 Cun Lin1
Cun Lin1- 1Faculty of Environmental Engineering and Graduate School of Environmental Engineering, The University of Kitakyushu, Kitakyushu, Japan
- 2International Photosynthesis Industrialization Research Center, The University of Kitakyushu, Kitakyushu, Japan
- 3University of Florence LINV Kitakyushu Research Center, Kitakyushu, Japan
- 4International Plant Neurobiology Laboratory, University of Florence, Sesto Fiorentino, Italy
- 5Institut des Energies de Demain (FRE3597), Université Paris Diderot, Sorbonne Paris Cité, Paris, France
- 6Université Paris Diderot, Sorbonne Paris Cité, Paris 7 Interdisciplinary Energy Research Institute, Paris, France
Al3+ toxicity in growing plants is considered as one of the major factors limiting the production of crops on acidic soils worldwide. In the last 15 years, it has been proposed that Al3+ toxicity are mediated with distortion of the cellular signaling mechanisms such as calcium signaling pathways, and production of cytotoxic reactive oxygen species (ROS) causing oxidative damages. On the other hand, zinc is normally present in plants at high concentrations and its deficiency is one of the most widespread micronutrient deficiencies in plants. Earlier studies suggested that lack of zinc often results in ROS-mediated oxidative damage to plant cells. Previously, inhibitory action of Zn2+ against lanthanide-induced superoxide generation in tobacco cells have been reported, suggesting that Zn2+ interferes with the cation-induced ROS production via stimulation of NADPH oxidase. In the present study, the effect of Zn2+ on Al3+-induced superoxide generation in the cell suspension cultures of tobacco (Nicotiana tabacum L., cell-line, BY-2) and rice (Oryza sativa L., cv. Nipponbare), was examined. The Zn2+-dependent inhibition of the Al3+-induced oxidative burst was observed in both model cells selected from the monocots and dicots (rice and tobacco), suggesting that this phenomenon (Al3+/Zn2+ interaction) can be preserved in higher plants. Subsequently induced cell death in tobacco cells was analyzed by lethal cell staining with Evans blue. Obtained results indicated that presence of Zn2+ at physiological concentrations can protect the cells by preventing the Al3+-induced superoxide generation and cell death. Furthermore, the regulation of the Ca2+ signaling, i.e., change in the cytosolic Ca2+ ion concentration, and the cross-talks among the elements which participate in the pathway were further explored.
Introduction
Aluminum is the most abundant metal and the third most abundant chemical element in the Earth’s crust. The increase in free aluminum ions (chiefly, Al3+) accompanying soil acidification is considered to be toxic to plants (Poschenrieder et al., 2008) and animals (Markich et al., 2002). Al3+ toxicity in growing plants is considered as one of the major factors limiting the production of crops on acidic soils worldwide (Poschenrieder et al., 2008; Panda et al., 2009).
A number of studies documented the toxic impact of Al3+ especially on roots (Le Van et al., 1994; Lukaszewski and Blevins, 1996; Sanzonowicz et al., 1998). It has been proposed that early effects of Al3+ toxicity at growing root apex, such as those on cell division, cell extension or nutrient transport, involve the binding to (Ma et al., 1999) or uptake of Al3+ by plants (Lazof et al., 1994; Babourina and Rengel, 2009). Accordingly, actin cytoskeleton and vesicle trafficking are primary targets for Al3+ toxicity in the root tips of the sensitive variety (Amenós et al., 2009).
In the last 15 years, it has been proposed that Al3+ toxicity are mediated with distortion of the cellular signaling mechanisms such as calcium signaling pathways (Kawano et al., 2003a, 2004; Rengel and Zhang, 2003; Lin et al., 2005, 2006a), and production of cytotoxic reactive oxygen species (ROS) causing oxidative damages (Yamamoto et al., 2002; Kawano et al., 2003a). Recently, Al3+-induced DNA damages in the root cells of Allium cepa was shown to be blocked by calcium channel blockers suggesting that Al3+-stimulated influx of extracellular Ca2+ into cytosol causes the programmed cell death-like decomposition of DNA (Achary et al., 2013).
To date, two independent groups have proposed the likely modes of ROS production in Al3+-treated plant cells. While Yamamoto et al. (2002), propounded the role of mitochondria challenged by Al3+ using the cultured cells of tobacco (Nicotiana tabacum L., cell line SL) and the roots of pea (Pisum sativum L.); our group (Kawano et al., 2003a) emphasized the involvement of NADPH oxidase, thus sensitive to an inhibitor of NADPH oxidase, diphenylene iodonium (DPI) in tobacco BY-2 cells. While ROS is slowly produced through mitochondrial dysfunction (ca. 12 h after Al3+ treatment; Yamamoto et al., 2002), the NADPH oxidase-mediated production of superoxide anion radical (O2•–) takes place immediately after Al3+ treatment (Kawano et al., 2003a).
The action of Al3+ for induction of O2•– generation which is sensitive to DPI was recently confirmed in the cells of Arabidopsis thaliana (Kunihiro et al., 2011). Furthermore, the Al3+-induced oxidative burst showed biphasic signature consisted with an acute transient spike and a slow but long-lasting wave of O2•– generation. In addition, among six respiratory burst oxidase homologs (Atrbohs) coding for plant NADPH oxidase, solely AtrbohD was shown to be responsive to Al3+ in biphasic manner by showing rapid (1 min) and long-lasting (24 h) expression profiles (Kunihiro et al., 2011).
Interestingly, the mechanism of Al3+-induced oxidative burst (production of O2•–) is highly analogous to the response of tobacco cell suspension culture to other metal cations, chiefly trivalent cations of lanthanides such as La3+ and Gd3+ (Kawano et al., 2001). Therefore, we assume that some known chemical factors reportedly interfere with the lanthanide-induced plant oxidative burst might be active for protection of plant cells from Al3+-induced oxidative stress. Such chemicals of interest to be tested include zinc and manganese (Kawano et al., 2002).
Zinc is normally present in plants at high concentrations (Santa-Maria and Cogliatti, 1988) and its deficiency is one of the most widespread micronutrient deficiencies in plants, causing severe reductions in crop production (Cakmak, 2000). Increasing studies indicate that oxidative damage to cellular components caused in plants being challenged by ROS, is highly due to the deficiency of zinc (Pinton et al., 1994; Cakmak, 2000).
Previously, inhibitory action of Zn2+ against lanthanide-induced O2•– generation in tobacco cells have been reported (Kawano et al., 2002). Pretreatments with Zn2+ reportedly interferes the La3+- and Gd3+-induced O2•– generation in tobacco cells. In the tobacco model, Zn2+ was shown to minimize the earlier phase of lanthanide-induced O2•– production while allowing the release of O2•– in the later phase, thus causing the retardation of the lanthanide actions on O2•– generation.
Although this process is well known, if it is preserved in higher plants and the specific mechanism of action have is not still clear. For this reason, in the present study, effect of Zn2+ on Al3+-induced O2•– generation in the suspension cultures of tobacco BY-2 cells and rice (Oryza sativa L., cv. Nipponbare) cells, was examined. Furthermore, the regulation of the Ca2+ signaling, i.e., change in the cytosolic Ca2+ ion concentration ([Ca2+]c), and the cross-talks among the elements which participate in the pathway were further explored. Finally the possible use of Zn2+ for protection of plant cells from Al3+ toxicity is discussed.
Materials and Methods
Chemicals
O2•–-specific chemiluminescence (CL) probe, Cypridina luciferin analog (CLA; 2-methyl-6-phenyl-3,7-dihydroimidazo[1,2-a]pyrazin-3-one) designated as CLA was purchased from Tokyo Kasei Kogyo Co. (Tokyo, Japan). Aluminum (III) chloride hexahydrate (AlCl3·6H20), zinc sulfate heptahydrate (ZnSO4·7H2O), gadolinium chloride hexahydrate (GdCl3·6H2O), and salicylic acid (SA) were from Wako Pure Chemical Industries (Osaka, Japan). Lanthanum chloride heptahydrate (LaCl3·7H2O) was from Kanto Chemical Co., Inc (Tokyo, Japan). DPI chloride, Evans blue, 4,5-dihydroxy-1,3-benzene-disulfonic acid (Tiron), N,N′-dimethylthiourea (DMTU), were from Sigma (St. Louis, MO, USA). Coelenterazine was a gift from Prof. M. Isobe (Nagoya University).
Plant Cell Culture
Tobacco (Nicotiana tabacum L. cv. Bright Yellow-2) suspension-culture cells (cell line, BY-2, expressing the aequorin gene; Figure 1A) were propagated as previously described (Kawano et al., 1998). Briefly, the culture was maintained in Murashige–Skoog liquid medium (pH 5.8) supplemented with 3% (w/v) sucrose and 0.2 μg ml–1 of 2,4-dichlorophenoxyacetic acid. The culture was propagated with shaking on a gyratory shaker in darkness at 23°C. For sub-culturing, 1.0 ml of confluent stationary culture was suspended in 30 ml of fresh culture medium and incubated at 23°C with shaking at 130 rpm on a gyratory shaker in darkness until used.
Figure 1. Microscopic images of plant cells used in this study. (A) Tobacco BY-2 cells. (B) Rice M1 cells. Scale bars, 50 μm.
Rice callus tissues (Oryza sativa L., cv. Nipponbare, cell line, M1; Figure 1B) were obtained from root explants derived from young seedlings and transferred in AA liquid medium to develop a suspension-culture. The cells were maintained and propagated at 23°C with shaking at 130 rpm on a gyratory shaker in darkness. For sub-culturing, with 2-week intervals, 10 ml of stationary culture was suspended in 100 ml of fresh culture medium.
Detection of O2•– with CLA
To detect the production of O2•– in plant cells, the 200 μl of plant cell suspension (either of tobacco or rice) was placed in glass cuvettes and CLA was added at final concentration of 2 μM (in tobacco cells) and 4 μM (in rice cells). The glass cuvettes containing 200 μl of plant cell suspension were placed in luminometers (CHEM-GLOW Photometer, American Instrument Co., Silver Spring, MD, USA; or PSN AB-2200-R Luminescensor, Atto, Tokyo). Generation of O2•– in cell suspension culture was monitored by CLA-CL, and expressed as relative chemiluminescence units (rcu) as previously described (Kawano et al., 1998). CLA-CL specifically indicates the generation of O2•– (and of 1O2 with a minor extent) but not that of O3, H2O2 or hydroxy radicals (Nakano et al., 1986).
Aequorin Ca2+ Detection
To detect the changes in [Ca2+]c in tobacco cells, 10 mL of plant cell suspension were pre-treated for 8 h with 10 μL of coelenterazine in the dark, then used for the experiments as previously described (Kawano et al., 1998). Also in this case, 200 μl plant of cell suspension was placed in glass cuvettes and placed in luminometers (as above). Increase in [Ca2+]c, reflecting the induced Ca2+ into cells, was monitored as luminescence derived upon binding of Ca2+ to aequorin (the recombinant gene over-expressed in the cytosol) and expressed as rcu.
Treatments with Aluminum, Zinc, and Other Stimuli
Tobacco BY-2 cells were harvested various days after sub-culturing (as indicated), and used for experiments with CLA or aequorin. AlCl3 was dissolved in distilled water and diluted with fresh culture media unless indicated, and 10–20 μl of the AlCl3 solution was added to 180–190 μl of cell suspension in glass cuvettes, and level of [Ca2+]c (aequorin experiment) or generation of O2•– (CLA experiment) were monitored. For comparison, effects of SA and hypo-osmotic shock (induced by dilution of media giving Δ100 mOsmol of hypo-osmolarity difference) on calcium homeostasis with and without zinc was monitored. Inhibition of events induced by Al3+ and other stimuli, monitored with CLA CL, aequorin luminescence, and cell death staining, was performed by addition of indicated concentration of Zn2+ to the cells prior to treatments with Al3+, SA, and hypo-osmotic shock.
Monitoring of Cell Death
Al3+-induced cell death in BY-2 tobacco cell suspension culture was allowed to develop in the presence of Evans blue, a lethal staining dye (0.1%, w/v). Evans blue was added to the cell suspension culture, 6 h after Al3+ application unless indicated or at the time indicated (0–8 h after Al addition). Then, the cells were further incubated for 1 h for fully developing and detecting the cell death as described (Kadono et al., 2006). After terminating the staining process by washing, stained cells were counted under microscopes. For statistical analyses, four different digital images of cells under the microscope (each covering 50 cells to be counted) were acquired and stained cells were counted.
Results and Discussions
Induction of O2•–
The effect of Al3+ concentration on induction of O2•– generation has been tested both in tobacco and rice cell suspension cultures (Figure 2). For this analysis, BY-2 tobacco cells have been tested 4 days after inoculation (DAI) unless indicated whereas the rice cell line M1 suspension culture was used 14 DAI, since the tobacco BY-2 cells grow at faster rate compared to rice M1 cells. In tobacco BY-2 cells, the active Al3+ concentrations for induction of O2•– generation ranged from 0.1 mM to 30 mM (optimally at 3 mM).
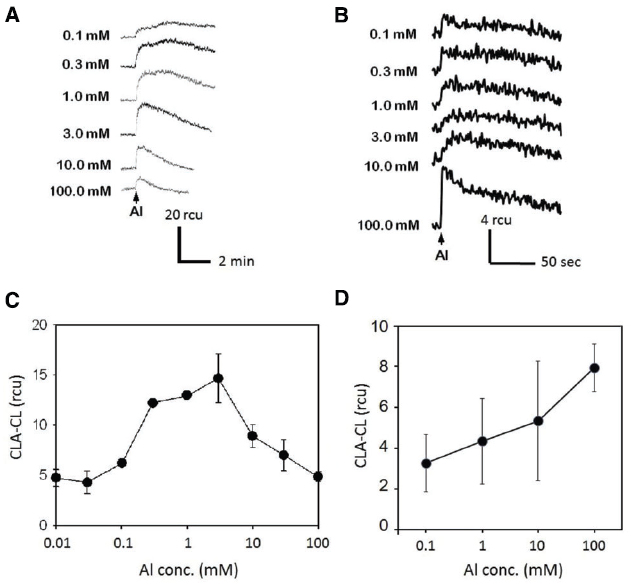
Figure 2. Effect of Al3+ concentration on O2•– induction. Typical records of Al3+-induced O2•– measured with CLA-CL in the cell suspension cultures of tobacco (A) and rice (B). Effect of Al3+ concentration on O2•– generation (C, tobacco; D, rice). Vertical error bars, SD; n = 3.
Notably, higher concentration of Al3+ was shown to be inhibitory to induction of O2•– generation in the tobacco cells (Figures 2A,C), while the rice cells showed only the proportional increase in generation of O2•– with the increase in Al3+ up to 100 mM (Figures 2B,D). In order to analyze the impact of Zn2+ against Al3+-induced O2•– generation, the concentration of Al3+ was fixed to at 3 mM for the tobacco cells and 100 mM for rice cells. Different concentrations have been chosen since the tobacco cells showed higher sensitivity to relatively lower concentrations of Al3+.
Effect of Pretreatment with Zn2+
To assess the effect of Zn2+, the cells of tobacco and rice were pre-treated with various concentration of ZnSO4 for 5 min and then AlCl3 was added to the cells (Figure 3). In tobacco cell, the O2•– generation induced by 3 mM Al3+ was significantly inhibited by 1 mM or higher concentrations of Zn2+, whilst in rice cell, 0.1 mM of Zn2+ was high enough to achieve a significant inhibition of O2•– generation induced by 100 mM Al3+. Although Zn2+-dependent retardation of lanthanide-induced O2•– production has been reported (Kawano et al., 2002), the Al3+-induced oxidative burst was simply inhibited without allowing the onset of slower increase in O2•– production.
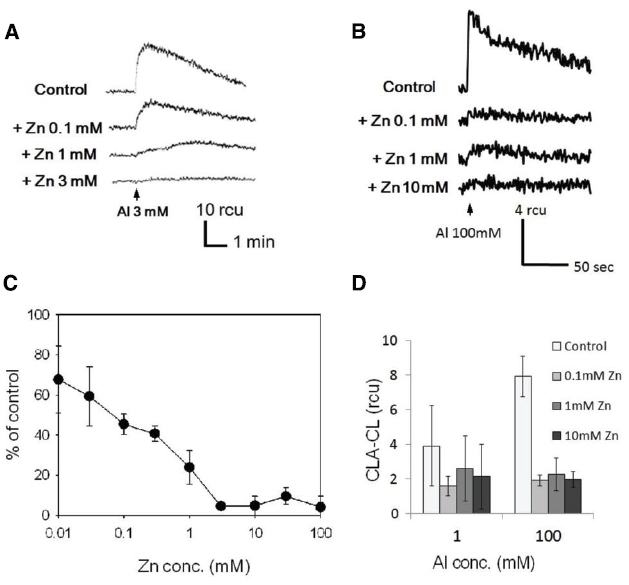
Figure 3. Inhibition of the Al3+-induced generation of O2•– in the presence of Zn2+. Typical records of Al3+-induced O2•– generation in the absence and the presence of ZnSO4 measured with CLA (A, tobacco; B, rice). Arrows indicate the time points for addition of AlCl3 (3 mM, tobacco; 100 mM, rice). Effects of Zn2+ concentration on Al3+-induced O2•– generation (C, tobacco; D, rice). Vertical error bars, SD; n = 3.
The Zn2+-dependent inhibition was observed in both model cells selected from the monocots and dicots (rice and tobacco), suggesting that this phenomenon (Al3+/Zn2+ interaction) can be observed universally in the wide range of higher plants. Since the sensitivity was higher in tobacco BY-2 cells, this cell line was chosen to be used in the further experiments examining the mode of Al3+/Zn2+ interaction.
Effect of Culture Age on O2•– Production
Prior to treatment with Al3+, tobacco BY-2 cell suspension culture was aged for 1, 3, 5, 8, and 13 DAI of the fresh media (30 ml) with 0.5 ml of confluent culture (at 10 DAI). The cultures at 1 and 3 DAI were smooth and colorless. The 4 and 5 DAI cultures were also smooth but colored slightly yellowish. The 8 and 12 DAI cultures were highly dense and colored yellow. The growth of the culture was assessed by measuring the changes in fresh cell weight at each time point. Figure 4A shows a typical growth curve for tobacco BY-2 cell culture. Effect of culture age of tobacco BY-2 cells on the sensitivity to Al3+ was examined using the differently aged cultures (Figure 4B), and the high sensitivity to Al3+ was observed in 2 and 4 DAI of tobacco BY-2 cells.
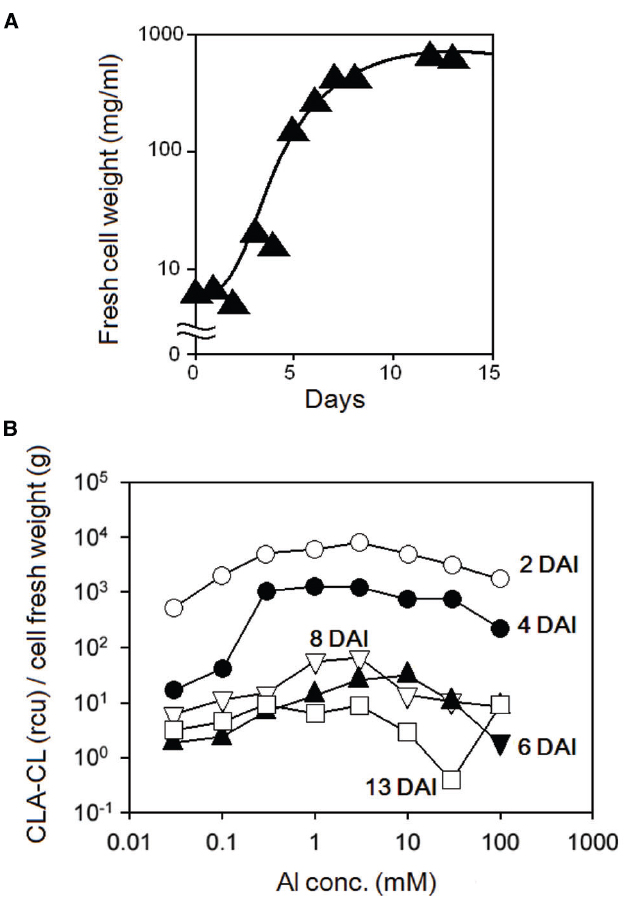
Figure 4. Effect of culture age on Al3+-induced O2•– generation in tobacco cell suspension culture. (A) Growth curve for tobacco cells. (B) Four different patterns of CLA-CL reflecting the Al3+-induced O2•– generation.
Competition Between Zn2+ and Al3+
Application of double-reciprocal plot analysis for studying the behavior of living plants or cells, so-called in vivo Lineweaver–Burk plot analysis was carried out to assess the mode of Al3+/Zn2+ interaction according to the procedure described elsewhere (Kawano et al., 2003b). By making use of linear dose-dependency in the limited range of Al3+ concentrations (up to 3 mM) in 4 DAI culture of tobacco BY-2 cell, the in vivo kinetic analysis was carried out by assuming Al3+ as a ligand to the putative Al3+ receptors on the cells and Zn2+ as an inhibitor (Figure 5A). The reciprocals of the CLA-CL yields (1/CLA-CL) were plotted against the reciprocals of Al3+ concentrations (1/[Al3+]). Linear relationship between 1/CLA-CL and 1/[Al3+] were obtained both in the presence and absence of Zn2+ (Figure 5B). In the presence of Zn2+, the apparent Km for Al3+ was elevated from 113 μM (control) to 376 μM (0.1 mM Zn2+; ca. 3.3-fold increase), while Vmax for Al3+-induced CLA-CL was not drastically altered. Vmax for Al3+-induced response in the absence of Zn2+ was calculated to be 14.3 rcu. In the presence of 0.1 mM Zn2+, Vmax was 18.6 rcu (ca. 30% increase). Therefore, the mode of Zn2+ action against Al3+ can be considered as a typical competitive inhibition.
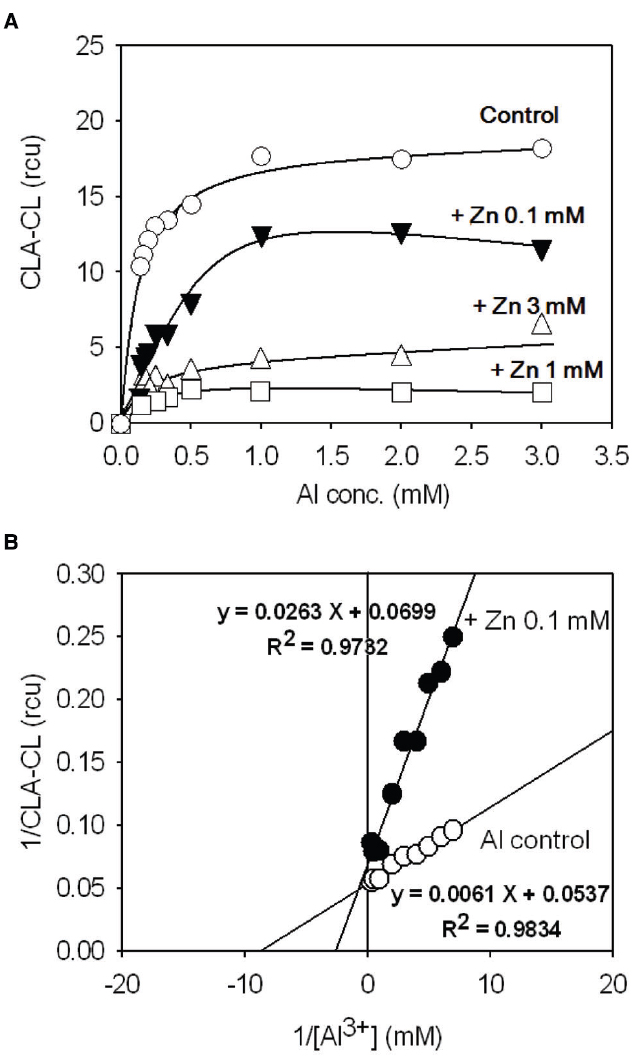
Figure 5. Competitive inhibition of the Al3+-induced O2•– generation by Zn2+ in tobacco cell suspension culture. (A) Effect of Zn2+ on Al3+-induced O2•– generation. (B) In vivo Lineweaver–Burk plot analysis.
According to Kawano et al. (2003a) the Al3+-induced generation of O2•– in tobacco cells is catalyzed by Al3+-stimulated NADPH oxidase which is sensitive to DPI. The cation-dependent enhancement in NADPH oxidase-catalyzed O2•– production is also known in human neutrophils in which binding of metal cations possibly results in spontaneous activation of the O2•–-generating activity of the membrane-bound enzyme (Cross et al., 1999). We can assume that NADPH oxidase itself, localized on the surface of cells (or other factors associated with NADPH oxidase), behaves as the receptor for Al3+ ions. The competitive mode of Zn2+ action against the Al3+-induced oxidative burst suggests us to consider that the binding site for Al3+ and Zn2+ on the NADPH oxidase or on the factors associated nearby must be identical.
Al3+-Induced Cell Death and its Inhibition by Zn2+
As shown in Figures 6A,B, treatment of tobacco BY-2 cells with various concentrations of AlCl3 resulted in cell death induction. Notably, the presence of Zn2+ significantly protected the cells from the induction of cell death by Al3+ (Figure 6C), as predicted by the action of Zn2+ against Al3+-induced oxidative burst.
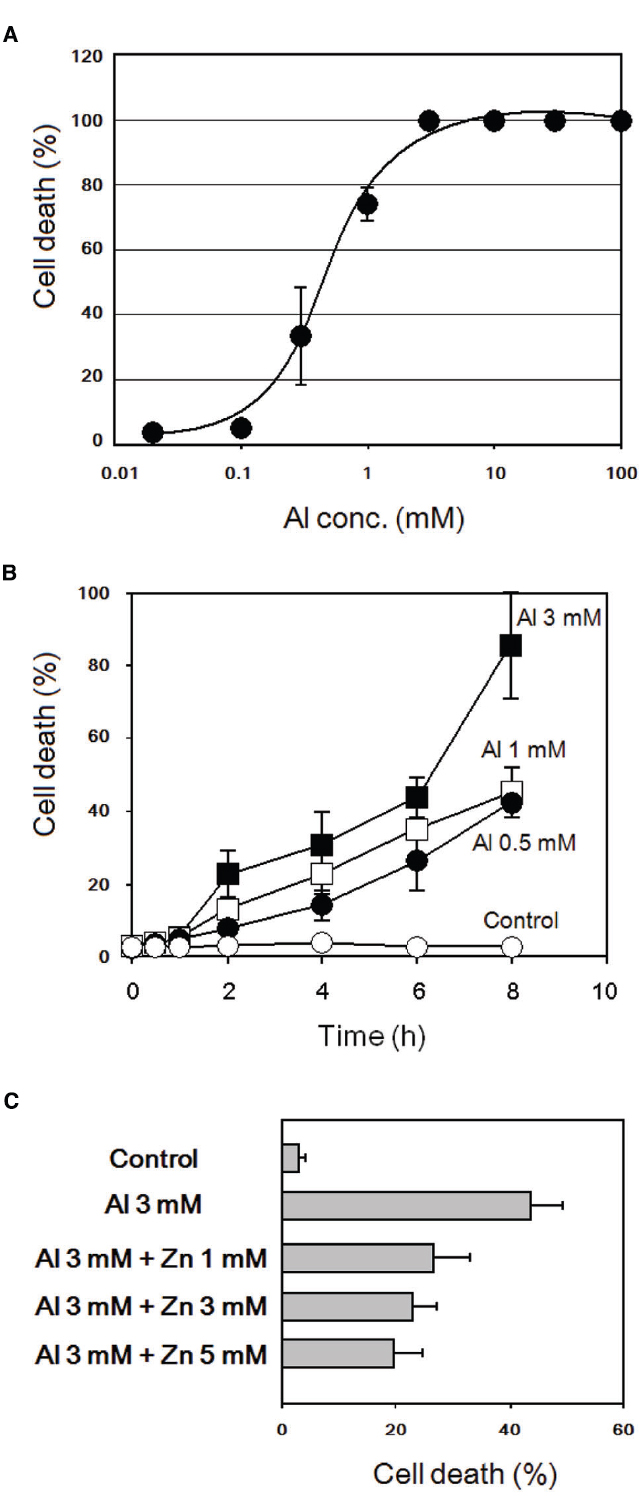
Figure 6. Al3+-induced cell death and its inhibition by addition of Zn2+ in tobacco BY-2 cells. (A) Effect of AlCl3 concentration on cell death induction. Cell death was assessed by Evans blue staining 8 h after addition of AlCl3. (B) Effect of post-Al3+ incubation (0.5–8 h) on development of cell death. (C) Effect of ZnSO4 on Al3+-induced cell death. Cell death was assessed 6 h after Al3+ treatments. Vertical error bars, SD; n = 3.
Effect of Pretreatment with Mn2+
Manganese is another micronutrient possibly protecting the living cells from oxidative damage (Ledig et al., 1991) and reportedly blocks the lanthanide-induced oxidative burst (Kawano et al., 2002). In fact, Mn2+ is often employed as a scavenger of O2•– for preventing the biochemical reactions involving O2•– (Momohara et al., 1990).
Therefore, we tested the effect of MnSO4 (up to 3 mM) for comparison. The results obtained suggested no inhibitory effect of Mn2+ against Al3+-induced generation of O2•–. Instead, low concentrations of Mn2+ slightly elevated the level of Al3+-induced oxidative burst (data not shown). For inhibition of Al3+-induced oxidative burst, much higher concentrations of MnSO4 (10–100 mM) were required. Since the range of Mn2+ concentrations required for lowering the level of Al3+-induced generation of O2•– was at phytotoxic range (Caldwell, 1989) and thus inducing cell death even in the absence of Al3+ in BY-2 cells (ca. 40% of cells died in the presence of 30 mM MnSO4), the use of Mn2+ is not suitable for preventing the production of O2•– induced by Al3+.
Anti-Oxidative Role for Zn2+
Plants require trace amounts of specific metals known as trace nutrients including Zn2+, supporting the essential functions of plant cells ranging from respiration to photosynthesis, and molecular biological studies on the mechanism for uptake of these metals by plants have been documented (Delhaize, 1996). One of the important roles for Zn2+ in living plants is anti-oxidative action against ROS (Kawano et al., 2002) as the present study successfully demonstrated that extracellular supplementation of Zn2+ inhibits the generation of O2•– (Figure 4) and cell death (Figure 6C) induced by Al3+.
In contrast to manganese, zinc is normally present in plants at high concentrations. For example, in roots of wheat seedlings, the cytoplasmic concentration of total Zn has been estimated to be approximately 0.4 mM (Santa-Maria and Cogliatti, 1988), and Zn-deficiency often results in inhibition of growth, as Zn reportedly protects the plants by preventing the oxidative damages to DNA, membranes, phospholipids, chlorophylls, proteins, SH-containing enzymes, and indole-3-acetic acid (Cakmak, 2000).
Here, Zn2+ at sub-mM concentrations showed strong inhibitory action against the toxicity of Al3+ (oxidative burst and cell death). The levels of Zn2+ naturally present in soil or plant tissues may be contributing to the prevention of Al3+-induced cellular damages but further studies on living plants are needed to evaluate this mechanism in living tissue and the possible applications to increase plant tolerance.
Oxidative and Calcium Crosstalk
Al3+ is known to inhibit plant calcium channels similarly to the action of various lanthanide ions (Lin et al., 2006b). The calcium channels sensitive to Al3+ could be identical to those involved in responses to ROS (Kawano et al., 2003a, 2004), cold shock (Lin et al., 2005, 2006a, 2007), and heat shock (Lin et al., 2007), but not responsive to osmotic shock (Lin et al., 2005, 2006b, 2007), as examined in transgenic cell lines of rice (Oryza sativa L., cv. Nipponbare) and tobacco (cell-lines, BY-2, Bel-B, and Bel-W3) all expressing aequorin in the cytosolic space.
To support the hypothesis that Al3+-induced distortion in [Ca2+]c involves the members of ROS derived from the action of NADPH oxidase, and calcium channel opening leading to transient [Ca2+]c elevation, the effect of DPI (NADPH oxidase inhibitor), ROS scavengers (Tiron, DMTU), and trivalent cations (La3+ and Gd3+) have been tested in tobacco cells expressing aequorin and compared with the antagonistic action of zinc protecting the cells (Figure 7A and inset).
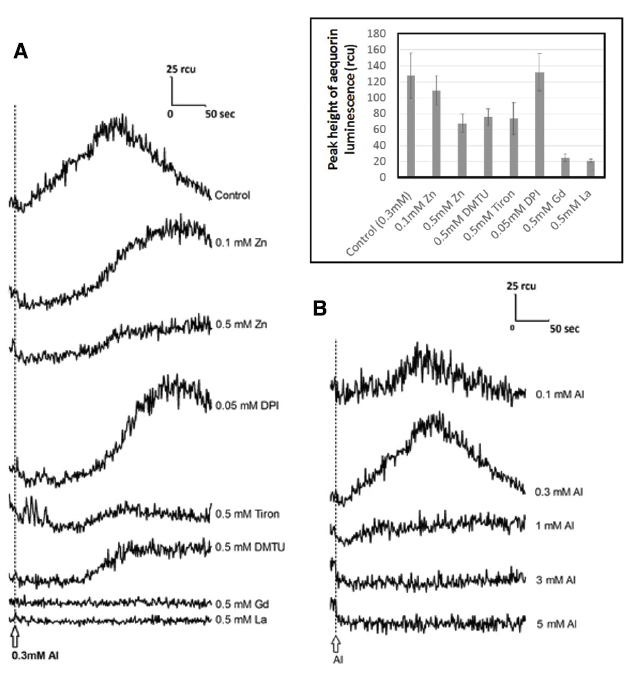
Figure 7. Monitoring of Al3+-responsive [Ca2+]c with aequorin luminescence in the presence and absence of various inhibitors. (A) Effect of high- and low-dose Zinc, DPI (NADPH oxidase inhibitor), ROS scavengers (Tiron, DMTU), and trivalent cations (La3+ and Gd3+) on Al-responsive [Ca2+]c elevation in tobacco cell suspension culture expressing aequorin. (Inset) Quantitative comparison of the action of inhibitors against the peak height of Al-responsive [Ca2+]c elevation (n = 3; error bars, SD). (B) Effect of Al3+ concentration on induction of [Ca2+]c elevation. Typical records of Ca2+-responsive aequorin luminescence measured in the presence of different Aluminum concentrations are shown.
As expected, Tiron, DMTU, and high concentration of zinc (0.5 mM) effectively lowered the level of Al3+-induced [Ca2+]c elevation. Especially, temporal patterns in which Al3+ induces an increase in [Ca2+]c was shown to be sensitive to both zinc and DPI. In fact, these chemicals significantly retarded the Al3+-responsive calcium influx, thus, time required for attaining the peak of Al3+-responsive [Ca2+]c elevation was shown to be longer, suggesting the zinc and DPI might share the common mode of action.
On the other hand, La3+ and Gd3+ strongly reduce the signal as we observed for high concentration of Al3+ (Figure 7A) supporting the view that they can concurrently act inhibiting the Ca2+ channel.
Previously, we have propose a model that Al3+ plays dual roles acting for and against the Ca2+ influx, by releasing O2•– and by inhibiting the Ca2+ channel(s), respectively (Kawano et al., 2003a). Al3+-dependent distortion in calcium signaling in plant cells can be dissected into two opposing modes of Al3+ actions, viz., (i) stimulation of ROS-responsive calcium channels via induction of O2•– and (ii) inhibition of calcium channels. At low Al3+ concentrations, the ROS-responsive Ca2+ influx potency is high but the driving force (due to ROS) is not sufficient. At high Al3+concentrations, the Ca2+ influx-driving force is at sufficient level but the channel’s Ca2+ permeability is low. This effect is showed in Figure 7B, where [Ca2+]c elevation could be manifested only in the range of Al3+ concentration in which the two opposing effects eventually compromise (Kawano et al., 2003a). Zn2+ hardly blocks the calcium influx in model plant cells unless the event of interest is dependent on the ROS generating events (Figure 8). Therefore, we view here that Zn2+ might target only the earlier phase of Al3+ action involved in induction of O2•– as illustrated in Figure 9.
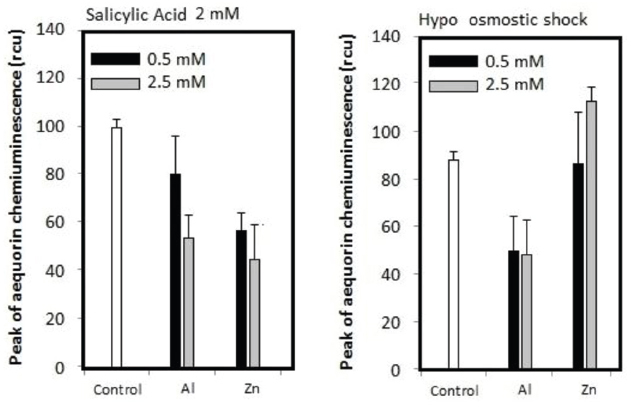
Figure 8. Effect of Al3+ and Zn2+ against SA- responsive and hypo-osmotic shock-responsive [Ca2+]c elevation in tobacco cells. Error bars, SD; n = 3.
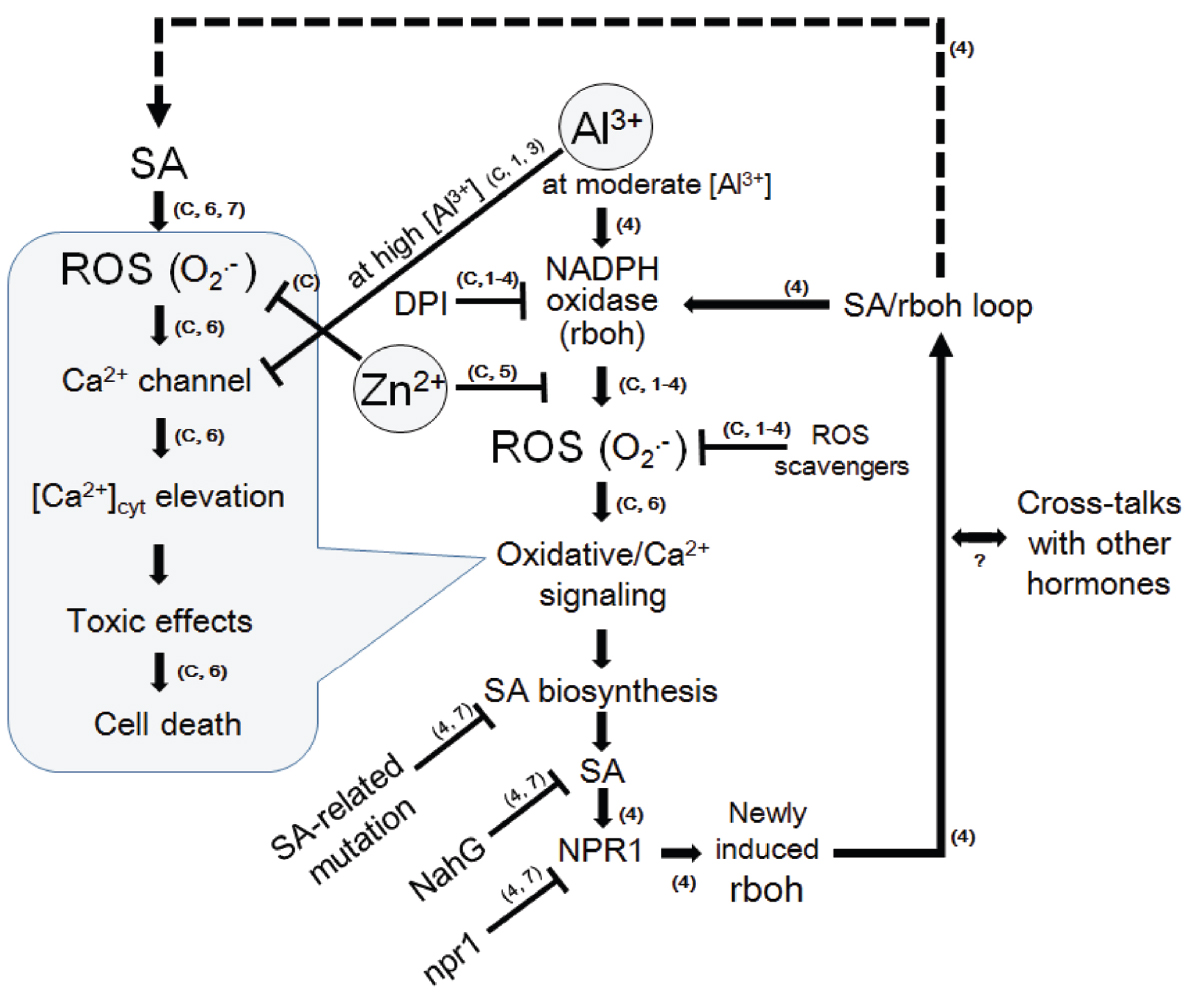
Figure 9. A model for Al3+-induced signaling leading to oxidative cell death and its inhibition by Zn2+ in model plant cells. Knowledge from present experiments using the aequorin-expressing cell lines of tobacco (BY-2) were strengthened by previously reported models. As suggested in the cells of Arabidopsis thaliana (Kunihiro et al., 2011), SA signaling pathway is activated in the downstream of ROS such as O2•– (and possibly of calcium signaling). The loop of ROS-stimulated the SA signaling and expression of NADPH oxidase which produces further ROS, may eventually lead to development of cell death. Zinc may block the loop by interfering the NADPH oxidase-catalyzed ROS production similarly to the model for lanthanide-induced oxidative burst and cell death in tobacco cells (Kawano et al., 2002). Arrows indicate the stimulation and the T-shaped lines indicate the steps inhibited by agent indicated. The numbers and the symbol “C” in small brackets next to arrows or T-shaped lines denote the source of knowledge, namely; C, data confirmed here; 1, Kawano et al. (2003a; BY-2 cells); 2, Kawano et al. (2004; BY-2 cells); 3, Lin et al. (2005; BY-2 cells); 4, Kunihiro et al. (2011; Arabidopsis thaliana); 5, Kawano et al. (2002; BY-2 cells); 6, Kawano et al. (1998; BY-2 cells); 7, Kawano et al. (2013; review on higher plants).
The Likely Signaling Paths
In Arabidopsis thaliana, Al3+-induced prolonged ROS generation requires the expression of AtrbohD coding for NADPH oxidase (Kunihiro et al., 2011). This work suggested that biosynthesis and signal transduction pathway for SA is involved in Al3+-mediated oxidative burst since the Al3+-induced AtrbohD expression and cell death were inhibited in the mutant and transgenic cell lines lacking SA biosynthesis, accumulation of SA, and SA-specific signaling components (sid2, NahG and npr1, respectively). It has been proposed that loop of SA signal transduction, involving the activity and further induction of NADPH oxidase, forms a signaling circuit enabling an amplification of SA-mediated signaling (Figure 9). This type of oxidative signal amplification was designated as SA/rboh loop (Kunihiro et al., 2011).
By analogy, there would be a similar mechanism in response to Al3+ in the cells of tobacco and rice since both the ROS production and cell death were commonly shown to be induced by Al3+ in these cells.
Lastly, we propose a likely mode of Zn2+ action against Al3+-induced cell death. Zn2+ may competitively antagonize the action of Al3+ by targeting the NADPH oxidase-catalyzed ROS production at upstream of SA signaling mechanism. As a consequence, activation of SA/rboh loop responsible for long-lasting oxidative burst releasing cytotoxic ROS could be prevented (Figure 9).
By assessing the action of Zn2+ against SA-induced [Ca2+]c elevation which is known to be one of the key events in the SA-induced O2•–-mediated signaling path, involving the activation of Ca2+ channel identified as TPC1 channel (Kawano et al., 2013; Lin et al., 2005), we understood that target of antioxidant activity of zinc is not limited to the Al3+-induced NADPH oxidase-catalyzed mechanism (Figure 8). It is known that SA-induced rapid O2•– is catalyzed by extracellular (cell-wall bound) peroxidase, while SA-induced long-lasting oxidative burst requires the induction of rboh genes coding for NADPH oxidase (Kawano et al., 1998; Yoshioka et al., 2008). In contrast, Zn2+ failed to block the Ca2+ influx induced by hypo-osmotic shock possibly involving the mechanosensitive-cation channel (Takahashi et al., 1997).
Taken together, target of Zn2+ is specifically against the ROS-generating mechanisms (both NADPH oxidase-mediated and peroxidase-mediated) eventually leading to ROS-responsive calcium signaling (Figure 9).
Furthermore, the action of Al3+ may form a loop of repeated reaction involving the action of SA which further induces specific type of NADPH oxidase (in case of Arabidopsis thaliana, only AtrbohD is Al3+-responsive, Kunihiro et al., 2011).
Author Contributions
TK designed and supervised the experiments and some key data for plant age and ROS production were obtained by him. CL conducted most tobacco experiments (mostly calcium signaling and cell viability tests), AH and DC performed additional experiments. AH was in charge of rice cell experiments (both ROS detection and calcium signaling). DC and FB contributed on the analysis of data and writing of MS. All authors actively contributed in the discussion.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This work was supported by Regional Innovation Cluster Program by Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.
Abbreviations
CL, chemiluminescence; CLA, Cypridina luciferin analog; DAI, days after inoculation; DPI, diphenylene iodonium; O2•–, superoxide anion radical; rcu, relative chemiluminescence units; ROS, reactive oxygen species; SA, salicylic acid.
References
Achary, V. M. M., Parinandi, N. L., and Panda, B. B. (2013). Calcium channel blockers protect against aluminium-induced DNA damage and block adaptive response to genotoxic stress in plant cells. Mutat. Res. 751, 130–138. doi: 10.1016/j.mrgentox.2012.12.008
Amenós, M., Corrales, I., Poschenrieder, C., Illéš, P., Baluška, F., and Barceló, J. (2009). Different effects of aluminum on the actin cytoskeleton and brefeldin A-sensitive vesicle recycling in root apex cells of two maize varieties differing in root elongation rate and aluminum tolerance. Plant Cell Physiol. 50, 528–540. doi: 10.1093/pcp/pcp013
Babourina, O., and Rengel, Z. (2009). Uptake of aluminium into Arabidopsis root cells measured by fluorescent lifetime imaging. Ann. Bot. 104, 189–195. doi: 10.1093/aob/mcp098
Cakmak, I. (2000). Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 146, 185–205. doi: 10.1046/j.1469-8137.2000.00630.x
Caldwell, C. R. (1989). Analysis of aluminum and divalent cation binding to wheat root plasma membrane proteins using terbium phosphorescence. Plant Physiol. 91, 233–241. doi: 10.1104/pp.91.1.233
Cross, A. R., Erichson, R. W., Eliss, B. A., and Curnutte, J. T. (1999). Spontaneous activation of NADPH oxidase in a cell-free system: unexpected multiple effects of magnesium ion concentrations. Biochem. J. 338, 229–233. doi: 10.1042/bj3380229
Delhaize, E. (1996). A metal-accumulator mutant of Arabidopsis thaliana. Plant Physiol. 111, 849–855. doi: 10.1104/pp.111.3.849
Kadono, T., Yamaguchi, Y., Furuichi, T., Hirono, M., Garrec, J. P., and Kawano, T. (2006). Ozone-induced cell death mediated with oxidative and calcium signaling pathways in tobacco Bel-W3 and Bel-B cell suspension cultures. Plant Signal. Behav. 1, 312–322. doi: 10.4161/psb.1.6.3518
Kawano, T., Hiramatsu, T., and Bouteau, F. (2013). “Signaling role of salicylic acid in abiotic stress responses in plants,” in Salicylic Acid, eds S. Hayat, A. Ahmad, and M. N. Alyemeni (Dordrecht: Springer), 249–275.
Kawano, T., Kadono, T., Fumoto, K., Lapeyrie, F., Kuse, M., Isobe, M., et al. (2004). Aluminum as a specific inhibitor of plant TPC1 Ca2+ channels. Biochem. Biophys. Res. Commun. 324, 40–45. doi: 10.1016/j.bbrc.2004.09.015
Kawano, T., Kadono, T., Furuichi, T., Muto, S., and Lapeyrie, F. (2003a). Aluminum-induced distortion in calcium signaling involving oxidative bursts and channel regulations in tobacco BY-2 cells. Biochem. Biophys. Res. Commun. 308, 35–42. doi: 10.1016/S0006-291X(03)01286-5
Kawano, N., Kawano, T., and Lapeyrie, F. (2003b). Inhibition of the indole-3-acetic acid-induced epinastic curvature in tobacco leaf strips by 2,4-dichlorophenoxyacetic acid. Ann. Bot. 91, 456–471. doi: 10.1093/aob/mcg043
Kawano, T., Kawano, N., Muto, S., and Lapeyrie, F. (2001). Cation-induced superoxide generation in tobacco cell suspension culture is dependent on ion valence. Plant Cell Environ. 24, 1235–1241. doi: 10.1046/j.1365-3040.2001.00766.x
Kawano, T., Kawano, N., Muto, S., and Lapeyrie, F. (2002). Retardation and inhibition of the cation-induced superoxide generation in BY-2 tobacco cell suspension culture by Zn2+ and Mn2+. Physiol. Plant. 114, 395–404. doi: 10.1034/j.1399-3054.2002.1140309.x
Kawano, T., Sahashi, N., Takahashi, K., Uozumi, N., and Muto, S. (1998). Salicylic acid induces extracellular superoxide generation followed by an increase in cytosolic calcium ion in tobacco suspension culture: the earliest events in salicylic acid signal transduction. Plant Cell Physiol. 39, 721–730. doi: 10.1093/oxfordjournals.pcp.a029426
Kunihiro, S., Hiramatsu, T., and Kawano, T. (2011). Involvement of salicylic acid signal transduction in aluminum-responsive oxidative burst in Arabidopsis thaliana cell suspension culture. Plant Signal. Behav. 6, 611–616. doi: 10.4161/psb.6.5.14895
Lazof, D. B., Goldsmith, J. G., Rufty, T. W., and Linton, R. W. (1994). Rapid uptake of aluminum into cells of intact soybean root tips. A microanalytical study using secondary ion mass spectrometry. Plant Physiol. 106, 1107–1114.
Ledig, M., Tholey, G., Megias-Megias, L., Kopp, P., and Wedler, F. (1991). Combined effects of ethanol and manganese on cultured neurons and glia. Neurochem. Res. 16, 591–596. doi: 10.1007/BF00974879
Le Van, H., Kuraishi, S., and Sakurai, N. (1994). Aluminum-induced rapid root inhibition and changes in cell-wall components of squash seedlings. Plant Physiol. 106, 971–976.
Lin, C., Kadono, T., Suzuki, T., Yoshizuka, K., Furuichi, T., Yokawa, K., et al. (2006a). Mechanism for temperature-shift-responsive acute Ca2+ uptake in suspension-cultured tobacco and rice cells. Cryobiol. Cryotechnol. 52, 83–89.
Lin, C., Kadono, T., Yoshizuka, K., Furuichi, T., and Kawano, T. (2006b). Effects of fifteen rare earth metals on Ca2+ influx in tobacco cells. Z. Naturforsch. 61, 74–80. doi: 10.1515/znc-2006-1-214
Lin, C., Yoshizuka, K., and Kawano, T. (2007). Effect of rare earth elements on cold-responsive Ca2+ signaling in tobacco cells. Cryobiol. Cryotechnol. 53, 7–11.
Lin, C., Yu, Y., Kadono, T., Iwata, M., Umemura, K., Furuichi, T., et al. (2005). Action of aluminum, novel TPC1-type channel inhibitor, against salicylate-induced and cold shock-induced calcium influx in tobacco BY-2 cells. Biochem. Biophys. Res. Commun. 332, 823–830. doi: 10.1016/j.bbrc.2005.05.030
Lukaszewski, K. M., and Blevins, D. G. (1996). Root growth inhibition in boron-deficient or aluminum-stressed squash may be a result of impaired ascorbate metabolism. Plant Physiol. 112, 1135–1140.
Ma, J. F., Yamamoto, R., Nevin, D. J., Matsumoto, H., and Brown, P. H. (1999). Al binding in the epidermis cell wall inhibits cell elongation of okra hypocotyl. Plant Cell Physiol. 40, 549–556. doi: 10.1093/oxfordjournals.pcp.a029576
Markich, S. J., Warne, M. S. J., Westbury, A. M., and Roberts, C. J. (2002). A compilation of data on the toxicity of chemicals to species in Australasia. Part 3: metal. Aust. J. Ecotoxicol. 8, 1–137.
Momohara, I., Matsumoto, Y., and Ishizu, A. (1990). Involvement of veratryl alcohol and active oxygen species in degradation of a quinone compound by lignin peroxidase. FEBS Lett. 273, 159–162. doi: 10.1016/0014-5793(90)81074-X
Nakano, M., Sugioka, K., Ushijima, Y., and Goto, T. (1986). Chemiluminescence probe with Cypridina luciferin analog, 2-methyl-6-phenyl-3,7-dihydroimidazo[1,2-a]-pyrazin-3-one, for estimating the ability of human granulocytes to generate O2–. Anal. Biochem. 159, 363–369. doi: 10.1016/0003-2697(86)90354-4
Panda, S. K., Baluska, F., and Matsumoto, H. (2009). Aluminum stress signaling in plants. Plant Signal. Behav. 4, 592–597. doi: 10.4161/psb.4.7.8903
Pinton, R., Cakmak, I., and Marschner, H. (1994). Zinc deficiency enhanced NAD(P)H-dependent superoxide radical production in plasma membrane vesicles isolated from roots of bean plants. J. Exp. Bot. 45, 45–50. doi: 10.1093/jxb/45.1.45
Poschenrieder, C., Gunséa, B., Corralesa, I., and Barcelóa, J. (2008). A glance into aluminum toxicity and resistance in plants. Sci. Total Environ. 400, 356–368. doi: 10.1016/j.scitotenv.2008.06.003
Rengel, Z., and Zhang, W. H. (2003). Role of dynamics of intracellular calcium in aluminum-toxicity syndrome. New Phytol. 159, 295–314. doi: 10.1046/j.1469-8137.2003.00821.x
Santa-Maria, G. E., and Cogliatti, D. H. (1988). Bidirectional Zn-fluxes and compartmentation in wheat seedling roots. J. Plant Physiol. 132, 312–315. doi: 10.1016/S0176-1617(88)80112-3
Sanzonowicz, C., Smyth, T. J., and Israel, D. W. (1998). Calcium alleviation of hydrogen and aluminum inhibition of soybean root extension from limed soil into acid subsurface solutions. J. Plant Nutr. 21, 785–804. doi: 10.1080/01904169809365442
Takahashi, K., Isobe, M., Knight, M. R., Trewavas, A. J., and Muto, S. (1997). Hypoosmotic shock induces increases in cytosolic Ca2+ in tobacco suspension-culture cells. Plant Physiol. 113, 587–594.
Yamamoto, Y., Kobayashi, Y., Devi, S. R., Rikiishi, S., and Matsumoto, H. (2002). Aluminum toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells. Plant Physiol. 128, 63–72. doi: 10.1104/pp.010417
Keywords: aluminum, zinc, BY-2, Nicotiana tabacum L., Oryza sativa L., ROS
Citation: Lin C, Hara A, Comparini D, Bouteau F and Kawano T (2015) Zinc-Dependent Protection of Tobacco and Rice Cells From Aluminum-Induced Superoxide-Mediated Cytotoxicity. Front. Plant Sci. 6:1079. doi: 10.3389/fpls.2015.01079
Received: 09 September 2015; Accepted: 18 November 2015;
Published: 01 December 2015.
Edited by:
Zhulong Chan, Wuhan Botanic Garden, Chinese Academy of Sciences, ChinaCopyright © 2015 Lin, Hara, Comparini, Bouteau and Kawano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tomonori Kawano, kawanotom@kitakyu-u.ac.jp