulates dopaminergic transmission (Ip et al., 2016). In Gomafu knockout (KO) mice, a significant increase in dopamine levels was detected in the brain after exposure to the psychostimulant methamphetamine (MAP). Notably, the increased dopamine was correlated with hyperactivity of the KO mice, as measured by open field and light-dark transition tests (Ip et al., 2016). This study directly links lncRNA regulation of dopamine signaling with neurobehavioral phenotypes, although the precise mechanism is not yet elucidated.
Conclusion
Epigenetic regulation of dopamine signaling by ncRNAs is a vital component to neuronal homeostasis. Correspondingly, impaired dopaminergic transmission provokes a range of cognitive, motor, and behavioral phenotypes in CNS disorders. The untapped pharmacological potential of ncRNA modulators to normalize aberrant synaptic transmission provides a wide-open opportunity for drug discovery efforts. Furthermore, as the dopamine pathway is extensive and partially driven by undefined mechanisms, the pleiotropic capacity of many implicated ncRNAs may enhance their efficacy.
Author Contributions
WC, BB, MC, and JK all wrote this manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Anderegg, A., Lin, H. P., Chen, J. A., Caronia-Brown, G., Cherepanova, N., Yun, B., et al. (2013). An Lmx1b-miR135a2 regulatory circuit modulates Wnt1/Wnt signaling and determines the size of the midbrain dopaminergic progenitor pool. PLoS Genet. 9:e1003973. doi: 10.1371/journal.pgen.1003973
Baek, S., Choi, H., and Kim, J. (2014). Ebf3-miR218 regulation is involved in the development of dopaminergic neurons. Brain Res. 1587, 23–32. doi: 10.1016/j.brainres.2014.08.059
Banday, A. A., and Lokhandwala, M. F. (2008). Dopamine receptors and hypertension. Curr. Hypertens. Rep. 10, 268–275.
Bannon, M. J., Savonen, C. L., Jia, H., Dachet, F., Halter, S. D., Schmidt, C. J., et al. (2015). Identification of long noncoding RNAs dysregulated in the midbrain of human cocaine abusers. J. Neurochem. 135, 50–59. doi: 10.1111/jnc.13255
Barreto-Valer, K., López-Bellido, R., Macho Sánchez-Simón, F., and Rodríguez, R. E. (2012). Modulation by cocaine of dopamine receptors through miRNA-133b in zebrafish embryos. PLoS ONE 7:e52701. doi: 10.1371/journal.pone.0052701
Bartel, D. P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136, 215–233. doi: 10.1016/j.cell.2009.01.002
Belin, A. C., and Westerlund, M. (2008). Parkinson's disease: a genetic perspective. FEBS J. 275, 1377–1383. doi: 10.1111/j.1742-4658.2008.06301.x
Bergman, O., Håkansson, A., Westberg, L., Nordenström, K., Carmine Belin, A., Sydow, O., et al. (2010). PITX3 polymorphism is associated with early onset Parkinson's disease. Neurobiol. Aging 31, 114–117. doi: 10.1016/j.neurobiolaging.2008.03.008
Briggs, C. E., Wang, Y., Kong, B., Woo, T. U., Iyer, L. K., and Sonntag, K. C. (2015). Midbrain dopamine neurons in Parkinson's disease exhibit a dysregulated miRNA and target-gene network. Brain Res. 1618, 111–121. doi: 10.1016/j.brainres.2015.05.021
Carrieri, C., Forrest, A. R., Santoro, C., Persichetti, F., Carninci, P., Zucchelli, S., et al. (2015). Expression analysis of the long non-coding RNA antisense to Uchl1 (AS Uchl1) during dopaminergic cells' differentiation in vitro and in neurochemical models of Parkinson's disease. Front. Cell. Neurosci. 9:114. doi: 10.3389/fncel.2015.00114
Centonze, D., Rossi, S., Napoli, I., Mercaldo, V., Lacoux, C., Ferrari, F., et al. (2007). The brain cytoplasmic RNA BC1 regulates dopamine D2 receptor-mediated transmission in the striatum. J. Neurosci. 27, 8885–8892. doi: 10.1523/JNEUROSCI.0548-07.2007
Chan, A. W., and Kocerha, J. (2012). The Path to microRNA therapeutics in psychiatric and neurodegenerative disorders. Front. Genet. 3:82. doi: 10.3389/fgene.2012.00082
Chu, C. T., Levinthal, D. J., Kulich, S. M., Chalovich, E. M., and DeFranco, D. B. (2004). Oxidative neuronal injury. The dark side of ERK1/2. Eur. J. Biochem. 271, 2060–2066. doi: 10.1111/j.1432-1033.2004.04132.x
Chugh, G., Lokhandwala, M. F., and Asghar, M. (2012). Altered functioning of both renal dopamine D1 and angiotensin II type 1 receptors causes hypertension in old rats. Hypertension 59, 1029–1036. doi: 10.1161/HYPERTENSIONAHA.112.192302
Chugh, G., Pokkunuri, I., and Asghar, M. (2013). Renal dopamine and angiotensin II receptor signaling in age-related hypertension. Am. J. Physiol. Renal Physiol. 304, F1–F7. doi: 10.1152/ajprenal.00441.2012
Chun, S., Westmoreland, J. J., Bayazitov, I. T., Eddins, D., Pani, A. K., Smeyne, R. J., et al. (2014). Specific disruption of thalamic inputs to the auditory cortex in schizophrenia models. Science 344, 1178–1182. doi: 10.1126/science.1253895
Cuellar, T. L., Davis, T. H., Nelson, P. T., Loeb, G. B., Harfe, B. D., Ullian, E., et al. (2008). Dicer loss in striatal neurons produces behavioral and neuroanatomical phenotypes in the absence of neurodegeneration. Proc. Natl. Acad. Sci. U.S.A. 105, 5614–5619. doi: 10.1073/pnas.0801689105
García-Pérez, D., Sáez-Belmonte, F., Laorden, M. L., Núñez, C., and Milanés, M. V. (2013). Morphine administration modulates expression of Argonaute 2 and dopamine-related transcription factors involved in midbrain dopaminergic neurons function. Br. J. Pharmacol. 168, 1889–1901. doi: 10.1111/bph.12083
Gerfen, C. R., Miyachi, S., Paletzki, R., and Brown, P. (2002). D1 dopamine receptor supersensitivity in the dopamine-depleted striatum results from a switch in the regulation of ERK1/2/MAP kinase. J. Neurosci. 22, 5042–5054.
German, C. L., Baladi, M. G., McFadden, L. M., Hanson, G. R., and Fleckenstein, A. E. (2015). Regulation of the dopamine and vesicular monoamine transporters: pharmacological targets and implications for disease. Pharmacol. Rev. 67, 1005–1024. doi: 10.1124/pr.114.010397
Girault, J. A., and Greengard, P. (2004). The neurobiology of dopamine signaling. Arch. Neurol. 61, 641–644. doi: 10.1001/archneur.61.5.641
Greengard, P. (2001). The neurobiology of dopamine signaling. Biosci. Rep. 21, 247–269. doi: 10.1023/A:1013205230142
Han, F., Konkalmatt, P., Chen, J., Gildea, J., Felder, R. A., Jose, P. A., et al. (2015). MiR-217 mediates the protective effects of the dopamine D2 receptor on fibrosis in human renal proximal tubule cells. Hypertension 65, 1118–1125. doi: 10.1161/HYPERTENSIONAHA.114.05096
Hasbi, A., Fan, T., Alijaniaram, M., Nguyen, T., Perreault, M. L., O'Dowd, B. F., et al. (2009). Calcium signaling cascade links dopamine D1-D2 receptor heteromer to striatal BDNF production and neuronal growth. Proc. Natl. Acad. Sci. U.S.A. 106, 21377–21382. doi: 10.1073/pnas.0903676106
Henstridge, C. M., Pickett, E., and Spires-Jones, T. L. (2016). Synaptic pathology: a shared mechanism in neurological disease. Ageing Res. Rev. 28, 72–84. doi: 10.1016/j.arr.2016.04.005
Hisahara, S., and Shimohama, S. (2011). Dopamine receptors and Parkinson's disease. Int. J. Med. Chem. 2011:403039. doi: 10.1155/2011/403039
Huang, W., and Li, M. D. (2009). Differential allelic expression of dopamine D1 receptor gene (DRD1) is modulated by microRNA miR-504. Biol. Psychiatry 65, 702–705. doi: 10.1016/j.biopsych.2008.11.024
Hurley, M. J., Mash, D. C., and Jenner, P. (2001). Dopamine D(1) receptor expression in human basal ganglia and changes in Parkinson's disease. Brain Res. Mol. Brain Res. 87, 271–279.
Hurley, M. J., Mash, D. C., and Jenner, P. (2003). Markers for dopaminergic neurotransmission in the cerebellum in normal individuals and patients with Parkinson's disease examined by RT-PCR. Eur. J. Neurosci. 18, 2668–2672. doi: 10.1046/j.1460-9568.2003.02963.x
Ip, J. Y., Sone, M., Nashiki, C., Pan, Q., Kitaichi, K., Yanaka, K., et al. (2016). Gomafu lncRNA knockout mice exhibit mild hyperactivity with enhanced responsiveness to the psychostimulant methamphetamine. Sci. Rep. 6:27204. doi: 10.1038/srep27204
Kaasinen, V., and Rinne, J. O. (2002). Functional imaging studies of dopamine system and cognition in normal aging and Parkinson's disease. Neurosci. Biobehav. Rev. 26, 785–793.
Kaasinen, V., Ruottinen, H. M., Nagren, K., Lehikoinen, P., Oikonen, V., and Rinne, J. O. (2000). Upregulation of putaminal dopamine D2 receptors in early Parkinson's disease: a comparative PET study with [11C] raclopride and [11C]N-methylspiperone. J. Nucl. Med. 41, 65–70.
Kim, J., Inoue, K., Ishii, J., Vanti, W. B., Voronov, S. V., Murchison, E., et al. (2007). A MicroRNA feedback circuit in midbrain dopamine neurons. Science 317, 1220–1224. doi: 10.1126/science.1140481
Kim, W., Lee, Y., McKenna, N. D., Yi, M., Simunovic, F., Wang, Y., et al. (2014). miR-126 contributes to Parkinson's disease by dysregulating the insulin-like growth factor/phosphoinositide 3-kinase signaling. Neurobiol. Aging 35, 1712–1721. doi: 10.1016/j.neurobiolaging.2014.01.021
Kocerha, J., Dwivedi, Y., and Brennand, K. J. (2015). Noncoding RNAs and neurobehavioral mechanisms in psychiatric disease. Mol. Psychiatry 20, 677–684. doi: 10.1038/mp.2015.30
Kocerha, J., Kauppinen, S., and Wahlestedt, C. (2009). microRNAs in CNS disorders. Neuromolecular Med. 11, 162–172. doi: 10.1007/s12017-009-8066-1
Kong, Y., Liang, X., Liu, L., Zhang, D., Wan, C., Gan, Z., et al. (2015). High Throughput sequencing identifies MicroRNAs mediating alpha-synuclein toxicity by targeting neuroactive-ligand receptor interaction pathway in early stage of Drosophila Parkinson's Disease Model. PLoS ONE 10:e0137432. doi: 10.1371/journal.pone.0137432
Kuan, W. L., Poole, E., Fletcher, M., Karniely, S., Tyers, P., Wills, M., et al. (2012). A novel neuroprotective therapy for Parkinson's disease using a viral noncoding RNA that protects mitochondrial complex I activity. J. Exp. Med. 209, 1–10. doi: 10.1084/jem.20111126
Li, J., Li, J., Liu, X., Qin, S., Guan, Y., Liu, Y., et al. (2013). MicroRNA expression profile and functional analysis reveal that miR-382 is a critical novel gene of alcohol addiction. EMBO Mol. Med. 5, 1402–1414. doi: 10.1002/emmm.201201900
Li, L., Chen, H. Z., Chen, F. F., Li, F., Wang, M., Wang, L., et al. (2013). Global microRNA expression profiling reveals differential expression of target genes in 6-hydroxydopamine-injured MN9D cells. Neuromolecular Med. 15, 593–604. doi: 10.1007/s12017-013-8244-z
Lokhandwala, M. F., and Amenta, F. (1991). Anatomical distribution and function of dopamine receptors in the kidney. FASEB J. 5, 3023–3030.
Lokhandwala, M. F., and Barrett, R. J. (1982). Cardiovascular dopamine receptors: physiological, pharmacological and therapeutic implications. J. Auton. Pharmacol. 2, 189–215.
Maccarrone, M., Rossi, S., Bari, M., De Chiara, V., Rapino, C., Musella, A., et al. (2010). Abnormal mGlu 5 receptor/endocannabinoid coupling in mice lacking FMRP and BC1 RNA. Neuropsychopharmacology 35, 1500–1509. doi: 10.1038/npp.2010.19
Macias, S., Cordiner, R. A., and Cáceres, J. F. (2013). Cellular functions of the microprocessor. Biochem. Soc. Trans. 41, 838–843. doi: 10.1042/BST20130011
McHugh, P. C., and Buckley, D. A. (2015). The structure and function of the dopamine transporter and its role in CNS diseases. Vitam. Horm. 98, 339–369. doi: 10.1016/bs.vh.2014.12.009
Merico, D., Zarrei, M., Costain, G., Ogura, L., Alipanahi, B., Gazzellone, M. J., et al. (2015). Whole-genome sequencing suggests schizophrenia risk mechanisms in humans with 22q11.2 Deletion Syndrome. G3 (Bethesda) 5, 2453–2461. doi: 10.1534/g3.115.021345
Navratilova, E., Atcherley, C. W., and Porreca, F. (2015). Brain circuits encoding reward from pain relief. Trends Neurosci. 38, 741–750. doi: 10.1016/j.tins.2015.09.003
Perreault, M. L., Hasbi, A., O'Dowd, B. F., and George, S. R. (2011a). The dopamine d1-d2 receptor heteromer in striatal medium spiny neurons: evidence for a third distinct neuronal pathway in Basal Ganglia. Front. Neuroanat. 5:31. doi: 10.3389/fnana.2011.00031
Perreault, M. L., Hasbi, A., O'Dowd, B. F., and George, S. R. (2014a). Heteromeric dopamine receptor signaling complexes: emerging neurobiology and disease relevance. Neuropsychopharmacology 39, 156–168. doi: 10.1038/npp.2013.148
Perreault, M. L., O'Dowd, B. F., and George, S. R. (2011b). Dopamine receptor homooligomers and heterooligomers in schizophrenia. CNS Neurosci. Ther. 17, 52–57. doi: 10.1111/j.1755-5949.2010.00228.x
Perreault, M. L., O'Dowd, B. F., and George, S. R. (2014b). Dopamine D(1)-D(2) receptor heteromer regulates signaling cascades involved in addiction: potential relevance to adolescent drug susceptibility. Dev. Neurosci. 36, 287–296. doi: 10.1159/000360158
Pinsonneault, J. K., Han, D. D., Burdick, K. E., Kataki, M., Bertolino, A., Malhotra, A. K., et al. (2011). Dopamine transporter gene variant affecting expression in human brain is associated with bipolar disorder. Neuropsychopharmacology 36, 1644–1655. doi: 10.1038/npp.2011.45
Planells-Cases, R., Lerma, J., and Ferrer-Montiel, A. (2006). Pharmacological intervention at ionotropic glutamate receptor complexes. Curr. Pharm. Des. 12, 3583–3596.
Rangel-Barajas, C., Coronel, I., and Florán, B. (2015). Dopamine receptors and neurodegeneration. Aging Dis. 6, 349–368. doi: 10.14336/AD.2015.0330
Saba, R., Störchel, P. H., Aksoy-Aksel, A., Kepura, F., Lippi, G., Plant, T. D., et al. (2012). Dopamine-regulated microRNA MiR-181a controls GluA2 surface expression in hippocampal neurons. Mol. Cell. Biol. 32, 619–632. doi: 10.1128/MCB.05896-11
Sanchez-Simon, F. M., Zhang, X. X., Loh, H. H., Law, P. Y., and Rodriguez, R. E. (2010). Morphine regulates dopaminergic neuron differentiation via miR-133b. Mol. Pharmacol. 78, 935–942. doi: 10.1124/mol.110.066837
Saraiva, C., Paiva, J., Santos, T., Ferreira, L., and Bernardino, L. (2016). MicroRNA-124 loaded nanoparticles enhance brain repair in Parkinson's disease. J. Control. Release 235, 291–305. doi: 10.1016/j.jconrel.2016.06.005
Schaefer, A., Im, H.-I., Venø, M. T., Fowler, C. D., Min, A., Intrator, A., et al. (2010). Argonaute 2 in dopamine 2 receptor-expressing neurons regulates cocaine addiction. J. Exp. Med. 207, 1843–1851. doi: 10.1084/jem.20100451
Schlaudraff, F., Grundemann, J., Fauler, M., Dragicevic, E., Hardy, J., and Liss, B. (2014). Orchestrated increase of dopamine and PARK mRNAs but not miR-133b in dopamine neurons in Parkinson's disease. Neurobiol. Aging 35, 2302–2315. doi: 10.1016/j.neurobiolaging.2014.03.016
Seeman, P., and Niznik, H. B. (1990). Dopamine receptors and transporters in Parkinson's disease and schizophrenia. FASEB J. 4, 2737–2744.
Sery, O., Paclt, I., Drtilkova, I., Theiner, P., Kopeckova, M., Zvolsky, P., et al. (2015). A 40-bp VNTR polymorphism in the 3′-untranslated region of DAT1/SLC6A3 is associated with ADHD but not with alcoholism. Behav. Brain Funct. 11, 21. doi: 10.1186/s12993-015-0066-8
Shi, S., Leites, C., He, D., Schwartz, D., Moy, W., Shi, J., et al. (2014). MicroRNA-9 and microRNA-326 regulate human dopamine D2 receptor expression, and the microRNA-mediated expression regulation is altered by a genetic variant. J. Biol. Chem. 289, 13434–13444. doi: 10.1074/jbc.M113.535203
Song, W., Zukor, H., Lin, S. H., Hascalovici, J., Liberman, A., Tavitian, A., et al. (2012). Schizophrenia-like features in transgenic mice overexpressing human HO-1 in the astrocytic compartment. J. Neurosci. 32, 10841–10853. doi: 10.1523/JNEUROSCI.6469-11.2012
Tan, C. L., Plotkin, J. L., Venø, M. T., von Schimmelmann, M., Feinberg, P., Mann, S., et al. (2013). MicroRNA-128 governs neuronal excitability and motor behavior in mice. Science 342, 1254–1258. doi: 10.1126/science.1244193
Tobón, K. E., Chang, D., and Kuzhikandathil, E. V. (2012). MicroRNA 142-3p mediates post-transcriptional regulation of D1 dopamine receptor expression. PLoS ONE 7:e49288. doi: 10.1371/journal.pone.0049288
Wang, H., Iacoangeli, A., Popp, S., Muslimov, I. A., Imataka, H., Sonenberg, N., et al. (2002). Dendritic BC1 RNA: functional role in regulation of translation initiation. J. Neurosci. 22, 10232–10241. doi: 10.1083/jcb.200506006
Westerlund, M., Hoffer, B., and Olson, L. (2010). Parkinson's disease: exit toxins, enter genetics. Prog Neurobiol 90, 146–156. doi: 10.1016/j.pneurobio.2009.11.001
Yang, D., Li, T., Wang, Y., Tang, Y., Cui, H., Tang, Y., et al. (2012). miR-132 regulates the differentiation of dopamine neurons by directly targeting Nurr1 expression. J Cell Sci 125(Pt 7), 1673–1682. doi: 10.1242/jcs.086421
Zhang, Y., Cheng, C., He, D., Shi, W., Fu, C., Wang, X., et al. (2016). Transcriptional gene silencing of dopamine D3 receptor caused by let-7d mimics in immortalized renal proximal tubule cells of rats. Gene 580, 89–95. doi: 10.1016/j.gene.2015.12.071
Zhang, Y., Wang, Y., Wang, L., Bai, M., Zhang, X., and Zhu, X. (2015). Dopamine Receptor D2 and associated microRNAs are involved in stress susceptibility and resistance to escitalopram treatment. Int. J. Neuropsychopharmacol. 18:pyv025. doi: 10.1093/ijnp/pyv025
Zhang, Y., Zhu, X., Bai, M., Zhang, L., Xue, L., and Yi, J. (2013). Maternal deprivation enhances behavioral vulnerability to stress associated with miR-504 expression in nucleus accumbens of rats. PLoS ONE 8:e69934. doi: 10.1371/journal.pone.0069934
Zhao, D., Lin, M., Chen, J., Pedrosa, E., Hrabovsky, A., Fourcade, H. M., et al. (2015). MicroRNA profiling of neurons generated using induced pluripotent stem cells derived from patients with schizophrenia and schizoaffective disorder, and 22q11.2 Del. PLoS ONE 10:e0132387. doi: 10.1371/journal.pone.0132387
Zhong, J., Chuang, S. C., Bianchi, R., Zhao, W., Lee, H., Fenton, A. A., et al. (2009). BC1 regulation of metabotropic glutamate receptor-mediated neuronal excitability. J. Neurosci. 29, 9977–9986. doi: 10.1523/JNEUROSCI.3893-08.2009
Keywords: microRNAs, psychiatric, neurodegeneration, noncoding RNA, dopaminergic, schizophrenia, dopamine, epigenetic
Citation: Carrick WT, Burks B, Cairns MJ and Kocerha J (2016) Noncoding RNA Regulation of Dopamine Signaling in Diseases of the Central Nervous System. Front. Mol. Biosci. 3:69. doi: 10.3389/fmolb.2016.00069
Received: 06 August 2016; Accepted: 07 October 2016;
Published: 25 October 2016.
Edited by:
Ashok Sharma, Augusta University, USAReviewed by:
Pedro José Carlos Rondot Radío, University of Buenos Aires, ArgentinaMichal Mielcarek, Imperial College London, UK
Copyright © 2016 Carrick, Burks, Cairns and Kocerha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jannet Kocerha, rkocerha@georgiasouthern.edu; jkocerha@gmail.com