Motor–language coupling in Huntington’s disease families
 Lucila Kargieman1,2,3†
Lucila Kargieman1,2,3†
 Eduar Herrera1,2,4†
Eduar Herrera1,2,4†
 Sandra Baez1,2,3
Sandra Baez1,2,3
 Adolfo M. García1,2,3,5
Adolfo M. García1,2,3,5
 Martin Dottori1
Martin Dottori1
 Carlos Gelormini1
Carlos Gelormini1
 Facundo Manes1,2,6
Facundo Manes1,2,6
 Oscar Gershanik1,2
Oscar Gershanik1,2  Agustín Ibáñez1,2,3,4*
Agustín Ibáñez1,2,3,4*- 1Laboratory of Experimental Psychology and Neuroscience (LPEN), Institute of Cognitive Neurology (INECO), Favaloro University, Buenos Aires, Argentina
- 2National Scientific and Technical Research Council (CONICET), Buenos Aires, Argentina
- 3UDP-INECO Foundation Core on Neuroscience (UIFCoN), Diego Portales University, Santiago, Chile
- 4Universidad Autónoma del Caribe, Barranquilla, Colombia
- 5School of Languages, National University of Córdoba (UNC), Córdoba, Argentina
- 6Australian Research Council (ARC) Centre of Excellence in Cognition and its Disorders, Sydney, NSW, Australia
Traditionally, Huntington’s disease (HD) has been known as a movement disorder, characterized by motor, psychiatric, and cognitive impairments. Recent studies have shown that motor and action–language processes are neurally associated. The cognitive mechanisms underlying this interaction have been investigated through the action compatibility effect (ACE) paradigm, which induces a contextual coupling of ongoing motor actions and verbal processing. The present study is the first to use the ACE paradigm to evaluate action–word processing in HD patients (HDP) and their families. Specifically, we tested three groups: HDP, healthy first-degree relatives (HDR), and non-relative healthy controls. The results showed that ACE was abolished in HDP as well as HDR, but not in controls. Furthermore, we found that the processing deficits were primarily linguistic, given that they did not correlate executive function measurements. Our overall results underscore the role of cortico-basal ganglia circuits in action–word processing and indicate that the ACE task is a sensitive and robust early biomarker of HD and familial vulnerability.
Introduction
Huntington’s disease (HD) is an autosomal dominant neurodegenerative disorder resulting from the expansion of CAG trinucleotide repeats (36 or more repeats) within a gene on the short arm of chromosome 4, which codes for the protein huntingtin (Ho et al., 2001). HD is clinically diagnosed by symptoms such as chorea, bradykinesia, dystonia, and incoordination (Tröster, 2006). These typically become evident between the ages of 35 and 44, but, depending on the number of CAG repeats, disease onset ranges from childhood to late adulthood (Bates and Jones, 2002). Due to developments in genetics, HD can now be diagnosed in the absence of motor symptoms. However, in pre-HD and early HD stages, the pathology may be present without any of its typical (motor, psychiatric, or otherwise cognitive) signs (Stout et al., 2011), which underscores the need for more sensitive measures. In such cases, damage is usually restricted to basal ganglia structures, especially the caudate nucleus (Harris et al., 1999; Kipps et al., 2005; Kloppel et al., 2008; Henley et al., 2009). As in other neurodegenerative disorders (Ibanez and Manes, 2012), early detection would enable more effective diagnosis and treatment.
Recent reports of other motor disorders in their early-stages have found deficits in verbal domains, including action–language. This is true of progressive supranuclear palsy (Bak et al., 2005), frontotemporal dementia (Rhee et al., 2001; D’Honincthun and Pillon, 2008), Parkinson’s disease, and amyotrophic lateral sclerosis (Neary et al., 2000; Bak et al., 2001; Peran et al., 2003, 2009; Bak and Hodges, 2004; Boulenger et al., 2008). Thus, action–language deficits are an emergent agenda in motor conditions.
In HD, typical motor alterations and cognitive deficits (Bachoud-Levi et al., 2001; Montoya et al., 2006) are accompanied by disorders in verbal production (Podoll et al., 1988; Murray and Lenz, 2001), including reduced verbal fluency (Chenery et al., 2002; Ho et al., 2002; Azambuja et al., 2012), affixation errors (Ullman et al., 1997), and syntactic processing difficulties (Teichmann et al., 2005, 2008). Language comprehension deficits have also been reported (Teichmann et al., 2008; Azambuja et al., 2012). Moreover, late-stage HD has been associated with action–verb generation deficits (Peran et al., 2004), and action naming was found to be the task that best discriminated controls and HD patients (HDP) (Azambuja et al., 2012). Nevertheless, the relationship between action–language and motor action remains poorly explored.
Nowadays, research into action–verb processing has focused on frontal and motor cortices (Federmeier et al., 2000; Pulvermuller et al., 2001; Yokoyama et al., 2006; Boulenger et al., 2008; Cappelletti et al., 2008; Kemmerer et al., 2008; Tomasino et al., 2008). However, the basal ganglia may also be involved in both motor representation and initiation of semantic integration during action–verb processing (Cardona et al., 2013). In this sense, a crucial unresolved issue is whether subcortical motor networks are actively engaged in action–language. The present study addresses this question by examining motor–language coupling during sentence comprehension in HDP. Furthermore, we explore this sentence comprehension mechanism in asymptomatic HD patients’ relatives (HDR). Cognitive impairments have been documented in HDP during pre-clinical stages near the onset of disease (Lawrence et al., 1998a). Moreover, biological, motor, and cognitive vulnerability have also been reported in HDR (Markianos et al., 2008; Dorsey, 2012). Building on these findings, we evaluated (a) whether motor–language coupling is affected by subcortical motor affectation, and (b) whether this coupling is a marker of early-stage HD or HD vulnerability. To the best of our knowledge, this is the first study on motor–language coupling in HDP and their relatives.
Materials and Methods
Subjects
In total, 73 subjects received a full protocol evaluation (see Table 1 for demographic and clinical data).
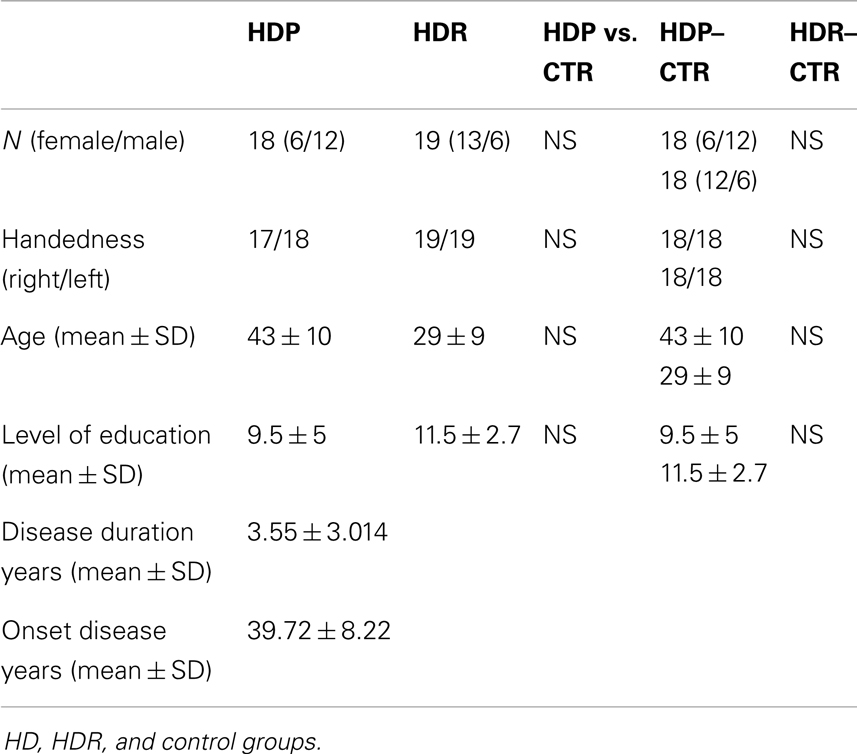
Table 1. Demographic and clinical data.
Huntington’s disease patients group
The HDP group consisted of 18 symptomatic patients genetically and clinically diagnosed with HD. They presented both clinical and cognitive manifestations as well as a positive family history of HD, according to the criteria proposed by Folstein et al. (1986). Patients with different neurological or psychiatric signs, or structural brain abnormalities compatible with diagnoses other than HD, were excluded from the study. To evaluate daily-life functional capacity in HDP, we used the Shoulson–Fahn Functional Capacity Scale (HDFCS) (Shoulson and Fahn, 1979). This instrument measures independence in daily activities such as dressing, eating, managing personal finances, and engagement in occupation. Functional capacity is ranked from 0 to 13 points, the latter score representing the most independent level of function (Shoulson and Fahn, 1979). Most of the participants in the HDP group were off medication (73%). Only five patients were under fluoxetine, clonazepam, and tetrabenazine treatment. Patients and relatives were recruited from the small rural town of Juan de Acosta, Colombia, a region possessing the second largest concentration of individuals with HD in the world (Pradilla et al., 2003).
Huntington’s disease relatives group
The HDR group was composed of 19 subjects with a positive family history of HD to the first-degree of consanguinity. They did not present any HD symptoms, and had not been diagnosed with HD or other neuropsychiatric diseases. Both the HDP and the HDR groups underwent a neurological examination and were assessed using the unified Huntington’s disease rating scale (UHDRS) (Huntington Study Group, 1996). UHDRS total motor scores were >5 for HDP and <5 for HDR, resembling the scores reported by Tabrizi et al. (2009). Patients and relatives had no history of any other major neurological illness, psychiatric disorders, or alcohol/drug abuse.
Control groups
Thirty-seven healthy comparison subjects were recruited and assigned to two control groups. One (n = 18) was matched with HDP and the other (n = 19) with HDR. Matching criteria were sex, age (±2 years), and years of education (±2 years). The HDP control group (HDP–CTR) had a mean age of 43.22 (±10.530), and a mean educational level of 10.16 (±4.218) years of schooling. For the HDR control group (HDR–CTR), the mean age was 29.50 (±10.245) and the mean level of education was 11.44 (±2.661) years of schooling. None of the subjects had a history of neurodegenerative disease, psychiatric disorders, or drug abuse. The demographic, clinical, and neuropsychological characteristics of the participants are summarized in Table 1. All participants gave written informed consent in agreement with the Helsinki declaration. Also, the study was approved by the Ethics Committee of the Institute of Cognitive Neurology.
Clinical and Neuropsychological Assessment
All participants completed a series of psychiatric questionnaires to establish a clinical symptom profile. The Beck depression inventory-II (BDI-II) (Beck et al., 1996) was used to rate depression. Anxiety symptoms were assessed by means of the Hamilton anxiety rating scale (HAM-A) (Hamilton, 1959). In addition, the Montreal cognitive assessment (MOCA) (Nasreddine et al., 2005) was used to assess the participants’ overall cognitive state, including short-term memory, visuospatial/executive skills (e.g., alternation, phonetic fluency, and abstraction), attention, working memory, language, and orientation.
Furthermore, all participants completed the INECO frontal screening (IFS) test (Torralva et al., 2009), which has been shown to successfully detect executive dysfunction (Torralva et al., 2009; Gleichgerrcht et al., 2011). The IFS includes the following eight subtests: (1) motor programing (Luria series, “fist, edge, palm”); (2) conflicting instructions (subjects are asked to hit the table once when the examiner hits it twice, or vice versa); (3) motor inhibitory control; (4) numerical working memory (backward digit span); (5) verbal working memory (months backwards); (6) spatial working memory (modified Corsi tapping test); (7) abstraction capacity (inferring the meaning of proverbs); and (8) verbal inhibitory control (modified Hayling test). The maximum possible score on the IFS is 30 points. Additionally, participants were evaluated with the Wechsler abbreviated scale of intelligence (WASI). This test includes vocabulary and similarities subtests and provides a verbal estimated IQ (Wechsler, 1999). Finally, participants were also administered the WAIS-III similarities subtest (Wechsler, 1997) to evaluate abstract thinking, and the Stroop test (Treisman and Fearnley, 1969) to assess mental speed, selective attention, and inhibitory control. To control for the influence of clinical symptoms (depression and anxiety) or cognitive state on experimental tasks, we applied ANCOVA tests adjusted for BDI-II, HAM-A, and MOCA scores. We report only those effects that were still significant after covariation.
Experimental Tasks
Semantic association of nouns
To assess the semantic association of nouns, we employed the picture version of the Pyramids and Palm Trees Test (PPT) (Howard et al., 1992). The PPT consists of 52 triplets of pictures depicting different objects. Each triplet is composed of a cue object–picture (e.g., spectacles) and two semantically related pictures (e.g., eye and ear). Participants are asked to point to the picture that is most closely related to the cue. Their goal is to discover the relation between the cue and the response picture, which varies across trials.
Action–verb processing
To assess selective action–verb processing, we used the picture version of the kissing and dancing test (KDT) (Bak and Hodges, 2003). The KDT uses 52 triads of images to assess access to semantic representations of verbs. It has proven useful in detecting subtle impairments in other subcortical motor diseases, such as Parkinson’s disease (Cardona et al., 2013; Ibanez et al., 2013).
Action sentence compatibility effect
We evaluated the interaction between language and motor processes using the action compatibility effect (ACE) paradigm. The ACE paradigm is well-suited for this study, since it recruits both motor and semantic brain areas, and has proved sensitive to deficits in other subcortical motor disorders (e.g., Parkinson’s disease) (Aravena et al., 2010; Ibanez et al., 2013). Moreover, the ACE task is unaffected by peripheral motor impairments (Cardona et al., 2014), which further underscores its relevance to the present study. Participants listened to sentences that described an action performed with the hand in a particular shape (open: OH, n = 52, or closed: CH, n = 52), as well as neutral sentences alluding to non-manual actions (neutral: N, n = 52). Participants indicated as quickly as possible when they understood each sentence by pressing a button with a pre-assigned hand-shape (open and closed; counterbalanced in two blocks). The combination of response type and sentence-type generates compatible (OH sentence and OH response or CH sentence and CH response), incompatible (OH sentence and CH response or vice versa), and neutral (N sentence with either response) trials. The ACE is defined as a longer reaction time (RT) in the incompatible than in the compatible conditions (Glenberg and Kaschak, 2002; Masumoto et al., 2004; Borreggine and Kaschak, 2006; Glenberg, 2006; Glenberg et al., 2008a,b; Aravena et al., 2010).
All participants performed the task with their dominant hand, although both hands were positioned in the required shape. By controlling the position of both hands, we controlled for possible bilateral manual interference, since posture modulates semantic processing (Glenberg et al., 2008b; Lindeman et al., 2008; van Elk et al., 2008; Badets et al., 2010). To ensure that all participants had understood the meaning of the sentences, they were asked to complete an offline questionnaire after finishing the task.
Data Analysis
For statistical analysis, we used the repeated measures analysis of variance (ANOVA) and χ2 for neuropsychological assessment. In the ACE paradigm, mean RTs were calculated for each subject for each type of trial (compatible, incompatible, and neutral) and each type of sentence (OH, CH, and N). Single trials eliciting outlier values with RTs outside 2.5 SD were excluded from the analyses. A mixed-repeated measure ANOVAs had group as a between-subject factor (HDP vs. controls, HDR vs. controls) and compatibility (compatible, incompatible, and neutral) as a within-subject factor. An additional factor was introduced, namely sentence-type (N, OH, and CH). Moreover, RTs in ACE were normalized by subtracting the mean RT of the neutral trials from the mean RTs of the compatible and incompatible trials. The N sentences are more predictable and frequent than OH and CH sentences, eliciting shorter RTs (Aravena et al., 2010; Ibanez et al., 2013). If either HDP or HDR evidences preserved sentence-type modulation (shorter RTs for N than OH and CH sentences), then the ACE in HD cannot be explained as a general motor impairment or as an artifact of response variability (Aravena et al., 2010; Ibanez et al., 2013; Cardona et al., 2014). Tukey’s HSD test was used in the calculation of post hoc contrasts.
We further explored individual differences in ACE; a global score of the ACE was defined by the subtraction of the mean RT for the incompatible and compatible conditions (Ibanez et al., 2013; Cardona et al., 2014). In the patient group, these global scores were tested for correlation with age and years of illness through Spearman’s rank correlations. We also performed Spearman’s rank correlations between the ACE and IFS total scores to evaluate involvement of executive functions in all groups.
Results
Demographic and Clinical Evaluation
There were no significant differences in age between HDP and controls [F(1, 34) = 0.030, p = 0.86], or HDR and controls [F(1, 35) = 0.005, p = 0.94]. Neither did the education level differ between HDP and controls [F(1, 34) = 0.15, p = 0.69] or HDR and controls [F(1, 35) = 0.008, p = 0.94]. No differences in gender were observed in comparing HDP and controls [χ2(1) = 0.00, p = 1.00] or HDR and controls [χ2(1) = 0.012, p = 0.90]. The control groups’ intellectual levels were similar to those of HDP [F(1, 34) = 0.004, p = 0.94] and HDR [F(1, 35) = 1.80, p = 0.18]. Descriptive data are provided in Table 1.
Psychopathological and Neuropsychological Assessment
Data regarding the neuropsychological, cognitive, and motor performance of the participants are displayed in Table 2.
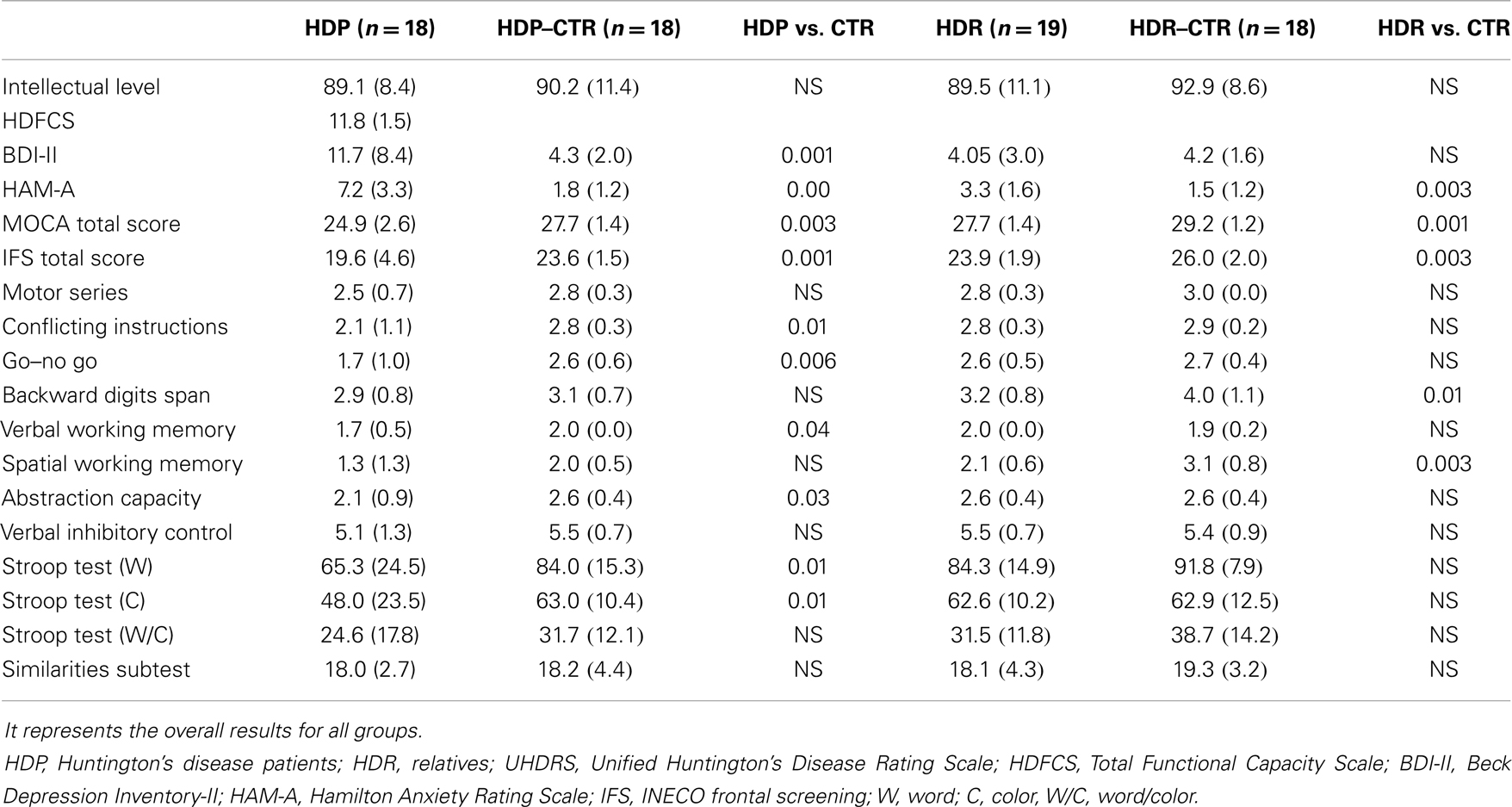
Table 2. Psychopathological and neuropsychological data.
HD patients
Relative to controls, HDP evidenced higher levels of anxiety [F(1, 34) = 40.19, p < 0.01], as measured by the HAM-A. In addition, HDP showed higher levels of depression symptoms [F(1, 34) = 12.73, p < 0.01].
Also, as compared to their controls, HDP had lower total scores on the MOCA [F(1, 34) = 15.94, p < 0.01] and the IFS [F(1, 34) = 12.05, p < 0.01]. A detailed comparison of performance on the eight IFS subtests indicated that both HDP and HDR exhibited deficits in verbal working memory. HDP also showed impairments in conflictive instructions, motor inhibitory control, and abstraction capacity. In addition, HDP were outperformed by controls in the word [F(1, 34) = 7.42, p < 0.05] and color naming [F(1, 34) = 6.09, p < 0.05] conditions of the Stroop test. However, no differences were observed in the word/color condition [F(1, 34) = 1.9, p = 0.16] or the similarities subtest [F(1, 34) = 0.01, p = 0.89]. See Table 2 for further details.
Relatives
Higher levels of anxiety were also observed in HDR [F(1, 35) = 15.92, p < 0.01] as compared to their controls. However, no differences between HDR and controls were observed in the BDI-II total score [F(1, 35) = 0.84, p < 0.77].
Huntington’s disease patients’ relatives had lower total scores than controls in both the MOCA [F(1, 35) = 11.36, p < 0.01] and the IFS [F(1, 35) = 9.67, p < 0.01]. Analysis of the eight IFS subtests revealed verbal working memory impairments in HDR. No differences between groups were observed in the word [F(1, 35) = 3.61, p = 0.06], color [F(1, 35) = 0.06, p = 0.93], or word/color [F(1, 35) = 2.80, p = 0.10] conditions of the Stroop test. Both groups also obtained comparable scores on the similarities subtest [F(1, 35) = 1.02, p = 0.31]. See Table 2 for further details.
Action–Language in HDP
ACE is impaired in HDP
We observed (Figure 1A) an interaction of Group × Compatibility [F(1, 34) = 8.38. p < 0.01]. Post hoc comparisons (Tukey’s HSD test, MS = 3560.4; DF = 68.00; ηp2 = 0. 2) revealed an ACE in controls: incompatible trials elicited longer RTs than both compatible (p < 0.002) and neutral trials (p < 0.0002). Conversely, ACE was abolished in HDP: RT differences were observed only between the neutral trials and the compatible (p < 0.0002) and incompatible trials (p < 0.0002), there being no differences between compatible and incompatible trials (p = 0.93). These results suggest that motor impairment affects action–language processing (Table 3).

Table 3. ACE-RTs.
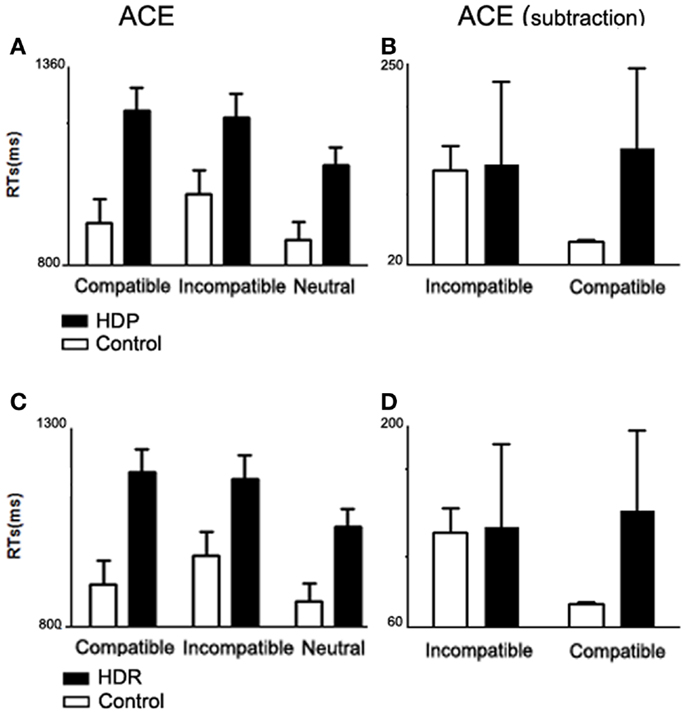
Figure 1. Action compatibility effect in HDP and HDR. (A) Mean RTs from compatible, incompatible, and neutral trials for HDP. HDP did not show an ACE (compatible facilitation and incompatible delay of RTs). (B) ACE subtraction, group comparison of ACE normalized by subtracting mean RT from the neutral trials from the mean RTs from the compatible and incompatible trials. (C) Mean RTs for HDR participants. HDR did not show ACE. (D) ACE subtraction. In all panels, the bars depict the SD.
Subtraction analysis. In order to assess performance while controlling for general differences between HDP and controls, neutral RTs were subtracted from compatible and incompatible categories. After subtraction (Figure 1B), group differences became larger [Group × Compatibility interaction, F(1, 34) = 11.699. p < 0.002]. In particular, the control group showed a larger difference between compatible (M = 47.32 ms, SD = 1.38) and incompatible trials (M = 128.53 ms, SD = 27, p < 0.0002), whereas in HDP the means for compatible (M = 153.35 ms, SD = 90.83) and incompatible (M = 134.55 ms, SD = 94.27) trials were quite similar (p = 0.6).
Preserved ACE motor responses to linguistic variables in HDP
Stimulus content analysis. Importantly, both HDP and controls responded faster to N sentences than to OH and CH sentences [F(2, 68) = 19.919; p < 0.00001]. Post hoc comparisons (Tukey’s HSD test, MS = 871.44, DF = 68.00) yielded significant differences between N and OH (p < 0.002) and between N and CH (p < 0.002). No difference was detected between hand-shape sentences (OH vs. CH; p < 0.97) (Table 4). This result confirms that motor impairment in HDP was not so severe as to preclude effects of linguistic variables. Consequently, the ACE deficits in HD cannot be explained by a general motor or language impairment.
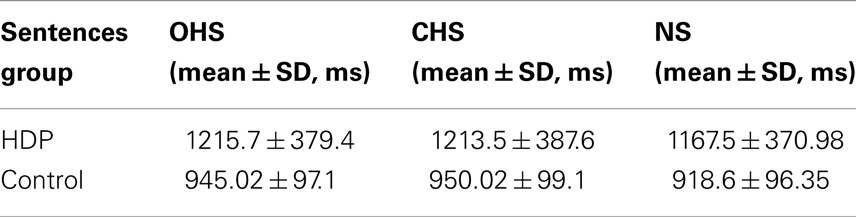
Table 4. Mean and SD of each sentence list in HDP.
Action–Language in HDR
ACE is impaired in HDR
We identified (Figure 1C) an interaction of Group × Compatibility between HDR and control groups [F(2, 70) = 6.32, p < 0.01]. Post hoc comparisons (MS = 1843.0; DF = 70.00) showed that HDR had shorter RTs on neutral trials as compared to compatible (p < 0.002) and incompatible trials (p < 0.002); however, they showed no differences between compatible and incompatible trials (p < 0.99) (Table 5). These results show that there is no ACE in HDR, suggesting an impairment in the use of the motor system to simulate language content.
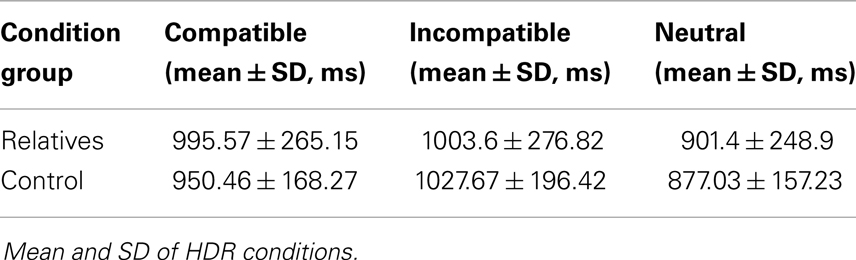
Table 5. ACE-RTs.
Subtraction analysis
In order to examine the performance of HDR and their controls while controlling for general group differences, neutral RTs were subtracted from compatible and incompatible categories. After subtraction (Figure 1D), group differences became larger [Group × Compatibility interaction, F(1, 35) = 23.669; p < 0.00002]. In particular, the control group showed a larger difference between compatible (M = 73.42 ms, SD = 70.83) and incompatible trials (M = 150.66 ms, SD = 90.18, p < 0.0002), whereas in HDR the means for compatible (M = 94.16 ms, SD = 51.09) and incompatible (M = 102.20 ms, SD = 53.05) trials were similar (p = 0.5).
Preserved ACE motor responses to linguistic variables in HDR
Both HDR and their controls responded faster to N sentences than to OH and CH sentences [F(2, 70) = 22.611, p < 0.00001]. Post hoc comparisons (Tukey’s HSD test, MS = 50,068, DF = 35.431) yielded significant differences between N and CH (p < 0.01). No differences were observed between hand-shape sentences (OH vs. CH, p < 0.50; N vs. OH, p < 0.23) (Table 6). As in HDP, this effect confirms that motor impairment in HDR was not so severe as to preclude effects of linguistic variables. Consequently, as was the case in HDP, the ACE deficits in HDR cannot be explained by a general motor or language impairment.

Table 6. Mean and SD of each sentence list in HDR.
Verbal (KDT) and Noun (PPT) Processing
Having found significant group differences in the KDT [F(3, 69) = 11.270; p < 0.00001], we performed ANOVAs for each group comparison: HDP vs. HDCTR [F(1, 34) = 14.842; p < 0.0005], HDR vs. HDRCTR [F(1, 35) = 11.400; p < 0.001], and HDP vs. HDR [F(1, 35) = 4.2285, p < 0.05]. The KDT score (percentage of correct responses) was significantly reduced in HDPs (M = 45.50; SD = 0.85) as compared to their controls (M = 50.16; SD = 0.85). The same was true of HDR (M = 48.31; SD = 0.53) relative to their own controls (M = 50.88, SD = 0.54). This result suggests specific action–language impairment in both the HDP and HDR groups (Figure 2).
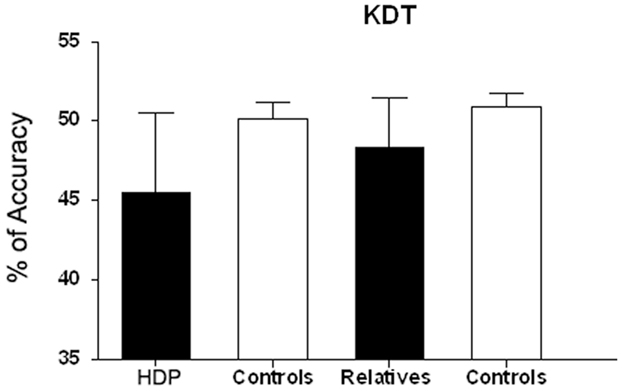
Figure 2. Verbal processing (KDT) in HDP and RHD. KDT scale denotes percentage of correct responses (percentage accuracy) in HDP vs. controls. **p < 0.0005; HDR vs. controls, p < 0.002; HDP vs. HDR,*p < 0.01.
An ANOVA on the PPT scores revealed differences between HDP and their controls [F(1, 34) = 6.526; p < 0.01], but not between HDR and their controls (p = 0.5).
Correlations
To evaluate whether executive function performance would explain the observed action–language profiles, we performed Spearman’s rank correlations between the ACE and IFS total scores. Considering all groups, significant correlation coefficients were obtained between the ACE and IFS total scores (N = 72, R = 0.3; p < 0.05). Nevertheless, no correlation was observed for HDP (N = 18, R = 0.2; p = 0.5) or HDR (N = 19, R = 0.3; p = 0.2). Finally, in HDP, we did not find significant correlation coefficients between ACE and age/years of illness (R = 0.12, p = 0.3; R = 0.05, p = 0.9, respectively). In brief, these correlations suggest that executive functions do not explain the HDP and HDR results. In addition, ACE impairments in HDP were most likely unrelated to age/years of illness, although more complex design is needed to further investigate this issue.
Discussion
This study sought to further current understanding of motor–language coupling in HD families and to establish whether basal ganglia networks are engaged in motor–language coupling. A strong association between action–language comprehension and motor processes has been reported in previous studies assessing normal participants or other motor conditions (Pulvermuller and Shtyrov, 2003; Hauk et al., 2004; Buccino et al., 2005; Tettamanti et al., 2005; Zwaan and Taylor, 2006). However, there is a dearth of studies exploring this issue in HD populations. This study provides novel results relevant for both clinical research on HD and the cognitive neuroscience of action–language.
Motor–Language Impairments as an Early Marker of HD
Motor–language coupling was evaluated by the ACE paradigm at an early-stage of HD. The present ACE results showed shorter RTs for healthy volunteers in the compatible condition, leading to faster, more accurate movement. Action was effectively facilitated by compatible sentences, as previously reported (Fischler and Bloom, 1980; Glenberg and Kaschak, 2002; Borghi et al., 2004; Kaschak et al., 2005; Tseng, 2005; Borreggine and Kaschak, 2006; Zwaan and Taylor, 2006; Havas et al., 2007; Glenberg et al., 2008b; Aravena et al., 2010; Bergen and Wheeler, 2010; de Vega et al., 2013). Conversely, we did not find an ACE in HD subjects (i.e., no facilitation was observed). These disturbances were not associated with the absence of language discrimination or the existence of motor–response variability. Then, the stimulus content analysis showed that, although HDP were slower (but not significantly so), they preserved the ability to discriminate linguistic aspects of the sentences.
For several years, the neural basis of word–meaning processing has been a topic of interest in cognitive neuroscience. Although there is substantial evidence for the involvement of sensory–motor systems in conceptual processing, it is still unclear whether these play a causal role in such a function (Fernandino et al., 2013). Our findings suggest that action–verb processing partially depends on basal ganglia activation, since the ACE was completely abolished in patients with atrophy of those structures (HDP). This result is consistent with previous studies showing reduced ACE in the early-stages of Parkinson’s disease (Amoruso et al., 2013; Cardona et al., 2013; Ibanez et al., 2013), and action–verb impairments in degenerative brain diseases compromising the motor system, such as motor neuron disease (Bak et al., 2001; Cotelli et al., 2006, 2007; Boulenger et al., 2008) and the frontal variant of frontotemporal dementia (Cotelli et al., 2006).
The earliest and most prominent neuropathological changes in HD are found in the neostriatum (Vonsattel et al., 1985). Loss of basal ganglia volume has been reported in pre-clinical cases (Aylward et al., 1994; Harris et al., 1999). Neuroimaging studies have consistently revealed cortico-basal ganglia compromise in pre-clinical HD (Antonini et al., 1996; Paulsen et al., 2004; Rosas et al., 2008) and a decrease in dopamine receptor binding (Antonini et al., 1996; Lawrence et al., 1998b).
Interestingly, we also found that HDP have an impairment in action–verb (KDT) and noun (PPT) processing. Nevertheless, as compared to controls, relatives presented only KDT deficits with preserved PPT performance. The discrepancy in the performance between KDT and PPT tasks may be explained by the pattern of atrophy in patients as compared to their relatives. Anatomically, tasks involving semantic association of nouns (PPT) result in focal activation of the anterior inferior temporal lobe, the parahippocampal gyrus, and the inferior occipital cortex (Vandenberghe et al., 1996; Ricci et al., 1999; Butler et al., 2009). In contrast, semantic processing of verbs (KDT) is linked to basal ganglia, left frontal cortex (Perani et al., 1999; Cappa et al., 2002), and Brodmann’s areas 44 and 45 (Bak et al., 2001). In view of these findings, the detection of early cognitive changes may be better served by a measure of cognitive impairment subsequent to basal ganglia lesions than by instruments tapping functions subserved by frontal cortical areas.
Altogether, the present results provide evidence for the involvement of a motor-related basal ganglia–cortical circuit in the processing of action–language. Given that our HDP were in an early-stage of the disease and that their relatives had no motor symptoms, the observed action–word processing impairment seems prior to motor symptoms.
Executive Functions and ACE
Classical cognitive theories propose that the role of subcortical structures in language processing is limited to executive functions (Sambin et al., 2012). Typically, language disturbances in HDP have been assumed to result from damage to frontostriatal and frontotemporal areas (Nadeau, 2008; Lepron et al., 2009). HDP perform poorly on some neuropsychological tests which are sensitive to frontostriatal dysfunction. In this sense, it should be noted that HDP showed mild cognitive deficits, especially in subtests of the IFS – namely, conflictive instructions, Go–No Go, verbal and spatial working memory, and abstraction capacity. Nevertheless, these deficits seem not to be directly related to ACE, since there were no correlations between ACE results and total IFS scores in either HDP or HDR. Thus, our results suggest that ACE impairments in HDP and their relatives were related to a sui generis motor–language coupling, independent of executive function involvement. Further research is required to clarify to what extent motor–language processing is unrelated to other deficits in HD families.
Motor–Language Impairments as a Marker of HD Vulnerability
Nowadays, genetic tests can establish the presence of the mutation causing HD. However, the diagnosis of HD is still based on clinical evidence, such as outward signs and family history. As HD is genetically transmitted, the children of an affected individual have 50% chance of inheriting the abnormal huntingtin gene and eventually developing the disease (Folstein et al., 1985).
Vulnerability to HD means that individuals with a family history of HD have a high probability of developing the disease or some unspecific deficits related to it (Panegyres and Goh, 2011). The CAG repeat length normally varies from 6 to 35 CAG units. Repeat lengths from 27 to 35 are considered “high normal” and may expand in subsequent generations (Rubinsztein et al., 1996; Quarrell et al., 2007). Repeat lengths from 36 to 39 exhibit reduced penetrance, with manifestations occurring at a later age or not at all (ACMG/ASHG statement) (McNeil et al., 1997; The American College of Medical Genetics/American Society of Human Genetics Huntington Disease Genetic Testing Working Group, 1998; Quarrell et al., 2007). Alleles with 40 repeats are fully penetrant and inevitably associated with progressive motor, cognitive, and behavioral features of HD (Hendricks et al., 2009). It has been observed that longer CAG repeat expansions are associated with earlier disease manifestation (Duyao et al., 1993; Stine et al., 1993; Langbehn et al., 2010), and that age of onset varies considerably for any given CAG repeat expansion (Dennhardt and LeDoux, 2010). Nevertheless, there is growing evidence that cognitive changes occur in individuals who carry an expanded allele prior to the clinical (motor) diagnosis of HD (Dorsey, 2012). Also, recent studies suggest that environmental factors can modify the onset and progression of HD (van Dellen et al., 2005). Moreover, clinical and neuropsychiatric manifestations have been reported in relatives of HDR, irrespective of whether they were HD gene carriers or not (Markianos et al., 2008).
In this study, we showed that HDP and their first-degree relatives all performed poorly on the ACE task. Interestingly, the ACE task results for HDR fall right between those of HDP and controls. The HDR group represents familial HD subjects at risk of developing HD, who did not receive genetic evaluation. Hence, this group would include both HD gene carriers and non-carriers. In this context, an intermediate pattern in cognitive differences between subjects with HD gene expression and subjects at risk without the HD gene could mean that part of the subjects will not develop HD in the future. Nevertheless, familial vulnerability has been reported even in the absence of HD genetic alleles (Markianos et al., 2008; Dorsey, 2012), and previous studies failed to find differences in cognition between prodromal carriers and mutation negative relatives in HD (Blackmore et al., 1995; Giordani et al., 1995; Campodonico et al., 1996, 1998; de Boo et al., 1999; Soliveri et al., 2002). The asymptomatic relatives assessed here represent a group with vulnerability to HD or some unspecific-related deficits (Panegyres and Goh, 2011). Although this group might include both HD gene carriers and non-carriers, its performance in action–language was significantly lower than controls, which suggests that even non-carriers may have selective impairments. Our data are consistent with previous studies on HD reporting other cognitive deficits without clinical motor signs (Henley et al., 2008; Tabrizi et al., 2009), and with findings of familial vulnerability factors even in the absence of HD mutation (Markianos et al., 2008; Dorsey, 2012). Although the probability of being a non-manifest carrier is 50%, all participants in this group were subclinical; however, even non-carriers can present vulnerability factors. Thus, two levels of vulnerability (one represented by gene-carrier relatives with subclinical manifestations, and another by non-carrier relatives with diffuse vulnerability factors) might be reasonably proposed. Taken together, these data indicate that action–language, in general, and the ACE paradigm, in particular, might tap familial vulnerability to HD.
Relevance for Theoretical Models of Subcortical Involvement in Motor–Language Coupling
Classically, language production and comprehension have been related to brain networks in the left inferior frontal and superior temporal cortices, such as Wernicke’s and Broca’s areas (Blank et al., 2002). However, a growing body of clinical and neuroimaging evidence shows that language processing activates a much more complex and widely distributed network (Mesulam, 1990; Price et al., 1996; Pulvermuller, 1999, 2002, 2005).
It is well-established that the motor system plays a fundamental role in action–verb comprehension/production (Pulvermuller, 2005; Pulvermuller and Fadiga, 2010). There is abundant evidence showing that the processing of action–verbs implicates the frontal and motor cortices (Federmeier et al., 2000; Pulvermuller et al., 2001; Yokoyama et al., 2006; Boulenger et al., 2008; Cappelletti et al., 2008; Kemmerer et al., 2008; Tomasino et al., 2008). However, numerous studies have failed to find a somatotopic distribution of action–verbs or sentences in the motor cortex (Aziz-Zadeh et al., 2006; Ruschemeyer et al., 2007; Postle et al., 2008; Raposo et al., 2009). Accordingly, theories of embodied cognition propose that language comprehension is based on perceptual and motor processes (Bak, 2013). For its own part, a weak view of the embodied cognition hypothesis proposes that other cortical regions are indeed required (Brass et al., 2007). Besides, evidence from Parkinson’s disease studies suggests an intricate connection between language and the motor system by a bidirectional influence of motor and language areas, including subcortical motor areas and even non-motor regions (Bak, 2013; Cardona et al., 2013).
The primary spot on HD neuropathology is the basal ganglia. Hence, this disorder provides an important model for the role of the human basal ganglia in motor–language coupling. Atrophy of caudate and putamen nuclei in HDP is a well-established fact (Vonsattel et al., 1985; Mann et al., 1993; Loh et al., 1994; Aylward et al., 1997). This degenerative process is present even in the early-stages of the disease (Antonini et al., 1996), and it has been reported in some studies in pre-symptomatic individuals who carry the HD mutation (Aylward et al., 1994). Some frontal neocortical atrophy may also occur later in the course of the disease (Aylward et al., 1998). Therefore, early HDP and HDR constitute ideal models to study the role of subcortical structures in motor–language coupling.
Selective action–verb impairments (using non-motor linguistic tasks, such as action naming) in other motor diseases have been reported in progressive supranuclear palsy, amyotrophic lateral sclerosis, and cortico-basal degeneration. Instead, the ACE paradigm may offer more sensitive, discriminatory measurements of action–language interaction already reported in Parkinson’s disease (Ibanez et al., 2013) and now in Huntington families. Recently, Cardona et al. (2014) showed that ACEs are abolished in Parkinson’s disease, but not in neuromyelitis optica and acute transverse myelitis (two models of preserved brain motor areas and musculoskeletal system injury). Additional comparative studies including other brain-affected (e.g., progressive supranuclear palsy, amyotrophic lateral sclerosis) and musculoskeletal-affected motor diseases (e.g., glutamine expansion diseases such as muscle–spinal atrophy) would expose which disorders present action–language impairments as reported with the ACE task.
Available evidence indicates that the basal ganglia participate in multiple parallel segregated circuits or “thalamo-cortical loops” involving connections with motor, sensory, and cognitive areas of the cerebral cortex (Alexander et al., 1986; Hoover and Strick, 1993; Middleton and Strick, 2002). The motor network is a complex circuit that includes primary motor and sensory cortices, pre-motor, parietal, precuneal, and dorsal lateral pre-frontal cortical regions, the basal ganglia, and the cerebellum (Cardona et al., 2013). The present findings support the notion that motor system involvement during language processing engages subcortical areas. Action–language seems to rely not only on the motor cortex, but also on neuronal circuits involving the basal ganglia network.
Limitations and Directions for Further Research
One limitation of this work is that the HDR group did not receive genetic screening. Thus, it may have included both genetic pre-symptomatic and healthy relatives without HD genetic heredity. Nevertheless, familial vulnerability has been reported even in the absence of HD genetic alleles (Markianos et al., 2008; Dorsey, 2012). Therefore, the HDRs in this study represent a vulnerability group including pre-symptomatic HDP as well as individuals with a diffuse vulnerability, not restricted to HD expanded alleles. Further studies are needed to explore differences in action–language impairments between relatives with and without HD non-expanded alleles.
Also, it would be useful to elaborate on this study by quantifying the ACE task according to the number of triplets’ expansion. Longitudinal studies that evaluate disease development based on ACE results could support the role of the ACE task as a predictor of HD onset.
Conclusion
The ACE task unmasked the initiation of action–language deficits subsequent to basal ganglia network damage. To our knowledge, this is the first study showing that motor–language coupling is impaired in HD relatives. These findings highlight the key role of a cortico-basal ganglia network in motor–language impairment – a distinct cognitive deficit in HD.
This overall result has important clinical implications. There is increasing evidence that cognitive impairments precede the phenotypic expression of HD. Studies describing the transition from health to disease phenotype are important to understand the nature of the disease and to outline possible therapies for different stages of the disease.
Here, we established that the ACE task could be useful to uncover asymptomatic cognitive dysfunction in HD, since the ACE is impaired in HDR preceding other cognitive and motor impairments. Our findings demonstrate that the ACE paradigm constitutes a sensitive method for the assessment of subcortical cognitive damage, which may be of critical importance for neurocognitive biomarker research, as well as for drug testing in clinical trials.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was partially supported by grants from CONICET, CONICYT/FONDECYT Regular (1130920), COLCIENCIAS (1115-545-31374, contract: 392), FONCyT-PICT 2012-0412, FONCyT-PICT 2012-1309, and the INECO Foundation.
References
Alexander, G. E., Delong, M. R., and Strick, P. L. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9, 357–381. doi: 10.1146/annurev.ne.09.030186.002041
Amoruso, L., Gelormini, C., Aboitiz, F., Alvarez Gonzalez, M., Manes, F., Cardona, J. F., et al. (2013). N400 ERPs for actions: building meaning in context. Front. Hum. Neurosci. 7:57. doi:10.3389/fnhum.2013.00057
Antonini, A., Leenders, K. L., Spiegel, R., Meier, D., Vontobel, P., Weigell-Weber, M., et al. (1996). Striatal glucose metabolism and dopamine D2 receptor binding in asymptomatic gene carriers and patients with Huntington’s disease. Brain 119(Pt 6), 2085–2095. doi:10.1093/brain/119.6.2085
Aravena, P., Hurtado, E., Riveros, R., Cardona, J. F., Manes, F., and Ibanez, A. (2010). Applauding with closed hands: neural signature of action-sentence compatibility effects. PLoS ONE 5:e11751. doi:10.1371/journal.pone.0011751
Aylward, E. H., Anderson, N. B., Bylsma, F. W., Wagster, M. V., Barta, P. E., Sherr, M., et al. (1998). Frontal lobe volume in patients with Huntington’s disease. Neurology 50, 252–258. doi:10.1212/WNL.50.1.252
Aylward, E. H., Brandt, J., Codori, A. M., Mangus, R. S., Barta, P. E., and Harris, G. J. (1994). Reduced basal ganglia volume associated with the gene for Huntington’s disease in asymptomatic at-risk persons. Neurology 44, 823–828. doi:10.1212/WNL.44.5.823
Aylward, E. H., Li, Q., Stine, O. C., Ranen, N., Sherr, M., Barta, P. E., et al. (1997). Longitudinal change in basal ganglia volume in patients with Huntington’s disease. Neurology 48, 394–399. doi:10.1212/WNL.48.2.394
Azambuja, M. J., Radanovic, M., Haddad, M. S., Adda, C. C., Barbosa, E. R., and Mansur, L. L. (2012). Language impairment in Huntington’s disease. Arq. Neuropsiquiatr. 70, 410–415. doi:10.1590/S0004-282X2012000600006
Aziz-Zadeh, L., Wilson, S. M., Rizzolatti, G., and Iacoboni, M. (2006). Congruent embodied representations for visually presented actions and linguistic phrases describing actions. Curr. Biol. 16, 1818–1823. doi:10.1016/j.cub.2006.07.060
Bachoud-Levi, A. C., Maison, P., Bartolomeo, P., Boisse, M. F., Dalla Barba, G., Ergis, A. M., et al. (2001). Retest effects and cognitive decline in longitudinal follow-up of patients with early HD. Neurology 56, 1052–1058. doi:10.1212/WNL.56.8.1052
Badets, A., Pesenti, M., and Olivier, E. (2010). Response-effect compatibility of finger-numeral configurations in arithmetical context. Q. J. Exp. Psychol. (Hove) 63, 16–22. doi:10.1080/17470210903134385
Bak, T., and Hodges, J. R. (2003). Kissing and dancing – a test to distinguish lexical and conceptual contributions to noun/verb and action/object dissociation. Preliminary results in patients with frontotemporal dementia. J. Neurolinguistics 16, 169–181. doi:10.1016/S0911-6044(02)00011-8
Bak, T. H. (2013). The neuroscience of action semantics in neurodegenerative brain diseases. Curr. Opin. Neurol. 26, 671–677. doi:10.1097/WCO.0000000000000039
Bak, T. H., Crawford, L. M., Hearn, V. C., Mathuranath, P. S., and Hodges, J. R. (2005). Subcortical dementia revisited: similarities and differences in cognitive function between progressive supranuclear palsy (PSP), corticobasal degeneration (CBD) and multiple system atrophy (MSA). Neurocase 11, 268–273. doi:10.1080/13554790590962997
Bak, T. H., and Hodges, J. R. (2004). The effects of motor neurone disease on language: further evidence. Brain Lang. 89, 354–361. doi:10.1016/S0093-934X(03)00357-2
Bak, T. H., O’Donovan, D. G., Xuereb, J. H., Boniface, S., and Hodges, J. R. (2001). Selective impairment of verb processing associated with pathological changes in Brodmann areas 44 and 45 in the motor neurone disease-dementia-aphasia syndrome. Brain 124, 103–120. doi:10.1093/brain/124.1.103
Beck, A., Brown, G., and Steer, R. (1996). Manual for the Beck Depression Inventory-II. San Antonio: The Psychological Corporation.
Bergen, B., and Wheeler, K. (2010). Grammatical aspect and mental simulation. Brain Lang. 112, 150–158. doi:10.1016/j.bandl.2009.07.002
Blackmore, L., Simpson, S. A., and Crawford, J. R. (1995). Cognitive performance in UK sample of presymptomatic people carrying the gene for Huntington’s disease. J. Med. Genet. 32, 358–362. doi:10.1136/jmg.32.5.358
Blank, S. C., Scott, S. K., Murphy, K., Warburton, E., and Wise, R. J. (2002). Speech production: Wernicke, Broca and beyond. Brain 125, 1829–1838. doi:10.1093/brain/awf191
Borghi, A. M., Glenberg, A. M., and Kaschak, M. P. (2004). Putting words in perspective. Mem. Cognit. 32, 863–873. doi:10.3758/BF03196865
Borreggine, K. L., and Kaschak, M. P. (2006). The action-sentence compatibility effect: it’s all in the timing. Cogn. Sci. 30, 1097–1112. doi:10.1207/s15516709cog0000_91
Boulenger, V., Mechtouff, L., Thobois, S., Broussolle, E., Jeannerod, M., and Nazir, T. A. (2008). Word processing in Parkinson’s disease is impaired for action verbs but not for concrete nouns. Neuropsychologia 46, 743–756. doi:10.1016/j.neuropsychologia.2007.10.007
Brass, M., Schmitt, R. M., Spengler, S., and Gergely, G. (2007). Investigating action understanding: inferential processes versus action simulation. Curr. Biol. 17, 2117–2121. doi:10.1016/j.cub.2007.11.057
Buccino, G., Riggio, L., Melli, G., Binkofski, F., Gallese, V., and Rizzolatti, G. (2005). Listening to action-related sentences modulates the activity of the motor system: a combined TMS and behavioral study. Brain Res. Cogn. Brain Res. 24, 355–363. doi:10.1016/j.cogbrainres.2005.02.020
Butler, C. R., Brambati, S. M., Miller, B. L., and Gorno-Tempini, M. L. (2009). The neural correlates of verbal and nonverbal semantic processing deficits in neurodegenerative disease. Cogn. Behav. Neurol. 22, 73–80. doi:10.1097/WNN.0b013e318197925d
Campodonico, J. R., Aylward, E., Codori, A. M., Young, C., Krafft, L., Magdalinski, M., et al. (1998). When does Huntington’s disease begin? J. Int. Neuropsychol. Soc. 4, 467–473. doi:10.1017/S1355617798455061
Campodonico, J. R., Codori, A. M., and Brandt, J. (1996). Neuropsychological stability over two years in asymptomatic carriers of the Huntington’s disease mutation. J. Neurol. Neurosurg. Psychiatr. 61, 621–624. doi:10.1136/jnnp.61.6.621
Cappa, S. F., Sandrini, M., Rossini, P. M., Sosta, K., and Miniussi, C. (2002). The role of the left frontal lobe in action naming: rTMS evidence. Neurology 59, 720–723. doi:10.1212/WNL.59.5.720
Cappelletti, M., Fregni, F., Shapiro, K., Pascual-Leone, A., and Caramazza, A. (2008). Processing nouns and verbs in the left frontal cortex: a transcranial magnetic stimulation study. J. Cogn. Neurosci. 20, 707–720. doi:10.1162/jocn.2008.20045
Cardona, J., Kargieman, L., Sinay, V., Gershanik, O., Gelormini, C., Amoruso, L., et al. (2014). How embodied is action language? Neurological evidence from motor diseases. Cognition 131, 311–322. doi:10.1016/j.cognition.2014.02.001
Cardona, J. F., Gershanik, O., Gelormini-Lezama, C., Houck, A. L., Cardona, S., Kargieman, L., et al. (2013). Action-verb processing in Parkinson’s disease: new pathways for motor-language coupling. Brain Struct. Funct. 218, 1355–1373. doi:10.1007/s00429-013-0510-1
Chenery, H. J., Copland, D. A., and Murdoch, B. E. (2002). Complex language functions and subcortical mechanisms: evidence from Huntington’s disease and patients with non-thalamic subcortical lesions. Int. J. Lang. Commun. Disord. 37, 459–474. doi:10.1080/1368282021000007730
Cotelli, M., Borroni, B., Manenti, R., Alberici, A., Calabria, M., Agosti, C., et al. (2006). Action and object naming in frontotemporal dementia, progressive supranuclear palsy, and corticobasal degeneration. Neuropsychology 20, 558–565. doi:10.1037/0894-4105.20.5.558
Cotelli, M., Borroni, B., Manenti, R., Zanetti, M., Arevalo, A., Cappa, S. F., et al. (2007). Action and object naming in Parkinson’s disease without dementia. Eur. J. Neurol. 14, 632–637. doi:10.1111/j.1468-1331.2007.01797.x
de Boo, G. M., Tibben, A. A., Hermans, J. A., Jennekens-Schinkel, A., Maat-Kievit, A., and Roos, R. A. (1999). Memory and learning are not impaired in presymptomatic individuals with an increased risk of Huntington’s disease. J. Clin. Exp. Neuropsychol. 21, 831–836. doi:10.1076/jcen.21.6.831.854
de Vega, M., Moreno, V., and Castillo, D. (2013). The comprehension of action-related sentences may cause interference rather than facilitation on matching actions. Psychol. Res. 77, 20–30. doi:10.1007/s00426-011-0356-1
Dennhardt, J., and LeDoux, M. S. (2010). Huntington disease in a nonagenarian mistakenly diagnosed as normal pressure hydrocephalus. J. Clin. Neurosci. 17, 1066–1067. doi:10.1016/j.jocn.2009.11.011
D’Honincthun, P., and Pillon, A. (2008). Verb comprehension and naming in frontotemporal degeneration: the role of the static depiction of actions. Cortex 44, 834–847. doi:10.1016/j.cortex.2007.04.003
Dorsey, E. (2012). Characterization of a large group of individuals with Huntington disease and their relatives enrolled in the COHORT study. PLoS ONE 7:e29522. doi:10.1371/journal.pone.0029522
Duyao, M., Ambrose, C., Myers, R., Novelletto, A., Persichetti, F., Frontali, M., et al. (1993). Trinucleotide repeat length instability and age of onset in Huntington’s disease. Nat. Genet. 4, 387–392. doi:10.1038/ng0893-387
Federmeier, K. D., Segal, J. B., Lombrozo, T., and Kutas, M. (2000). Brain responses to nouns, verbs and class-ambiguous words in context. Brain 123(Pt 12), 2552–2566. doi:10.1093/brain/123.12.2552
Fernandino, L., Conant, L. L., Binder, J. R., Blindauer, K., Hiner, B., Spangler, K., et al. (2013). Parkinson’s disease disrupts both automatic and controlled processing of action verbs. Brain Lang. 127, 65–74. doi:10.1016/j.bandl.2012.07.008
Fischler, I., and Bloom, P. A. (1980). Rapid processing of the meaning of sentences. Mem. Cognit. 8, 216–225. doi:10.3758/BF03197609
Folstein, S. E., Leigh, R. J., Parhad, I. M., and Folstein, M. F. (1986). The diagnosis of Huntington’s disease. Neurology 36, 1279–1283. doi:10.1212/WNL.36.10.1279
Folstein, S. E., Phillips, J. A. III, Meyers, D. A., Chase, G. A., Abbott, M. H., Franz, M. L., et al. (1985). Huntington’s disease: two families with differing clinical features show linkage to the G8 probe. Science 229, 776–779. doi:10.1126/science.2992086
Giordani, B., Berent, S., Boivin, M. J., Penney, J. B., Lehtinen, S., Markel, D. S., et al. (1995). Longitudinal neuropsychological and genetic linkage analysis of persons at risk for Huntington’s disease. Arch. Neurol. 52, 59–64. doi:10.1001/archneur.1995.00540250063014
Gleichgerrcht, E., Roca, M., Manes, F., and Torralva, T. (2011). Comparing the clinical usefulness of the institute of cognitive neurology (INECO) frontal screening (IFS) and the frontal assessment battery (FAB) in frontotemporal dementia. J. Clin. Exp. Neuropsychol. 33, 997–1004. doi:10.1080/13803395.2011.589375
Glenberg, A. M. (2006). Naturalizing cognition: the integration of cognitive science and biology. Curr. Biol. 16, R802–R804. doi:10.1016/j.cub.2006.08.044
Glenberg, A. M., and Kaschak, M. P. (2002). Grounding language in action. Psychon. Bull. Rev. 9, 558–565. doi:10.3758/BF03196313
Glenberg, A. M., Sato, M., and Cattaneo, L. (2008a). Use-induced motor plasticity affects the processing of abstract and concrete language. Curr. Biol. 18, R290–R291. doi:10.1016/j.cub.2008.02.036
Glenberg, A. M., Sato, M., Cattaneo, L., Riggio, L., Palumbo, D., and Buccino, G. (2008b). Processing abstract language modulates motor system activity. Q. J. Exp. Psychol. (Hove) 61, 905–919. doi:10.1080/17470210701625550
Hamilton, M. (1959). The assessment of anxiety states by rating. Br. J. Med. Psychol. 32, 50–55. doi:10.1111/j.2044-8341.1959.tb00467.x
Harris, G. J., Codori, A. M., Lewis, R. F., Schmidt, E., Bedi, A., and Brandt, J. (1999). Reduced basal ganglia blood flow and volume in pre-symptomatic, gene-tested persons at-risk for Huntington’s disease. Brain 122(Pt 9), 1667–1678. doi:10.1093/brain/122.9.1667
Hauk, O., Johnsrude, I., and Pulvermuller, F. (2004). Somatotopic representation of action words in human motor and premotor cortex. Neuron 41, 301–307. doi:10.1016/S0896-6273(03)00838-9
Havas, D. A., Glenberg, A. M., and Rinck, M. (2007). Emotion simulation during language comprehension. Psychon. Bull. Rev. 14, 436–441. doi:10.3758/BF03194085
Hendricks, A. E., Latourelle, J. C., Lunetta, K. L., Cupples, L. A., Wheeler, V., MacDonald, M. E., et al. (2009). Estimating the probability of de novo HD cases from transmissions of expanded penetrant CAG alleles in the Huntington disease gene from male carriers of high normal alleles (27-35 CAG). Am. J. Med. Genet. A 149A, 1375–1381. doi:10.1002/ajmg.a.32901
Henley, S. M., Wild, E. J., Hobbs, N. Z., Frost, C., Macmanus, D. G., Barker, R. A., et al. (2009). Whole-brain atrophy as a measure of progression in premanifest and early Huntington’s disease. Mov. Disord. 24, 932–936. doi:10.1002/mds.22485
Henley, S. M., Wild, E. J., Hobbs, N. Z., Warren, J. D., Frost, C., Scahill, R. I., et al. (2008). Defective emotion recognition in early HD is neuropsychologically and anatomically generic. Neuropsychologia 46, 2152–2160. doi:10.1016/j.neuropsychologia.2008.02.025
Ho, A. K., Sahakian, B. J., Robbins, T. W., Barker, R. A., Rosser, A. E., and Hodges, J. R. (2002). Verbal fluency in Huntington’s disease: a longitudinal analysis of phonemic and semantic clustering and switching. Neuropsychologia 40, 1277–1284. doi:10.1016/S0028-3932(01)00217-2
Ho, L. W., Carmichael, J., Swartz, J., Wyttenbach, A., Rankin, J., and Rubinsztein, D. C. (2001). The molecular biology of Huntington’s disease. Psychol. Med. 31, 3–14. doi:10.1017/S0033291799002871
Hoover, J. E., and Strick, P. L. (1993). Multiple output channels in the basal ganglia. Science 259, 819–821. doi:10.1126/science.7679223
Howard, D., Patterson, K., Wise, R., Brown, W. D., Friston, K., Weiller, C., et al. (1992). The cortical localization of the lexicons. Positron emission tomography evidence. Brain 115(Pt 6), 1769–1782. doi:10.1093/brain/115.6.1769
Huntington Study Group. (1996). Unified Huntington’s disease rating scale: reliability and consistency. Huntington Study Group. Mov. Disord. 11, 136–142. doi:10.1002/mds.870110204
Ibanez, A., Cardona, J. F., Dos Santos, Y. V., Blenkmann, A., Aravena, P., Roca, M., et al. (2013). Motor-language coupling: direct evidence from early Parkinson’s disease and intracranial cortical recordings. Cortex 49, 968–984. doi:10.1016/j.cortex.2012.02.014
Ibanez, A., and Manes, F. (2012). Contextual social cognition and the behavioral variant of frontotemporal dementia. Neurology 78, 1354–1362. doi:10.1212/WNL.0b013e3182518375
Kaschak, M. P., Madden, C. J., Therriault, D. J., Yaxley, R. H., Aveyard, M., Blanchard, A. A., et al. (2005). Perception of motion affects language processing. Cognition 94, B79–B89. doi:10.1016/j.cognition.2004.06.005
Kemmerer, D., Castillo, J. G., Talavage, T., Patterson, S., and Wiley, C. (2008). Neuroanatomical distribution of five semantic components of verbs: evidence from fMRI. Brain Lang. 107, 16–43. doi:10.1016/j.bandl.2007.09.003
Kipps, C. M., Duggins, A. J., Mahant, N., Gomes, L., Ashburner, J., and McCusker, E. A. (2005). Progression of structural neuropathology in preclinical Huntington’s disease: a tensor based morphometry study. J. Neurol. Neurosurg. Psychiatr. 76, 650–655. doi:10.1136/jnnp.2004.047993
Kloppel, S., Draganski, B., Golding, C. V., Chu, C., Nagy, Z., Cook, P. A., et al. (2008). White matter connections reflect changes in voluntary-guided saccades in pre-symptomatic Huntington’s disease. Brain 131, 196–204. doi:10.1093/brain/awm275
Langbehn, D. R., Hayden, M. R., and Paulsen, J. S. (2010). CAG-repeat length and the age of onset in Huntington disease (HD): a review and validation study of statistical approaches. Am. J. Med. Genet. B Neuropsychiatr. Genet. 153B, 397–408. doi:10.1002/ajmg.b.30992
Lawrence, A. D., Hodges, J. R., Rosser, A. E., Kershaw, A., Ffrench-Constant, C., Rubinsztein, D. C., et al. (1998a). Evidence for specific cognitive deficits in preclinical Huntington’s disease. Brain 121(Pt 7), 1329–1341. doi:10.1093/brain/121.7.1329
Lawrence, A. D., Weeks, R. A., Brooks, D. J., Andrews, T. C., Watkins, L. H., Harding, A. E., et al. (1998b). The relationship between striatal dopamine receptor binding and cognitive performance in Huntington’s disease. Brain 121(Pt 7), 1343–1355. doi:10.1093/brain/121.7.1343
Lepron, E., Peran, P., Cardebat, D., and Demonet, J. F. (2009). A PET study of word generation in Huntington’s disease: effects of lexical competition and verb/noun category. Brain Lang. 110, 49–60. doi:10.1016/j.bandl.2009.05.004
Lindeman, M., Cederstrom, S., Simola, P., Simula, A., Ollikainen, S., and Riekki, T. (2008). Sentences with core knowledge violations increase the size of N400 among paranormal believers. Cortex 44, 1307–1315. doi:10.1016/j.cortex.2007.07.010
Loh, E. A., Roberts, J. K., and Mohr, E. (1994). Structural correlates of neurological signs in Huntington’s disease. Behav. Neurol. 7, 127–134. doi:10.3233/BEN-1994-73-404
Mann, D. M., Oliver, R., and Snowden, J. S. (1993). The topographic distribution of brain atrophy in Huntington’s disease and progressive supranuclear palsy. Acta Neuropathol. 85, 553–559. doi:10.1007/BF00230496
Markianos, M., Panas, M., Kalfakis, N., and Vassilopoulos, D. (2008). Low plasma total cholesterol in patients with Huntington’s disease and first-degree relatives. Mol. Genet. Metab. 93, 341–346. doi:10.1016/j.ymgme.2007.10.002
Masumoto, K., Takai, T., Tsuneto, S., and Kashiwagi, T. (2004). Influence of motoric encoding on forgetting function of memory for action sentences in patients with Alzheimer’s disease. Percept. Mot. Skills 98, 299–306. doi:10.2466/pms.98.1.299-306
McNeil, S. M., Novelletto, A., Srinidhi, J., Barnes, G., Kornbluth, I., Altherr, M. R., et al. (1997). Reduced penetrance of the Huntington’s disease mutation. Hum. Mol. Genet. 6, 775–779. doi:10.1093/hmg/6.5.775
Mesulam, M. M. (1990). Large-scale neurocognitive networks and distributed processing for attention, language, and memory. Ann. Neurol. 28, 597–613. doi:10.1002/ana.410280502
Middleton, F. A., and Strick, P. L. (2002). Basal-ganglia ‘projections’ to the prefrontal cortex of the primate. Cereb. Cortex 12, 926–935. doi:10.1093/cercor/12.9.926
Montoya, A., Price, B. H., Menear, M., and Lepage, M. (2006). Brain imaging and cognitive dysfunctions in Huntington’s disease. J. Psychiatry Neurosci. 31, 21–29.
Murray, L. L., and Lenz, L. P. (2001). Productive syntax abilities in Huntington’s and Parkinson’s diseases. Brain Cogn. 46, 213–219. doi:10.1016/S0278-2626(01)80069-5
Nadeau, S. E. (2008). The thalamus and working memory. J. Int. Neuropsychol. Soc. 14, 900–901. doi:10.1017/S1355617708081149
Nasreddine, Z. S., Phillips, N. A., Bedirian, V., Charbonneau, S., Whitehead, V., Collin, I., et al. (2005). The Montreal cognitive assessment, MoCA: a brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 53, 695–699. doi:10.1111/j.1532-5415.2005.53221.x
Neary, D., Snowden, J. S., and Mann, D. M. (2000). Cognitive change in motor neurone disease/amyotrophic lateral sclerosis (MND/ALS). J. Neurol. Sci. 180, 15–20. doi:10.1016/S0022-510X(00)00425-1
Panegyres, P. K., and Goh, J. G. (2011). The neurology and natural history of patients with indeterminate CAG repeat length mutations of the Huntington disease gene. J. Neurol. Sci. 301, 14–20. doi:10.1016/j.jns.2010.11.015
Paulsen, J. S., Zimbelman, J. L., Hinton, S. C., Langbehn, D. R., Leveroni, C. L., Benjamin, M. L., et al. (2004). fMRI biomarker of early neuronal dysfunction in presymptomatic Huntington’s disease. AJNR Am. J. Neuroradiol. 25, 1715–1721.
Peran, P., Cardebat, D., Cherubini, A., Piras, F., Luccichenti, G., Peppe, A., et al. (2009). Object naming and action-verb generation in Parkinson’s disease: a fMRI study. Cortex 45, 960–971. doi:10.1016/j.cortex.2009.02.019
Peran, P., Demonet, J. F., Pernet, C., and Cardebat, D. (2004). Verb and noun generation tasks in Huntington’s disease. Mov. Disord. 19, 565–571. doi:10.1002/mds.10706
Peran, P., Rascol, O., Demonet, J. F., Celsis, P., Nespoulous, J. L., Dubois, B., et al. (2003). Deficit of verb generation in nondemented patients with Parkinson’s disease. Mov. Disord. 18, 150–156. doi:10.1002/mds.10306
Perani, D., Schnur, T., Tettamanti, M., Gorno-Tempini, M., Cappa, S. F., and Fazio, F. (1999). Word and picture matching: a PET study of semantic category effects. Neuropsychologia 37, 293–306. doi:10.1016/S0028-3932(98)00073-6
Podoll, K., Caspary, P., Lange, H. W., and Noth, J. (1988). Language functions in Huntington’s disease. Brain 111(Pt 6), 1475–1503. doi:10.1093/brain/111.6.1475
Postle, N., McMahon, K. L., Ashton, R., Meredith, M., and De Zubicaray, G. I. (2008). Action word meaning representations in cytoarchitectonically defined primary and premotor cortices. Neuroimage 43, 634–644. doi:10.1016/j.neuroimage.2008.08.006
Pradilla, A. G., Vesga, A. B., and Leon-Sarmiento, F. E. (2003). National neuroepidemiological study in Colombia (EPINEURO). Rev. Panam. Salud Publica 14, 104–111. doi:10.1590/S1020-49892003000700005
Price, C. J., Wise, R. J., and Frackowiak, R. S. (1996). Demonstrating the implicit processing of visually presented words and pseudowords. Cereb. Cortex 6, 62–70. doi:10.1093/cercor/6.1.62
Pulvermuller, F. (1999). Words in the brain’s language. Behav. Brain Sci. 22, 253–279. doi:10.1017/S0140525X9900182X
Pulvermuller, F. (2002). A brain perspective on language mechanisms: from discrete neuronal ensembles to serial order. Prog. Neurobiol. 67, 85–111. doi:10.1016/S0301-0082(02)00014-X
Pulvermuller, F. (2005). Brain mechanisms linking language and action. Nat. Rev. Neurosci. 6, 576–582. doi:10.1038/nrn1706
Pulvermuller, F., and Fadiga, L. (2010). Active perception: sensorimotor circuits as a cortical basis for language. Nat. Rev. Neurosci. 11, 351–360. doi:10.1038/nrn2811
Pulvermuller, F., Harle, M., and Hummel, F. (2001). Walking or talking? Behavioral and neurophysiological correlates of action verb processing. Brain Lang. 78, 143–168. doi:10.1006/brln.2000.2390
Pulvermuller, F., and Shtyrov, Y. (2003). Automatic processing of grammar in the human brain as revealed by the mismatch negativity. Neuroimage 20, 159–172. doi:10.1016/S1053-8119(03)00261-1
Quarrell, O. W., Rigby, A. S., Barron, L., Crow, Y., Dalton, A., Dennis, N., et al. (2007). Reduced penetrance alleles for Huntington’s disease: a multi-centre direct observational study. J. Med. Genet. 44, e68. doi:10.1136/jmg.2006.045120
Raposo, A., Moss, H. E., Stamatakis, E. A., and Tyler, L. K. (2009). Modulation of motor and premotor cortices by actions, action words and action sentences. Neuropsychologia 47, 388–396. doi:10.1016/j.neuropsychologia.2008.09.017
Rhee, J., Antiquena, P., and Grossman, M. (2001). Verb comprehension in frontotemporal degeneration: the role of grammatical, semantic and executive components. Neurocase 7, 173–184. doi:10.1093/neucas/7.2.184
Ricci, P. T., Zelkowicz, B. J., Nebes, R. D., Meltzer, C. C., Mintun, M. A., and Becker, J. T. (1999). Functional neuroanatomy of semantic memory: recognition of semantic associations. Neuroimage 9, 88–96. doi:10.1006/nimg.1998.0386
Rosas, H. D., Salat, D. H., Lee, S. Y., Zaleta, A. K., Pappu, V., Fischl, B., et al. (2008). Cerebral cortex and the clinical expression of Huntington’s disease: complexity and heterogeneity. Brain 131, 1057–1068. doi:10.1093/brain/awn025
Rubinsztein, D. C., Leggo, J., Coles, R., Almqvist, E., Biancalana, V., Cassiman, J. J., et al. (1996). Phenotypic characterization of individuals with 30-40 CAG repeats in the Huntington disease (HD) gene reveals HD cases with 36 repeats and apparently normal elderly individuals with 36-39 repeats. Am. J. Hum. Genet. 59, 16–22.
Ruschemeyer, S. A., Brass, M., and Friederici, A. D. (2007). Comprehending prehending: neural correlates of processing verbs with motor stems. J. Cogn. Neurosci. 19, 855–865. doi:10.1162/jocn.2007.19.5.855
Sambin, S., Teichmann, M., De Diego Balaguer, R., Giavazzi, M., Sportiche, D., Schlenker, P., et al. (2012). The role of the striatum in sentence processing: disentangling syntax from working memory in Huntington’s disease. Neuropsychologia 50, 2625–2635. doi:10.1016/j.neuropsychologia.2012.07.014
Shoulson, I., and Fahn, S. (1979). Huntington disease: clinical care and evaluation. Neurology 29, 1–3. doi:10.1212/WNL.29.1.1
Soliveri, P., Monza, D., Piacentini, S., Paridi, D., Nespolo, C., Gellera, C., et al. (2002). Cognitive and psychiatric characterization of patients with Huntington’s disease and their at-risk relatives. Neurol. Sci. 23(Suppl. 2), S105–S106. doi:10.1007/s100720200091
Stine, O. C., Pleasant, N., Franz, M. L., Abbott, M. H., Folstein, S. E., and Ross, C. A. (1993). Correlation between the onset age of Huntington’s disease and length of the trinucleotide repeat in IT-15. Hum. Mol. Genet. 2, 1547–1549. doi:10.1093/hmg/2.10.1547
Stout, J. C., Paulsen, J. S., Queller, S., Solomon, A. C., Whitlock, K. B., Campbell, J. C., et al. (2011). Neurocognitive signs in prodromal Huntington disease. Neuropsychology 25, 1–14. doi:10.1037/a0020937
Tabrizi, S. J., Langbehn, D. R., Leavitt, B. R., Roos, R. A., Durr, A., Craufurd, D., et al. (2009). Biological and clinical manifestations of Huntington’s disease in the longitudinal TRACK-HD study: cross-sectional analysis of baseline data. Lancet Neurol. 8, 791–801. doi:10.1016/S1474-4422(09)70170-X
Teichmann, M., Dupoux, E., Cesaro, P., and Bachoud-Levi, A. C. (2008). The role of the striatum in sentence processing: evidence from a priming study in early stages of Huntington’s disease. Neuropsychologia 46, 174–185. doi:10.1016/j.neuropsychologia.2007.07.022
Teichmann, M., Dupoux, E., Kouider, S., Brugieres, P., Boisse, M. F., Baudic, S., et al. (2005). The role of the striatum in rule application: the model of Huntington’s disease at early stage. Brain 128, 1155–1167. doi:10.1093/brain/awh472
Tettamanti, M., Buccino, G., Saccuman, M. C., Gallese, V., Danna, M., Scifo, P., et al. (2005). Listening to action-related sentences activates fronto-parietal motor circuits. J. Cogn. Neurosci. 17, 273–281. doi:10.1162/0898929053124965
The American College of Medical Genetics/American Society of Human Genetics Huntington Disease Genetic Testing Working Group. (1998). ACMG/ASHG statement. Laboratory guidelines for Huntington disease genetic testing. The American College of Medical Genetics/American Society of Human Genetics Huntington Disease Genetic Testing Working Group. Am. J. Hum. Genet. 62, 1243–1247.
Tomasino, B., Fink, G. R., Sparing, R., Dafotakis, M., and Weiss, P. H. (2008). Action verbs and the primary motor cortex: a comparative TMS study of silent reading, frequency judgments, and motor imagery. Neuropsychologia 46, 1915–1926. doi:10.1016/j.neuropsychologia.2008.01.015
Torralva, T., Roca, M., Gleichgerrcht, E., Lopez, P., and Manes, F. (2009). INECO frontal screening (IFS): a brief, sensitive, and specific tool to assess executive functions in dementia. J. Int. Neuropsychol. Soc. 15, 777–786. doi:10.1017/S1355617709990415
Treisman, A., and Fearnley, S. (1969). The Stroop test: selective attention to colours and words. Nature 222, 437–439. doi:10.1038/222437a0
Tröster, A. I. (ed.) (2006). Assessment of Movement and Demyelinating Disorders. Washington, DC: American Psychological Association.
Tseng, M. B. (2005). “Lexical processing drives motor simulation,” in Proceedings of the Twenty-Seventh Annual Conference of the Cognitive Science Society, (Mahwah, NJ: Erlbaum). p. 2206–2211.
Ullman, M. T., Corkin, S., Coppola, M., Hickok, G., Growdon, J. H., Koroshetz, W. J., et al. (1997). A neural dissociation within language: evidence that the mental dictionary is part of declarative memory, and that grammatical rules are processed by the procedural system. J. Cogn. Neurosci. 9, 266–276. doi:10.1162/jocn.1997.9.2.266
van Dellen, A., Grote, H. E., and Hannan, A. J. (2005). Gene-environment interactions, neuronal dysfunction and pathological plasticity in Huntington’s disease. Clin. Exp. Pharmacol. Physiol. 32, 1007–1019. doi:10.1111/j.1440-1681.2005.04313.x
van Elk, M., Van Schie, H. T., and Bekkering, H. (2008). Semantics in action: an electrophysiological study on the use of semantic knowledge for action. J. Physiol. Paris 102, 95–100. doi:10.1016/j.jphysparis.2008.03.011
Vandenberghe, R., Price, C., Wise, R., Josephs, O., and Frackowiak, R. S. (1996). Functional anatomy of a common semantic system for words and pictures. Nature 383, 254–256. doi:10.1038/383254a0
Vonsattel, J. P., Myers, R. H., Stevens, T. J., Ferrante, R. J., Bird, E. D., and Richardson, E. P. Jr. (1985). Neuropathological classification of Huntington’s disease. J. Neuropathol. Exp. Neurol. 44, 559–577. doi:10.1097/00005072-198511000-00003
Wechsler, D. (1997). Wechsler Adult Intelligent Scale III. San Antonio, TX: The Psychological Corporation.
Wechsler, D. (1999). Wechsler Abbreviated Scale of Intelligence. San Antonio, TX: The Psychological Corporation.
Yokoyama, S., Miyamoto, T., Riera, J., Kim, J., Akitsuki, Y., Iwata, K., et al. (2006). Cortical mechanisms involved in the processing of verbs: an fMRI study. J. Cogn. Neurosci. 18, 1304–1313. doi:10.1162/jocn.2006.18.8.1304
Keywords: Huntington’s disease, action–language, ACE, KDT, motor–language coupling, familial vulnerability
Citation: Kargieman L, Herrera E, Baez S, García AM, Dottori M, Gelormini C, Manes F, Gershanik O and Ibáñez A (2014) Motor–language coupling in Huntington’s disease families. Front. Aging Neurosci. 6:122. doi: 10.3389/fnagi.2014.00122
Received: 04 March 2014; Accepted: 27 May 2014;
Published online: 12 June 2014.
Edited by:
Rodrigo Orlando Kuljiš, Zdrav Mozak Limitada, ChileCopyright: © 2014 Kargieman, Herrera, Baez, García, Dottori, Gelormini, Manes, Gershanik and Ibáñez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Agustín Ibáñez, Laboratory of Experimental Psychology and Neuroscience (LPEN), Institute of Cognitive Neurology (INECO) and CONICET, Pacheco de Melo 1860, Buenos Aires, C1126AAB, Argentina e-mail: aibanez@ineco.org.ar
†First authors