 Nilsa E. Gonzalez-Britez1,2
Nilsa E. Gonzalez-Britez1,2 Hernán J. Carrasco2*
Hernán J. Carrasco2* Clara Elena Martínez Purroy2
Clara Elena Martínez Purroy2 M. Dora Feliciangeli3
M. Dora Feliciangeli3 Marisel Maldonado1
Marisel Maldonado1 Elsa López1
Elsa López1 Maikell J. Segovia2
Maikell J. Segovia2 Antonieta Rojas de Arias4
Antonieta Rojas de Arias4
- 1Departamento de Medicina Tropical, Instituto de Investigaciones en Ciencias de la Salud (IICS), Universidad Nacional de Asunción, Asunción, Paraguay
- 2Laboratorio de Biología Molecular de Protozoarios, Facultad de Medicina, Instituto de Medicina Tropical, Universidad Central de Venezuela, Caracas, Venezuela
- 3Instituto de Investigaciones Biomédicas (BIOMED), Universidad de Carabobo, Maracay, Venezuela
- 4Centro para el Desarrollo de la Investigación Científica (CEDIC)/Díaz Gill Medicina Laboratorial/Fundación Moisés Bertoni, Asunción, Paraguay
Triatoma sordida is widely distributed throughout the Chaco and the Eastern Region of Paraguay. It is associated to palm trees and artificial ecotopes located in peridomestic environments. The aim of this work was to determine genetic and morphometric variability and feeding behavior among population of T. sordida captured in domicile and peridomicile areas of Paraguay. Feeding contents and levels of genetic and morphometric variation were determined in 124 T. sordida from domicile and peridomicile populations of San Pedro and Paraguarí departments of the Eastern Region and Boquerón and Presidente Hayes departments of the Western region using Double Diffusion Gel, random amplified polymorphic DNA (RAPD), and head and wings morphometry. Morphometric analysis revealed isolation of populations by geographic region and larger size in triatomine populations from the Western Region. RAPD showed no specific patterns for domicile and peridomicile populations. The estimator of diversity (FST; 0.08) and high gene flow obtained (Nm; 5.7) did not allow the establishment of genetic differentiation within the same region. The blood meal source showed that poultry feeding was 38% of host preferences, and human blood was the second feeding preference (24%) in the insects from the Eastern Region while poultry feeding was predominant in those from the Western Region (30%). This work showed homogeneity between T. sordida populations of the same region and between domicile and peridomicile. The genetic diversity was determined among T. sordida populations of both geographical regions suggesting differentiation associated to eco-geographical isolation by distance. It is important to notice that pattern feedings were different between the two regions. Further studies should be focused on how phenetic and genetic variations could be related to the adaptation capacity of these triatomine populations to domicile, increasing their vector potentiality in the transmission of Chagas disease.
Introduction
The subfamily Triatominae (Hemiptera: Reduviidae) includes over 144 species of strictly hematophagous insects, considered potential vectors of Trypanosoma cruzi among mammals. However, not all of them are epidemiologically important (1–4). In Paraguay, 11 species of triatomines have been registered and from them Triatoma infestans (5) and T. sordida (6) have been found naturally infected with T. cruzi (7, 8).
In the Southern Cone countries, the most important hematophagous vector involved in the transmission of Chagas disease is T. infestans. T. sordida of wild origin seems to have been disseminated from Brazilian plateaus toward the south, and now is found in Argentina, Bolivia, Paraguay, and Uruguay where it occupies extensive geographical areas but generally in small populations of individuals (9, 10). T. sordida is considered as a ubiquitous species with high ecological potential living in various ecotopes and feeding from different sources. This insect could withstand large environmental changes that cause the disappearance of his competitors and could widen its ecotopes to dead and dry trees (11). However, these ecotopes usually do not offer feeding sources, stimulating its dispersion to peridomiciles and domiciles and there is ever-greater contact inside and around houses with species other than T. infestans that were not very important for vector transmission in the past because they used to be found only in natural ecotopes, as Triatoma sordida. Their epidemiological importance regarding vector transmission of Chagas disease is still low, but they may become a bigger problem if they become domesticated, thereby occupying the empty place left by T. infestans (12, 13).
On the other hand, the sympatry with T. infestans in domicile and peridomicile is known as well as the diversity of the ecotopes it occupies and the difficulty this has meant for its control. T. sordida is associated with re-infestation sources of dwellings treated with insecticides and currently is considered a potential vector of Chagas disease (11, 14–16).
Morphometric and molecular analyses are important tools that provide evidence of the population structure of insect vectors. Enzymatic and genetic studies performed on this triatomine species have confirmed the variability of loci in two groups (17–19). T. sordida group 2 seems to be restricted to the Chaco and group 1 is widely distributed in Bolivia and Brazil (18, 20). Besides, the genetic distances between both populations led to infer the hypothesis of recent cryptic speciation (21).
The study based on morphometric analysis and the molecular patterns of random amplified polymorphic DNA (RAPD) in T. brasiliensis have reported the existence of a common relationship between wild and domiciliary populations (22). Similarity, the gene-flow index and reduced genetic divergence found between different populations of Triatoma rubida support sub-specific designation for this species (23). In relation to T. sordida, low levels of genetic variation among populations of southeastern Brazil have been reported through the analysis of 28 allozyme loci. None of these loci presented significant differences between any pair of populations, and only two showed polymorphism, accounting for low levels of heterozygosity (10). Similarly, the genetic study of T. sordida from different ecotopes of Paraguay revealed low genetic diversity levels suggesting that extra-household populations could represent an important epidemiological link to maintain the transmission of trypanosomatids (24).
Thus, triatomines studies based on diverse molecular markers have been used to clarify phylogenetic and evolutionary relationships between species, apart from inferring divergences and population structure, as it has been demonstrated previously in others triatomines, where wide polymorphism between discrete populations suggest the existence of a species complex (25, 26). These facts suggest an increment of the epidemiological significance of vectors considered secondary (12, 22). In the case of T. sordida, the polymorphism levels and its implication in the infestation of dwellings in endemic areas for Chagas disease in Paraguay are still unknown. The objective of the present work was to determine the feeding behavior and genetic and morphometric variability among population of T. sordida captured in domicile and peridomicile of the two geographical regions of Paraguay. Finally, this study contributes to improve surveillance strategies embracing this potential vector.
Materials and Methods
Study Sites, Bug Collection, and Parasitological Search
The Eastern Region is humid, sub-tropical, composed by valleys, small hills, and wooded areas. The average annual temperature is 24.3°C and the average annual rainfall is between 1000 and 1600 mm. (27). The Western or Chaco Region is characterized by extreme temperatures ranging from 45°C in spring and summer to 27°C in winter with annual minimum rainfall of 100–900 mm. (28). Both regions are separated by an important ecological barrier, the Paraguay River.
The specimens were collected by manual capture in poultry house, stables, and pigsties of peridomicile and intra-domicile areas of San Pedro (SP) and Paraguarí (PA) departments of the Eastern Region of Paraguay; Boquerón (BO) and Presidente Hayes (PH) departments of the Western Region (Figure 1). All triatomines were maintained alive and classified previously as T. sordida according to Lent and Wygodzinsky (29).
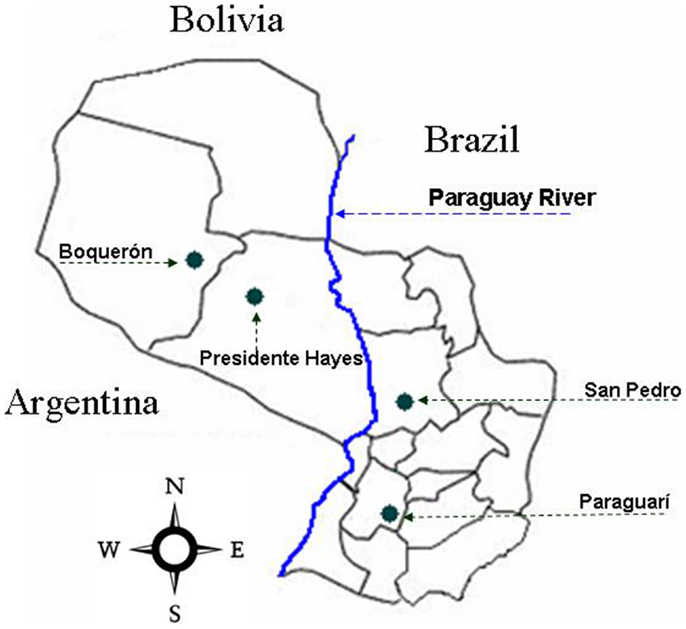
Figure 1. Geographical localization of the study area. The green dots show the capture places in the different departments of the Republic of Paraguay. Departments: Boquerón (BO), Presidente Hayes (PH), Paraguarí (PA), and San Pedro.
One hundred twenty-four specimens were analyzed: 63 males and 61 females (Table 1). All insects were studied by morphometric analysis and half of them were studied by molecular methods. Parasitological search was also carried out microscopically in all insects by the direct observation of their feces and morphological identification performed after staining with Giemsa at 400× in an Olympus microscope in order to identify trypanosomatids. Characteristic morphological features of T. cruzi were identified as described by Hoare (30).
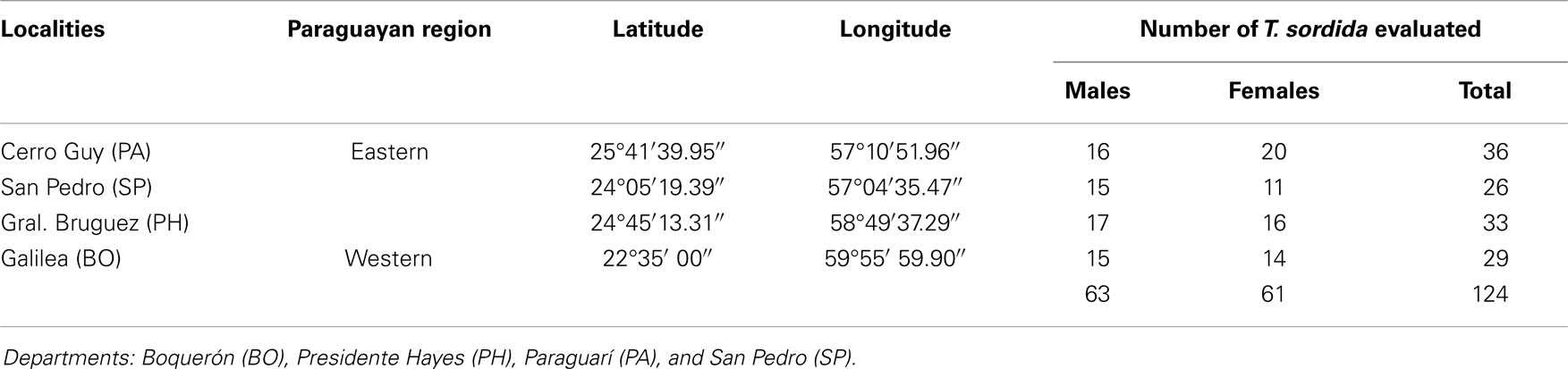
Table 1. Distribution of T. sordida captured in different localities of the Paraguayan regions.
Morphometric Analyses
Head and wings were selected according to protocols previously described (31–33). In the head, seven homologous points were selected (Figures 2A,B), while six distances measurements between the points of intersection of the veins were used for wings (Figure 2C). All measurements were made in duplicate by the same researcher and the images were captured using a lucid camera connected to an Olympus stereoscopic microscope DF Plan 1×.
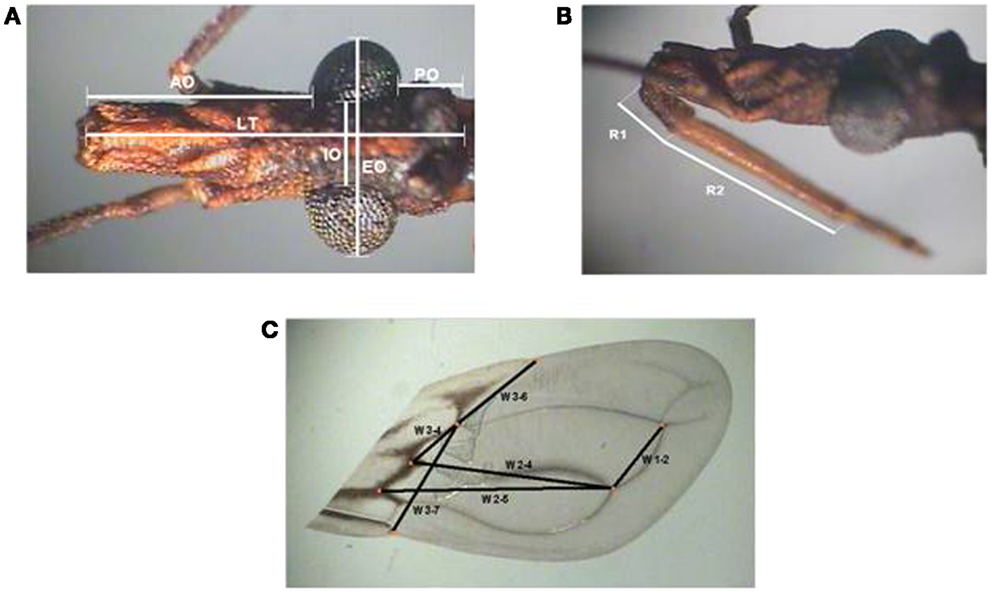
Figure 2. Diagram of head and wings measurements used for the morphometry of Triatoma sordida is shown. (A) Dorsal (left) and (B) lateral (right) views of the head indicating morphometric measurements taken. AO, anteocular distance; PO, post-ocular distance; LT, total length; IO, inner distance between eyes; EO, outer distance between eyes; R1, length of first rostral segment; R2, length of second rostral segment. (C) Dorsal view of right wing with measurements taken for the distance between the following points: W1–2 between point 1 and 2, W2–4 between point 2 and 4, W2–5 between point 2 and 5, W3–6 between points 3 and 6, W3–4 between points 3 and 4, and W3–7 between points 3 and 7.
The matrixes were tabulated by sex and population. The sexual dimorphism and Guillaumin profile were determined to obtain information about the general size of a group respect to other (34, 35). The principal components analysis (PCA) was carried out using covariance matrix from which a factorial map was constructed to show the differences in size and shape among sexes and populations (36). The free-allometry analysis for shape differences was performed after the discriminant analysis (DA) made on the set of common principal components (CPC), excepting the first common principal component (CPC1), according to a protocol described by Dujardin et al. (37). For this, it was indispensable to check the compatibility with the model of CPC using a Chi-square goodness-of-fit test (X2). All parameters were calculated using the JMP 4.0.0 (38) and NTSYSp.c version 2.10p (39) statistical packages.
Feeding Source Analyses
Extraction of blood content
The intestinal content of 62 adult specimens (29 males and 33 females) was extracted; 34 of them were from the Eastern Region (PA) and 28 from the Western Region (Pte. Hayes). In order to do this, a section was made in the front third of the abdomen of the specimens. When the content volume was insufficient, the complete promesenteron was transferred to a vial (40). Each vial had the same blood sample with 180 μL of 4% saline solution and 20 μL of 10% crystal violet and the mixture was maintained at 4°C for 24 h (41).
Determination of the feeding source
The Gel Double Diffusion method was carried out in a glass slide (7.5 cm × 5 cm) using 3.5 mL of 1.3% agar (I.D. Oxoid Agar) diluted in veronal hydrochloride buffer (pH 8.6). This preparation was maintained in a humid chamber for 24 h (40, 42). In the agar, there was a central hole that was filled up with the specific antiserum and six peripheral holes, five containing the diluted antigen (blood sample from different triatomines) and one with saline solution (negative control).
The antisera used for the identification of the feeding source was against human, poultry (chicken), dog, cat, goat, mice, and guinea pig blood and were also put in contact with the intestinal content of the triatomines searching for the corresponding antigen. All the antisera were prepared and tested previously in the Laboratory of General Ecology of the University of Buenos Aires, Argentina.
Random Amplified Polymorphic DNA
The extraction of DNA was carried out according to the protocol of Promega Wizard Genomic Purification Kit, USA (43) in five legs of each insect (25).
The amplification reaction was performed according to the protocol of Williams et al. (44) modified by Carrasco et al. (45). The DNA of 62 specimens: PH (8 males, 8 females), BO (7 males, 8 females), PA (8 males, 8 females), SP (8 males, 7 females) was amplified with four primers to distinguish triatomine species of the same genus or identify affinities between species. The primers were: A1 (5′-TCACGATGCA-3′), A2 (5′-GAAACGGGTG-3′), L4 (5′-GTGGATGCGA-3′), and L5 (5′-AAGAGCCCGT-3′). The PCR was set up as follows: a final volume of 25 μL PCR mixture that contained 0.25 mM dNTPs (Pharmacy Biotechnology, Sweden), 10 pmol of primers, 1.0 unit of Taq polymerase (Gibco Life Technology), 5 ng of DNA template in a buffer with 2 mM MgCl; 50 mM KCl, 10 mM Tris–HCl, pH 8.8 was used; each amplification included a DNA-free negative control. The visualization of the products was obtained using 2.5% ultra-pure agarose gel electrophoresis stained with ethidium bromide. The bands obtained were digitalized by KODAK 1D (Kodak Digital Science) software.
Data Analysis
The binary matrix was built using the specimens that generated better band reproducibility and intensity. For the analysis, it was assumed that the T. sordida populations were in Hardy–Weinberg equilibrium and that there were no selection processes favoring any particular genotype. All loci were entered in a binary matrix and a similarity index was obtained from this matrix (46) in order to build a UPGMA (unweighted pair group method of arithmetic mean) dendogram. The genetic distance was based on Nei (47) and the index of genetic differentiation (FST) was determined according to Nei and Chakraborty (48), which is used to examine the level of genetic divergence among sub-populations and provides an estimation of the genetic flow (Nm). These parameters were analyzed using the software POPGENE (version 1.31) (49).
Results
Parasitological Assays
Feces of 124 insects were analyzed looking for T. cruzi. None of them showed natural T. cruzi infection, confirming in this occasion the low infection rates of T. sordida specimens in both regions (data not shown).
Morphometric Assays
Size analysis
The Guillaumin profile allowed, in general, determining that individuals from the Western Region were larger than those from the Eastern Region. The sexual dimorphism of the analyzed structures (W2–5, W3–6, W3–7, W2–4, W1–2, W3–4, AO, R2, R1, EO) was significant (p = 0.01–0.0001), excepting the post-ocular (PO) distance of the head (p = 0.08). The Bonferroni correction showed that the characters of the wings differentiated the sexual dimorphism better than those of the head.
The size analysis using the principal components showed that, although females were consistently larger than males, the populations from BO department had significantly larger wings and heads than those from the other studied localities (Figure 3). The significance of the size differences among insects of different departments was determined by Kruskal–Wallis non-parametric ANOVA test, using the mean of each group separately (p = 0.0001) for head and (p = 0.001) for wings.
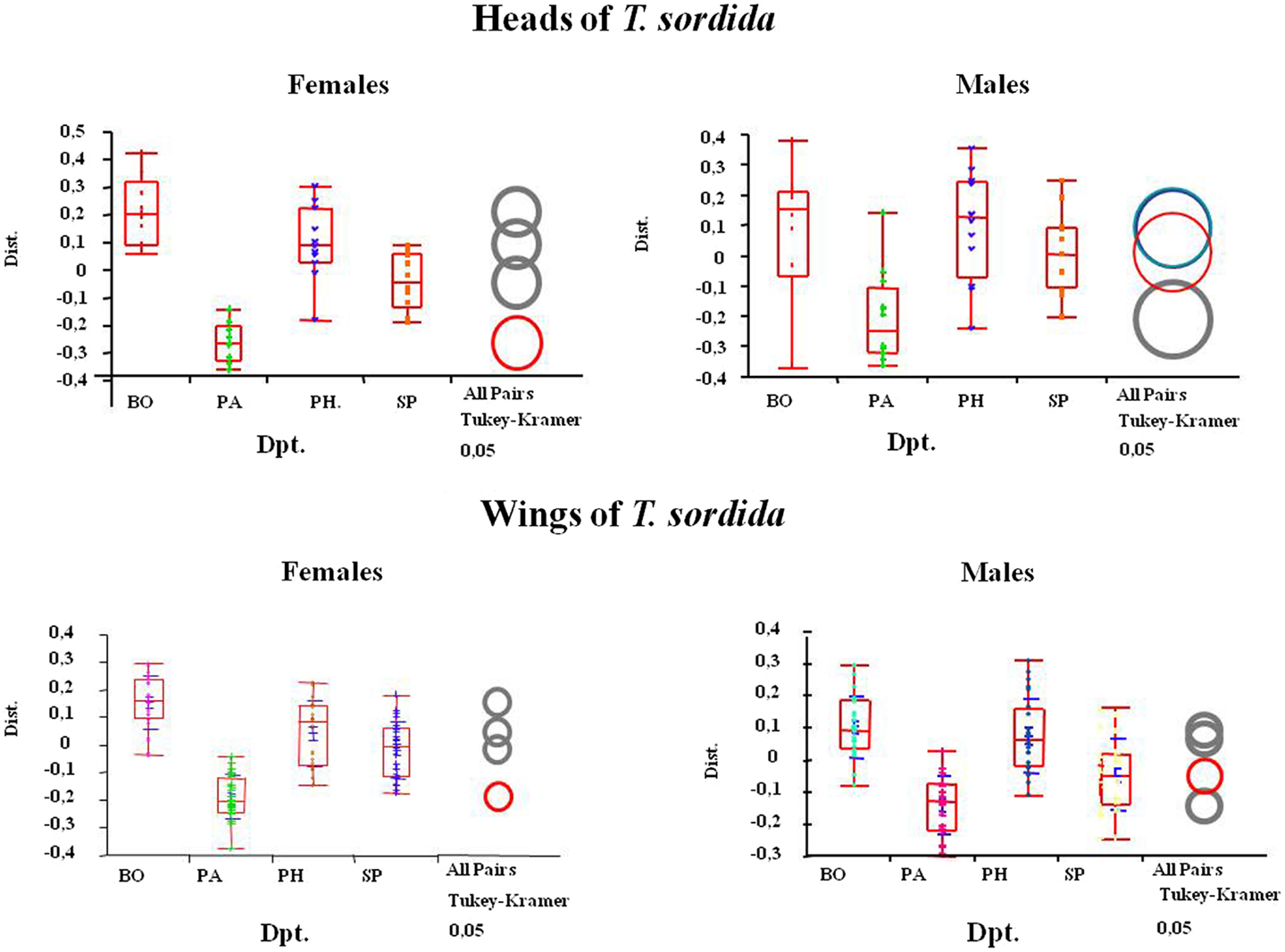
Figure 3. Size differences between geographical populations (from the indicated departments) in heads (up) and wings (below) of T. sordida males and females are shown. The arithmetic mean can be observed as a horizontal line that divides the boxes in two. The ends of the boxes correspond to 25th and 75th quantiles while the vertical lines show the maximum and minimum values of the distribution. BO, Boqueron Department; PA, Paraguari Department; PH, Presidente Hayes department; SP: San Pedro Department.
Conformation and shape analysis
The DA for isometry-free variables evidenced the significant separation of the triatomine populations of the two geographical regions, better reflected by females according to the values of Wilks lambda = 036, p = 0.0003 (head) and Wilks lambda = 042, p = 0.0001 (wings). With the elimination of the allometric size, only the head variables were compatible with the CPC (common principal component) model (x: 33.57, p = 0.2982 in females and x: 36.29, p = 0.1987 in males). Through the DA, the specimens were correctly classified into their respective groups with considerable concordance (Kappa between 0.66 and 0.83). This canonical variation analysis also showed the isolation of PA population for both sexes (Figure 4).
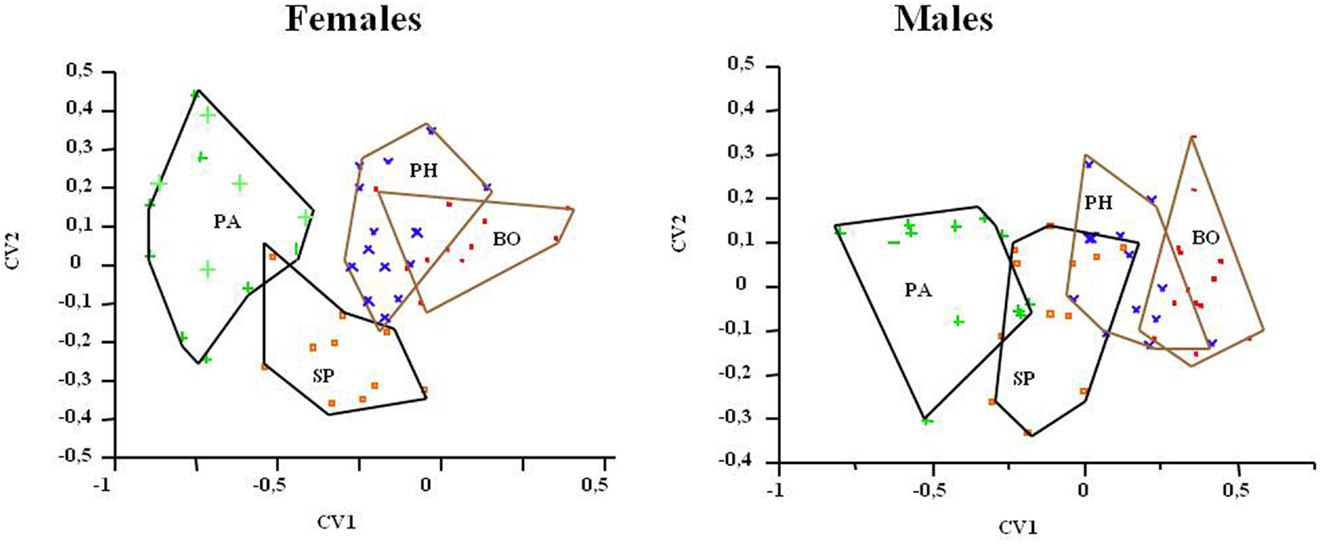
Figure 4. Discriminant analysis (allometry-free), derivate of variables of T. sordida heads in four different populations. The polygons contain the specimens of each T. sordida population. BO, Boquerón Department; PA, Paraguarí Department; PH, Presidente Hayes Department; SP, San Pedro Department.
Feeding Source Analyses
Figure 5 shows the percentages of T. sordida intestinal content that reacted with different vertebrate hosts. The preferred feeding source was varied and included the finding of blood from pets, poultry, and even rodents. The most frequent feeding source of the specimens collected inside and around the houses was poultry blood (hen or chicken): 30% for Gral; Bruguez community of Western Region and 38% for Cerro Guy community of Eastern Region. In the latter, the most frequent second blood source was human (24%) followed by cat and dog blood. In the Western Region, the second frequent blood source was multiple blood (feeding on several animals) where the most common blood mixtures were poultry-human, poultry-rat, and poultry-dog-cat.
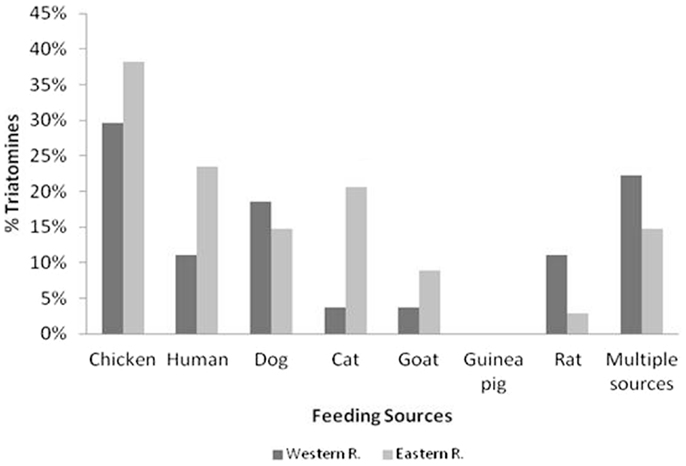
Figure 5. Frequency of blood ingestion for T. sordida in the Eastern and Western Regions is shown. The results of most common vertebrates are included. In the category “Multiple Sources,” the specimens with blood ingestion from several vertebrates are included.
Molecular Analysis
The RAPD profiles were complex (Figure 6A), and the size variation of amplified fragments ranged from 200 to 2500 bp. A total of 98 polymorphic loci generated by four primers were selected for comparative analysis according to their intensity, resolution, and reproducibility. The remarkable polymorphism showed patterns of different bands, but specific patterns were neither observed for insects collected in intra-domicile/peridomicile environments nor for the different departments. The similarity matrix was calculated in accordance with the degree of paired band between each pair of individuals (46) from which the UPGMA tree was generated (Figure 6B). On the other hand, the grouping of the means of allelic frequency showed two particular groups, corresponding to specimens from Western (BO and PH) and Eastern (SP and PA) Regions. The Nei’s genetic distance (1978) indicated a larger separation between the populations separated by larger geographic distance (approximately 520 km), i.e., between BO and PA. The analysis with the POPGENE software showed an estimate of the genetic diversity value (Gst) of 0.08 while the migration index value (Nm) was 5.7 individuals per generation.
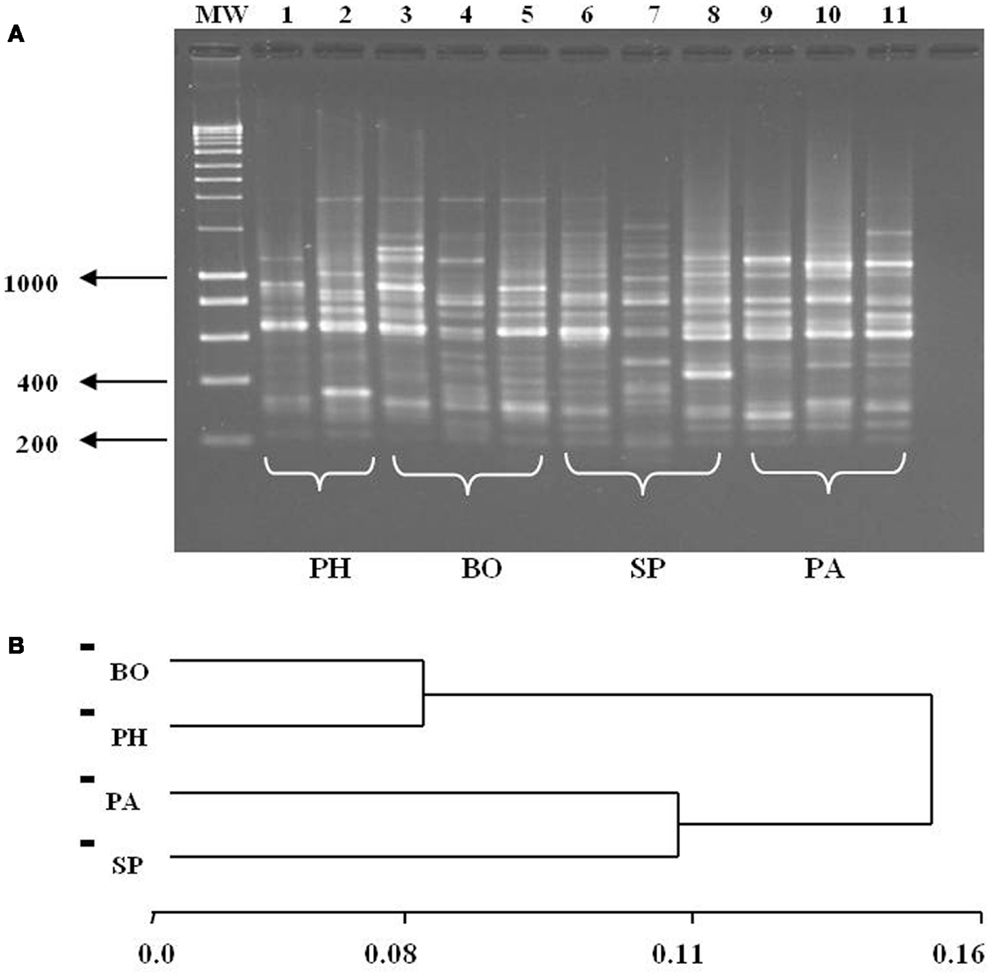
Figure 6. (A) RAPD profiles with DNA extracted from different T. sordida populations using the primer A2. Lanes 1–2: Pte. Hayes (PH), Lanes 3–5: Boquerón (BO), Lanes 6–8: San Pedro (SP), Lanes 9–11: Paraguarí (PA), CN, negative control. PM, Hyperladder I Bioline. (B) Dendrogram of grouping four T. sordida populations by genetic distance based on Nei’s genetic distance. BO, Boquerón; PH, Pte. Hayes; PA, Paraguarí; SP, San Pedro.
Discussion
This is the first report of T. sordida invading and trying to colonize houses in both Paraguayan regions. These areas have been intensively sprayed and a tendency of invasion has been observed when residual insecticide activity ends. Previous reports demonstrated very low rates of domiciliary colonization of T. sordida in Argentina and Brazil (11, 15), but the scenario is quite different in some areas of Bolivia (50).
We have explored the intraspecific relationships among T. sordida populations from different endemic areas for Chagas disease in Paraguay, considering the limited information on their behavior and based on reports about their wide dispersion and high peridomicile infestation (51). A previous study referred that human blood is the second more important feeding source of T. sordida in endemic areas of Paraguay (52), which suggests an increment in the transmission risk of the parasite without the necessity of establishing colonies in rural dwellings. In this study, we still observed human blood as the second feeding source in T. sordida from the Eastern Region but insects captured at the Western Region mainly showed a peridomestic pattern feeding where blood from animals, including sylvatic ones, was detected. Although triatomines were not found positive for T. cruzi infection, this new scenario should be taken into consideration in locations where T. sordida is frequently found inside the houses where T. infestans is absent.
Both morphometric and molecular analyses were carried out to determine the genetic structure of triatomines in order to generate useful information to establish more effective strategies for vector surveillance, incorporating information on their phenotypical variations and sexual dimorphism, excluding changes caused by environmental factors (53) that were corroborated by RAPDs techniques.
Our study did not show any differences in the sexual dimorphism of specimens from peridomicile and domicile, suggesting a continuous exchange with the sylvatic triatomine populations and no transition from sylvatic to domestic habitats or a domiciliation process and adaptation to new habitats were found as it was demonstrated for T. infestans and Panstrongylus geniculatus (53–55). That is to say that in spite of the frequent finding of T. sordida species in the domicile, specific changes related to domiciliation process were not observed, suggesting their temporary presence in dwellings, which was already reported by other authors (56). According to Jaramillo et al. (57), the size of triatomines can be modified in response to environmental changes, thus the significant variation shown by the multivariate analysis of the isometric size in triatomines from Western and Eastern Regions corroborates the influence of micro-environmental conditions, like fourrage disposition in peridomiciles, which leads to the permanency of the insects in such ecotopes. The triatomines from PA department presented significant morphometric variations in relation to the other populations, suggesting a recent adaptation to peridomiciliary ecotopes. Although Chaco populations have conditions for colonization, the triatomines remain in sylvatic ecotopes that are more unstable (58–60). On the other hand, the decrease of size in PA can be attributed to the less favorable modified environments due to the big density of insects and the competition for the nutritional source available in that ecotope (61). The migration of triatomines to peridomicile is produced in response to agricultural habits and destruction of natural forests for anthropic action, even more if we consider that the PA department has been subjected to frequent modifications (agricultural area) or faced control interventions with insecticides. This leads to the dispersion of triatomines and the later adaptation to “new ecotopes”; or simply this adaptation is compatible with the hypothesis of restricted migrations if such ecotopes have enough food sources available (7, 62). According to Dujardin et al. (61), this adaptation to different ecotopes (ecological pressure) is the main mechanism that drives the speciation in the sub family Triatominae. It is important to notice that Chagas Disease Control Program in Paraguay recently showed more frequent domiciliary infestation of T. sordida in several localities of the PA department during the monitoring man/hour search carried out by its technical personnel.
With the elimination of the allometric changes, the DA evidenced the separation of the type morphologies of each region, which can be associated with environmental differences, geographical distances, or the intervention of genetic factors. The significant discrimination observed between the populations of PA and BO suggests a separation due to the distance among both in agreement with what was reported in a previous study made with sensilla patterns between populations of R. prolixus of the Andean area and oriental plains of Colombia (63). On the other hand, the observed overlapping of the factorial map among populations of the same region leads us to think of a process of passive migration.
The genetic variability can be a consequence of the metric differences observed in specimens from different habitats, i.e., that the metric characteristics are almost exclusively under environmental control and the genetic variations could be the result of the contribution of genetic and environmental features (64). The RAPD method showed genetic structuring between T. sordida populations of both geographical regions and the genetic similarity was bigger among populations of the same region, suggesting the existence of a constant gene flow among them. This seems logical but such grouping may be reflecting recent events with few codon changes caused by the adaptation process of sylvatic populations to artificial ecotopes.
The similarity analysis shown by the FST index suggests an exchange among insect populations from neighboring departments that gets reduced among regions. Therefore, the FST estimator shows little genetic differentiation and according to Nei’s classification (1973), this fact seems logical as they are insects of the same species. However, we suppose that the separation between regions and the morphobiometric differences of PA populations could be related to the genetic changes caused by local selective pressures. The grouping observed in the dendrogram could be associated with epidemiological differences in their respective origin focuses, considering that the Western Region is an area with high pressure of triatomine infestation (16, 58). However, to confirm this we suggest increasing the study of T. sordida populations in Paraguay and the use of more sensitive molecular markers to compare these findings with cryptic speciation groups previously described (18, 34).
The dendrogram is similar to the result previously obtained for T. infestans populations that demonstrated allelic differences among neighboring localities, which increase among populations more distant from each other (65). It has been suggested that the genetic isolation by geographical distance greatly contributes to the genetic variability of triatomine populations caused by the passive dispersal of the insects in association with human migrations, resulting in the founder effects and subsequent genetic drift (33). The migration index obtained in this study suggests the mobility of T. sordida between neighboring populations and the results presented suggest a genetic homogeneity between T. sordida from the same region, which is due to the permanent genetic flow between neighboring populations. However, the observed heterogeneity between specimens from Western and Eastern Regions could be associated with the big distances and even with the presence of the Paraguay River as a geographical barrier, which would be in agreement with the separation obtained with the morphometric analyses, i.e., that the differences between PA and BO involves a differentiation process, possibly associated with the eco-geographic isolation by distance and absence of genetic flow. Feeding behavior also confirm differences in these two populations, while triatomines from the Western Regions showed a poultry feeding profile, patterns of the insects from Eastern regions were associated with poultry feeding and human blood feeding profiles in triatomines captured in domicile. Mixed feeding showed an intense mobilization behavior of these triatomines between peridomicile and domicile areas.
It is important to notice that the genetic analysis has shown intraspecific divergences that allow us to think of possible gene variations involved in the shape expression, although we could not discard the possibility that the separation reflected in the dendrogram is influenced by the variation of allelic frequencies of the individuals. In this case, new questions arise and further studies will be required with models of population genetics to obtain better markers related to the infestation risk of potentials triatomines, mainly as a consequence of the control of T. infestans in Paraguay as other secondary vector species such as T. sordida are more frequently detected in the studied region.
Further studies should be focused on how phenetic and genetic variations could be related to the adaptation capacity of these triatomine populations to domicile, increasing their vector potentiality in the transmission of Chagas disease.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work is part of the Master thesis of the author and was conducted with the financial support of PAHO/HDP/HDR/RG/VEN3 223 and FONACIT-G2005000827 projects. We specially would like to thank Marlene Rodriguez for technical help, Dr. Servio Urdaneta for the critical revision of the manuscript, and Dr. Ricardo Gurtler for kindly providing the antisera.
References
1. Galvão C, Carcavallo R, Da Silva Rocha D, Jurberg J. A checklist of the current valid species of the subfamily Triatominae Jeannel, 1919 (Hemiptera, Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa (2003) 36:1–36.
2. Da Rosa J, Rocha C, Gardim S, Pinto M, Mendonça V, Ferreira Filho J, et al. Description of Rhodnius montenegrensis n. sp. (Hemiptera: Reduviidae: Triatominae) from the state of Rondônia, Brazil. Zootaxa (2012) 3478:62–76.
3. Abad-Franch F, Pavan M, Jaramillo N, Palomeque F, Dale C, Chaverra D, et al. Rhodnius barretti, a new species of Triatominae (Hemiptera: Reduviidae) from western Amazonia. Mem Inst Oswaldo Cruz (2013) 108(Suppl I):92–9. doi:10.1590/0074-0276130434
4. Gonçalves T, Teves-Neves S, Mallet J, Carbajal A, Lopes C. Triatoma jatai sp. nov. in the state of Tocantins, Brazil (Hemiptera: Reduviidae: Triatominae). Mem Inst Oswaldo Cruz (2013) 108(4):429–37. doi:10.1590/0074-0276108042013006
5. Klug F. In Reise um die Erde. In den Jahren 1830, 1831, und 1832 ausgefuert von F.J.F. Meyen. Teil 1. Berlin (1834). p. 412.
6. Stål C. Hemiptera species novas descripsit. In: Kongliga Svenska Fregatten Eugenies Resa omkring jorden, III. Zoologi, Insekter (1859). p. 219–98.
7. Canese A. Encuesta sobre vectores de la Enfermedad de Chagas en el Paraguay, años 1979 a 1980. Rev Parag Microbiol (1981) 16(1):7–8.
8. Sánchez Z. Rol Potencial de T. sordida en la enfermedad de Chagas en Paraguay [Tesis de Maestría]. Asunción: IICS, UNA (2011). p. 34–8.
9. Bar ME, Damborsky MP, Oscherov EB, Alvarez BM, Mizdraji G, Avalos G. Household infestation by triatomines and human seroprevalence in Empedrado Department, Corrientes, Argentina. Cad Saude Publica (1997) 13(2):305–12. doi:10.1590/S0102-311X1997000200020
10. Monteiro F, Jurberg J, Lazoski C. Very low levels of genetic variation in natural peridomestic populations of the Chagas disease vector Triatoma sordida (Hemiptera: Reduviidae) in Southeastern Brazil. Am J Trop Med Hyg (2009) 81(2):223–7.
11. Diotaiuti L, Paula OR, Falcão PL, Dias JCP. Avaliação do programa de controle vectorial da doença de Chagas em Minas Gerais, Brasil, com referência especial ao Triatoma sordida. Bol Oficina Sanit Panam (1995) 118(3):211–9.
12. Diotaiuti L, Faria Filho O, Carneiro F, Dias JC, Pires H, Schofield CJ. Aspectos operacionais do controle do Triatoma brasiliensis. Cad Saude Publica (2000) 16:61–7. doi:10.1590/S0102-311X2000000800006
13. Dias J, Bastos C, Araújo E, Mascarenhas A, Netto E, Grassi F, et al. Acute Chagas disease outbreak associated with oral transmission. Rev Soc Brasil Med Trop (2008) 41(3):296–300. doi:10.1590/S0037-86822008000300014
14. Garcia-Zapata M, Marsden P. Control of the transmission of Chagas’ disease in Mambai, Goiás, Brazil (1980-1988). Am J Trop Med Hyg (1992) 46:440–3.
15. Gürtler R, Cecere M, Canale D, Castañera M, Chuit R, Cohen J. Monitoring house reinfestation by vectors of Chagas disease: a comparative trial of detection methods during a four-year follow-up. Acta Trop (1999) 72:213–34. doi:10.1016/S0001-706X(98)00096-5
16. Rojas de Arias A, Abad-Franch F, Acosta N, López E, González N, Zerba E, et al. Post-control surveillance of Triatoma infestans and Triatoma sordida with chemically-baited sticky traps. PLoS Negl Trop Dis (2012) 6:e1822. doi:10.1371/journal.pntd.0001822
17. Panzera F, Hornos S, Pereira J, Cestau R, Canale D, Diotaiuti L, et al. Genetic variability and geographic differentiation among three species of triatomine bugs (Hemiptera: Reduviidae). Am J Trop Med Hyg (1997) 57:732–9.
18. Noireau F, Zegarra M, Ordoñez Y, Gutierrez T, Dujardin JP. Genetic structure of Triatoma sordida (Hemiptera: Reduviidae) domestic populations from Bolivia: application on control interventions. Mem Inst Oswaldo Cruz (1999) 94:347–51. doi:10.1590/S0074-02761999000300011
19. Acosta N, López E, González-Britez N, Fernández MJ, Rojas de Arias A. Perfiles isoenzimáticos de poblaciones de Triatoma infestans de la región oriental y occidental del Paraguay. Mem Inst Investig Cienc Salud (2001) 1(1): 39–41.
20. Forattini O, Rocha D, Silva E, Ferreira O, Rabello E, Pattoli D. Aspectos ecológicos da tripanossomose americana. III. Dispersão local de triatomíneos, com especial referência ao Triatoma sordida. Rev Saude Publica (1971) 5:193–205. doi:10.1590/S0034-89101971000200002
21. Gurgel-Gonçalves R, Ferreira J, Rosa A, Bar M, Galvao O. Geometric morphometrics and ecological niche modelling for delimitation of near-sibling triatomine species. Med Vet Entomol (2011) 25:84–93. doi:10.1111/j.1365-2915.2010.00920.x
22. Borges EC, Dujardin J-P, Schofield CJ, Romanha AJ, Diotaiuti L. Dynamics between sylvatic, peridomestic and domestic populations of Triatoma brasiliensis (Hemiptera: Reduviidae) in Ceará State, Northeastern Brazil. Acta Trop (2005) 93:119–26. doi:10.1016/j.actatropica.2004.10.002
23. Pfeiler E, Bitler B, Ramsey J, Palacios-Cardiel C, Markow T. Genetic variation, population structure, and phylogenetic relationships of Triatoma rubida and T. recurva (Hemiptera: Reduviidae: Triatominae) from the Sonoran Desert, insect vectors of the Chagas’ disease parasite Trypanosoma cruzi. Mol Phylogenet Evol (2006) 41:209–21. doi:10.1016/j.ympev.2006.07.001
24. González-Britez N, Martínez C, Feliciangeli M, Carrasco HJ. Estructura genética de poblaciones domésticas y peridomésticas de Triatoma sordida (Hemiptera: Reduviidae) provenientes de dos regiones endémicas del Paraguay. Arch Venezol Med Trop (2006) 4(1):32–6.
25. Garcia A, Carrasco HJ, Schofield C, Stothard J, Frame I, Valente S, et al. Random amplification of polymorphic DNA as a tool for taxonomic studies of triatomine bugs (Hemiptera: Reduviidae). J Med Entomol (1998) 35:38–45.
26. Monteiro F, Donnelly M, Beard C, Costa J. Nested clade and phylogeographic analyses of the Chagas disease vector Triatoma brasiliensis in Northeast Brazil. Mol Phylogenet Evol (2004) 32:46–56. doi:10.1016/j.ympev.2003.12.011
27. Duarte N, Maciel J, Sosa Z. Atlas de Necesidades Básicas Insatisfechas-Dirección General de Estadística y Censo del Paraguay. Paraguay: DGEE Publicaciones (2005). p. 143–92.
28. Atlas Climático. Fundación para el Desarrollo Sustentable del Chaco & Latin America and the Caribean, U.S. Agency for International Development. Fortalecimiento del Manejo Sustentable de las Ecorregiones Chaco y Pantanal. Cooperative Agrement – Nu 526-00-A-00-00125-00. Asunción: Atlas Climático del Chaco Paraguayo (2005). 92 p.
29. Lent H, Wygodzinsky P. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas disease. Bull Am Mus Nat Hist (1979) 163:123–201.
30. Hoare CA. The Trypanosomes of Mammals: A Zoological Monograph. Oxford: Blackwell Scientific Publications (1972).
31. Chavez T. Tipificación del género Rhodnius mediante la morfometría. Licenciatura en Bioquímica, Tesis. Bolivia: Universidad Mayor Real y Pontificia de San Francisco Xavier de Chuquisaca (1998). p. 13–8.
32. Dujardin JP, Bermudez H, Schofield C. The use of morphometrics in entomological surveillance of sylvatic foci of Triatoma infestans in Bolivia. Acta Trop (1997) 66:145–53. doi:10.1016/S0001-706X(97)00038-7
33. Dujardin JP, Muñoz M, Chavez T, Ponce C, Moreno J, Schofield C. The origin of Rhodnius prolixus in Central America. Med Vet Entomol (1998) 12:113–5. doi:10.1046/j.1365-2915.1998.00092.x
34. Noireau F, Gutiérrez T, Zegarra M, Flores R, Breniere F, Cardozo L, et al. Cryptic speciation in Triatoma sordida (Hemiptera: Reduviidae) from the Bolivian Chaco. Trop Med Int Health (1998) 3:364–72. doi:10.1046/j.1365-3156.1998.00219.x
35. Silveira A, Vinhaes M. Elimination of vector-borne transmission of Chagas disease. Mem Inst Oswaldo Cruz (1999) 94(Suppl 1):405–11. doi:10.1590/S0074-02761999000700080
36. Lyman D, Monteiro F, Escalante A, Cordon-Rosales C, Wesson D, Dujardin J, et al. Mitochondrial DNA sequence variation among triatomine vectors of Chagas’ disease. Am J Trop Med Hyg (1999) 60:377–86.
37. Dujardin JP, Bermudez H, Casini C, Schofield C, Tibayrenc M. Metric differences between sylvatic and domestic Triatoma infestans (Heteroptera: Reduviidae) in Bolivia. J Med Entomol (1997) 34:544–51.
38. Sall J, Jones B. JMP statistical discovery software. Wiley Interdiscip Rev Comput Stat (2011) 3(3):188–94. doi:10.1002/wics.162
39. Rohlf FJ. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, Versión 2.1 p. User Guide. New York: Exceter Software (2001).
40. Gurtler R, Cecere M, Vazquez D, Chuit R, Cohen J. Host-feeding patterns of domiciliary Triatoma infestans (Hemiptera: Reduviidae) in Northwest Argentina: seasonal and instar variation. J Med Entomol (1996) 33: 15–26.
41. Pant CP, Houba V, Henger HD. Bloodmeal identification in vectors. Parasitol Today (1987) 3(11):324–6. doi:10.1016/0169-4758(87)90114-1
42. Correa R, Aguilar AO. Teste de precipitina na identificacao da fonte alimentar do Triatoma infestans (Hemiptera Reduviidae). Arq Hig Saude Publica (1952) 17:3–8.
43. Miller S, Dykes D, Polesky H. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res (1988) 16:1215–7. doi:10.1093/nar/16.3.1215
44. Williams J, Kubelik A, Livak K, Rafalski J, Tingey S. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res (1990) 18:6531–5. doi:10.1093/nar/18.22.6531
45. Carrasco HJ, Frame I, Valente S, Miles MA. Genetic exchange as a possible source of genomic diversity in sylvatic populations of Trypanosoma cruzi. Am J Trop Med Hyg (1996) 54:418–24.
46. Dice LR. Measures of the amount of ecological association between species. Ecology (1945) 26:297–302. doi:10.2307/1932409
47. Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics (1978) 89:583–90.
48. Nei M, Chakraborty R. Genetic distance and electrophoretic identity of proteins between taxa. J Mol Evol (1973) 2:323–8. doi:10.1007/BF01654100
49. Yeh F, Yang R, Boyle T. POPGENE pc, Version 1.31. Edmonton: University of Alberta and Centre for International Forestry Research (1999).
50. Noireau F, Breniere F, Ordonez J, Cardozo L, Morochi W, Gutierrez T, et al. Low probability of transmission of Trypanosoma cruzi to humans by domiciliary Triatoma sordida in Bolivia. Trans R Soc Trop Med Hyg (1997) 91:653–6. doi:10.1016/S0035-9203(97)90508-3
51. Noireau F, Dujardin JP. Flight and nutritional status of sylvatic Triatoma sordida and Triatoma guasayana. Mem Inst Oswaldo Cruz (2001) 96:385–9. doi:10.1590/S0074-02762001000300018
52. González N, Gurtler R, Rojas de Arias A, De Marco R, Cuosiño B. Fuentes de alimentación de triatominos domesticos (Hemiptera-reduviidae) en una localidad endémica para la enfermedad de Chagas. Annual Reports, IICS. Ed. EFACIM. (1997). p. 71–6.
53. Jaramillo ON, Castillo D, Wolff EM. Geometric morphometric differences between Panstrongylus geniculatus from field and laboratory. Mem Inst Oswaldo Cruz (2002) 97:667–73. doi:10.1590/S0074-02762002000500015
54. Dujardin JP, Panzera P, Schofield C. Triatominae as a model of morphological plasticity under ecological pressure. Mem Inst Oswaldo Cruz (1999) 94(Suppl 1):223–8. doi:10.1590/S0074-02761999000700036
55. Dujardin JP, Forgues G, Torrez M, Martinez E, Cordoba C, Gianella A. Morphometrics of domestic Panstrongylus rufotuberculatus in Bolivia. Ann Trop Med Parasitol (1998) 92:219–28. doi:10.1080/00034989860076
56. Gorla D, Jurberg J, Catalá S, Schofield C. Systematics of Triatoma sordida, Triatoma guasayana, and T. patagónica (Hemiptera: Reduviidae). Mem Inst Oswaldo Cruz (1993) 88:379–85. doi:10.1590/S0074-02761993000300006
57. Jaramillo N, Calle D, Harling C, Calle J, Ortega E. Diferencias Morfométricas Asociadas a la Distribución Geográfica de Rhodnius pallescens Provenientes de siete Localidades de Colombia y Panamá. Taller Técnico de Estudio Sobre Rhodnius pallescens, su Vigilancia y Control. Panamá: Publicación deOrganización Panamericana de la Salud OPS (2002). p. 15–23.
58. Canese J, Brice E. Elevado índice de serología positiva para la enfermedad de Chagas en el Chaco Paraguayo. Rev Parag Microbiol (1978) 13:3–9.
59. Rodríguez C, Mora V. Secretaría Técnica de Planificación (STP). Dirección General de Estadísticas, Encuestas y Censos. Paraguay: Atlas Censal-Paraguay (1993). p. 35–76.
60. Rolón M, Vega M, Román F, Gómez A, Rojas de Arias A. First report of colonies of sylvatic Triatoma infestans (Hemiptera: Reduviidae) in the Paraguayan Chaco, using a trained dog. PLoS Negl Trop Dis (2011) 5:e1026. doi:10.1371/journal.pntd.0001026
61. Dujardin JP, Chavez T, Moreno J, Machane M, Noireau F, Schofield C. Comparison of isoenzyme electrophoresis and morphometric analysis for phylogenetic reconstruction of the Rhodniini (Hemiptera: Reduviidae: Triatominae). J Med Entomol (1999) 36:653–9.
62. Forattini O, Ferreira O, Silva E, Rabello E. Aspectos ecológicos da tripanossomíase americana. VI. Persistência do Triatoma sordida pós alteração ambiental e suas possíveis relações com dispersão da espécie. Rev Saude Publica (1974) 6:265–82. doi:10.1590/S0034-89101974000300003
63. Esteban L. Variación fenotípica antenal de poblaciones domésticas de R. prolixus (Hemiptera-Reduviidae) de Colombia. Resúmenes: l XXXII Congreso de la Sociedad Colombiana de Entomología: Julio. Ibagué: Sociedad Colombiana de Entomología (2005).
64. Cassini C, Dujardin J, Martinez A, Bentos A, Salvatella R. Morphometric differentiation between two geographic populations of Triatoma infestans in Uruguay. Res Rev Parasitol (1995) 55:25–30.
Keywords: Triatoma sordida, Chagas disease, RAPD, morphometric analysis, feeding content
Citation: Gonzalez-Britez NE, Carrasco HJ, Martínez Purroy CE, Feliciangeli MD, Maldonado M, López E, Segovia MJ and Rojas de Arias A (2014) Genetic and morphometric variability of Triatoma sordida (Hemiptera: Reduviidae) from the eastern and western regions of Paraguay. Front. Public Health 2:149. doi: 10.3389/fpubh.2014.00149
Received: 18 June 2014; Accepted: 03 September 2014;
Published online: 19 September 2014.
Edited by:
Juan-Carlos Navarro, Universidad Central de Venezuela, VenezuelaCopyright: © 2014 Gonzalez-Britez, Carrasco, Martínez Purroy, Feliciangeli, Maldonado, López, Segovia and Rojas de Arias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hernán J. Carrasco, Laboratorio de Biología Molecular de Protozoarios (N° 224), 1er piso, Instituto de Medicina Tropical, Universidad Central de Venezuela, Los Chaguaramos, Caracas 1050, Venezuela e-mail: hjcarrasco@yahoo.com