Chemical cues mediate species recognition in field crickets
 Frances Tyler
Frances Tyler David Fisher
David Fisher Patrizia d'Ettorre
Patrizia d'Ettorre Rolando Rodríguez-Muñoz2
Rolando Rodríguez-Muñoz2  Tom Tregenza
Tom Tregenza- 1Department of Biological Sciences, University of Cincinnati, Cincinnati, OH, USA
- 2Centre for Ecology and Conservation, College of Life and Environmental Sciences, University of Exeter, Penryn, UK
- 3Laboratoire d'Ethologie Expérimentale et Comparée, Université Paris 13, Villetaneuse, France
Cuticular hydrocarbons (CHCs) are important in mate choice in many insects, and may be used for species recognition if CHC profiles differ between potentially hybridizing species. In the sibling field cricket species Gryllus campestris and G. bimaculatus, females of G. bimaculatus are tolerant toward G. campestris males and can mate with them. However, G. campestris females are highly aggressive toward heterospecific G. bimaculatus males, and matings between them never happen. We examined whether cricket females might use CHCs to determine the species identity of their potential mates. We firstly analyzed the cuticular chemical profile by gas chromatography and mass spectrometry to assess the potential of CHCs to be used for species recognition in these crickets. We then manipulated females' ability to detect chemical cues by carrying out chemical ablation of the antennae, and measured changes in aggressive responses to heterospecific males. We show that there are significant interspecies differences in CHC expression for both sexes, and that females with chemically ablated antennae reduce aggressive behavior toward heterospecific males. Our findings support the prediction that cuticular semiochemicals can play a key role in reproductive isolation between closely related insect species.
Introduction
Mate choice decisions are based upon the traits of potential partners. If gene flow is restricted between two populations, mating traits and preferences may diverge (Panhuis et al., 2001). Eventually, individuals from divergent populations may no longer be recognized as potential mates, at which point speciation has occurred (Mayr, 1942). Information about the traits used in species recognition can be received through a range of sensory modalities, for example using auditory, visual or tactile cues, likely in combination (Hebets and Papaj, 2005). Chemoreception (comprised of olfactory and gustatory systems) is thought to be the most ubiquitous of the sensory modalities (Ache and Young, 2005), and is used for species recognition across a broad range of taxa (reviewed by Smadja and Butlin, 2008). For example, swordtail fish can scent the water around them with olfactory cues, with female Xiphophorus nigrensis more attracted to the cues from conspecific males than to those from heterospecific X. cortezi or X. montezumae (McLennan and Ryan, 1999). In mice, olfactory signals found in urine are used to discriminate between individuals, with Mus musculus musculus preferring the urine of conspecifics to urine of heterospecific M. m. domesticus (Smadja et al., 2004). Wall lizards investigate chemical signals through tongue flicks, with males of both Podarcis bocagei and P. hispanica preferentially investigating the signals of conspecific females (Barbosa et al., 2006). The cuticles of insect species are covered with molecules that can be detected by chemoreceptors. For example male Drosophila mauritania reduce their efforts to court conspecific females that carry chemical signals transferred from heterospecific D. sechellia females (Coyne and Charlesworth, 1997). The males of leaf beetles Pyrrhalta maculicollis and P. aenescens preferentially mate with females bearing conspecific chemical profiles (Zhang et al., 2014). Three species of parasitoid wasps Leptopilina heterotoma, L. boulardi, and L. victoriae use combinations of different chemical cues to identify conspecific mates (Weiss et al., 2015).
Species of true crickets, which have been intensively studied in the context of reproductive isolation (reviewed by Veen et al., 2013), are rarely reported to use chemoreception for species recognition (but see Maroja et al., 2014). Like all other insect species, the exoskeletons of crickets bear lipid molecules known as cuticular hydrocarbons (CHCs). Primarily functioning to prevent desiccation (Gibbs et al., 1997; Gibbs, 1998), they also serve as cues for short-range chemical signaling (Blomquist and Bagnères, 2010; Howard and Blomquist, 2005). There are numerous molecular forms of CHCs, and individuals can gain information about one another through the relative expression of these molecules, primarily detected through the chemosensory hairs covering the antennae. In crickets, CHCs are involved in many aspects of mate choice. Female Gryllodes sigillatus can detect their own chemical signature on males they have already mated with, and can use this self-referent cue to mate polyandrously (Ivy et al., 2005). Male Telelogryllus oceanicus can detect the mating status of females, with non-virgin females carrying the CHCs of males previously mated with (Thomas and Simmons, 2009). Gryllus bimaculatus and T. oceanicus both use CHCs to determine the genetic relatedness of potential mates (Simmons, 1989, 1990; Thomas and Simmons, 2011). G. bimaculatus, Acheta domesticus, G. integer, T. oceanicus, and T. commodus respond to sex differences in chemosensory cues (von Hörmann-Heck, 1957; Otte and Cade, 1976; Rence and Loher, 1977; Hardy and Shaw, 1983; Tregenza and Wedell, 1997; Nagamoto et al., 2005; Leonard and Hedrick, 2009), and in some cases CHC profiles have been shown to differ between the sexes (Warthen and Uebel, 1980; Tregenza and Wedell, 1997; Thomas and Simmons, 2008). Given their general importance in mate choice in crickets, we might also expect CHCs to play an important role in species recognition. There are a few studies implying that this may be the case in G. integer, G. lineaticeps, and Allonemobius species (Otte and Cade, 1976; Paul, 1976; Leonard and Hedrick, 2009), and differences in CHC profiles have been found among species of rapidly diverging Laupala (Mullen et al., 2007, 2008), however, other than Maroja et al. (2014), no studies have explicitly tested this prediction. Here, we investigate the role of chemical cues in species recognition between a pair of European field crickets. We primarily focus upon CHCs, though other chemicals may also be used in chemical communication (Kühbandner et al., 2012).
G. campestris and G. bimaculatus are sister species (Huang et al., 2000) that are considerably divergent; their morphologies differ such that they are easily identifiable (von Hörmann-Heck, 1957), and interspecies matings in the laboratory rarely result in viable offspring (Tyler et al., 2013b; Veen et al., 2013). They have overlapping distributions through southern Europe and further east (Popov and Shuvalov, 1977; Pardo et al., 1993; Gorochov and Llorente, 2001). In Spain G. bimaculatus are distributed along the Mediterranean coast occupying relatively arid conditions (Gorochov and Llorente, 2001). They can be found sheltering under rocks or logs, or in grass, and are predominantly nocturnal. They are capable of flight, and are multivoltine, producing many generations through the year. G. campestris occupy less arid conditions, and are more common in northern and central Spain (Gorochov and Llorente, 2001). They are more abundant in grass lands, where they dig burrows in which to shelter. They are territorial and flightless, which limits dispersal. They are diurnal and univoltine. Despite these differences between the species, their sequences of behaviors leading up to and during mating are similar. The males produce long-range calling song to attract mates. Once a female has encountered a male, the male produces a courtship song that is distinct from the calling song (Alexander, 1961). They touch antennae, and explore the surface of each other's bodies with their antennae, through which they receive both mechanosensory and chemosensory information (Balakrishnan and Pollack, 1997). The male then presents himself to the female, and the female mounts the male to begin copulation.
The characteristics of the long range calling songs differ between the species. Despite these differences, females only show weak (or no) preference for conspecific song. Female G. bimaculatus show a moderate increase in turning effort toward synthetic song typical of conspecific males, whereas G. campestris females show no such change in turning effort (Veen et al., 2013). Instead, close range species recognition seems to play an important role (Veen et al., 2011, 2013). Though G. bimaculatus females preferentially mate with conspecific males, they will frequently engage in hybrid matings in the laboratory, and only very rarely show aggressive behavior toward males of either species. In contrast, G. campestris females will very rarely mate with heterospecifics (Cousin, 1933; von Hörmann-Heck, 1957; Veen et al., 2011, 2013). Despite regular attempts by G. bimaculatus males to court G. campestris females, we have only ever observed the females responding indifferently or aggressively, evidence of discrimination against heterospecific males in mate choice (Veen et al., 2011, 2013). In contrast, when presented with a conspecific male, these females will readily mount and mate. This discrimination at close-range may be achieved through a number of sensory modalities, with detection of species-specific chemical cues a likely contender.
We firstly assess the potential of CHCs to be used for species recognition in these crickets through comparison of CHC profiles using gas chromatography coupled with mass spectrometry, predicting both sex and species differences among individuals. Given that females are the choosy sex in these crickets, sexual selection is likely to be acting on the males. If sexual selection has driven divergence, we might expect to find greater species differences between the males than between the females. We then aim to confirm that G. campestris females discriminate between conspecific and heterospecific males using aggressive behavior to indicate species recognition, and finally, determine whether their ability to detect chemical cues alters the extent of aggression toward heterospecific males. We manipulate females' ability to detect chemical cues through chemical ablation of the antennae, a method that prevents chemoreception without impairing mechanoreception (Balakrishnan and Pollack, 1997).
Methods
Study Population
CHCs were sampled from 94 laboratory reared G. campestris (49 male, 45 female), and 72 laboratory reared G. bimaculatus (38 male, 34 female). G. campestris were the first generation offspring of adults collected near Gijon, northern Spain, in spring 2013. G. bimaculatus were from a population that had been maintained under laboratory conditions for 2 years, originally collected near Valencia, southern Spain. Lab populations were kept at 28°C under a 16:8 light dark cycle, with food and water provided ad libitum. Females were provided with damp sand to lay eggs in, the progeny from which were reared to adulthood. All individuals were kept until death, after which they were preserved at −20°C. Behavioral assays were carried out with laboratory reared G. bimaculatus from the same population and with wild-caught G. campestris collected in spring 2012 and transferred to the laboratory at least 2 weeks prior to use in trials. CHC extraction and behavioral trials were carried out on different groups of individuals due to the logistics of working with wild-caught G. campestris. However, individuals were collected only one generation apart, and from the same location. All individuals were sexually mature, and had not mated for at least 7 days. Female G. campestris will re-mate within ~1–3 days (Tyler et al., 2013a), and so a delay of at least 7 days is ample for them to overcome the influence that a previous mating may have on their behavior.
CHC Extraction and Analysis
38 G. bimaculatus males, 34 G. bimaculatus females, 49 G. campestris males and 46 female G campestris were assessed. To extract the cuticular hydrocarbons we completely submerged each cricket in 4 ml of HPLC grade hexane. The hexane contained dodecane as an internal standard at a concentration of 10 ppm. The crickets were soaked for 5 min before being removed. 2 μl of each sample was injected into a GC-MS [Agilent 7890A gas chromatograph coupled with an Agilent 5975B mass spectrometer with electron impact ionization (70eV)]. This was fitted with a DB-1 ms column (30 × 0.25 mm × 0.25 μm); helium (99.9% research grade) was used as a carrier gas. The inlet was set at 250°C, and the injection was run in constant flow mode at 1.1 ml/min. Separation of the extract was optimized by using an oven program starting at 100°C for 1 min, increasing at 20°C/min to 250°C, then increasing at 5°C/min to 320°C. It was then held at 320°C for 2 min. The MS transfer line was kept at 300°C. Data were analyzed using MSD Chemstation software version E.02.00.493 (Agilent Technologies).
Forty-four CHCs were quantified. Compounds were identified on the basis of their retention time and mass spectra, the diagnostic ions of each compound are given in Table 1. Relative peak size was calculated by dividing the peak areas of a given compound by the peak area of the internal standard in that sample (Table 2). These relative peak sizes were normalized with a log transformation, and principal components analyses (PCA) were run (JMP v9). We used a correlation matrix to extract PCs with eigenvectors greater than 1 (Norman and Streiner, 1984) (Table 3). PC factor loadings greater than 0.25, or less than -0.25, were considered biologically important (Tabachnick and Fidell, 1989) (Table 4). We also calculated diagnostic power (DP) for each of the peaks. The standardized peak areas were used to calculate a standard deviation across all of the individuals, and a pooled standard deviation within each sex or species. The global standard deviation was divided by the pooled standard deviation to obtain a DP value (Table 4) (Christensen et al., 2005; van Zweden et al., 2009). We tested for species and sex differences in CHC expression using a multivariate analysis of variance (MANOVA) in R v3.0.2 (R Core Team, 2013). The first four PCs of CHC expression were entered as response variables, and species, sex, and the species:sex interaction entered as fixed effects (Table 5).
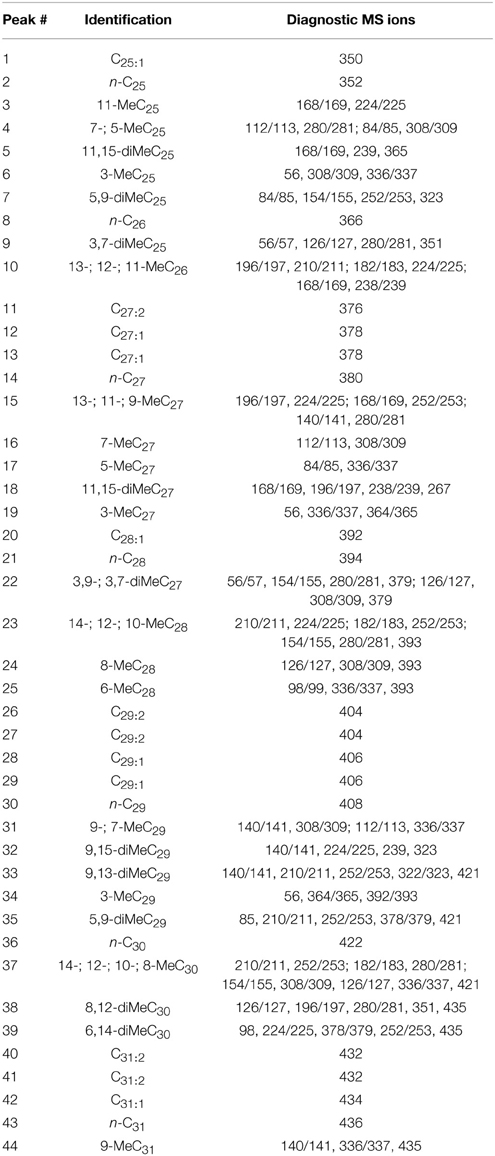
Table 1. CHC profile, displaying hydrocarbon identity and diagnostic ions.
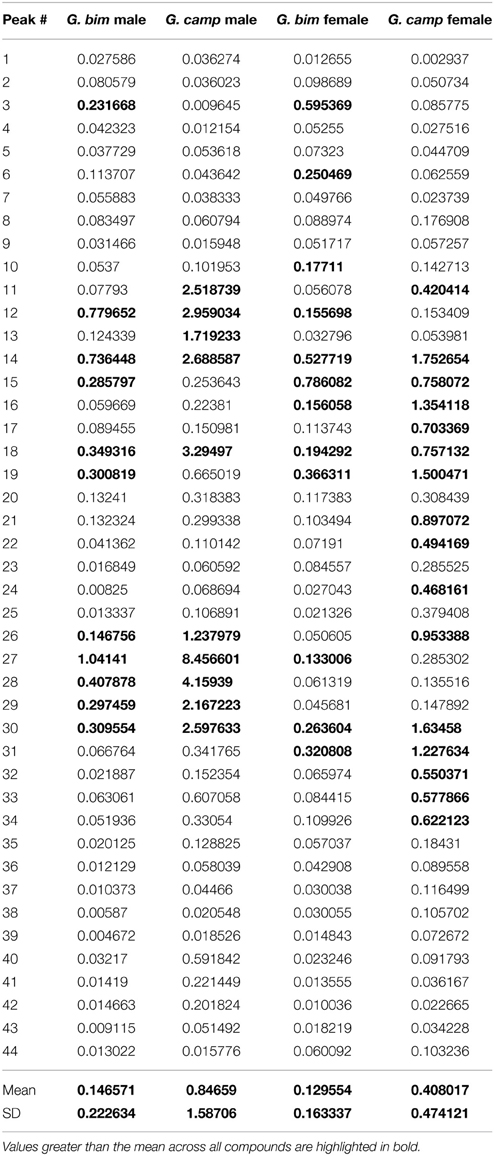
Table 2. Amount of each compound relative to the internal standard, individuals split by species and by sex.
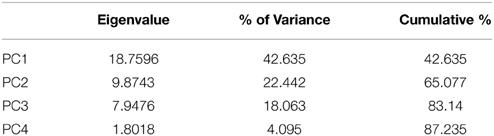
Table 3. Principle component analysis of CHCs.

Table 4. Factor loadings for each of the peaks in the 4 leading PCs (factor loadings of >0.25 are highlighted in bold), and diagnostic power (compounds with DP values greater than the mean across all compounds are highlighted in bold).
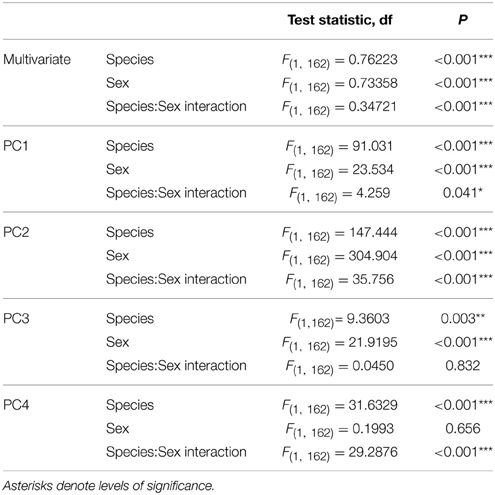
Table 5. Species and sex differences in CHC expression, shown as output from the multivariate ANOVA, as well as the univariate contribution of each principal component.
Behavioral Assays
Males were silenced prior to use in behavioral trials so that the females had to use cues other than song to assess species identity of the males. To do this they were anesthetized by exposing them to CO2 gas until they stopped moving (~20 s), and the small sections of the wings which are used to create song (the file and scraper) were removed with fine dissection scissors, leaving the wings predominantly intact. The procedure was carried out at least 24 h before use in trials.
Behavioral trials were carried out in 20 cm ⊘ plastic arenas, which were cleaned between trials. A silenced male, of either G. bimaculatus or G. campestris, and a G. campestris female were placed either side of a temporary divider. They were allowed to settle for at least 1 min before the divider was removed. The trial started when the pair made first physical contact (including antennal contact), after which they were observed for 5 min (trials were terminated if the female mounted the male, and the pair immediately separated to prevent mating). The number of times the female flared her mandibles at the male was recorded, as well as the number of seconds spent performing aggressive behavior. Aggressive behavior was defined as antennal fencing, mandible flaring, biting, or attempts to attack the male (Alexander, 1961; Wilson et al., 2010). Where possible, for each female two pre-treatment trials were carried out, one with a heterospecific male and one with a conspecific male, in either order, allowing us to confirm that female aggression is dependent upon male species rather than being directed toward all males.
After the pre-treatment trials, females were assigned to chemical ablation or control treatment groups. Balakrishnan and Pollack (1997) demonstrated, through the use of electrophysical assays and tactile reflex tests, that immersion of cricket antennae in zinc sulfate solution eliminates responsiveness of chemosensory hairs, without affecting tactile reflex of the antennae. Whereas the chemosensory sensilla have pores through which the zinc sulfate can reach neurons, the sensilla of mechanoreceptors are poreless, and are therefore unaffected. To ensure that both groups included a similar range of aggressive behavior, we matched individuals according to the number of flares performed during their pre-treatment trial with the heterospecific male, and divided these pairs across the groups. Females were anesthetized using CO2 gas to prevent them from moving during antennal immersion. They were held upside down in a plastic tube and suspended so that their antennae were immersed in the solution, but it did not touch their head or body. The antennae of the females in the chemical ablation group were immersed in 0.4 mol/l of zinc sulfate (ZnSO·7H2O, Sigma Aldrich) dissolved in 0.3% Triton-X (Sigma Aldrich). Triton-X reduces surface tension, allowing penetration though the fine hairs that cover the antennae. The antennae of females in the control group were immersed in 0.3% Triton-X (Balakrishnan and Pollack, 1997; Ryan and Sakaluk, 2009). Antennae were soaked for 10 min, after which they were washed clean with water.
Finally, a post-treatment aggression trial was carried out using a heterospecific male so that we could measure the change in female response toward heterospecific males, and how this change differed between treatment groups. Where possible, the heterospecific male used in the post-treatment trial was the same as for the pre-treatment trial. Some males were paired with more than one female. The post-treatment trial was not carried out until the day following antennal treatment to allow the female to recover.
A total of 44 G. campestris females were used in trials. Any female that did not behave aggressively toward the male during her pre-treatment trial was removed from the dataset. One of the females was extremely aggressive, and her trials were terminated early to prevent the male from being killed. Data for this female were also removed, leaving data for 27 females in the dataset. These 27 females were exposed to 24 G. bimaculatus males (3 males used more than once), and 20 G. campestris males (4 males used more than once). 3 of the females were not exposed to a conspecific male in the pre-treatment trials, due to availability of crickets (results do not change if these females are omitted).
Behavioral data were analyzed in R. We firstly made a comparison of females' pre-treatment responses to conspecific and heterospecific males using Wilcoxon signed-rank tests. We then used paired t-tests to analyze a female's change in aggression (pre-treatment score compared to post-treatment score) for each of the treatment groups. This analysis was firstly conducted using mandible flares as the measure of aggression, and then repeated with seconds spent performing aggressive behavior as the measure.
Results
CHC Profiling
The PCA across all individuals returned 4 PCs, with eigenvectors that cumulatively explained 87.2% of the variance (Table 3) (for PCAs split by species and by sex, see the Supplementary Material). The majority of factor loadings in PC1 were of a magnitude that is likely to be biologically important (Tabachnick and Fidell, 1989) and positively loaded (35 positive, 1 negative), suggesting that overall investment in CHCs may contribute to much of the variation in expression among the species and sexes. There were biologically important factor loadings spread over PC2 and PC3, with no clear pattern in positive or negative loadings, suggesting that investment in particular combinations of CHCs is more important than total investment (Table 4). Overall, there was a significant interaction between species and sex in the expression of CHCs [MANOVA: Approx F(1, 8) = 0.347, P = <0.001, Figure 1]. The difference between the species was driven by all 4 PCs. The difference between the sexes was only driven by PCs 1, 2, and 3 (Table 5). Based on values of diagnostic power, we found a subset (48%) of the CHCs to be >2.17 times more variable between species (mean DP for species) than within species. Many of these compounds with high DP were dimethyl alkanes or n-alkanes. Another subset of CHCs was found to be >2.04 (mean DP for sex) more variable between sexes than within sexes, most of which were monoenes, n-alkanes or methyl alkanes, but no dienes. These are most likely to be used in recognition of species differences, or sex differences, respectively. Of the CHCs likely to play a role in either species or sex recognition, 29.5% were found to occur in both subsets, and may therefore be generally important in communicating chemosensory information about individuals. These were predominantly n-alkanes.
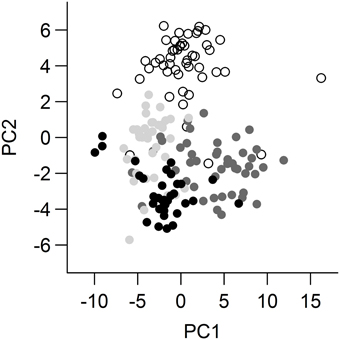
Figure 1. First and second principal components from statistical analysis of cuticular hydrocarbon profiles. G. campestris males shown in white, G. bimaculatus males shown in light gray, G. campestris females shown in dark gray G. bimaculatus females shown in black.
Behavioral Assays
G. campestris females behaved far more aggressively toward heterospecific than conspecific males, with mandible flares rarely being directed toward conspecific males [V(24) = 300, P < 0.001, Figure 2]. Females with chemically ablated antennae reduced the number of times they flared their mandibles at heterospecific males [t(15) = 2.42, P = 0.028] whereas control females did not [t(10) = 0.39, P = 0.71, Figure 3]. When aggression was measured as the number of seconds spent performing aggressive behavior, females were not found to reduce their level of aggression in either of the treatment groups [chemically ablated: t(15) = 2.02, P = 0.061, control: t(10) = 0.23, P = 0.83].
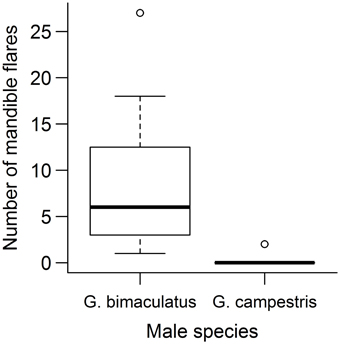
Figure 2. Female aggression toward heterospecific and conspecific males, prior to chemical treatment of their antennae. G. bimaculatus males are heterospecific, G. campestris males are conspecific. Aggression was measured as the number of times that a female flared her mandibles at a male during a 5 min trial. Boxes show the upper and lower quartiles, and central lines show medians. Whiskers show maximum and minimum values, excluding outliers. Dots show outliers.
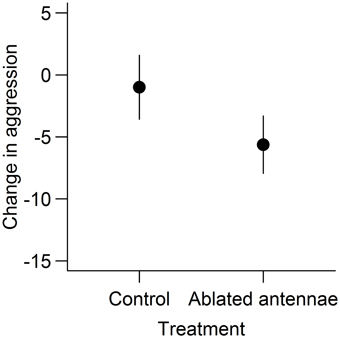
Figure 3. The change in aggression after female antennae were treated with a control or chemically ablated. Change in aggression is presented as the change in counts of mandible flares performed by the G. campestris females at G. bimaculatus males between a pre-treatment and post-treatment trial. Data are presented as means ± standard errors.
Discussion
We show that there are significant species differences in CHC expression for both sexes, and that species identity could be determined on the basis of an individual's CHC profile. We confirm that female G. campestris behave more aggressively toward heterospecific than conspecific males, and that females incapable of receiving chemosensory information via their antennae reduce aggressive behavior toward heterospecific males.
Since these species tend to be found in different habitats, it is possible that initial divergence in CHC expression was due to adaptation to the environment. This may have been a relatively fast process, for example, divergence in CHCs among Laupala populations is rapid, with cryptic species on different islands having different profiles (Mullen et al., 2007). We found that a large proportion of the CHCs produced by these crickets have the potential to be used for species discrimination, and many of these were identified as dimethyl alkanes and long-chain hydrocarbons. These heavier, less volatile compounds may be important in desiccation resistance, and so it is plausible that initial divergence could have been driven through adaptation to the different temperatures and humidities that these species experience. As well as divergence through natural selection, sexual selection may have driven divergence in CHC expression between the species. We predicted that males, as the less choosy sex, might be subject to sexual selection and have greater differences in CHC expression than females (Chenoweth and Blows, 2005; Rundle et al., 2005). Indeed, we found a large proportion of CHCs likely be important is sex recognition. Many of these were identified as monoenes and n-alkanes, and were found spread across the CHC profile. Despite showing weak (or no) discrimination between long-range calling songs (Veen et al., 2013), G. campestris females almost never accept heterospecific mating opportunities (Cousin, 1933; von Hörmann-Heck, 1957; Veen et al., 2011, 2013). This suggests that short range cues are important to females in assessing potential mates, exerting selection on males to strengthen these cues.
Although we have identified species differences in CHC expression, we are unable to rule out the possibility that it is compounds other than CHCs that drive the bias in aggressive behavior toward heterospecifics. Future studies could establish this link more conclusively by demonstrating that the extracts taken from the crickets do in fact elicit the aggressive behavior. The chemical identity of males could be manipulated by washing away their CHCs and/or substituting CHCs from other individuals (for example, Tregenza and Wedell, 1997; Simmons et al., 2003; Nagamoto et al., 2005). Furthermore, extracts could be fractionated to isolate the CHC from other cuticular compounds, followed by demonstrations that it is this fraction eliciting aggression (Liang and Silverman, 2000; Akino et al., 2004).
There is uncertainty as to whether the reduction in aggression was due to an impaired ability to assess male species, or whether the loss of chemosensory information as a whole led females to become indifferent to any cricket. If the latter then we might expect chemically ablated females to respond indifferently to conspecific males, as well as reducing aggression to heterospecific males. Although not explicitly studied, we observed some of these females mating with conspecific males after the trials, indicating that they were not indifferent to them. Ryan and Sakaluk (2009) found that chemically ablated female G. sigillatus took longer to respond to courting males, but would still mount them. This indicates that while there is a suppression of response to males, they are not indifferent to them. This supports the notion that the reduced aggression observed in our study is not due to females being entirely indifferent toward the males. Rather, the reduced aggression is likely due to chemically ablated females no longer sensing the species identity of the males.
The discrepancy in the results obtained from the two measures of aggression is intriguing. While counts of mandible flares revealed a significant difference between the treated and control groups, this difference was only a trend when measured as time spent performing aggressive behaviors. The discrepancy is likely due to the inaccuracy of the latter measurement. A cricket performs each behavior for only a short time, making the capture of this information difficult, introducing measurement error. In future studies this inaccuracy could be overcome by making video recordings of behavioral trials. Additionally, the interpretation of the behaviors was not always clear. Though we had a pre-defined list of behaviors categorized as aggressive, there is variation in how each of these behaviors is performed, and there may be cases of non-aggressive behavior being considered as aggressive and vice versa. Of all the behaviors categorized as aggressive, mandible flaring was the least ambiguous. There are no similar behaviors with which to confuse flaring, and, as far as we are aware, flares are not used in other behavioral contexts.
Although chemical ablation of antennae reduced the aggression of G. campestris females toward heterospecific males, aggression was not eliminated and females did not begin to accept courtship attempts from heterospecific males. This suggests that either our method of chemical ablation was only moderately successful, or that additional cues are informing species recognition. Zinc sulfate is known to be effective in the chemical ablation of antennae, disabling the electrophysiological response of the chemosensory sensilla that are found along the length of antennae (Balakrishnan and Pollack, 1997). It is possible that some of the chemosensory sensilla closest to the cricket's head, which we avoided immersing in the solution, remained active. Also, there are other areas of the body such as the palps that carry chemosensory sensilla (Klein, 1981), and may therefore detect CHCs. These might be particularly important for the detection of volatile airborne compounds, cues that can be passed between individuals without physical contact. Females are likely using additional cues to assess the males, which may explain why aggression was not entirely eliminated. While we disabled chemoreception of their antennae, mechanoreceptors were unaffected by the zinc sulfate. Females could therefore still detect any tactile differences between the males, such as differences in morphology, or in movements made by the antennae or body. Though the males were silenced to prevent females from using auditory cues, there may be characteristic movements of the wings made by singing males that could be used to differentiate between species, or other visual cues.
The change in behavior attributed to loss of chemoreception suggests that chemical cues are involved in species recognition between G. campestris and G. bimaculatus. While females of many species use characteristics of male advertisement calls to distinguish between conspecifics and heterospecifics, in these crickets calling song represents a relatively weak barrier to hybridization. Cues provided through other sensory modalities received at short range, such as chemoreception, may therefore be important in the maintenance of species boundaries. The use of chemical cues for species recognition may well be important between other species of interbreeding crickets, and we encourage studies explicitly investigating this.
Author Contributions
FT, RR, and TT conceived and designed the study. RR and DF carried out field work. FT and DF carried out laboratory work. FT, Pd'E, RR, and TT analyzed and interpreted data. FT, DF, Pd'E, RR, and TT drafted and revised the manuscript, and gave final approval to the version to be published.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Luke Meadows for help collecting wild crickets, Christopher Mitchell and Melia Burdon for assistance in processing samples, Fiona Ingleby and Megan Head for advice on statistical analyses, and Jelle van Zweden for help with chemical identification of compounds. Anonymous reviewers gave constructive advice for which we are thankful. This work was supported by the European Social Fund and Natural Environment Research Council (grant no.: NE/H02364X/1; studentship no.: NE/H02249X/1). CHC and behavioral data can be accessed at https://ore.exeter.ac.uk/repository/handle/10871/17117.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/article/10.3389/fevo.2015.00048/abstract
References
Ache, B. W., and Young, J. M. (2005). Olfaction: diverse species, conserved principles. Neuron 48, 417–430. doi: 10.1016/j.neuron.2005.10.022
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Akino, T., Yamamura, K., Wakamura, S., and Yamaoka, R. (2004). Direct behavioral evidence for hydrocarbons as nestmate recognition cues in Formica japonica (Hymenoptera: Formicidae). Appl. Entomol. Zool. 39, 381–387. doi: 10.1303/aez.2004.381
Alexander, R. D. (1961). Aggressiveness, territoriality, and sexual behavior in field crickets (Orthoptera: Gryllidae). Behaviour 17, 130–223. doi: 10.1163/156853961X00042
Balakrishnan, R., and Pollack, G. (1997). The role of antennal sensory cues in female responses to courting males in the cricket Teleogryllus oceanicus. J. Exp. Biol. 200, 511–522.
Barbosa, D., Font, E., Desfilis, E., and Carretero, M. (2006). Chemically mediated species recognition in closely related Podarcis wall lizards. J. Chem. Ecol. 32, 1587–1598. doi: 10.1007/s10886-006-9072-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Blomquist, G. C., and Bagnères, A. G. (eds.). (2010). Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology. Cambridge: Cambridge University Press.
Chenoweth, S. F., and Blows, M. W. (2005). Contrasting mutual sexual selection on homologous signal traits in Drosophila serrata. Am. Nat. 165, 281–289. doi: 10.1017/CBO9780511711909
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Christensen, J. H., Hansen, A. B., Karlson, U., Mortensen, U., and Andersen, O. (2005). Multivariate statistical methods for evaluating biodegradation of mineral oil. J. Chromatogr. A 1090, 133–145. doi: 10.1016/j.chroma.2005.07.025
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cousin, G. (1933). Sur l'hybridation de deux espèces de Gryllidae (Acheta campestris et bimaculata). Bull. Soc. Entomol. Fr. 12, 189–193.
Coyne, J. A., and Charlesworth, B. (1997). Genetics of a pheromonal difference affecting sexual isolation between Drosophila mauritiana and D. sechellia. Genetics 145, 1015–1030.
Gibbs, A. G. (1998). Water-proofing properties of cuticular lipids. Am. Zool. 38, 471–482. doi: 10.1093/icb/38.3.471
Gibbs, A. G., Chippindale, A. K., and Rose, M. R. (1997). Physiological mechanisms of evolved desiccation resistance in Drosophila melanogaster. J. Exp. Biol. 200, 1821–1832.
Gorochov, A. V., and Llorente, V. (2001). Estudio taxonómico preliminar de los Grylloidea de España (Insecta, Orthoptera). Graellsia 57, 95–139. doi: 10.3989/graellsia.2001.v57.i2.281
Hardy, T. N., and Shaw, K. C. (1983). The role of chemoreception in sex recognition by male crickets: Acheta domesticus and Teleogryllus oceanicus. Physiol. Entomol. 8, 151–166. doi: 10.1111/j.1365-3032.1983.tb00344.x
Hebets, E. A., and Papaj, D. R. (2005). Complex signal function: developing a framework of testable hypotheses. Behav. Ecol. Sociobiol. 57, 197–214. doi: 10.1007/s00265-004-0865-7
Howard, R. W., and Blomquist, G. J. (2005). Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 50, 371–393. doi: 10.1146/annurev.ento.50.071803.130359
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Huang, Y., Ortí, G., Sutherlin, M., Duhachek, A., and Zera, A. (2000). Phylogenetic relationships of North American field crickets inferred from mitochondrial DNA data. Mol. Phylogenet. Evol. 17, 48–57. doi: 10.1006/mpev.2000.0815
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ivy, T. M., Weddle, C. B., and Sakaluk, S. K. (2005). Females use self-referent cues to avoid mating with previous mates. Proc. Biol. Sci. 272, 2475–2478. doi: 10.1098/rspb.2005.3222
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Klein, U. (1981). Sensilla of the cricket palp. Cell Tissue Res. 219, 229–252. doi: 10.1007/BF00210145
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kühbandner, S., Sperling, S., Mori, K., and Ruther, J. (2012). Deciphering the signature of cuticular lipids with contact sex pheromone function in a parasitic wasp. J. Exp. Biol. 215, 2471–2478. doi: 10.1242/jeb.071217
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Leonard, A. S., and Hedrick, A. V. (2009). Single versus multiple cues in mate discrimination by males and females. Anim. Biol. 77, 151–159. doi: 10.1016/j.anbehav.2008.09.029
Liang, D., and Silverman, J. (2000). “You are what you eat”: diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepithema humile. Naturwissenschaften 87, 412–416. doi: 10.1007/s001140050752
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maroja, L. S., McKenzie, Z. M., Hart, E., Jing, J., Larson, E. L., and Richardson, D. P. (2014). Barriers to gene exchange in hybridizing field crickets: the role of male courtship effort and cuticular hydrocarbons. BMC Evol. Biol. 14:65. doi: 10.1186/1471-2148-14-65
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mayr, E. (1942). Systematics and the Origin of Species, from the Viewpoint of a Zoologist. Cambridge, MA: Harvard University Press.
McLennan, D. A., and Ryan, M. J. (1999). Interspecific recognition and discrimination based upon olfactory cues in northern swordtails. Evolution 53, 880–888. doi: 10.2307/2640728
Mullen, S. P., Mendelson, T. C., Schal, C., and Shaw, K. L. (2007). Rapid evolution of cuticular hydrocarbons in a species radiation of acoustically diverse Hawaiian crickets (Gryllidae: Trigonidiinae: Laupala). Evolution 61, 223–231. doi: 10.1111/j.1558-5646.2007.00019.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mullen, S. P., Millar, J., Schal, C., and Shaw, K. L. (2008). Identification and characterization of cuticular hydrocarbons from a rapid species radiation of Hawaiian swordtailed crickets (Gryllidae: Trigonidiinae: Laupala). J. Chem. Ecol. 34, 198–204. doi: 10.1007/s10886-007-9419-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nagamoto, J., Aonuma, H., and Hisada, M. (2005). Discrimination of conspecific individuals via cuticular pheromones by males of the cricket Gryllus bimaculatus. Zool. Sci. 22, 1079–1088. doi: 10.2108/zsj.22.1079
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Otte, D., and Cade, W. (1976). On the role of olfaction in sexual and interspecies recognition in crickets (Acheta and Gryllus). Anim. Behav. 24, 1–6. doi: 10.1016/S0003-3472(76)80091-7
Panhuis, T. M., Butlin, R., Zuk, M., and Tregenza, T. (2001). Sexual selection and speciation. Trends. Ecol. Evol. 16, 364–371. doi: 10.1016/S0169-5347(01)02160-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pardo, J. E., Gomes, R., and Del Cerro, A. (1993). Orthopteroidea from the mountain chains of Castilla-La Mancha (Spain). II. Ensifera. Zool. Baetica 4, 113–148.
Paul, R. (1976). Acoustic response to chemical stimuli in ground crickets. Nature 263, 404–405. doi: 10.1038/263404a0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Popov, A. V., and Shuvalov, V. F. (1977). Phonotactic behavior of crickets. J. Comp. Physiol. A 119, 111–126.
R Core Team. (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rence, B., and Loher, W. (1977). Contact chemoreceptive sex recognition in the male cricket, Teleogryllus commodus. Physiol. Entomol. 2, 225–236. doi: 10.1111/j.1365-3032.1977.tb00108.x
Rundle, H. D., Chenoweth, S. F., Doughty, P., and Blows, M. W. (2005). Divergent selection and the evolution of signal traits and mating preferences. PLoS Biol. 3:e368. doi: 10.1371/journal.pbio.0030368
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ryan, K. M., and Sakaluk, S. K. (2009). Dulling the senses: the role of the antennae in mate recognition, copulation and mate guarding in decorated crickets. Anim. Behav. 77, 1345–1350. doi: 10.1016/j.anbehav.2009.02.011
Simmons, L. W. (1989). Kin recognition and its influence on mating preferences of the field cricket, Gryllus bimaculatus (de Geer). Anim. Behav. 38, 68–77. doi: 10.1016/S0003-3472(89)80066-1
Simmons, L. W. (1990). Pheromonal cues for the recognition of kin by female field crickets, Gryllus bimaculatus. Anim. Behav. 40, 192–195. doi: 10.1016/S0003-3472(05)80683-9
Simmons, L. W., Alcock, J., and Reeder, A. (2003). The role of cuticular hydrocarbons in male attraction and repulsion by female Dawson's burrowing bee, Amegilla dawsoni. Anim. Behav. 66, 677–685. doi: 10.1006/anbe.2003.2240
Smadja, C., and Butlin, R. K. (2008). On the scent of speciation: the chemosensory system and its role in premating isolation. Heredity 102, 77–97. doi: 10.1038/hdy.2008.55
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Smadja, C., Catalan, J., and Ganem, G. (2004). Strong premating divergence in a unimodal hybrid zone between two subspecies of the house mouse. J. Evolution. Biol. 17, 165–176. doi: 10.1046/j.1420-9101.2003.00647.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tabachnick, B. G., and Fidell, L. S. (1989). Using Multivariate Statistics. Philadelphia: Harper and Row.
Thomas, M. L., and Simmons, L. W. (2008). Sexual dimorphism in cuticular hydrocarbons of the Australian field cricket Teleogryllus oceanicus (Orthoptera: Gryllidae). J. Insect Physiol. 54, 1081–1089. doi: 10.1016/j.jinsphys.2008.04.012
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thomas, M. L., and Simmons, L. W. (2009). Male derived cuticular hydrocarbons signal sperm competition intensity and affect ejaculate expenditure in crickets. Proc. Biol. Sci. 276, 383–388. doi: 10.1098/rspb.2008.1206
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thomas, M. L., and Simmons, L. W. (2011). Crickets detect the genetic similarity of mating partners via cuticular hydrocarbons. J. Evolution. Biol. 24, 1793–1800. doi: 10.1111/j.1420-9101.2011.02319.x
Tregenza, T., and Wedell, N. (1997). Definitive evidence for cuticular pheromones in a cricket. Anim. Behav. 54, 979–984. doi: 10.1006/anbe.1997.0500
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tyler, F., Harrison, X. A., Bretman, A., Veen, T., Rodríguez− Muñoz, R., and Tregenza, T. (2013a). Multiple post−mating barriers to hybridization in field crickets. Mol. Ecol. 22, 1640–1649. doi: 10.1111/mec.12187
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tyler, F., Rodríguez-Muñoz, R., and Tregenza, T. (2013b). Fertilisation and early developmental barriers to hybridisation in field crickets. BMC Evol. Biol. 13:43. doi: 10.1186/1471-2148-13-43
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van Zweden, J., Dreier, S., and d'Ettorre, P. (2009). Disentangling environmental and heritable nestmate recognition cues in a carpenter ant. J. Insect Physiol. 55, 158–163. doi: 10.1016/j.jinsphys.2008.11.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Veen, T., Faulks, J., Rodríguez-Muñoz, R., and Tregenza, T. (2011). Premating reproductive barriers between hybridising cricket species differing in their degree of polyandry. PLoS ONE 6:e19531. doi: 10.1371/journal.pone.0019531
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Veen, T., Faulks, J., Tyler, F., Lloyd, J., and Tregenza, T. (2013). Diverse reproductive barriers in hybridizing crickets suggests extensive variation in the evolution and maintenance of isolation. Evol. Ecol. 27, 993–1015. doi: 10.1007/s10682-012-9610-2
von Hörmann-Heck, S. (1957). Untersuchungen über den Erbgang eininger Verhaltensweisen bei Grillenbastarden. Z. Tierpsychol. 14, 137–183. doi: 10.1111/j.1439-0310.1957.tb00529.x
Warthen, J. D., and Uebel, E. C. (1980). Comparison of the unsaturated cuticular hydrocarbons of male and female house crickets, Acheta domesticus (L.) (Orthoptera: Gryllidae). Insect Biochem. 10, 435–439. doi: 10.1016/0020-1790(80)90015-3
Weiss, I., Hofferberth, J., Ruther, J., and Stökl, J. (2015). Varying importance of cuticular hydrocarbons and iridoids in the species-specific mate recognition pheromones of three closely related Leptopilina species. Front. Ecol. Evol. 3:19. doi: 10.3389/fevo.2015.00019
Wilson, A. D., Whattam, E. M., Bennett, R., Visanuvimol, L., Lauzon, C., and Bertram, S. M. (2010). Behavioral correlations across activity, mating, exploration, aggression, and antipredator contexts in the European house cricket, Acheta domesticus. Behav. Ecol. Sociobiol. 64, 703–715. doi: 10.1007/s00265-009-0888-1
Zhang, B., Xue, H. J., Song, K. Q., Liu, J., Li, W. Z., Nie, R. E., et al. (2014). Male mate recognition via cuticular hydrocarbons facilitates sexual isolation between sympatric leaf beetle sister species. J. Insect Physiol. 70, 15–21. doi: 10.1016/j.jinsphys.2014.08.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: Gryllus bimaculatus, Gryllus campestris, speciation, reproductive isolation, chemoreception, hybridization, aggression
Citation: Tyler F, Fisher D, d'Ettorre P, Rodríguez-Muñoz R and Tregenza T (2015) Chemical cues mediate species recognition in field crickets. Front. Ecol. Evol. 3:48. doi: 10.3389/fevo.2015.00048
Received: 22 February 2015; Accepted: 21 April 2015;
Published: 08 May 2015.
Edited by:
Sebastien Lebreton, Swedish University of Agricultural Sciences, SwedenReviewed by:
Joachim Ruther, University of Regensburg, GermanyAnne-Geneviève Bagnères, Centre National de la Recherche Scientifique, France
Copyright © 2015 Tyler, Fisher, d'Ettorre, Rodríguez-Muñoz and Tregenza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frances Tyler, Department of Biological Sciences, University of Cincinnati, Rieveschl Hall, Cincinnati, OH 45221-0006, USA, frances.a.tyler@gmail.com