Endogenous cortisol levels are associated with an imbalanced striatal sensitivity to monetary versus non-monetary cues in pathological gamblers
 Yansong Li
Yansong Li Guillaume Sescousse
Guillaume Sescousse Jean-Claude Dreher
Jean-Claude Dreher- 1Reward and Decision Making Team, Centre de Neurosciences Cognitives, CNRS, UMR 5229, Lyon, France
- 2Neuroscience Department, Université Claude Bernard Lyon 1, Lyon, France
Pathological gambling is a behavioral addiction characterized by a chronic failure to resist the urge to gamble. It shares many similarities with drug addiction. Glucocorticoid hormones including cortisol are thought to play a key role in the vulnerability to addictive behaviors, by acting on the mesolimbic reward pathway. Based on our previous report of an imbalanced sensitivity to monetary versus non-monetary incentives in the ventral striatum of pathological gamblers (PGs), we investigated whether this imbalance was mediated by individual differences in endogenous cortisol levels. We used functional magnetic resonance imaging (fMRI) and examined the relationship between cortisol levels and the neural responses to monetary versus non-monetary cues, while PGs and healthy controls were engaged in an incentive delay task manipulating both monetary and erotic rewards. We found a positive correlation between cortisol levels and ventral striatal responses to monetary versus erotic cues in PGs, but not in healthy controls. This indicates that the ventral striatum is a key region where cortisol modulates incentive motivation for gambling versus non-gambling related stimuli in PGs. Our results extend the proposed role of glucocorticoid hormones in drug addiction to behavioral addiction, and help understand the impact of cortisol on reward incentive processing in PGs.
Introduction
Glucocorticoid hormones (cortisol in humans and corticosterone in rodents) are produced by the adrenal cortex after the hypothalamic-pituitary-adrenal (HPA) axis is stimulated by psychologically or physiologically arousing stimuli (Sapolsky et al., 2000; Herman et al., 2005; Ulrich-Lai and Herman, 2009). These hormones have essential roles in normal physiological processes, such as acting on anti-stress and anti-inflammatory pathways, and, by doing so, have wide-ranging effects on behavior. Over the past few years, the potential role of glucocorticoid hormones on mental disorders has gained increased attention (Meewisse et al., 2007; Wingenfeld and Wolf, 2011). In particular, in the search for risk factors for drug addiction, increasing evidence points to an interaction between HPA functioning and drug exposure (Stephens and Wand, 2012). For example, a positive correlation between glucocorticoid levels and self-administration of psychostimulants has been observed in rodents (Goeders and Guerin, 1996; Deroche et al., 1997). In addition, drug administration produces stress-like cortisol responses (Broadbear et al., 2004) and similarly, acute administration of cortisol promotes cocaine craving in cocaine-dependent individuals (Elman et al., 2003). These findings not only point to the link between glucocorticoid hormones and addiction (Lovallo, 2006), but also emphasize the need to develop integrative theories explaining the mechanisms by which they affect addictive behavior.
Animal and human neuroimaging studies have demonstrated that addiction involves altered functioning of the mesolimbic reward system (Koob and Le Moal, 2008; Koob and Volkow, 2010; Schultz, 2011). Another line of research has shown that altered HPA response is associated with changes in dopaminergic regulation (Oswald and Wand, 2004; Alexander et al., 2011) and that glucocorticoid hormones have modulatory effects on dopamine release in the mesolimbic pathway, especially in the nucleus accumbens (NAcc; Oswald et al., 2005; Wand et al., 2007). Building on this evidence, it has been proposed that glucocorticoid hormones have facilitatory effects on behavioral responses to drugs of abuse, and that these effects are implemented via action on the mesolimbic reward system (Marinelli and Piazza, 2002; de Jong and de Kloet, 2004). Furthermore, on the basis of the incentive sensitization theory stating that the mesolimbic reward system mediates addiction-related cue hypersensitivity (Robinson and Berridge, 1993; Vezina, 2004, 2007; Robinson and Berridge, 2008), it has been proposed that glucocorticoid hormones contribute to drug addiction by modulating this neural system directly (Goodman, 2008; Vinson and Brennan, 2013).
Pathological gambling is a behavioral addiction characterized by compulsive gambling behavior and loss of control, which has gained much attention recently (van Holst et al., 2010; Conversano et al., 2012; Achab et al., 2013; Clark and Limbrick-Oldfield, 2013; Petry et al., 2013; Potenza, 2013). Since pathological gambling behavior shares many similarities with drug addiction in terms of clinical phenomenology (e.g., craving, tolerance, compulsive use, or withdrawal symptoms), heritability, and neurobiological profile (Potenza, 2006, 2008; Petry, 2007; Wareham and Potenza, 2010; Leeman and Potenza, 2012), it may be similarly under the influence of glucocorticoid hormones. However, little is known about the interaction between glucocorticoid hormones and incentive reward processing in pathological gambling. In the present study, we examined how endogenous cortisol modulates the processing of monetary and non-monetary cues in PGs. To achieve this goal, we re-analyzed previously published data using an incentive delay task manipulating both monetary and erotic rewards in PGs and healthy controls (Sescousse et al., 2013), and performed further correlation analyses between basal cortisol levels and neural responses. Based on the role of glucocorticoid hormones in drug addiction, we expected endogenous cortisol levels to be associated with neural responses to addiction-related cues versus non-addiction related cues. Specifically, since our previously published analysis found a differential response to monetary versus erotic cues in the ventral striatum of gamblers (Sescousse et al., 2013), we expected that higher cortisol levels would be associated with an increased differential response in anticipation of monetary versus erotic rewards in PGs.
Materials and Methods
Subjects
We evaluated 20 healthy control subjects and 20 PGs. All were right-handed heterosexual males. We chose to study only men because men generally respond more to visual sexual stimuli than women (Hamann et al., 2004; Rupp and Wallen, 2008) and because there is a higher prevalence of pathological gambling among men than among women (Blanco et al., 2006; Kessler et al., 2008). The dataset from these subjects has already been used in our published functional magnetic resonance imaging (fMRI) study aiming at comparing primary and secondary rewards in healthy controls and pathological gamblers (PGs; Sescousse et al., 2013). Our current analysis focuses specifically on the relationship with cortisol levels and is therefore entirely original. As described in Sescousse et al. (2013), our published analysis excluded data from two PGs, due to technical problems with the task presentation in one case, and due to a highly inconsistent behavior in terms of hedonic ratings throughout the task in the other case. In the current analysis, we further discarded the data from one pathological gambler, because of a failure in successfully collecting blood samples. Therefore, the results reported are based on 20 healthy control subjects and 17 PGs. All subjects gave written informed consent to participate in the experiment. The study was approved by the local ethics committee (Centre Léon Bérard, Lyon, France).
Subjects underwent a semi-structured interview (Nurnberger et al., 1994) performed by a psychiatrist. All PGs met the DSM-IV-TR [Diagnostic and Statistical Manual of Mental Disorders (fourth edition, text revision)] criteria for pathological gambling diagnosis. Patients had a minimum score of 5 on the South Oaks Gambling Screen questionnaire (SOGS; range: 5–14) (Lesieur and Blume, 1987). Importantly, all were active gamblers, and none were under therapy or treatment of any type. Healthy control subjects had a score of 0 on the SOGS questionnaire, except one subject who had a score of 1. In both groups, a history of major depressive disorder or substance abuse/dependence (except nicotine dependence) in the past year was considered an exclusion criterion. All other DSM-IV-TR axis I disorders were excluded based on lifetime diagnosis.
We used a number of questionnaires to assess our subjects. The Fagerstrom Test for Nicotine Dependence (FTND; Heatherton et al., 1991) measured their nicotine dependence severity; the Alcohol Use Disorders Identification Test (AUDIT; Saunders et al., 1993) was employed to estimate their alcohol consumption; the Hospital Anxiety and Depression scale (HAD; Zigmond and Snaith, 1983) was used to evaluate current depressive and anxiety symptoms; and finally the Sexual Arousability Inventory (SAI; Hoon and Chambless, 1998) was used to assess their sexual arousal. Both groups were matched on age, nicotine dependence, education, alcohol consumption, and depressive symptoms (Table 1). PGs scored slightly higher on the anxiety subscale of the HAD questionnaire. Importantly, the two groups did not differ on income level and sexual arousability (Table 1), thereby ensuring a comparable motivation across groups for monetary and erotic rewards.
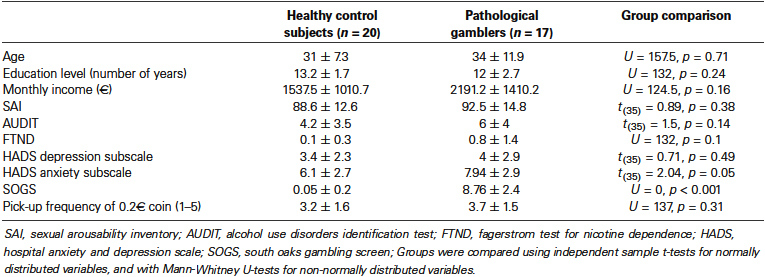
Table 1. Demographic and clinical characteristics of PGs and healthy controls.
To assess the subjects’ motivation for money, we asked them about the frequency with which they would pick up a 0.20€ coin from the street on a scale from 1 to 5 (Tobler et al., 2007) and matched the two groups based on this criterion (Table 1). To ensure that all subjects would be in a similar state of motivation to see erotic stimuli, we asked them to avoid any sexual contact during a period of 24 h before the scanning session. Finally, we also sought to enhance the motivation for money by telling subjects that the financial compensation for their participation would add up the winnings accumulated in one of the three runs. For ethical reasons, however, and unbeknownst to the subjects, they all received a fixed amount of cash at the end of the experiment.
All subjects were medication-free and instructed not to use any substance of abuse other than cigarettes on the day of the scan.
Experimental Task
We used an incentive delay task with both erotic and monetary rewards (Figure 1A). The total number of trials was 171. Each of them consisted of two phases: reward anticipation and reward outcome. During anticipation, subjects saw one of 12 cues announcing the type (monetary/erotic), probability (25/50/75%) and intensity (low/high) of an upcoming reward. An additional control cue was associated with a null reward probability. After a variable delay period (question mark representing a pseudo-random draw), subjects were asked to perform a visual discrimination task. If they answered correctly within less than 1 s, they were then allowed to view the outcome of the pseudo-random draw. In rewarded trials, the outcome was either an erotic image (with high or low erotic content) or the picture of a safe mentioning the amount of money won (high [10/11/12€] or low [1/2/3€]). Following each reward outcome, subjects were asked to provide a hedonic rating on a 1–9 continuous scale (1 = very little pleased; 9 = very highly pleased). In non-rewarded and control trials, subjects were presented with “scrambled” pictures. A fixation cross was finally used as an inter-trial interval of variable length.
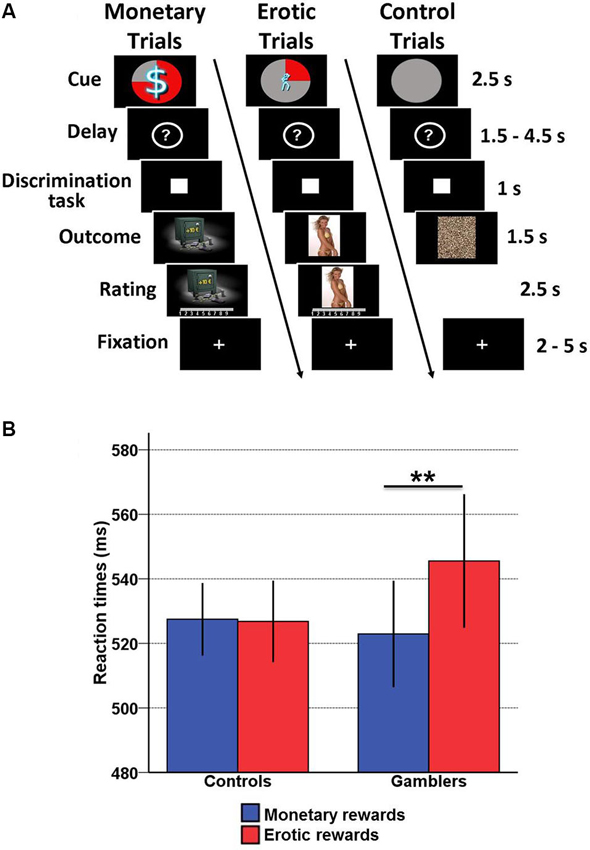
Figure 1. Incentive delay task and behavioral results. (A) Subjects first saw a cue informing them about the type (pictogram), intensity (size of pictogram) and probability (pie chart) of an upcoming reward. Three cases are represented here: a 75% chance of receiving a large amount of money (left), a 25% chance of seeing a low erotic content picture (middle) and a sure chance of getting nothing (control trials, right). Then the cue was replaced by a question mark, symbolizing a delay period during which a pseudorandom draw was performed according to the announced probability. Following this anticipation phase, participants had to perform a target discrimination task within <1 s. The target was either a triangle (left button press required) or a square (right button press required). Both their performance and the result of the pseudorandom draw determined the nature of the outcome. In rewarded trials, subjects saw a monetary amount displayed on a safe (high or low amount, left) or an erotic picture (with high or low erotic content, middle), and had to provide a hedonic rating on a continuous scale. In non-rewarded and control trials, subjects saw a scrambled picture (right). (B) Plot of mean reaction times according to reward type (monetary/erotic) and group (healthy controls/gamblers) in the discrimination task. There is a significant interaction between group and reward type, driven by slower reaction times for erotic compared to monetary cues in gamblers. Error bars indicate SEM. Asterisks denote significance of Tukey’s HSD tests (** p < 0.01).
Stimuli
Two categories (high and low intensity) of erotic pictures and monetary gains were used. Nudity being the main criteria driving the reward value of erotic stimuli, we separated them into a “low intensity” group displaying females in underwear or bathing suits and a “high intensity” group displaying naked females in an inviting posture. Each erotic picture was presented only once during the course of the task to avoid habituation. A similar element of surprise was introduced for monetary rewards by randomly varying the amounts at stake (low amounts: 1, 2, or 3€; high amounts: 10, 11, or 12€). The pictures displayed in non-rewarded and control trials were scrambled versions of the pictures used in rewarded trials and hence contained the same information in terms of chromaticity and luminance.
Plasma Cortisol Measurements
In order to minimize the effect of circadian hormone rhythms, we conducted all fMRI sessions between 8.50 and 11.45 AM. Just prior to the scanning session, blood samples were collected (mean time, 9.24 AM ± 0.27 mn) to measure the levels of plasma cortisol for each subject. Cortisol concentrations were measured by radioimmunoassay using an antiserum raised in rabbit immunized with cortisol 3-O (carboxy-methyl oxime) bovine serum albumin conjugate, 125I cortisol as tracer and buffer containing 8-anilino-1-naphtalene sulfonic acid (ANS) for cortisol-corticosteroid-binding globulin dissociation. Below is the description of the procedure. 100 μL of 125I cortisol (10000 dpm) was mixed with the standard or the sample (10 μL), buffer (500 μL) and 100 μL of antiserum solution. Samples were incubated for 45 min at 37°C and 1 h at 4°C. Bound and free cortisol was separated by addiction of a mixture PEG—anti-rabbit gamma globulin. After centrifugation, the radioactivity of the supernatant, containing the cortisol bound to antibody, was counted in a gamma-counter. The within and inter-assay coefficients of variation were less than 3.5 and 5.0% respectively at 300 nmol/L cortisol level. This method has been validated by gas chromatography/mass spectrometry measurements (Chazot et al., 1984).
Functional Magnetic Resonance Imaging (fMRI) Data Acquisition
Imaging was conducted on a 1.5 T Siemens Sonata scanner, using an eight-channel head coil. The scanning session was divided into three runs. Each of them included four repetitions of each cue, with the exception of the control condition, repeated nine times. This yielded a total of 171 trials. Within each run, the order of the different conditions was pseudorandomized and optimized to improve signal deconvolution. The order of the runs was counterbalanced between subjects. Before scanning, all subjects were given oral instructions and familiarized with the cognitive task in a short training session. Each of the three functional runs consisted of 296 volumes. Twenty-six interleaved slices parallel to the anterior commissure–posterior commissure line were acquired per volume (field of view, 220 mm; matrix, 64 × 64; voxel size, 3.4 × 3.4 × 4 mm; gap, 0.4 mm), using a gradient-echoechoplanar imaging (EPI) T2*-weighted sequence (repetition time, 2500 ms; echo time, 60 ms; flip angle, 90°). To improve the local field homogeneity and hence minimize susceptibility artifacts in the orbitofrontal area, a manual shimming was performed within a rectangular region including the orbitofrontal cortex (OFC) and the basal ganglia. A high-resolution T1-weighted structural scan was subsequently acquired in each subject.
Functional Magnetic Resonance Imaging (fMRI) Data Analysis
The analysis of the data was conducted using Statistical Parametric Mapping (SPM2). The pre-processing procedure included the deletion of the first four functional volumes of each run, slice-timing correction for the remaining volumes and spatial realignment to the first image of each time series. Subsequently, we used tsdiffana utility1 to search for residual artifacts in the time series and modeled them with dummy regressors in our general linear model. Then, the functional images were normalized to the Montreal Neurological Institute (MNI) stereotaxic space using the EPI template of SPM2 and spatially smoothed with a 10 mm full-width at half-maximum isotropic Gaussian kernel. Anatomical scans were normalized to the MNI space using the icbm152 template brain and averaged across the subjects. The averaged anatomical image was used as a template to display the functional activations.
Following the preprocessing step, the functional data from each subject was subjected to an event-related statistical analysis. Responses to monetary and erotic cues were modeled separately with 2.5 s box-car functions time-locked to the onset of the cue. For each cue, two orthogonal parametric regressors were added to account for the trial-to-trial variations in reward probability and intensity. The control condition was modeled in a separate regressor. Outcome-related responses were modeled as events time-locked to the appearance of the reward. The two rewards (monetary/erotic) and two possible outcomes (rewarded/non-rewarded) were modeled as four separate conditions. Two covariates linearly modeling the probability and the ratings were further added to each rewarded condition, while another covariate modeling the probability was added to each of the non-rewarded conditions. A last regressor modeled the appearance of a scrambled picture in the control condition. All regressors were subsequently convolved with the canonical hemodynamic response function and entered in a first level analysis. A high-pass filter with a cut-off of 128 s was applied to the time series. Contrast images were calculated based on the parameter estimates output by the general linear model, and were then passed in a second level group analysis.
Second-level analyses focused on the anticipation phase. First, we examined the contrast “monetary > erotic cue” in gamblers minus control subjects. This contrast was thresholded using a cluster-wise family-wise error (FWE) corrected p < 0.05. Then, based on our hypothesis, we investigated the relationship between basal cortisol levels and the differential brain response to monetary versus erotic cues. This correlation was computed separately for each group, and was then compared between groups. Based on our a priori hypotheses regarding the role of the ventral striatum in attributing incentive salience to reward cues, we used a small volume correction (SVC) based on 7 mm radius spheres centered around the peak voxels reported in a recent meta-analysis on reward processing (x, y, z = 12, 10, −6; x, y, z = −10, 8, −4) (Liu et al., 2011). We used a cluster-wise FWE corrected threshold of p ≤ 0.05. To further describe the patterns of activation, we used the EasyROI toolbox to extract the parameter estimates from significant clusters in the ventral striatum.
Results
Hormonal data
No significant differences between PGs and healthy control subjects were observed in basal cortisol levels (PGs: mean = 511.59, SD = 137.46; Healthy controls: mean = 588.7, SD = 121.61; t(35) = −1.81, p > 0.05). This is consistent with findings from recent studies reporting no difference in basal cortisol levels between recreational and PGs (Franco et al., 2010; Paris et al., 2010a,b). In addition, we performed a correlation analysis between cortisol levels and gambling symptom severity in PGs as indexed by the SOGS scale. Our result did not reveal a significant correlation between these variables (r = −0.35, p = 0.17).
Behavior
In our previous study (Sescousse et al., 2013), the main behavioral finding was a group × reward type interaction in the reaction time data, reflecting a weaker motivation for erotic compared with monetary rewards in gamblers. Given that one subject was discarded from our current analysis due to a failure to collect hormonal data, we performed this analysis again without this subject. The previous group × reward type interaction remained significant without this subject (F(1, 35) = 7.85, p < 0.01). In addition, Tukey’s post-hoc t-tests confirmed that the interaction was due to slower reaction times for erotic (mean = 547.54, SD = 17.22) compared with monetary rewards (mean = 522.91, SD = 14.29) in gamblers relative to healthy controls (p < 0.01) (Figure 1B). However, there was no significant correlation between basal cortisol levels and the performance on the discrimination task in either group.
Brain-Cortisol Correlation
Our previously published analysis revealed a group × reward type interaction in the ventral striatum, reflecting a larger differential response to monetary versus erotic cues in PGs compared with controls (Sescousse et al., 2013). In our current analysis, the results of the group × reward type interaction were still significant after removing the discarded subject (x, y, z = −9, 0, 3, T = 4.11; 18, 0, 0, T = 3.88; p(SVC) < 0.05, FWE). The present analysis focused on how this differential response relates to endogenous cortisol levels. Between-subject correlation analyses revealed a positive relationship between cortisol levels and BOLD responses to monetary versus erotic cues in the ventral striatum of gamblers (x, y, z = 3, 6, −6, T = 4.76, p(SVC) < 0.05, FWE; Figure 2A), but no such relationship in healthy controls. The direct comparison between groups was also significant (x, y, z = −3, 6, −6, T = 3.10, p(SVC) ≤ 0.05, FWE; Figure 2B). We additionally examined whether cortisol levels were correlated with brain activity elicited by each reward cue separately, as compared to the control cue. This analysis did not reveal any significant correlation in the ventral striatum in either group (at p < 0.001 uncorrected).
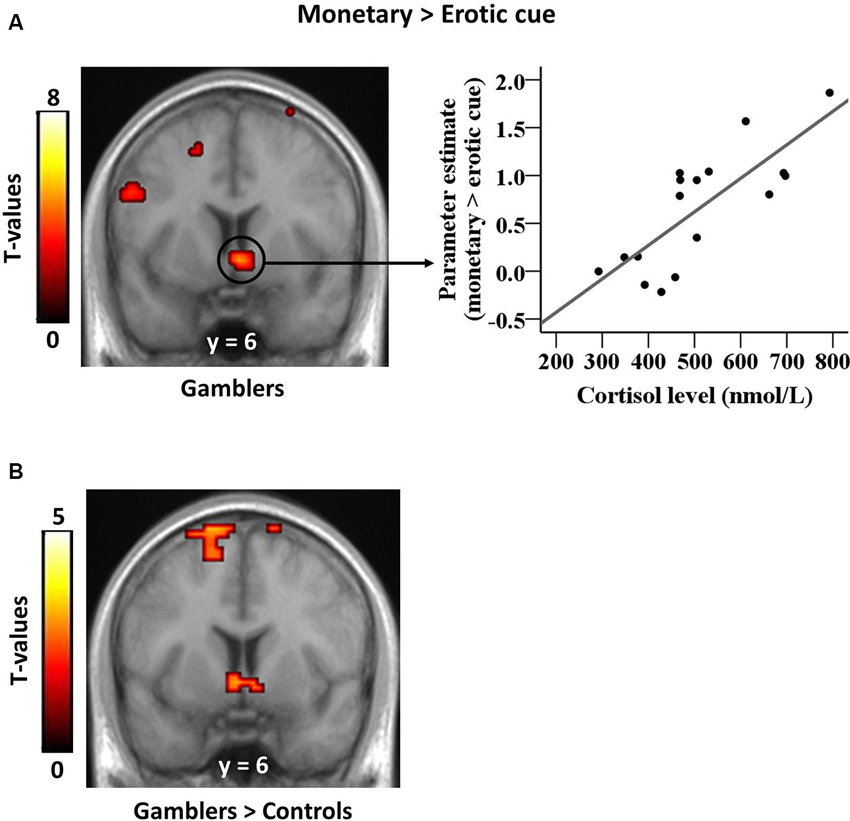
Figure 2. Correlation between striatal cue reactivity and basal cortisol levels in gamblers. (A) Ventral striatal responses to monetary versus erotic cues in gamblers are positively correlated with basal cortisol levels. The scatter plot illustrates this positive correlation within significant striatal voxels extracted from the whole-brain map. (B) This correlation between ventral striatal responses to monetary versus erotic cues and basal cortisol levels is stronger in gamblers than in healthy controls.
Discussion
To the best of our knowledge, this is the first study exploring the relationship between cortisol levels and brain activation during an incentive delay task in PGs. In line with our a priori hypothesis, we observed that higher endogenous cortisol levels were associated with an increased differential neural response to monetary versus erotic cues in the ventral striatum of gamblers as compared to healthy controls. This indicates a specific role of cortisol in biasing gamblers’ motivation towards monetary relative to non-monetary cues. Thus, cortisol may contribute to the addictive process in PGs by enhancing the saliency of gambling-related cues over other stimuli. Because enhanced incentive salience of gambling-related cues in PGs triggers gambling urges, this supports a link between cortisol and PGs’ motivation to pursue monetary rewards.
One potential mechanism through which cortisol might act to influence cue-elicited brain activity is glucocorticoid receptors in the NAcc. It has been shown that glucocorticoid hormones act on the brain through binding with two main intracellular receptors: the mineralocorticoid receptor (MR) and the glucocorticoid receptor. Glucocorticoid hormones play a fundamental role in reward-related behavior via their influence on mesolimbic dopamine circuitry and the NAcc in particular. For example, animal evidence shows that glucocorticoid hormones facilitate dopamine transmission in the NAcc shell through glucocorticoid receptors (Marinelli and Piazza, 2002). Microdialysis studies reported that corticosterone has stimulant effects on dopamine transmission in the NAcc (Piazza et al., 1996). Furthermore, infusion of glucocorticoid receptor antagonists has inhibitory effect on drug-induced dopamine release in the NAcc (Marinelli et al., 1998). In line with these findings in animals, human studies found evidence that cortisol levels were positively associated with amphetamine-induced dopamine release in the ventral striatum (Oswald et al., 2005).
It is important to note that we did not observe differences in basal cortisol levels between PGs and controls. Although this finding is in agreement with previous reports showing no difference in basal cortisol levels between PG and recreational gamblers (Meyer et al., 2004; Paris et al., 2010a,b), it does not imply that there is no HPA dysfunction in PGs. Indeed, while most previous studies investigating cortisol levels in PGs have focused on HPA responses to stress-inducing cues, such as gambling cues (Ramirez et al., 1988; Meyer et al., 2000; Franco et al., 2010), in the current study we measured baseline cortisol and its relationship with striatal activations. Moreover, other factors, such as the time of the day when blood or saliva are collected for cortisol level assessment, need to be considered because there are known endogenous diurnal variation in cortisol levels, which may vary between PGs and healthy controls or recreational gamblers. In particular, PGs may have a greater cortisol rise following waking than do recreational gamblers (Wohl et al., 2008).
Another important aspect to consider is that although cortisol is frequently used as a biomarker of psychological stress, a linear relationship between cortisol and other measures of HPA related endocrine signals does not necessarily exist (Hellhammer et al., 2009). Moreover, the absence of relationship between reward-related activity and basal cortisol levels in healthy controls is consistent with the variable effects of both acute stress and cortisol levels observed in the neuroimaging literature on reward processing in healthy individuals. For example, a recent study reported that stress reduces NAcc activation in response to reward cues, but that cortisol suppresses this relationship, as high cortisol was related to stronger NAcc activation in response to reward (Oei et al., 2014). Another study reported that acute stress decreased the response of the dorsal (not ventral) striatum and OFC to monetary outcomes (Porcelli et al., 2012), while no difference was observed in the NAcc between a stress group and control group using an emotion-induction procedure (Ossewaarde et al., 2011). Together, the evidence from fMRI studies indicates non-trivial relationships between stress, cortisol levels and brain activation and suggest that stress and cortisol may play distinct mediating roles in modulating sensitivity to potentially rewarding stimuli through the ventral striatum.
Several limitations of the present study need to be considered. First, only male PG were involved in the current study. It remains unclear whether our current findings would extend to female gamblers. This is an important question because sex differences exist in several aspects of gambling activity (Tschibelu and Elman, 2010; Grant et al., 2012; González-Ortega et al., 2013; van den Bos et al., 2013). Moreover, the modulatory effect of a number of hormonal factors on cognitive functioning varies between sexes (Kivlighan et al., 2005; Reilly, 2012; Vest and Pike, 2013). The current study only included men because they are generally more responsive to visual sexual stimuli than women (Stevens and Hamann, 2012; Wehrum et al., 2013) and show an elevated risk for gambling problems or severity of gambling compared to women (Toneatto and Nguyen, 2007; Wong et al., 2013). Second, we cannot make causal inferences regarding the effects of cortisol on neural responses because our results are based on correlational analyses. A pharmacological design with external cortisol administration compared to a placebo condition would be needed to assess the causal role of cortisol on gambling addiction. Despite these limitations, we believe that our current findings provide a foundation for further research on the interaction between cortisol and brain responses to incentive cues.
Conclusions
We have found that, in PGs, endogenous cortisol levels are associated with a differential activation of the ventral striatum in response to gambling-related incentives relative to non-gambling-related incentives. Our results point to the importance of integrating endocrinology with a cognitive neuroscience approach to elucidate the neural mechanisms underlying maladaptive gambling behavior. Finally, this study may have important implications for further research investigating the role of cortisol on vulnerability to develop behavioral addictions such as pathological gambling.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was performed within the framework of the LABEX ANR-11-LABEX-0042 of Université de Lyon, within the program “Investissements d’Avenir” (ANR-11-IDEX-0007) operated by the French National Research Agency (ANR). Yansong Li was supported by a PhD fellowship from Pari Mutuel Urbain (PMU). Guillaume Sescousse was funded by a scholarship from the French Ministry of Research and the Medical Research Foundation. We thank P. Domenech and G. Barbalat for clinical assessment of PGs. We thank Dr. I. Obeso for helpful revision on the manuscript and the staff of CERMEP–Imagerie du Vivant for helpful assistance with data collection.
Footnotes
References
Achab, S., Karila, L., and Khazaal, Y. (2013). Pathological gambling: update of decision making and neuro-functional correlates in clinical samples. Curr. Pharm. Des. [Epub ahead of print].
Alexander, N., Osinsky, R., Mueller, E., Schmitz, A., Guenthert, S., Kuepper, Y., et al. (2011). Genetic variants within the dopaminergic system interact to modulate endocrine stress reactivity and recovery. Behav. Brain Res. 216, 53–58. doi: 10.1016/j.bbr.2010.07.003
Blanco, C., Hasin, D. S., Petry, N., Stinson, F. S., and Grant, B. F. (2006). Sex differences in subclinical and DSM-IV pathological gambling: results from the National Epidemiologic Survey on Alcohol and related conditions. Psychol. Med. 36, 943–953. doi: 10.1017/s0033291706007410
Broadbear, J. H., Winger, G., and Woods, J. H. (2004). Self-administration of fentanyl, cocaine and ketamine: effects on the pituitary–adrenal axis in rhesus monkeys. Psychopharmacology (Berl) 176, 398–406. doi: 10.1007/s00213-004-1891-x
Chazot, G., Claustrat, B., Brun, J., Jordan, D., Sassolas, G., and Schott, B. (1984). A chronobiological study of melatonin, cortisol growth hormone and prolactin secretion in cluster headache. Cephalalgia 4, 213–220. doi: 10.1046/j.1468-2982.1984.0404213.x
Clark, L., and Limbrick-Oldfield, E. H. (2013). Disordered gambling: a behavioral addiction. Curr. Opin. Neurobiol. 23, 655–659. doi: 10.1016/j.conb.2013.01.004
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G., and Dell’osso, L. (2012). Pathological gambling: a systematic review of biochemical, neuroimaging, and neuropsychological findings. Harv. Rev. Psychiatry 20, 130–148. doi: 10.3109/10673229.2012.694318
Deroche, V., Marinelli, M., Le Moal, M., and Piazza, P. V. (1997). Glucocorticoids and behavioral effects of psychostimulants. II: cocaine intravenous self-administration and reinstatement depend on glucocorticoid levels. J. Pharmacol. Exp. Ther. 281, 1401–1407.
Elman, I., Lukas, S. E., Karlsgodt, K. H., Gasic, G. P., and Breiter, H. C. (2003). Acute cortisol administration triggers craving in individuals with cocaine dependence. Psychopharmacol. Bull. 37, 84–89.
Franco, C., Paris, J., Wulfert, E., and Frye, C. (2010). Male gamblers have significantly greater salivary cortisol before and after betting on a horse race, than do female gamblers. Physiol. Behav. 99, 225–229. doi: 10.1016/j.physbeh.2009.08.002
Goeders, N. E., and Guerin, G. F. (1996). Role of corticosterone in intravenous cocaine self-administration in rats. Neuroendocrinology 64, 337–348. doi: 10.1159/000127137
González-Ortega, I., Echeburúa, E., Corral, P., Polo-López, R., and Alberich, S. (2013). Predictors of pathological gambling severity taking gender differences into account. Eur. Addict. Res. 19, 146–154. doi: 10.1159/000342311
Goodman, A. (2008). Neurobiology of addiction. An integrative review. Biochem. Pharmacol. 75, 266–322. doi: 10.1016/j.bcp.2007.07.030
Grant, J. E., Chamberlain, S. R., Schreiber, L., and Odlaug, B. L. (2012). Gender-related clinical and neurocognitive differences in individuals seeking treatment for pathological gambling. J. Psychiatr. Res. 46, 1206–1211. doi: 10.1016/j.jpsychires.2012.05.013
Hamann, S., Herman, R. A., Nolan, C. L., and Wallen, K. (2004). Men and women differ in amygdala response to visual sexual stimuli. Nat. Neurosci. 7, 411–416. doi: 10.1038/nn1208
Heatherton, T. F., Kozlowski, L. T., Frecker, R. C., and Fagerstrom, K. O. (1991). The Fagerström test for nicotine dependence: a revision of the Fagerstrom Tolerance Questionnaire. Br. J. Addict. 86, 1119–1127. doi: 10.1111/j.1360-0443.1991.tb01879.x
Hellhammer, D. H., Wüst, S., and Kudielka, B. M. (2009). Salivary cortisol as a biomarker in stress research. Psychoneuroendocrinology 34, 163–171. doi: 10.1016/j.psyneuen.2008.10.026
Herman, J. P., Ostrander, M. M., Mueller, N. K., and Figueiredo, H. (2005). Limbic system mechanisms of stress regulation: hypothalamo-pituitary-adrenocortical axis. Prog. Neuropsychopharmacol. Biol. Psychiatry 29, 1201–1213. doi: 10.1016/j.pnpbp.2005.08.006
Hoon, E. F., and Chambless, D. (1998). “Sexual arousability inventory and sexual arousability inventory-expanded,” in Handbook of Sexuality-Related Measures, eds C. Davis, W. Yarber, R. Bauserman, R. Schreer and S. Davis (Thousand Oaks, CA: Sage), 71–74.
de Jong, I. E., and de Kloet, E. R. (2004). Glucocorticoids and vulnerability to psychostimulant drugs: toward substrate and mechanism. Ann. N Y Acad. Sci. 1018, 192–198. doi: 10.1196/annals.1296.022
Kessler, R. C., Hwang, I., Labrie, R., Petukhova, M., Sampson, N. A., Winters, K. C., et al. (2008). DSM-IV pathological gambling in the National Comorbidity Survey Replication. Psychol. Med. 38, 1351–1360. doi: 10.1017/s0033291708002900
Kivlighan, K. T., Granger, D. A., and Booth, A. (2005). Gender differences in testosterone and cortisol response to competition. Psychoneuroendocrinology 30, 58–71. doi: 10.1016/j.psyneuen.2004.05.009
Koob, G. F., and Le Moal, M. (2008). Addiction and the brain antireward system. Annu. Rev. Psychol. 59, 29–53. doi: 10.1146/annurev.psych.59.103006.093548
Koob, G. F., and Volkow, N. D. (2010). Neurocircuitry of addiction. Neuropsychopharmacology 35, 217–238. doi: 10.1038/npp.2009.110
Leeman, R. F., and Potenza, M. N. (2012). Similarities and differences between pathological gambling and substance use disorders: a focus on impulsivity and compulsivity. Psychopharmacology (Berl) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Lesieur, H. R., and Blume, S. B. (1987). The south oaks gambling screen (SOGS): a new instrument for the identification of pathological gamblers. Am. J. Psychiatry 144, 1184–1188.
Liu, X., Hairston, J., Schrier, M., and Fan, J. (2011). Common and distinct networks underlying reward valence and processing stages: a meta-analysis of functional neuroimaging studies. Neurosci. Biobehav. Rev. 35, 1219–1236. doi: 10.1016/j.neubiorev.2010.12.012
Lovallo, W. R. (2006). Cortisol secretion patterns in addiction and addiction risk. Int. J. Psychophysiol. 59, 195–202. doi: 10.1016/j.ijpsycho.2005.10.007
Marinelli, M., Aouizerate, B., Barrot, M., Le Moal, M., and Piazza, P. V. (1998). Dopamine-dependent responses to morphine depend on glucocorticoid receptors. Proc. Natl. Acad. Sci. U S A 95, 7742–7747. doi: 10.1073/pnas.95.13.7742
Marinelli, M., and Piazza, P. V. (2002). Interaction between glucocorticoid hormones, stress and psychostimulant drugs*. Eur. J. Neurosci. 16, 387–394. doi: 10.1046/j.1460-9568.2002.02089.x
Meewisse, M. L., Reitsma, J. B., De Vries, G. J., Gersons, B. P., and Olff, M. (2007). Cortisol and post-traumatic stress disorder in adults: systematic review and meta-analysis. Br. J. Psychiatry 191, 387–392. doi: 10.1192/bjp.bp.106.024877
Meyer, G., Hauffa, B. P., Schedlowski, M., Pawlak, C., Stadler, M. A., and Exton, M. S. (2000). Casino gambling increases heart rate and salivary cortisol in regular gamblers. Biol. Psychiatry 48, 948–953. doi: 10.1016/s0006-3223(00)00888-x
Meyer, W. N., Keifer, J., Korzan, W. J., and Summers, C. H. (2004). Social stress and corticosterone regionally upregulate limbic N-methyl-D-aspartatereceptor (NR) subunit type NR(2A) and NR(2B) in the lizard Anolis carolinensis. Neuroscience 128, 675–684. doi: 10.1016/j.neuroscience.2004.06.084
Nurnberger, J. I., Blehar, M. C., Kaufmann, C. A., and York-Cooler, C. (1994). Diagnostic interview for genetic studies: rationale, unique features and training. Arch. Gen. Psychiatry 51, 849–859. doi: 10.1001/archpsyc.1994.03950110009002
Oei, N. Y., Both, S., Van Heemst, D., and Van Der Grond, J. (2014). Acute stress-induced cortisol elevations mediate reward system activity during subconscious processing of sexual stimuli. Psychoneuroendocrinology 39, 111–120. doi: 10.1016/j.psyneuen.2013.10.005
Ossewaarde, L., Qin, S., Van Marle, H. J., Van Wingen, G. A., Fernández, G., and Hermans, E. J. (2011). Stress-induced reduction in reward-related prefrontal cortex function. Neuroimage 55, 345–352. doi: 10.1016/j.neuroimage.2010.11.068
Oswald, L. M., and Wand, G. S. (2004). Opioids and alcoholism. Physiol. Behav. 81, 339–358. doi: 10.1016/j.physbeh.2004.02.008
Oswald, L. M., Wong, D. F., Mccaul, M., Zhou, Y., Kuwabara, H., Choi, L., et al. (2005). Relationships among ventral striatal dopamine release, cortisol secretion and subjective responses to amphetamine. Neuropsychopharmacology 30, 821–832. doi: 10.1038/sj.npp.1300667
Paris, J. J., Franco, C., Sodano, R., Freidenberg, B., Gordis, E., Anderson, D. A., et al. (2010b). Sex differences in salivary cortisol in response to acute stressors among healthy participants, in recreational or pathological gamblers and in those with posttraumatic stress disorder. Horm. Behav. 57, 35–45. doi: 10.1016/j.yhbeh.2009.06.003
Paris, J. J., Franco, C., Sodano, R., Frye, C., and Wulfert, E. (2010a). Gambling pathology is associated with dampened cortisol response among men and women. Physiol. Behav. 99, 230–233. doi: 10.1016/j.physbeh.2009.04.002
Petry, N. M. (2007). Gambling and substance use disorders: current status and future directions. Am. J. Addict. 16, 1–9. doi: 10.1080/10550490601077668
Petry, N. M., Blanco, C., Auriacombe, M., Borges, G., Bucholz, K., Crowley, T. J., et al. (2013). An overview of and rationale for changes proposed for pathological gambling in DSM-5. J. Gambl. Stud. doi: 10.1007/s10899-013-9370-0. [Epub ahead of print].
Piazza, P. V., Rouge-Pont, F., Deroche, V., Maccari, S., Simon, H., and Le Moal, M. (1996). Glucocorticoids have state-dependent stimulant effects on the mesencephalic dopaminergic transmission. Proc. Natl. Acad. Sci. U S A 93, 8716–8720. doi: 10.1073/pnas.93.16.8716
Porcelli, A. J., Lewis, A. H., and Delgado, M. R. (2012). Acute stress influences neural circuits of reward processing. Front. Neurosci. 6:157. doi: 10.3389/fnins.2012.00157
Potenza, M. N. (2006). Should addictive disorders include non-substance-related conditions? Addiction 101, 142–151. doi: 10.1111/j.1360-0443.2006.01591.x
Potenza, M. N. (2008). Review. The neurobiology of pathological gambling and drug addiction: an overview and new findings. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3181–3189. doi: 10.1098/rstb.2008.0100
Potenza, M. N. (2013). Neurobiology of gambling behaviors. Curr. Opin. Neurobiol. 23, 660–667. doi: 10.1016/j.conb.2013.03.004
Ramirez, L. F., Mccormick, R. A., and Lowy, M. T. (1988). Plasma cortisol and depression in pathological gamblers. Br. J. Psychiatry 153, 684–686. doi: 10.1192/bjp.153.5.684
Reilly, D. (2012). Exploring the science behind sex and gender differences in cognitive abilities. Sex Roles 67, 247–250. doi: 10.1007/s11199-012-0134-6
Robinson, T. E., and Berridge, K. C. (1993). The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-p
Robinson, T. E., and Berridge, K. C. (2008). The incentive sensitization theory of addiction: some current issues. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3137–3146. doi: 10.1098/rstb.2008.0093
Rupp, H. A., and Wallen, K. (2008). Sex differences in response to visual sexual stimuli: a review. Arch. Sex. Behav. 37, 206–218. doi: 10.1007/s10508-007-9217-9
Sapolsky, R. M., Romero, L. M., and Munck, A. U. (2000). How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 21, 55–89. doi: 10.1210/er.21.1.55
Saunders, J. B., Aasland, O. G., Babor, T. F., de la Fuente, J. R., and Grant, M. (1993). Development of the alcohol use disorders identification test (AUDIT): WHO collaborative project on early detection of persons with harmful alcohol consumption–II. Addiction 88, 791–804. doi: 10.1111/j.1360-0443.1993.tb02093.x
Schultz, W. (2011). Potential vulnerabilities of neuronal reward, risk, and decision mechanisms to addictive drugs. Neuron 69, 603–617. doi: 10.1016/j.neuron.2011.02.014
Sescousse, G., Barbalat, G., Domenech, P., and Dreher, J. C. (2013). Imbalance in the sensitivity to different types of rewards in pathological gambling. Brain 136, 2527–2538. doi: 10.1093/brain/awt126
Stephens, M. A. C., and Wand, G. (2012). Stress and the HPA axis: role of glucocorticoids in alcohol dependence. Alcohol. Res. 34, 468–483.
Stevens, J. S., and Hamann, S. (2012). Sex differences in brain activation to emotional stimuli: a meta-analysis of neuroimaging studies. Neuropsychologia 50, 1578–1593. doi: 10.1016/j.neuropsychologia.2012.03.011
Tobler, P. N., Fletcher, P. C., Bullmore, E. T., and Schultz, W. (2007). Learning-related human brain activations reflecting individual finances. Neuron 54, 167–175. doi: 10.1016/j.neuron.2007.03.004
Toneatto, T., and Nguyen, L. (2007). “Individual characteristics and problem gambling behavior,” in Research and Measurement Issues in Gambling Studies, eds G. Smith, D. C. Hodgins and R. Williams (New York: Elsevier), 279–303.
Tschibelu, E., and Elman, I. (2010). Gender differences in psychosocial stress and in its relationship to gambling urges in individuals with pathological gambling. J. Addict. Dis. 30, 81–87. doi: 10.1080/10550887.2010.531671
Ulrich-Lai, Y. M., and Herman, J. P. (2009). Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 10, 397–409. doi: 10.1038/nrn2647
van den Bos, R., Davies, W., Dellu-Hagedorn, F., Goudriaan, A. E., Granon, S., Homberg, J., et al. (2013). Cross-species approaches to pathological gambling: a review targeting sex differences, adolescent vulnerability and ecological validity of research tools. Neurosci. Biobehav. Rev. 37, 2454–2471.
van Holst, R. J., van den Brink, W., Veltman, D. J., and Goudriaan, A. E. (2010). Why gamblers fail to win: a review of cognitive and neuroimaging findings in pathological gambling. Neurosci. Biobehav. Rev. 34, 87–107. doi: 10.1016/j.neubiorev.2009.07.007
Vest, R. S., and Pike, C. J. (2013). Gender, sex steroid hormones and Alzheimer’s disease. Horm. Behav. 63, 301–307. doi: 10.1016/j.yhbeh.2012.04.006
Vezina, P. (2004). Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci. Biobehav. Rev. 27, 827–839. doi: 10.1016/j.neubiorev.2003.11.001
Vezina, P. (2007). Sensitization, drug addiction and psychopathology in animals and humans. Prog. Neuropsychopharmacol. Biol. Psychiatry 31, 1553–1555. doi: 10.1016/j.pnpbp.2007.08.030
Vinson, G. P., and Brennan, C. H. (2013). Addiction and the adrenal cortex. Endocr. Connect. doi: 10.1530/ec-13-0028. [Epub ahead of print].
Wand, G. S., Oswald, L. M., Mccaul, M. E., Wong, D. F., Johnson, E., Zhou, Y., et al. (2007). Association of amphetamine-induced striatal dopamine release and cortisol responses to psychological stress. Neuropsychopharmacology 32, 2310–2320. doi: 10.1038/sj.npp.1301373
Wareham, J. D., and Potenza, M. N. (2010). Pathological gambling and substance use disorders. Am. J. Drug Alcohol Abuse 36, 242–247. doi: 10.3109/00952991003721118
Wehrum, S., Klucken, T., Kagerer, S., Walter, B., Hermann, A., Vaitl, D., et al. (2013). Gender commonalities and differences in the neural processing of visual sexual stimuli. J. Sex. Med. 10, 1328–1342. doi: 10.1111/jsm.12096
Wingenfeld, K., and Wolf, O. T. (2011). HPA axis alterations in mental disorders: impact on memory and its relevance for therapeutic interventions. CNS Neurosci. Ther. 17, 714–722. doi: 10.1111/j.1755-5949.2010.00207.x
Wohl, M. J. A., Matheson, K., Young, M. M., and Anisman, H. (2008). Cortisol rise following awakening among problem gamblers: dissociation from comorbid symptoms of depression and impulsivity. J. Gambl. Stud. 24, 79–90. doi: 10.1007/s10899-007-9080-6
Wong, G., Zane, N., Saw, A., and Chan, A. K. K. (2013). Examining gender differences for gambling engagement and gambling problems among emerging adults. J. Gambl. Stud. 29, 171–189. doi: 10.1007/s10899-012-9305-1
Keywords: cortisol, reward, pathological gambling, fMRI, ventral striatum, addiction, incentive, glucocorticoid hormones
Citation: Li Y, Sescousse G and Dreher J-C (2014) Endogenous cortisol levels are associated with an imbalanced striatal sensitivity to monetary versus non-monetary cues in pathological gamblers. Front. Behav. Neurosci. 8:83. doi: 10.3389/fnbeh.2014.00083
Received: 14 November 2013; Accepted: 25 February 2014;
Published online: 25 March 2014.
Edited by:
Mike James Ferrar Robinson, Wesleyan University, USAReviewed by:
Guido Van Wingen, Academic Medical Center Amsterdam, NetherlandsEve Limbrick-Oldfield, University of Cambridge, UK
Copyright ©2014 Li, Sescousse and Dreher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yansong Li, Reward and Decision Making Team, Centre de Neurosciences Cognitives, CNRS, UMR 5229, 67 Boulevard Pinel, 69675, Lyon, France e-mail: yansong.li@isc.cnrs.fr
†Present address: Guillaume Sescousse, Donders Institute for Brain, Cognition and Behavior, Radboud University Nijmegen, Nijmegen, Netherlands