 Dragomir Milovanovic
Dragomir Milovanovic Reinhard Jahn
Reinhard Jahn- Department of Neurobiology, Max Planck Institute for Biophysical Chemistry, Göttingen, Germany
Our view of the lateral organization of lipids and proteins in the plasma membrane has evolved substantially in the last few decades. It is widely accepted that many, if not all, plasma membrane proteins and lipids are organized in specific domains. These domains vary widely in size, composition, and stability, and they represent platforms governing diverse cell functions. The presynaptic plasma membrane is a well-studied example of a membrane which undergoes rearrangements, especially during exo- and endocytosis. Many proteins and lipids involved in presynaptic function are known, and major efforts have been made to understand their spatial organization and dynamics. Here, we focus on the mechanisms underlying the organization of SNAREs, the key proteins of the fusion machinery, in distinct domains, and we discuss the functional significance of these clusters.
Introduction: SNAREs as Tools to Understand the Physical Priniciples Behind Membrane Patterning
Half a century ago biological membranes were envisioned as lipid bilayers which are covered on both faces with proteins that hydrophilically interact with the phospholipid head-groups (Robertson, 1963). However, with an increasing understanding of hydrophobic interactions between proteins and lipids (Kauzmann, 1959) it became evident that the plasma membrane is not fully covered by protein layers, but rather contains two types of proteins: the ones which span the entire bilayer (integral) and the ones which are embedded in only a single of the monolayers (peripheral). These membrane proteins were shown to undergo lateral mobility in the membrane (Lenard and Singer, 1966; Wallach and Zahler, 1968; Frye and Edidin, 1970; Singer and Nicolson, 1972). Equally important were data on the structure of lipid bilayers and on the existence of different lipid phases (i.e., solid, liquid-crystalline and liquid-ordered), as well as dependence of this phase transition on the temperature and acyl chain saturation (Steim et al., 1969; Melchior et al., 1970; Wilkins et al., 1971). Together, these developments led to a major change of concept: Membranes were considered as dynamic structures in which all components were able to diffuse laterally, with the membrane proteins “floating like icebergs in a sea” of membrane lipids (fluid mosaic model; Singer and Nicolson, 1972). This model still forms the foundation of our present understanding of biological membranes, with its major tenets being confirmed in numerous studies. However, it is becoming evident that there is an added layer of complexity in the lateral organization of proteins and lipids: Membrane contains subdomains whose properties are only slowly emerging.
The first hint for membrane subdomains resulted from experiments in which biological membranes were treated with certain detergents. Surprisingly, not all components were solubilized, i.e., incorporated individually in detergent micelles. Rather, detergent-resistant proteolipid complexes were isolated that were highly enriched in specific components. It was proposed that these complexes represent specific lipid domains in the plasma membrane, so-called lipid rafts, to which certain proteins preferentially associate (Simons and Ikonen, 1997). Certain lipid mixtures (dependent on the degree of side chain saturation and the amount of cholesterol) undergo multiple phase transitions, with equilibrium between different phases (most prominently Lo and Ld phases) coexisting at physiological temperatures. Insertion of proteins then revealed that proteins mostly partition into the Ld phase; however some proteins accumulate in the Lo-phase that is also referred as a raft (de Almeida et al., 2003, 2004). Such phase transitions were also observable in vesicles formed from blebs of the plasma membrane (Levental et al., 2011; Sezgin et al., 2012), supporting the idea that lipid phase partitioning is a major factor in generating subdomains within the membrane plane. However, single particle tracking and fluorescence correlation spectroscopy experiments suggested that the formation of such large phases is prevented in plasma membranes. This may partially be due to an underlying actin meshwork that acts as a fence together with some of the membrane proteins docked into this meshwork as pickets (hence, picket-fence model) (Jacobson et al., 1981; Dietrich et al., 2002; Fujiwara et al., 2002; Owen et al., 2009; Kusumi et al., 2010, 2011). Finally, owing to the development of superresolution optical microscopy, it is becoming clear that many membrane proteins are predominantly organized in nanometer-sized clusters and that multiple different clusters are co-existing in the plasma membrane (Sieber et al., 2007; Saka et al., 2014; Wilhelm et al., 2014).
Evidently, the presence of many different domains in the plasma membrane with distinct compositions cannot be explained by only one, or just a few, parameters. The plasma membrane contains thousands of different lipid and protein species (van Meer et al., 2008); hence a two-component system such as liquid ordered and liquid disordered phases does not provide a satisfactory explanation. Intriguingly, the protein occupancy of the bilayer volume (~20%) as indicated by the analysis of organelles (Takamori et al., 2006) and plasma membranes (Dupuy and Engelman, 2008) is much higher than originally anticipated. Thus, proteins may form phases on their own, together with certain lipids (Anderson and Jacobson, 2002). Additionally, one of the main characteristics of the plasma membrane is the asymmetry of the bilayer with the inner leaflet containing substantial amount of charged phospholipids (Bigay and Antonny, 2012). Therefore, new concepts are emerging, which consider electrostatic interactions, protein-protein (homo- and heterotypic) interactions, and hydrophobic interactions between bilayer core and protein transmembrane domains as parameters that contribute to segregation of proteins and lipids in distinct domains. Recently, a few studies have tried to merge these different mechanisms into a single model (Diaz-Rohrer et al., 2014; Milovanovic et al., 2015). In addition, it is becoming apparent that plasma membrane domains represent local hot spots that are essential for the functional segregation of distinct cellular processes. Due to technical limitations, progress in our understanding of membrane domains is largely confined to the plasma membrane. However, it will be interesting to understand if the same protein and lipid organizing principles also apply to organelles where they may play a role in processes such as sorting during trafficking and in organelle biogenesis.
Proteins involved in synaptic vesicle release have served as excellent models for analyzing the patterning of the plasma membrane. Synaptic vesicle release itself is a well-orchestrated process where a neurotransmitter-loaded vesicle attaches to the plasma membrane (a process known as docking), after which the fusion machinery enters a “preparatory” phase (known as priming) and then, once there is a calcium influx, the vesicle fuses with the plasma membrane (Südhof, 2004). Membrane fusion, the key step in neurotransmitter release is mediated by the interaction between protein members of the soluble NSF-attached protein receptor (SNARE) family that reside in the donor membrane with their cognate partners in the target membrane (Jahn and Scheller, 2006; Hong and Lev, 2014). SNARE proteins posses a central 60-70 AA-long motif (SNARE domain) that forms a coiled coil upon the interaction with the cognate SNARE partners. This coiled coil is connected by sixteen layers of interacting amino acid side chains that are hydrophobic (the flanking are all polar or charged) except of the amino acids in the central layer, which are either glutamine (Q) or arginine (R). Generally, the coiled-coil SNARE complex has three domains that contain glutamine together with one that contains arginine (QabcR) (Sutton et al., 1998; Antonin et al., 2002; Stein et al., 2009). The SNAREs involved in neuronal exocytosis include the plasma membrane residents syntaxin 1A (Qa) and SNAP 25 that contributes with two SNARE motifs (Qbc), and synaptobrevin 2 at the synaptic vesicle (R).
In recent years, an increasing body of evidence has revealed that SNAREs form clusters in both plasma membranes and intracellular membranes. Multiple approaches have yielded an increasingly refined picture of the forces and of the other biophysical parameters responsible for SNARE clustering, which will be discussed in the following chapters.
Segregation within the Hydrophoibc Core of the Membrane
Clustering Induced by Lipid Phases
As discussed above, lipid-based domain segregation was first postulated based on the observation that certain proteins tend to associate with specific lipid species (most conspicuously with cholesterol and sphingomyelin) and resist extraction by some nonionic detergents (Simons and van Meer, 1988; Brown and Rose, 1992; Schroeder et al., 1994). For many years, it has been controversially discussed whether these “detergent-resistant membranes (DRMs)” represent pre-existing structural entities that are preserved during extraction or are artificial aggregates generated by the extraction itself (Silvius et al., 1996; London and Brown, 2000; Heerklotz and Seelig, 2002; Sot et al., 2002; de Almeida et al., 2003). Presently, consensus is emerging that DRMs are largely created during extraction but many of the components enriched in DRMs appear to also associate with each other in an intact membrane. Thus, DRMs continue to be a useful tool for identifying candidates for membrane domains, but they do not represent a homogeneous population. For instance, GPI-anchored thymocyte antigen 1, ganglioside GM-1, and the membrane spanning linker for activation of T cells are all enriched when purified using DRMs, but are shown to form distinct clusters in the membrane (Wilson et al., 2004; Lichtenberg et al., 2005).
Although initial studies reported enrichment of SNAREs within DRMs (Lafont et al., 1999; Chamberlain et al., 2001; Chamberlain and Gould, 2002; Predescu et al., 2005; Salaün et al., 2005), it soon became clear that by applying different detergents, SNAREs were not co-floating with the classical DRM markers (Lang et al., 2001; Ohara-Imaizumi et al., 2004a). However, similar to DRMs cholesterol is required for the integrity of SNARE clusters (Lang et al., 2001; Lang, 2007). Furthermore, cholesterol depletion inhibits exocytosis in both neuronal (Lang et al., 2001) and non-neuronal cells such as epithelial (Chintagari et al., 2006) and endothelial (Predescu et al., 2005) cells, but it is still unclear whether dispersal of SNARE clusters and inhibition of fusion are causally related. Beyond neurotransmitter release, SNARE clusters are shown to be the release sites for cytokines at the phagocytic cup (Kay et al., 2006) and insulin (Ohara-Imaizumi et al., 2004a,b). Additionally, in vitro reconstitution of neuronal SNARE proteins into giant unilamellar liposomes capable of undergoing phase segregation suggested that SNAREs distribute in the liquid disordered phase (unsaturated phospholipids, cholesterol depleted regions), rather than in the liquid ordered phases (rich in saturated phospholipids and cholesterol). Although such simple phase-separation may not reflect phase-partitioning in the plasma membranes, these studies confirmed that SNAREs do not associate with sphingomyelin and saturated phospholipids (Saslowsky et al., 2003; Bacia et al., 2004). On the other hand they demonstrate that SNARE proteins are sensitive to such phase partitioning, raising the possibility that phase heterogeneity may contribute to SNARE segregation.
Clustering Induced by Hydrophobic Mismatch
Hydrophobic mismatch occurs when the length of the protein transmembrane domains (TMDs) does not match the bilayer thickness. In this case, it is energetically favorable to cluster the TMDs of similar length in the same region rather than to accommodate each of the TMDs separately. In a theoretical paper, Mouritsen and Bloom proposed that proteins may cluster in order to minimize membrane mismatch (Mouritsen and Bloom, 1984). Pioneering research showed that certain enzymes have the highest activity when reconstituted in bilayers of particular thickness, whereas in both thinner and thicker bilayers the activity drops (Johannsson et al., 1981a,b; Kusumi and Hyde, 1982). This implied that hydrophobic mismatch affects enzyme conformation that subsequently reduces its activity. Moreover, the aggregation state of some of these proteins such as rhodopsin is shown to depend on the acyl-chain length of lipids that were used for the reconstitution (Kusumi and Hyde, 1982). More recently, it was also shown that the overlap between the TMD length of the perfringolysin O, a multispanning barrel protein, and the width of lipid bilayer also affects the proteins' distribution and functionality in proteoliposomes (Lin and London, 2013). Protein clustering driven by hydrophobic mismatch was first shown directly for synthetic TMD peptides (de Planque et al., 1998; Sparr et al., 2005). The phospholipid acyl-chains are flexible and their lateral organization depends on the neighboring lipid molecules (i.e., cholesterol restricts the flexibility due to the pronounced hydrophobic planar structure). Hence, lipids can adopt to a range of different thicknesses (Zaccai, 2000). On the other hand, proteins exhibit less flexibility in length distortion in the case of membrane mismatch (Petrache et al., 2002). Caution is needed when interpreting experiments based on altering acyl chain lengths because these changes also affect the lipid packing, curvature and surface charge distribution. Hence, the protein function may be affected by many of these parameters (Anderson and Jacobson, 2002).
Hydrophobic mismatch appears to play a role in defining the final destination of membrane components in intracellular trafficking. It is well-established that sorting of proteins and lipids in polarized, epithelial cells is mediated by both the lipid environment and the cytoskeleton, and that lipid domains coalesce prior to vesicle formation (Brown and Rose, 1992; Lipowsky, 1993; Yoshimori et al., 1996; Roux et al., 2005). Additionally, altering the TMD length of peptides affected their trafficking from ER, Golgi to the plasma membrane (Munro, 1991, 1995; Pelham and Munro, 1993; Nilsson et al., 1996). Considering that the average thickness of the membrane increases from ER (~3.75 nm) to the plasma membrane (~4.25 nm; Mitra et al., 2004), it is reasonable to expect that proteins destined to the plasma membrane have longer TMDs. Indeed, in a comprehensive screen of the TMDs sequences from different species, Sharpe et al. demonstrate that an average length of TMDs is about 5 amino acids shorter for proteins destined to ER compared to the proteins of the plasma membrane (Sharpe et al., 2010).
Intriguingly, thickness differences within the plane of the plasma membrane have recently been shown to be important for lateral sorting of proteins into distinct domains (Milovanovic et al., 2015). The crystal structure of the neuronal SNARE complex revealed that the TMD of syntaxin 1 might be too short to span the entire plasma membrane (Stein et al., 2009). In comparison to syntaxin 1 (involved in calcium regulated exocytosis), syntaxin 4 (involved in the constitutive exocytosis) has a slightly (1–2 residues) longer TMD. We have recently shown that the TMD length difference of a single amino acid between syntaxin 1 and syntaxin 4 can drive the proteins into separate clusters in the plasma membrane (Milovanovic et al., 2015). Here, the effect of cholesterol on local alterations of membrane thickness plays an important role. The average plasma membrane thickness is estimated to be around 4 nm (Mitra et al., 2004) and incorporation of physiological amounts (30%) of cholesterol increases the membrane thickness by about 0.35 nm (Milovanovic et al., 2015). Therefore, cholesterol can drive protein clustering by inducing or enhancing local hydrophobic mismatch in the membrane (Figure 1). This probably explains why cholesterol depletion causes the dispersal of SNARE domains in the membrane and a concomitant decrease in exocytosis (Lang et al., 2001; Chamberlain and Gould, 2002; Ohara-Imaizumi et al., 2004a; Kay et al., 2006).
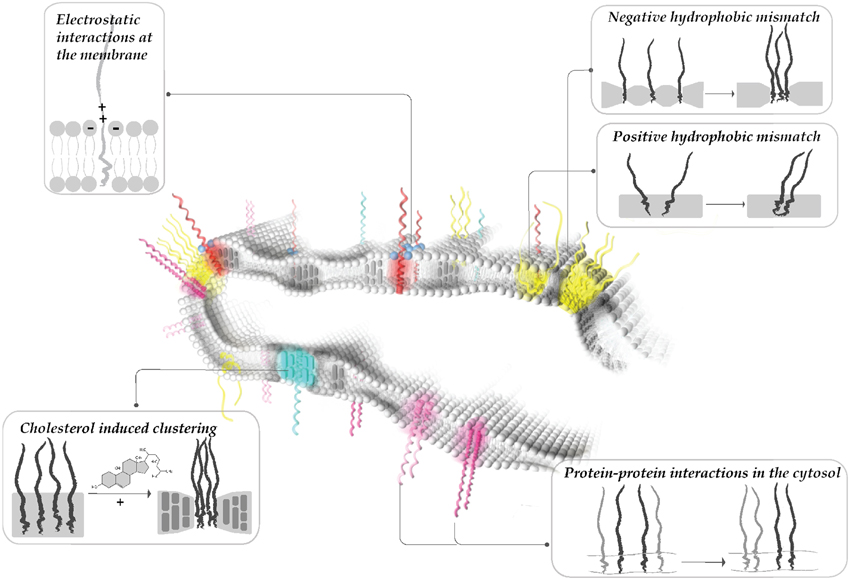
Figure 1. Modularity of protein clustering. Hydrophobic interactions between the core of the lipid bilayer and the protein TMDs (i.e., hydrophobic mismatch, interactions mediated by cholesterol), electrostatic interactions at the membrane surface (i.e., positive patches of proteins and polyphosphoinositides) and specific protein-protein interactions at the cytoplasmic surface all affect membrane patterning. Multiple mechanisms affect the protein/lipid organization simultaneously, leading to dynamic reorganization of protein clusters.
Palmitoylation Modulates Attachment of Soluble Proteins to the Membrane
Posttranslational modifications further modulate SNARE patterning (for detailed review see van den Bogaart et al., 2013). Most attention has been paid to palmitoylation, i.e., the covalent addition of the acyl chain palmitate (C16:0) to a cysteine residue in the protein. For instance, the Qbc SNAREs SNAP 23 and 25 are palmitoylated (Prescott et al., 2009) at five and four cysteine residues, respectively, which is required for membrane attachment. Proteomics analyses suggested that many other synaptic proteins undergo palmitoylation including proteins containing TMDs (Kang et al., 2008), among these are the SNAREs syntaxin 1 and synaptobrevin 2. It has been suggested that SNAREs are reversibly targeted to cholesterol and sphingomyelin rich regions via palmitoylation (Fukata and Fukata, 2010; Levental et al., 2010), which would add another mechanism contributing to cluster formation. Support for this concept is provided by the recent finding that a fraction of amyloid precursor is palmitoylated, which further modulates its association with cholesterol-rich regions in the presynaptic membrane (Bhattacharyya et al., 2013).
Segregation Caused by Interactions at the Hydrophobic-Hydrophilic Boundary
Clustering of SNAREs is influenced by electrostatic interactions between positively charged side chains adjacent to the hydrophobic TMD with negatively charged polyphosphoinositides (Di Paolo et al., 2004; Do Heo et al., 2006; van den Bogaart et al., 2011). Tamm and colleagues (Wagner and Tamm, 2001) showed that diffusion of syntaxin 1 decreases upon incorporation of PI(4,5)P2 in the lipid monolayer. PI(4,5)P2 is enriched in regions of the plasma membrane where secretory vesicles dock (Laux et al., 2000; Aoyagi et al., 2005), and it is essential for exocytosis (Hay and Martin, 1993; Milosevic et al., 2005; James et al., 2008; Wen et al., 2011). Although PI(4,5)P2 comprises only 1% of total lipids of the plasma membrane (Di Paolo and De Camilli, 2006), it can reach concentrations of more than 80% of total lipids in clusters (van den Bogaart et al., 2011). Association between syntaxin 1 and PI(4,5)P2 was clearly shown both in vitro reconstituted systems (Murray and Tamm, 2009, 2011) and in cells (van den Bogaart et al., 2011). Responsible for this strong interaction is a cluster of positively charged arginines and lysines directly adjacent to the TMD of syntaxin 1 (van den Bogaart et al., 2011; Khuong et al., 2013).
Ionic interactions between macromolecules are strongly influenced by mobile ions. The ionic composition at the surface of a membrane is highly complex (Wang et al., 2012, 2014), rendering it difficult to quantify the influence of ions on domain formation. Ions present at high concentrations on the cytoplasmic surface (K+, Mg2+, glutamate, ATP) are able to shield the charge of both lipid head-groups and proteins involved in exocytosis (Park et al., 2012). It is worth noting that calcium increases syntaxin 1 clustering in the plasma membrane of PC12 cells (Zilly et al., 2011), and this mechanism might involve the interaction with negatively charged lipids. Polybasic clusters on the cytoplasmic face adjacent to transmembrane proteins are common among many membrane proteins (von Heijne, 2006). Thus, it is possible that such ionic interactions play a major role in patterning of the plasma membrane and possibly also of intracellular membranes (Figure 1).
Segregation due to Interactions in the Hydrophilic Space
Both homophilic and heterophilic interactions have been described for Qa SNARE family members. For instance, syntaxin 1 and syntaxin 4 are involved in regulated and constitutive exocytosis, respectively. Interactions between the SNARE motifs at the cytoplasmic surface has been suggested to contribute to the segregation of these proteins into distinct domains (Sieber et al., 2006). Hence, in case of syntaxin isoforms homotypic protein interactions contribute to the functional segregation (Figure 1). Similarly, in non-neuronal cells, syntaxin isoforms segregate in different regions of the membrane. In highly polarized epithelial cells, syntaxin 3, and syntaxin 4 are trafficked distinctly to the apical and basolateral membrane, respectively. Even the deletion of the targeting signal of syntaxin 3 does not eliminate its distinct segregation from syntaxin 4 enriched regions (Low et al., 2006).
Heterotypic protein interactions are important for both the spatial sorting of proteins in the presynapse, as well as for catalyzing the fusion reaction. For instance, some presynaptic membrane proteins bind to the actin meshwork (Torregrosa-Hetland et al., 2011, 2013; Villanueva et al., 2012). This binding to the cytoskeleton can be direct as in the case of syntaxin 4 (Jewell et al., 2008; Woronowicz et al., 2010) and SNAP 25 (Torregrosa-Hetland et al., 2013). Alternatively, binding to actin can be mediated by adaptor proteins such as myosin V that connects syntaxin 1 to actin (Watanabe et al., 2005), and α-fodrin that connects syntaxins 3 and 4 to actin (Nakano et al., 2001). Another example for heterotypic interactions includes binding of regulatory proteins to SNAREs. The SM-protein Munc 18 that binds to syntaxin 1 is not only essential for exocytosis (Verhage et al., 2000) but also necessary for trafficking of syntaxin 1 to the plasma membrane (Voets et al., 2001; Yang et al., 2006; Kurps and de Wit, 2012). Indeed, if syntaxin 1 clusters serve as reservoir of the protein for fusion, Munc 18 may be needed to pry a individual syntaxin 1 molecules away from the cluster (Bar-On et al., 2012). Munc 18 bound syntaxin 1 is able to recruit SNAP 25 in the cell lawns and synaptobrevin 2 containing vesicles can bind to this complex (Zilly et al., 2006).
Functional Relevance of SNARE Clustering
SNARE Clustering may be Important for Exocytosis
Several lines of evidence suggest three important physiological roles for SNARE clustering in exocytosis (Figure 2). First, the high local concentrations of SNAREs at the plasma membrane may provide the functional pools of proteins necessary for the formation of SNARE complexes. Clustering of SNAREs may also prevent nonproductive side-reactions of the highly reactive SANREs such as the formation of so-called “dead-end” complexes between syntaxin 1 and SNAP 25 incapable of fusion (Fasshauer and Margittai, 2004). It has been shown that the plasma membrane of chromaffin cells lacks these dead-end complexes (Halemani et al., 2010). Also, removal of cholesterol does not only affect the clustering of SNAREs, but also reduces the number of functionally active syntaxin 1/SNAP 25 complexes ready for ternary complex formation with synaptobrevin 2 (Rickman et al., 2010). Second, Q-SNARE domains [together with PI(4,5)P2] may represent docking platforms for vesicles (James et al., 2008; de Wit et al., 2009; Imig et al., 2014). PI(4,5)P2 was shown to be enriched at the sites of vesicle fusion, and altering the amount of PI(4,5)P2 affects the release capacities (Milosevic et al., 2005; de Wit et al., 2009). Therefore, PI(4,5)P2 domains have been proposed to act as molecular beacons for vesicle recruitment to the membrane. Indeed, synaptotagmin 1, the main calcium sensor for exocytosis, binds to the syntaxin 1/PI(4,5)P2 clusters in the plasma domains (Honigmann et al., 2013). Finally, clustering may decrease the energy barrier that needs to be overcome for membrane fusion in two ways. First, line tension around the cluster locally destabilizes the membrane. Based on several studies, such micro-destabilized regions decrease the energy barrier needed for membrane remodeling (Boucrot et al., 2012; Kozlov et al., 2014; Risselada et al., 2014). The lower the energy barrier, the less SNARE complexes are needed for successful fusion (Mohrmann et al., 2010; van den Bogaart et al., 2010; Hernandez et al., 2014). Secondly, initial experiments emphasized that the interaction of not only SNARE motifs but also of the C-terminal TMDs of syntaxin 1 and synaptobrevin 2 was seen as necessary for the processing from the hemifused to the fully fused state (i.e., fusion pore opening) (Grote et al., 2000; Han et al., 2004; Fdez et al., 2010). Recent work from Südhof's lab indicated that this is not the case since fusion was possible in cells where the syntaxin 1 TMD was replaced with a lipid anchor (Zhou et al., 2013). Interestingly, bioactive molecules such as anesthetics (e.g., isoflurane, etomidate and propofol) reduce the release capacities of chromaffin cells (Herring et al., 2009, 2011; Xie et al., 2013). Since they interact with SANREs and SNARE-associated proteins, they may be altering the lateral organization of SNARE clusters (Herring et al., 2011). Clustered SNAREs are in dynamic exchange with the surrounding membrane, and diffusion of SNARE molecules between clusters is rather high (Sieber et al., 2007; Barg et al., 2010; Knowles et al., 2010; Wilhelm et al., 2014). As discussed above, syntaxin clusters may serve as molecular beacons (or hot spots) for vesicle docking. However, the exact assembly of fusion competent SNARE complexes most probably takes place adjacent to these clusters (Rickman et al., 2005; Bar-On et al., 2012; Gandasi and Barg, 2014).
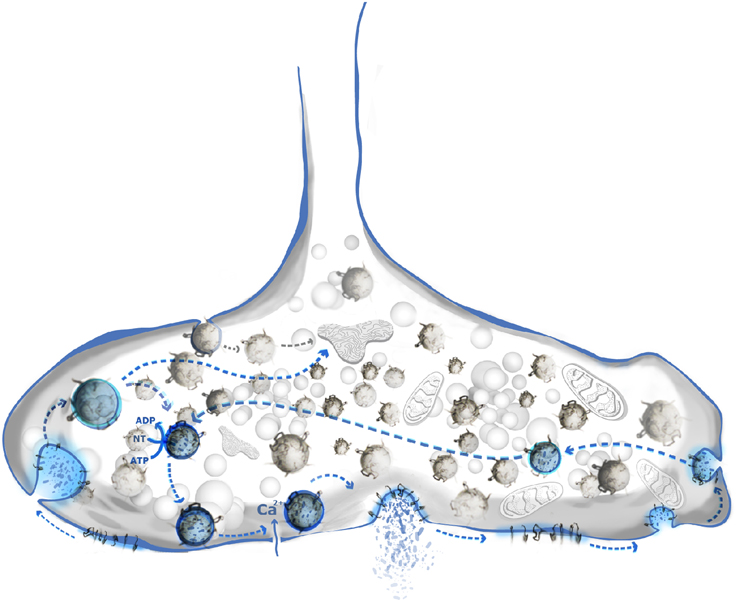
Figure 2. SNARE clustering has multiple roles in synaptic vesicle cycle. Classical synaptic vesicle cycle (blue shade) includes: neurotransmitter loading, docking, priming, calcium triggered release, subsequent engulfing, and recycling of vesicle. Syntaxin 1/PI(4,5)P2 domains are shown to play an important role in vesicle docking. Components of vesicle's membrane remain clustered or are re-clustered prior to endocytosis. SNARE domains may be a sorting determinant, especially in the case of vesicles engulfed by rapid endocytosis.
SNARE Clustering may be Important for Endocytotic Retrieval of Vesicles
Neurotransmitter release is a rapid and repetitive process. In order to maintain membrane balance vesicle fusion and fission have to be tightly spatially and temporally coupled (Figure 2). During endocytosis, vesicle-specific proteins are selectively retrieved while plasma membrane residents are excluded. Even during sustained, high activity, the composition of synaptic vesicles needs to remain constant. While some flexibility may be tolerated for abundant proteins such as synaptobrevin 2 (~70 copies/vesicle) some of the functionally essential proteins are present only 1–2 copies/vesicle, e.g., the vacuolar ATPase required for neurotransmitter uptake (Takamori et al., 2006). Using STED microscopy, Willig et al. (2006) suggested that that SV proteins remain clustered after exocytosis. Alternatively, SV proteins may be sorted and re-clustered prior to endocytosis (Hua et al., 2011). Further, SNAREs are connected to the proteins that regulate the vesicle release such as soluble SM proteins and tethering factors. The region of the presynaptic membrane where vesicles dock and fuse is distinguished by defined structural elements (Szule et al., 2012; Fernandez-Busnadiego et al., 2013; Harlow et al., 2013; Imig et al., 2014). These “active zones” contain multiple protein complexes that regulate tethering and docking (Fernandez-Busnadiego et al., 2013; Imig et al., 2014), but the details of the structure and dynamics of the underlying protein-protein interactions are only slowly emerging. Interestingly, a study that combined the electron microscopy and the STED nanoscopy showed that synaptic vesicle proteins such as synaptotagmin remain clustered even within the early endosome, thus being a marker for synaptic vesicle retrieval (Hoopmann et al., 2010). Generally, there appear to be at least two main pathways for vesicle endocytosis: (i) slow, clathrin-mediated endocytosis (CME), and (ii) fast, mostly clathrin-independent, endocytosis. CME has been extensively studied (Jung and Haucke, 2007; Dittman and Ryan, 2009). The relatively slow kinetics of CME (~20 s) cannot fully explain fast vesicle turnover at the synaptic bouton (Heuser and Reese, 1973; Gandhi and Stevens, 2003). Using a combination of optogenetics and high-pressure freezing electron microscopy, Jorgensen and colleagues showed that a second type of endocytosis co-exists in neurons that can be very rapid (~30 ms) but is likely to be less accurate than CME, resulting in endocytotic membrane vesicles larger than SV (Watanabe et al., 2014a,b, 2013a,b). Apart from speed, the availability of endocytotic machinery might be the limiting step in CME during the sustained SV release. Indeed, quantitative analysis of the synaptic bouton showed that there are about five folds less endocytotic than exocytotic proteins (Wilhelm et al., 2014). This problem may be overcome by fast, bulk endocytosis that requires fewer proteins to be involved in vesicle engulfing (Watanabe et al., 2013b).
It is still debated to which extent endocytosed vesicles need to pass through an additional endosomal sorting step before re-entering the SV pool. It is conceivable that the fate of the endocytosed membrane is determined by its protein and lipid components (Rizzoli, 2014). Shortly after exocytosis the protein content of the synaptic vesicle either remains clustered (Willig et al., 2006), or it diffuses in the plane of the membrane which is followed by immediate re-clustering (Wienisch and Klingauf, 2006; Hua et al., 2011). Specific adaptor proteins such as AP2, stonin and AP 180 specifically bind to synaptic vesicle proteins such as synaptobrevin 2 (AP 180) (Granseth et al., 2006) or synaptotagmin 1 (AP2, stonin) (Collins et al., 2002; Jung et al., 2007), ensuring their clustering in a coated pit (Glyvuk et al., 2010). It is conceivable that a clathrin-coated vesicle separating from the plasma membrane matches the membrane composition of synaptic vesicles (as already suggested earlier; Maycox et al., 1992), allowing for immediate re-use after uncoating without an intermediate sorting step (Watanabe et al., 2014b). In contrast, it is highly unlikely that vesicles retrieved from the plasma membrane by ultrarapid endocytosis are sorted with similarly high accuracy. It is conceivable that these vesicles need to “proof-read” by cytoplasmic factors after endocytosis (Figure 2). If the protein and lipid content of such an endocytosed vesicle meets the requirements for a functional synaptic vesicle, the vesicle might be loaded with NT and can be immediately used for the next round of the release. Otherwise, the vesicle is targeted to recycling endosomes for further sorting. The precise sorting mechanism is far from understood and the sorting signals involved in vesicle recycling and the maintenance of the vesicle identity still need to be identified.
Conclusion
SNARE proteins have turned out to be excellent paradigms for studying the biochemical and biophysical mechanisms that govern domain formation in biological membranes. The picture that is emerging reveals a modular interplay of different forces and interactions that are all required for the complex patterning of a membrane into multiple different clusters (Figure 1). These parameters include: (i) hydrophobic interactions between the core of the lipid bilayer and the hydrophobic membrane anchors of the proteins such as hydrophobic mismatch, phase partitioning, and cholesterol-mediated interactions, (ii) interactions between negatively charged lipid head-groups with positively charged clusters frequently observed at the membrane surface, (iii) specific protein–protein interactions at the cytoplasmic surface. Many of these clusters are not static features of the membrane but rather represent dynamic entities where clustered and thus mostly immobile proteins and lipids are in equilibrium with their rapidly diffusing monomeric or oligomeric counterparts. Future work is needed to appreciate the heterogeneity of the clusters, their molecular composition, and to clarify whether only some or most membrane proteins are organized in such domains.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Geert van den Bogaart and Akihiro Kusumi for the valuable discussion and input during the preparation of this review; Christian Hoffmann, Achintya Prahlad for careful reading and comments on the manuscript. We are grateful to Tanja Grubic (www.tgau-design.com) for giving the visual identity to the illustrations. DM is a graduate student at the International Max Planck Research School for Molecular Biology. This article is based on work supported by grants (to RJ) of the Deutsche Forschungsgemeinschaft (SFB803) and of the US National Institutes of Health (P01 GM072694).
References
Anderson, R. G. W., and Jacobson, K. (2002). A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science 296, 1821–1825. doi: 10.1126/science.1068886
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Antonin, W., Fasshauer, D., Becker, S., Jahn, R., and Schneider, T. R. (2002). Crystal structure of the endosomal SNARE complex reveals common structural principles of all SNAREs. Nat. Struct. Biol. 9, 107–111. doi: 10.1038/nsb746
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Aoyagi, K., Sugaya, T., Umeda, M., Yamamoto, S., Terakawa, S., and Takahashi, M. (2005). The activation of exocytotic sites by the formation of phosphatidylinositol 4,5-bisphosphate microdomains at syntaxin clusters. J. Biol. Chem. 280, 17346–17352. doi: 10.1074/jbc.M413307200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bacia, K., Schuette, C. G., Kahya, N., Jahn, R., and Schwille, P. (2004). SNAREs prefer liquid-disordered over “raft” (liquid-ordered) domains when reconstituted into giant unilamellar vesicles. J. Biol. Chem. 279, 37951–37955. doi: 10.1074/jbc.M407020200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Barg, S., Knowles, M. K., Chen, X., Midorikawa, M., and Almers, W. (2010). Syntaxin clusters assemble reversibly at sites of secretory granules in live cells. Proc. Natl. Acad. Sci. U.S.A. 107, 20804–20809. doi: 10.1073/pnas.1014823107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bar-On, D., Wolter, S., van de Linde, S., Heilemann, M., Nudelman, G., Nachliel, E., et al. (2012). Super-resolution imaging reveals the internal architecture of nano-sized syntaxin clusters. J. Biol. Chem. 287, 27158–27167. doi: 10.1074/jbc.M112.353250
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bhattacharyya, R., Barren, C., and Kovacs, D. M. (2013). Palmitoylation of amyloid precursor protein regulates amyloidogenic processing in lipid rafts. J. Neurosci. 33, 11169–11183. doi: 10.1523/JNEUROSCI.4704-12.2013
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bigay, J., and Antonny, B. (2012). Curvature, lipid packing, and electrostatics of membrane organelles: defining cellular territories in determining specificity. Dev. Cell 23, 886–895. doi: 10.1016/j.devcel.2012.10.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boucrot, E., Pick, A., Çamdere, G., Liska, N., Evergren, E., McMahon, H. T., et al. (2012). Membrane fission is promoted by insertion of amphipathic helices and is restricted by crescent BAR domains. Cell 149, 124–136. doi: 10.1016/j.cell.2012.01.047
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brown, D. A., and Rose, J. K. (1992). Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell-surface. Cell 68, 533–544. doi: 10.1016/0092-8674(92)90189-J
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chamberlain, L. H., Burgoyne, R. D., and Gould, G. W. (2001). SNARE proteins are highly enriched in lipid rafts in PC12 cells: implications for the spatial control of exocytosis. Proc. Natl. Acad. Sci. U.S.A. 98, 5619–5624. doi: 10.1073/pnas.091502398
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chamberlain, L. H., and Gould, G. W. (2002). The vesicle- and target-SNARE proteins that mediate Glut4 vesicle fusion are localized in detergent-insoluble lipid rafts present on distinct intracellular membranes. J. Biol. Chem. 277, 49750–49754. doi: 10.1074/jbc.M206936200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chintagari, N. R., Jin, N., Wang, P., Narasaraju, T. A., Chen, J., and Liu, L. (2006). Effect of cholesterol depletion on exocytosis of alveolar type II cells. Am. J. Respir. Cell Mol. Biol. 34, 677–687. doi: 10.1165/rcmb.2005-0418OC
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Collins, B. M., McCoy, A. J., Kent, H. M., Evans, P. R., and Owen, D. J. (2002). Molecular architecture and functional model of the endocytic AP2 complex. Cell 109, 523–535. doi: 10.1016/S0092-8674(02)00735-3
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Almeida, R. F. M., Fedorov, A., and Prieto, M. (2003). Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: boundaries and composition of lipid rafts. Biophys. J. 85, 2406–2416. doi: 10.1016/S0006-3495(03)74664-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Almeida, R., Loura, L., Prieto, M., Watts, A., Fedorov, A., and Barrantes, F. J. (2004). Cholesterol modulates the organization of the gamma M4 transmembrane domain of the muscle nicotinic acetylcholine receptor. Biophys. J. 86, 2261–2272. doi: 10.1016/S0006-3495(04)74284-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Planque, M. R., Greathouse, D. V., Koeppe, R. E., Schäfer, H., Marsh, D., and Killian, J. A. (1998). Influence of lipid/peptide hydrophobic mismatch on the thickness of diacylphosphatidylcholine bilayers. A 2H NMR and ESR study using designed transmembrane alpha-helical peptides and gramicidin A. Biochemistry 37, 9333–9345. doi: 10.1021/bi980233r
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Wit, H., Walter, A. M., Milosevic, I., Gulyas-Kovacs, A., Riedel, D., Sørensen, J. B., et al. (2009). Synaptotagmin-1 docks secretory vesicles to syntaxin-1/SNAP-25 acceptor complexes. Cell 138, 935–946. doi: 10.1016/j.cell.2009.07.027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Diaz-Rohrer, B. B., Levental, K. R., Simons, K., and Levental, I. (2014). Membrane raft association is a determinant of plasma membrane localization. Proc. Natl. Acad. Sci. U.S.A. 111, 8500–8505. doi: 10.1073/pnas.1404582111
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dietrich, C., Yang, B., Fujiwara, T., Kusumi, A., and Jacobson, K. (2002). Relationship of lipid rafts to transient confinement zones detected by single particle tracking. Biophys. J. 82, 274–284. doi: 10.1016/S0006-3495(02)75393-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Di Paolo, G., and De Camilli, P. (2006). Phosphoinositides in cell regulation and membrane dynamics. Nature 443, 651–657. doi: 10.1038/nature05185
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Di Paolo, G., Moskowitz, H. S., Gipson, K., Wenk, M. R., Voronov, S., Obayashi, M., et al. (2004). Impaired PtdIns(4,5)P-2 synthesis in nerve terminals produces defects in synaptic vesicle trafficking. Nature 431, 415–422. doi: 10.1038/nature02896
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dittman, J., and Ryan, T. A. (2009). Molecular circuitry of endocytosis at nerve terminals. Annu. Rev. Cell Dev. Biol. 25, 133–160. doi: 10.1146/annurev.cellbio.042308.113302
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Do Heo, W., Inoue, T., Park, W. S., Kim, M. L., Park, B. O., Wandless, T. J., et al. (2006). PI(3,4,5)P3 and PI(4,5)P2 lipids target proteins with polybasic clusters to the plasma membrane. Science 314, 1458–1461. doi: 10.1126/science.1134389
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dupuy, A. D., and Engelman, D. M. (2008). Protein area occupancy at the center of the red blood cell membrane. Proc. Natl. Acad. Sci. U.S.A. 105, 2848–2852. doi: 10.1073/pnas.0712379105
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fasshauer, D., and Margittai, M. (2004). A transient N-terminal interaction of SNAP-25 and syntaxin nucleates SNARE assembly. J. Biol. Chem. 279, 7613–7621. doi: 10.1074/jbc.M312064200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fdez, E., Martínez-Salvador, M., Beard, M., Woodman, P., and Hilfiker, S. (2010). Transmembrane-domain determinants for SNARE-mediated membrane fusion. J. Cell Sci. 123, 2473–2480. doi: 10.1242/jcs.061325
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fernandez-Busnadiego, R., Asano, S., Oprisoreanu, A. M., Sakata, E., Doengi, M., Kochovski, Z., et al. (2013). Cryo-electron tomography reveals a critical role of RIM1α in synaptic vesicle tethering. J. Cell Biol. 201, 725–740. doi: 10.1083/jcb.201206063
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Frye, L. D., and Edidin, M. (1970). The rapid intermixing of cell surface antigens after formation of mouse-human heterokaryons. J. Cell. Sci. 7, 319–335.
Fujiwara, T., Ritchie, K., Murakoshi, H., Jacobson, K., and Kusumi, A. (2002). Phospholipids undergo hop diffusion in compartmentalized cell membrane. J. Cell Biol. 157, 1071–1081. doi: 10.1083/jcb.200202050
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fukata, Y., and Fukata, M. (2010). Protein palmitoylation in neuronal development and synaptic plasticity. Nat. Rev. Neurosci. 11, 161–175. doi: 10.1038/nrn2788
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gandasi, N. R., and Barg, S. (2014). Contact-induced clustering of syntaxin and munc18 docks secretory granules at the exocytosis site. Nat. Commun. 5, 3914. doi: 10.1038/ncomms4914
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gandhi, S. P., and Stevens, C. F. (2003). Three modes of synaptic vesicular recycling revealed by single-vesicle imaging. Nature 423, 607–613. doi: 10.1038/nature01677
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Glyvuk, N., Tsytsyura, Y., Geumann, C., D'Hooge, R., Hüve, J., Kratzke, M., et al. (2010). AP-1/sigma1B-adaptin mediates endosomal synaptic vesicle recycling, learning and memory. EMBO J. 29, 1318–1330. doi: 10.1038/emboj.2010.15
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Granseth, B., Odermatt, B., Royle, S. J., and Lagnado, L. (2006). Clathrin-mediated endocytosis is the dominant mechanism of vesicle retrieval at hippocampal synapses. Neuron 51, 773–786. doi: 10.1016/j.neuron.2006.08.029
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grote, E., Baba, M., Ohsumi, Y., and Novick, P. J. (2000). Geranylgeranylated SNAREs are dominant inhibitors of membrane fusion. J. Cell Biol. 151, 453–466. doi: 10.1083/jcb.151.2.453
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Halemani, N. D., Bethani, I., Rizzoli, S. O., and Lang, T. (2010). Structure and dynamics of a two-helix SNARE complex in live cells. Traffic 11, 394–404. doi: 10.1111/j.1600-0854.2009.01020.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Han, X., Wang, C. T., Bai, J., Chapman, E. R., and Jackson, M. B. (2004). Transmembrane segments of syntaxin line the fusion pore of Ca2+-triggered exocytosis. Science 304, 289–292. doi: 10.1126/science.1095801
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Harlow, M. L., Szule, J. A., Xu, J., Jung, J. H., Marshall, R. M., and McMahan, U. (2013). Alignment of synaptic vesicle macromolecules with the macromolecules in active zone material that direct vesicle docking. PLoS ONE 8:e69410. doi: 10.1371/journal.pone.0069410
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hay, J. C., and Martin, T. F. (1993). Phosphatidylinositol transfer protein required for ATP-dependent priming of Ca(2+)-activated secretion. Nature 366, 572–575. doi: 10.1038/366572a0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heerklotz, H., and Seelig, J. (2002). Application of pressure perturbation calorimetry to lipid bilayers. Biophys. J. 82, 1445–1452. doi: 10.1016/S0006-3495(02)75498-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hernandez, J. M., Kreutzberger, A. J. B., Kiessling, V., Tamm, L. K., and Jahn, R. (2014). Variable cooperativity in SNARE-mediated membrane fusion. Proc. Natl. Acad. Sci. U.S.A. 111, 12037–12042. doi: 10.1073/pnas.1407435111
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Herring, B. E., McMillan, K., Pike, C. M., Marks, J., Fox, A. P., and Xie, Z. (2011). Etomidate and propofol inhibit the neurotransmitter release machinery at different sites. J. Physiol. (Lond.) 589, 1103–1115. doi: 10.1113/jphysiol.2010.200964
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Herring, B. E., Xie, Z., Marks, J., and Fox, A. P. (2009). Isoflurane inhibits the neurotransmitter release machinery. J. Neurophysiol. 102, 1265–1273. doi: 10.1152/jn.00252.2009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heuser, J. E., and Reese, T. S. (1973). Evidence for recycling of synaptic vesicle membrane during transmitter release at the frog neuromuscular junction. J. Cell Biol. 57, 315–344. doi: 10.1083/jcb.57.2.315
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hong, W., and Lev, S. (2014). Tethering the assembly of SNARE complexes. Trends Cell Biol. 24, 35–43. doi: 10.1016/j.tcb.2013.09.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Honigmann, A., van den Bogaart, G., Iraheta, E., Risselada, H. J., Milovanovic, D., Mueller, V., et al. (2013). Phosphatidylinositol 4,5-bisphosphate clusters act as molecular beacons for vesicle recruitment. Nat. Struct. Mol. Biol. 20, 679–686. doi: 10.1038/nsmb.2570
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hoopmann, P., Punge, A., Barysch, S. V., Westphal, V., Bückers, J., Opazo, F., et al. (2010). Endosomal sorting of readily releasable synaptic vesicles. Proc. Natl. Acad. Sci. U.S.A. 107, 19055–19060. doi: 10.1073/pnas.1007037107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hua, Y., Sinha, R., Thiel, C. S., Schmidt, R., Hüve, J., Martens, H., et al. (2011). A readily retrievable pool of synaptic vesicles. Nat. Neurosci. 14, 833–839. doi: 10.1038/nn.2838
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Imig, C., Min, S.-W., Krinner, S., Arancillo, M., Rosenmund, C., Südhof, T. C., et al. (2014). The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 84, 416–431. doi: 10.1016/j.neuron.2014.10.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jacobson, K., Hou, Y., Derzko, Z., Wojcieszyn, J., and Organisciak, D. (1981). Lipid lateral diffusion in the surface membrane of cells and in multibilayers formed from plasma membrane lipids. Biochemistry 20, 5268–5275. doi: 10.1021/bi00521a027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jahn, R., and Scheller, R. H. (2006). SNAREs–engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 7, 631–643. doi: 10.1038/nrm2002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
James, D. J., Khodthong, C., Kowalchyk, J. A., and Martin, T. F. J. (2008). Phosphatidylinositol 4,5-bisphosphate regulates SNARE-dependent membrane fusion. J. Cell Biol. 182, 355–366. doi: 10.1083/jcb.200801056
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jewell, J. L., Luo, W., Oh, E., Wang, Z., and Thurmond, D. C. (2008). Filamentous actin regulates insulin exocytosis through direct interaction with Syntaxin 4. J. Biol. Chem. 283, 10716–10726. doi: 10.1074/jbc.M709876200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Johannsson, A., Keightley, C. A., Smith, G. A., Richards, C. D., Hesketh, T. R., and Metcalfe, J. C. (1981a). The effect of bilayer thickness and n-alkanes on the activity of the (Ca2+ and Mg2+)-dependent ATPase of sarcoplasmic reticulum. J. Biol. Chem. 256, 1643–1650.
Johannsson, A., Smith, G. A., and Metcalfe, J. C. (1981b). The effect of bilayer thickness on the activity of (Na+ and K+)-ATPase. Biochim. Biophys. Acta 641, 416–421. doi: 10.1016/0005-2736(81)90498-3
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jung, N., and Haucke, V. (2007). Clathrin-mediated endocytosis at synapses. Traffic 8, 1129–1136. doi: 10.1111/j.1600-0854.2007.00595.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jung, N., Wienisch, M., Gu, M., Rand, J. B., Müller, S. L., Krause, G., et al. (2007). Molecular basis of synaptic vesicle cargo recognition by the endocytic sorting adaptor stonin 2. J. Cell Biol. 179, 1497–1510. doi: 10.1083/jcb.200708107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kang, R., Wan, J., Arstikaitis, P., Takahashi, H., Huang, K., Bailey, A. O., et al. (2008). Neural palmitoyl-proteomics reveals dynamic synaptic palmitoylation. Nature 456, 904–909. doi: 10.1038/nature07605
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kauzmann, W. (1959). Some factors in the interpretation of protein denaturation. Adv. Protein Chem. 14, 1–63. doi: 10.1016/S0065-3233(08)60608-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kay, J. G., Murray, R. Z., Pagan, J. K., and Stow, J. L. (2006). Cytokine secretion via cholesterol-rich lipid raft-associated SNAREs at the phagocytic cup. J. Biol. Chem. 281, 11949–11954. doi: 10.1074/jbc.M600857200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Khuong, T. M., Habets, R. L. P., Kuenen, S., Witkowska, A., Kasprowicz, J., Swerts, J., et al. (2013). Synaptic PI(3,4,5)P3 is required for Syntaxin1A clustering and neurotransmitter release. Neuron 77, 1097–1108. doi: 10.1016/j.neuron.2013.01.025
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Knowles, M. K., Barg, S., Wan, L., Midorikawa, M., Chen, X., and Almers, W. (2010). Single secretory granules of live cells recruit syntaxin-1 and synaptosomal associated protein 25 (SNAP-25) in large copy numbers. Proc. Natl. Acad. Sci. U.S.A. 107, 20810–20815. doi: 10.1073/pnas.1014840107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kozlov, M. M., Campelo, F., Liska, N., Chernomordik, L. V., Marrink, S. J., and McMahon, H. T. (2014). Mechanisms shaping cell membranes. Curr. Opin. Cell Biol. 29C, 53–60. doi: 10.1016/j.ceb.2014.03.006
Kurps, J., and de Wit, H. (2012). The role of munc18-1 and its orthologs in modulation of cortical F-actin in chromaffin cells. J. Mol. Neurosci. 48, 339–346. doi: 10.1007/s12031-012-9775-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kusumi, A., and Hyde, J. S. (1982). Spin-label saturation-transfer electron spin resonance detection of transient association of rhodopsin in reconstituted membranes. Biochemistry 21, 5978–5983. doi: 10.1021/bi00266a039
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kusumi, A., Shirai, Y. M., Koyama-Honda, I., Suzuki, K. G. N., and Fujiwara, T. K. (2010). Hierarchical organization of the plasma membrane: investigations by single-molecule tracking vs. fluorescence correlation spectroscopy. FEBS Lett. 584, 1814–1823. doi: 10.1016/j.febslet.2010.02.047
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kusumi, A., Suzuki, K. G. N., Kasai, R. S., Ritchie, K., and Fujiwara, T. K. (2011). Hierarchical mesoscale domain organization of the plasma membrane. Trends Biochem. Sci. 36, 604–615. doi: 10.1016/j.tibs.2011.08.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lafont, F., Verkade, P., Galli, T., Wimmer, C., Louvard, D., and Simons, K. (1999). Raft association of SNAP receptors acting in apical trafficking in Madin-Darby canine kidney cells. Proc. Natl. Acad. Sci. U.S.A. 96, 3734–3738. doi: 10.1073/pnas.96.7.3734
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lang, T. (2007). SNARE proteins and ‘membrane rafts’. J. Physiol. (Lond.) 585, 693–698. doi: 10.1113/jphysiol.2007.134346
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lang, T., Bruns, D., Wenzel, D., Riedel, D., Holroyd, P., Thiele, C., et al. (2001). SNAREs are concentrated in cholesterol−dependent clusters that define docking and fusion sites for exocytosis. EMBO J. 20, 2202–2213. doi: 10.1093/emboj/20.9.2202
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Laux, T., Fukami, K., Thelen, M., Golub, T., Frey, D., and Caroni, P. (2000). GAP43, MARCKS, and CAP23 modulate PI(4,5)P(2) at plasmalemmal rafts, and regulate cell cortex actin dynamics through a common mechanism. J. Cell Biol. 149, 1455–1472. doi: 10.1083/jcb.149.7.1455
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lenard, J., and Singer, S. J. (1966). Protein conformation in cell membrane preparations as studied by optical rotatory dispersion and circular dichroism. Proc. Natl. Acad. Sci. U.S.A. 56, 1828–1835. doi: 10.1073/pnas.56.6.1828
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Levental, I., Grzybek, M., and Simons, K. (2010). Greasing their way: lipid modifications determine protein association with membrane rafts. Biochemistry 49, 6305–6316. doi: 10.1021/bi100882y
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Levental, I., Grzybek, M., and Simons, K. (2011). Raft domains of variable properties and compositions in plasma membrane vesicles. Proc. Natl. Acad. Sci. U.S.A. 108, 11411–11416. doi: 10.1073/pnas.1105996108
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lichtenberg, D., Goñi, F. M., and Heerklotz, H. (2005). Detergent-resistant membranes should not be identified with membrane rafts. Trends Biochem. Sci. 30, 430–436. doi: 10.1016/j.tibs.2005.06.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lin, Q., and London, E. (2013). Altering hydrophobic sequence lengths shows that hydrophobic mismatch controls affinity for ordered lipid domains (rafts) in the multitransmembrane strand protein perfringolysin O. J. Biol. Chem. 288, 1340–1352. doi: 10.1074/jbc.M112.415596
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lipowsky, R. (1993). Domain-induced budding of fluid membranes. Biophys. J. 64, 1133–1138. doi: 10.1016/S0006-3495(93)81479-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
London, E., and Brown, D. A. (2000). Insolubility of lipids in triton X-100: physical origin and relationship to sphingolipid/cholesterol membrane domains (rafts). Biochim. Biophys. Acta 1508, 182–195. doi: 10.1016/S0304-4157(00)00007-1
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Low, S. H., Vasanji, A., Nanduri, J., He, M., Sharma, N., Koo, M., et al. (2006). Syntaxins 3 and 4 are concentrated in separate clusters on the plasma membrane before the establishment of cell polarity. Mol. Biol. Cell 17, 977–989. doi: 10.1091/mbc.E05-05-0462
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maycox, P. R., Link, E., Reetz, A., Morris, S. A., and Jahn, R. (1992). Clathrin-coated vesicles in nervous tissue are involved primarily in synaptic vesicle recycling. J. Cell Biol. 118, 1379–1388. doi: 10.1083/jcb.118.6.1379
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Melchior, D. L., Morowitz, H. J., Sturtevant, J. M., and Tsong, T. Y. (1970). Characterization of the plasma membrane of Mycoplasma laidlawii. VII. Phase transitions of membrane lipids. Biochim. Biophys. Acta 219, 114–122. doi: 10.1016/0005-2736(70)90066-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Milosevic, I., Sørensen, J. B., Lang, T., Krauss, M., Nagy, G., Haucke, V., et al. (2005). Plasmalemmal phosphatidylinositol-4,5-bisphosphate level regulates the releasable vesicle pool size in chromaffin cells. J. Neurosci. 25, 2557–2565. doi: 10.1523/JNEUROSCI.3761-04.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Milovanovic, D., Honigmann, A., Koike, S., Göttfert, F., Pähler, G., Junius, M., et al. (2015). Hydrophobic mismatch sorts SNARE proteins into distinct membrane doimains. Nat. Commun. 5:5984 doi: 10.1038/ncomms6984
Mitra, K., Ubarretxena-Belandia, I., Taguchi, T., Warren, G., and Engelman, D. M. (2004). Modulation of the bilayer thickness of exocytic pathway membranes by membrane proteins rather than cholesterol. Proc. Natl. Acad. Sci. U.S.A. 101, 4083–4088. doi: 10.1073/pnas.0307332101
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mohrmann, R., de Wit, H., Verhage, M., Neher, E., and Sørensen, J. B. (2010). Fast vesicle fusion in living cells requires at least three SNARE complexes. Science 330, 502–505. doi: 10.1126/science.1193134
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mouritsen, O. G., and Bloom, M. (1984). Mattress model of lipid-protein interactions in membranes. Biophys. J. 46, 141–153. doi: 10.1016/S0006-3495(84)84007-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Munro, S. (1991). Sequences within and adjacent to the transmembrane segment of alpha-2,6-sialyltransferase specify Golgi retention. EMBO J. 10, 3577–3588.
Munro, S. (1995). An investigation of the role of transmembrane domains in Golgi protein retention. EMBO J. 14, 4695–4704.
Murray, D. H., and Tamm, L. K. (2009). Clustering of syntaxin-1A in model membranes is modulated by phosphatidylinositol 4,5-bisphosphate and cholesterol. Biochemistry 48, 4617–4625. doi: 10.1021/bi9003217
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Murray, D. H., and Tamm, L. K. (2011). Molecular mechanism of cholesterol- and polyphosphoinositide-mediated syntaxin clustering. Biochemistry 50, 9014–9022. doi: 10.1021/bi201307u
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nakano, M., Nogami, S., Sato, S., Terano, A., and Shirataki, H. (2001). Interaction of syntaxin with alpha-fodrin, a major component of the submembranous cytoskeleton. Biochem. Biophys. Res. Commun. 288, 468–475. doi: 10.1006/bbrc.2001.5795
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nilsson, T., Rabouille, C., Hui, N., Watson, R., and Warren, G. (1996). The role of the membrane-spanning domain and stalk region of N-acetylglucosaminyltransferase I in retention, kin recognition and structural maintenance of the Golgi apparatus in HeLa cells. J. Cell. Sci. 109, 1975–1989.
Ohara-Imaizumi, M., Nishiwaki, C., Kikuta, T., Kumakura, K., Nakamichi, Y., and Nagamatsu, S. (2004a). Site of docking and fusion of insulin secretory granules in live MIN6 cells analyzed by TAT-conjugated anti-syntaxin 1 antibody and total internal reflection fluorescence microscopy. J. Biol. Chem. 279, 8403–8408. doi: 10.1074/jbc.M308954200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ohara-Imaizumi, M., Nishiwaki, C., Nakamichi, Y., Kikuta, T., Nagai, S., and Nagamatsu, S. (2004b). Correlation of syntaxin-1 and SNAP-25 clusters with docking and fusion of insulin granules analysed by total internal reflection fluorescence microscopy. Diabetologia 47, 2200–2207. doi: 10.1007/s00125-004-1579-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Owen, D. M., Williamson, D., Rentero, C., and Gaus, K. (2009). Quantitative microscopy: protein dynamics and membrane organisation. Traffic 10, 962–971. doi: 10.1111/j.1600-0854.2009.00908.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Park, Y., Hernandez, J. M., van den Bogaart, G., Ahmed, S., Holt, M., Riedel, D., et al. (2012). Controlling synaptotagmin activity by electrostatic screening. Nat. Struct. Mol. Biol. 19, 991–997. doi: 10.1038/nsmb.2375
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pelham, H. R., and Munro, S. (1993). Sorting of membrane proteins in the secretory pathway. Cell 75, 603–605. doi: 10.1016/0092-8674(93)90479-A
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Petrache, H. I., Zuckerman, D. M., Sachs, J. N., Killian, J. A., Koeppe, R. E., and Woolf, T. B. (2002). Hydrophobic matching mechanism investigated by molecular dynamics simulations. Langmuir 18, 1340–1351. doi: 10.1021/la011338p
Predescu, S. A., Predescu, D. N., Shimizu, K., Klein, I. K., and Malik, A. B. (2005). Cholesterol-dependent syntaxin-4 and SNAP-23 clustering regulates caveolar fusion with the endothelial plasma membrane. J. Biol. Chem. 280, 37130–37138. doi: 10.1074/jbc.M505659200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Prescott, G. R., Gorleku, O. A., Greaves, J., and Chamberlain, L. H. (2009). Palmitoylation of the synaptic vesicle fusion machinery. J. Neurochem. 110, 1135–1149. doi: 10.1111/j.1471-4159.2009.06205.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rickman, C., Hu, K., Carroll, J., and Davletov, B. (2005). Self-assembly of SNARE fusion proteins into star-shaped oligomers. Biochem. J. 388, 75–79. doi: 10.1042/BJ20041818
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rickman, C., Medine, C. N., Dun, A. R., Moulton, D. J., Mandula, O., Halemani, N. D., et al. (2010). t-SNARE protein conformations patterned by the lipid microenvironment. J. Biol. Chem. 285, 13535–13541. doi: 10.1074/jbc.M109.091058
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Risselada, H. J., Bubnis, G., and Grubmüller, H. (2014). Expansion of the fusion stalk and its implication for biological membrane fusion. Proc. Natl. Acad. Sci. U.S.A. 111, 11043–11048. doi: 10.1073/pnas.1323221111
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rizzoli, S. O. (2014). Synaptic vesicle recycling: steps and principles. EMBO J. 33, 788–822. doi: 10.1002/embj.201386357
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Robertson, J. D. (1963). The occurrence of a subunit pattern in the unit membranes of club endings in Mauthner cell synapses in goldfish brains. J. Cell Biol. 19, 201–221. doi: 10.1083/jcb.19.1.201
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Roux, A., Cuvelier, D., Nassoy, P., Prost, J., Bassereau, P., and Goud, B. (2005). Role of curvature and phase transition in lipid sorting and fission of membrane tubules. EMBO J. 24, 1537–1545. doi: 10.1038/sj.emboj.7600631
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Saka, S. K., Honigmann, A., Eggeling, C., Hell, S. W., Lang, T., and Rizzoli, S. O. (2014). Multi-protein assemblies underlie the mesoscale organization of the plasma membrane. Nat. Commun. 5, 4509. doi: 10.1038/ncomms5509
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Salaün, C., Gould, G. W., and Chamberlain, L. H. (2005). Lipid raft association of SNARE proteins regulates exocytosis in PC12 cells. J. Biol. Chem. 280, 19449–19453. doi: 10.1074/jbc.M501923200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Saslowsky, D. E., Lawrence, J. C., Henderson, R. M., and Edwardson, J. M. (2003). Syntaxin is efficiently excluded from sphingomyelin-enriched domains in supported lipid bilayers containing cholesterol. J. Membr. Biol. 194, 153–164. doi: 10.1007/s00232-003-2035-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schroeder, R., London, E., and Brown, D. (1994). Interactions between saturated acyl chains confer detergent resistance on lipids and glycosylphosphatidylinositol (GPI)-anchored proteins: GPI-anchored proteins in liposomes and cells show similar behavior. Proc. Natl. Acad. Sci. U.S.A. 91, 12130–12134. doi: 10.1073/pnas.91.25.12130
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sezgin, E., Kaiser, H.-J., Baumgart, T., Schwille, P., Simons, K., and Levental, I. (2012). Elucidating membrane structure and protein behavior using giant plasma membrane vesicles. Nat. Protoc. 7, 1042–1051. doi: 10.1038/nprot.2012.059
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sharpe, H. J., Stevens, T. J., and Munro, S. (2010). A comprehensive comparison of transmembrane domains reveals organelle-specific properties. Cell 142, 158–169. doi: 10.1016/j.cell.2010.05.037
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sieber, J. J., Willig, K. I., Heintzmann, R., Hell, S. W., and Lang, T. (2006). The SNARE Motif Is Essential for the Formation of Syntaxin Clusters in the Plasma Membrane. Biophys. J. 90, 2843–2851. doi: 10.1529/biophysj.105.079574
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sieber, J. J., Willig, K. I., Kutzner, C., Gerding-Reimers, C., Harke, B., Donnert, G., et al. (2007). Anatomy and dynamics of a supramolecular membrane protein cluster. Science 317, 1072–1076. doi: 10.1126/science.1141727
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Silvius, J. R., del Giudice, D., and Lafleur, M. (1996). Cholesterol at different bilayer concentrations can promote or antagonize lateral segregation of phospholipids of differing acyl chain length. Biochemistry 35, 15198–15208. doi: 10.1021/bi9615506
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Simons, K., and Ikonen, E. (1997). Functional rafts in cell membranes. Nature 387, 569–572. doi: 10.1038/42408
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Simons, K., and van Meer, G. (1988). Lipid sorting in epithelial cells. Biochemistry 27, 6197–6202. doi: 10.1021/bi00417a001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Singer, S. J., and Nicolson, G. L. (1972). The fluid mosaic model of the structure of cell membranes. Science 175, 720–731. doi: 10.1126/science.175.4023.720
Sot, J., Collado, M. I., Arrondo, J. L. R., Alonso, A., and Goñi, F. M. (2002). Triton X-100-resistant bilayers: effect of lipid composition and relevance to the raft phenomenon. Langmuir 18, 2828–2835. doi: 10.1021/la011381c
Sparr, E., Ganchev, D. N., Snel, M. M. E., Ridder, A. N. J. A., Kroon-Batenburg, L. M. J., Chupin, V., et al. (2005). Molecular organization in striated domains induced by transmembrane alpha-helical peptides in dipalmitoyl phosphatidylcholine bilayers. Biochemistry 44, 2–10. doi: 10.1021/bi048047a
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Steim, J. M., Tourtellotte, M. E., Reinert, J. C., McElhaney, R. N., and Rader, R. L. (1969). Calorimetric evidence for the liquid-crystalline state of lipids in a biomembrane. Proc. Natl. Acad. Sci. U.S.A. 63, 104–109. doi: 10.1073/pnas.63.1.104
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Stein, A., Weber, G., Wahl, M. C., and Jahn, R. (2009). Helical extension of the neuronal SNARE complex into the membrane. Nature 460, 525–528. doi: 10.1038/nature08156
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Südhof, T. C. (2004). The synaptic vesicle cycle. Annu. Rev. Neurosci. 27, 509–547. doi: 10.1146/annurev.neuro.26.041002.131412
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sutton, R. B., Fasshauer, D., Jahn, R., and Brunger, A. T. (1998). Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 A resolution. Nature 395, 347–353. doi: 10.1038/26412
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Szule, J. A., Harlow, M. L., Jung, J. H., De-Miguel, F. F., Marshall, R. M., and McMahan, U. (2012). Regulation of synaptic vesicle docking by different classes of macromolecules in active zone material. PLoS ONE 7:e33333. doi: 10.1371/journal.pone.0033333
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Takamori, S., Holt, M., Stenius, K., Lemke, E. A., Grønborg, M., Riedel, D., et al. (2006). Molecular anatomy of a trafficking organelle. Cell 127, 831–846. doi: 10.1016/j.cell.2006.10.030
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Torregrosa-Hetland, C. J., Villanueva, J., Garcia-Martinez, V., Expósito-Romero, G., Francés, M. D. M., and Gutiérrez, L. M. (2013). Cortical F-actin affects the localization and dynamics of SNAP-25 membrane clusters in chromaffin cells. Int. J. Biochem. Cell Biol. 45, 583–592. doi: 10.1016/j.biocel.2012.11.021
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Torregrosa-Hetland, C. J., Villanueva, J., Giner, D., López-Font, I., Nadal, A., Quesada, I., et al. (2011). The F-actin cortical network is a major factor influencing the organization of the secretory machinery in chromaffin cells. J. Cell Sci. 124, 727–734. doi: 10.1242/jcs.078600
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van den Bogaart, G., Holt, M. G., Bunt, G., Riedel, D., Wouters, F. S., and Jahn, R. (2010). One SNARE complex is sufficient for membrane fusion. Nat. Struct. Mol. Biol. 17, 358–364. doi: 10.1038/nsmb.1748
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van den Bogaart, G., Lang, T., and Jahn, R. (2013). Microdomains of SNARE proteins in the plasma membrane. Curr. Top. Membr. 72, 193–230. doi: 10.1016/B978-0-12-417027-8.00006-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van den Bogaart, G., Meyenberg, K., Risselada, H. J., Amin, H., Willig, K. I., Hubrich, B. E., et al. (2011). Membrane protein sequestering by ionic protein-lipid interactions. Nature 479, 552–555. doi: 10.1038/nature10545
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van Meer, G., Voelker, D. R., and Feigenson, G. W. (2008). Membrane lipids: where they are and how they behave. Nat. Rev. Mol. Cell Biol. 9, 112–124. doi: 10.1038/nrm2330
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Verhage, M., Maia, A. S., Plomp, J. J., Brussaard, A. B., Heeroma, J. H., Vermeer, H., et al. (2000). Synaptic assembly of the brain in the absence of neurotransmitter secretion. Science 4, 864–869. doi: 10.1126/science.287.5454.864
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Villanueva, J., Torregrosa-Hetland, C. J., Garcia-Martinez, V., del Mar Francés, M., Viniegra, S., and Gutiérrez, L. M. (2012). The F-actin cortex in chromaffin granule dynamics and fusion: a minireview. J. Mol. Neurosci. 48, 323–327. doi: 10.1007/s12031-012-9718-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Voets, T., Toonen, R. F., Brian, E. C., de Wit, H., Moser, T., Rettig, J., et al. (2001). Munc18-1 promotes large dense-core vesicle docking. Neuron 31, 581–591. doi: 10.1016/S0896-6273(01)00391-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
von Heijne, G. (2006). Membrane-protein topology. Nat. Rev. Mol. Cell. Biol. 7, 909–918. doi: 10.1038/nrm2063
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wagner, M. L., and Tamm, L. K. (2001). Reconstituted syntaxin1a/SNAP25 interacts with negatively charged lipids as measured by lateral diffusion in planar supported bilayers. Biophys. J. 81, 266–275. doi: 10.1016/S0006-3495(01)75697-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wallach, D. F., and Zahler, P. H. (1968). Infrared spectra of plasma membrane and endoplasmic reticulum of Ehrlich ascites carcinoma. Biochim. Biophys. Acta 150, 186–193. doi: 10.1016/0005-2736(68)90162-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wang, Y.-H., Collins, A., Guo, L., Smith-Dupont, K. B., Gai, F., Svitkina, T., et al. (2012). Divalent cation-induced cluster formation by polyphosphoinositides in model membranes. J. Am. Chem. Soc. 134, 3387–3395. doi: 10.1021/ja208640t
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wang, Y.-H., Slochower, D. R., and Janmey, P. A. (2014). Counterion-mediated cluster formation by polyphosphoinositides. Chem. Phys. Lipids 182, 38–51. doi: 10.1016/j.chemphyslip.2014.01.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watanabe, M., Nomura, K., Ohyama, A., Ishikawa, R., Komiya, Y., Hosaka, K., et al. (2005). Myosin-Va regulates exocytosis through the submicromolar Ca2+-dependent binding of syntaxin-1A. Mol. Biol. Cell 16, 4519–4530. doi: 10.1091/mbc.E05-03-0252
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watanabe, S., Lehmann, M., Hujber, E., Fetter, R. D., Richards, J., Söhl-Kielczynski, B., et al. (2014a). Nanometer-resolution fluorescence electron microscopy (nano-EM) in cultured cells. Methods Mol. Biol. 1117, 503–526. doi: 10.1007/978-1-62703-776-1_22
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watanabe, S., Liu, Q., Davis, M. W., Hollopeter, G., Thomas, N., Jorgensen, N. B., et al. (2013a). Ultrafast endocytosis at Caenorhabditis elegans neuromuscular junctions. Elife 2:e00723. doi: 10.7554/eLife.00723
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watanabe, S., Rost, B. R., Camacho-Pérez, M., Davis, M. W., Söhl-Kielczynski, B., Rosenmund, C., et al. (2013b). Ultrafast endocytosis at mouse hippocampal synapses. Nature 504, 242–247. doi: 10.1038/nature12809
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watanabe, S., Trimbuch, T., Camacho-Pérez, M., Rost, B. R., Brokowski, B., Söhl-Kielczynski, B., et al. (2014b). Clathrin regenerates synaptic vesicles from endosomes. Nature 515, 228–233. doi: 10.1038/nature13846
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wen, P. J., Osborne, S. L., Zanin, M., Low, P. C., Wang, H.-T. A., Schoenwaelder, S. M., et al. (2011). Phosphatidylinositol(4,5)bisphosphate coordinates actin-mediated mobilization and translocation of secretory vesicles to the plasma membrane of chromaffin cells. Nat. Commun. 2, 491. doi: 10.1038/ncomms1500
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wienisch, M., and Klingauf, J. (2006). Vesicular proteins exocytosed and subsequently retrieved by compensatory endocytosis are nonidentical. Nat. Neurosci. 9, 1019–1027. doi: 10.1038/nn1739
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wilhelm, B. G., Mandad, S., Truckenbrodt, S., Kröhnert, K., Schäfer, C., Rammner, B., et al. (2014). Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science 344, 1023–1028. doi: 10.1126/science.1252884
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wilkins, M. H., Blaurock, A. E., and Engelman, D. M. (1971). Bilayer structure in membranes. Nature 230, 72–76.
Willig, K. I., Rizzoli, S. O., Westphal, V., Jahn, R., and Hell, S. W. (2006). STED microscopy reveals that synaptotagmin remains clustered after synaptic vesicle exocytosis. Nature 440, 935–939. doi: 10.1038/nature04592
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wilson, B. S., Steinberg, S. L., Liederman, K., Pfeiffer, J. R., Surviladze, Z., Zhang, J., et al. (2004). Markers for detergent-resistant lipid rafts occupy distinct and dynamic domains in native membranes. Mol. Biol. Cell 15, 2580–2592. doi: 10.1091/mbc.E03-08-0574
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Woronowicz, K., Dilks, J. R., Rozenvayn, N., Dowal, L., Blair, P. S., Peters, C. G., et al. (2010). The platelet actin cytoskeleton associates with SNAREs and participates in α-granule secretion. Biochemistry 49, 4533–4542. doi: 10.1021/bi100541t
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Xie, Z., McMillan, K., Pike, C. M., Cahill, A. L., Herring, B. E., Wang, Q., et al. (2013). Interaction of anesthetics with neurotransmitter release machinery proteins. J. Neurophysiol. 109, 758–767. doi: 10.1152/jn.00666.2012
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yang, X., Xu, P., Xiao, Y., Xiong, X., and Xu, T. (2006). Domain requirement for the membrane trafficking and targeting of syntaxin 1A. J. Biol. Chem. 281, 15457–15463. doi: 10.1074/jbc.M513246200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yoshimori, T., Keller, P., Roth, M. G., and Simons, K. (1996). Different biosynthetic transport routes to the plasma membrane in BHK and CHO cells. J. Cell Biol. 133, 247–256. doi: 10.1083/jcb.133.2.247
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zaccai, G. (2000). How soft is a protein? A protein dynamics force constant measured by neutron scattering. Science 288, 1604–1607. doi: 10.1126/science.288.5471.1604
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhou, P., Bacaj, T., Yang, X., Pang, Z. P., and Südhof, T. C. (2013). Lipid-anchored SNAREs lacking transmembrane regions fully support membrane fusion during neurotransmitter release. Neuron 80, 470–483. doi: 10.1016/j.neuron.2013.09.010
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zilly, F. E., Halemani, N. D., Walrafen, D., Spitta, L., Schreiber, A., Jahn, R., et al. (2011). Ca2+ induces clustering of membrane proteins in the plasma membrane via electrostatic interactions. EMBO J. 30, 1209–1220. doi: 10.1038/emboj.2011.53
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zilly, F. E., Sørensen, J. B., Jahn, R., and Lang, T. (2006). Munc18-bound syntaxin readily forms SNARE complexes with synaptobrevin in native plasma membranes. PLoS Biol 4:e330. doi: 10.1371/journal.pbio.0040330
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: SNARE proteins, clustering, membrane domains, hydrophobic interactions, ionic interactions, modularity of cluster formation
Citation: Milovanovic D and Jahn R (2015) Organization and dynamics of SNARE proteins in the presynaptic membrane. Front. Physiol. 6:89. doi: 10.3389/fphys.2015.00089
Received: 11 January 2015; Paper pending published: 03 February 2015;
Accepted: 05 March 2015; Published: 19 March 2015.
Edited by:
Annemiek Van Spriel, Radboud University Nijmegen Medical Centre, NetherlandsReviewed by:
Josep Rizo, University of Texas Southwestern Medical Center, USAJoseph Szule, Texas A&M University, USA
Copyright © 2015 Milovanovic and Jahn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Reinhard Jahn, Department of Neurobiology, Max Planck Institute for Biophysical Chemistry, Am Fassberg 11, 37077 Göttingen, Germany rjahn@gwdg.de