 Rebecca Dumbell
Rebecca Dumbell Olga Matveeva
Olga Matveeva Henrik Oster
Henrik Oster- Chronophysiology Group, Medical Department I, University of Lübeck, Lübeck, Germany
In mammals, molecular circadian clocks are present in most cells of the body, and this circadian network plays an important role in synchronizing physiological processes and behaviors to the appropriate time of day. The hypothalamic–pituitary–adrenal endocrine axis regulates the response to acute and chronic stress, acting through its final effectors – glucocorticoids – released from the adrenal cortex. Glucocorticoid secretion, characterized by its circadian rhythm, has an important role in synchronizing peripheral clocks and rhythms downstream of the master circadian pacemaker in the suprachiasmatic nucleus. Finally, glucocorticoids are powerfully anti-inflammatory, and recent work has implicated the circadian clock in various aspects and cells of the immune system, suggesting a tight interplay of stress and circadian systems in the regulation of immunity. This mini-review summarizes our current understanding of the role of the circadian clock network in both the HPA axis and the immune system, and discusses their interactions.
Introduction
Life on Earth has evolved in the context of a rhythmic environment, characterized largely by the regular succession of night and day. This has led to the evolution of intrinsic circadian (from Latin circa diem – about the day) clock systems, in order to optimally time physiological and behavioral processes. Disruption of circadian timing, such as with inter-time zone travel, shift work, and mistimed eating, can have consequences for cardiovascular, metabolic, and mental health and, crucially, immune function. The hypothalamic–pituitary–adrenal (HPA) axis and the immune system show extensive crosstalk, in particular with regard to the strong anti-inflammatory effects of glucocorticoids (cortisol in humans and corticosterone in rodents). However, less well studied is the interaction of the HPA axis and immune system with regard to the circadian clock. This mini-review will summarize current knowledge regarding the role of the circadian clock in each of these systems, and the interactions that can occur in the context of disrupted circadian rhythmicity.
The Circadian Clock
Circadian rhythms are synchronized to external time by cues known as zeitgebers (German for time givers), such as light and food. In mammals, the clock system is organized in a hierarchical manner, with a master pacemaker residing in the hypothalamic suprachiasmatic nuclei (SCN), acting to synchronize peripheral clocks in all other tissues via endocrine and autonomic signals (1). At the cellular level, circadian clocks coordinate gene expression programs to control physiological processes over the course of the day. The primary zeitgeber for the SCN is light. From the eye, signals are relayed via the retinohypothalamic tract to the SCN, which in turn coordinates peripheral tissue clocks. Other zeitgebers can influence circadian rhythms, with mistimed feeding in particular being able to reset peripheral clocks independent from the SCN (2, 3).
The molecular circadian clock consists of interlocked transcriptional–translational feedback loops [TTLs; discussed in detail elsewhere (4, 5)]. Briefly, during the day, the transcription factors CLOCK (circadian locomotor output cycles kaput) or NPAS2 (neuronal PAS domain-containing protein 2) in complex with BMAL1 (brain and muscle aryl hydrocarbon receptor nuclear translocator-like 1) bind to E-box (enhancer box) promoter elements to drive expression of Period (Per1-3) and Cryptochrome (Cry1/2), along with other clock controlled genes (inset in Figure 1). PER/CRY protein complexes accumulate in the cytoplasm over the day and later relocate into the nucleus where they inhibit the activity of the CLOCK–BMAL1 (or NPAS2–BMAL1) complex. This shuts down Per/Cry transcription during the night. After degradation of nuclear PER/CRY complexes toward the next morning, the inhibition of CLOCK–BMAL1 is released and a new cycle begins.
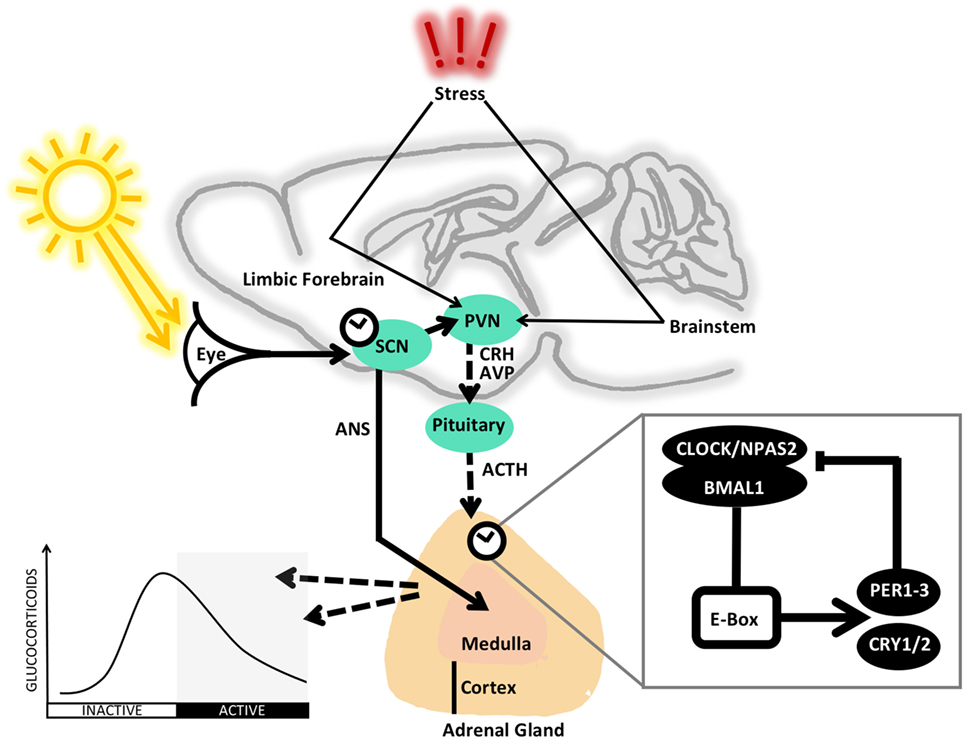
Figure 1. The rhythmic control of the HPA axis is regulated at several levels. The master clock residing in the suprachiasmatic nucleus (SCN) is synchronized by light information received via the retinohypothalamic tract from the eye in order to exert autonomic (ANS) and hormonal influence on the clocks and rhythms of downstream tissues of the body. In addition to the direct innervation of the adrenal, the SCN influences the paraventricular nucleus (PVN) to secrete corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP), which reach the pituitary via the blood portal system to stimulate secretion of adrenocorticotropic hormone (ACTH), which activates production and release of glucocorticoids. In addition, local adrenal clocks are thought to regulate responsiveness to ACTH in a circadian fashion. The baseline circadian rhythm of circulating glucocorticoids peaks just before the beginning of the active phase (day in humans and night in rodents). Stress-induced stimulation of the HPA axis acts via afferent signals from the limbic forebrain and brainstem to the PVN. Inset: the core transcriptional–translational feedback loop (TTL) that makes up the molecular circadian clockwork. In the positive arm of the clock, CLOCK or NPAS2 form a complex with BMAL1 and bind to E-Box elements in the gene promotors of PERs and CRYs, which make up the negative arm and act to inhibit the activity of CLOCK–BMAL1 or NPAS2–BMAL1, with a cycle of roughly 24 h. For further detail, see the main text.
Circadian and Stress Regulation of the HPA Axis
The HPA axis is a key in the regulation of stress responses, with glucocorticoids mediating intermediate and chronic adaptation to stressful stimuli, complementing the rapid response of catecholamines, both secreted from the adrenal gland. The rhythmic regulation of catecholamines and other adrenal hormones is discussed elsewhere (6). Rhythmic regulation of glucocorticoid release (Figure 1) allows for anticipation of daily timing of energy-demanding situations. In addition, glucocorticoid rhythms play a key role in the systemic coordination of circadian rhythms by resetting cellular clocks downstream of the SCN.
During stress, the brainstem and limbic forebrain stimulate corticotrophin-releasing hormone (CRH) and arginine vasopressin (AVP) secretion from neurosecretory neurons of the paraventricular nucleus of the hypothalamus (PVN) (7). Via the hypophyseal portal system, these reach anterior pituitary corticotrophs, which secrete adrenocorticotrophic hormone (ACTH). ACTH then acts at melanocortin type-2 receptors (MC2R) in the adrenal cortex to stimulate production and release of glucocorticoids. Negative feedback from glucocorticoids acts at the level of CRH in the hypothalamus and ACTH in the pituitary (8, 9). In addition, PVN CRH expression is indirectly controlled by the SCN (8–11).
Glucocorticoids bind to glucocorticoid (GR) and mineralocorticoid receptors (MR) in target tissues. Unlike the MR, which is almost constantly activated by glucocorticoids, GR – widely expressed in the brain and periphery, but not in the SCN (12) – activation occurs only during glucocorticoid peak levels (13). This means that GR activation may occur during the peak of the circadian rhythm even during the ultradian trough, but ultradian peaks during the circadian nadir may not be sufficient for activation (14, 15). GR binds to glucocorticoid response elements (GRE or nGRE) to regulate transcription of target genes (16). GRE are present in the promoter region of the clock genes Per1, Per2, Npas2, and various clock controlled genes involved in the synchronization of peripheral circadian rhythms (17).
Glucocorticoid circadian rhythms peak slightly before the onset of the active phase, which is during the night for most rodent species and during the day for humans (18). This rhythm overlays a more dynamic ultradian pattern for both ACTH and glucocorticoid secretion (19) driven by feedback between glucocorticoids and ACTH release at the pituitary (20) and intra-adrenal feedback of glucocorticoids (21). The presence of GR in the adrenal cortex (22, 23) and the demonstrated inhibitory effects of exogenous corticosterone on the ACTH-stimulated corticosteroid synthesis could potentially play a role in local regulation of glucocorticoid secretion in the adrenal gland. Circadian glucocorticoid rhythms can persist independent of the SCN (24–26). Given the influence that circadian rhythmicity of glucocorticoids may have on peripheral clock function, it is perhaps not surprising that the HPA axis, which is acutely activated in stressful situations, is unlikely to be the main driver of the circadian rhythm of these hormones. Indeed, there are several components of the HPA axis which, although they express circadian rhythmicity, do not synchronize well enough to explain downstream endocrine rhythms (3, 10, 27). The circadian influence of the SCN on the HPA axis also occurs through autonomic innervation of the adrenal gland (28, 29), with SCN-dependent rapid induction of Per1 expression being stimulated in the adrenal gland following a light pulse (30). In line with this, splanchnic nerve transection results in dampened circadian glucocorticoid rhythm in rats (31, 32).
Local Regulation of Glucocorticoid Rhythms
A circadian rhythm of steroid release was first demonstrated in isolated Syrian hamster adrenals [Mesocricetus auratus, See Ref. (33)], and rhythmic glucocorticoid concentrations persist under constant peripheral CRH infusion in CRH knockout mice (34), or in hamsters with natural loss of ACTH rhythm (35). Adrenal circadian rhythms can however be altered by ACTH, which stimulates PER1 and BMAL1 expression ex vivo in human tissue (36) and shifts clock rhythms in isolated adrenals of Per2:LUC reporter mice (37).
Circadian clocks within the adrenal gland also play a role in the rhythmic regulation of the HPA axis, both in glucocorticoid production and in sensitivity to ACTH. Robust clock gene expression rhythms have been demonstrated in the adrenal cortex of rodents and primates (38–43), and several steroidogenic genes show circadian expression (31, 40, 44). Work in transgenic mice has shown that those lacking genes of the positive arm of the TTL produce lower levels of corticosterone (45, 46), while those with mutations in the negative arm are chronical hypersecreters (47, 48). Evidence for the importance of adrenocortical clocks in regulating ACTH sensitivity comes from isolated adrenal tissue responses to ACTH, which differs across the day and is very low in mice deficient for Per2 and Cry1 (41) or Bmal1 (46). This is further supported by evidence from primate adrenal explant studies, where knockdown of Cry2 and subsequent downregulation of Bmal1 lead to attenuated ACTH responses (49). Together, these studies suggest that the local adrenal clock is important for regulating the circadian glucocorticoid rhythm independent of systemic influences such as during stress, and may explain the high amplitude of glucocorticoid rhythm in the face of comparably low variations in ACTH concentrations.
HPA Axis Interaction with the Immune System
A bidirectional communication exists between the HPA axis and the immune system (Figure 2). It is well understood that immune cells can activate the HPA axis via cytokines such as tumor necrosis factor-alpha (TNF-α), interleukins (IL-1, IL-6), and the type-I interferons (IFNs) (50–53). Interestingly, some cytokines can activate the HPA axis via different mechanisms. Although primarily acting on the PVN to stimulate CRH release (54–56), they also have direct action at the level of the pituitary and adrenal (57, 58).
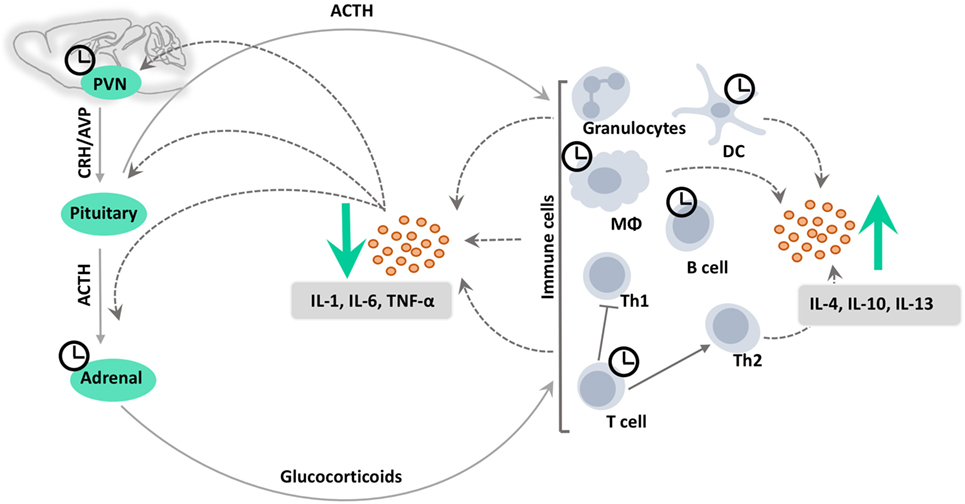
Figure 2. Circadian clocks in HPA axis-immune system crosstalk. Immune cells can activate the HPA axis via cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukins (IL-1/6) at the level of the paraventricular nucleus (PVN) of the hypothalamus as well as at the pituitary and adrenal, stimulating the production of glucocorticoids. Glucocorticoids in turn act on the receptors on the surface or in the cytoplasm of immune cells to suppress the induction of pro-inflammatory responses, and to promote a shift from T helper cell type 1 (Th1) toward T helper cell type 2 (Th2)-mediated humoral immunity. This inhibits the production of pro-inflammatory cytokines, while promoting the production of anti-inflammatory cytokines, such as interleukin-4, interleukin-10, and interleukin-13 (IL-4/10/13) by various immune cells. In addition, ACTH exerts direct anti-inflammatory and immune-modulating effects via the melanocortin system. CRH, corticotropin-releasing hormone; AVP, arginine vasopressin; DC, dendritic cell; MΦ, macrophage.
At the same time, glucocorticoids can affect viability and function of many immune cell types, including T cells, B cells, monocytes, macrophages, and granulocytes (59, 60). Glucocorticoids suppress the synthesis and release of cytokines, thereby protecting the host organism from the detrimental consequences of a long-term hyperactivity of the immune system [reviewed in Ref. (61)]. Pioneering work by Hench, Kendall, and Reichstein demonstrated the immunosuppressive actions of glucocorticoids almost 70 years ago (62). Nevertheless, glucocorticoids are still the most widely used and most effective treatment to control allergic, autoimmune, inflammatory, and hematological disorders (63). GR are found in almost all types of immune cells, and upon activation tether and trans-represses pro-inflammatory regulators such as nuclear factor kappa-light-chain-enhancer of activated B-cells (NF-κB) and activator protein 1 (AP-1) (61, 64) by activating anti-inflammatory molecules such as glucocorticoid-induced-leucine zipper [GILZ (65)], MAPK phosphatase-1 [MKP-1 (66)], annexin-1 (67), mitogen-inducible gene-6 [Mig-6 (68)], and SRC-like adaptor protein 1 [SLAP (69)].
In contrast to the well-described immunosuppressive effects recent studies indicate that glucocorticoids can also have permissive or even stimulatory effects on immune processes [reviewed in Ref. (70–72)]. It is thought that acute stress enhances, while chronic stress suppresses the peripheral immune response (70), but the mechanism of this dual role is not well understood. Several studies report that glucocorticoids induce expression of innate immune-related genes, including members of the toll-like receptor (TLR) family, such as TLR2 and TLR4 (73–75). Glucocorticoids also rapidly induce a central component of the inflammasome, NLRP3, in macrophages, which stimulates secretion of pro-inflammatory cytokines (76).
In addition, glucocorticoids regulate adaptive immune responses by influencing cell trafficking to the sites of inflammation and by suppressing T helper cell type 1 (Th1) and enhancing T helper cell type 2 (Th2) cytokine-driven responses (77, 78). Consequently, in contrast to the traditional view of glucocorticoids as generally immunosuppressive hormones, glucocorticoids are now more accurately regarded as immune modulators.
As an alternative to the glucocorticoid treatment of chronic inflammatory diseases such as multiple sclerosis, the use of ACTH has recently been re-employed, appearing to act not only indirectly by stimulating glucocorticoid production but also by a direct anti-inflammatory effect via the melanocortin system [reviewed in Ref. (79–81)]. Melanocortin receptors (MCRs) are found on lymphocytes and macrophages (82–84). Anti-inflammatory effects of ACTH are mediated primarily by MC1R and MC3R, while immune-regulatory effects rely on MC5R (80). Such glucocorticoid-independent effects of ACTH have been demonstrated after lipo-polysaccharide (LPS)-stimulated production of IL-1ß and TNF-α in human blood samples (85), in a rat gout model (86), and in TNF-α-induced acute kidney disease in rats (87). Furthermore, ACTH can reduce neutrophil infiltration via MC3R (88).
Circadian Interaction of the HPA Axis with Immune Function
In mammals, the circadian clock is an important regulator of the immune system, allowing the organism to anticipate daily changes in activity and the associated risk of antigen encounter. Circadian rhythms are found in multiple aspects of immune function, such as recruitment of immune cells to tissues, antigen presentation, lymphocyte proliferation, TLR function, and cytokine gene expression (89, 90). Furthermore, several inflammatory diseases, such as bronchial asthma and rheumatoid arthritis vary in severity over the course of the day, implicating a circadian regulation of vulnerability (91, 92). Animal studies have revealed that circadian rhythm disruption by shift work or chronic jet lag leads to a dysregulation of the immune system and a higher risk for several pathologies (93, 94).
Molecular clocks have been characterized in various immune cells, including macrophages, dendritic cells, and T and B lymphocytes (95–97). In humans, under constant routine conditions, administration of exogenous glucocorticoids 10 h after awakening can entrain circadian rhythms in peripheral blood mononuclear cells (PBMCs) without changing plasma melatonin and cortisol rhythms, thus linking HPA axis regulation to immune cell function. In line with this, oral administration of synthetic hydrocortisone shifts the expression of BMAL1 and PER2/3 in PBMCs by 9.5–11.5 h (98).
Stress-induced alterations of HPA axis rhythmicity can lead to wide-spread alterations in innate and adaptive immune responses and contribute to the development and progression of some types of cancer in animals (99, 100). In humans, data are less clear, with a positive association between stress and breast cancer observed in some (101), but not in other studies (102). In another context, stress-induced inflammatory priming of microglia was influenced by time of day in rats. Animals exposed to stress during the rest phase showed enhanced neuroinflammatory responses to an LPS challenge compared with animals experiencing stress during the active phase (103). Whether these effects involve circadian alterations remains to be shown. Of interest in this context, pulmonary antibacterial responses in mice appear to be gated by circadian clocks residing in the epithelial club cells lining the pulmonary airways, entrained by glucocorticoids (104).
Not only does the circadian clock regulate the immune system but immune status also feeds back on circadian rhythms. For example, administration of LPS resets activity rhythms in mice (105), and transiently suppresses Per2 and Dbp expression in the SCN and liver of rats (106). LPS treatment increases AVP release from SCN explants (107), and TNF-α treatment downregulates SCN Dbp expression and causes prolonged rest periods during the active phase in rodents (108).
Recent studies reveal blunted circadian cortisol rhythms and attenuated stress responses in patients with allergic diseases, such as bronchial asthma, allergic rhinitis, atopic dermatitis, and extensive nasal polyposis (109–112). In patients with sepsis hypercortisolism persists despite low ACTH levels, suggesting that non-ACTH-mediated mechanisms are involved in the maintenance of high glucocorticoid levels. Interestingly, circadian rhythm of both ACTH and cortisol secretion were shown to be blunted in these patients (113). Furthermore, neonatal endotoxin exposure reprograms HPA axis development in rats, leading to ACTH and corticosterone hyper-responsiveness later in life (114).
Conclusion
Cellular circadian clocks in central and peripheral tissues interact to regulate the activity of the main endocrine axes. Our improved understanding about the systemic regulation of HPA axis circadian rhythms and the interplay of clock and stress functions in this context may help to optimize current treatment strategies for many immunological disorders. For example, in chronic diseases such as rheumatoid arthritis, timed administration of exogenous glucocorticoids at specific times of day may improve therapeutic effectiveness and reduce negative side effects, since lower doses are required (115). At the same time, the importance of glucocorticoids in the coordination of the circadian timing system itself has so far been largely neglected in clinical settings. Stabilizing circadian HPA axis regulation may protect against adverse external influences such as stress and infection, thus protecting the body against some of the most frequent threats of the 24/7 globalized society.
Author Contributions
RD, OM, and HO discussed the concept, compiled the literature, and wrote the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from the Volkswagen Foundation (Lichtenberg Fellowship to HO) and the German Research Association (DFG; SFB-654 and a single project grant to HO).
References
1. Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol (2010) 72:517–49. doi: 10.1146/annurev-physiol-021909-135821
2. Damiola F, Le Minh N, Preitner N, Kornmann B, Fleury-Olela F, Schibler U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev (2000) 14(23):2950–61. doi:10.1101/gad.183500
3. Girotti M, Weinberg MS, Spencer RL. Diurnal expression of functional and clock-related genes throughout the rat HPA axis: system-wide shifts in response to a restricted feeding schedule. Am J Physiol Endocrinol Metab (2009) 296(4):E888–97. doi:10.1152/ajpendo.90946.2008
4. Zhang EE, Kay SA. Clocks not winding down: unravelling circadian networks. Nat Rev Mol Cell Biol (2010) 11(11):764–76. doi:10.1038/nrm2995
5. Brown SA, Kowalska E, Dallmann R. (Re)inventing the circadian feedback loop. Dev Cell (2012) 22(3):477–87. doi:10.1016/j.devcel.2012.02.007
6. Leliavski A, Dumbell R, Ott V, Oster H. Adrenal clocks and the role of adrenal hormones in the regulation of circadian physiology. J Biol Rhythms (2015) 30(1):20–34. doi:10.1177/0748730414553971
7. Ulrich-Lai YM, Herman JP. Neural regulation of endocrine and autonomic stress responses. Nat Rev Neurosci (2009) 10(6):397–409. doi:10.1038/nrn2647
8. Watts AG. Glucocorticoid regulation of peptide genes in neuroendocrine CRH neurons: a complexity beyond negative feedback. Front Neuroendocrinol (2005) 26(3–4):109–30. doi:10.1016/j.yfrne.2005.09.001
9. Dallman MF, Akana SF, Cascio CS, Darlington DN, Jacobson L, Levin N. Regulation of ACTH secretion: variations on a theme of B. Recent Prog Horm Res (1987) 43:113–73.
10. Watts AG, Tanimura S, Sanchez-Watts G. Corticotropin-releasing hormone and arginine vasopressin gene transcription in the hypothalamic paraventricular nucleus of unstressed rats: daily rhythms and their interactions with corticosterone. Endocrinology (2004) 145(2):529–40. doi:10.1210/en.2003-0394
11. Girotti M, Weinberg MS, Spencer RL. Differential responses of hypothalamus-pituitary-adrenal axis immediate early genes to corticosterone and circadian drive. Endocrinology (2007) 148(5):2542–52. doi:10.1210/en.2006-1304
12. Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science (2000) 289(5488):2344–7. doi:10.1126/science.289.5488.2344
13. de Kloet ER, Joels M, Holsboer F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci (2005) 6(6):463–75. doi:10.1038/nrn1683
14. Reul JM, de Kloet ER. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology (1985) 117(6):2505–11. doi:10.1210/endo-117-6-2505
15. Spencer RL, Miller AH, Moday H, Stein M, McEwen BS. Diurnal differences in basal and acute stress levels of type I and type II adrenal steroid receptor activation in neural and immune tissues. Endocrinology (1993) 133(5):1941–50. doi:10.1210/en.133.5.1941
16. Surjit M, Ganti KP, Mukherji A, Ye T, Hua G, Metzger D, et al. Widespread negative response elements mediate direct repression by agonist-liganded glucocorticoid receptor. Cell (2011) 145(2):224–41. doi:10.1016/j.cell.2011.03.027
17. So AY, Bernal TU, Pillsbury ML, Yamamoto KR, Feldman BJ. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc Natl Acad Sci U S A (2009) 106(41):17582–7. doi:10.1073/pnas.0909733106
18. Lightman SL, Conway-Campbell BL. The crucial role of pulsatile activity of the HPA axis for continuous dynamic equilibration. Nat Rev Neurosci (2010) 11(10):710–8. doi:10.1038/nrn2914
19. Dickmeis T, Weger BD, Weger M. The circadian clock and glucocorticoids – interactions across many time scales. Mol Cell Endocrinol (2013) 380(1–2):2–15. doi:10.1016/j.mce.2013.05.012
20. Walker JJ, Spiga F, Waite E, Zhao Z, Kershaw Y, Terry JR, et al. The origin of glucocorticoid hormone oscillations. PLoS Biol (2012) 10(6):e1001341. doi:10.1371/journal.pbio.1001341
21. Walker JJ, Spiga F, Gupta R, Zhao Z, Lightman SL, Terry JR. Rapid intra-adrenal feedback regulation of glucocorticoid synthesis. J R Soc Interface (2015) 12(102):20140875. doi:10.1098/rsif.2014.0875
22. Paust HJ, Loeper S, Else T, Bamberger AM, Papadopoulos G, Pankoke D, et al. Expression of the glucocorticoid receptor in the human adrenal cortex. Exp Clin Endocrinol Diabetes (2006) 114(1):6–10. doi:10.1055/s-2005-873007
23. Loose DS, Do YS, Chen TL, Feldman D. Demonstration of glucocorticoid receptors in the adrenal cortex: evidence for a direct dexamethasone suppressive effect on the rat adrenal gland. Endocrinology (1980) 107(1):137–46. doi:10.1210/endo-107-1-137
24. Moore RY, Eichler VB. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res (1972) 42(1):201–6. doi:10.1016/0006-8993(72)90054-6
25. Stephan FK, Zucker I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci U S A (1972) 69(6):1583–6. doi:10.1073/pnas.69.6.1583
26. Husse J, Leliavski A, Tsang AH, Oster H, Eichele G. The light-dark cycle controls peripheral rhythmicity in mice with a genetically ablated suprachiasmatic nucleus clock. FASEB J (2014) 28(11):4950–60. doi:10.1096/fj.14-256594
27. Henley DE, Russell GM, Douthwaite JA, Wood SA, Buchanan F, Gibson R, et al. Hypothalamic-pituitary-adrenal axis activation in obstructive sleep apnea: the effect of continuous positive airway pressure therapy. J Clin Endocrinol Metab (2009) 94(11):4234–42. doi:10.1210/jc.2009-1174
28. Buijs RM, Wortel J, Van Heerikhuize JJ, Feenstra MG, Ter Horst GJ, Romijn HJ, et al. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur J Neurosci (1999) 11(5):1535–44. doi:10.1046/j.1460-9568.1999.00575.x
29. Engeland WC. Sensitization of endocrine organs to anterior pituitary hormones by the autonomic nervous system. Handb Clin Neurol (2013) 117:37–44. doi:10.1016/B978-0-444-53491-0.00004-3
30. Ishida A, Mutoh T, Ueyama T, Bando H, Masubuchi S, Nakahara D, et al. Light activates the adrenal gland: timing of gene expression and glucocorticoid release. Cell Metab (2005) 2(5):297–307. doi:10.1016/j.cmet.2005.09.009
31. Ulrich-Lai YM, Arnhold MM, Engeland WC. Adrenal splanchnic innervation contributes to the diurnal rhythm of plasma corticosterone in rats by modulating adrenal sensitivity to ACTH. Am J Physiol Regul Integr Comp Physiol (2006) 290(4):R1128–35. doi:10.1152/ajpregu.00042.2003
32. Wotus C, Lilley TR, Neal AS, Suleiman NL, Schmuck SC, Smarr BL, et al. Forced desynchrony reveals independent contributions of suprachiasmatic oscillators to the daily plasma corticosterone rhythm in male rats. PLoS One (2013) 8(7):e68793. doi:10.1371/journal.pone.0068793
33. Andrews RV, Folk GE Jr. Circadian metabolic patterns in cultured hamster adrenal glands. Comp Biochem Physiol (1964) 11:393–409. doi:10.1016/0010-406X(64)90006-4
34. Muglia LJ, Jacobson L, Weninger SC, Luedke CE, Bae DS, Jeong KH, et al. Impaired diurnal adrenal rhythmicity restored by constant infusion of corticotropin-releasing hormone in corticotropin-releasing hormone-deficient mice. J Clin Invest (1997) 99(12):2923–9. doi:10.1172/JCI119487
35. Lilley TR, Wotus C, Taylor D, Lee JM, de la Iglesia HO. Circadian regulation of cortisol release in behaviorally split golden hamsters. Endocrinology (2012) 153(2):732–8. doi:10.1210/en.2011-1624
36. Campino C, Valenzuela FJ, Torres-Farfan C, Reynolds HE, Abarzua-Catalan L, Arteaga E, et al. Melatonin exerts direct inhibitory actions on ACTH responses in the human adrenal gland. Horm Metab Res (2011) 43(5):337–42. doi:10.1055/s-0031-1271693
37. Yoder JM, Brandeland M, Engeland WC. Phase-dependent resetting of the adrenal clock by ACTH in vitro. Am J Physiol Regul Integr Comp Physiol (2014) 306(6):R387–93. doi:10.1152/ajpregu.00519.2013
38. Bittman EL, Doherty L, Huang L, Paroskie A. Period gene expression in mouse endocrine tissues. Am J Physiol Regul Integr Comp Physiol (2003) 285(3):R561–9. doi:10.1152/ajpregu.00783.2002
39. Lemos DR, Downs JL, Urbanski HF. Twenty-four-hour rhythmic gene expression in the rhesus macaque adrenal gland. Mol Endocrinol (2006) 20(5):1164–76. doi:10.1210/me.2005-0361
40. Oster H, Damerow S, Hut RA, Eichele G. Transcriptional profiling in the adrenal gland reveals circadian regulation of hormone biosynthesis genes and nucleosome assembly genes. J Biol Rhythms (2006) 21(5):350–61. doi:10.1177/0748730406293053
41. Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab (2006) 4(2):163–73. doi:10.1016/j.cmet.2006.07.002
42. Fahrenkrug J, Hannibal J, Georg B. Diurnal rhythmicity of the canonical clock genes Per1, Per2 and Bmal1 in the rat adrenal gland is unaltered after hypophysectomy. J Neuroendocrinol (2008) 20(3):323–9. doi:10.1111/j.1365-2826.2008.01651.x
43. Valenzuela FJ, Torres-Farfan C, Richter HG, Mendez N, Campino C, Torrealba F, et al. Clock gene expression in adult primate suprachiasmatic nuclei and adrenal: is the adrenal a peripheral clock responsive to melatonin? Endocrinology (2008) 149(4):1454–61. doi:10.1210/en.2007-1518
44. Son GH, Chung S, Choe HK, Kim HD, Baik SM, Lee H, et al. Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production. Proc Natl Acad Sci U S A (2008) 105(52):20970–5. doi:10.1073/pnas.0806962106
45. Turek FW, Joshu C, Kohsaka A, Lin E, Ivanova G, McDearmon E, et al. Obesity and metabolic syndrome in circadian clock mutant mice. Science (2005) 308(5724):1043–5. doi:10.1126/science.1108750
46. Leliavski A, Shostak A, Husse J, Oster H. Impaired glucocorticoid production and response to stress in Arntl-deficient male mice. Endocrinology (2014) 155(1):133–42. doi:10.1210/en.2013-1531
47. Lamia KA, Papp SJ, Yu RT, Barish GD, Uhlenhaut NH, Jonker JW, et al. Cryptochromes mediate rhythmic repression of the glucocorticoid receptor. Nature (2011) 480(7378):552–6. doi:10.1038/nature10700
48. Barclay JL, Shostak A, Leliavski A, Tsang AH, Johren O, Muller-Fielitz H, et al. High fat diet-induced hyperinsulinemia and tissue-specific insulin resistance in Cry deficient mice. Am J Physiol Endocrinol Metab (2013) 304(10):E1053–63. doi:10.1152/ajpendo.00512.2012
49. Torres-Farfan C, Abarzua-Catalan L, Valenzuela FJ, Mendez N, Richter HG, Valenzuela GJ, et al. Cryptochrome 2 expression level is critical for adrenocorticotropin stimulation of cortisol production in the capuchin monkey adrenal. Endocrinology (2009) 150(6):2717–22. doi:10.1210/en.2008-1683
50. Besedovsky H, del Rey A, Sorkin E, Dinarello CA. Immunoregulatory feedback between interleukin-1 and glucocorticoid hormones. Science (1986) 233(4764):652–4. doi:10.1126/science.3014662
51. van der Meer MJ, Sweep CG, Rijnkels CE, Pesman GJ, Tilders FJ, Kloppenborg PW, et al. Acute stimulation of the hypothalamic-pituitary-adrenal axis by IL-1 beta, TNF alpha and IL-6: a dose response study. J Endocrinol Invest (1996) 19(3):175–82. doi:10.1007/BF03349862
52. Turnbull AV, Rivier CL. Regulation of the hypothalamic-pituitary-adrenal axis by cytokines: actions and mechanisms of action. Physiol Rev (1999) 79(1):1–71.
53. Saphier D. Neuroendocrine effects of interferon-alpha in the rat. Adv Exp Med Biol (1995) 373:209–18. doi:10.1007/978-1-4615-1951-5_29
54. Berkenbosch F, van Oers J, del Rey A, Tilders F, Besedovsky H. Corticotropin-releasing factor-producing neurons in the rat activated by interleukin-1. Science (1987) 238(4826):524–6. doi:10.1126/science.2443979
55. Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W. Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing factor. Science (1987) 238(4826):522–4. doi:10.1126/science.2821621
56. Kovacs KJ, Elenkov IJ. Differential dependence of ACTH secretion induced by various cytokines on the integrity of the paraventricular nucleus. J Neuroendocrinol (1995) 7(1):15–23. doi:10.1111/j.1365-2826.1995.tb00662.x
57. Harbuz MS, Stephanou A, Sarlis N, Lightman SL. The effects of recombinant human interleukin (IL)-1 alpha, IL-1 beta or IL-6 on hypothalamo-pituitary-adrenal axis activation. J Endocrinol (1992) 133(3):349–55. doi:10.1677/joe.0.1330349
58. Ehrhart-Bornstein M, Hinson JP, Bornstein SR, Scherbaum WA, Vinson GP. Intraadrenal interactions in the regulation of adrenocortical steroidogenesis. Endocr Rev (1998) 19(2):101–43. doi:10.1210/edrv.19.2.0326
59. Ashwell JD, Lu FW, Vacchio MS. Glucocorticoids in T cell development and function*. Annu Rev Immunol (2000) 18:309–45. doi:10.1146/annurev.immunol.18.1.309
60. Caramori G, Adcock I. Anti-inflammatory mechanisms of glucocorticoids targeting granulocytes. Curr Drug Targets Inflamm Allergy (2005) 4(4):455–63. doi:10.2174/1568010054526331
61. Vandevyver S, Dejager L, Tuckermann J, Libert C. New insights into the anti-inflammatory mechanisms of glucocorticoids: an emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology (2013) 154(3):993–1007. doi:10.1210/en.2012-2045
62. Hench PS, Kendall EC, Slocumb CH, Polley HF. The effect of a hormone of the adrenal cortex (17-hydroxy-11-dehydrocorticosterone; compound E) and of pituitary adrenocorticotropic hormone on rheumatoid arthritis. Proc Staff Meet Mayo Clin (1949) 24(8):181–97.
63. Rhen T, Cidlowski JA. Antiinflammatory action of glucocorticoids – new mechanisms for old drugs. N Engl J Med (2005) 353(16):1711–23. doi:10.1056/NEJMra050541
64. De Bosscher K, Vanden Berghe W, Haegeman G. Cross-talk between nuclear receptors and nuclear factor kappaB. Oncogene (2006) 25(51):6868–86. doi:10.1038/sj.onc.1209935
65. D’Adamio F, Zollo O, Moraca R, Ayroldi E, Bruscoli S, Bartoli A, et al. A new dexamethasone-induced gene of the leucine zipper family protects T lymphocytes from TCR/CD3-activated cell death. Immunity (1997) 7(6):803–12. doi:10.1016/S1074-7613(00)80398-2
66. Lasa M, Abraham SM, Boucheron C, Saklatvala J, Clark AR. Dexamethasone causes sustained expression of mitogen-activated protein kinase (MAPK) phosphatase 1 and phosphatase-mediated inhibition of MAPK p38. Mol Cell Biol (2002) 22(22):7802–11. doi:10.1128/MCB.22.22.7802-7811.2002
67. Perretti M, D’Acquisto F. Annexin A1 and glucocorticoids as effectors of the resolution of inflammation. Nat Rev Immunol (2009) 9(1):62–70. doi:10.1038/nri2470
68. Xu D, Makkinje A, Kyriakis JM. Gene 33 is an endogenous inhibitor of epidermal growth factor (EGF) receptor signaling and mediates dexamethasone-induced suppression of EGF function. J Biol Chem (2005) 280(4):2924–33. doi:10.1074/jbc.M408907200
69. Park SK, Beaven MA. Mechanism of upregulation of the inhibitory regulator, src-like adaptor protein (SLAP), by glucocorticoids in mast cells. Mol Immunol (2009) 46(3):492–7. doi:10.1016/j.molimm.2008.10.011
70. Dhabhar FS. Stress-induced augmentation of immune function – the role of stress hormones, leukocyte trafficking, and cytokines. Brain Behav Immun (2002) 16(6):785–98. doi:10.1016/S0889-1591(02)00036-3
71. Cruz-Topete D, Cidlowski JA. One hormone, two actions: anti- and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation (2015) 22(1–2):20–32. doi:10.1159/000362724
72. Dhabhar FS. Enhancing versus suppressive effects of stress on immune function: implications for immunoprotection and immunopathology. Neuroimmunomodulation (2009) 16(5):300–17. doi:10.1159/000216188
73. Galon J, Franchimont D, Hiroi N, Frey G, Boettner A, Ehrhart-Bornstein M, et al. Gene profiling reveals unknown enhancing and suppressive actions of glucocorticoids on immune cells. FASEB J (2002) 16(1):61–71. doi:10.1096/fj.01-0245com
74. Homma T, Kato A, Hashimoto N, Batchelor J, Yoshikawa M, Imai S, et al. Corticosteroid and cytokines synergistically enhance toll-like receptor 2 expression in respiratory epithelial cells. Am J Respir Cell Mol Biol (2004) 31(4):463–9. doi:10.1165/rcmb.2004-0161OC
75. Chinenov Y, Rogatsky I. Glucocorticoids and the innate immune system: crosstalk with the toll-like receptor signaling network. Mol Cell Endocrinol (2007) 275(1–2):30–42. doi:10.1016/j.mce.2007.04.014
76. Busillo JM, Azzam KM, Cidlowski JA. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J Biol Chem (2011) 286(44):38703–13. doi:10.1074/jbc.M111.275370
77. Daynes RA, Araneo BA. Contrasting effects of glucocorticoids on the capacity of T cells to produce the growth factors interleukin 2 and interleukin 4. Eur J Immunol (1989) 19(12):2319–25. doi:10.1002/eji.1830191221
78. Agarwal SK, Marshall GD Jr. Dexamethasone promotes type 2 cytokine production primarily through inhibition of type 1 cytokines. J Interferon Cytokine Res (2001) 21(3):147–55. doi:10.1089/107999001750133159
79. Catania A, Gatti S, Colombo G, Lipton JM. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol Rev (2004) 56(1):1–29. doi:10.1124/pr.56.1.1
80. Ross AP, Ben-Zacharia A, Harris C, Smrtka J. Multiple sclerosis, relapses, and the mechanism of action of adrenocorticotropic hormone. Front Neurol (2013) 4:21. doi:10.3389/fneur.2013.00021
81. Montero-Melendez T. ACTH: the forgotten therapy. Semin Immunol (2015) 27(3):216–26. doi:10.1016/j.smim.2015.02.003
82. Johnson EW, Hughes TK Jr, Smith EM. ACTH receptor distribution and modulation among murine mononuclear leukocyte populations. J Biol Regul Homeost Agents (2001) 15(2):156–62.
83. Andersen GN, Hagglund M, Nagaeva O, Frangsmyr L, Petrovska R, Mincheva-Nilsson L, et al. Quantitative measurement of the levels of melanocortin receptor subtype 1, 2, 3 and 5 and pro-opio-melanocortin peptide gene expression in subsets of human peripheral blood leucocytes. Scand J Immunol (2005) 61(3):279–84. doi:10.1111/j.1365-3083.2005.01565.x
84. Cooper A, Robinson SJ, Pickard C, Jackson CL, Friedmann PS, Healy E. Alpha-melanocyte-stimulating hormone suppresses antigen-induced lymphocyte proliferation in humans independently of melanocortin 1 receptor gene status. J Immunol (2005) 175(7):4806–13. doi:10.4049/jimmunol.175.7.4806
85. Catania A, Garofalo L, Cutuli M, Gringeri A, Santagostino E, Lipton JM. Melanocortin peptides inhibit production of proinflammatory cytokines in blood of HIV-infected patients. Peptides (1998) 19(6):1099–104. doi:10.1016/S0196-9781(98)00055-2
86. Getting SJ, Christian HC, Flower RJ, Perretti M. Activation of melanocortin type 3 receptor as a molecular mechanism for adrenocorticotropic hormone efficacy in gouty arthritis. Arthritis Rheum (2002) 46(10):2765–75. doi:10.1002/art.10526
87. Si J, Ge Y, Zhuang S, Wang LJ, Chen S, Gong R. Adrenocorticotropic hormone ameliorates acute kidney injury by steroidogenic-dependent and -independent mechanisms. Kidney Int (2013) 83(4):635–46. doi:10.1038/ki.2012.447
88. Getting SJ, Gibbs L, Clark AJ, Flower RJ, Perretti M. POMC gene-derived peptides activate melanocortin type 3 receptor on murine macrophages, suppress cytokine release, and inhibit neutrophil migration in acute experimental inflammation. J Immunol (1999) 162(12):7446–53.
89. Scheiermann C, Kunisaki Y, Frenette PS. Circadian control of the immune system. Nat Rev Immunol (2013) 13(3):190–8. doi:10.1038/nri3386
90. Geiger SS, Fagundes CT, Siegel RM. Chrono-immunology: progress and challenges in understanding links between the circadian and immune systems. Immunology (2015) 146(3):349–58. doi:10.1111/imm.12525
91. Gibbs JE, Ray DW. The role of the circadian clock in rheumatoid arthritis. Arthritis Res Ther (2013) 15(1):205. doi:10.1186/ar4146
92. Durrington HJ, Farrow SN, Loudon AS, Ray DW. The circadian clock and asthma. Thorax (2014) 69(1):90–2. doi:10.1136/thoraxjnl-2013-203482
93. Castanon-Cervantes O, Wu M, Ehlen JC, Paul K, Gamble KL, Johnson RL, et al. Dysregulation of inflammatory responses by chronic circadian disruption. J Immunol (2010) 185(10):5796–805. doi:10.4049/jimmunol.1001026
94. Knutsson A. Health disorders of shift workers. Occup Med (Lond) (2003) 53(2):103–8. doi:10.1093/occmed/kqg048
95. Silver AC, Arjona A, Hughes ME, Nitabach MN, Fikrig E. Circadian expression of clock genes in mouse macrophages, dendritic cells, and B cells. Brain Behav Immun (2012) 26(3):407–13. doi:10.1016/j.bbi.2011.10.001
96. Keller M, Mazuch J, Abraham U, Eom GD, Herzog ED, Volk HD, et al. A circadian clock in macrophages controls inflammatory immune responses. Proc Natl Acad Sci U S A (2009) 106(50):21407–12. doi:10.1073/pnas.0906361106
97. Bollinger T, Leutz A, Leliavski A, Skrum L, Kovac J, Bonacina L, et al. Circadian clocks in mouse and human CD4+ T cells. PLoS One (2011) 6(12):e29801. doi:10.1371/journal.pone.0029801
98. Cuesta M, Cermakian N, Boivin DB. Glucocorticoids entrain molecular clock components in human peripheral cells. FASEB J (2015) 29(4):1360–70. doi:10.1096/fj.14-265686
99. Reiche EM, Nunes SO, Morimoto HK. Stress, depression, the immune system, and cancer. Lancet Oncol (2004) 5(10):617–25. doi:10.1016/S1470-2045(04)01597-9
100. Ben-Eliyahu S, Page GG, Yirmiya R, Shakhar G. Evidence that stress and surgical interventions promote tumor development by suppressing natural killer cell activity. Int J Cancer (1999) 80(6):880–8. doi:10.1002/(SICI)1097-0215(19990315)80:6<880::AID-IJC14>3.0.CO;2-Y
101. Nielsen NR, Zhang ZF, Kristensen TS, Netterstrom B, Schnohr P, Gronbaek M. Self reported stress and risk of breast cancer: prospective cohort study. BMJ (2005) 331(7516):548. doi:10.1136/bmj.38547.638183.06
102. Protheroe D, Turvey K, Horgan K, Benson E, Bowers D, House A. Stressful life events and difficulties and onset of breast cancer: case-control study. BMJ (1999) 319(7216):1027–30. doi:10.1136/bmj.319.7216.1027
103. Fonken LK, Weber MD, Daut RA, Kitt MM, Frank MG, Watkins LR, et al. Stress-induced neuroinflammatory priming is time of day dependent. Psychoneuroendocrinology (2016) 66:82–90. doi:10.1016/j.psyneuen.2016.01.006
104. Gibbs J, Ince L, Matthews L, Mei J, Bell T, Yang N, et al. An epithelial circadian clock controls pulmonary inflammation and glucocorticoid action. Nat Med (2014) 20(8):919–26. doi:10.1038/nm.3599
105. Marpegan L, Bekinschtein TA, Costas MA, Golombek DA. Circadian responses to endotoxin treatment in mice. J Neuroimmunol (2005) 160(1–2):102–9. doi:10.1016/j.jneuroim.2004.11.003
106. Okada K, Yano M, Doki Y, Azama T, Iwanaga H, Miki H, et al. Injection of LPS causes transient suppression of biological clock genes in rats. J Surg Res (2008) 145(1):5–12. doi:10.1016/j.jss.2007.01.010
107. Nava F, Carta G, Haynes LW. Lipopolysaccharide increases arginine-vasopressin release from rat suprachiasmatic nucleus slice cultures. Neurosci Lett (2000) 288(3):228–30. doi:10.1016/S0304-3940(00)01199-X
108. Cavadini G, Petrzilka S, Kohler P, Jud C, Tobler I, Birchler T, et al. TNF-alpha suppresses the expression of clock genes by interfering with E-box-mediated transcription. Proc Natl Acad Sci U S A (2007) 104(31):12843–8. doi:10.1073/pnas.0701466104
109. Buske-Kirschbaum A, von Auer K, Krieger S, Weis S, Rauh W, Hellhammer D. Blunted cortisol responses to psychosocial stress in asthmatic children: a general feature of atopic disease? Psychosom Med (2003) 65(5):806–10. doi:10.1097/01.PSY.0000095916.25975.4F
110. Wamboldt MZ, Laudenslager M, Wamboldt FS, Kelsay K, Hewitt J. Adolescents with atopic disorders have an attenuated cortisol response to laboratory stress. J Allergy Clin Immunol (2003) 111(3):509–14. doi:10.1067/mai.2003.140
111. Fei GH, Liu RY, Zhang ZH, Zhou JN. Alterations in circadian rhythms of melatonin and cortisol in patients with bronchial asthma. Acta Pharmacol Sin (2004) 25(5):651–6.
112. Fidan V, Alp HH, Kalkandelen S, Cingi C. Melatonin and cortisol rhythm in patients with extensive nasal polyposis. Am J Otolaryngol (2013) 34(1):61–4. doi:10.1016/j.amjoto.2012.09.001
113. Michalaki M, Margeli T, Tsekouras A, Gogos CH, Vagenakis AG, Kyriazopoulou V. Hypothalamic-pituitary-adrenal axis response to the severity of illness in non-critically ill patients: does relative corticosteroid insufficiency exist? Eur J Endocrinol (2010) 162(2):341–7. doi:10.1530/EJE-09-0883
114. Shanks N, Larocque S, Meaney MJ. Neonatal endotoxin exposure alters the development of the hypothalamic-pituitary-adrenal axis: early illness and later responsivity to stress. J Neurosci (1995) 15(1 Pt 1):376–84.
115. Buttgereit F, Doering G, Schaeffler A, Witte S, Sierakowski S, Gromnica-Ihle E, et al. Efficacy of modified-release versus standard prednisone to reduce duration of morning stiffness of the joints in rheumatoid arthritis (CAPRA-1): a double-blind, randomised controlled trial. Lancet (2008) 371(9608):205–14. doi:10.1016/S0140-6736(08)60132-4
Keywords: circadian clock, HPA axis, immune system, glucocorticoids, stress
Citation: Dumbell R, Matveeva O and Oster H (2016) Circadian Clocks, Stress, and Immunity. Front. Endocrinol. 7:37. doi: 10.3389/fendo.2016.00037
Received: 15 February 2016; Accepted: 18 April 2016;
Published: 02 May 2016
Edited by:
Nicole Gallo-Payet, University of Sherbrooke, CanadaReviewed by:
Patrick J. Ronan, Veterans Administration Research, USARobert Spencer, University of Colorado Boulder, USA
John Watson Funder, Prince Henry’s Institute, Australia
Stefan Richard Bornstein, University of Dresden, Germany
Copyright: © 2016 Dumbell, Matveeva and Oster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Henrik Oster, henrik.oster@uni-luebeck.de
†Rebecca Dumbell and Olga Matveeva contributed equally.