Proximity to Woodland and Landscape Structure Drives Pollinator Visitation in Apple Orchard Ecosystem
 Neelendra K. Joshi1,2,3*‡
Neelendra K. Joshi1,2,3*‡  Mark Otieno1,4*†‡
Mark Otieno1,4*†‡  Edwin G. Rajotte1
Edwin G. Rajotte1  Shelby J. Fleischer1 David J. Biddinger1,3
Shelby J. Fleischer1 David J. Biddinger1,3- 1Department of Entomology, Pennsylvania State University, University Park, PA, USA
- 2Entomology, Fruit Research and Extension Center, Pennsylvania State University, Biglerville, PA, USA
- 3Department of Entomology, University of Arkansas, Fayetteville, AR, USA
- 4Department of Agricultural Resource Management, Embu University College, Embu, Kenya
Landscapes of farms and adjacent areas are known to influence abundance of various arthropods such as pollinators in commercial agricultural ecosystems. In this context, we examined the effect of heterogeneous landscapes surrounding and including commercial apple orchards on pollinator visitation and foraging distance during bloom period from 2011 to 2013 in Pennsylvania. Our results showed that the frequency of feral honeybees and solitary bee visits within an apple orchard depends on the proximity of the orchard to an unmanaged habitat (primarily comprised of forest). At the landscape scale, we found that the Mean Proximity Index, the Largest Patch Index, and the Number of Patches positively correlated with the visitation rate of dominant bee taxa (Apis mellifera, Bombus spp., and solitary bees) visiting apple flowers at low spatial scales (up to 500 m around the orchards). The Mean Proximity Index at 500 m was related to bee visitation patterns, especially for solitary bees and A. mellifera. Bees in all our study sites preferred to forage in areas with large homogenous patches up to 500 m around an apple orchard. This effect can be attributed to the mass flowering of apples that formed the largest proportion of the 500 m spatial scale. The Number of Patches at 250 m spatial scale was positively correlated with bee visitation, especially Bombus spp., probably because these areas had more habitats and more resources required by these bees. We conclude that retaining unmanaged habitats closer to commercial apple orchards will maintain biodiversity within the landscapes and insure pollination services to apples.
Introduction
Agricultural crop production relies heavily on insects to provide pollination services (Zhang et al., 2007; Potts et al., 2010). Among several species of insects, managed bees, mainly honey bees (Apis mellifera), and bumble bees (Bombus spp.), commonly make up for shortages in wild pollinators and feral honey bees that provide pollination services in various crops (Potts et al., 2010). Wild bees are crucial in pollination of several cultivated and wild flowering plants, and recent research has shown that they are efficient crop pollinators (Tepedino et al., 2007; Aebi et al., 2012; Christmann and Aw-Hassan, 2012; Garibaldi et al., 2013).
During the last few years, population decline in honey bees and other wild bees like bumble bees have been reported in North America and Europe (Grixti et al., 2009; Cameron et al., 2011), although similar trends have not been documented in other parts of the world such as Australia and Africa. Among several interacting stressors that have been reported in the recent past (Smith et al., 2013), agricultural intensification is thought to be one of the major causes of pollinator decline (Inouye, 2007; Marini et al., 2012) mainly through the destruction of nesting and foraging habitats, and frequent use of pesticides. Preserving natural and semi-natural habitats surrounding farmlands under commercial crop production may provide nesting and forage resources and buffer pollinators from adverse conditions. Such natural resource management strategy may increase the amounts of ecosystem services provided by pollinators, resulting in increased crop yields, and quality of produce (Holzschuh et al., 2012).
Apple (Malus domestica L., Rosaceae) is a major pollinator-dependent and high-value fruit crop which relies heavily on insect pollinators to set fruit (Tepedino et al., 2007; Garratt et al., 2014; Sheffield, 2014). In 2007, apples were grown on more than 145,765 hectares in the U.S. that produced 4.73 million tons ($2.22 billion US) (see USDA1 economic research service 2010). Like many other roseaous tree fruits (for instance, cherries, plums, pears etc.), bloom in apple occurs in the early spring when most solitary bee species are inactive and some social species such as bumble bees have not had time to build colony membership. During this period apples produce a large number of blossoms, a phenomenon referred to as mass flowering (Westphal et al., 2003) which increases the demand for insects, mainly bees, to pollinate them.
Orchard management in commercial apple production is quite intense and may create unsuitable conditions for nesting bees. Vegetation between tree rows in the apple orchard is regularly mowed and herbicides are applied beneath trees to reduce competition for water and nutrients (Merwin, 2003), which may also affect plant species competition for floral visitors such as bees in the orchard landscape. The orchards are also sprayed pre-bloom with some common organophosphate and neonicotinoid insecticides and during bloom with many types of fungicides to control various insect pests and diseases (Hull et al., 2009; Penn State Extension, 2014). However, semi-natural habitats that surround these intensively managed commercial orchards offer refuge that may protect bees from on-farm disturbances while providing floral resources and nesting habitats (Wilson and Thomson, 1991; Javorek et al., 2002; Williams and Thomson, 2003; Winfree et al., 2007). Several studies have recently examined the effect of habitat around apple orchards or other crops in relation to wild bee visitation and the potential of A. mellifera to provide comparable pollination services in the absence of wild bees (e.g., Ricketts et al., 2008; Garibaldi et al., 2011, 2013; Kennedy et al., 2013). However, these studies have rarely considered apples grown in heterogeneous landscapes like those found in the Mid-Atlantic region of the eastern Untited States. Such differences in landscape, weather conditions, and bee community composition makes it difficult to draw relevant conclusions from other systems and regions, and highlight the need for regional studies that are relevant and realistic for pollinator management. Additionally, these regional studies contribute to a comprehensive understanding of pollinator declines.
Proximity of crop fields to semi-natural vegetation is important in enhancing pollinator diversity and the level of ecosystem services provided by pollinators to crops (Karanja et al., 2010; Blitzer et al., 2012). However, effects of proximity to semi natural vegetation may vary with the landscape context (Steffan-Dewenter et al., 2002; Ricketts et al., 2008; Jha and Kremen, 2013). Other studies measure proximity to unmanaged land, (e.g., Bailey et al., 2014), but few consider the shape and size of those unmanaged land parcels or landscape contexts. Some studies suggest that the shape and size of unmanagaed landscape parcels matter in determining the diversity and foraging activity of pollinators (Steffan-Dewenter et al., 2002; Otieno et al., 2011; Marini et al., 2012). The Mid-Atlantic region, specifically Pennsylvania provides an opportunity to evaluate the effects of proximity to natural or semi-natural habitat in heterogeneous landscapes. In this context, here we examine the effects of diverse landscapes surrounding commercial apple orchards on pollinator visitation over a 3 years period. We explore the following three questions: (1) Does proximity to natural woodland affect pollinator foraging distances into an orchard? (2) How does the surrounding landscape context affect pollinator visitation on apple flowers? (3) How do different pollinator species differ in their responses to landscape structure in and around apple orchards?
Materials and Methods
This study was conducted in Adams County, Pennsylvania during apple bloom periods of April–May 2011, 2012, and 2013. Apples in this county are grown in orchards covering ca. 8903 hectares, and it is the leading tree fruit producing area in the state, making Pennsylvania the fourth leading apple producing state in the USA.
Description of Study Orchards
The orchards in the region are generally established in south facing slopes on well drained soils and surrounded by a heterogeneous landscape comprising forests (>50%), shrubs, field crops and developed land, which is a typical geography of the eastern Appalachian region (Egan and Mortensen, 2012).
For this study, we selected five commercial orchards with similar management programs (such as pesticide, herbicide, and fruit thinning practices) and which did not stock managed honey bees, bumble bees, or solitary bees for pollination. The orchards were mature at 15–20 years, on semi-dwarf rootstocks, with an average spacing of ~4.5 m between trees by ~6 m between rows (average of about 600–700 trees/ha). Within the Golden Delicious, York, and Honey Crisp varieties of each orchard, apple trees (n = 120/orchard, i.e., 600 trees from five orchards) were individually selected by distance from the edge closest to woodland toward the center of the orchard, marked and referenced with a GPS unit for sampling and bee observations. The distance gradients were same in each study orchard, and were 15, 35, 55, 100, and 200 m from the edge of the adjacent wooded area into the orchard.
Pollinator Visitation
Observations were made during optimal weather conditions [i.e., low wind speed (>3.5 m/s), temperature above 16°C, partly cloudy or bright overcast for bee flight (Supplementary Material S1). For each of the two sampling dates per orchard, all flowers on each tree were watched by two trained observers for 1 min between 1000 and 1400 h, during good weather conditions. Each observer stood on either side of the tree to capture all visitations to that side of the tree. Three main pollinator taxa were recorded—A. mellifera, Bombus spp., and solitary bees. In addition, syrphid flies were also counted, but were low in numbers across all study years. Immediately after the pollinator visit observations, we net-collected vouchers of all non-Apis bees for 30 min each at 15, 35, 55, 100, and 200 m from the edge of the adjacent wooded area into the orchard. These bee samples were placed in vials, labeled, and chilled in the field and frozen in the lab until they could be pinned, labeled, and identified. All bees were identified to species (see Supplementary Material S2).
Landscape Analysis
The surrounding landscape was assessed with the National Agricultural Statistical Service (NASS) map layer 2012 and groundtruthed in October 2013. The NASS land-use maps were 30 m raster dateset. The maps were first uploaded in ArcGIS 10.1 and buffer layers were created at increasing spatial scales. These buffers started from the edges of the wooded areas where they were adjacent to the orchards and extended out to 250, 500, and 1000 m to reflect the flight ranges of bees from the wooded adjacent areas that are generally nesting habitats of wild bees (Greenleaf et al., 2007; Zurbuchen et al., 2010). These buffers were then uploaded in Fragstats 4.0 and the metrics were generated at the landscape level for each scale (McGarigal et al., 2012; McGarigal, 2014). We assessed collinearity by using a correlation matrix to eliminate collinear metrics. The following landscape metrics were retained for further analyses: (i) number of patches; (ii) edge density; (iii) largest patch index; (iv) mean proximity index (v) landscape shape index; and (vi) contagion. A description of these metrics is available from http://www.umass.edu/landeco/teaching/landscape_ecology/schedule/chapter9_metrics.pdf (accessed November 2013) and in Supplementary Material S3.
Data Analyses
Data from 120 individual trees (the sum from the two observers on a given sampling date) were averaged across the sampling dates within each year. We used simple linear regression models to determine the relationship between the number of bee visits and distance from the woodland into the apple orchards.
To test how landscape composition affects the apple pollinator community, we used redundancy analyses to assess the strengths of correlations between various landscape metrics and bee taxa (CANOCO 5.0,ter Braak and Šmilauer, 2012); we calculated the proportion of variance explained by an ordination of the metrics in the selected groups against all the response bee taxa selected (Pakeman, 2011), and then a forward selection technique to obtain a subset of landscape variables to model multivariate bee community composition (Legendre and Legendre, 1998).
For each year, the mean pollinator visitation per 2 min per tree per year were fitted to landscape factors using generalized linear mixed effects models (glimmer) in lme4 library (Bates et al., 2011) using R (R Development Core Team, 2013). We used a log-link function to account for Poisson distribution in our data. We fitted the model with “site” as a random term to account for the variance in the response variables. The entire model was fitted with all the landscape variables as fixed factors first, then the model was progressively simplified by deleting fixed factors that caused a significant reduction in the Akaike Information Criterion (AIC), a measure of the quality of a statistical model useful in model selection, until a minimum adequate model was attained (Otieno et al., 2011). The minimum adequate model was taken to be a model from which the removal of any of the fixed factors caused an increase in the AIC (Crawley, 2007). To assess the significance of each predictor in the minimum adequate model, we used a log-likelihood ratio test (L-Ratio test). We used the same modeling structure to test for each pollinator taxa (Bombus spp., A. mellifera, and solitary bees) as a response variable. The L-Ratio test was done by applying a two-tier process: first we fitted the minimum adequate model with all predictors, then fitted a second model excluding the one predictor variable at a time. Each reduced model was compared to the minimum adequate model to determine the significance based on its Chi-square statistic and p-value. The response variables tested were mean visits per apple tree by A. mellifera, Bombus spp., and solitary bee taxa. We did not include syrphid flies due to very low frequency of observation.
Results
A total of 2083 bees were observed over the 3 years of sampling (872, 628, and 584 bees in 2011, 2012, and 2013, respectively). We recorded 32 species: 26 species of solitary bees, 5 species of Bombus, and A. mellifera (the full list of bee species is appended as Supplementary Material S2). Wild honey bees (A. mellifera) were the most abundant visitors, accounting for 60% of the total visits recorded across the 3 years of sampling. Solitary bees accounted for 33% of the total visits, while Bombus spp. made only 7% of total visits.
Proximity to Natural Woodland and Foraging Distance
Most foraging activity was recorded closer to the natural woodland; lowest activity was recorded farther into the orchard. The proximity of an apple tree to woodland significantly affected distances that bees foraged into apple flowers in 2011 (R2 = 0.601, P < 0.001), and 2012 (R2 = 0.186, P = 0.008), but the effect was not significant in 2013 (R2 = 0.0164, P > 0.05) (Figure 1).
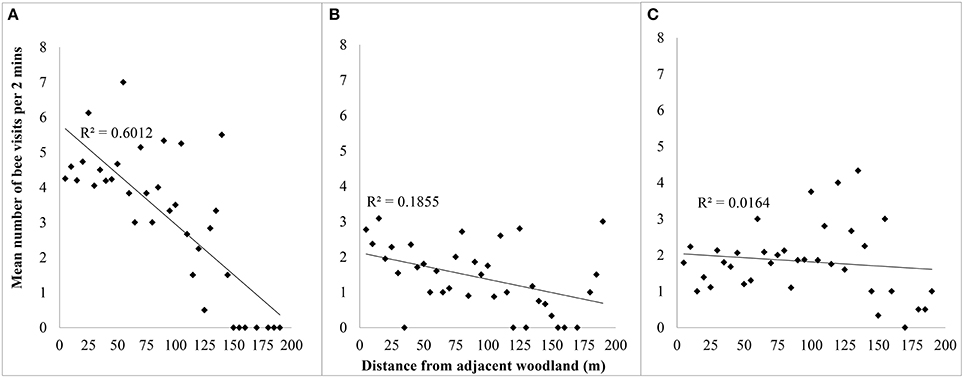
Figure 1. Relationship between number of bee visits to apple flowers per unit time against distance (in meters) from woodland adjacent to orchard for (A) 2011, (B) 2012, and (C) 2013.
Honey bees and solitary bees were responsible for the significant decline in foraging activity into the orchard from the woodland edge during the mass flowering period of apples in both 2011 and 2012 (Table 1, Figure 2). Bombus spp. contributed to the overall decline in foraging activity into the orchard in 2011, and were responsible for a slight and significant decline observed in 2013 (P < 0.027, Table 1). During 2013, Apis and solitary bee foraging activity did not significantly change with increasing distance from the orchard edge.
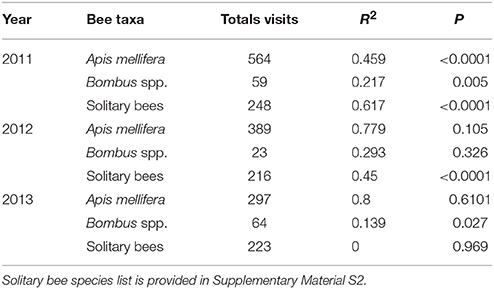
Table 1. Summary of regression outputs of the relationship between bee taxa foraging on apple flowers (mean visits per 2 min count per tree per year) and distance (meters) from natural woodland.
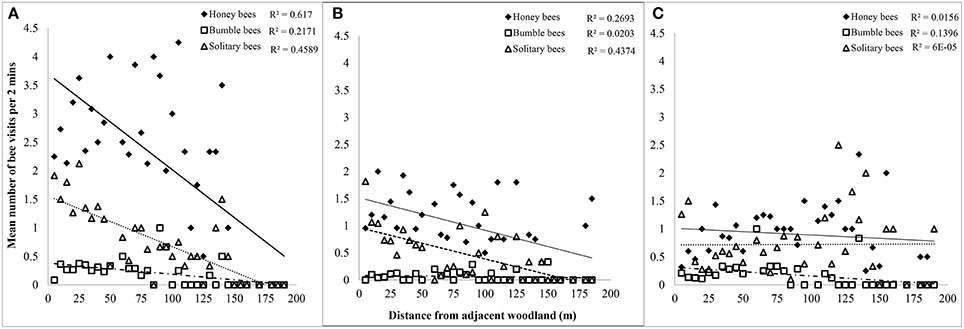
Figure 2. Relationship between number of bee visits by taxa to apple flowers per unit time vs. distance (in meters) from woodland adjacent to orchard for (A) 2011, (B) 2012, and (C) 2013.
Impact of Landscape Context on Bee Visitation
Overall, the landscape at low spatial scales (250–500 m) significantly affected numbers of bee visits to apple flowers. The Mean Proximity Index at 500 m had the highest effect on numbers of bee visits, explaining 48.5% of the total variation in the data (Figure 3).
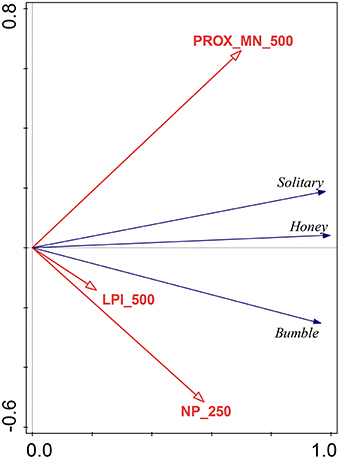
Figure 3. Results of RDA with pollinator taxa and landscape factors forward selected. The variables shown in the figure significantly account for 99.5% of adjusted explained variation. Mean Proximity Index at 500 m accounted for 48.5% of the variation. Number of Patches at 250 m explains 42.6% and Largest Patch Index at 500 m explains 8.8% of the variation.
The Number of Patches at 250 m and Largest Patch Index at 500 m explained 42.6 and 8.8% of the total explained variation, respectively. No other landscape factor significantly affected the number of pollinator visits. Solitary bees had a strong relationship with the Mean Proximity Index while Bombus spp. responded more to Largest Patch Index at 500 m and Number of Patches at 250 m in the RDA ordination. A. mellifera were largely unaffected, but showed a weak relationship with the Mean Proximity Index at 500 m (Figure 3).
Using a univariate linear mixed effects model approach to determine how the above landscape factors affected each bee taxa independent of the other taxas, we found all the bee taxa to be significantly affected by Number of Patches at 250 m, Largest Patch Index at 500 m, and Mean Proximity Index at 500 m (Table 2 and Figure 4).
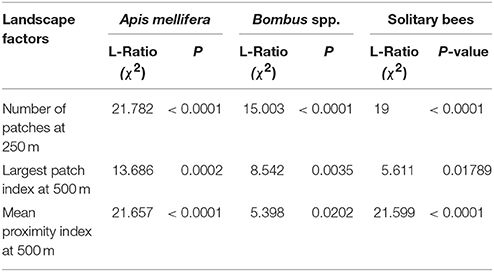
Table 2. Linear mixed effects models showing L-Ratio (χ2) outputs of the effects of landscape heterogeneity at varying scales on pollinator taxa visiting apple orchards.
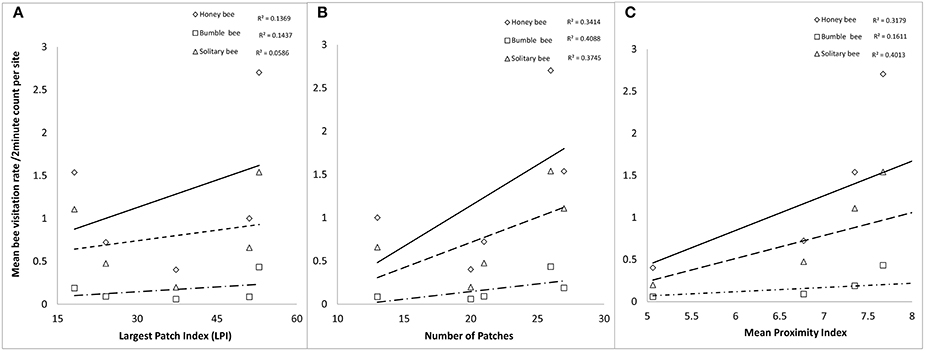
Figure 4. Relationship between mean number of bee visits by taxa to apple flowers per unit time against (A) Largest Patch Index, (B) Number of Patches, and (C) Mean Proximity Index.
Discussion
Our study showed that the distance of apple orchards from surrounding heterogeneous landscape has significant effects on the foraging activity of bees that provide an important ecosystem service during bloom period. We also showed that landscape variables were more significant at much smaller spatial scales than previously reported (Kremen et al., 2004). Feral A. mellifera and solitary bee foragers were significantly more abundant closer to the natural woodland than they were in the center of the orchard. Bees prefer to forage in the most rewarding patches (in terms of quality and distance) in the landscape (Olsson et al., 2015), and the higher abundance of bees near natural woodland could be due to this reason. Similar to a previous study on bee community composition in apple orchards in this region (Joshi et al., 2015), we also observed high diversity of wild bees visiting apple flowers during bloom period. These bee species, especially solitary bees, commonly live within natural, or semi-natural vegetation. Cavity-nesting bees have been shown to respond negatively to intense agriculture, presumably in response to loss of nesting habitat availability (Sheffield et al., 2013). The area bees forage is dependent on their nesting site as central-placed foragers. The distance they fly depends on their size and flight capability (Greenleaf et al., 2007; Zurbuchen et al., 2010), as well as floral resource availability (Biddinger et al., 2013). Presumably, if adequate food is available nearby, foragers will not fly beyond that distance.
Proximity to Natural Woodland and Foraging Distance
Generally, solitary bees can fly a few hundred meters, depending on availability of floral resources near their nests (Greenleaf et al., 2007). A. mellifera can forage much further (1100 m) from their nest (Gary et al., 1981) while Bombus spp. forage further away than most species (Greenleaf et al., 2007). Most bee species generally fly further from their nest if food (as floral resources) is in short supply, but fly a short distance to reach a plentiful supply of flowers (such as a mass-flowering apple crop) (Rao and Strange, 2012; Jha and Kremen, 2013; Sheffield, 2014). Pollinators may use natural and semi-natural habitats within agricultural landscapes as refugia. These refugia are important for bees and other insects because they provide alternative habitats for foraging, mating, and nesting (Westrich, 1996). As such, habitats such as woodlands adjacent to apple orchards may be an important source habitat of pollinators that forage on an apple crop (Watson et al., 2011).
In ideal environmental conditions during apple bloom, pollinators constantly move between the natural habitats and crop fields (Garibaldi et al., 2011). Insects that migrate into crop fields to forage for pollen and nectar usually effect pollination in the process (Blitzer et al., 2012). Wild bee species that use semi-natural vegetation will move to crop fields when more resources become available within the crop (such as the mass-flowering apples) than in the natural vegetation (Blitzer et al., 2012). This phenomenon is illustrated in temperate and tropical agroecosystems. In the latter, wild bees from neighboring natural forest fragments increase yields and quality of coffee crops (Klein et al., 2003; Karanja et al., 2010).
Yearly variation in weather and bloom time affected overall bee foraging during our study. In 2011 we observed the highest number of bees foraging on apples. Sampling was done in bright, sunny and warm days and rain did not fall during this period. However, in 2012, spring occurred very early; flowers bloomed early and bees emerged early. Just before apple bloom, temperatures plummeted, and frost occurred (Supplementary Material S1). The bloom period was cold and rainy. Similarly in 2013 frost did not occur during bloom, but was dominated by overcast weather (Supplementary Material S1) and rain. These are not favorable conditions for bee flights (especially smaller solitary bees). Bees require optimal environmental conditions (temperature, humidity, wind) to effectively forage. At the time of apple bloom the only bumble bees we observed were queens in relatively low numbers, so we did not detect a significant association with semi-natural vegetation.
Impact of Landscape Context on Bee Visitation
Because bees have limited flight ranges from their nesting locations (Vicens and Bosch, 2000; Kremen et al., 2004; Biddinger et al., 2013), the landscape surrounding a farmland and the resources that the landscape provides, are important for our understanding of bee foraging patterns, bee conservation, and ecosystem services provided by bees. Examination of the landscapes that surround a particular crop has interested landscape ecologists and conservation biologists in order to understand the patterns of bee visitation and pollination services (Kremen et al., 2004; Greenleaf and Kremen, 2006; Winfree et al., 2008; Klein et al., 2012; Cariveau et al., 2013). In this study, we found that landscape composition and configuration positively affected the dominant bee taxa (A. mellifera, Bombus spp., and solitary bees) visiting apple flowers at low spatial scales (up to 500 m around the orchards). Mean Proximity Index at 500 m, the Number of Patches at 250 m, and the Largest Patch Index at 500 m were the three main landscape metrics that correlated positively with bee visitation.
The Mean Proximity Index measures the connectivity of patches within the landscape; it takes into account patch size and distance to neighboring patches (Schweiger et al., 2010). In our study, landscape connectivity at 500 m was positively correlated with bee visitation patterns, especially for solitary bees and A. mellifera, but Bombus spp. visitation to flowers was very low (constituted only 7% of the total 2083 visits by all bees). Habitats that are highly interconnected, i.e., that have a higher Mean Proximity Index, have a better chance to retain high populations of bees and therefore increase pollination activity compared with habitats that are disconnected from one another.
The Largest Patch Index measures the percentage of the total area made up by the largest homogenous patch. Bees in all our study sites preferred to forage in areas with a large homogenous patch up to 500 m around a central point within an apple orchard. This effect can be attributed to the mass flowering of apples that formed the largest proportion of the 500 m spatial scale. In apple production system, large orchards (more than 10 ha per grower, which is >100,000 m2) could easily span beyond 500 m from an orchard's center. Apple bloom period is temporally partitioned so that the mass flowering occurs when few other wild plants or crops are flowering. Furthermore, in our landscape it is not uncommon for mass flowering to have occurred from other rosaceous orchard crops (plum, peach, apricot) in close proximity to apple. However, they do not have much overlap in time. Thus, bees have the opportunity to move temporally among a series of mass flowering tree fruits.
The Number of Patches per unit area at 250 m was positively correlated with bee visitation. More bees, especially Bombus spp., responded to landscapes with more habitat patches, probably because these areas had more nutritional or nesting resources required by these bees. Bombus spp., Apis and some solitary bees are generalist foragers and may require additional resources beyond apple pollen and nectar to supplement their nutritional needs. In areas with multiple habitats, but within the vicinity of a large focal patch (i.e., the mass flowering apple crop), pollinators are moving across crop/non-crop interfaces. The largest diversity and abundance of pollinators is highest at the interface between crop fields and natural vegetation (the ecotone) because these habitats offer more floral and/or nesting resources compared to natural vegetation (e.g., forests, Hagen and Kraemer, 2010). Agricultural and other managed landscapes surely affect the pollinator-plant interactions of adjacent natural habitats.
Results of this study reveal that the effects of landscape configuration on bee visitation were more apparent at smaller spatial scales in apple production system. In general, the landscape in Pennsylvania and the other Mid-Atlantic States is quite heterogeneous. The Mid-Atlantic region is unique in that the landscape therein is a mosaic of different habitats and various land cover types are closer to each other, potentially providing season-long diverse floral resources required by pollinators, which explains the lack of significant results of landscape factors of bee visitation beyond 500 m of an orchard. This landscape structure supports the results of Winfree et al. (2007) with watermelon in the Mid-Atlantic region, who reported no effect of proximity to semi-natural habitats at a high spatial scale (>1000 m), and attributed it to the high heterogeneity of the Pennsylvania and New Jersey landscape. Similar patterns were reported by Lonsdorf et al. (2009) who used a predictive model to map pollination services based on pollinators nesting and floral resource availability and foraging ranges. Their model best fit areas dominated by highly intensive agriculture with low natural or semi-natural cover left (i.e., California and Costa Rica), but not our region (Pennsylvania and New Jersey). They attributed their results for this region to the small-scale heterogeneity defined by high plant diversity and interconnection between similar land cover types offering more nesting and floral resources than in highly intensified landscapes (Tscharntke et al., 2005). Bee foraging behavior in complex heterogeneous landscape (such as Pennsylvania apple orchards) may be more accurately predicted by the refined version of Lonsdorf model, which consider behavioral component into the model (Olsson et al., 2015). At larger spatial scales (e.g., more than 500 m), within heterogeneous landscapes, resources become more abundant and have marginal effects on pollinator visitation because the bees are already meeting their needs without having to forage far from their nests.
In regions with less heterogeneous landscapes such as California, for example, the abundance and diversity of bees strongly influenced by spatial characteristics of the landscape (Collinge, 2010). Farms that are surrounded by a low proportion natural habitat cover (<1% within 1 km of the farm) has a low diversity and abundance of bees and low pollination of crops as a consequence (Collinge, 2010). Bees in such landscapes must fly further from their nests to meet their nutritional requirements. In such scenarios, they may be influenced by landscape context at much higher spatial scales than bees in heterogeneous settings such as Pennsylvania.
To conclude, our study revealed that the magnitude of feral A. mellifera and solitary bee foraging within an apple orchard depends on the proximity of the orchard to an unmanaged habitat such as natural woodland. For purposes of conservation, retention of unmanaged habitats near agricultural areas seems useful in maintaining biodiversity within the landscapes and therefore may help insure pollination. Furthermore, conserving unmanaged habitats in such landscape would also provide additional resources required by pollinators and other beneficial insects such as syrphids and other predatory flies, predatory beetles, spiders, and parasitoids, which in turn help to maintain a healthier community in and near the orchard. Economically, conserving natural areas near the apple orchard system and maintaining standard management practices that are safe to pollinators will ensure sustainable pollination and steady economic returns due to crops receiving sufficient pollination. Our results from landscape structure studies further support and complement the findings of the relationship between bee foraging activity during apple bloom period and the proximity of orchards to unmanaged habitats. We found landscape configuration at smaller spatial scales to be a significant positive determinant of bee visitation. Measures aimed at restoring natural pollinator visitation in and around apple orchards would be more successful if suitable habitats were provided at a smaller spatial scale (i.e., farm level) in heterogeneous landscapes such as Pennsylvania.
Author Contributions
NJ, DB, ER, and MO conceived the study and designed the experiments. NJ, DB, and ER conducted field research and observations. MO collected landscape data and performed statistical analysis. SF contributed in data analysis and manuscript development strategy. MO, NJ, SF, DB, and ER wrote the manuscript. All authors reviewed the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge funding provided by the USDA-NIFA Specialty Crop Research Initiative, from project PEN04398: Determining the Role of and Limiting Factors Facing Native Pollinators in Assuring Quality Apple Production in Pennsylvania; a Model for the Mid-Atlantic Tree Fruit Industry, and from project 2012-51181-20105: Developing Sustainable Pollination Strategies for U.S. Specialty Crops, and the State Horticultural Association of Pennsylvania for conducting this research. Authors are thankful to K. Wholaver, K. Ellis, L. Miller, A. Ritz, J. Fissel for assisting in the fieldwork, E. Winzler for assistance in GIS mapping of field sites, R. Donovall, Dr. J. Gibbs and Dr. R. Jean for helping with bee identification, and fruit growers: D. Slaybaugh, E. Diveley Jr., S. Slaybaugh and B. Knouse for allowing us to use their orchards to conduct this study. Authors are also thankful to K. Watrous for editing and C. S. Sidhu for reviewing an earlier draft of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/article/10.3389/fevo.2016.00038
Footnotes
1. ^USDA (2012). Economic Research Service: Apple statistics. Available online at: http://usda.mannlib.cornell.edu/MannUsda/viewDocumentInfo.do?documentID=1002 (Accessed April 2014).
References
Aebi, A., Vaissière, B. E., Delaplane, K. S., Roubik, D. W., and Neumann, P. (2012). Back to the future: Apis versus non-Apis pollination. Trends Ecol. Evol. 27, 142–143. doi: 10.1016/j.tree.2011.11.017
Bailey, S., Requier, F., Nusillard, B., Roberts, S. P., Potts, S. G., and Bouget, C. (2014). Distance from forest edge affects bee pollinators in oilseed rape fields. Ecol. Evol. 4, 370–380. doi: 10.1002/ece3.924
Bates, D., Maechler, M., and Bolker, B. (2011). me4: Linear Mixed-Effects Models Using S4 Classes. R Package Version 0.999375–42. Available online at: http://CRAN.R-project.org/package=lme4
Biddinger, D. J., Joshi, N. K., Rajotte, E. G., Halbrendt, N. O., Pulig, C., Naithani, K. J., et al. (2013). An immunomarking method to determine the foraging patterns of Osmia cornifrons and resulting fruit set in a cherry orchard. Apidologie 44, 738–749. doi: 10.1007/s13592-013-0221-x
Blitzer, E. J., Dormann, C. F., Holzschuh, A., Klein, A. M., Rand, T. A., and Tscharntke, T. (2012). Spillover of functionally important organisms between managed and natural habitats. Agric. Ecosyst. Environ. 146, 34–43. doi: 10.1016/j.agee.2011.09.005
Cameron, S. A., Lozier, J. D., Strange, J. P., Koch, J. B., Cordes, N., Solter, L. F., et al. (2011). Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. U.S.A. 108, 662–227. doi: 10.1073/pnas.1014743108
Cariveau, D. P., Williams, N. M., Benjamin, F. E., and Winfree, R. (2013). Response diversity to land use occurs but does not consistently stabilise ecosystem services provided by native pollinators. Ecol. Lett. 16, 903–911. doi: 10.1111/ele.12126
Christmann, S., and Aw-Hassan, A. A. (2012). Farming with alternative pollinators (FAP)—an overlooked win–win strategy for climate change adaptation. Agric. Ecosyst. Environ. 161, 161–164. doi: 10.1016/j.agee.2012.07.030
Egan, J. F., and Mortensen, D. A. (2012). A comparison of land-sharing and land-sparing strategies for plant richness conservation in agricultural landscapes. Ecol. Appl. 22, 459–471. doi: 10.1890/11-0206.1
Garibaldi, L. A., Steffan-Dewenter, I., Kremen, C., Morales, J. M., Bommarco, R., Cunningham, S. A., et al. (2011). Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 14, 1062–1072. doi: 10.1111/j.1461-0248.2011.01669.x
Garibaldi, L. A., Steffan-Dewenter, I., Winfree, R., Aizen, M. A., Bommarco, R., Cunningham, S. A., et al. (2013). Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339, 1608–1611. doi: 10.1126/science.1230200
Garratt, M. P. D., Truslove, C. L., Coston, D. J., Evans, R. L., Moss, E. D., Dodson, C., et al. (2014). Pollination deficits in UK apple orchards. J. Pollinat. Ecol. 12, 9–14.
Gary, N. E., Witherell, P. C., and Lorenzen, K. (1981). Effect of age on honey bee (Hymenoptera, Apidae) foraging distance and pollen collection. Environ. Entomol. 10, 950–952. doi: 10.1093/ee/10.6.950
Greenleaf, S. S., and Kremen, C. (2006). Wild bees enhance Apis mellifera pollination of hybrid sunflower. Proc. Natl. Acad. Sci. U.S.A. 103, 13890–13895. doi: 10.1073/pnas.0600929103
Greenleaf, S. S., Williams, N. M., Winfree, R., and Kremen, C. (2007). Bee foraging ranges and their relationship to body size. Oecologia 153, 589–596. doi: 10.1007/s00442-007-0752-9
Grixti, J. C., Wong, L. T., Cameron, S. A., and Favret, C. (2009). Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 142, 75–84. doi: 10.1016/j.biocon.2008.09.027
Hagen, M., and Kraemer, M. (2010). Agricultural surroundings support flower-visitor networks in an Afrotropical rain forest. Biol. Conserv. 143, 1654–1663. doi: 10.1016/j.biocon.2010.03.036
Holzschuh, J., Dudenhöffer, H., and Tscharntke, T. (2012). Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biol. Conserv. 153, 101–107. doi: 10.1016/j.biocon.2012.04.032
Hull, L. A., Joshi, N. K., and Zaman, F. (2009). Management of internal feeding lepidopteran pests in apple, 2008. Arthropod Manage. Tests 34. doi: 10.4182/amt.2009.A8
Inouye, D. W. (2007). The value of bees. Biol. Conserv. 140, 198–199. doi: 10.1016/j.biocon.2007.09.011
Javorek, S., Mackenzie, K., and Vander Kloet, S. (2002). Comparative pollination effectiveness among bees (Hymenoptera: Apoidea) on lowbush blueberry (Ericaceae: Vaccinium angustifolium). Ann. Entomol. Soc. Am. 95, 345–351. doi: 10.1603/0013-8746(2002)095[0345:CPEABH]2.0.CO;2
Jha, S., and Kremen, C. (2013). Resource diversity and landscape-level homogeneity drive native bee foraging. Proc. Natl. Acad. Sci. U.S.A. 110, 555–558. doi: 10.1073/pnas.1208682110
Joshi, N. K., Leslie, T., Rajotte, E. G., Kammerer, M. A., Otieno, M., and Biddinger, D. J. (2015). Comparative trapping efficiency to characterize bee abundance, diversity and community composition in apple orchards. Ann. Entomol. Soc. 108, 785–799. doi: 10.1093/aesa/sav057
Karanja, R. H., Njoroge, G. N., Gikungu, M. W., and Newton, L. E. (2010). Bee interactions with wild flora around organic and conventional coffee farms in Kiambu district, central Kenya. J. Pollinat. Ecol. 2, 7–12.
Kennedy, C. M., Lonsdorf, E., Neel, M. C., Williams, N. M., Ricketts, T. H., Winfree, R., et al. (2013). A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 16, 584–599. doi: 10.1111/ele.12082
Klein, A. M., Brittain, C., Hendrix, S. D., Thorp, R., Williams, N., and Kremen, C. (2012). Wild pollination services to California almond rely on semi-natural habitat. J. Appl. Ecol. 49, 723–732. doi: 10.1111/j.1365-2664.2012.02144.x
Klein, A. M., Steffan-Dewenter, I., and Tscharntke, T. (2003). Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc. B. 270, 955–961. doi: 10.1098/rspb.2002.2306
Kremen, C., Williams, N. M., Bugg, R. L., Fay, J. P., and Thorp, R. W. (2004). The area requirements of an ecosystem service: crop pollination by native bee communities in California. Ecol. Lett. 7, 1109–1119. doi: 10.1111/j.1461-0248.2004.00662.x
Lonsdorf, E., Kremen, C., Ricketts, T., Winfree, R., Williams, N., and Greenleaf, S. (2009). Modelling pollination services across agricultural landscapes. Ann. Bot. 103, 1589–1600. doi: 10.1093/aob/mcp069
Marini, L., Quaranta, M., Fontana, P., Biesmeijer, J. C., and Bommarco, R. (2012). Landscape context and elevation affect pollinator communities in intensive apple orchards. Basic Appl. Ecol. 13, 681–689. doi: 10.1016/j.baae.2012.09.003
McGarigal, K. (2014). FRAGSTATS HELP. Available online at: http://www.umass.edu/landeco/research/fragstats/documents/fragstats.help.4.2.pdf
McGarigal, K., Cushman, S. A and Ene, E. (2012). “FRAGSTATS V4: spatial pattern analysis program for categorical and continuous maps,” in Computer Software Program Produced (University of Massachusetts, Amherst). Available online at: http://www.umass.edu/landeco/research/fragstats/fragstats.html
Merwin, I. A. (2003). “Orchard floor management systems,” in Apples: Botany, Production, and Uses, ed D. C. Ferree (Wallingford: CABI Publications), 303–318.
Olsson, O., Bolin, A., Smith, H. G., and Lonsdorf, E. V. (2015). Modeling pollinating bee visitation rates in heterogeneous landscapes from foraging theory. Ecol. Model. 316, 133–143. doi: 10.1016/j.ecolmodel.2015.08.009
Otieno, M., Woodcock, B. A., Wilby, A., Vogiatzakis, I. N., Mauchline, A. L., Gikungu, M. W., et al. (2011). Local management and landscape drivers of pollination and biological control services in a Kenyan agro-ecosystem. Biol. Conserv. 144, 2424–2431. doi: 10.1016/j.biocon.2011.06.013
Pakeman, R. J. (2011). Multivariate identification of plant functional response and effect traits in an agricultural landscape. Ecology 92, 1353–1365. doi: 10.1890/10-1728.1
Penn State Extension (2014). Tree Fruit Production Guide. Available online at: http://extension.psu.edu/plants/tree-fruit/tfpg (Accessed October 05, 2015).
Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O., and Kunin, W. E. (2010). Global pollinator declines; trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353. doi: 10.1016/j.tree.2010.01.007
R Development Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rao, S., and Strange, J. P. (2012). Bumble bee (Hymenoptera: Apidae) foraging distance and colony density associated with a late-season mass flowering crop. Environ. Entomol. 41, 905–915. doi: 10.1603/EN11316
Ricketts, T. H., Regetz, J., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C., Bogdanski, A., et al. (2008). Landscape effects on crop pollination services: are there general patterns? Ecol. Lett. 11, 499–515. doi: 10.1111/j.1461-0248.2008.01157.x
Schweiger, O., Biesmeijer, J. C., Bommarco, R., Hickler, T., Hulme, P. E., Klotz, S., et al. (2010). Multiple stressors on biotic interactions: how climate change and alien species interact to affect pollination. Biol. Rev. 85, 777–795. doi: 10.1111/j.1469-185x.2010.00125.x
Sheffield, C. S. (2014). Pollination, seed set and fruit quality in apple: studies with Osmia lignaria (Hymenoptera: Megachilidae) in the Annapolis Valley, Nova Scotia, Canada. J. Pollinat. Ecol. 12, 120–128.
Sheffield, C. S., Kevan, P. G., Pindar, A., and Packer, L. (2013). Bee (Hymenoptera: Apoidea) diversity within apple orchards and old fields in the Annapolis Valley, Nova Scotia, Canada. Can. Entomol. 145, 94–114. doi: 10.4039/tce.2012.89
Smith, K. M., Loh, E. H., Rostal, M. K., Zambrana-Torrelio, C. M., Mendiola, L., and Daszak, P. (2013). Pathogens, pests, and economics: Drivers of honey bee colony declines and losses. Ecohealth 10, 434–445. doi: 10.1007/s10393-013-0870-2
Steffan-Dewenter, I., Münzenberg, U., Bürger, C., Thies, C., and Tscharntke, T. (2002). Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83, 1421–1432. doi: 10.1890/0012-9658(2002)083[1421:SDEOLC]2.0.CO;2
Tepedino, V. J., Alston, D. G., Bradley, B. A., Toler, T. R., and Griswold, T. L. (2007). Orchard pollination in Capitol Reef National Park, Utah, USA. Apis mellifera or native bees? Biodivers. Conserv. 16, 3083–3094. doi: 10.1007/s10531-007-9164-8
ter Braak, C. J. F., and Šmilauer, P. (2012). CANOCO Reference Manual and User's Guide: Software for Ordination (Version 5.0). Microcomputer Power. Ithaca, NY.
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I., and Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity—ecosystem service management. Ecol. Lett. 8, 857–874. doi: 10.1111/j.1461-0248.2005.00782.x
Vicens, N., and Bosch, J. (2000). Weather-Dependent Pollinator Activity in an Apple Orchard, with Special Reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ. Entomol. 29, 413–420. doi: 10.1603/0046-225X-29.3.413
Watson, J. C., Wolf, A. T., and Ascher, J. S. (2011). Forested landscapes promote richness and abundance of native bees (Hymenoptera: Apoidea: Anthophila) in Wisconsin apple orchards. Environ. Entomol. 40, 621–632. doi: 10.1603/EN10231
Westphal, C., Steffan-Dewenter, I., and Tscharntke, T. (2003). Mass flowering crops enhance pollinator densities at landscape scale. Ecol. Lett. 6, 961–965. doi: 10.1046/j.1461-0248.2003.00523.x
Westrich, P. (1996). “Habitat requirements of Central European bees and the problems of partial habitats,” in The Conservation of Bees. Linnean Society Symposium Series, eds A. Matheson, I. H. Buchmann, C. O'Toole, P. Westrichand, and I. H. Williams (London: Academic Press), 1–16.
Williams, N. M., and Thomson, J. D. (2003). “Comparing pollinator quality of Apis mellifera (Hymemoptera: Apidae) and native bees using pollen removal and deposition measures,” in For Nonnative crops, Whence Pollinators of the Future?, eds K. Strickler and J. H. Cane (Lanham, MD: Entomol Soc Amer), 163–179.
Wilson, P., and Thomson, J. (1991). Heterogeneity among floral visitors leads to discordance between removal and deposition of pollen. Ecology 72, 1503–1507. doi: 10.2307/1941124
Winfree, R., Williams, N. M., Dushoff, J., and Kremen, C. (2007). Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 10, 1105–1113. doi: 10.1111/j.1461-0248.2007.01110.x
Winfree, R., Williams, N. M., Gaines, H., Ascher, J. S., and Kremen, C. (2008). Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J. Appl. Ecol. 45, 793–802. doi: 10.1111/j.1365-2664.2007.01418.x
Zhang, W., Ricketts, T. H., Kremen, C., Carney, K., and Swinton, S. M. (2007). Ecosystem services and dis-services to agriculture. Ecol. Econ. 64, 253–260. doi: 10.1016/j.ecolecon.2007.02.024
Keywords: apple, bees, heterogeneous landscape, pollination, largest patch index, mean proximity index, ecosystem service
Citation: Joshi NK, Otieno M, Rajotte EG, Fleischer SJ and Biddinger DJ (2016) Proximity to Woodland and Landscape Structure Drives Pollinator Visitation in Apple Orchard Ecosystem. Front. Ecol. Evol. 4:38. doi: 10.3389/fevo.2016.00038
Received: 31 December 2015; Accepted: 31 March 2016;
Published: 22 April 2016.
Edited by:
Evangelia G. Drakou, University of West Brittany, FranceCopyright © 2016 Joshi, Otieno, Rajotte, Fleischer and Biddinger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Neelendra K. Joshi, nkjoshi@uark.edu;
Mark Otieno, mmarkotieno@gmail.com
†Present Address: Mark Otieno, Department of Agricultural Resource Management, Embu University College, Embu, Kenya
‡These authors have contributed equally to this work and are co-first authors.