Selection of Bacillus spp. for cellulase and xylanase production as direct-fed microbials to reduce digesta viscosity and Clostridium perfringens proliferation using an in vitro digestive model in different poultry diets
 Juan D. Latorre1
Juan D. Latorre1
 Xochitl Hernandez-Velasco2
Xochitl Hernandez-Velasco2
 Vivek A. Kuttappan1
Vivek A. Kuttappan1
 Ross E. Wolfenden3
Ross E. Wolfenden3
 Jose L. Vicente3
Jose L. Vicente3
 Amanda D. Wolfenden1
Amanda D. Wolfenden1
 Lisa R. Bielke1
Lisa R. Bielke1
 Omar F. Prado-Rebolledo4
Eduardo Morales5
Billy M. Hargis1
Omar F. Prado-Rebolledo4
Eduardo Morales5
Billy M. Hargis1  Guillermo Tellez1*
Guillermo Tellez1*
- 1Department of Poultry Science, University of Arkansas, Fayetteville, AR, USA
- 2Departamento de Medicina y Zootecnia de Aves, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autonoma de Mexico, Mexico City, Mexico
- 3Pacific Vet Group-USA, Inc., Fayetteville, AR, USA
- 4Facultad de Medicina Veterinaria y Zootecnia, Universidad de Colima, Colima, Mexico
- 5Departamento de Produccion Agricola y Animal, Universidad Autonoma Metropolitana, Mexico City, Mexico
Previously, our laboratory has screened and identified Bacillus spp. isolates as direct-fed microbials (DFM). The purpose of the present study was to evaluate the cellulase and xylanase production of these isolates and select the most appropriate Bacillus spp. candidates for DFM. Furthermore, an in vitro digestive model, simulating different compartments of the gastrointestinal tract, was used to determine the effect of these selected candidates on digesta viscosity and Clostridium perfringens proliferation in different poultry diets. Production of cellulase and xylanase were based on their relative enzyme activity. Analysis of 16S rRNA sequence classified two strains as Bacillus amyloliquefaciens and one of the strains as Bacillus subtilis. The DFM was included at a concentration of 108 spores/g of feed in five different sterile soybean-based diets containing corn, wheat, rye, barley, or oat. After digestion time, supernatants from different diets were collected to measure viscosity, and C. perfringens proliferation. Additionally, from each in vitro simulated compartment, samples were taken to enumerate viable Bacillus spores using a plate count method after heat-treatment. Significant (P < 0.05) DFM-associated reductions in supernatant viscosity and C. perfringens proliferation were observed for all non-corn diets. These results suggest that antinutritional factors, such as non-starch polysaccharides from different cereals, can enhance viscosity and C. perfringens growth. Remarkably, dietary inclusion of the DFM that produce cellulase and xylanase reduced both viscosity and C. perfringens proliferation compared with control diets. Regardless of diet composition, 90% of the DFM spores germinated during the first 30 min in the crop compartment of the digestion model, followed by a noteworthy increased in the intestine compartment by ~2log10, suggesting a full-life cycle development. Further studies to evaluate in vivo necrotic enteritis effects are in progress.
Introduction
Necrotic enteritis (NE) in broilers is a multi-factorial disease with severe economic implications (1). It is caused by type A strains of Clostridium perfringens that are specific to poultry with toxin types alpha and NetB (2, 3). Coccidia infections are the most common pre-requisite for NE to occur (4), however, dysbacteriosis associated with diet ingredients, changes in feed ration, immunosuppression, Salmonella infections, and/or removal of the use of quimioterapeutics are known to predispose birds to NE (1–5). Antibiotic growth promoters (AGPs) are commonly used to mitigate the incidence of enteric diseases, such as NE. Nevertheless, concerns regarding the development of antibiotic-resistant microorganisms and social pressures have led to a tendency to ban AGPs in poultry production (6). In this scenario, there is an imperative necessity to find feasible alternatives for AGPs to maintain poultry health (7). In fact, the use of selected strains of various beneficial microorganisms from the genus Bacillus and Lactobacillus have shown to be a suitable option for the poultry industry (8). Bacillus spp. are Gram-positive, aerobe, motile, and usually found in soil and water sources, as well as in the gastrointestinal tract of animals and humans (9). Different Bacillus spp. have already been studied and extensively used as a source of industrial enzymes as well as antibiotics by biotechnology companies (10). However, the production of most of these enzymes depends on the intense metabolic changes associated with environmental conditions (11–13). During extreme environmental conditions, vegetative cells of Bacillus spp. form endospores, which are considered, the toughest way of life on Earth (14).
The use of spores from selected Bacillus strains, as direct-fed microbials (DFM), are shown to have the capacity to germinate and sporulate in the gastrointestinal tract of different animal species including poultry. Thus, they become metabolically active in vivo, imparting numerous nutritional benefits including the production of extracellular enzymes, such as protease, lipase, cellulase, xylanase, phytase, and keratinase (15, 16), and other chemical compounds beneficial for the host (17).
In most of the USA and other countries, including Brazil, broiler feed is based primarily on corn and soybean meal. However, sometimes it is difficult to formulate least-cost diets using corn. Consequently, other cereals or ethanol by-products with variable concentrations of antinutritional factors are used as alternatives. When chickens are fed alternative grains with high levels of non-starch polysaccharides (NSP), an increase in digesta viscosity, poor nutrient digestibility, reduced bone mineralization, and occurrence of enteric diseases, such as NE, have been reported (18, 19). Hence, utilization of these feedstuffs in poultry diets usually result in decreased growth performance, intestinal dysbacteriosis, and detrimental litter conditions caused by sticky droppings (20, 21). For that reason, the inclusion of enzymes, such as carbohydrases, is a routine practice in poultry diets that contain grains with elevated NSP concentration values in comparison to corn (22, 23). However, there are inconveniences related to dietary inclusion of some enzymes, due to denaturation and lost of activity under high-pelletization temperatures commonly used in poultry rations. Therefore, the objective of the present study was to perform a selection of Bacillus spp. for cellulase and xylanase production as DFM, and evaluate them on digesta viscosity and C. perfringens proliferation in different poultry diets using an in vitro digestive model. The practical implication of the results will be to utilize cost-effective alternative grains in poultry feed formulation, and at the same time improve digestibility as well as production performance in birds using a more thermostable DFM product.
Materials and Methods
Diets
Five mash soybean-based broiler grower diets containing different cereals, such as corn, wheat, rye, barley, or oat, were used as substrate for bacterial growth during the in vitro digestive model. Experimental diets were formulated to approximate the nutritional requirements of broiler chickens as recommended by the NRC (24), and adjusted to breeder’s recommendations (25). No antibiotics, coccidiostats, or enzymes were added to the feed (Table 1). All diets were autoclaved and confirmed negative for Bacillus spp. spores. Later, these diets were inoculated with the respective spores (108 spores/g of feed) of the Bacillus-DFM candidate according to various treatments.
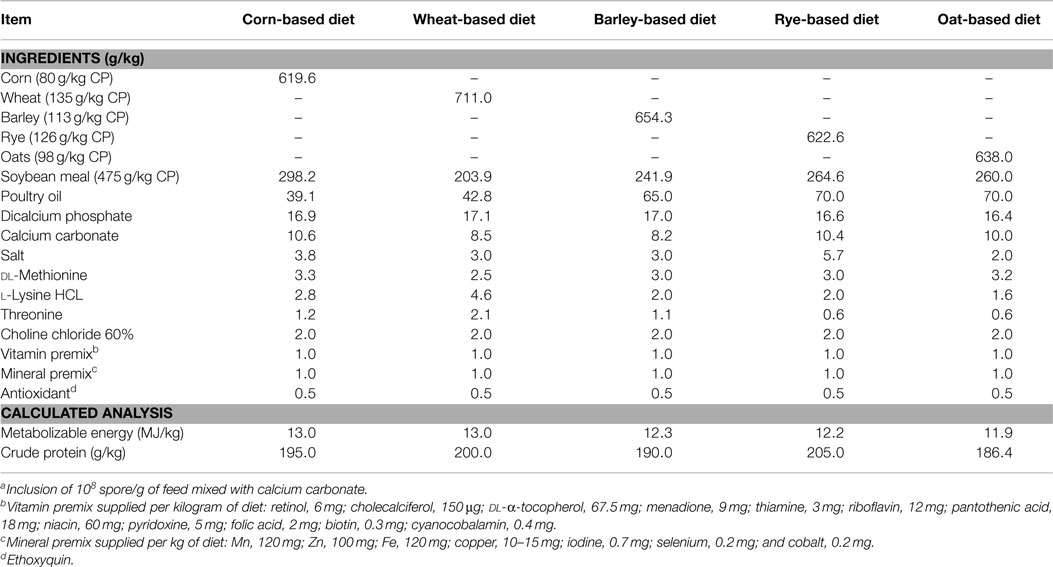
Table 1. Ingredient composition and nutrient content of different broiler chicken diets used for in vitro digestion with or without inclusion of Bacillus-DFM candidate spore on as-is basisa.
In Vitro Assessment of Cellulase and Xylanase Production
Previous research conducted in our laboratory focused on isolation of several Bacillus spp. from environmental and poultry sources (26, 27). Isolates were then screened for production of cellulase and xylanase. For evaluation of cellulase activity, the cellulose-Congo red agar was used and consisted of 0.50 g of K2HPO4 (Fisher Scientific, San Francisco, CA, USA), 0.25 g of MgSO4 (Sigma Chemical Co, St. Louis, MO, USA), 1.88 g of ashed, acid-washed cellulose powder (J. T. Baker Chemical Inc, Phillipsburg, NJ, USA), 0.20 g of Congo red (J. T. Baker Chemical Inc, Phillipsburg, NJ, USA), 20 g of noble agar (Difco Laboratories, Detroit, MI, USA), and 1000 mL of distilled water (15). For evaluation of xylanase activity, the medium used to screen Bacillus isolates contained 3 g of NaNO3, 0.5 g of K2HPO4, 0.2 g of MgSO4⋅7H2O, 0.02 g of MnSO4⋅H2O, 0.02 g of FeSO4⋅H2O, 0.02 g of CaCl2⋅2H2O with 20 g of noble agar (Difco Laboratories, Detroit, MI, USA), and 1000 mL of distilled water. Besides, 1 g yeast extract and 5 g beechwood xylan (Sigma Chemical Co, St. Louis, MO, USA) were used as carbon sources (28). During the screening process, 10 μL of each Bacillus isolate were placed on the center of each plate containing cellulose or xylan media. After 24 h of incubation at 37°C, all plates were evaluated and the diameters of the zones of clearance were measured removing the diameter of the bacterial colony. The relative enzyme activity (REA) was calculated by using the formula: REA = diameter of zone of clearance divided by the diameter of the bacterial colony in millimeters. Based on REA test in each group, organisms were categorized in to excellent (REA > 5.0), good (REA > 2.0 to 5.0), or poor (REA < 2.0) REA (29). Each Bacillus strain was evaluated by triplicate, and the average measurements are presented in Table 2.
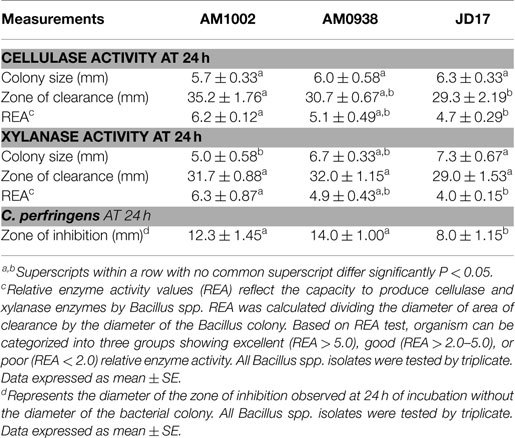
Table 2. Relative enzyme activity values (REA) and Clostridium perfringens zone of inhibition produced by different Bacillus spp. strains present in the Bacillus-DFM candidate treatment.
DFM Culture Identification
Based on the REA results, three Bacillus-DFM candidates with excellent to good REA were selected. These candidates were then identified and characterized using a bioMerieux API 50 CHB test kit (bioMerieux, Marcy l’Etoile, FRA). Individual strain was also subjected to 16S rRNA sequence analysis to a specialized laboratory (Midi labs, Newark, DE, USA). Generally recognized as safe (GRAS) status of these three isolates was affirmed, as described by Wolfenden et al. (30). One of the three Bacillus strains (AM1002) was identified as Bacillus subtilis, and the other two isolates (AM0938 and JD17) were identified as B. amyloliquefaciens (Table 3). Following the identification, all three Bacillus candidate strains were sporulated and mixed in equal amounts during the Bacillus-DFM preparation process as described below and incorporated to the experimental diets.
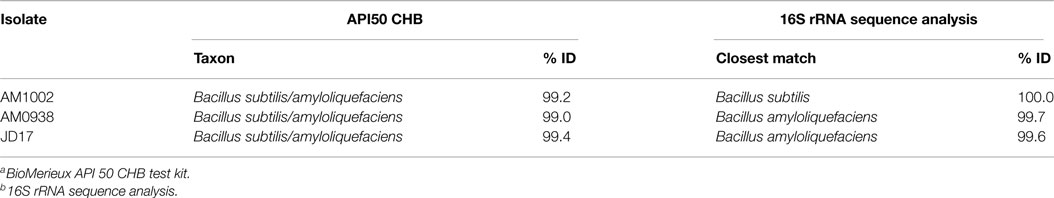
Table 3. Identification of Bacillus spp. isolates by bioMerieux API 50 CHBa and 16S rRNA sequence analysesb present in the Bacillus-DFM candidate treatment.
Preparation of Spore-Based DFM
In an effort to grow high numbers of viable spores, modified version of a solid state fermentation media (SS) developed by Zhao et al. (31) was used. Briefly, to prepare the SS fermentation media, ammonia broth was added to a mixture of 70% rice straw and 30% wheat bran at the rate of 40% by weight. Then, the SS fermentation media was added to 250 mL Erlenmeyer flasks and sterilized by autoclaving for 30 min at 121°C. Each of the three Bacillus strains candidates was grown, individually, overnight at 37°C in test tubes containing 10 mL of tryptic soy broth (TSB, Becton Dickinson, Sparks, MD, USA). After incubation, 2 mL of each candidate culture were added separately to the previously prepared SS fermentation media flasks. The inoculated flasks were incubated for 24 h at 37°C to promote growth of the Bacillus spp. candidates, and incubated for another 72 h at 30°C to trigger the initiation of the sporulation process. Following this, the inoculated SS fermentation media was removed from the Erlenmeyer flasks, placed onto Petri dishes, and dried at 60°C for 18 h. Then, the SS fermentation media was aseptically ground into a fine powder that contained stable Bacillus spores (~1011 spores/g). One gram of spores from each isolate (1:1:1) was combined to produce the Bacillus-DFM candidate final product containing ~3 × 1011 spores/g. Bacillus-DFM candidate was included into each experimental diet to reach a final concentration of 108 spores/g using a rotary mixer for 15 min. Samples of feed containing the DFM candidate were subjected to 100°C for 10 min to eliminate vegetative cells and validate the amount of spores per gram of feed after inclusion and mixing steps. Following heat-treatment, 10-fold dilutions of the same feed samples from the glass tubes were plated on tryptic soy agar plates (TSA, Becton Dickinson, Sparks, MD, USA); letting spores in the feed sample germinate to vegetative cells after incubation at 37°C for 24 h, hence representing the number of spores present per gram of feed.
Clostridium perfringens Strain
A strain of C. perfringens previously described in a NE challenge model was kindly donated by Dr. Jack. L. McReynolds, USDA-ARS, College Station, TX, USA (32). A frozen aliquot was shipped on ice to our laboratory and was amplified in TSB with sodium thioglycolate (Sigma-Aldrich, St Louis, MO, USA). The broth culture was plated on phenylethyl alcohol agar plates (PEA, Becton Dickinson, Sparks, MD, USA) with 5% sheep blood (Remel, Lenexa, KS, USA) to confirm purity, aliquots were made with 25% sterile glycerol and stored at −80°C until further use. A single aliquot was individually amplified in TSB with sodium thioglycolate overnight for the in vitro proliferation studies and the final dose was confirmed by plating 10-fold dilutions on TSA plates with sodium thioglycolate.
In Vitro Assessment of Antimicrobial Activity Against Clostridium perfringens
The three Bacillus isolates present in the Bacillus-DFM candidate treatment were individually cultured aerobically overnight on TSA and screened for in vitro antimicrobial activity against C. perfringens as reported previously (33). Briefly, 10 μL of each Bacillus isolate were placed on the center of TSA plates, and incubated for 24 h at 37°C. Then, the plates with visible Bacillus colonies were overlaid with TSA with sodium thioglycolate containing 106 cfu/mL of C. perfringens, and all plates were incubated anaerobically at 37°C. After 24 h of incubation, all plates were evaluated and the diameters of the zones of inhibition were measured removing the diameter of the bacterial colony. Each Bacillus strain was evaluated by triplicate, and the average measurements of antimicrobial activity against C. perfringens are presented in Table 2.
In Vitro Digestion Assay
The in vitro digestion model used in the present study was based on previous publications, with minor modifications (34, 35), and the assay was performed with five different experimental diets, with or without Bacillus-DFM candidate, in quintuplicates. Briefly, for all the gastrointestinal compartments simulated during the in vitro digestion model, a biochemical oxygen demand incubator (VWR, Houston, TX, USA) set at 40°C (to simulate poultry body temperature), customized with an standard orbital shaker (19 rpm; VWR, Houston, TX, USA) was used for mixing the feed content. Additionally, all tube samples were held at an angle of 30° inclination to facilitate proper blending of feed particles and the enzyme solutions in the tube. The first gastrointestinal compartment simulated was the crop, where 5 g of feed and 10 ml of 0.03M hydrochloric acid (HCL, EMD Millipore Corporation, Billerica, MA, USA) were placed in 50 mL polypropylene centrifuge tubes and mixed vigorously reaching a pH value around 5.2. Tubes were then incubated for 30 min. Following this time, all tubes were removed from the incubator. To simulate the proventriculus as the next gastrointestinal compartment, 3000 U of pepsin per gram of feed (Sigma-Aldrich, St Louis, MO, USA) and 2.5 mL of 1.5M HCl were added to each tube to reach a pH of 1.4–2.0. All tubes were incubated for additional 45 min. The third and the final steps were intended to simulate the intestinal section of the gastrointestinal tract. For that, 6.84 mg of 8× pancreatin (Sigma-Aldrich, St Louis, MO, USA) in 6.5 mL of 1.0M sodium bicarbonate (Sigma-Aldrich, St Louis, MO, USA) were added, and the pH was adjusted to range between 6.4 and 6.8 with 1.0M sodium bicarbonate. All tube samples were further incubated for 2 h. Hence, the complete in vitro digestion process took 3 h and 15 min. After the digestion, supernatants from all the diets were obtained by centrifugation for 30 min at 2000 × g. All supernatants were then tested for viscosity and C. perfringens proliferation, as described below.
Viscosity
Viscosity was measured using a Brookfield digital cone-plate viscometer fitted with a CP-40 spindle (Brookfield Engineering Laboratories Inc., Stoughton, MA, USA). From each supernatant, 0.5 mL was taken to measure viscosity at a shear rate of 42.5/s at 40°C to mimic body temperature of poultry. Viscosity was evaluated by quintuplicate per diet with or without inclusion of the Bacillus-DFM candidate and reported in centipoise (cP = 1/100 dyne s/cm2).
Clostridium perfringens Proliferation
Proliferation of C. perfringens was performed according to previously published methods (35), with minor modifications. A suspension of 105 cfu/mL of C. perfringens was added to five replicates of each of the following groups: (1) 6 mL TSB with sodium thioglycolate as a positive control group; (2) 3 mL TSB with sodium thioglycolate plus 3 mL supernatant from each digested control non-DFM diet; (3) 3 mL TSB with sodium thioglycolate plus 3 mL supernatant from digested diets supplemented with Bacillus-DFM. Samples were incubated anaerobically at 40°C, with tubes set at 30° angle with constant shaking (200 rpm) for 4 h. After incubation, 10-fold serial dilutions were made from all treatment groups in 0.85% sterile saline. Then, 10 μL was plated on TSA with sodium thioglycolate and incubated for 24 h at 40°C, anaerobically. Results were expressed as log10 cfu of C. perfringens/mL.
In Vitro Determination of Spore Persistence
Persistence of the Bacillus-DFM spores in the in vitro digestive model was also evaluated (five replicates per diet treatment). At each time point during the digestive simulation process (crop, proventriculus, and intestine), 0.2 mL was immediately loaded into 0.5 mL sterile centrifuge tubes and heat-treated (pasteurized) at 75°C for 10 min to eliminate the presence of vegetative cells (36). After pasteurization, samples were loaded into sterile 96-well flat bottom plate and 10-fold dilutions were made and plated on TSA. Plates were incubated for 24 h at 37°C on aerobic conditions to enumerate spores per gram of sample.
Statistical Analysis
Data from all measurements were subjected to One-way analysis of variance as a completely randomized design using the General Linear Models procedure of SAS (SAS version 9.1) (37). Means were separated with Duncan’s multiple-range test at P < 0.05 considered as significant. Data were reported as mean ± SE.
Results
Isolates AM1002, AM0938, and JD17 were selected from a pooled of Bacillus isolates in our laboratory, based on the REA values for cellulose and xylanase, and the zone of inhibition for C. perfringens (Table 2). Isolate AM1002 showed a REA value of 6.2 and AM0938 showed a REA value of 5.1, both considered excellent REA values (>5.0) for cellulase activity (29); additionally, isolate JD17 showed a REA value of 4.7, which is considered good (>2.0–5.0) for cellulase production. A similar trend was observed for xylanase activity where isolate AM1002 showed a REA value of 6.3 (excellent); AM0938 showed a REA value of 4.8 (good), and isolate JD17 showed a REA value of 4.0 (good) for xylanase production. In the case of antimicrobial activity against C. perfringens, isolate AM0938 generated the largest diameter of the zone of inhibition with 14 mm, followed by isolates AM1002 and JD17 with 12 and 8 mm, respectively. Although enzyme production and antimicrobial activity were observed for all the isolates, individual differences were evident even in bacteria of the same species (Table 2). The API 50 CHB system characterized all three isolates as B. subtilis/amyloliquefaciens (Table 3). Analysis of 16S rRNA sequence classified two strains (AM0938, JD17) as B. amyloliquefaciens and one of the strains (AM1002) as B. subtilis, which was consistent with the results observed by the carbohydrate fermentation profile of the biochemical test.
The results of the evaluation of digesta viscosity of different diets with or without inclusion of a Bacillus-DFM candidate after in vitro digestion are summarized in Table 4. An evident increase in viscosity was observed in soybean-based diets containing wheat, barley, rye, and oats when compared to corn, being rye, and oat diets with the highest viscosity values. However, it was noteworthy to observe that dietary inclusion of the Bacillus-DFM candidate significantly (P < 0.05) reduced viscosity in all diets containing cereals different to corn in comparison to control diets without DFM inclusion (Table 4).
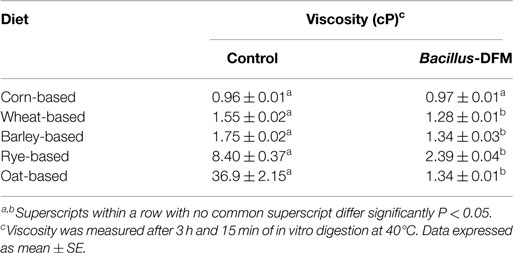
Table 4. Evaluation of in vitro viscosity of different diets with or without inclusion of a Bacillus-DFM candidate.
Table 5 summarizes the results of the proliferation of C. perfringens in the supernatant from different digested diets with or without inclusion of a Bacillus-DFM candidate. A significant increase in C. perfringens proliferation was observed in supernatants collected from control diets that contained wheat, barley, rye, and oat compared to the TSB positive control group. Startlingly, dietary inclusion of a Bacillus-DFM candidate in non-corn diets significantly reduced C. perfringens proliferation when compared to the control non-DFM supplemented diets. The corn-based diet showed similar cfu values of C. perfringens with or without inclusion of the Bacillus-DFM candidate.
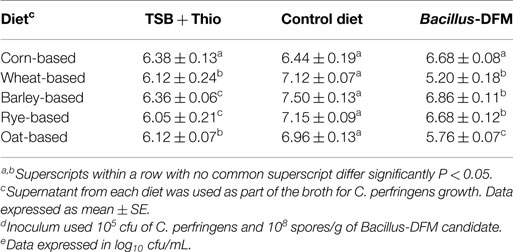
Table 5. Proliferation of Clostridium perfringensd in different digested diets with or without inclusion of Bacillus-DFM candidate sporee.
Persistence of the Bacillus-DFM candidate spores in the different gastrointestinal compartments simulated in the in vitro digestive model is presented in Table 6. Regardless of diet composition, on average, a reduction of more than half of a log10 was observed in the crop compartment during the first 30 min of incubation, and it was followed by a further significant ~2log10 reduction of spore counts in the proventriculus. Remarkably, in all diets, a significant increment in spore numbers, ~2log10 was observed during the final digestion step simulating intestinal conditions (Table 6).
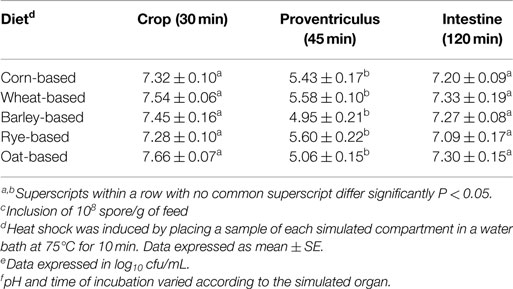
Table 6. Persistence of Bacillus-DFM candidatec spore during in vitro digestionf in different diets under variable biochemical conditions simulating different sections of the gastrointestinal tract of poultrye.
Discussion
High-energy diets have been utilized to maximize growth during starter, grower, and finisher phases of production. Consequently, the primary dietary energy sources in commercial broiler diets have been traditional cereal grains such as corn and sorghum. However, with the recent price volatility of common feed ingredients, the animal industry seeks alternative grains or industry by-products to include in diet formulations (38, 39). Wheat, barley, rye, and oat contain lower bioavailable energy, and elevated NSP levels in comparison to corn (40) are alternative options. However, these cereals have limited use in monogastric diets, because often high-inclusion results in relatively poor performance, detrimental litter conditions, and increase predisposition for NE (41–43). Hence, supplemental carbohydrases, such as NSP-degrading enzymes, have allowed to increase the utilization of these alternative ingredients by reducing their antinutritional effects (40, 44, 45). The carbohydrase market is accounted by two dominant enzymes: xylanases and cellulases. Other commercially available carbohydrases include α-amylase, α-galactosidase, β-glucanase, β-mannanase, and pectinase (46).
In the present study, the Bacillus spp. strains that conform the DFM candidate treatment were identified as either B. subtilis or B. amyloliquefaciens (Table 3), therefore being feasible for in vivo evaluation studies as they are GRAS candidates (12, 26–28). Furthermore, the three selected Bacillus spp. isolates showed a variable ability to produce cellulase and xylanase (Table 2), hence, in addition to the benefits that spores or vegetative cells can provide as probiotics (9, 12), they may improve the digestibility of cereals with high-soluble NSP (47).
The Bacillus-DFM candidate treatment also demonstrated effective antimicrobial properties against C. perfringens, which could be due to production of antimicrobial-like compounds and/or competition for nutrients (Tables 2 and 5). Little is known about the mechanisms underlying the higher incidence of NE in broilers fed diets containing cereals with elevated levels of NSP, but it could be related to a prolonged feed rate of passage and a reduction in the digestion of nutrients that later in the hind gut will be available for bacteria to growth (48). For in vitro evaluation of C. perfringens proliferation, TSB with sodium thioglycolate (positive control) groups were included. In the TSB group (positive control), the C. perfringens inoculum was increased ~0.5log10, after 4 h of incubation. However, it was interesting to observe a significant increase in C. perfringens proliferation in the supernatants collected from control non-DFM diets that contained wheat, barley, rye, and oat, compared with the enrichment TSB medium with sodium thioglycolate group (Table 5).
These results suggest that partial digestion of NSP grains and increased digesta viscosity provides a favorable nutritional environment that supports the growth of C. perfringens. Interestingly, dietary inclusion of a Bacillus-DFM candidate in non-corn diets significantly reduced both viscosity (Table 4) and C. perfringens proliferation (Table 5), when compared to control diets without DFM inclusion. This result shows the capacity of certain Bacillus isolates to inhibit the growth of pathogenic bacteria like C. perfringens, probably due to competition for nutrients, production of antimicrobial-like compounds, or changes in environmental conditions. Proliferation of C. perfringens in the corn-based diet remained constant with or without the inclusion of the Bacillus-DFM candidate and in the TSB positive control group (Table 5). This outcome could be related to the lower concentration of NSP usually found in corn grains in comparison to other cereals, which was also supported by low digesta viscosity values (Table 4). These results are in accordance with previous reports (35), however, it is important to mention that diet ingredients are just one of the multiple predisposing factors that could affect the incidence of NE in commercial conditions (49, 50).
Beneficial bacterial spores are popular as DFM, though little is known about their mode of action. Previous studies conducted in our laboratory, have demonstrated that ~90% of Bacillus spores of a selected strain germinate within 30 min under in vitro and in vivo model conditions, with relatively constant numbers of spores in each gastrointestinal compartment evaluated, hence, suggesting that full-life cycle may occurs (51). In the present study, regardless of the diet, similar in vitro persistence of the Bacillus-DFM candidate spores was observed in the different simulated compartments (Table 6). On average, a half log10 reduction in spore numbers were detected in the crop compartment suggesting spore germination. In the proventriculus compartment, a further ~2log10 reduction was shown, supporting our previous findings (27, 51), which suggest that further germination of spores occurs even at low pH environments. However, it was particularly interesting to observe a ~2log10 increment in spore counts in the intestinal simulated compartment (Table 6). The increment in the numbers of spores could be a response to bacterial metabolites, competition for oxygen and nutrients available, resulting in resporulation (17). The above observations also support previous reports suggesting that spore transiting through the gastrointestinal tract could potentially undergo a full-life cycle of germination and resporulation (36, 52). Moreover, it has been demonstrated that germination of spores into metabolically and functionally active vegetative cells, within a similar time frame, produced beneficial metabolic and immunological effects in different animal species (53–57).
In summary, our results confirm that poultry diets containing cereal grains with a higher content of NSP in comparison to corn can enhance viscosity and C. perfringens growth (18–21). Remarkably, the dietary inclusion of a selected Bacillus-DFM candidate in non-corn-based diets significantly reduced both viscosity and C. perfringens proliferation when compared with the control non-supplemented-diets. Additionally, Bacillus-DFM candidate spore persisted and change their amount according to the variable biochemical conditions of the in vitro digestive model; therefore, supporting the hypothesis of the possible full-life cycle development in the gastrointestinal tract. The results from the present in vitro study encourage us to further evaluate the utilization of this Bacillus-DFM candidate in an in vivo NE model that we have developed in our laboratory (5), as well as to purify, characterize, and measure the international units of cellulase and xylanase that these Bacillus isolates produce. This knowledge will provide a valuable tool to use a stable DFM that produce exogenous enzymes in poultry diets.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Hofacre CL. Necrotic enteritis, currently a billion dollar disease: is there anything new on the horizon? In: Lyons TP, Jacques KA, editors. Science and Technology in the Feed Industry: Proceedings of Alltech’s 17th Annual Symposium. Nottingham: Nottingham University Press (2001). p. 79–86.
2. Keyburn AL, Sheedy SA, Ford ME, Williamson MM, Awad MM, Rood JI, et al. Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens. Infect Immun (2006) 74:6496–500. doi: 10.1128/IAI.00806-06
3. Keyburn AL, Boyce JD, Vaz P, Bannam TL, Ford ME, Parker D, et al. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog (2008) 4:e26. doi:10.1371/journal.ppat.0040026
4. Schoepe H, Pache C, Neubauer A, Potschka H, Schlapp T, Wieler LH, et al. Naturally occurring Clostridium perfringens nontoxic alpha-toxin variant as a potential vaccine candidate against alpha-toxin-associated diseases. Infect Immun (2001) 69:7194–6. doi:10.1128/IAI.69.11.7194-7196.2001
5. Shivaramaiah S, Wolfenden RE, Barta JR, Morgan MJ, Wolfenden AD, Hargis BM, et al. The role of an early Salmonella typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model. Avian Dis (2011) 55:319–23. doi:10.1637/9604-112910-ResNote.1
6. Castanon JI. History of the use of antibiotic as growth promoters in European poultry feeds. Poult Sci (2007) 86:2466–71. doi:10.3382/ps.2007-00249
7. Alvarez-Olmos MI, Oberhelman RA. Probiotic agents and infectious diseases: a modern perspective on a traditional therapy. Clin Infect Dis (2001) 32:1567–76. doi:10.1086/320518
8. Tellez G, Pixley C, Wolfenden R, Layton S, Hargis B. Probiotics/direct fed microbials for Salmonella control in poultry. Food Res Int (2012) 45:628–33. doi:10.1016/j.foodres.2011.03.047
9. Hong HA, Khaneja R, Tam NM, Cazzato A, Tan S, Urdaci M, et al. Bacillus subtilis isolated from the human gastrointestinal tract. Res Microbiol (2009) 160:134–43. doi:10.1016/j.resmic.2008.11.002
10. Kunst F, Ogasawara N, Moszer I, Albertini AM, Alloni G, Azevedo V, et al. The complete genome sequence of the Gram-positive bacterium Bacillus subtilis. Nature (1997) 390:249–56. doi:10.1038/36786
11. González-Pastor JE, Hobbs EC, Losick R. Cannibalism by sporulating bacteria. Science (2003) 301:510–3. doi:10.1126/science.1086462
12. Hong HA, Duc LH, Cutting SM. The use of bacterial spore formers as probiotics. FEMS Microbiol Rev (2005) 29:813–35. doi:10.1016/j.femsre.2004.12.001
13. López D, Vlamakis H, Losick R, Kolter R. Cannibalism enhances biofilm development in Bacillus subtilis. Mol Microbiol (2009) 74:609–18. doi:10.1111/j.1365-2958.2009.06882.x
14. Vreeland RH, Rosenzweig WD, Powers DW. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal. Nature (2000) 407:897–900. doi:10.1038/35038060
15. Hendricks CW, Doyle JD, Hugley B. A new solid medium for enumerating cellulose-utilizing bacteria in soil. Appl Environ Microbiol (1995) 61:2016–9.
16. Sen S, Ingale SL, Kim YW, Kim JS, Kim KH, Lohakare JD, et al. Effect of supplementation of Bacillus subtilis LS 1-2 to broiler diets on growth performance, nutrient retention, caecal microbiology and small intestinal morphology. Res Vet Sci (2012) 93:264–8. doi:10.1016/j.rvsc.2011.05.021
17. Jadamus A, Vahjen W, Simon O. Growth behaviour of a spore forming probiotic strain in the gastrointestinal tract of broiler chicken and piglets. Arch Tierernahr (2001) 54:1–17. doi:10.1080/17450390109381962
18. MacAuliffe T, Pietraszek A, McGinnis J. Variable rachitogenic effects of grain and alleviation by extraction or supplementation with Vitamin D, fat and antibiotics. Poult Sci (1976) 55:2142–7. doi:10.3382/ps.0552142
19. Choct M, Hughes RJ, Wang J, Bedford MR, Morgan AJ, Annison G. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens. Br Poult Sci (1996) 37:609–21. doi:10.1080/00071669608417891
20. Fengler AI, Marquardt RR. Water-soluble pentosans from rye: II. Effects on rate of dialysis and on the retention of nutrients by the chick. Cereal Chem (1988) 65:298–302.
21. Bedford MR, Classen HL. An in vitro assay for prediction of broiler intestinal viscosity and growth when fed rye-based diets in the presence of exogenous enzymes. Poult Sci (1993) 72:137–43. doi:10.3382/ps.0720137
22. Adeola O, Cowieson AJ. Board-invited review: opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J Anim Sci (2011) 89:3189–218. doi:10.2527/jas.2010-3715
23. Slominski BA. Recent advances in research on enzymes for poultry diets. Poult Sci (2011) 90:2013–23. doi:10.3382/ps.2011-01372
24. NRC. Nutrient Requirements of Poultry. 9th rev. ed. Washington, DC: National Academy Press (1994).
25. Cobb-Vantress, Inc. Cobb 500 Broiler Performance and Nutrition Supplement (2013). Available from: http://www.cobb-vantress.com/docs/default-source/cobb-500-guides/cobb500-broiler-performance-nutrition-supplement-(english).pdf
26. Shivaramaiah S, Pumford NR, Morgan MJ, Wolfenden RE, Wolfenden AD, Torres-Rodríguez A, et al. Evaluation of Bacillus species as potential candidates for direct-fed microbials in commercial poultry. Poult Sci (2011) 90:1574–80. doi:10.3382/ps.2010-00745
27. Menconi A, Morgan MJ, Pumford NR, Hargis BM, Tellez G. Physiological properties and Salmonella growth inhibition of probiotic Bacillus strains isolated from environmental and poultry sources. Int J Bacteriol (2013) 2013:1–8. doi:10.1155/2013/958408
28. Monisha R, Uma MV, Murthy VK. Partial purification and characterization of Bacillus pumilus xylanase from soil source. KUSET (2009) 5:137–48.
29. Jani SA, Chudasama CJ, Patel DB, Bhatt PS, Patel HN. Optimization of extracellular protease production from alkali thermos tolerant Actinomycetes: Sacharomonospora viridis SJ-21. Bull Environ Pharmacol Life Sci (2012) 1:84–92.
30. Wolfenden RE, Pumford NR, Morgan MJ, Shivaramaiah S, Wolfenden AD, Pixley CM, et al. Evaluation of selected direct-fed microbial candidates on live performance and Salmonella reduction in commercial turkey brooding houses. Poult Sci (2011) 90:2627–31. doi:10.3382/ps.2011-01360
31. Zhao S, Deng L, Hu N, Zhao B, Liang Y. Cost-effective production of Bacillus licheniformis using simple netting bag solid bioreactor. World J Microbiol Biotechnol (2008) 24:2859–63. doi:10.1007/s11274-008-9820-5
32. McReynolds JL, Byrd JA, Anderson RC, Moore RW, Edrington TS, Genovese KJ, et al. Evaluation of immunosuppressants and dietary mechanisms in an experimental disease model for necrotic enteritis. Poult Sci (2004) 83:1948–52. doi:10.1093/ps/83.12.1948
33. Layton SL, Hernandez-Velasco X, Shivaramaiah C, Xavier J, Menconi A, Latorre JD, et al. The effect of a Lactobacillus-based probiotic for the control of necrotic enteritis in broilers. Food Nutr Sci (2013) 4:1–7. doi:10.4236/fns.2013.411A001
34. Zyla K, Ledoux DR, Garcia A, Veum TL. An in vitro procedure for studying enzymic dephosphorylation of phytate in maize-soybean feeds for turkey poults. Br J Nutr (1995) 74:3–17. doi:10.1079/BJN19950102
35. Annett CB, Viste JR, Chirino-Trejo M, Classen HL, Middleton DM, Simko E. Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A. Avian Pathol (2002) 31:598–601. doi:10.1080/0307945021000024544
36. Barbosa TM, Serra CR, La Ragione RM, Woodward MJ, Henriques AO. Screening for Bacillus isolates in the broiler gastrointestinal tract. Appl Environ Microbiol (2005) 71:968–78. doi:10.1128/AEM.71.2.968-978.2005
38. Friesen OD, Guenter W, Marquardt RR, Rotter BA. The effect of enzyme supplementation on the apparent metabolizable energy and nutrient digestibilities of wheat, barley, oats, and rye for the young broiler chick. Poult Sci (1992) 71:1710–21. doi:10.3382/ps.0711710
39. Kiarie E, Romero LF, Nyachoti CM. The role of added feed enzymes in promoting gut health in swine and poultry. Nutr Res Rev (2013) 26:71–88. doi:10.1017/S0954422413000048
40. Bach Kundsen KE. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim Feed Sci Technol (1997) 67:319–38. doi:10.1016/S0377-8401(97)00009-6
41. Bedford MR, Classen HL, Campbell GL. The effect of pelleting, salt, and pentosanase on the viscosity of intestinal contents and the performance of broilers fed rye. Poult Sci (1991) 70:1571–7. doi:10.3382/ps.0701571
42. Choct M, Hughes RJ, Trimble RP, Angkanaporn K, Annison G. Non-starch polysaccharide-degrading enzymes increase the performance of broiler chickens fed wheat of low apparent metabolizable energy. J Nutr (1995) 125:485–92.
43. Mahmood K, Rahman SU, Hussain I, Abbas RZ, Khaliq T, Arif J, et al. Non-antibiotic strategies for the control of necrotic enteritis in poultry. Worlds Poult Sci J (2014) 70:865–79. doi:10.1017/S0043933914000919
44. Shirzadi H, Moravej H, Shivazad M. Influence of non starch polysaccharide-degrading enzymes on the meat yield and viscosity of jejunal digesta in broilers fed wheat/barley-based diet. Afr J Biotechnol (2010) 9:1517–22. doi:10.5897/AJB09.1483
45. Zou J, Zheng P, Zhang K, Ding X, Bai S. Effects of exogenous enzymes and dietary energy on performance and digestive physiology of broilers. J Anim Sci Biotechnol (2013) 4:14. doi:10.1186/2049-1891-4-14
46. Bedford MR, Schulze H. Exogenous enzymes for pigs and poultry. Nutr Res Rev (1998) 11:91–114. doi:10.1079/NRR19980007
47. Wang ZR, Qiao SY, Lu WQ, Li DF. Effects of enzyme supplementation on performance, nutrient digestibility, gastrointestinal morphology, and volatile fatty acid profiles in the hindgut of broilers fed wheat-based diets. Poult Sci (2005) 84:875–81. doi:10.1093/ps/84.6.875
48. Palliyeguru MWCD, Rose SP. Sub-clinical necrotic enteritis: its aetiology and predisposing factors in commercial broiler production. Worlds Poult Sci J (2014) 70:803–16. doi:10.1017/S0043933914000865
49. Murphy TC, McCracken JK, McCann ME, George J, Bedford MR. Broiler performance and in vivo viscosity as influenced by a range of xylanases, varying in ability to effect wheat in vitro viscosity. Br Poult Sci (2009) 50:716–24. doi:10.1080/00071660903389950
50. Lee KW, Lillehoj HS, Jeong W, Jeoung HY, An DJ. Avian necrotic enteritis: experimental models, host immunity, pathogenesis, risk factors, and vaccine development. Poult Sci (2011) 90:1381–90. doi:10.3382/ps.2010-01319
51. Latorre JD, Hernandez-Velasco X, Kallapura G, Menconi A, Pumford NR, Morgan MJ, et al. Evaluation of germination, distribution, and persistence of Bacillus subtilis spore through the gastrointestinal tract of chickens. Poult Sci (2014) 93:1793–800. doi:10.3382/ps.2013-03809
52. Cartman ST, La Ragione RM, Woodward MJ. Bacillus subtilis spore germinate in the chicken gastrointestinal tract. Appl Environ Microbiol (2008) 74:5254–8. doi:10.1128/AEM.00580-08
53. Hoa TT, Duc LH, Isticato R, Baccigalupi L, Ricca E, Van PH, et al. Fate and dissemination of Bacillus subtilis spore in a murine model. Appl Environ Microbiol (2001) 67:3819–23. doi:10.1128/AEM.67.9.3819-3823.2001
54. La Ragione RM, Woodward MJ. Competitive exclusion by Bacillus subtilis spore of Salmonella enterica serotype enteritidis and Clostridium perfringens in young chickens. Vet Microbiol (2003) 94:245–56. doi:10.1016/S0378-1135(03)00077-4
55. Leser TD, Knarreborg A, Worm J. Germination and outgrowth of Bacillus subtilis and Bacillus licheniformis spore in the gastrointestinal tract of pigs. J Appl Microbiol (2008) 104:1025–33. doi:10.1111/j.1365-2672.2007.03633.x
56. Huang JM, Hong HA, Van Tong H, Hoang TH, Brisson A, Cutting SM. Mucosal delivery of antigens using adsorption to bacterial spore. Vaccine (2010) 28:1021–30. doi:10.1016/j.vaccine.2009.10.127
Keywords: Clostridium perfringens, Bacillus-DFM, spore, enzymes, viscosity
Citation: Latorre JD, Hernandez-Velasco X, Kuttappan VA, Wolfenden RE, Vicente JL, Wolfenden AD, Bielke LR, Prado-Rebolledo OF, Morales E, Hargis BM and Tellez G (2015) Selection of Bacillus spp. for cellulase and xylanase production as direct-fed microbials to reduce digesta viscosity and Clostridium perfringens proliferation using an in vitro digestive model in different poultry diets. Front. Vet. Sci. 2:25. doi: 10.3389/fvets.2015.00025
Received: 11 May 2015; Accepted: 03 August 2015;
Published: 17 August 2015
Edited by:
Ryan Arsenault, University of Delaware, USAReviewed by:
Jason Kindrachuk, Frederick Integrated Research Facility, USARamesh Selvaraj, Ohio Agricultural Research and Development Center, USA
Copyright: © 2015 Latorre, Hernandez-Velasco, Kuttappan, Wolfenden, Vicente, Wolfenden, Bielke, Prado-Rebolledo, Morales, Hargis and Tellez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guillermo Tellez, Department of Poultry Science, Center of Excellence for Poultry Science, University of Arkansas, 1260 West Maple, POSC 0-114, Fayetteville, AR 72701, USA, gtellez@uark.edu