 Alejandro Pérez-de-Luque
Alejandro Pérez-de-Luque Fernando Flores
Fernando Flores Diego Rubiales
Diego Rubiales- 1Area de Mejora y Biotecnologia, IFAPA, Centro Alameda del Obispo, Córdoba, Spain
- 2Departamento de Ciencias Agroforestales, ETSI La Rábida, Universidad de Huelva, Huelva, Spain
- 3Institute for Sustainable Agriculture, Consejo Superior de Investigaciones Científicas (CSIC), Córdoba, Spain
Root parasitic weeds are a major limiting production factor in a number of crops, and control is difficult. Genetic resistance and chemical control lead the fight, but without unequivocal success. Models that help to describe and even predict the evolution of parasitism underground are a valuable tool for herbicide applications, and even could help in breeding programs. Legumes are heavily affected by Orobanche crenata (crenate broomrape) in the Mediterranean basin. This work presents a descriptive model based on thermal time and correlating growing degree days (GDD) with the different developmental stages of the parasite. The model was developed in three different legume crops (faba bean, grass pea and lentil) attacked by crenate broomrape. The developmental stages of the parasite strongly correlated with the GDD and differences were found depending on the host crop.
Introduction
Broomrapes (Orobanche and Phelipanche spp.) are holoparasitic plants which attach to the host roots and grow at the expense of the host plant's resources. They parasitize a large number of crops, legumes being some of the most severely affected (Joel et al., 2007; Parker, 2013). Because the pathogenesis and most of the growing process take place underground long before diagnosis of the infection, it hampers the development of effective control strategies. In addition, the large amount of seeds released by a single individual (more than 100,000) provides the parasite with a great genetic adaptability to environmental changes (Parker and Riches, 1993; Press and Graves, 1995; Joel et al., 2007). Orobanche crenata Forsk. (crenate broomrape) is widely spread through the Mediterranean basin, and remains to date as the most important threat against legumes in the area (Parker, 2009, 2013; Rubiales and Heide-Jørgensen, 2011). Other broomrape species such Orobanche foetida Poir. or Phelipanche aegyptiaca (Pers.) Pomel can also infect legumes, but are of local importance only.
Although O. crenata has a broad host range infecting most legume crops, levels of infection vary between the different species and between accessions within species (Rubiales et al., 2003a,b; Pérez-de-Luque et al., 2004; Román et al., 2007). No complete resistance to O. crenata is available in any legume so far, but existing levels of incomplete resistance in some accessions (Rubiales et al., 2006, 2014; Pérez-de-Luque et al., 2007; Fernández-Aparicio et al., 2008, 2012) can delay the growth of already established broomrapes, because the presence of defense mechanisms interferes with the normal development of the parasite (Pérez-de-Luque et al., 2004, 2005).
In addition to the host, the environment can greatly influence the development of the parasitic weed on a given host. Although water availability may be an important factor for broomrape development, particularly in Mediterranean areas (Rubiales et al., 2003a,b; Pérez-de-Luque et al., 2004), it seems that temperature is the main factor affecting broomrape development (Mesa-García and García-Torres, 1986; Castejón-Muñoz et al., 1993; Eizenberg et al., 2004; Ephrath and Eizenberg, 2010). Increased temperature is often associated with an increase in broomrape parasitism, whereas low temperature correlates with lower infections. Cool winters reduce broomrape infection in several crops such as sunflower (Helianthus annuus L.), chickpea (Cicer arietinum L.), lentil (Lens culinaris Med.), pea (Pisum sativum L.), vetch (Vicia sativa L.) or faba bean (Vicia faba L.) (Arjona-Berral et al., 1987; Castejón-Muñoz et al., 1993; Rubiales et al., 2003a,b).
In recent years, several experiments accomplished by Eizenberg and coworkers have correlated the growth of broomrape (Phelipanche aegyptiaca, O. minor and O. cumana) with the thermal time, measured as growing day degrees (GDD) (Eizenberg et al., 2004, 2005; Ephrath and Eizenberg, 2010; Ephrath et al., 2012). Even more, the thermal time can be used to predict broomrape growth on crop roots under the soil, and mathematical models can be developed based on GDD for optimization of herbicide timing application (Eizenberg et al., 2006). This can be a powerful tool for the chemical control of broomrapes, because one of the main problems to date has been establishing the right time for herbicide applications, as to date no consistent relationships between the growth stages of the different hosts and the parasite development have been found (Arjona-Berral et al., 1987).
However, there are no works to date using a thermal time relation as a tool for showing differences in parasite development on different hosts. If temperature affects both, host and parasite development, a thermal time model could be used as a descriptive method for differences in infection levels integrating the two main factors affecting broomrape development: the host and the temperature. For that reason, the relationship between soil temperature and O. crenata growth on three different legume crops has been studied and a putative descriptive model based on work on O. minor / Trifolium by Eizenberg et al. (2005) is proposed. The hypotheses are that a) there are differences in parasite virulence between different host species, b) a sigmoidal relation between thermal time and parasite development exists and c) such relation will show the differences in parasite development between hosts.
Materials and Methods
Plant Material
Lentils, faba beans and grass peas (Lathyrus sativus L.) were included in the study. The three tested species are natural hosts for O. crenata, and the cultivars selected (“Rubia de la Armuña” for lentil, “Prothabon” for faba bean, breeding line “ICARDA 2” for grass pea) were susceptible to the parasitic weed attack.
Field Assays
Field trials were conducted over two cropping seasons (2006–2007 and 2007–2008) at Alameda del Obispo experimental station, in Córdoba (southern Spain). The fields located there present a deep loam soil (Typic Xerofluvent). The selected crops were sown by the end of November in a field highly infested with O. crenata, in 2 m length rows, with 0.7 m between rows and 20 plants per row. Four plots were set for each crop and every year (12 plots per year), surrounded and delimited by faba bean rows. The experiment was performed as a randomized, complete block design with four replications.
Temperature Data Recording
Soil temperature was recorded hourly starting January 1st and using data loggers (Gemini Tinytag Plus Data Logger) buried at 5 cm depth and converted to GDD according to McMaster and Wilhelm (1997) using the following equation:
where Tmax and Tmin are the maximum and minimum daily temperatures, respectively, and Tbase is the base temperature, all of them measured in degree Celsius (°C). Tbase was fixed as 0°C for faba bean and grass pea (Stützel, 1995; Rao and Northup, 2008), and 1.5°C for lentil (Ellis and Barrett, 1994).
Plant Sampling and Broomrape Development
Beginning the first week of February, five plants were extracted from each crop plot every week. The number of attached broomrapes was recorded and their developmental phase classified according to seven stages (Pérez-de-Luque et al., 2004): Stage 1 (T1), tubercles smaller than 2 mm; Stage 2 (T2), tubercles greater than 2 mm, without root development; Stage 3 (T3), tubercles with crown roots, without shoot formation; Stage 4 (T4), shoot formation, remaining underground; Stage 5 (T5), shoot emergence; Stage 6 (T6), flowering; Stage 7 (T7), setting of seeds.
Statistical Analysis
Broomrape tubercle number was log transformed (log[number of broomrapes per plant]) and the transformed data presented because of nonhomogeneity in variance. In order to model each stage (T1 to T7) of broomrape development, log-transformed broomrape number was nonlinearly regressed to GDD using a three-parameter logistic function (Brown and Mayer, 1988):
where Y represents log-transformed broomrape tubercle number, a represents the upper asymptote (maximum broomrape number), x represents GDD, xo represents the GDD when the Y is 50% of maximum, and b represents the slope at xo. A t-test was used for comparison of the regression parameter estimates in each parasitism stage between crops (Zar, 1999). A t-test compares a difference with the standard error of that difference. We used the standard error reported by nonlinear regression. The numerator is the difference between fit values. The denominator is an estimate of the standard error of that difference, computed as the square root of the sum of the squares of the two standard error values. This is a reasonable estimate if the number of data points in the two curves is equal, or nearly so.
Results
The combined analysis of variance showed that the interaction between species and year was significant for all the developmental stages of broomrape. A strong relation was found for the broomrape stage and GDD (Figure 1; Tables 1, 2). The model can be divided into three phases for each parasite stage: lag, log and maximum. During the lag phase, the parasite population grows very slowly. After that, the log phase covers an exponential growth of the parasite population. Finally, a maximum is reached and the parasite population remains stable. The lag phase varied slightly with the crop, the parasite stage and the season. For example, the lag phase in faba bean for T3 stage lasted until 850 GDD in 2007 and 750 GDD in 2008; in grass pea until 750 GDD in 2007 and 650 GDD in 2008; and in lentil until 850 GDD in 2007 and 750 GDD in 2008. In general terms, a delay of about 100 GDD was observed in the 2007 season compared to the 2008 season, and the attachments started to develop first in grass pea and later in faba bean and lentil. For that same stage (T3), de log phase ended at about 1150 GDD in 2007 and 1050 GDD in 2008 for faba bean (lasted for about 300 GDD); at 1050 GDD in 2007 and 950 GDD in 2008 for grass pea (lasted for 300 GDD); and at 1050 GDD in 2007 and 950 GDD in 2008 for lentil (lasted for 200 GDD).
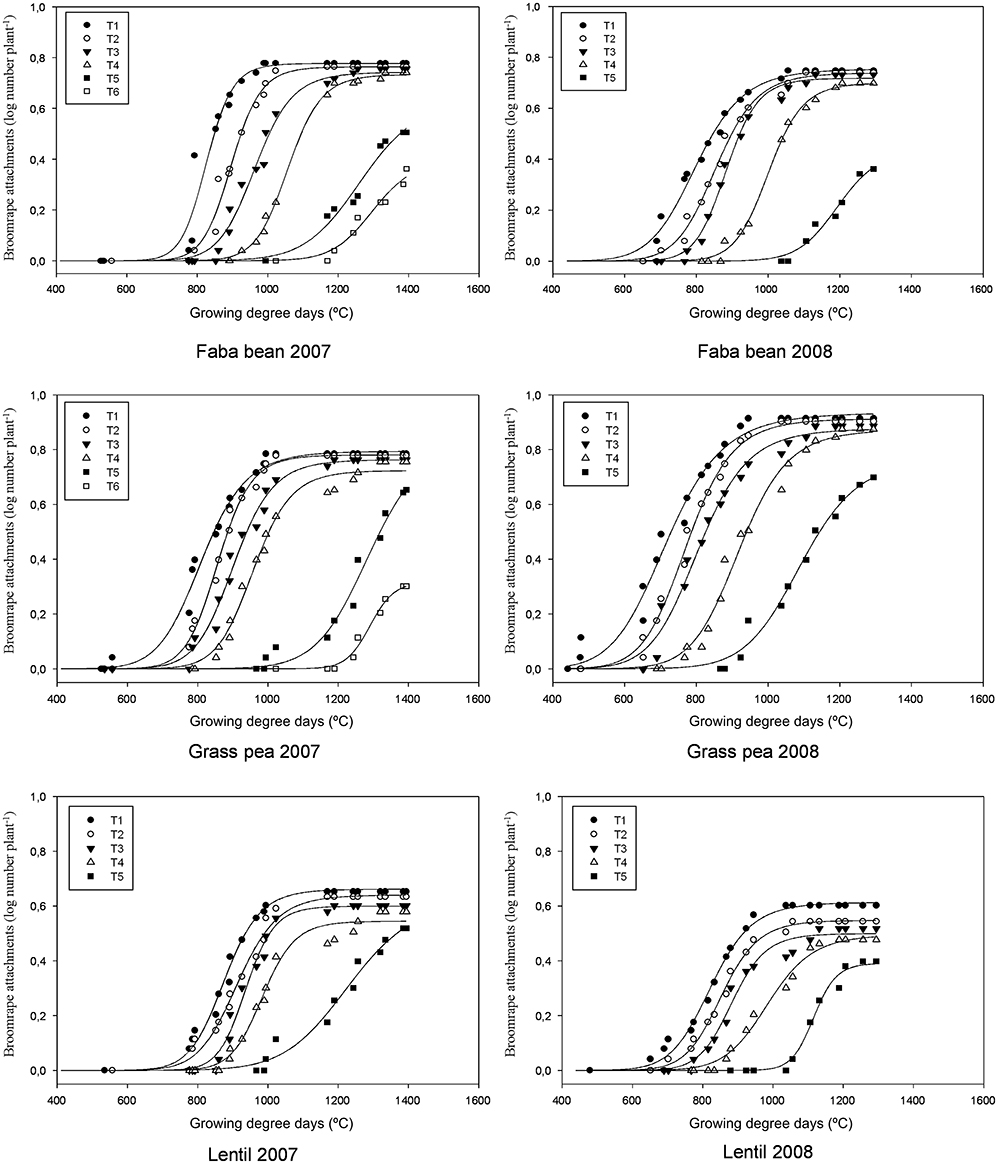
Figure 1. Three-parameter sigmoid nonlinear regressions between log-transformed broomrape attachment number and growing degree days (GDD) in three crops (faba bean, grass pea and lentil) grouped by broomrape growth stage in two years (2007 and 2008).
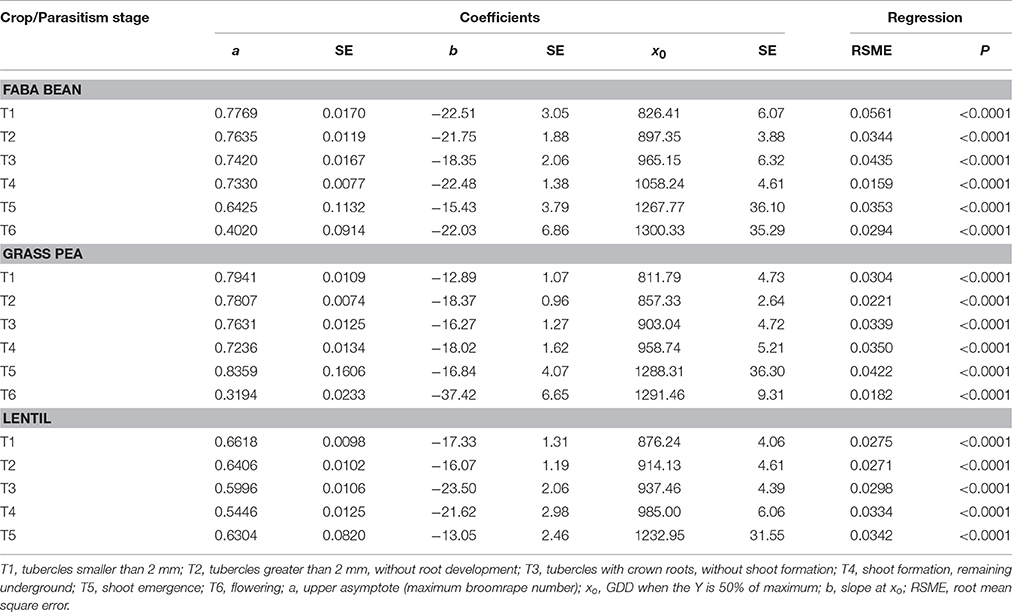
Table 1. Coefficients of a three-parameter sigmoid nonlinear regression between log-transformed broomrape number and GDD for season 2006-07.
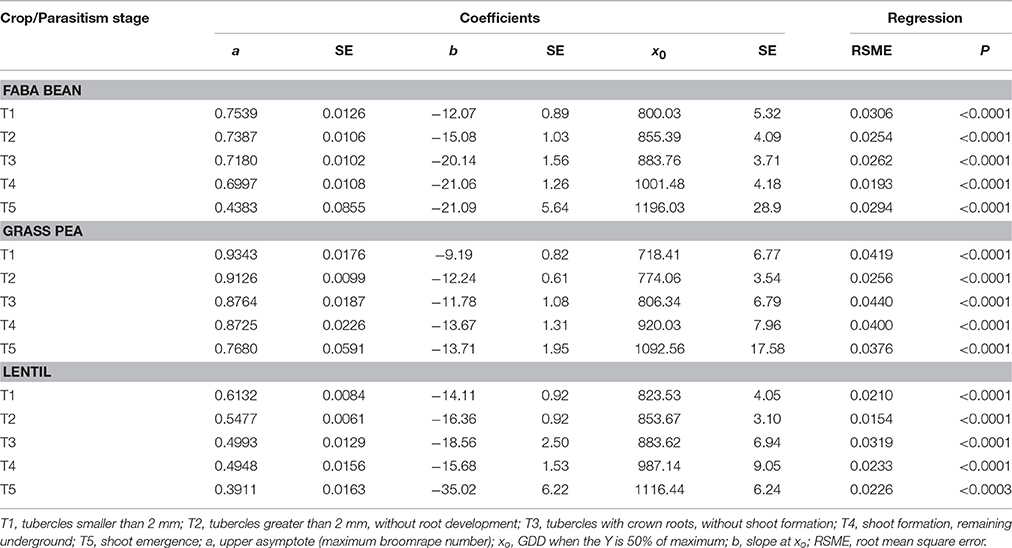
Table 2. Coefficients of a three-parameter sigmoid nonlinear regression between log-transformed broomrape number and GDD for season 2007–08.
The median (xo) in crenate broomrape parasitism (the GDD in the log phase where the infection was 50% of the maximum) varied also with the crop, the parasitism stage and the season (Tables 1, 2). This index gives an indication about the speed of parasite growth on the host: a low value means that less GDD are needed to reach 50% of the maximum number of attachments, so the parasite grows faster. In general, grass pea presented the lower values, followed by lentil and then faba bean, with some exceptions mainly in season 2007. Broomrape grew faster on grass pea than on the other crops and needed less GDD for developing the shoot (T4, 920–958) than those on lentil (985–987) or faba bean (1001–1058). During season 2007, broomrapes needed some more GDD to reach the median (xo) of all the developmental stages (except T4 in lentil) than during season 2008. Such differences were more accentuated in grass pea (near 100 GDD for most of the stages) than in faba bean or lentil (near 50–60 GDD for most of the stages).
The maximum parasite number was reached in stages T1 to T4 at about 1000 GDD for T1 and T2, 1100–1200 GDD for T3, and 1200 GDD for T4. It was similar for the three crops and the two seasons. However, the number of broomrapes per plant was different depending on the crop: grass pea and faba bean supported a higher number of broomrapes than lentil (Figure 1). In addition, the upper asymptote (or maximum broomrape number, a) reached by stages T1 to T4 for each crop was similar between years: 0.7–0.8 (5.0–6.3 broomrapes per plant) for faba bean, 0.7–0.9 (5.0–7.9 broomrapes per plant) for grass pea, 0.5–0.6 (3.2–4.0 broomrapes per plant) for lentil (Tables 1, 2). With respect to the emergence of broomrape (>T5), although the upper asymptote was not reached completely at the end of the experiment for all the crops, the tendency indicates a maximum below the threshold reached by the other stages: 04–0.6 (2.5–4.0 broomrapes per plant) for faba bean, 0.7–0.8 (5.0–6.3 broomrapes per plant) for grass pea, 0.4–0.6 (2.5–4.0 broomrapes per plant) for lentil. Finally, only in faba bean and grass pea during the 2007 season, broomrapes at the T6 stage were detected, with a lag phase at about 1100–1200 GDD, a median (xo) of 1300 and 1292 (respectively), and a maximum of 0.4 (2.5 broomrapes per plant) and 0.3 (2.0 broomrapes per plant) (respectively).
When the model was compared between crops, significant differences were found for the three parameters (a, b, and xo) regarding broomrape development (Table 3). The maximum broomrape number (a) was significantly different for the underground stages (T1–T4) in lentils respect to the other two crops in 2007 and for the three crops in 2008. For aboveground stage (T5) only grass pea was significantly different from faba bean and lentil in 2008. The median (xo) was significantly different for the three crops considering the underground stages in 2007, except for stage T1 in faba bean and grass pea. In 2008, grass pea values were different from those of faba bean and lentil, except for T1 stage, which values were significant for all the crops. The values for aboveground stages (T5) were significant only in 2008 for faba bean respect to the other two crops. The slope at xo (b) showed the most variable results, with significant differences varying between the season and the parasitism stage considered.
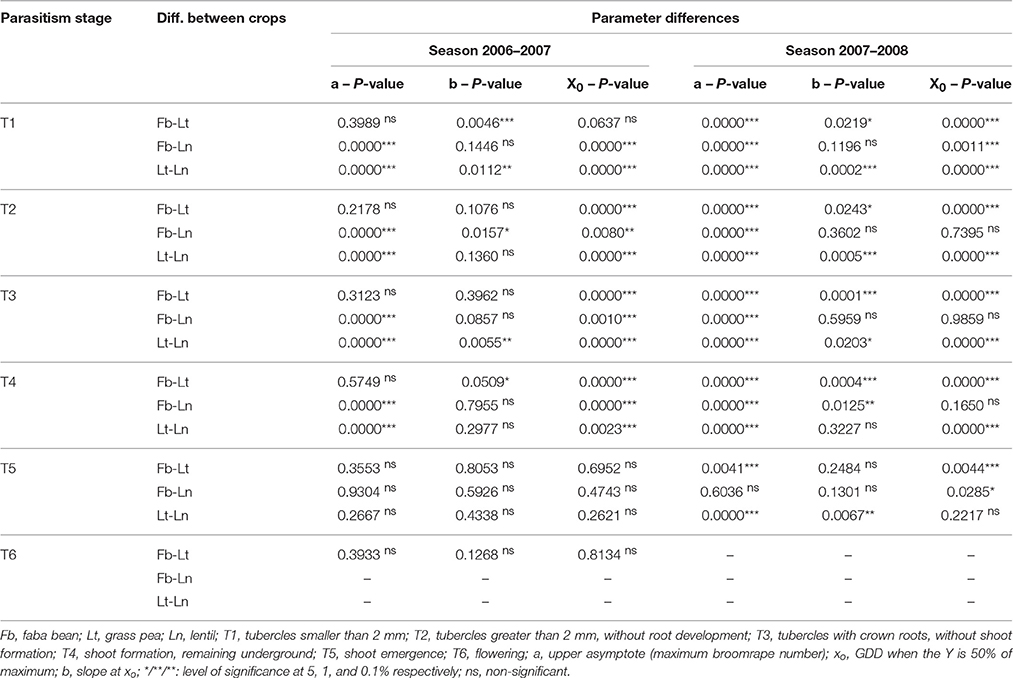
Table 3. Comparison of the coefficients for the three-parameter sigmoid nonlinear regression between the different crops.
Discussion
The present study confirms that the growth of crenate broomrape on the three tested legume crops is highly temperature-related. The thermal time measured as GDD appears as a valuable tool for describing the parasite growth and establishing the developmental stage of the infection, as previously shown for other crops and broomrape species (Eizenberg et al., 2004, 2005; Ephrath and Eizenberg, 2010; Ephrath et al., 2012).
Although we found differences between years, such differences do not seem to be a problem for developing a descriptive model because, in general terms, they were about 100 GDD or less. Broomrapes growing on grass pea appear as the most affected by year to year variation, with a faster rate and a higher number of infections in the second season (2008) than in the previous one (2007). There are previous reports about differences in broomrape infection depending on the crop and the environmental conditions (Pérez-de-Luque et al., 2004). Such differences could be explained by the uneven rainfall during both seasons. The distribution of the rainfall during both seasons showed some differences (Supplementary Figure 1), with a remarkable higher precipitation during January and April in 2008 compared to 2007, and a low rainfall during March 2008 respect to 2007. The higher rainfall during January 2008 could positively have affected germination of broomrape and the beginning of the infection (allowing a better spreading of germination stimulants), because such conditions have been previously reported as a factor increasing parasitism (Pérez-de-Luque et al., 2004). It is possible that including soil moisture measurements, in addition to thermal time, might improve this kind of models, mainly in dryland fields with no irrigation (highly dependent on rainfall).
Regarding crop species, differences were found in the development of the parasite stages. Most of the parasites evolved from one stage into the next one (T1 to T2, etc.). However, the upper asymptote (a) (the maximum) was lower as the developmental stage advances. In the T5 stage emergence differences between crop species become more marked: a lower proportion of broomrapes emerged in faba bean compared with the other two crops. It seems that broomrapes attached to grass pea or lentil can evolve more easily into more advanced developmental stages compared to faba bean, which could be explained by a better availability of resources from such host. It is possible that severity of the attack and the development of broomrapes could be related to physiological traits of the host plant determining the allocation of nutrients into the parasite, as has been shown in Striga spp. (Arnaud et al., 1999). In addition, the size of emerged broomrapes on faba bean was bigger than those on grass pea and lentil. As big individuals are more competitive for resources than small ones, intra-specific competition might play a role here.
The comparison of the descriptive thermal model between the three crops confirms significant differences regarding the evolution of the developmental stages of broomrape. The most consistent differences are found when the parameters (mainly a and xo) of the underground stages (T1-T4) are considered. The thermal models are usually associated to a specific host (Eizenberg et al., 2004, 2005; Ephrath and Eizenberg, 2010; Ephrath et al., 2012) and they link phenological events with temperature (GDD). This means that the models include physiological traits of the host (for example, taking into account the crop's base temperature). Considering this, differences in the parameters of the model for different crops are pointing out differences in the virulence of the parasite against each crop. In the same way, such differences could be used for differentiating susceptible from resistant genotypes: resistant accessions have been shown to delay broomrape development in the field (Rubiales et al., 2003a; Pérez-de-Luque et al., 2004). Additionally, resistance to broomrape has been reported as temperature dependent in some cases (Eizenberg et al., 2003), so a model considering the environmental temperature would be desirable for evaluating sources of resistance in a breeding programme. Future work should involve validation of this kind of descriptive model in order to develop a predictive one, using different locations and cultivars with different degrees of resistance against crenate broomrape, in order to use it as a tool for plant breeding.
Author Contributions
AP and DR: Conceived and designed the experiments; AP: Developed the experiment; FF performed the statistical analysis; AP, FF, and DR: Discussed and interpreted the results; AP: Wrote a first draft of the manuscript and DR and FF helped with the writing and further edition and correction process; DR: Contributed materials, equipment, and analysis tools.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge the financial support from the EU project FP6-2002-FOOD-1-5062232004-2008 and Spanish Ministerio de Economia and Competitividad project ALG2014-52871-R confinanced by FEDER.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.01910/full#supplementary-material
References
Arjona-Berral, A., Mesa-García, J., and García-Torres, L. (1987). Phenology and growth of Orobanche crenata Forsk. (broomrape) in four legume crops. Weed Res. 27, 349–360. doi: 10.1111/j.1365-3180.1987.tb00772.x
Arnaud, M. C., Véronési, C., and Thalouarn, P. (1999). Physiology and histology of resistance to Striga hermonthica in Sorghum bicolor var. Framida. Aust. J. Plant Physiol. 26, 63–70. doi: 10.1071/PP98070
Brown, R. F., and Mayer, D. G. (1988). Representing cumulative germination. The use of the Weibull function and other empirically derived curves. Ann. Bot. 61, 127–138.
Castejón-Muñoz, M., Romero-Muñoz, F., and García-Torres, L. (1993). Effect of planting date on broomrape (Orobanche cernua Loefl.) infection on sunflower (Helianthus annuus L.). Weed Res. 33, 171–176. doi: 10.1111/j.1365-3180.1993.tb01930.x
Eizenberg, H., Colquhoun, J. B., and Mallory-Smith, C. A. (2004). The relationship between growing degree days and small broomrape (Orobanche minor) parasitism in red clover (Trifolium pratense). Weed Sci. 52, 735–741. doi: 10.1614/WS-03-157R
Eizenberg, H., Colquhoun, J. B., and Mallory-Smith, C. A. (2005). A predictive degree-days model for small broomrape (Orobanche minor) parasitism in red clover in Oregon. Weed Sci. 53, 37–40. doi: 10.1614/WS-04-018R1
Eizenberg, H., Colquhoun, J. B., and Mallory-Smith, C. A. (2006). Imazamox application timing for small broomrape (Orobanche minor) control in red clover. Weed Sci. 54, 923–927. doi: 10.1614/WS-05-151R.1
Eizenberg, H., Plakhine, D., Hershenhorn, J., Kleifeld, Y., and Rubin, B. (2003). Resistance to broomrape (Orobanche spp.) in sunflower (Helianthus annuus L.) is temperature dependent. J. Exp. Bot. 54, 1305–1311. doi: 10.1093/jxb/erg129
Ellis, R. H., and Barrett, S. (1994). Alternating temperatures and rate of seed germination in lentil. Ann. Bot. 74, 519–524. doi: 10.1006/anbo.1994.1149
Ephrath, J. E., and Eizenberg, H. (2010). Quantification of the dynamics of Orobanche cumana and Phelipanche aegyptiaca parasitism in confectionery sunflower. Weed Res. 50, 140–152. doi: 10.1111/j.1365-3180.2010.00768.x
Ephrath, J. E., Hershenhorn, J., Achdari, G., Bringer, S., and Eizenberg, H. (2012). Use of logistic equation for detection of the initial parasitism phase of Egyptian broomrape (Phelipanche aegyptiaca) in tomato. Weed Sci. 60, 57–63. doi: 10.1614/WS-D-11-00070.1
Fernández-Aparicio, M., Flores, F., and Rubiales, D. (2012). Escape and true resistance to crenate broomrape (Orobanche crenata Forsk.) in grass pea (Lathyrus sativus L.) germplasm. Field Crops Res. 125, 92–97. doi: 10.1016/j.fcr.2011.09.003
Fernández-Aparicio, M., Sillero, J. C., Pérez-de-Luque, A., and Rubiales, D. (2008). Identification of sources of resistance to crenate broomrape (Orobanche crenata) in Spanish lentil (Lens culinaris) germplasm. Weed Res. 48, 85–94. doi: 10.1111/j.1365-3180.2008.00604.x
Joel, D. M., Hershenhorn, J., Eizenberg, H., Aly, R., Ejeta, G., Rich, P. J., et al. (2007). Biology and management of weedy root parasites. Hortic. Rev. 33, 267–349. doi: 10.1002/9780470168011.ch4
McMaster, G. S., and Wilhelm, W. W. (1997). Growing degree-days: one equation, two interpretations. Agric. Forest Meteorol. 87, 291–300. doi: 10.1016/S0168-1923(97)00027-0
Mesa-García, J., and García-Torres, L. (1986). Effect of planting date on parasitism of broadbean (Vicia faba) by crenate broomrape (Orobanche crenata). Weed Sci. 34, 544–550.
Parker, C. (2009). Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag. Sci. 65, 453–459. doi: 10.1002/ps.1713
Parker, C. (2013). “The parasitic weeds of the Orobanchaceae,” in Parasitic Orobanchaceae, eds D. M. Joel, J. Gressel, and L. Musselman (Berlin: Springer-Verlag), 313–344.
Parker, C., and Riches, C. R. (1993). Parasitic Weeds of the World: Biology and Control. Wallingford, CT: CAB International.
Pérez-de-Luque, A., Jorrín, J., Cubero, J. I., and Rubiales, D. (2005). Orobanche crenata resistance and avoidance in pea (Pisum spp.) operate at different developmental stages of the parasite. Weed Res. 45, 379–387. doi: 10.1111/j.1365-3180.2005.00464.x
Pérez-de-Luque, A., Lozano, M. D., Moreno, M. T., Testillano, P. S., and Rubiales, D. (2007). Resistance to broomrape (Orobanche crenata) in faba bean (Vicia faba): cell wall changes associated with pre-haustorial defensive mechanisms. Ann. Appl. Biol. 151, 89–98. doi: 10.1111/j.1744-7348.2007.00164.x
Pérez-de-Luque, A., Sillero, J. C., Moral, A., Cubero, J. I., and Rubiales, D. (2004). Effect of sowing date and host resistance on the establishment of Orobanche crenata in faba bean and common vetch. Weed Res. 44, 282–288. doi: 10.1111/j.1365-3180.2004.00401.x
Rao, S., and Northup, B. (2008). Planting date affects production and quality of grass pea forage. Crop Sci. 48, 1629–1635. doi: 10.2135/cropsci2007.11.0622
Román, B., Satovic, Z., Alfaro, C., Moreno, M. T., Kharrat, M., Pérez-de-Luque, A., et al. (2007). Host differentiation in Orobanche foetida Poir. Flora 202, 201–208. doi: 10.1016/j.flora.2006.07.003
Rubiales, D., Alcántara, C., Pérez-de-Luque, A., Gil, J., and Sillero, J. C. (2003a). Infection of chickpea (Cicer arietinum) by crenate broomrape (Orobanche crenata) as influenced by sowing date and weather conditions. Agronomie 23, 359–362. doi: 10.1051/agro:2003016
Rubiales, D., Flores, F., Emeran, A. S., Kharrat, M., Amri, M., Rojas-Molina, M. M., et al. (2014). Identification and multi-environment validation of resistance against broomrapes (Orobanche crenata and O. foetida) in faba bean (Vicia faba). Field Crops Res. 166, 58–65. doi: 10.1016/j.fcr.2014.06.010
Rubiales, D., and Heide-Jørgensen, H. S. (2011). “Parasitic plants,” in: eLS (Chichester: John Wiley & Sons Ltd). Available online at: http://www.els.net; http://onlinelibrary.wiley.com/doi/10.1002/9780470015902.a0021271/full
Rubiales, D., Pérez-de-Luque, A., Cubero, J. I., and Sillero, J. C. (2003b). Crenate broomrape (Orobanche crenata) infection in field pea cultivars. Crop Prot. 22, 865–872. doi: 10.1016/S0261-2194(03)00070-X
Rubiales, D., Pérez-de-Luque, A., Sillero, J. C., Román, B., Kharrat, M., Khalil, S., et al. (2006). Screening techniques and sources of resistance against parasitic weeds in grain legumes. Euphytica 147, 187–199. doi: 10.1007/s10681-006-7399-1
Stützel, H. (1995). A simple model for simulation of growth and development in faba beans (Vicia faba L.), 1. Model description. Eur. J. Agron. 4, 175–185. doi: 10.1016/S1161-0301(14)80044-0
Keywords: Orobanche crenata, parasitic weeds, GDD, faba bean, lentil, grass pea, legumes
Citation: Pérez-de-Luque A, Flores F and Rubiales D (2016) Differences in Crenate Broomrape Parasitism Dynamics on Three Legume Crops Using a Thermal Time Model. Front. Plant Sci. 7:1910. doi: 10.3389/fpls.2016.01910
Received: 08 July 2016; Accepted: 02 December 2016;
Published: 15 December 2016.
Edited by:
Paul Christiaan Struik, Wageningen University and Research Centre, NetherlandsReviewed by:
Grama Nanjappa Dhanapal, University of Agricultural Sciences, Bangalore, IndiaHanan Eizenberg, Agricultural Research Organization, Israel
Copyright © 2016 Pérez-de-Luque, Flores and Rubiales. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alejandro Pérez-de-Luque, alejandro.perez.luque@juntadeandalucia.es