 Gemma Navarro1,2,3
Gemma Navarro1,2,3 Paula Morales4,5
Paula Morales4,5 Carmen Rodríguez-Cueto2,6,7
Carmen Rodríguez-Cueto2,6,7 Javier Fernández-Ruiz2,6,7
Javier Fernández-Ruiz2,6,7 Nadine Jagerovic4
Nadine Jagerovic4 Rafael Franco1,2,3*
Rafael Franco1,2,3*- 1Department of Biochemistry and Molecular Biomedicine, University of Barcelona, Barcelona, Spain
- 2Centro de Investigación en Red sobre Enfermedades Neurodegenerativas (CIBERNED), Instituto de Salud Carlos III, Madrid, Spain
- 3Cell and Molecular Neuropharmacology, Institut de Biomedicina (IBUB), Universitat de Barcelona, Barcelona, Spain
- 4Instituto de Química Médica, Consejo Superior de Investigaciones Científicas, Madrid, Spain
- 5Center for Drug Discovery, University of North Carolina at Greensboro, Greensboro, NC, USA
- 6Departamento de Bioquímica, Facultad de Medicina, Instituto Universitario de Investigación en Neuroquímica, Universidad Complutense, Madrid, Spain
- 7Instituto Ramón y Cajal de Investigación Sanitaria, Madrid, Spain
Endocannabinoids activate two types of specific G-protein-coupled receptors (GPCRs), namely cannabinoid CB1 and CB2. Contrary to the psychotropic actions of agonists of CB1 receptors, and serious side effects of the selective antagonists of this receptor, drugs acting on CB2 receptors appear as promising drugs to combat CNS diseases (Parkinson's disease, Huntington's chorea, cerebellar ataxia, amyotrohic lateral sclerosis). Differential localization of CB2 receptors in neural cell types and upregulation in neuroinflammation are keys to understand the therapeutic potential in inter alia diseases that imply progressive neurodegeneration. Medicinal chemistry approaches are now engaged to develop imaging tools to map receptors in the living human brain, to develop more efficacious agonists, and to investigate the possibility to develop allosteric modulators.
Introduction
To date only two cannabinoid receptors have been identified and completely accepted as key members of the endocannabinoid signaling. The CB1 receptor (CB1R) is mainly expressed in the central nervous system (CNS) (Hu and Mackie, 2015), whereas, the CB2 receptor (CB2R) is mainly expressed in the periphery, especially in blood cells, and in blood-cell producing organs (Onaivi et al., 1999; Atwood and Mackie, 2010; Atwood et al., 2012). Other receptors, e.g., GPR55, the cation channel TRPV1 and the nuclear receptors of the PPAR family, are also under discussion as possible members of the endocannabinoid receptor family. CB1R and CB2R belong to the most populated family of the human proteome, i.e., to the family of receptors coupled to heterotrimeric G proteins (GPCRs). More specifically they are members of class A GPCRs, which are characterized by being structurally similar to rhodopsin, for having an extracellular N-terminal domain, a seven α-helical transmembrane domain, and a C-terminal domain of 73 (for CB1R) or of 59 (for CB2R) amino acids. Total length of the most common1 protein products is 472 for CB1R and 360 for CB2R. The difference in receptor length comes from the bigger N-terminal domain of the CB1R (116 vs. 33 amino acids).
Soon after its discovery and the realization of the relevant role of endogenous cannabinoids, the CB1R was considered a potential target to combat CNS diseases. In fact, the CB1R is considered the class A GPCR member with the highest expression in the CNS. In sharp contrast, controversy surrounds expression of CB2R in the CNS, and until recently this receptor was not considered as target for neurological or neuropsychiatric diseases (Atwood and Mackie, 2010; Atwood et al., 2012). This paper scans the literature that supports the view that CB2R may have now more potential than CB1R to combat some CNS disorders, in particular those related to neuroinflammatory, and neurodegenerative events. The paper also informs on current developments in medicinal chemistry aspects of CB2R-based CNS drug discovery.
Better Prospects for CB2R than for CB1R in CNS Diseases
GPCRs constitute the target of approximately 40% of approved drugs. Drug development programs are still heavily relying on the potential of GPCRs for a huge variety of diseases. Agonists, which are able to activate the receptor and compete with the endogenous agonist, and antagonists, which block the receptor and impede activation by the endogenous agonist, have therapeutic potential. However, the number of medications that consist of GPCR antagonists outnumbers that of GPCR agonists. In general terms, the higher success of antagonists means that they have fewer side effects than agonists, although other causes overlay. The endocannabinoid system is a very special case as endogenous compounds produced by neurons and acting on central CB1Rs are absolutely required for higher brain functions, but any synthetic or natural (e.g., Δ9-tetrahydrocannabinol) agonist reaching the brain and hitting CB1R has proved to have psychotropic actions in animal models of disease and in humans. Therefore, the potential of CB1Rs as targets for diseases of the CNS, and also peripheral disorders, has been limited by the psychoactive side effects derived from their agonists, and for the need to consider the risk-benefit balance. In this context, some researchers wanted to develop CB1R antagonists (including inverse agonists) as a safer alternative in those pathologies having an overactivity of the endocannabinoid system (e.g., obesity, addiction, schizophrenia), although side effects were also evident with such strategy (see below).
The first two molecules targeting CB1R that reached the therapeutic market (in the 80s) were Δ9-tetrahydrocannabinol, also known as dronabinol (marketed as Marinol®), and nabilone (marketed as Cesamet®) (Figure 1), both prescribed to combat nausea and vomiting, as well anorexia, derived from cancer, and AIDS treatments, respectively (Green et al., 1989), but their use was limited. By contrast, a CB1R antagonist/inverse agonist, rimonabant (Acomplia®), was approved in 2006 to treat obesity, and metabolic syndrome (Carai et al., 2006) and generated extremely high expectations. Unfortunately, the drug had to be retired due to side effects, especially due to reports of suicide (Sam et al., 2011). Consequently, chances, that other CB1R selective drug may advance though regulatory bodies, and reach the market have dramatically diminished. In this context, the CB2R has taken the lead in the race to find novel cannabinoid-related drugs for CNS diseases. On the one hand, CB1R is expressed in almost any brain region, and in many neuronal cell types, whereas CB2R expression in neurons is restricted to few areas. Accordingly, fewer side effects are expected when drugs are targeting receptors with restricted expression than when drugs are targeting receptors widely expressed in the CNS. Furthermore, CB2R are upregulated in a variety of CNS diseases that course with activated microglia or astroglia. Then the CB2R but not the CB1R is a promising candidate to consider in diseases with a neuroinflammatory component. It is even possible that the activation of CB2Rs may explain recent controversies in relation with the consumption of cannabis as a factor either increasing risk or preventing against spontaneous brain insults (e.g., intracerebral hemorrhage). Recent epidemiological studies suggest a potential protective effect of cannabis to the modulation of C-reactive protein response in intracerebral hemorrhage (Di Napoli et al., 2012, 2016; Alshaarawy and Anthony, 2015), an effect that could be possibly related to CB2R activation, although this has not been investigated. Advantages of developing CB2R selective drugs to prevent neurodegeneration in cases of neuroinflammation are presented later in this article.
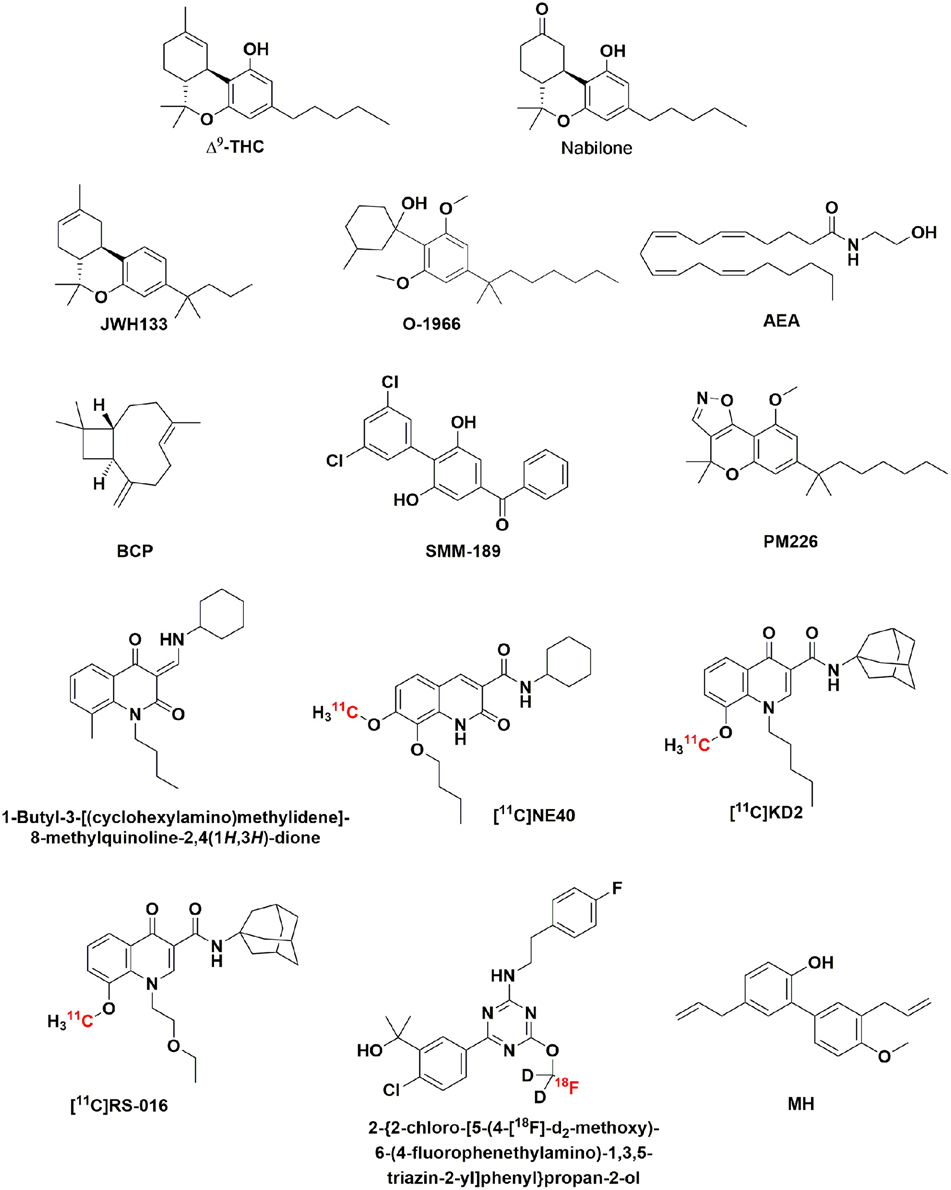
Figure 1. Chemical structure of Δ9-THC, nabilone, and the CB2R ligands: JWH133, 0-1966, AEA, BCP, SMM-189, PM226, 1-butyl-3-[(cyclohexylamino)methylidene]-8-methylquinoline-2,4(1H,3H)-dione, [11C]NE40, [11C]KD2, [11C]RS-016, 2-{2-chloro-[5-(4-[18F]-d2-methoxy)-6-(4-fluorophenethylamino)-1,3,5-triazin-2-yl]phenyl}propan-2-ol.
As macrophages express CB2R and microglia is somehow a similar cell type, these receptors were soon identified in microglial cells, but further research demonstrated that they can be also found in other types of glial cells (see below). There is however, some controversy on the degree of CB2R expression in resting vs. activated microglial cells. Also the activated microglial phenotype is different in macrophages filtered from the blood into the CNS and in resident microglia that becomes activated due to, inter alia, accumulation of protein aggregates such as alpha-synuclein, or ß-amyloid. Remarkably, (see Franco and Fernández-Suárez, 2015 and references therein) a better understanding of the expression and role of CB2R in the different microglial phenotypes (M0, M1, M2) will help in designing CB2R selective ligands able to induce the neuroprotective/anti-inflammatory-skewed phenotype(s).
CB2R may be also expressed by CNS neurons. The role of CB2Rs in schizophrenia, depression, food consumption, and drug addiction has been demonstrated in different laboratories and the results are consistent with neuronal expression of the receptor (Onaivi et al., 2008a,b,c; Hu et al., 2009; García-Gutiérrez et al., 2010; Ishiguro et al., 2010a,b; García-Gutiérrez and Manzanares, 2011; Ortega-Alvaro et al., 2011; Aracil-Fernández et al., 2012; Navarrete et al., 2012, 2013; Bahi et al., 2014; Blanco-Calvo et al., 2014; Ortega-Álvaro et al., 2015; Rodríguez-Arias et al., 2015; García-Cabrerizo and García-Fuster, 2016). The receptor is significantly expressed in neurons in the brain stem (Van Sickle et al., 2005), in the cerebellum (Skaper et al., 1996; Ashton et al., 2006; Gong et al., 2006; Rodríguez-Cueto et al., 2014) in the internal and the external segments of the globus pallidus of the non-human primate (Lanciego et al., 2011), and in the substantia nigra (in humans, not in rodents) (García et al., 2016; Gómez-Gálvez et al., 2016). Different laboratories working with rodents or primates have also identified receptor expression in neurons of the prefrontal cortex and hippocampus (Callén et al., 2012; den Boon et al., 2012; Sierra et al., 2015; García-Cabrerizo and García-Fuster, 2016). Expression of CB2R in the basal ganglia show promise in Parkinson's disease and Huntington's chorea; the presence of the receptor in hippocampus and prefrontal cortex makes it attractive for Alzheimer's disease and the expression in brain stem and cerebellum opens novel therapeutic avenues for a variety of diseases such as hereditary spinocerebellar ataxias. Last but not least, the data on CB2R-mediated endocannabinoid regulation of microglial activation makes the receptor attractive for diseases with a neuroinflammatory component.
Cannabinoid neuroregulation is mainly based on retrograde signaling (Alger, 2002), i.e., endocannabinoids come from post-synaptic elements to activate presynaptic receptors. However, postsynaptic CB2Rs have been also reported (Brusco et al., 2008). The combination of restricted neuronal expression with the possibility of targeting pre- or postsynaptic receptors, makes the CB2R a really attractive target.
CB2R in Neurodegenerative Disorders. Relevance of Differential Expression of CB2R in Neural Cells
The preservation of neuronal integrity and survival is one of the most promising therapeutic possibilities of CB2R-targeting cannabinoids (Atwood et al., 2012). There is potential in pain and in numerous acute or chronic neurodegenerative/neuroinflammatory conditions (Jhaveri et al., 2007; Micale et al., 2007; Campillo and Páez, 2009). The neuroprotective potential of compounds targeting the CB2R is, first of all, the logical consequence of their location in key cell types (e.g., in specific neuronal subsets, activated astrocytes, reactive microglia, perivascular microglia, oligodendrocytes, and neural progenitor cells), and also in some structures (e.g., the blood-brain barrier (BBB)) that are critical for the maintenance of the CNS integrity (Amenta et al., 2012; Chung et al., 2016) (Figure 2A). Such variety of locations enable compounds capable to selectively activate the CB2R to exert a selective control over the specific functions fulfilled by these cells in degeneration, protection and/or repair (Fernández-Ruiz et al., 2014). For example, BBB function is under the control of CB2R-mediated signals (Fujii et al., 2014), which maintain the integrity of tight junctions, inhibit leukocyte infiltration, and facilitate β-amyloid clearance (Vendel and de Lange, 2014).
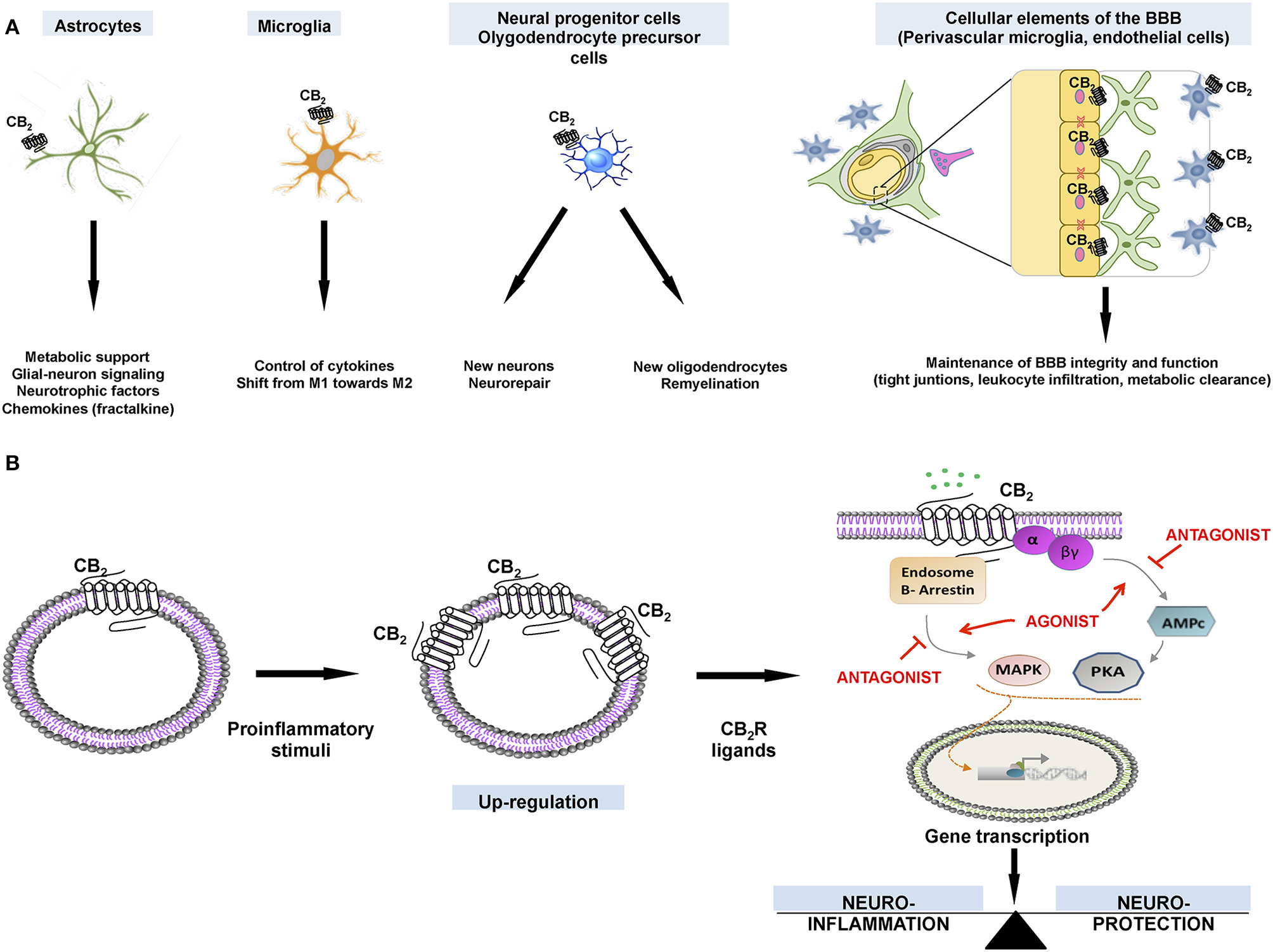
Figure 2. (A). Expression of CB2Rs in different neural cell types and how receptor activation may impact on cell-specific functions. (B) Cellular events that explain the therapeutic possibilities for ligands that target CB2Rs, which are upregulated in activated glial cells.
CB2Rs in glial cells recruited to the site of the neurodegeneration, appear to be critical for preserving the neuronal integrity and function (Savonenko et al., 2015). In fact, CB2R may be absent of these cells in resting conditions, with a weak expression in the healthy brain. As the receptors are strongly up-regulated when glial cells are activated in conditions of neurodegeneration (Fernández-Ruiz et al., 2007, 2015), they have potential from a therapeutic point of view (Figure 2B). Up-regulation may occur in both astrocytes and microglial cells, but the CB2R-mediated signaling may vary depending inter alia on the type of pathology and the experimental model. CB2R-mediated neuroprotection/neurorestoration mechanisms are of special interest in disorders that affect movement-related areas, such as (i) Parkinson's and Huntington's diseases (affecting the basal ganglia, and producing rigidity, postural instability, bradykinesia, tremor, and chorea), (ii) autosomal dominant spinocerebellar ataxias (affecting the cerebellum and its afferent and efferent connections, and producing loss of balance, and motor incoordination), and (iii) amyotrophic lateral sclerosis (ALS) (affecting upper and lower spinal motor neurons, and producing muscle denervation and atrophy, which results in a progressive weakness and paralysis affecting voluntary muscles). For example, in this last disorder, CB2Rs become up-regulated in microglial cells recruited at the spinal cord of patients (Yiangou et al., 2006), a fact corroborated by studies in the TDP-43 mouse model of the disease (Espejo-Porras et al., 2015). However, apart from microglial cells, other CB2R-positive cells were found in this murine model (Espejo-Porras et al., 2015). In another murine model of ALS, (the SOD-1 mouse), CB2R also become up-regulated, but the study did not characterize the type of cell that was expressing the receptors (Shoemaker et al., 2007).
Interestingly, microglial CB2Rs appear up-regulated in the cerebellum of patients with different autosomal dominant cerebellar ataxias, but such trend was also found in activated astrocytes located in the cerebellar parenchyma and in the periphery of blood vessels, and in certain neuronal subpopulations (Rodríguez-Cueto et al., 2014). Similarly, increased levels of CB2R are found in both striatal activated astrocytes and reactive microglial cells after an insult with malonate in rats, an experimental model of Huntington's disease (Sagredo et al., 2009). Although data collected from Huntington's disease patients or obtained in genetic models of the disease (e.g., R6/1, R6/2) indicated that CB2R were located and up-regulated only in microglial cells (Palazuelos et al., 2009), a more recent study situated the up-regulation of these receptors in vascular cells, not in activated glial cells, in HD patients (Dowie et al., 2014).
In yet another neurodegenerative condition affecting the basal ganglia circuits, (Price et al., 2009) were the first to demonstrate up-regulation of CB2R in microglial cells recruited at the substantia nigra in MPTP-lesioned mice. In the study it was not addressed whether there were other CB2R-positive cells that do not correspond to reactive microglia. We investigated the issue in parkinsonian patients using postmortem samples and identified such up-regulation in microglial cells (labeled with Iba-1) and in another unidentified cell type (Gómez-Gálvez et al., 2016).
CB2R has potential in demyelinating disorders (e.g., multiple sclerosis; Molina-Holgado et al., 2002; Gomez et al., 2010, 2011). In fact, CB2R are present in oligodendrocytes, and more importantly, in their natural precursor cells, so that they may play a role in their survival, proliferation, and differentiation. CB2Rs have been also identified in neural progenitor cells, and it appears that they can play a role in the proliferation and differentiation of these precursors (Palazuelos et al., 2006, 2012; Goncalves et al., 2008; Avraham et al., 2014), opening the possibility to facilitate neurorestoration by pharmacologically manipulating this receptor. Lastly, the identification of CB2Rs in perivascular microglial cells in the cerebellum (Núñez et al., 2004) may be possibly related to the role attributed to these receptors at the level of the BBB (see above).
Challenges in CB2R-Based Drug Design
Pharmacology of cannabinoid receptors is complex due to the lipophilic nature of many natural and synthetic agonists. Endogenous agonists of many class A GPCRs are hydrophilic, which contrast with the lipophilic nature of endocannabinoids. Pharmacological characterization by radioligand binding to CB2R is especially complex. On the one hand, the binding site extends deeply within the seven transmembrane domain of the receptor, and the two available radiolabeled ligands (tritiated CP-55940 and tritiated WIN-55212-2) do not interact with exactly the same amino acid residues in the orthosteric center; in particular CP-55940 does not interact with a conserved lysine residue in the binding site (Tao et al., 1999). Furthermore, it is hypothesized that cannabinoids may not reach the binding site from the outside of the cells but by lateral diffusion via the lipid bilayer of the plasma membrane (Guo et al., 2003; Makriyannis et al., 2005; Hurst et al., 2010). These features suggest that newly synthesized drugs or newly discovered natural cannabinoids have qualitatively different modes of binding to CB2Rs. On the other hand, the nonspecific binding to membranes from natural CNS sources is high and leads to low-confidence values of the amount of receptor in neural cells. This problem is partially solved by performing the assays in heterologous cells expressing the human receptor; such approach provides reliable parameters for drug discovery. The complex pharmacology is also slowing the discovery of allosteric centers, and accordingly, of allosteric CB2R modulators.
GPCR pharmacology must somehow be revisited due to the occurrence of receptor heteromers (Cordomí et al., 2015; Franco et al., 2016). Each heteromer is unique and functionally different from the two constituting receptors. In fact, affinity of agonists/antagonists may change when a given receptor is forming heteroreceptor complexes, and more importantly, signaling cascades may be heteromer-specific (Ferré et al., 2009; Franco et al., 2016). Also relevant is the fact that presynaptic heteromers seem to be different from those in post-synaptic locations, i.e., a given GPCR may form different heteromers in pre- or post-synaptic membranes. Cannabinoid receptors may form a variety of heteromers with other class a GPCRs (see www.gpcr-hetnet.com; Borroto-Escuela et al., 2014). Interestingly, the two cannabinoid receptors may interact and give rise to CB1R-CB2R heteromers (Callén et al., 2012; Sierra et al., 2015). In agreement with the widespread distribution of CB2Rs in brain and the robust expression of CB2Rs in the globus pallidus, CB1R-CB2R heteromers are abundant in basal ganglia output neurons; available data indicate that these CB1R-CB2R heteromers are mainly post-synaptic. Pallidal expression of heteromers investigated in a primate model of Parkinson's disease was evident in naïve and parkinsonian animals, but it was markedly reduced in the levodopa-induced dyskinetic group (Sierra et al., 2015). Although likely, cannabinoid-receptor containing heteromers have not been identified and characterized in glial cells. Heteromer expression is worth considering on designing drugs targeting CB2R. In particular pallidal CB1R-CB2R heteromers constitute a specific target in Parkinson's disease. A main advantage of selectively targeting GPCR heteromers, i.e., to use drugs that preferentially act on heteromer-expressing cells, is the reduction of side effects.
CB2R Ligands as Therapeutic Agents in CNS Diseases
Positron Emission Tomography Reagents for Brain Imaging
Studies of CB2R ligands as diagnostic agents for noninvasive brain imaging have been reported. Positron emission tomography (PET) provides a sensitive and non-invasive imaging technique to quantify CB2R expression in the CNS. This technique requires radioligands with high affinity and high specificity toward CB2R. Despite the development of highly selective CB2R ligands (Han et al., 2014), a limited number of PET radiotracers for imaging CB2R have been reported. Whereas, novel PET tracers for CB1R in brain imaging have been evaluated in clinical trials, few CB2R radioligands have been tested in humans. Few years ago, the first PET tracers for CB2R were presented as candidates for the in vivo imaging of neuroinflammatory events (Evens and Bormans, 2010). Preliminary clinical assays of the first CB2R radioligand, [11C]NE40 (Figure 1), showed appropriate fast brain kinetics in the healthy human brain (Ahmad et al., 2013). A major challenge is the development of CB2R PET agents with maximized brain penetration and minimized non-specific binding. In this sense, structural optimization of [11C]KD2 (Figure 1) (Mu et al., 2013), a potential PET tracer with poor brain penetration, led to the discovery of [11C]RS-016 (Figure 1), which showed slightly improved blood-brain penetration, and higher specific CB2R binding in murine spleen tissues and postmortem ALS patient spinal cord tissues (Contartese et al., 2012; Slavik et al., 2015a,b). A promising PET tracer candidate for the in vivo evaluation of neuroinflammation and disease progression has been recently described (Hortala et al., 2014). A triazine derivative labeled with the long-lasting radionucleotide fluorine-18 (Figure 1), 2-{2-chloro-[5-(4-[18F]-d2-methoxy)-6-(4-fluorophenethylamino)-1,3,5-triazin-2-yl]phenyl}propan-2-ol, showed in rhesus macaques, and baboons significant brain uptake and moderate washout.
Current Medicinal Chemistry Approaches
Often, increased levels of the endogenous cannabinoid, anandamide (AEA, Figure 1), correlate with neurodegenerative conditions. In recent studies, AEA has been shown to alleviate lipopolysaccharide-induced neuroinflammation in rat primary microglial cultures. Even though AEA can activate CB1R, CB2R, and other receptors such as GPR55, GPR18, TRPV1, or PPARs, the anti-inflammatory effects seem to be CB2R-mediated, although a possible functional cross talk with GPR18/GPR55 cannot be ruled out (Malek et al., 2015). Accordingly, AEA may have potential therapeutic action on managing microglial-derived neuroinflammation and may regulate many aspects of the brain's inflammatory response. However, from a medicinal chemistry perspective, drug development is more securely based on designing novel and selective CB2R ligands.
Despite the increasing number of reports on selective CB2R ligands and the high expectations with this cannabinoid target, only a few synthetic CB2R agonists have reached clinical trials (Han et al., 2014; Aghazadeh Tabrizi et al., 2016). CB2R agonists, namely GW842166X, CP55940, S-777469, and JTE-907, completed phase II for treatment of different pain conditions, but none of them has been evaluated in humans for neurodegenerative or neuroinflammatory diseases. However, preclinical data of CB2R agonists and inverse agonists have been described within this therapeutic perspective (Dhopeshwarkar and Mackie, 2014; Zhang et al., 2014).
Administration of a selective CB2R agonist, JWH-133 (Figure 1), to an animal model of brain infarction improved infarct outcome and neurological impairment through inhibition of different subpopulations of microglia and macrophages (Zarruk et al., 2012). Repeated treatments with the resorcinol-based CB2R agonist, O-1966, resulted in attenuated BBB disruption and neuronal degeneration as shown in a traumatic brain injury model (Amenta et al., 2012).
Trans-caryophyllene (BCP, Figure 1), a bicyclic sesquiterpene with selective CB2R agonist properties, has been reported as a therapeutic target for the treatment of cerebral ischemia (Guo et al., 2014). This sesquiterpene suppressed hypoxia-induced neuroinflammatory responses by inhibiting NF-κB activation in microglia. Effectively, studies performed in the microglial cell line BV-2 and in primary cultures of microglia indicated that the inhibitory action of both cannabinoid receptor agonists and antagonists was mediated by extracellular signal regulated kinase 1/2 (ERK1/2), cytosolic phospholipase A2 (cPLA2), and activation of nuclear factor kappa (NF-κB) (Ribeiro et al., 2013).
New potentially neuroprotective CB2R ligands have been recently described. Among them, the novel CB2R inverse agonist SMM-189 (Figure 1) (Ki(CB2) = 121 nM; Ki(CB1) = 4780 nM; EC50 = 153 nM) showed in a murine model of mild traumatic brain injury efficacy in reducing the motor, visual, and emotional deficits; such neuroprotection was seemingly achieved by modulating microglial activation (Reiner et al., 2015) and chemokine expression. Reduction of the proinflammatory markers, oetaxin, MCP-1, and IP-10 by SMM-189 suggests that SMM-189 would decrease infiltration of peripheral macrophage and other cells of the immune system implicated in neurodegeneration events (Presley et al., 2015). The chromenoisoxazole PM226 (Figure 1) has been described as a selective CB2R agonist (Ki(CB2) = 13 nM; Ki(CB1R) > 40 μM; EC50 = 39 nM) with neuroprotective properties in vitro and in vivo evaluations (Gómez-Cañas et al., 2016). In this study, the beneficial effects of PM226 against the toxicity caused by conditioned media generated from LPS-treated cultured BV2 cells and exposed to a striatal neuron-derived cell line in culture was shown to be mediated by CB2R. This neuroprotective potential was confirmed in an in vivo model of mitochondrial damage of striatal neurons in rats. Structure-activity relationship studies on the quinoline-2,4(1H,3H)-dione scaffold allowed the discovery of the CB2R agonist 1-butyl-3-[(cyclohexylamino)methylidene]-8-methylquinoline-2,4(1H,3H)-dione (Figure 1) (EC50(CB2) = 92 nM; EC50(CB1) > 10 μM) that significantly reduced the clinical symptoms of experimental autoimmune encephalomyelitis in a mouse model of multiple sclerosis (Han et al., 2015). As shown by histological analysis, oral administration of this quinoline-2,4(1H,3H)-dione(10 mg/Kg) decreased leukocyte infiltration in the spinal cord and demyelination in white matter.
New strategies involving the targeting of CB2R have been recently proposed for neurodegenerative and neuroinflammatory diseases. One of them has been proposed recently after reporting the mechanisms that could led to the beneficial effects of 4′-O-methylhokiol (MH, Figure 1), the major bioactive component of Magnolia grandiflora L., in animal models of neurodegeneration (Chicca et al., 2015). MH exerts dual actions on the endocannabinoid system by acting as CB2R modulator and COX-2 substrate-specific inhibitor.
Another strategy that needs to be explored is targeting CB2R homo o heterodimers. Homobivalent and heterobivalent ligands have been explored for several GPCRs such as opioid (Fulton et al., 2010), dopamine (Gogoi et al., 2012), or histamine receptors (Birnkammer et al., 2012). CB1R homobivalent and heterobivalent ligands have been designed and reported in the literature (Nimczick and Decker, 2015). In what concerns CB2R dimers, the first structurally bivalent compounds was designed and synthesized in 2014 (Nimczick et al., 2014). Unfortunately, these molecules have less activity and selectivity compared to their monomeric compound. Bivalent molecules showed to be weak antagonists/inverse agonists of CB1 and CB2 receptors whereas the monomeric parent was selective CB2R agonist (Nimczick et al., 2014). It appears that the development of bivalent drugs for CB2Rs is still a complex task as commented very recently (Glass et al., 2016). Reported bivalent CB1 receptor ligands are too short to bind both receptors simultaneously. The strategy for CB1 or CB2 receptor dimers need to be reviewed due to the fact that the ligand reaches the binding site through the lipid bilayer and the linkers are unlikely to be at the external receptor face.
Despite the promising therapeutic potential offered by CB2R agonists, their translational success depends on overcoming some limitations, such as immune suppression upon chronic use- or pro-inflammatory actions. There is growing evidence that CB1Rs are subject to ligand-biased signaling (Khajehali et al., 2015). However, ligand-biased signaling profiles of ligands at CB2R are still under scrutiny; certainly, upon validation, they could open new therapeutic approaches. For example, the endocannabinoid 2-arachidonoylglycerol is very potent activating the ERK1/2-MAPK pathway at low concentration, whereas the inhibition of the adenylyl cyclase and calcium pathways needs higher concentrations (Dhopeshwarkar and Mackie, 2014). In the near future allosteric modulation at CB2R may offer a novel therapeutic approach as allosteric modulators may both fine-tune the receptor response and minimize side-effects. Signaling-specific allosteric modulation as well as orthosteric probe dependence at CB1R is currently under intense focus (Morales et al., 2016). In what concerns the CB2R, positive and negative CB2R allosteric modulators still need to be discovered.
Targeting CB2R in Neurodegenerative Disorders
As above mentioned, drugs specifically targeting CB2R in pallidal neurons may provide symptomatic relief in Parkinson's disease. However, neuroprotection is more likely afforded by guiding glial cells to protect or restore neuronal damage. The expression of CB2R by glia enables these receptors to participate in the control by glial cells of the neuronal homeostasis, integrity and survival, particularly when glial cells become reactive (Fernández-Ruiz et al., 2007, 2015). Such potential situates cannabinoid ligands acting on CB2Rs in a promising position for being used in neuroprotection (Figure 2B) (Fernández-Ruiz et al., 2015). Such pharmacological manipulations may be the best way to modulate the endogenous response provoked by these receptors, which are up-regulated in activated astrocytes and reactive microglia in response to inflammatory, excitotoxic and traumatic insults. Accordingly, preserving healthy neurons, or rescuing damaged neurons may be likely achieved by selecting the right agonist or allosteric modulator of CB2R (see Figure 2B).
In the case of activated astrocytes, the benefits derived from the activation of CB2R may be associated with: (i) increasing the trophic role exerted by these glial cells, including the supply of metabolic substrates to neurons (Köfalvi et al., 2016); (ii) enhancing the generation of neurotrophins (e.g., GDNF), anti-inflammatory mediators (e.g., interleukin-10, interleukin-1 receptor antagonist), and/or pro-survival factors (e.g., transforming growth factor-β) (Smith et al., 2000; Molina-Holgado et al., 2003); and (iii) inhibiting the production of chemokines (e.g., fractalkine) which contribute to neuronal damage (Sheng et al., 2009). All these effects should be likely dependent on the activation of CB2R, either working alone or in conjunction with CB1R (Stella, 2010).
Microglial cells have an added value as they are recruited to the lesion site where they become reactive and change morphology and molecular phenotype. Accordingly, CB2Rs are concentrated surrounding the site of action of the therapeutic drug. The benefits derived from targeting CB2R in activated microglia may be associated with: i) regulation of migration and proliferation at lesion sites (Walter et al., 2003; Carrier et al., 2004); (ii) regulation in the production of TNF-α and other microglia-derived neurotoxic factors (Fernández-Ruiz et al., 2007, 2015; Stella, 2010); and (iii) regulation of the balance M1 (pro-inflammatory) vs. M2 (neuroprotective) phenotypes (Mecha et al., 2013; Franco and Fernández-Suárez, 2015; Malek et al., 2015; Jia et al., 2016).
Concluding Remarks and Future Perspectives
The aim of this article was to collect evidence generated in the last years in support of the therapeutic potential of compounds selectively targeting the CB2R. We placed emphasis in the potential relevance to provoke neuroprotection/neurorestoration in neurodegenerative disorders, particularly when activation of glial elements and occurrence of local inflammatory events are involved. We have compared the advantages of targeting CB2Rs over targeting other elements of the endocannabinoid signaling, in particular the CB1Rs. Right now there are a number of advantages based on the biochemical and signaling properties of CB2Rs, the characteristics of the binding site, their capability to form heteromers, and very importantly, to their differential expression and function depending on the CNS region and the neural cell type. Knowledge of the exact role of CB2R in activated glial cells will enhance the therapeutic potential of targeting these receptors in neuroinflammatory/neurodegenerative disorders.
It would be relevant to assess which among those disorders may receive more benefit from the targeting the receptor. Also relevant are the new perspectives in the design and development of novel ligands targeting the receptor. Other issues that require additional investigation are those related to the necessary developments to translate the preclinical potential of CB2Rs and their ligands to the clinical scenario. This would be the major challenge in the next 5–10 years after which the first CB2R-based medications will, hopefully, be available. Expectations are that new formulations of selective CB2R ligands active at the orthosteric binding site, or acting as allosteric modulators, used alone or in combination with other licensed medicines, will be available to combat devastating neurological disorders such as Alzheimer's disease, Parkinson's disease, ataxias or amyotrophic lateral sclerosis.
Author Contributions
All authors have contributed to the writing and to design and preparation of figures. Coordination of efforts has been carried out by the senior authors (NJ, JF, RF) of the three participating laboratories.
Funding
Open access partially supported by Grant 201413-30 from Fundació La Marató de TV3. Authors declare that personal funds are needed to carry out our research and/or to elaborate didactical materials/papers in some of our Institutions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
1. ^Isoforms of endocannabinoid receptors have been identified (details available at www.uniprot.org.)
References
Aghazadeh Tabrizi, M., Baraldi, P. G., Borea, P. A., and Varani, K. (2016). Medicinal chemistry, pharmacology, and potential therapeutic benefits of cannabinoid CB2 Receptor Agonists. Chem. Rev. 116, 519–560. doi: 10.1021/acs.chemrev.5b00411
Ahmad, R., Koole, M., Evens, N., Serdons, K., Verbruggen, A., Bormans, G., et al. (2013). Whole-body biodistribution and radiation dosimetry of the cannabinoid type 2 receptor ligand [11C]-NE40 in healthy subjects. Mol. Imaging Bio. 15, 384–390. doi: 10.1007/s11307-013-0626-y
Alger, B. E. (2002). Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog. Neurobiol. 68, 247–286. doi: 10.1016/S0301-0082(02)00080-1
Alshaarawy, O., and Anthony, J. C. (2015). Cannabis smoking and serum C-reactive protein: a quantile regressions approach based on NHANES 2005-2010. Drug Alcohol Depend. 147, 203–207. doi: 10.1016/j.drugalcdep.2014.11.017
Amenta, P. S., Jallo, J. I., Tuma, R. F., and Elliott, M. B. (2012). A cannabinoid type 2 receptor agonist attenuates blood-brain barrier damage and neurodegeneration in a murine model of traumatic brain injury. J. Neurosci. Res. 90, 2293–2305. doi: 10.1002/jnr.23114
Aracil-Fernández, A., Trigo, J. M., García-Gutiérrez, M. S., Ortega-Álvaro, A., Ternianov, A., Navarro, D., et al. (2012). Decreased cocaine motor sensitization and self-administration in mice overexpressing cannabinoid CB2 receptors. Neuropsychopharmacology 37, 1749–1763. doi: 10.1038/npp.2012.22
Ashton, J. C., Friberg, D., Darlington, C. L., and Smith, P. F. (2006). Expression of the cannabinoid CB2 receptor in the rat cerebellum: an immunohistochemical study. Neurosci. Lett. 396, 113–116. doi: 10.1016/j.neulet.2005.11.038
Atwood, B. K., and Mackie, K. (2010). CB2: a cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 160, 467–479. doi: 10.1111/j.1476-5381.2010.00729.x
Atwood, B. K., Straiker, A., and Mackie, K. (2012). CB2: therapeutic target-in-waiting. Prog. Neuropsychopharmacol. Biol. Psychiatry 38, 16–20. doi: 10.1016/j.pnpbp.2011.12.001
Avraham, H. K., Jiang, S., Fu, Y., Rockenstein, E., Makriyannis, A., Zvonok, A., et al. (2014). The cannabinoid CB2 receptor agonist AM1241 enhances neurogenesis in GFAP/Gp120 transgenic mice displaying deficits in neurogenesis. Br. J. Pharmacol. 171, 468–479. doi: 10.1111/bph.12478
Bahi, A., Al Mansouri, S., Al Memari, E., Al Ameri, M., Nurulain, S. M., and Ojha, S. (2014). β-Caryophyllene, a CB2 receptor agonist produces multiple behavioral changes relevant to anxiety and depression in mice. Physiol. Behav. 135, 119–124. doi: 10.1016/j.physbeh.2014.06.003
Birnkammer, T., Spickenreither, A., Brunskole, I., Lopuch, M., Kagermeier, N., Bernhardt, G., et al. (2012). The bivalent ligand approach leads to highly potent and selective acylguanidine-type histamine H 2 receptor agonists. J. Med. Chem. 55, 1147–1160. doi: 10.1021/jm201128q
Blanco-Calvo, E., Rivera, P., Arrabal, S., Vargas, A., Pavón, F. J., Serrano, A., et al. (2014). Pharmacological blockade of either cannabinoid CB1 or CB2 receptors prevents both cocaine-induced conditioned locomotion and cocaine-induced reduction of cell proliferation in the hippocampus of adult male rat. Front. Integr. Neurosci. 7:106. doi: 10.3389/fnint.2013.00106
Borroto-Escuela, D. O., Brito, I., Romero-Fernandez, W., Di Palma, M., Oflijan, J., Skieterska, K., et al. (2014). The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 15, 8570–8590. doi: 10.3390/ijms15058570
Brusco, A., Tagliaferro, P., Saez, T., and Onaivi, E. S. (2008). Postsynaptic localization of CB2 cannabinoid receptors in the rat hippocampus. Synapse 62, 944–949. doi: 10.1002/syn.20569
Callén, L., Moreno, E., Barroso-Chinea, P., Moreno-Delgado, D., Cortés, A., Mallol, J., et al. (2012). Cannabinoid receptors CB1 and CB2 form functional heteromers in brain. J. Biol. Chem. 287, 20851–20865. doi: 10.1074/jbc.M111.335273
Campillo, N. E., and Páez, J. A. (2009). Cannabinoid system in neurodegeneration: new perspectives in Alzheimer's disease. Mini Rev. Med. Chem. 9, 539–559. doi: 10.2174/138955709788167628
Carai, M. A. M., Colombo, G., Maccioni, P., and Gessa, G. L. (2006). Efficacy of rimonabant and other cannabinoid CB1 receptor antagonists in reducing food intake and body weight: preclinical and clinical data. CNS Drug Rev. 12, 91–99. doi: 10.1111/j.1527-3458.2006.00091.x
Carrier, E. J., Kearn, C. S., Barkmeier, A. J., Breese, N. M., Yang, W., Nithipatikom, K., et al. (2004). Cultured rat microglial cells synthesize the endocannabinoid 2-arachidonylglycerol, which increases proliferation via a CB2 receptor-dependent mechanism. Mol. Pharmacol. 65, 999–1007. doi: 10.1124/mol.65.4.999
Chicca, A., Gachet, M. S., Petrucci, V., Schuehly, W., Charles, R.-P., and Gertsch, J. (2015). 4′-O-methylhonokiol increases levels of 2-arachidonoyl glycerol in mouse brain via selective inhibition of its COX-2-mediated oxygenation. J. Neuroinflammation 12, 89. doi: 10.1186/s12974-015-0307-7
Chung, Y. C., Shin, W.-H., Baek, J. Y., Cho, E. J., Baik, H. H., Kim, S. R., et al. (2016). CB2 receptor activation prevents glial-derived neurotoxic mediator production, BBB leakage and peripheral immune cell infiltration and rescues dopamine neurons in the MPTP model of Parkinson's disease. Exp. Mol. Med. 48:e205. doi: 10.1038/emm.2015.100
Contartese, A., Valoti, M., Corelli, F., Pasquini, S., Mugnaini, C., Pessina, F., et al. (2012). A novel CB2 agonist, COR167, potently protects rat brain cortical slices against OGD and reperfusion injury. Pharmacol. Res. 66, 555–563. doi: 10.1016/j.phrs.2012.08.003
Cordomí, A., Navarro, G., Aymerich, M. S., and Franco, R. (2015). Structures for G-protein-coupled receptor tetramers in complex with g proteins. Trends Biochem. Sci. 40, 548–551. doi: 10.1016/j.tibs.2015.07.007
den Boon, F. S., Chameau, P., Schaafsma-zhao, Q., Van Aken, W., Bari, M., Oddi, S., et al. (2012). Excitability of prefrontal cortical pyramidal neurons is modulated by activation of intracellular type-2 cannabinoid receptors. Proc. Natl. Acad. Sci. U.S.A. 109, 1–6. doi: 10.1073/pnas.1118167109
Dhopeshwarkar, A., and Mackie, K. (2014). CB2 Cannabinoid receptors as a therapeutic target-what does the future hold? Mol. Pharmacol. 86, 430–437. doi: 10.1124/mol.114.094649
Di Napoli, M., Godoy, D. A., Campi, V., Masotti, L., Smith, C. J., Parry Jones, A. R., et al. (2012). C-reactive protein in intracerebral hemorrhage: time course, tissue localization, and prognosis. Neurology 79, 690–699. doi: 10.1212/WNL.0b013e318264e3be
Di Napoli, M., Zha, A. M., Godoy, D. A., Masotti, L., Schreuder, F. H. B. M., Popa-Wagner, A., et al. (2016). Prior cannabis use is associated with outcome after intracerebral hemorrhage. Cerebrovasc. Dis. 41, 248–255. doi: 10.1159/000443532
Dowie, M. J., Grimsey, N. L., Hoffman, T., Faull, R. L. M., and Glass, M. (2014). Cannabinoid receptor CB2 is expressed on vascular cells, but not astroglial cells in the post-mortem human Huntington's disease brain. J. Chem. Neuroanat. 59, 62–71. doi: 10.1016/j.jchemneu.2014.06.004
Espejo-Porras, F., Piscitelli, F., Verde, R., Ramos, J. A., Di Marzo, V., de Lago, E., et al. (2015). Changes in the endocannabinoid signaling system in CNS structures of TDP-43 transgenic mice: relevance for a neuroprotective therapy in TDP-43-related disorders. J. Neuroimmune Pharmacol. 10, 233–244. doi: 10.1007/s11481-015-9602-4
Evens, N., and Bormans, G. M. (2010). Non-invasive imaging of the type 2 cannabinoid receptor, focus on positron emission tomography. Curr. Top. Med. Chem. 10, 1527–1543. doi: 10.2174/156802610793176819
Fernández-Ruiz, J., de Lago, E., Gómez-Ruiz, M., García, C., Sagredo, O., and García-Arencíbia, M. (2014). “Neurodegenerative disorders other than multiple sclerosis,” in Handbook of Cannabis, ed R. Pertwee (Oxford: Oxford University Press), 505–525. doi: 10.1093/acprof:oso/9780199662685.003.0027
Fernández-Ruiz, J., Romero, J., and Ramos, J. A. (2015). Endocannabinoids and neurodegenerative disorders: Parkinson's Disease, Huntington's Chorea, Alzheimer's Disease, and Others. Handb. Exp. Pharmacol. 231, 233–259. doi: 10.1007/978-3-319-20825-1_8
Fernández-Ruiz, J., Romero, J., Velasco, G., Tolón, R. M., Ramos, J. A., and Guzmán, M. (2007). Cannabinoid CB2 receptor: a new target for controlling neural cell survival? Trends Pharmacol. Sci. 28, 39–45. doi: 10.1016/j.tips.2006.11.0 01
Ferré, S., Baler, R., Bouvier, M., Caron, M. G., Devi, L. A., Durroux, T., et al. (2009). Building a new conceptual framework for receptor heteromers. Nat. Chem. Biol. 5, 131–134. doi: 10.1038/nchembio0309-131
Franco, R., and Fernández-Suárez, D. (2015). Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 131, 65–86. doi: 10.1016/j.pneurobio.2015.05.003
Franco, R., Martínez-Pinilla, E., Lanciego, J. L., and Navarro, G. (2016). Basic pharmacological and structural evidence for class a g-protein-coupled receptor heteromerization. Front. Pharmacol. 7:76. doi: 10.3389/fphar.2016.00076
Fujii, M., Sherchan, P., Krafft, P. R., Rolland, W. B., and Soejima, Y. Z. J. (2014). Cannabinoid type 2 receptor stimulation attenuates brain edema by reducing cerebral leukocyte infiltration following subarachnoid hemorrhage in rats. J. Neurol. Sci. 342, 101–106. doi: 10.1016/j.jns.2014.04.034
Fulton, B. S., Knapp, B. L., Bidlack, J. M., and Neumeyer, J. L. (2010). Effect of linker substitution on the binding of butorphan univalent and bivalent ligands to opioid receptors. Bioorganic Med. Chem. Lett. 20, 1507–1509. doi: 10.1016/j.bmcl.2010.01.101
García, C., Palomo-Garo, C., Gómez-Gálvez, Y., and Fernández-Ruiz, J. (2016). Cannabinoid-dopamine interactions in the physiology and physiopathology of the basal ganglia. Br. J. Pharmacol. 173, 2069–2079. doi: 10.1111/bph.13215
García-Cabrerizo, R., and García-Fuster, M. J. (2016). Opposite regulation of cannabinoid CB1 and CB2 receptors in the prefrontal cortex of rats treated with cocaine during adolescence. Neurosci. Lett. 615, 60–65. doi: 10.1016/j.neulet.2016.01.018
García-Gutiérrez, M. S., and Manzanares, J. (2011). Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 25, 111–120. doi: 10.1177/0269881110379507
García-Gutiérrez, M. S., Pérez-Ortiz, J. M., Gutiérrez-Adán, A., and Manzanares, J. (2010). Depression-resistant endophenotype in mice overexpressing cannabinoid CB(2) receptors. Br. J. Pharmacol. 160, 1773–1784. doi: 10.1111/j.1476-5381.2010.00819.x
Glass, M., Govindpani, K., Furkert, D. P., Hurst, D. P., Reggio, P. H., and Flanagan, J. U. (2016). One for the Price of two…are bivalent ligands targeting cannabinoid receptor dimers capable of simultaneously binding to both receptors? Trends Pharmacol. Sci. 37, 353–363. doi: 10.1016/j.tips.2016.01.010
Gogoi, S., Biswas, S., Modi, G., Antonio, T., Reith, M. E. A., and Dutta, A. K. (2012). Novel bivalent ligands for D2/D3 dopamine receptors: significant cooperative gain in D2 affinity and potency. ACS Med. Chem. Lett. 3, 991–996. doi: 10.1021/ml3002117
Gómez-Cañas, M., Morales, P., García-Toscano, L., Navarrete, C., Muñoz, E., Jagerovic, N., et al. (2016). Biological characterization of PM226, a chromenoisoxazole, as a selective CB2 receptor agonist with neuroprotective profile. Pharmacol. Res. 110, 205–215. doi: 10.1016/j.phrs.2016.03.021
Gómez-Gálvez, Y., Palomo-Garo, C., Fernández-Ruiz, J., and García, C. (2016). Potential of the cannabinoid CB2 receptor as a pharmacological target against inflammation in Parkinson's disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 64, 200–208. doi: 10.1016/j.pnpbp.2015.03.017
Gomez, O., Arevalo-Martin, A., Garcia-Ovejero, D., Ortega-Gutierrez, S., Cisneros, J. A., Almazan, G., et al. (2010). The constitutive production of the endocannabinoid 2-arachidonoylglycerol participates in oligodendrocyte differentiation. Glia 58, 1913–1927. doi: 10.1002/glia.21061
Gomez, O., Sanchez-Rodriguez, A., Le, M., Sanchez-Caro, C., Molina-Holgado, F., and Molina-Holgado, E. (2011). Cannabinoid receptor agonists modulate oligodendrocyte differentiation by activating PI3K/Akt and the mammalian target of rapamycin (mTOR) pathways. Br. J. Pharmacol. 163, 1520–1532. doi: 10.1111/j.1476-5381.2011.01414.x
Goncalves, M. B., Suetterlin, P., Yip, P., Molina-Holgado, F., Walker, D. J., Oudin, M. J., et al. (2008). A diacylglycerol lipase- CB2 cannabinoid pathway regulates adult subventricular zone neurogenesis in an age-dependent manner. Mol. Cell. Neurosci. 38, 526–536. doi: 10.1016/j.mcn.2008.05.001
Gong, J.-P., Onaivi, E. S., Ishiguro, H., Liu, Q.-R., Tagliaferro, P. A., Brusco, A., et al. (2006). Cannabinoid CB2 receptors: immunohistochemical localization in rat brain. Brain Res. 1071, 10–23. doi: 10.1016/j.brainres.2005.11.035
Green, S. T., Nathwani, D., Goldberg, D. J., and Kennedy, D. H. (1989). Nabilone as effective therapy for intractable nausea and vomiting in AIDS. Br. J. Clin. Pharmacol. 28, 494–495. doi: 10.1111/j.1365-2125.1989.tb03533.x
Guo, J., Pavlopoulos, S., Tian, X., Lu, D., Nikas, S. P., Yang, D., et al. (2003). Conformational study of lipophilic ligands in phospholipid model membrane systems by solution NMR. J. Med. Chem. 46, 4838–4846. doi: 10.1021/jm020385r
Guo, K., Mou, X., Huang, J., Xiong, N., and Li, H. (2014). Trans-caryophyllene suppresses hypoxia-induced neuroinflammatory responses by inhibiting NF-κB activation in microglia. J. Mol. Neurosci. 54, 41–48. doi: 10.1007/s12031-014-0243-5
Han, S., Chen, J. J., and Chen, J.-Z. (2014). Latest progress in the identification of novel synthetic ligands for the cannabinoid CB2 receptor. Mini Rev. Med. Chem. 14, 426–443. doi: 10.2174/1389557514666140428105753
Han, S., Zhang, F.-F., Qian, H.-Y., Chen, L.-L., Pu, J.-B., Xie, X., et al. (2015). Development of quinoline-2,4(1 H,3 H)-diones as potent and selective ligands of the cannabinoid type 2 receptor. J. Med. Chem. 58, 5751–5769. doi: 10.1021/acs.jmedchem.5b00227
Hortala, L., Arnaud, J., Roux, P., Oustric, D., Boulu, L., Oury-Donat, F., et al. (2014). Synthesis and preliminary evaluation of a new fluorine-18 labelled triazine derivative for PET imaging of cannabinoid CB2 receptor. Bioorganic Med. Chem. Lett. 24, 283–287. doi: 10.1016/j.bmcl.2013.11.023
Hu, B., Doods, H., Treede, R.-D., and Ceci, A. (2009). Depression-like behaviour in rats with mononeuropathy is reduced by the CB2-selective agonist GW405833. Pain 143, 206–212. doi: 10.1016/j.pain.2009.02.018
Hu, S. S.-J., and Mackie, K. (2015). Distribution of the endocannabinoid system in the central nervous system. Handb. Exp. Pharmacol. 231, 59–93. doi: 10.1007/978-3-319-20825-1_3
Hurst, D. P., Grossfield, A., Lynch, D. L., Feller, S., Romo, T. D., Gawrisch, K., et al. (2010). A lipid pathway for ligand binding is necessary for a cannabinoid G protein-coupled receptor. J. Biol. Chem. 285, 17954–17964. doi: 10.1074/jbc.M109.041590
Ishiguro, H., Carpio, O., Horiuchi, Y., Shu, A., Higuchi, S., Schanz, N., et al. (2010a). A nonsynonymous polymorphism in cannabinoid CB2 receptor gene is associated with eating disorders in humans and food intake is modified in mice by its ligands. Synapse 64, 92–96. doi: 10.1002/syn.20714
Ishiguro, H., Horiuchi, Y., Ishikawa, M., Koga, M., Imai, K., Suzuki, Y., et al. (2010b). Brain cannabinoid CB2 receptor in schizophrenia. Biol. Psychiatry 67, 974–982. doi: 10.1016/j.biopsych.2009.09.024
Jhaveri, M. D., Sagar, D. R., Elmes, S. J. R., Kendall, D. A., and Chapman, V. (2007). Cannabinoid CB2 receptor-mediated anti-nociception in models of acute and chronic pain. Mol. Neurobiol. 36, 26–35. doi: 10.1007/s12035-007-8007-7
Jia, J., Peng, J., Li, Z., Wu, Y., Wu, Q., Tu, W., et al. (2016). Cannabinoid CB2 receptor mediates nicotine-induced anti-inflammation in n9 microglial cells exposed to β amyloid via protein kinase C. Mediat. Inflamm. 2016:4854378. doi: 10.1155/2016/4854378
Khajehali, E., Malone, D. T., Glass, M., Sexton, P. M., Christopoulos, A., and Leach, K. (2015). Biased agonism and biased allosteric modulation at the CB1 cannabinoid receptors. Mol. Pharmacol. 88, 368–379. doi: 10.1124/mol.115.099192
Köfalvi, A., Lemos, C., Martín-Moreno, A. M., Pinheiro, B. S., García-García, L., Pozo, M. A., et al. (2016). Stimulation of brain glucose uptake by cannabinoid CB2 receptors and its therapeutic potential in Alzheimer's disease. Neuropharmacology. doi: 10.1016/j.neuropharm.2016.03.015. [Epub ahead of print].
Lanciego, J. L., Barroso-Chinea, P., Rico, A. J., Conte-Perales, L., Callén, L., Roda, E., et al. (2011). Expression of the mRNA coding the cannabinoid receptor 2 in the pallidal complex of Macaca fascicularis. J. Psychopharmacol. 25, 97–104. doi: 10.1177/0269881110367732
Makriyannis, A., Tian, X., and Guo, J. (2005). How lipophilic cannabinergic ligands reach their receptor sites. Neurobiol. Dis. 77, 210–218. doi: 10.1016/j.prostaglandins.2004.01.010
Malek, N., Popiolek-Barczyk, K., Mika, J., Przewlocka, B., and Starowicz, K. (2015). Anandamide, acting via CB2 receptors, alleviates LPS-induced neuroinflammation in rat primary microglial cultures. Neural Plast. 2015:130639. doi: 10.1155/2015/130639
Mecha, M., Feliú, A., Iñigo, P. M., Mestre, L., and Guaza, C. (2013). Cannabidiol provides long-lasting protection against the deleterious effects of inflammation in a viral model of multiple sclerosis: a role for A2A receptors. Neurobiol. Dis. 59, 141–150. doi: 10.1016/j.nbd.2013.06.016
Micale, V., Mazzola, C., and Drago, F. (2007). Endocannabinoids and neurodegenerative diseases. Pharmacol. Res. 56, 382–392. doi: 10.1016/j.phrs.2007.09.008
Molina-Holgado, E., Vela, J. M., Arévalo-Martín, A., Almazán, G., Molina-Holgado, F., Borrell, J., et al. (2002). Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/Akt signaling. J. Neurosci. 22, 9742–9753.
Molina-Holgado, F., Pinteaux, E., Moore, J. D., Molina-Holgado, E., Guaza, C., Gibson, R. M., et al. (2003). Endogenous interleukin-1 receptor antagonist mediates anti-inflammatory and neuroprotective actions of cannabinoids in neurons and glia. J. Neurosci. 23, 6470–6474.
Morales, P., Goya, P., Jagerovic, N., and Hernandez-Folgado, L. (2016). Allosteric modulators of the CB 1 cannabinoid receptor : a structural update review. Cannabis Cannabinoid Res. 1, 22–30. doi: 10.1089/can.2015.0005
Mu, L., Bieri, D., Slavik, R., Drandarov, K., Muller, A., Cermak, S., et al. (2013). Radiolabeling and in vitro /in vivo evaluation of N-(1-adamantyl)-8-methoxy-4-oxo-1-phenyl-1,4-dihydroquinoline-3-carboxamide as a PET probe for imaging cannabinoid type 2 receptor. J. Neurochem. 126, 616–624. doi: 10.1111/jnc.12354
Navarrete, F., Pérez-Ortiz, J. M., and Manzanares, J. (2012). Cannabinoid CB2 receptor-mediated regulation of impulsive-like behaviour in DBA/2 mice. Br. J. Pharmacol. 165, 260–273. doi: 10.1111/j.1476-5381.2011.01542.x
Navarrete, F., Rodríguez-Arias, M., Martín-García, E., Navarro, D., García-Gutiérrez, M. S., Aguilar, M. A., et al. (2013). Role of CB2 cannabinoid receptors in the rewarding, reinforcing, and physical effects of nicotine. Neuropsychopharmacology 38, 2515–2524. doi: 10.1038/npp.2013.157
Nimczick, M., and Decker, M. (2015). New approaches in the design and development of cannabinoid receptor ligands: multifunctional and bivalent compounds. ChemMedChem 10, 773–786. doi: 10.1002/cmdc.201500041
Nimczick, M., Pemp, D., Darras, F. H., Chen, X., Heilmann, J., and Decker, M. (2014). Synthesis and biological evaluation of bivalent cannabinoid receptor ligands based on hCB2 R selective benzimidazoles reveal unexpected intrinsic properties. Bioorg. Med. Chem. 22, 3938–3946. doi: 10.1016/j.bmc.2014.06.008
Núñez, E., Benito, C., Pazos, M. R., Barbachano, A., Fajardo, O., González, S., et al. (2004). Cannabinoid CB2 receptors are expressed by perivascular microglial cells in the human brain: an immunohistochemical study. Synapse 53, 208–213. doi: 10.1002/syn.20050
Onaivi, E. S., Carpio, O., Ishiguro, H., Schanz, N., Uhl, G. R., and Benno, R. (2008a). Behavioral effects of CB2 cannabinoid receptor activation and its influence on food and alcohol consumption. Ann. N.Y. Acad. Sci. 1139, 426–433. doi: 10.1196/annals.1432.035
Onaivi, E. S., Chaudhuri, G., Abaci, A. S., Parker, M., Manier, D. H., Martin, P. R., et al. (1999). Expression of cannabinoid receptors and their gene transcripts in human blood cells. Prog. Neuropsychopharmacol. Biol. Psychiatry 23, 1063–1077.
Onaivi, E. S., Ishiguro, H., Gong, J.-P., Patel, S., Meozzi, P. A., Myers, L., et al. (2008b). Brain neuronal CB2 cannabinoid receptors in drug abuse and depression: from mice to human subjects. PLoS ONE 3:e1640. doi: 10.1371/journal.pone.0001640
Onaivi, E. S., Ishiguro, H., Gong, J.-P., Patel, S., Meozzi, P. A., Myers, L., et al. (2008c). Functional expression of brain neuronal CB2 cannabinoid receptors are involved in the effects of drugs of abuse and in depression. Ann. N.Y. Acad. Sci. 1139, 434–449. doi: 10.1196/annals.1432.036
Ortega-Alvaro, A., Aracil-Fernández, A., García-Gutiérrez, M. S., Navarrete, F., and Manzanares, J. (2011). Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 36, 1489–1504. doi: 10.1038/npp.2011.34
Ortega-Álvaro, A., Ternianov, A., Aracil-Fernández, A., Navarrete, F., García-Gutiérrez, M. S., and Manzanares, J. (2015). Role of cannabinoid CB2 receptor in the reinforcing actions of ethanol. Addict. Biol. 20, 43–55. doi: 10.1111/adb.12076
Palazuelos, J., Aguado, T., Egia, A., Mechoulam, R., Guzmán, M., and Galve-Roperh, I. (2006). Non-psychoactive CB2 cannabinoid agonists stimulate neural progenitor proliferation. FASEB J. 20, 2405–2407. doi: 10.1096/fj.06-6164fje
Palazuelos, J., Aguado, T., Pazos, M. R., Julien, B., Carrasco, C., Resel, E., et al. (2009). Microglial CB2 cannabinoid receptors are neuroprotective in Huntington's disease excitotoxicity. Brain 132, 3152–3164. doi: 10.1093/brain/awp239
Palazuelos, J., Ortega, Z., Díaz-Alonso, J., Guzmán, M., and Galve-Roperh, I. (2012). CB2 cannabinoid receptors promote neural progenitor cell proliferation via mTORC1 signaling. J. Biol. Chem. 287, 1198–1209. doi: 10.1074/jbc.M111.291294
Presley, C. S., Mustafa, S. M., Abidi, A. H., and Moore, B. M. (2015). Synthesis and biological evaluation of (3′,5′-dichloro-2,6-dihydroxy-biphenyl-4-yl)-aryl/alkyl-methanone selective CB2 inverse agonist. Bioorg. Med. Chem. 23, 5390–5401. doi: 10.1016/j.bmc.2015.07.057
Price, D. A., Martinez, A. A., Seillier, A., Koek, W., Acosta, Y., Fernandez, E., et al. (2009). WIN55,212-2, a cannabinoid receptor agonist, protects against nigrostriatal cell loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson's disease. Eur. J. Neurosci. 29, 2177–2186. doi: 10.1111/j.1460-9568.2009.06764.x
Reiner, A., Heldt, S. A., Presley, C. S., Guley, N. H., Elberger, A. J., Deng, Y., et al. (2015). Motor, visual and emotional deficits in mice after closed-head mild traumatic brain injury are alleviated by the novel CB2 inverse agonist SMM-189. Int. J. Mol. Sci. 16, 758–787. doi: 10.3390/ijms16010758
Ribeiro, R., Wen, J., Li, S., and Zhang, Y. (2013). Involvement of ERK1/2, cPLA2 and NF-κB in microglia suppression by cannabinoid receptor agonists and antagonists. Prostaglandins Other Lipid Mediat. 100–101, 1–14. doi: 10.1016/j.prostaglandins.2012.11.003
Rodríguez-Arias, M., Navarrete, F., Blanco-Gandia, M. C., Arenas, M. C., Aguilar, M. A., Bartoll-Andrés, A., et al. (2015). Role of CB2 receptors in social and aggressive behavior in male mice. Psychopharmacology 232, 3019–3031. doi: 10.1007/s00213-015-3939-5
Rodríguez-Cueto, C., Benito, C., Fernández-Ruiz, J., Romero, J., Hernández-Gálvez, M., and Gómez-Ruiz, M. (2014). Changes in CB(1) and CB(2) receptors in the post-mortem cerebellum of humans affected by spinocerebellar ataxias. Br. J. Pharmacol. 171, 1472–1489. doi: 10.1111/bph.12283
Sagredo, O., González, S., Aroyo, I., Pazos, M. R., Benito, C., Lastres-Becker, I., et al. (2009). Cannabinoid CB2 receptor agonists protect the striatum against malonate toxicity: relevance for Huntington's disease. Glia 57, 1154–1167. doi: 10.1002/glia.20838
Sam, A. H., Salem, V., and Ghatei, M. A. (2011). Rimonabant: from RIO to Ban. J. Obes. 2011:432607. doi: 10.1155/2011/432607
Savonenko, A. V., Melnikova, T., Wang, Y., Ravert, H., Gao, Y., Koppel, J., et al. (2015). Cannabinoid CB2 receptors in a mouse model of Aβ amyloidosis: immunohistochemical analysis and suitability as a PET biomarker of neuroinflammation. PLoS ONE 10:e0129618. doi: 10.1371/journal.pone.0129618
Sheng, W. S., Hu, S., Ni, H. T., Rock, R. B., and Peterson, P. K. (2009). WIN55,212-2 Inhibits production of CX3CL1 by human astrocytes: involvement of p38 MAP Kinase. J. Neuroimmune Pharmacol. 4, 244–248. doi: 10.1007/s11481-009-9147-5
Shoemaker, J. L., Seely, K. A., Reed, R. L., Crow, J. P., and Prather, P. L. (2007). The CB2 cannabinoid agonist AM-1241 prolongs survival in a transgenic mouse model of amyotrophic lateral sclerosis when initiated at symptom onset. J. Neurochem. 101, 87–98. doi: 10.1111/j.1471-4159.2006.04346.x
Sierra, S., Luquin, N., Rico, A. J., Gómez-Bautista, V., Roda, E., Dopeso-Reyes, I. G., et al. (2015). Detection of cannabinoid receptors CB1 and CB2 within basal ganglia output neurons in macaques: changes following experimental parkinsonism. Brain Struct. Funct. 220, 2721–2738. doi: 10.1007/s00429-014-0823-8
Skaper, S. D., Buriani, A., Dal Toso, R., Petrelli, L., Romanello, S., Facci, L., et al. (1996). The ALIAmide palmitoylethanolamide and cannabinoids, but not anandamide, are protective in a delayed postglutamate paradigm of excitotoxic death in cerebellar granule neurons. Proc. Natl. Acad. Sci. U.S.A. 93, 3984–3989. doi: 10.1073/pnas.93.9.3984
Slavik, R., Grether, U., Müller Herde, A., Gobbi, L., Fingerle, J., Ullmer, C., et al. (2015a). Discovery of a high affinity and selective pyridine analog as a potential positron emission tomography imaging agent for cannabinoid type 2 receptor. J. Med. Chem. 58, 4266–4277. doi: 10.1021/acs.jmedchem.5b00283
Slavik, R., Herde, A. M., Bieri, D., Weber, M., Schibli, R., Krämer, S. D., et al. (2015b). Synthesis, radiolabeling and evaluation of novel 4-oxo-quinoline derivatives as PET tracers for imaging cannabinoid type 2 receptor. Eur. J. Med. Chem. 92, 554–564. doi: 10.1016/j.ejmech.2015.01.028
Smith, S. R., Terminelli, C., and Denhardt, G. (2000). Effects of cannabinoid receptor agonist and antagonist ligands on production of inflammatory cytokines and anti-inflammatory interleukin-10 in endotoxemic mice. J. Pharmacol. Exp. Ther. 293, 136–50.
Stella, N. (2010). Cannabinoid and cannabinoid-like receptors in microglia, astrocytes, and astrocytomas. Glia 58, 1017–1030. doi: 10.1002/glia.20983
Tao, Q., McAllister, S. D., Andreassi, J., Nowell, K. W., Cabral, G. A., Hurst, D. P., et al. (1999). Role of a conserved lysine residue in the peripheral cannabinoid receptor (CB2): evidence for subtype specificity. Mol. Pharmacol. 55, 605–613.
Van Sickle, M. D., Duncan, M., Kingsley, P. J., Mouihate, A., Urbani, P., Mackie, K., et al. (2005). Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 310, 329–332. doi: 10.1126/science.1115740
Vendel, E., and de Lange, E. C. M. (2014). Functions of the CB1 and CB 2 receptors in neuroprotection at the level of the blood-brain barrier. Neuromolecular Med. 16, 620–642. doi: 10.1007/s12017-014-8314-x
Walter, L., Franklin, A., Witting, A., Wade, C., Xie, Y., Kunos, G., et al. (2003). Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J. Neurosci. 23, 1398–1405.
Yiangou, Y., Facer, P., Durrenberger, P., Chessell, I. P., Naylor, A., Bountra, C., et al. (2006). COX-2, CB2 and P2X7-immunoreactivities are increased in activated microglial cells/macrophages of multiple sclerosis and amyotrophic lateral sclerosis spinal cord. BMC Neurol. 6:12. doi: 10.1186/1471-2377-6-12
Zarruk, J. G., Fernandez-Lopez, D., Garcia-Yebenes, I., Garcia-Gutierrez, M. S., Vivancos, J., Nombela, F., et al. (2012). Cannabinoid type 2 receptor activation downregulates stroke-induced classic and alternative brain macrophage/microglial activation concomitant to neuroprotection. Stroke 49, 211–219. doi: 10.1161/STROKEAHA.111.631044
Keywords: heteromer, microglia, astroglia, M0/M1/M2 phenotype, neuroprotection, neurorestoration, GPCR, amyotrophic lateral sclerosis
Citation: Navarro G, Morales P, Rodríguez-Cueto C, Fernández-Ruiz J, Jagerovic N and Franco R (2016) Targeting Cannabinoid CB2 Receptors in the Central Nervous System. Medicinal Chemistry Approaches with Focus on Neurodegenerative Disorders. Front. Neurosci. 10:406. doi: 10.3389/fnins.2016.00406
Received: 25 May 2016; Accepted: 22 August 2016;
Published: 13 September 2016.
Edited by:
Marialessandra Contino, University of Bari Aldo Moro, ItalyReviewed by:
Aurel Popa-Wagner, University of Rostock, GermanyMichael Decker, University of Würzburg, Germany
Claudia Mugnaini, University of Siena, Italy
Copyright © 2016 Navarro, Morales, Rodríguez-Cueto, Fernández-Ruiz, Jagerovic and Franco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafael Franco, rfranco@ub.edu; rfranco123@gmail.com