 Margaux Deynoux
Margaux Deynoux Nicola Sunter
Nicola Sunter Olivier Hérault1,2
Olivier Hérault1,2 Frédéric Mazurier
Frédéric Mazurier- 1Génétique, Immunothérapie, Chimie et Cancer (GICC) UMR 7292, CNRS, UFR de Médecine, Université François-Rabelais de Tours, Tours, France
- 2Service d’Hématologie Biologique, Centre Hospitalier Régional Universitaire de Tours, Tours, France
Despite huge improvements in the treatment of leukemia, the percentage of patients suffering relapse still remains significant. Relapse most often results from a small number of leukemic stem cells (LSCs) within the bone marrow, which are able to self-renew, and therefore reestablish the full tumor. The marrow microenvironment contributes considerably in supporting the protection and development of leukemic cells. LSCs share specific niches with normal hematopoietic stem cells with the niche itself being composed of a variety of cell types, including mesenchymal stem/stromal cells, bone cells, immune cells, neuronal cells, and vascular cells. A hallmark of the hematopoietic niche is low oxygen partial pressure, indeed this hypoxia is necessary for the long-term maintenance of hematopoietic stem/progenitor cells. Hypoxia is a strong signal, principally maintained by members of the hypoxia-inducible factor (HIF) family. In solid tumors, it has been well established that hypoxia triggers intrinsic metabolic changes and microenvironmental modifications, such as the stimulation of angiogenesis, through activation of HIFs. As leukemia is not considered a “solid” tumor, the role of oxygen in the disease was presumed to be inconsequential and remained long overlooked. This view has now been revised since hypoxia has been shown to influence leukemic cell proliferation, differentiation, and resistance to chemotherapy. However, the role of HIF proteins remains controversial with HIFs being considered as either oncogenes or tumor suppressor genes, depending on the study and model. The purpose of this review is to highlight our knowledge of hypoxia and HIFs in leukemic development and therapeutic resistance and to discuss the recent hypoxia-based strategies proposed to eradicate leukemias.
Leukemias and Hypoxia
Leukemia is characterized by uncontrolled proliferation of hematopoietic cells within bone marrow (BM). Lymphoid leukemias can be distinguished from myeloid according to the abnormal cell lineage, and acute from chronic leukemias according to the maturity of the blood cells involved and progression rate. Acute leukemias are characterized by rapid proliferation of immature hematopoietic cells, termed blasts, which fail to differentiate into mature cells. Their accumulation in BM also prevents growth and differentiation of normal hematopoietic cells. The clinical evolution is fast (1). In contrast, in chronic leukemia, the growth advantage of neoplastic cells leads to the generation of a more mature cell population that outcompetes normal hematopoiesis, and clinical evolution is longer (several months to years) (2). To date, although the majority of pediatric acute lymphoid leukemia (ALL) and chronic myeloid leukemia (CML) cases (3) are cured or well controlled under treatment, chronic lymphoid leukemia (CLL) and, even more, acute myeloid leukemia (AML) have a high risk of relapse, despite therapeutic progressions (4). While treatments often target cycling cells, the idea that a small population of quiescent leukemic cells survive and trigger relapse regardless of treatment has emerged (5). In the early 90s, the team of John E. Dick established a hierarchy in leukemic cell populations which, by analogy with that of normal hematopoietic cells, led to the introduction of the concept of cancer stem cells for all cancers [reviewied in Ref. (5, 6)]. Their work identified a subpopulation of leukemic cells able to initiate leukemic growth after transplantation into immune-deficient mice. These stem-like cells, named leukemia-initiating cells (LICs) or leukemic stem cells (LSCs) (7, 8), arise from hematopoietic stem/progenitor cells (HSCs) that reside in the most hypoxic tissue areas within the normal HSC niche (9, 10). The oxygen partial pressure (ppO2) in tissues is much lower compared to that in the atmosphere (160 mmHg corresponding to around 21% oxygen). In particular, in BM an oxygen gradient exists ranging from <6% oxygen, close to the vessels, to anoxia in the most distant regions from blood vessels (11–15). However, O2 level differs according to the nature of the hematopoietic niche; the sinusoidal niche is around 10 mmHg (equivalent to 1.3% O2) (16).
In solid cancers, it is well established that uncontrolled proliferation leads to profound hypoxia, associated with tumor development, metabolic changes, metastatic propagation, immune response modulation, and increased mortality (17, 18). Consequently, it could be assumed that intense blast cell proliferation would eventually decrease the oxygen availability by high consumption. This assumption is particularly difficult to validate by direct measurement in human BM. However, Fiegl et al. (19) demonstrated in total BM aspirates from AML patients that oxygen percentage was highly comparable to the normal counterpart. Using a rat model of an acute AML subtype, the promyelocytic leukemia, Jensen et al. (20) noted an increasing level of hypoxia during disease progression, comparable to that observed with solid tumours. In this model, both normal and leukemic cells stained with 2-nitroimidazole (hypoxic marker) underwent decreased proliferation. In agreement with this observation, the hypoxic culture of normal hematopoietic (21–23) and CML cells (24–27) led to decreased proliferation. Nevertheless, a small fraction of leukemic cells remained insensitive to hypoxia-induced proliferation arrest (26), probably triggering tumor growth (20). Moreover, it has been established that mild hypoxia such as ≤3% O2 sustains both primary CML (24) and AML cell (28) maintenance longer than normoxia.
Low oxygen might also affect hematopoietic cells through the modulation of the stromal cells. Indeed, hypoxia has been shown to impact on survival, proliferation capability, and differentiation as well as metabolism of mesenchymal stem/stromal cells (MSCs) (29–31). Hypoxia triggers secretion by MSCs of numerous factors, including SDF-1, VEGF, and IL-6, known to promote HSC maintenance. Interestingly, even in normoxia, HSCs present a hypoxic profile when seeded on MSCs, suggesting appearance of “microhypoxic” regions (32, 33). Moreover, MSCs and hypoxic culture synergize to sustain in vitro normal stem cells (23) and primary AML cells (28). Finally, the poorly oxygenized niche enhances resistance to treatments (28, 34, 35), thus protecting from various stresses, such as DNA damage, cell death stimuli, or oxidative stress signals (36–38).
Hypoxia-Inducible Factors in Leukemias
The master regulators mediating cell responses to hypoxia are the hypoxia-inducible factors (HIFs). These heterodimer complexes are composed of one of three oxygen-regulated HIF-alpha subunits (HIF-1alpha, HIF-2alpha, and HIF-3alpha) and the constitutively expressed HIF-beta subunit [HIF-1-beta, also known as aryl hydrocarbon receptor nuclear translocator (ARNT)] (39–41). The HIF1A gene is ubiquitously expressed (42). HIF2A, also termed endothelial Per-ARNT-Sim (PAS) protein 1 (EPAS1), is expressed in a more tissue specific manner, particularly in blood vessels (39, 40, 43). Little is currently known about expression and function of HIF3A, but at least 10 splice variants have been described to date (44, 45). HIF-1alpha and HIF-2alpha proteins share similar structural domains such as an N-terminal basic helix-loop-helix (bHLH) domain involved in DNA binding, the two PAS domains allowing dimerization, an oxygen-dependent degradation domain (ODDD) plus N- and C-terminal transactivation domains (NTAD and CTAD). Although HIF-3alpha also exhibits high similarity in bHLH and PAS domains, the lack of the CTAD precludes binding to p300 coactivator (45, 46). Under atmospheric conditions, HIF-alpha subunits are differentially hydroxylated by prolyl hydroxylase domain 1–3 (PHDs) on two proline residues in the ODDD, with oxygen and α-ketoglutarate as substrates. The hydroxylated motif allows binding to von Hippel–Lindau (VHL) tumor suppressor, which leads to HIF-alpha ubiquitination and consequent degradation by 26S proteasome (39–41, 47). In parallel, the hydroxylation of HIF-1alpha by factor inhibiting HIF-1 (FIH1) triggers inhibition of p300/CBP coactivator recruitment (48). PHD activity falls with decreasing oxygen levels, thereby triggering HIF-alpha stabilization and nuclear translocation where it heterodimerizes with HIF-1beta. HIF complexes bind to specific HIF-response elements consisting of specific RCGTG sequences within target gene promoters. Although HIF-1 and HIF-2 share common targets, additional genomic regions and cofactor-binding specifically drive the transcriptional initiation of genes involved in many pathways, such as angiogenesis, differentiation, stem cells maintenance, apoptosis, and invasion (35, 39, 40).
HIF-1alpha mainly participates in the initial response to acute hypoxia, whereas HIF-2alpha responds to chronic exposure (47, 49). Additionally, even in prolonged hypoxia HIF-1alpha undergoes feedback control, whereas HIF-2alpha is stabilized. The multiple HIF-3alpha splice variants appear essentially to regulate HIF-1alpha and HIF-2alpha activity by sequestrating HIF-1beta or by acting as dominant negative regulators (40, 44, 45, 50). HIFs, moreover, can be regulated by oxygen-independent mechanisms. Factors involved in hematopoiesis such as MEIS1 (51), TPO (52), and SCF (53, 54) positively regulate expression of HIFs. Conversely, factors implicated in metabolic changes like the SIRT1 (55, 56) or SDH (57) inhibit the expression of HIFs. Furthermore, downregulation of HIFs may be achieved by tumor suppressor genes like p53 (58, 59) or GSK3 (60) and the upregulation by oncogenes such as PI3K/AKT (60, 61) or mTORC1 (62). Genetic abnormalities encountered in leukemia such as the IDH mutation decrease expression of HIFs by stimulating PHD activity (63) or conversely, FLT3-ITD stimulates translation of HIFs via the PI3K/AKT pathway (40).
Elevated expression of HIFs is considered to be a marker of poor prognosis in solid cancers (64–77). Overall, increased expression of HIFs is correlated with tumor growth and resistance to therapies, which leads to disease relapse (37). The subject is somewhat more complex and controversial in leukemia. Overexpression of HIF-1alpha in leukemia has been suggested as a marker of poor prognosis and chemotherapy outcomes (78–81). Elevated levels of HIF-1alpha are reported, in the majority of studies, in AML (79, 82–84), acute promyelocytic leukemia (APL) (85), ALL (82), and CML (86, 87). Disease severity and survival have been shown to be influenced by HIF-1alpha levels, in AML and myelodysplastic syndromes (84, 88, 89); the protein expression of HIF-2alpha, usually absent in normal cells, has been observed in both AML and ALL, but has not been correlated with outcome (82, 90, 91). Thus, leukemic subtype, disease stage or the molecular abnormality involved might explain the variability. To explore potential functions of HIFs in leukemogenesis, various mouse models have been proposed (Table 1). Several studies have shown that inhibition of HIF-1alpha, either by RNAi targeting or by small molecular inhibition, resulted in a failure of primary cells to form in vitro colonies and decreased tumor growth and leukemic progression. In vivo, dramatic decrease and potential eradication of primary AML cell xenografts have been shown and a complete absence of leukemic induction in secondary transplantation has been reported in cells in which HIF-1alpha was inhibited (79, 84, 85, 92). This has also been confirmed in ALL (93, 94) and CML (83, 87). Similarly, knockdown of HIF-2alpha with shRNA triggers leukemic inhibition (82, 85, 90), which is demonstrated in vitro by limited cellular proliferation as well as in vivo by absence or poor engraftment.
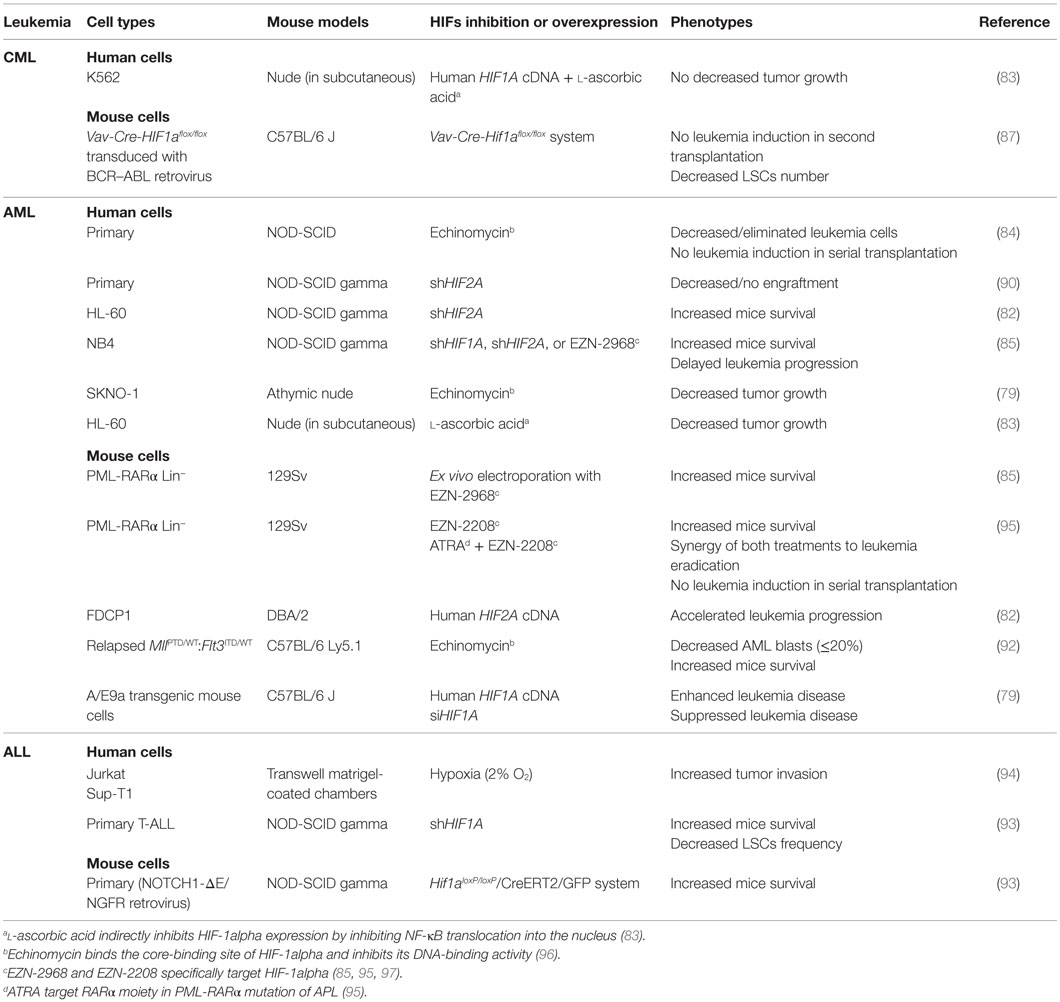
Table 1. Models used to characterize HIF1A and HIF2A as oncogenes in leukemias.
Hypoxia via HIFs may promote disease maintenance and progression through different mechanisms including energy metabolism (98–100), cycle and quiescence (101, 102), and immune function (103) that are important in normal physiology and deregulated in cancer (47, 104). On one hand, HIF-1alpha and HIF-2alpha influence signaling pathways relevant to leukemia maintenance and propagation. HIF-1alpha activates the Notch1 pathway, which leads to leukemia invasion (94), and promotes the Wnt pathway, consequently preserving LSCs (93). On the other hand, HIF-1alpha acts as an inhibitor of the expression of tumor suppressor genes, such as p15, p16, p19, and p53 (79, 87). Indeed, HIF-1alpha-transactivated DNMT3a methylates DNA, which inhibits tumor suppressors and leads to tumor growth (79). In AML, DNMT3a plays a crucial role since more than 20% of patients exhibit DNMT3A mutation (105), conferring a global hypomethylation of DNA and predisposition to developing hematological diseases (106). In contrast, in T-ALL this mutation confers hypermethylation, so the contribution of hypomethylation and hypermethylation to disease development remains to be elucidated (106). Interestingly, taken from non-hematopoietic tissue and cancers, studies have explored the role of hypoxia in epigenetic modifications, through HIF-1alpha stabilization, such as DNA methylation, histone modifications, and non-coding RNA expression (107). Promoter methylation is modified by hypoxia and regulates neural progenitor cell fate (108). The histone demethylase JMJD1A and 1B are targets of HIFs in normal and cancer cells (109–111). Finally in HIF-2alpha-deficient cells, transcriptomic approaches have identified deregulated genes involved in energetic and oxidative metabolisms, plus endoplasmic reticulum (ER) stress, indicating that HIF-2alpha protects AML cells from apoptosis induced by ER stress (90).
One consequences of the expression of HIFs is the promotion of quiescence, which favors chemoresistance. Hypoxia-induced HIF-1alpha promotes entry into G0/G1 and decreases S phase in AML cells through, in part, upregulation of p27 (112). Quiescence enhances chemoresistance of leukemic cells to cytosine arabinoside (Ara-C) (112, 113) and adriamycin (ADR) (88), since these agents target cycling cells. Coculture of primary AML cells with stromal cells in hypoxia (3% O2) confers resistance to Ara-C through stabilization of HIFs and induction of quiescence (28). Antiapoptotic signaling is observed through increased XIAP level, an apoptosis-inhibitory protein, and the activation of the PI3K/AKT pro-survival pathway (112). HIF-1alpha activation by a PHD inhibitor, cobalt chloride (CoCl2), protects HL-60 leukemic cells against arsenic trioxide (ATO) by inhibiting BAX and Caspase 3 and 9 and promoting HSP70 protein and p38/ERK pro-survival factors (114). In T-ALL, through Notch1 activation, HIF-1alpha induces BCL2 and BCL-XL upregulation and the downregulation of Caspase 3 and 9 activities, which decreased dexamethasone-induced apoptosis in leukemic cells (94).
Conversely, low oxygen and hypoxia-mimicking agents such as CoCl2 or desferrioxamine induce AML cell differentiation through HIF-1alpha accumulation (115, 116). In fact, HIF-1alpha mediates differentiation by binding to C/EBPα and promoting its transcriptional activity (115–117) as well as that of RUNX1 and PU.1 (118, 119). Additionally, C/EBPα/HIF-1alpha induces AML differentiation through c-MYC inhibition and further suppression of miR17 and miR20a expression. The knockdown of p21 and STAT3, two inhibitory targets of miR17 and miR20A, reverses HIF-1alpha-induced AML differentiation (120). In renal cell carcinoma, HIF-1alpha inhibits c-MYC/MAX association, which decreases c-MYC promoter binding and thus blocks cells in G1 (121). Conversely, HIF-2alpha triggers cell cycle progression and proliferation by enhancing the formation of c-MYC/MAX and its activity. Since HIF-2alpha and HIF-1alpha have dual effects on cell cycle progression according to cell types, more investigations are needed on their antagonistic effects in leukemias. Nguyen-Khac et al. (122) discovered a translocation involving TEL and ARNT in an AML patient exerting a dominant/negative activity on HIF-1alpha. The fusion protein blocks leukemic differentiation, thus conferring a tumor suppressor function for HIF-1alpha. In line with this, data have previously shown that intermittent hypoxia slows down leukemic development in mice (123). However, in vivo hypoxia may have unrelated consequences on leukemic cell physiology. More recently, Velasco-Hernandez et al. (124) using different AML models found reduced survival of mice transplanted with HIF-1alpha KO cells (Table 2). These observations were confirmed in myeloproliferative neoplasia through a FLT3ITD-induced mouse model (125). Overall, Hifa KO enhanced disease progression and severity, making it a tumor suppressor gene. However, authors show that HIF-1alpha deletion may promote compensatory effects via overexpression of HIF-2alpha, which may eventually mask the role of HIF-1alpha. This elevation was already seen in HIF-1alpha-deficient cells (90). The Hif2a KO in MLL-AF9-driven and Meis1/HoxA9-induced murine AML enhances LSCs development but, once leukemias are established, HIF-2alpha has no impact on their maintenance and propagation. Furthermore, double inhibition of HIF-1alpha and HIF-2alpha demonstrated that HIFs synergize to inhibit AML development, without any role in leukemic propagation (126). Transcriptomic analysis reveals that HIF-1alpha and HIF-2alpha promote a set of genes that fosters survival and proliferation of leukemic cells.
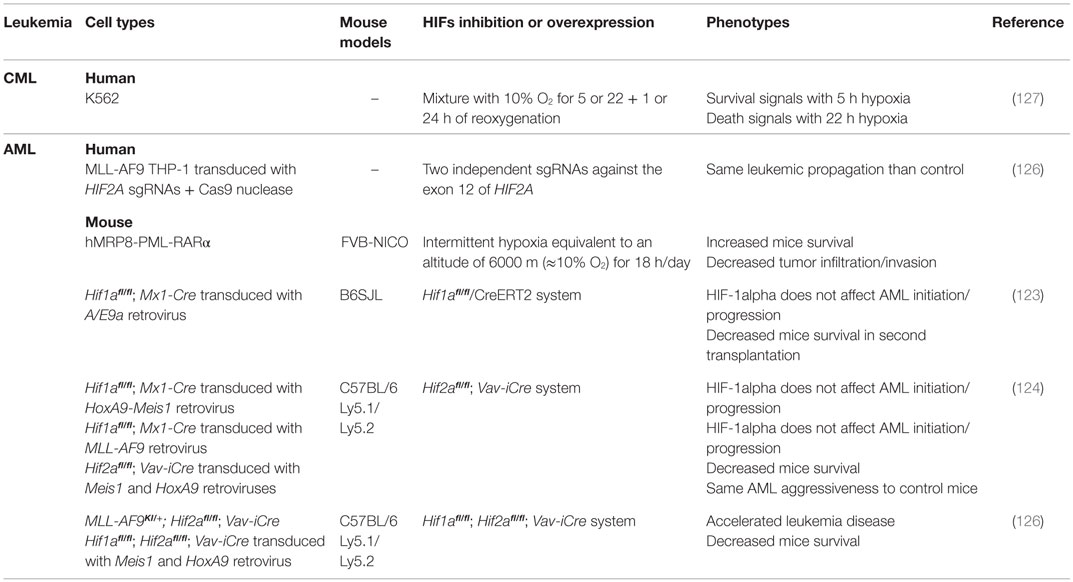
Table 2. Models used to characterize HIF1A and HIF2A as tumor suppressor genes in leukemias.
Off-target effects of shRNAs and poor specificity of drugs that inhibit HIFs compared to KO might explain this controversy between studies. However, KO may also induce slow cellular adaptation with compensatory effects. Differences may also arise from the model used, mouse versus human, and the different protumoral gene constructions used to generate the leukemia. It will be pertinent to assess the overexpression of HIFs in AML models, and thus observe whether increased HIF delays disease development and, even, eradicate leukemia. HIFs may also differently impact on LSCs and more mature blasts cells, conferring pro-survival effect on LSCs and differentiation on blasts. In favor of this hypothesis, intermittent hypoxia increases survival of APL mice (123). Similarly, in CML, HIF-1alpha induction following short (5 h) hypoxia exposure delivered a survival signal to cells, whereas it promoted cell death within a longer period (22 h) (127). In the ALL model, 24 h-exposure to hypoxia conferred chemoresistance in contrast to longer exposure (48–72 h) (128). These data suggest that duration of hypoxia incubation may promote or inhibit leukemia progression and maintenance, thus explaining the oncogenic or tumor suppressor activity. The link between HIFs and tumor suppressor activity has been previously demonstrated in other cancers (71, 129–131). Taken together, these data suggest that not only the level but also the duration of activity dictates HIF action and hence cellular response in leukemia.
Therapeutic Strategies: Targeting HIFs or Not?
In light of the results summarized above, it could be difficult to affirm that downregulation of HIFs could be a therapeutic approach. Nevertheless, chemical inhibitors have been tested and the proof-of-concept was first illustrated using echinomycin, which blocks HIF-1alpha-binding activity. This drug preferentially targets AML LSCs through apoptosis, decreasing leukemia burden, prolonging mouse survival, and abrogating disease development in secondary transplantation (84, 92). Echinomycin does not impact on self-renewal and differentiation of normal HSCs, which makes it an ideal molecule to treat leukemia (92). l-ascorbic acid has also been shown to inhibit expression of HIF-1alpha in CML cells and consequently reduces tumor growth. This effect is specific to HIF-1alpha since its overexpression in l-ascorbic acid-treated mice antagonizes leukemic growth inhibition (83). In APL, EZN-2968 and EZN-2208 confer antileukemic activity and prolong mouse survival; in combination with all-trans retinoic acid (ATRA), leukemia eradication was observed, along with survival of mice; fortunately, both compounds are non-toxic to normal HSCs (85, 95). Overall, these data offer new therapeutic options, targeting HIF in leukemia with no impact on normal hematopoiesis. Another approach will be the combination of HIF inhibitors with treatments capable of determining the departure of LSCs from quiescence, and then with treatments that target cycling cells, such as Ara-C.
An alternative strategy will consist of taking advantage of hypoxia to activate drugs and thus to target LSCs in the niche. TH-302 is a hypoxia-activated prodrug, which exhibits a specific cytotoxicity in hypoxia (132, 133). In primary AML, TH-302 hampers tumor growth through multiple mechanisms (cycle arrest, DNA cross-linking, DNA damage). In mouse models, it decreases leukemia burden and prolongs survival (133). PR-104 quickly undergoes alcohol hydrolysis and induces DNA cross-linking in hypoxic cells, impairs ALL progression, decreases tissue infiltration, and prolongs mice survival (134, 135). In a phase I/II study, PR-104 reduced the number of AML and ALL cells in refractory and relapsed patients (136). Despite some side effects, including myelosuppression, febrile neutropenia, and infections, collectively, these results propose innovative therapies for leukemia based on hypoxia-activated prodrugs.
Conclusion
Overall, these data argue that hypoxia and HIF-mediated signaling play a crucial role in leukemia and leukemogenic processes. However, they conflict in determining whether HIFs act as oncogenes or tumor suppressors, certainly because of the different leukemic models, study design, oxygen level, and hypoxia duration. However, therapies targeting hypoxia and HIFs have proven their efficacy in treating mouse models and may benefit leukemic patients.
Author Contributions
All authors designed, wrote, and revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
The authors acknowledge La Ligue contre le Cancer for funding NS, the Ministry of Research for Funding MD, and the Région Center. La Ligue contre le Cancer, CANCEN, and Les Sapins de l’Espoir associations supported this work.
References
1. Brown CMS, Larsen SR, Iland HJ, Joshua DE, Gibson J. Leukaemias into the 21st century: part 1: the acute leukaemias. Intern Med J (2012) 42(11):1179–86. doi: 10.1111/j.1445-5994.2012.02938.x
2. Gibson J, Iland HJ, Larsen SR, Brown CMS, Joshua DE. Leukaemias into the 21st century – part 2: the chronic leukaemias. Intern Med J (2013) 43(5):484–94. doi:10.1111/imj.12135
3. Freireich EJ, Wiernik PH, Steensma DP. The leukemias: a half-century of discovery. J Clin Oncol (2014) 32(31):3463–9. doi:10.1200/JCO.2014.57.1034
4. Döhner H, Weisdorf DJ, Bloomfield CD. Acute myeloid leukemia. N Engl J Med (2015) 373(12):1136–52. doi:10.1056/NEJMra1406184
5. Zagozdzon R, Golab J. Cancer stem cells in haematological malignancies. Contemp Oncol (2015) 19(1A):A1–6. doi:10.5114/wo.2014.47127
6. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Stem Cells (2001) 414(6859):105–11. doi:10.1007/978-1-60327-933-8
7. Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med (1997) 3(7):730–7. doi:10.1038/nm0797-730
8. Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature (1994) 367(6464):645–8. doi:10.1038/367645a0
9. Schepers K, Campbell TB, Passegué E. Normal and leukemic stem cell niches: insights and therapeutic opportunities. Cell Stem Cell (2015) 16(3):254–67. doi:10.1016/j.stem.2015.02.014
10. Tabe Y, Konopleva M. Advances in understanding the leukaemia microenvironment. Br J Haematol (2014) 164(6):767–78. doi:10.1111/bjh.12725
11. Chow DC, Wenning LA, Miller WM, Papoutsakis ET. Modeling pO(2) distributions in the bone marrow hematopoietic compartment. I. Krogh’s model. Biophys J (2001) 81(2):675–84. doi:10.1016/S0006-3495(01)75733-5
12. Chow DC, Wenning LA, Miller WM, Papoutsakis ET. Modeling pO(2) distributions in the bone marrow hematopoietic compartment. II. Modified Kroghian models. Biophys J (2001) 81(2):685–96. doi:10.1016/S0006-3495(01)75733-5
13. Harrison JS, Rameshwar P, Chang V, Bandari P. Oxygen saturation in the bone marow of healthy volunteers. Blood (2012) 99(1):394. doi:10.1182/blood.V98.1.250
14. Nombela-Arrieta C, Pivarnik G, Winkel B, Canty KJ, Mahoney JE, Park S-Y, et al. Quantitative imaging of hematopoietic stem and progenitor cell localization and hypoxic status in the bone marrow microenvironment. Nat Cell Biol (2013) 15(5):533–43. doi:10.1038/ncb2730
15. Parmar K, Mauch P, Vergilio J-A, Sackstein R, Down JD. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc Natl Acad Sci U S A (2007) 104(13):5431–6. doi:10.1073/pnas.0701152104
16. Spencer JA, Ferraro F, Roussakis E, Klein A, Wu J, Runnels JM, et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature (2014) 508(7495):269–73. doi:10.1038/nature13034
17. Quail D, Joyce J. Microenvironmental regulation of tumor progression and metastasis. Nat Med (2013) 19(11):1423–37. doi:10.1038/nm.3394
18. Semenza GL. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J Clin Invest (2013) 123(9):3664–71. doi:10.1172/JCI67230.3664
19. Fiegl M, Samudio I, Clise-Dwyer K, Burks JK, Mnjoyan Z, Andreeff M. CXCR4 expression and biologic activity in acute myeloid leukemia are dependent on oxygen partial pressure. Blood (2009) 113(7):1504–12. doi:10.1182/blood-2008-06-161539
20. Jensen PO, Mortensen BT, Hodgkiss RJ, Iversen APO, Christensen IJ, Helledie N, et al. Increased cellular hypoxia and reduced proliferation of both normal and leukaemic cells during progression of acute myeloid leukaemia in rats. Cell Prolif (2000) 33(6):381–95. doi:10.1046/j.1365-2184.2000.00183.x
21. Eliasson P, Rehn M, Hammar P, Larsson P, Sirenko O, Flippin LA, et al. Hypoxia mediates low cell-cycle activity and increases the proportion of long-term-reconstituting hematopoietic stem cells during in vitro culture. Exp Hematol (2010) 38(4):301–310.e2. doi:10.1016/j.exphem.2010.01.005
22. Ivanovic Z, Hermitte F, Brunet de La Grange P, Dazey B, Belloc F, Lacombe F, et al. Simultaneous maintenance of human cord blood SCID-repopulating cells and expansion of committed progenitors at low O2 concentration (3%). Stem Cells (2004) 22:716–24. doi:10.1634/stemcells.2004-0094
23. Hammoud M, Vlaski M, Duchez P, Chevaleyre J, Lafarge X, Boiron J-M, et al. Combination of low O2 concentration and mesenchymal stromal cells during culture of cord blood CD34+ cells improves the maintenance and proliferative capacity of hematopoietic stem cells. J Cell Physiol (2012) 227(6):2750–8. doi:10.1002/jcp.23019
24. Desplat V, Faucher J-L, Mahon FX, Dello Sbarba P, Praloran V, Ivanovic Z. Hypoxia modifies proliferation and differentiation of CD34+ CML cells. Stem Cells (2002) 20:347–54. doi:10.1634/stemcells.20-4-347
25. Giuntoli S, Rovida E, Barbetti V, Cipolleschi MG, Olivotto M, Dello Sbarba P. Hypoxia suppresses BCR/Abl and selects imatinib-insensitive progenitors within clonal CML populations. Leukemia (2006) 20(7):1291–3. doi:10.1038/sj.leu.2404224
26. Giuntoli S, Rovida E, Gozzini A, Barbetti V, Cipolleschi MG, Olivotto M, et al. Severe hypoxia defines heterogeneity and selects highly immature progenitors within clonal erythroleukemia cells. Stem Cells (2007) 25(5):1119–25. doi:10.1634/stemcells.2006-0637
27. Giuntoli S, Tanturli M, Di Gesualdo F, Barbetti V, Rovida E, Dello Sbarba P. Glucose availability in hypoxia regulates the selection of chronic myeloid leukemia progenitor subsets with different resistance to imatinib-mesylate. Haematologica (2011) 96(2):204–12. doi:10.3324/haematol.2010.029082
28. Griessinger E, Anjos-Afonso F, Pizzitola I, Rouault-Pierre K, Vargaftig J, Taussig D, et al. A niche-like culture system allowing the maintenance of primary human acute myeloid leukemia-initiating cells: a new tool to decipher their chemoresistance and self-renewal mechanisms. Stem Cells Transl Med (2014) 3:520–9. doi:10.5966/sctm.2013-0166
29. Beegle J, Lakatos K, Kalomoiris S, Stewart H, Isseroff RR, Nolta JA, et al. Hypoxic preconditioning of mesenchymal stromal cells induces metabolic changes, enhances survival and promotes cell retention in vivo. Stem Cells (2015) 33:1818–28. doi:10.1002/stem.1976
30. Holzwarth C, Vaegler M, Gieseke F, Pfister SM, Handgretinger R, Kerst G, et al. Low physiologic oxygen tensions reduce proliferation and differentiation of human multipotent mesenchymal stromal cells. BMC Cell Biol (2010) 11:11. doi:10.1186/1471-2121-11-11
31. Tamama K, Kawasaki H, Kerpedjieva SS, Guan J, Ganju RK, Sen CK. Differential roles of hypoxia inducible factor subunits in multipotential stromal cells under hypoxic condition. J Cell Biochem (2011) 112(3):804–17. doi:10.1002/jcb.22961
32. Jing D, Fonseca AV, Alakel N, Fierro FA, Muller K, Bornhauser M, et al. Hematopoietic stem cells in co-culture with mesenchymal stromal cells – modeling the niche compartments in vitro. Haematologica (2010) 95(4):542–50. doi:10.3324/haematol.2009.010736
33. Jing D, Wobus M, Poitz DM, Bornhauser M, Ehninger G, Ordemann R. Oxygen tension plays a critical role in the hematopoietic microenvironment in vitro. Haematologica (2012) 97(2):331–9. doi:10.3324/haematol.2011.050815
34. Benito J, Zeng Z, Konopleva M, Wilson WR. Targeting hypoxia in the leukemia microenvironment. Int J Hematol Oncol (2013) 2(4):279–88. doi:10.2217/IJH.13.32
35. Martin SK, Diamond P, Gronthos S, Peet DJ, Zannettino ACW. The emerging role of hypoxia, HIF-1 and HIF-2 in multiple myeloma. Leukemia (2011) 25(10):1533–42. doi:10.1038/leu.2011.122
36. Ishikawa F, Yoshida S, Saito Y, Hijikata A, Kitamura H, Tanaka S, et al. Chemotherapy-resistant human AML stem cells home to and engraft within the bone-marrow endosteal region. Nat Biotechnol (2007) 25(11):1315–21. doi:10.1038/nbt1350
37. Mimeault M, Batra SK. Hypoxia-inducing factors as master regulators of stemness properties and altered metabolism of cancer- and metastasis-initiating cells. J Cell Mol Med (2013) 17(1):30–54. doi:10.1111/jcmm.12004
38. Pollyea DA, Gutman JA, Gore L, Smith CA, Jordan CT. Targeting acute myeloid leukemia stem cells: a review and principles for the development of clinical trials. Haematologica (2014) 99(8):1277–84. doi:10.3324/haematol.2013.085209
39. Lee KE, Simon MC. From stem cells to cancer stem cells: HIF takes the stage. Curr Opin Cell Biol (2012) 24(2):232–5. doi:10.1016/j.ceb.2012.01.005
40. Peng G, Liu Y. Hypoxia-inducible factors in cancer stem cells and inflammation. Trends Pharmacol Sci (2015) 36(6):374–83. doi:10.1016/j.tips.2015.03.003
41. Semenza GL. Hypoxia-inducible factors: mediators of cancer progression and targets for cancer therapy. Trends Pharmacol Sci (2012) 33(4):207–14. doi:10.1016/j.tips.2012.01.005
42. Wang GL, Jiang BH, Rue EA, Semenza GL. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci U S A (1995) 92(12):5510–4. doi:10.1073/pnas.92.12.5510
43. Keith B, Johnson RS, Simon MC. HIF1α and HIF2α: sibling rivalry in hypoxic tumour growth and progression. Nat Rev Cancer (2012) 12(1):9–22. doi:10.1038/nrc3183
44. Duan C. Hypoxia-inducible factor 3 biology: complexities and emerging themes. Am J Physiol Cell Physiol (2016) 310(4):C260–9. doi:10.1152/ajpcell.00315.2015
45. Heikkilä M, Pasanen A, Kivirikko KI, Myllyharju J. Roles of the human hypoxia-inducible factor (HIF)-3α variants in the hypoxia response. Cell Mol Life Sci (2011) 68(23):3885–901. doi:10.1007/s00018-011-0679-5
46. Augstein A, Poitz DM, Braun-Dullaeus RC, Strasser RH, Schmeisser A. Cell-specific and hypoxia-dependent regulation of human HIF-3α: inhibition of the expression of HIF target genes in vascular cells. Cell Mol Life Sci (2011) 68(15):2627–42. doi:10.1007/s00018-010-0575-4
47. Koh MY, Powis G. Passing the baton: the HIF switch. Trends Biochem Sci (2012) 37(9):364–72. doi:10.1016/j.tibs.2012.06.004
48. Lando D, Peet DJ, Gorman JJ, Whelan DA, Whitelaw ML, Bruick RK. FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor. Genes Dev (2002) 16(12):1466–71. doi:10.1101/gad.991402.1466
49. Holmquist-Mengelbier L, Fredlund E, Löfstedt T, Noguera R, Navarro S, Nilsson H, et al. Recruitment of HIF-1α and HIF-2α to common target genes is differentially regulated in neuroblastoma: HIF-2α promotes an aggressive phenotype. Cancer Cell (2006) 10(5):413–23. doi:10.1016/j.ccr.2006.08.026
50. Li Z, Bao S, Wu Q, Wang H, Eyler C, Sathornsumetee S, et al. Hypoxia-inducible factors regulate tumorigenic capacity of glioma stem cells. Cancer Cell (2009) 15(6):501–13. doi:10.1016/j.ccr.2009.03.018
51. Kocabas F, Zheng J, Thet S, Copeland NG, Jenkins NA, DeBerardinis RJ, et al. Meis1 regulates the metabolic phenotype and oxidant defense of hematopoietic stem cells. Blood (2012) 120(25):4963–72. doi:10.1182/blood-2012-05-432260
52. Kirito K, Fox N, Komatsu N, Kaushansky K, Tpo T. Thrombopoietin enhances expression of vascular endothelial growth factor (VEGF) in primitive hematopoietic cells through induction of HIF-1α. Blood (2005) 105(11):4258–63. doi:10.1182/blood-2004-07-2712
53. Gibbs BF, Yasinska IM, Oniku AE, Sumbayev VV. Effects of stem cell factor on hypoxia-inducible factor 1 alpha accumulation in human acute myeloid leukaemia and LAD2 mast cells. PLoS One (2011) 6(7):e22502. doi:10.1371/journal.pone.0022502
54. Pedersen M, Löfstedt T, Sun J, Holmquist-Mengelbier L, Påhlman S, Rönnstrand L. Stem cell factor induces HIF-1α at normoxia in hematopoietic cells. Biochem Biophys Res Commun (2008) 377(1):98–103. doi:10.1016/j.bbrc.2008.09.102
55. Dioum EM, Chen R, Alexander MS, Zhang Q, Hogg RT, Gerard RD, et al. Regulation of hypoxia-inducible factor 2α signaling by the stress-responsive deacetylase sirtuin 1. Science (2009) 324(5932):1289–93. doi:10.1126/science.1169956
56. Lim J-H, Lee Y-M, Chun Y-S, Chen J, Kim J-E, Park J-W. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1α. Mol Cell (2010) 38(6):864–78. doi:10.1016/j.molcel.2010.05.023
57. Selak MA, Armour SM, MacKenzie ED, Boulahbel H, Watson DG, Mansfield KD, et al. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-α prolyl hydroxylase. Cancer Cell (2005) 7(1):77–85. doi:10.1016/j.ccr.2004.11.022
58. Ravi R, Mookerjee B, Bhujwalla ZM, Sutter CH, Artemov D, Zeng Q, et al. Regulation of tumor angiogenesis by p53-induced degradation of hypoxia-inducible factor 1α. Genes Dev (2000) 14(1):34–44. doi:10.1101/gad.14.1.34
59. Yamakuchi M, Lotterman CD, Bao C, Hruban RH, Karim B, Mendell JT, et al. P53-induced microRNA-107 inhibits HIF-1 and tumor angiogenesis. Proc Natl Acad Sci U S A (2010) 107(14):6334–9. doi:10.1073/pnas.0911082107
60. Mottet D, Dumont V, Deccache Y, Demazy C, Ninane N, Raes M, et al. Regulation of hypoxia-inducible factor-1alpha protein level during hypoxic conditions by the phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3beta pathway in HepG2 cells. J Biol Chem (2003) 278(33):31277–85. doi:10.1074/jbc.M300763200
61. Zhang Q, Oh CK, Messadi DV, Duong HS, Kelly AP, Soo C, et al. Hypoxia-induced HIF-1 α accumulation is augmented in a co-culture of keloid fibroblasts and human mast cells: involvement of ERK1/2 and PI-3K/Akt. Exp Cell Res (2006) 312(2):145–55. doi:10.1016/j.yexcr.2005.10.006
62. Bernardi R, Guernah I, Jin D, Grisendi S, Alimonti A, Teruya-Feldstein J, et al. PML inhibits HIF-1alpha translation and neoangiogenesis through repression of mTOR. Nature (2006) 442(7104):779–85. doi:10.1038/nature05029
63. Takubo K, Suda T. Roles of the hypoxia response system in hematopoietic and leukemic stem cells. Int J Hematol (2012) 95(5):478–83. doi:10.1007/s12185-012-1071-4
64. Bangoura G, Liu Z-S, Qian Q, Jiang C-Q, Yang G-F, Jing S. Prognostic significance of HIF-2alpha/EPAS1 expression in hepatocellular carcinoma. World J Gastroenterol (2007) 13(23):3176–82. doi:10.3748/wjg.v13.i23.3176
65. Birner P, Schindl M, Obermair A, Plank C, Breitenecker G, Oberhuber G. Overexpression of hypoxia-inducible factor 1alpha is a marker for an unfavorable prognosis in early-stage invasive cervical cancer. Cancer Res (2000) 60(17):4693–6.
66. Chi J-T, Wang Z, Nuyten DSA, Rodriguez EH, Schaner ME, Salim A, et al. Gene expression programs in response to hypoxia: cell type specificity and prognostic significance in human cancers. PLoS Med (2006) 3(3):e47. doi:10.1371/journal.pmed.0030047
67. Giatromanolaki A, Koukourakis MI, Sivridis E, Turley H, Talks K, Pezzella F, et al. Relation of hypoxia inducible factor 1 alpha and 2 alpha in operable non-small cell lung cancer to angiogenic/molecular profile of tumours and survival. Br J Cancer (2001) 85(6):881–90. doi:10.1054/bjoc.2001.2018
68. Hui EP, Chan ATC, Pezzella F, Turley H, To K-F, Poon TCW, et al. Coexpression of hypoxia-inducible factors 1alpha and 2alpha, carbonic anhydrase IX, and vascular endothelial growth factor in nasopharyngeal carcinoma and relationship to survival. Clin Cancer Res (2002) 8:2595–604.
69. Koukourakis MI, Giatromanolaki A, Sivridis E, Simopoulos C, Turley H, Talks K, et al. Hypoxia-inducible factor (HIF1A and HIF2A), angiogenesis, and chemoradiotherapy outcome of squamous cell head-and-neck cancer. Int J Radiat Oncol Biol Phys (2002) 53(5):1192–202. doi:10.1016/S0360-3016(02)02848-1
70. Matsuyama T, Nakanishi K, Hayashi T, Yoshizumi Y, Aiko S, Sugiura Y, et al. Expression of hypoxia-inducible factor-1alpha in esophageal squamous cell carcinoma. Cancer Sci (2005) 96(3):176–82. doi:10.1111/j.1349-7006.2005.00025.x
71. Noguera R, Fredlund E, Piqueras M, Pietras A, Beckman S, Navarro S, et al. HIF-1α and HIF-2α are differentially regulated in vivo in neuroblastoma: high HIF-1α correlates negatively to advanced clinical stage and tumor vascularization. Clin Cancer Res (2009) 15(23):7130–6. doi:10.1158/1078-0432.CCR-09-0223
72. Scrideli CA, Carlotti CG, Mata JF, Neder L, Machado HR, Oba-Sinjo SM, et al. Prognostic significance of co-overexpression of the EGFR/IGFBP-2/HIF-2A genes in astrocytomas. J Neurooncol (2007) 83(3):233–9. doi:10.1007/s11060-007-9328-0
73. Sivridis E, Giatromanolaki A, Gatter KC, Harris AL, Koukourakis MI. Association of hypoxia-inducible factors 1alpha and 2alpha with activated angiogenic pathways and prognosis in patients with endometrial carcinoma. Cancer (2002) 95(5):1055–63. doi:10.1002/cncr.10774
74. Theodoropoulos VE, Lazaris AC, Sofras F, Gerzelis I, Tsoukala V, Ghikonti I, et al. Hypoxia-inducible factor 1α expression correlates with angiogenesis and unfavorable prognosis in bladder cancer. Eur Urol (2004) 46(2):200–8. doi:10.1016/j.eururo.2004.04.008
75. Trastour C, Benizri E, Ettore F, Ramaioli A, Chamorey E, Pouysségur J, et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer (2007) 120(7):1451–8. doi:10.1002/ijc.22436
76. Wu X-H, Qian C, Yuan K. Correlations of hypoxia-inducible factor-1α/hypoxia-inducible factor-2α expression with angiogenesis factors expression and prognosis in non-small cell lung cancer. Chin Med J (2011) 124(1):11–8. doi:10.3760/cma.j.issn.0366-6999.2011.01.004
77. Yoshimura H, Dhar D, Kohno H, Kubota H, Fujii T, Ueda S, et al. Prognostic impact of hypoxia-inducible factors 1 alpha and 2 alpha in colorectal cancer patients: correlation with tumor angiogenesis and cyclooxygenase-2 expression. Clin Cancer Res (2004) 10(24):8554–60. doi:10.1158/1078-0432.CCR-0946-03
78. Deeb G, Vaughan MM, McInnis I, Ford LA, Sait SNJ, Starostik P, et al. Hypoxia-inducible factor-1α protein expression is associated with poor survival in normal karyotype adult acute myeloid leukemia. Leuk Res (2011) 35(5):579–84. doi:10.1016/j.leukres.2010.10.020
79. Gao XN, Yan F, Lin J, Gao L, Lu XL, Wei SC, et al. AML1/ETO cooperates with HIF1α to promote leukemogenesis through DNMT3a transactivation. Leukemia (2015) 29(8):1730–40. doi:10.1038/leu.2015.56
80. Li Y, Ye D. Cancer therapy by targeting hypoxia-inducible factor-1. Curr Cancer Drug Targets (2010) 10(7):782–96. doi:10.2174/156800910793605857
81. Zhe N, Wang J, Chen S, Lin X, Chai Q, Zhang Y, et al. Heme oxygenase-1 plays a crucial role in chemoresistance in acute myeloid leukemia. Hematology (2015) 20(7):384–91. doi:10.1179/1607845414Y.0000000212
82. Forristal CE, Brown AL, Helwani FM, Winkler IG, Nowlan B, Barbier V, et al. Hypoxia inducible factor (HIF)-2α accelerates disease progression in mouse models of leukemia and lymphoma but is not a poor prognosis factor in human AML. Leukemia (2015) 29(10):2075–85. doi:10.1038/leu.2015.102
83. Kawada H, Kaneko M, Sawanobori M, Uno T, Matsuzawa H, Nakamura Y, et al. High concentrations of l-ascorbic acid specifically inhibit the growth of human leukemic cells via downregulation of HIF-1α transcription. PLoS One (2013) 8(4):e62717. doi:10.1371/journal.pone.0062717
84. Wang Y, Liu Y, Malek SN, Zheng P, Liu Y. Targeting HIF1α eliminates cancer stem cells in hematological malignancies. Cell Stem Cell (2011) 8(4):399–411. doi:10.1016/j.stem.2011.02.006
85. Coltella N, Percio S, Valsecchi R, Cuttano R, Guarnerio J, Ponzoni M, et al. HIF factors cooperate with PML-RARα to promote acute promyelocytic leukemia progression and relapse. EMBO Mol Med (2014) 6(5):640–50. doi:10.1002/emmm.201303065
86. Chen H, Shen Y, Gong F, Jiang Y, Zhang R. HIF-α promotes chronic myelogenous leukemia cell proliferation by upregulating p21 expression. Cell Biochem Biophys (2015) 72(1):179–83. doi:10.1007/s12013-014-0434-2
87. Zhang H, Li H, Xi HS, Li S. HIF1α is required for survival maintenance of chronic myeloid leukemia stem cells. Blood (2012) 119(11):2595–607. doi:10.1182/blood-2011-10-387381
88. Song K, Li M, Xu X-J, Xuan L, Huang G-N, Song X-L, et al. HIF-1α and GLUT1 gene expression is associated with chemoresistance of acute myeloid leukemia. Asian Pac J Cancer Prev (2014) 15(4):1823–9. doi:10.7314/APJCP.2014.15.4.1823
89. Tong H, Hu C, Zhuang Z, Wang L, Jin J. Hypoxia-inducible factor-1α expression indicates poor prognosis in myelodysplastic syndromes. Leuk Lymphoma (2012) 53(12):2412–8. doi:10.3109/10428194.2012.696637
90. Rouault-Pierre K, Lopez-Onieva L, Foster K, Anjos-Afonso F, Lamrissi-Garcia I, Serrano-Sanchez M, et al. HIF-2α protects human hematopoietic stem/progenitors and acute myeloid leukemic cells from apoptosis induced by endoplasmic reticulum stress. Cell Stem Cell (2013) 13(5):549–63. doi:10.1016/j.stem.2013.08.011
91. Wellmann S, Guschmann M, Griethe W, Eckert C, Stackelberg A, Lottaz C, et al. Activation of the HIF pathway in childhood ALL, prognostic implications of VEGF. Leukemia (2004) 18(5):926–33. doi:10.1038/sj.leu.2403332
92. Wang Y, Liu Y, Tang F, Bernot KM, Schore R, Marcucci G, et al. Echinomycin protects mice against relapsed acute myeloid leukemia without adverse effect on hematopoietic stem cells. Blood (2014) 124(7):1127–36. doi:10.1182/blood-2013-12-544221.The
93. Giambra V, Jenkins CE, Lam SH, Hoofd C, Belmonte M, Wang X, et al. Leukemia stem cells in T-ALL require active Hif1α and Wnt signaling. Blood (2015) 125(25):3917–27. doi:10.1182/blood-2014-10-609370
94. Zou J, Li P, Lu F, Liu N, Dai J, Ye J, et al. Notch1 is required for hypoxia-induced proliferation, invasion and chemoresistance of T-cell acute lymphoblastic leukemia cells. J Hematol Oncol (2013) 6:3. doi:10.1186/1756-8722-6-3
95. Coltella N, Valsecchi R, Ponente M, Ponzoni M, Bernardi R. Synergistic leukemia eradication by combined treatment with retinoic acid and HIF inhibition by EZN-2208 (PEG-SN38) in preclinical models of PML-RAR and PLZF-RAR-driven leukemia. Clin Cancer Res (2015) 21(16):3685–94. doi:10.1158/1078-0432.CCR-14-3022
96. Vlaminck B, Toffoli S, Ghislain B, Demazy C, Raes M. Dual effect of echinomycin on hypoxia-inducible factor-1 activity under normoxic and hypoxic conditions. FEBS J (2007) 274:5533–42. doi:10.1111/j.1742-4658.2007.06072.x
97. Greenberger LM, Horak ID, Filpula D, Sapra P, Westergaard M, Frydenlund HF, et al. A RNA antagonist of hypoxia-inducible factor-1, EZN-2968, inhibits tumor cell growth. Mol Cancer Ther (2008) 7(11):3598–608. doi:10.1158/1535-7163.MCT-08-0510
98. Simsek T, Kocabas F, Zheng J, Deberardinis RJ, Mahmoud AI, Olson EN, et al. The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche. Cell Stem Cell (2010) 7(3):380–90. doi:10.1016/j.stem.2010.07.011
99. Suda T, Takubo K, Semenza GL. Metabolic regulation of hematopoietic stem cells in the hypoxic niche. Cell Stem Cell (2011) 9(4):298–310. doi:10.1016/j.stem.2011.09.010
100. Zhang CC, Sadek HA. Hypoxia and metabolic properties of hematopoietic stem cells. Antioxid Redox Signal (2014) 20(12):1891–901. doi:10.1089/ars.2012.5019
101. Forristal CE, Winkler IG, Nowlan B, Barbier V, Walkinshaw G, Levesque JP. Pharmacologic stabilization of HIF-1α increases hematopoietic stem cell quiescence in vivo and accelerates blood recovery after severe irradiation. Blood (2013) 121(5):759–69. doi:10.1182/blood-2012-02-408419
102. Takubo K, Goda N, Yamada W, Iriuchishima H, Ikeda E, Kubota Y, et al. Regulation of the HIF-1α level is essential for hematopoietic stem cells. Cell Stem Cell (2010) 7(3):391–402. doi:10.1016/j.stem.2010.06.020
103. Palazon A, Goldrath AW, Nizet V, Johnson RS. HIF transcription factors, inflammation, and immunity. Immunity (2014) 41(4):518–28. doi:10.1016/j.immuni.2014.09.008
104. Gezer D, Vukovic M, Soga T, Pollard PJ, Kranc KR. Concise review: genetic dissection of hypoxia signaling pathways in normal and leukemic stem cells. Stem Cells (2014) 32(6):1390–7. doi:10.1002/stem.1657
105. Yang L, Rau R, Goodell MA. DNMT3A in haematological malignancies. Nat Rev Cancer (2015) 15(3):152–65. doi:10.1038/nrc3895
106. Mayle A, Yang L, Rodriguez B, Zhou T, Chang E, Curry CV, et al. Dnmt3a loss predisposes murine hematopoietic stem cells to malignant transformation. Blood (2015) 125(4):629–39. doi:10.1182/blood-2014-08-594648
107. Watson JA, Watson CJ, McCann A, Baugh J. Epigenetics, the epicenter of the hypoxic response. Epigenetics (2010) 5(4):293–6. doi:10.4161/epi.5.4.11684
108. Mutoh T, Sanosaka T, Ito K, Nakashima K. Oxygen levels epigenetically regulate fate switching of neural precursor cells via hypoxia-inducible factor 1α-Notch signal interaction in the developing brain. Stem Cells (2012) 30(3):561–9. doi:10.1002/stem.1019
109. Beyer S, Kristensen MM, Jensen KS, Johansen JV, Staller P. The histone demethylases JMJD1A and JMJD2B are transcriptional targets of hypoxia-inducible factor HIF. J Biol Chem (2008) 283(52):36542–52. doi:10.1074/jbc.M804578200
110. Fu L, Chen L, Yang J, Ye T, Chen Y, Fang J. HIF-1alpha-induced histone demethylase JMJD2B contributes to the malignant phenotype of colorectal cancer cells via an epigenetic mechanism. Carcinogenesis (2012) 33(9):1664–73. doi:10.1093/carcin/bgs217
111. Krieg AJ, Rankin EB, Chan D, Razorenova O, Fernandez S, Giaccia AJ. Regulation of the histone demethylase JMJD1A by hypoxia-inducible factor 1 alpha enhances hypoxic gene expression and tumor growth. Mol Cell Biol (2010) 30(1):344–53. doi:10.1128/MCB.00444-09
112. Drolle H, Wagner M, Vasold J, Kütt A, Deniffel C, Sotlar K, et al. Hypoxia regulates proliferation of acute myeloid leukemia and sensitivity against chemotherapy. Leuk Res (2015) 39(7):779–85. doi:10.1016/j.leukres.2015.04.019
113. Matsunaga T, Imataki O, Torii E, Kameda T, Shide K, Shimoda H, et al. Elevated HIF-1α expression of acute myelogenous leukemia stem cells in the endosteal hypoxic zone may be a cause of minimal residual disease in bone marrow after chemotherapy. Leuk Res (2012) 36(6):e122–4. doi:10.1016/j.leukres.2012.02.028
114. Yook Y-J, Seo Y-J, Kang HJ, Ko S-H, Shin HY, Lee JJ, et al. Induction of hypoxia-inducible factor-1α inhibits drug-induced apoptosis in the human leukemic cell line HL-60. Korean J Hematol (2010) 45(3):158–63. doi:10.5045/kjh.2010.45.3.158
115. Huang Y, Du KM, Xue ZH, Yan H, Li D, Liu W, et al. Cobalt chloride and low oxygen tension trigger differentiation of acute myeloid leukemic cells: possible mediation of hypoxia-inducible factor-1α. Leukemia (2003) 17(11):2065–73. doi:10.1038/sj.leu.2403141
116. Jiang Y, Xue ZH, Shen WZ, Du KM, Yan H, Yu Y, et al. Desferrioxamine induces leukemic cell differentiation potentially by hypoxia-inducible factor-1α that augments transcriptional activity of CCAAT/enhancer-binding protein-α. Leukemia (2005) 19(7):1239–47. doi:10.1038/sj.leu.2403734
117. Kim JS, Cho EW, Chung HW, Kim IG. Effects of Tiron, 4,5-dihydroxy-1,3-benzene disulfonic acid, on human promyelotic HL-60 leukemia cell differentiation and death. Toxicology (2006) 223(1–2):36–45. doi:10.1016/j.tox.2006.03.004
118. Zhang J, Chen G-Q. Hypoxia-HIF-1α-C/EBPα/Runx1 signaling in leukemic cell differentiation. Pathophysiology (2009) 16(4):297–303. doi:10.1016/j.pathophys.2009.02.005
119. Zhang J, Song L-P, Huang Y, Zhao Q, Zhao K-W, Chen G-Q. Accumulation of hypoxia-inducible factor-1 protein and its role in the differentiation of myeloid leukemic cells induced by all-trans retinoic acid. Haematologica (2008) 93(10):1480–7. doi:10.3324/haematol.13096
120. He M, Wang QY, Yin QQ, Tang J, Lu Y, Zhou CX, et al. HIF-1α downregulates miR-17/20a directly targeting p21 and STAT3: a role in myeloid leukemic cell differentiation. Cell Death Differ (2013) 20(3):408–18. doi:10.1038/cdd.2012.130
121. Gordan JD, Bertout JA, Hu C-J, Diehl JA, Simon MC. HIF-2α promotes hypoxic cell proliferation by enhancing c-Myc transcriptional activity. Cancer Cell (2007) 11(4):335–47. doi:10.1016/j.ccr.2007.02.006
122. Nguyen-Khac F, Della Valle V, Lopez RG, Ravet E, Mauchauffé M, Friedman AD, et al. Functional analyses of the TEL-ARNT fusion protein underscores a role for oxygen tension in hematopoietic cellular differentiation. Oncogene (2006) 25(35):4840–7. doi:10.1038/sj.onc.1209503
123. Liu W, Guo M, Xu Y-B, Li D, Zhou Z-N, Wu Y-L, et al. Induction of tumor arrest and differentiation with prolonged survival by intermittent hypoxia in a mouse model of acute myeloid leukemia. Blood (2006) 107(2):698–707. doi:10.1182/blood-2005-03-1278
124. Velasco-Hernandez T, Hyrenius-Wittsten A, Rehn M, Bryder D, Cammenga J. HIF-1α can act as a tumor suppressor gene in murine acute myeloid leukemia. Blood (2014) 124(24):3597–607. doi:10.1182/blood-2014-04-567065
125. Velasco-Hernandez T, Tornero D, Cammenga J. Loss of HIF-1α accelerates murine FLT-3ITD-induced myeloproliferative neoplasia. Leukemia (2015) 29(12):2366–74. doi:10.1038/leu.2015.156
126. Vukovic M, Guitart AV, Sepulveda C, Villacreces A, O’Duibhir E, Panagopoulou TI, et al. Hif-1α and Hif-2α synergize to suppress AML development but are dispensable for disease maintenance. J Exp Med (2015) 212(13):2223–34. doi:10.1084/jem.20150452
127. Di Giacomo V, Rapino M, Miscia S, Di Giulio C, Cataldi A. Dual role of HIF-1alpha in delivering a survival or death signal in hypoxia exposed human K562 erythroleukemia cells. Cell Biol Int (2009) 33(1):49–56. doi:10.1016/j.cellbi.2008.10.014
128. Silveira VS, Freire BMR, Borges KS, Andrade AF, Cruzeiro GAV, Sabino JPJ, et al. Hypoxia-related gene expression profile in childhood acute lymphoblastic leukemia: prognostic implications. Leuk Lymphoma (2014) 55(8):1751–7. doi:10.3109/10428194.2013.858812
129. Acker T, Diez-Juan A, Aragones J, Tjwa M, Brusselmans K, Moons L, et al. Genetic evidence for a tumor suppressor role of HIF-2α. Cancer Cell (2005) 8:131–41. doi:10.1016/j.ccr.2005.07.003
130. Lidgren A, Herdberg Y, Grankvist K, Rasmuson T, Vasko J, Ljungberg B. The expression of hypoxia-inducible factor 1α is a favorable independent prognostic factor in renal cell carcinoma. Clin Cancer Res (2005) 11(3):1129–35.
131. Mazumdar J, Hickey MM, Pant DK, Durham AC, Sweet-Cordero A, Vachani A, et al. HIF-2alpha deletion promotes Kras-driven lung tumor development. Proc Natl Acad Sci U S A (2010) 107(32):14182–7. doi:10.1073/pnas.1001296107
132. Hu J, Handisides DR, Van Valckenborgh E, De Raeve H, Menu E, Vande Broek I, et al. Targeting the multiple myeloma hypoxic niche with TH-302, a hypoxia-activated prodrug. Blood (2010) 116(9):1524–7. doi:10.1182/blood-2010-02-269126
133. Portwood S, Lal D, Hsu YC, Vargas R, Johnson MK, Wetzler M, et al. Activity of the hypoxia-activated prodrug, TH-302, in preclinical human acute myeloid leukemia models. Clin Cancer Res (2013) 19(23):6506–19. doi:10.1158/1078-0432.CCR-13-0674
134. Benito J, Shi Y, Szymanska B, Carol H, Boehm I, Lu H, et al. Pronounced hypoxia in models of murine and human leukemia: high efficacy of hypoxia-activated prodrug PR-104. PLoS One (2011) 6(8):e23108. doi:10.1371/journal.pone.0023108
135. Patterson AV, Ferry DM, Edmunds SJ, Gu Y, Singleton RS, Patel K, et al. Mechanism of action and preclinical antitumor activity of the novel hypoxia-activated DNA cross-linking agent PR-104. Clin Cancer Res (2007) 13(13):3922–32. doi:10.1158/1078-0432.CCR-07-0478
Keywords: hypoxia, hypoxia-inducible factors, leukemia, microenvironment, cancer
Citation: Deynoux M, Sunter N, Hérault O and Mazurier F (2016) Hypoxia and Hypoxia-Inducible Factors in Leukemias. Front. Oncol. 6:41. doi: 10.3389/fonc.2016.00041
Received: 12 January 2016; Accepted: 08 February 2016;
Published: 26 February 2016
Edited by:
Christian Gomez, University of Mississippi Medical Center, USAReviewed by:
Persio Dello Sbarba, Università degli Studi di Firenze, ItalyDaniele Tibullo, University of Catania, Italy
Carine Michiels, University of Namur, Belgium
Copyright: © 2016 Deynoux, Sunter, Hérault and Mazurier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédéric Mazurier, frederic.mazurier@inserm.fr