An Experimental Test of a Causal Link between Problem-Solving Performance and Reproductive Success in Wild Great Tits
 Laure Cauchard1*
Laure Cauchard1*  Bernard Angers1
Bernard Angers1  Neeltje J. Boogert2
Neeltje J. Boogert2  Mélissa Lenarth3
Mélissa Lenarth3  Pierre Bize3,4†
Pierre Bize3,4†  Blandine Doligez5,6†
Blandine Doligez5,6†- 1Département de Sciences Biologiques, Université de Montréal, Montréal, QC, Canada
- 2Centre for Ecology and Conservation, University of Exeter, Penryn, United Kingdom
- 3Département D'écologie et Évolution, Université de Lausanne, Lausanne, Switzerland
- 4Institute of Biological and Environmental Sciences, University of Aberdeen, Aberdeen, United Kingdom
- 5Department of Biometry and Evolutionary Biology, Centre National de la Recherche Scientifique, University of Lyon 1, Villeurbanne, France
- 6Animal Ecology, Department of Ecology and Genetics, Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden
Recent studies have uncovered relationships between measures of various cognitive performances and proxies of fitness such as reproductive success in non-human animals. However, to better understand the evolution of cognition in the wild, we still have to determine the causality of these relationships and the underlying mechanisms. The cognitive ability of an individual may directly influence its ability to raise many and/or high quality young through for example its provisioning ability. Conversely, large and/or high quality broods may lead to high parental motivation to solve problems related to their care. To answer this question, we manipulated reproductive success through brood size and measured subsequent problem-solving performance in wild great tit parents. Our results show that brood size manipulation did not affect the probability to solve the task. Moreover, solver pairs fledged more young than non-solver pairs independently of brood size treatment in one of the two experimental years and they showed higher nestling provisioning rate in both years. Overall, it shows that problem-solving performance was not driven by motivation and suggest that problem-solvers may achieve higher fledging success through higher provisioning rates. Our study constitutes a first key step toward a mechanistic understanding of the consequences of innovation ability for individual fitness in the wild.
Introduction
Human activities currently generate major and rapid environmental changes at various spatio-temporal scales (e.g., climate change, urbanization, habitat fragmentation) that can strongly impact individual fitness in wild populations. To limit the negative impacts of these changes, animals may attempt to cope with the unexpected problems or situations by innovating, i.e., using novel or flexibly adjust established behaviors (Reader and Laland, 2003; Tebbich et al., 2010). Such ability shapes behavioral changes that are thought to facilitate a rapid response to novel environmental conditions before adaptive evolution can take place (Duckworth and Badyaev, 2007; Sutter and Kawecki, 2009). Yet, research on the consequences of innovation on behavioral adaptation in non-human animals and its contribution to fitness in the wild is still in its infancy.
Innovation may influence fitness through different ways. For example, innovation can increase survival by facilitating the exploitation of new food sources during harsh conditions (Kozlovsky et al., 2015; Edmunds et al., 2016). It can also enhance mating and reproductive success by facilitating the display of a new behavior or phenotypic trait favored during mate choice (Keagy et al., 2009; Mateos-Gonzalez et al., 2011; Isden et al., 2013) or the exploitation of resources that will directly influence young growth and/or survival (Cole et al., 2012; Cauchard et al., 2013). The first empirical evidence for a positive association between innovation and reproductive success in the wild came from two studies conducted on two different populations of great tits (Parus major) (Cole et al., 2012; Cauchard et al., 2013). In these studies, problem-solving performance (a proxy of innovation, Griffin and Guez, 2014), measured either in captivity (stick-pulling task motivated by food, Cole et al., 2012) or in the field (string-pulling task motivated by nestling provisioning, Cauchard et al., 2013), was linked to various measures of reproductive success. Nests where at least one parent solved the novel task laid larger clutches (Cole et al., 2012; Cauchard et al., 2013) and fledged more young (Cauchard et al., 2013), although this effect was counterbalanced by problem-solving females being more likely to abandon their nest after human perturbation in one of the populations (Cole et al., 2012).
However, the mechanisms underlying potential reproductive benefits of problem-solving performance remain poorly explored. Moreover, as our understanding of the proximal causes of among-individual differences in problem-solving performance has considerably improved in the last decade, it has become apparent that various other factors may generate an indirect link between problem-solving performance and reproductive success. For example, studies have shown that individual traits such as age (Loepelt et al., 2016; Quinn et al., 2016), novelty response (Sol et al., 2011), stress level (Bókony et al., 2013) or motivation (Laland and Reader, 1999) as well as external factors such as predation (Taylor et al., 2012), competition (Overington et al., 2009) or habitat quality (Quinn et al., 2016) can affect both problem-solving performance and reproductive success independently.
Therefore, an experimental manipulation is critically needed to disentangle cause and effect in the relationship between problem solving performance and reproductive success. Here we manipulated the reproductive success (i.e., reduced, control, or enlarged brood size) and recorded parents' subsequent problem-solving performance as well as nestling provisioning rate in a natural population of breeding great tits. We predicted that if reproductive success drives parental motivation to solve the task, parents with experimentally increased broods should be more successful at solving the task than parents with control broods and parents with control broods should be more successful than parents with decreased broods. Conversely, if higher problem-solving performance per se allows parents to raise more young, our experimental brood size manipulation should not affect parents' success in solving the task and solving pairs should still achieve higher reproductive success compared to non-solving pairs independently of the brood size manipulation. Moreover, beside the brood size manipulation, if problem-solvers are more efficient in exploiting their habitat, we predicted that solver pairs should achieve a higher nestling provisioning rate than non-solver pairs.
Materials and Methods
Study Site and Population Monitoring
Data were collected in a population of great tits breeding on the island of Gotland, Sweden (57°10'N, 18°20'E), between April and June 2012 and 2013. Great tits are small, monogamous passerines that breed readily in nest boxes, allowing us to record laying and hatching date, clutch size, hatching success, brood size at different ages and final number of fledglings. Nestlings were ringed at day 9, weighed and measured (tarsus length, to the nearest 0.1 mm) at day 14. Nestling body mass at day 14 is a good proxy of future recruitment in this species (Linden et al., 1992). Adults were caught within nest boxes when nestlings were 9–14 days old and identified using individually numbered rings.
Birds were caught, handled and ringed under a license from the Stockholm Museum Ringing Center. Behavioral experiments were authorized by the Swedish Committee for Experiments on Animals and conducted in accordance with international standards on animal welfare as well as being compliant with local and national regulations.
Brood Size Manipulation
In our great tit population, brood size range from 3 to 12 nestlings, with an average ± SE of 8.06 ± 0.13. In total, 150 broods were manipulated. We created enlarged (N = 57), reduced (N = 54) and control (N = 39) broods by adding or removing two nestlings (i.e., an average 25% increase or decrease in brood size) or exchanging two nestlings between broods without changing brood size. The difference between reduced and increased brood size treatments was thus on average 40% (6 vs. 9 nestlings, see Table 1), which seemed sufficient to allow detecting differences in parental care. Studies have shown that brood size manipulation using quantitatively similar changes in brood size has significant consequences on parental feeding behavior, with provisioning rate being lowest when broods were reduced in size and greatest when broods were enlarged (Sanz and Tinbergen, 1999; Garcia-Navas and Sanz, 2010). We excluded from this experiment nests with extreme brood sizes before the manipulation (<5 or more than 10 hatchlings on day 2) and assigned treatment (reduced, control, or enlarged brood) randomly with respect to initial brood size. Two days after hatching, nestlings were exchanged between broods that hatched on the same day and matched the same average weight (mean brood weight per nest ± SD: 2.72 g ± 0.47, nests were grouped when the mean difference among nests was <1 g). Whenever possible, we used triplets of broods: four nestlings were transferred from a first nest (nest A) to a second nest (nest B), then four other nestlings from nest B to a third nest (nest C), and finally two other nestlings from nest C to nest A. We thus reduced brood size by two nestlings in nests A, increased it by two nestlings in nests C and left it unchanged in nests B, which functioned as a control for having foreign nestlings without changing brood size. All broods thus contained either two (nests A) or four foreign nestlings (nests B and C). When we could not match three broods, we used duos by skipping the control treatment (nest B), and when more broods could be matched, we used quadruplets of nests by repeating the control treatment (nest B). In great tits, the number of native vs. foreign nestlings in a nest does not affect provisioning rate (Neuenschwander et al., 2003).
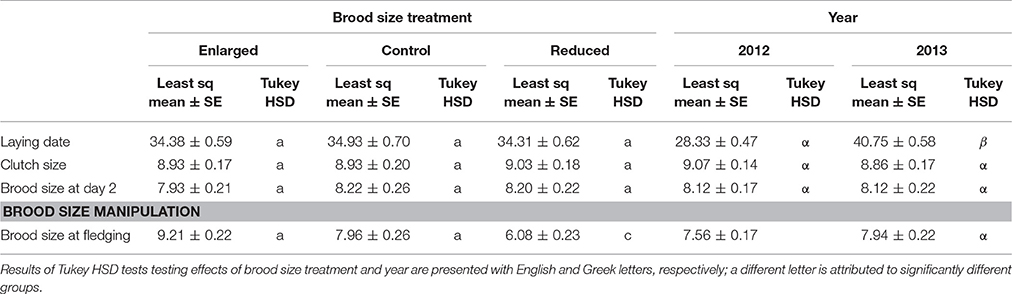
Table 1. Least square means ± SE for reproductive variables according to experimental treatments (reduced: N = 54; enlarged: N = 57; control: N = 39) and year (2012: N = 93; 2013: N = 57).
Provisioning Rate and Problem-Solving Performance
We recorded provisioning rate when nestlings were 6 days old, using a camouflaged video recorder placed at a distance of ~6 m from the nest box. The recording lasted 90 min and was performed during the daily peak of parental provisioning activity, i.e., between 06:00 AM and 02:00 PM. In great tits, both parents feed their young. Because distinguishing males from females on provisioning videos was difficult, we measured nestling provisioning rate per breeding pair. We calculated pair provisioning rate as the total number of parental visits to the nest during 1 h.
Problem-solving performance was measured directly during breeding. At this stage, it is however not possible to keep birds long enough to test them in captivity, in controlled conditions, without directly compromising their reproductive success. Therefore, we chose to conduct the problem-solving task directly in the wild. In this situation, food-motivated tasks cannot easily be used since food is more abundant in the environment at that time of the year than for the rest of the year and individuals show little motivation to interact with such tasks. To overcome this issue, problem-solving performance was thus measured using a string-pulling task attached in front of the nest box for which the solving motivation stems from parents' drive to feed their young during the nestling rearing period (Cauchard et al., 2013). The task consisted of a door placed in front of the entrance of the nest box. The door was by default closed. To enter, parents had to pull a string placed below the door using their feet to open it and then slip their body under the door. The door then closed automatically behind the bird, but could be simply pushed open from inside the nest box by parents to get out. The test was conducted during the peak of nestling food demand (i.e., when nestlings were 7–9 days old, between 07:00 AM and 04:00 PM), only when nestlings were satiated (e.g., not begging intensely at the beginning of the test). To avoid nestling starvation if parents were not able to solve the task, the test lasted 1 h but was repeated on two consecutive days. We randomly selected breeding pairs to be tested among pairs separated by at least 200 m from the nearest neighbors previously tested, to avoid social learning. We installed a camouflaged video recorder at a distance of ~6 m in front of the nest box to record all the movements and interactions of parents with the task. Video recordings were scored by observers blind to the brood size manipulation. Because the entrance of the nest was closed during the test, birds had to stop on the nest and the gender was thus identified thanks to plumage features. Individuals who succeeded in solving the task (i.e., opening the door and entering the box) were considered to be solvers, while those who contacted the nest box but failed to enter were considered to be non-solvers (i.e., we defined problem-solving status as a binary variable).
Statistical Analyses
We first checked whether nests in different brood size treatments (i.e., reduced, control, or enlarged broods) differed in reproductive parameters prior to the brood size manipulation (i.e., laying date, clutch size and number of nestlings at day 2) using linear models (LM) including brood size treatment, year (i.e., 2012, 2013) and their pairwise interactions as fixed effects. We proceeded as well to check whether the brood size manipulation was successful in affecting the final number of fledglings.
We then tested whether brood size manipulation affected parental problem-solving probability (i.e., solvers vs. non-solvers) using generalized linear mixed models (GLMMs) with a binomial error and logit link function. The model included brood size treatment, year, sex and the pairwise interactions between treatment and cofactors as fixed effects, and pair identity as a random effect to account for the non-independence of pair members.
Finally, we tested whether problem-solving performance affected measures of reproductive success and provisioning rate independently from the brood size manipulation. We conducted these analyses at the pair level to avoid pseudoreplication since both parents had the same measures of reproductive success and provisioning rate. We tested whether pairs where both parents were non-solvers (NN pairs), pairs where only one parent solved the task (NS pairs) and pairs where both parents solved the task (SS pairs) differed in nestling mean nestling body mass at day 14, final number of fledglings and provisioning rate at day 6 using LMs. The models included pair's problem-solving performance, brood size treatment, year and their pairwise interactions with problem-solving performance as fixed effects. When analyzing mean nestling body mass, we added mean nestling tarsus length as a covariate to control for the effect of structural size on body mass. To check whether reproductive parameters prior to the brood size manipulation may account for the differences in final reproduction success and provisioning rate between pairs of different problem-solving performance, we also included laying date and clutch size as fixed covariates in the models described above. Inclusion of these covariates did not change qualitatively our results on effects of pair solving performance. Hence, hereafter we are only presenting the reduced models.
Sample sizes varied slightly between analyses because of missing data. To avoid pseudoreplication, we removed from our data all 2013 pairs in which at least one parent was tested in 2012 (N = 10). Non-significant effects were backward eliminated from the starting models. Normality and homogeneity of variance as well as residuals were visually checked and data transformed when needed (in this case, the transformation used is mentioned in the results). All the analyses, including power analyses, were performed using JMP® (Version 11. SAS Institute Inc., Cary, NC, 1989–2007) at the exception of the GLMM tests that were performed using the glmer function in R cran (Bates et al., 2015). Tests were two-tailed, and P < 0.05 were considered significant.
Results
Differences between Treatment Groups Before and After the Brood Size Manipulation
Prior to brood size manipulation, nests of different experimental treatments did not differ in laying date [F(2, 146) = 0.25, P = 0.78], clutch size [F(2, 146) = 0.10, P = 0.90] or number of nestlings at day 2 [F(2, 143) = 0.55, P = 0.58], accounting for year (Table 1). As expected, brood size manipulation successfully affected the final number of fledglings [F(2, 146) = 50.59, P < 0.001], accounting for year. More nestlings fledged from increased broods (least square means ± SE: 9.2 ± 0.2) compared to control broods (8.0 ± 0.3), and more in control broods compared to decreased broods (6.1 ± 0.2) (Table 1).
Effect of Brood Size Manipulation on Problem-Solving Performance
Of the 150 pairs tested, nine males and eight females did not participate in the problem-solving test. The brood size manipulation treatment did not influence parental probability to solve the task either alone ( = 1.20, P = 0.55) or in interaction with year ( = 1.45, P = 0.48) or sex ( = 1.00, P = 0.61). The probability to solve the task only depended on sex ( = 11.21, P < 0.001), with females being more likely to solve compared to males (number of solvers: 58 of 142 (40.8%) females vs. 32 of 141 (22.7%) males).
Links between Problem-Solving Performance, Provisioning Rate and Reproductive Success Independently from the Brood Size Manipulation
Mean nestling body mass at day 14 did not differ between SS, NS and NN pairs [F(2, 137) = 0.95, P = 0.39], accounting for the positive effect of mean nestling tarsus length [F(1, 137) = 167.26, P < 0.001] and for differences between years [i.e., nestlings were heavier in 2012 than 2013; F(1, 137) = 5.82, P = 0.017]. Brood size treatment had no effect on mean nestling body mass at day 14 either alone [F(2, 137) = 0.68, P = 0.51] or in interaction with year [F(2, 137) = 0.13, P = 0.88].
The final number of fledglings differed between pairs of different problem-solving performance, but this effect depended on year [interaction between pair problem-solving status and year: F(2, 139) = 5.12, P = 0.007; Figure 1], after controlling for the effect of brood size treatment [F(2, 139) = 54.92, P < 0.001]. In 2012, SS pairs fledged more young than NS and NN pairs [F(2, 87) = 7.18, P = 0.001; Figure 1]. In 2013, there was no difference in final number of fledglings between pairs of different problem-solving performance [F(2, 50) = 1.46, P = 0.24]. Post-hoc power analyses suggest that the absence of difference in 2013 is also due to a lower effect of problem-solving performance on the final number of fledglings in 2013 than 2012 (δ = 0.35 vs. 0.58) rather than merely due to smaller sample sizes in 2013 than 2012 (N = 55 vs. 92). The effect of pair problem-solving performance on the final number of fledglings was independent of brood size treatment [i.e., non-significant interaction between problem-solving performance and brood-size treatment: F(4, 135) = 0.86, P = 0.49].
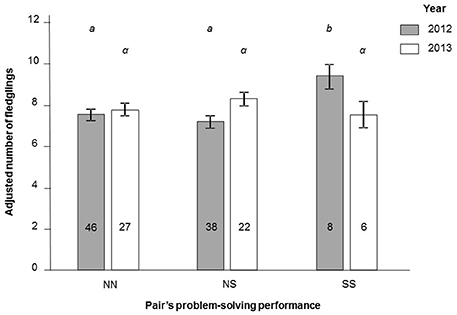
Figure 1. Final number of fledglings (least square means ± SE) according to pair's problem-solving performance (NN, pairs where both parents were non-solvers; NS, pairs with one solver; SS, pairs with two solvers) and year (2012, 2013) in a natural population of great tits. Values are adjusted for the other significant effect of the model, i.e., effect of brood size treatment. Letters represent results of Tukey HSD test, where different letters are attributed to significantly different groups. Numbers are sample sizes.
Provisioning rates also differed between pairs of different problem-solving performance [provisioning rates Box-Cox transformed; F(2, 103) = 5.20, P = 0.007; Figure 2], after controlling for the effects of brood size treatment [i.e., tendency for higher rates in enlarged vs. reduced broods; F(2, 103) = 2.98, P = 0.055] and year [i.e., higher rates in 2012 than in 2013; F(1, 103) = 10.38, P = 0.002]. SS pairs and NS pairs showed higher provisioning rates than NN pairs (SS vs. NN pairs: mean difference ± SE = 11.74 ± 4.40, P = 0.024; NS vs. NN pairs: 6.80 ± 2.78, P = 0.043; Figure 2). Because provisioning rate can vary between males and females in this species (Pagani-Núñez and Senar, 2013), we checked whether the sex of the solver influences the link between problem-solving performance and provisioning rate. We ran the same initial model using both female and male problem-solving status as fixed factors instead of pair problem-solving performance. Results showed that both were significant [males: F(1, 95) = 4.9, P = 0.028; Females: F(1, 95) = 4.3, P = 0.040].
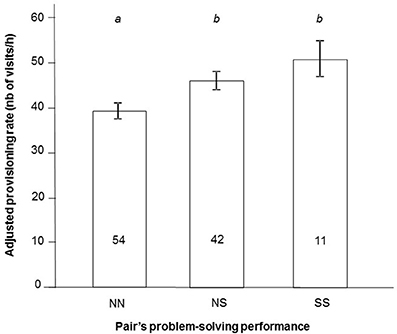
Figure 2. Pair's provisioning rates (least square means ± SE) according to its problem-solving performance (NN, pairs where both parents were non-solvers; NS, pairs with one solver; SS, pairs with two solvers) in a natural population of great tits. Values are adjusted for the other significant effects of the model, i.e., effects of brood size treatment and year. Letters represent results of Tukey HSD test, where different letters are attributed to significantly different groups. Numbers are sample size.
Discussion
Our first aim was to test the causality of the relationship between problem-solving performance and reproductive success in our study population. While the cognitive features of problem-solving (e.g., the cognitive and neurological processes of problem-solving) still need to be indentified, we experimentally showed here that parental motivation to solve the task, manipulated through the brood size manipulation (Sanz and Tinbergen, 1999; Garcia-Navas and Sanz, 2010), did not affect problem-solving performance, and thus could not generate the observed relation between problem-solving performance and reproductive success. Our results thus support the hypothesis that higher problem-solving performance per se might allow parents to raise more young. Accordingly, pairs' problem-solving performance was positively correlated to the final number of fledglings for each brood size treatment, although this relation was observed only in one of the two experimental years: in 2012, solver pairs fledged more young than pairs with at least one non-solver parent, beyond the brood size manipulation. Moreover, this positive correlation did not depend on brood size treatment (no significant interaction between problem-solving performance and brood size treatment). One could have expected problem-solving performance to affect the ability of the pair to cope with the manipulated reproductive effort differently depending on treatment, for example if solver and non-solver parents differ in their ability to cope with a change in parental work load and stress. On the one hand, all pairs could have achieved a similar reproductive success when brood size was decreased, i.e., when reproductive effort and thus the level of stress were low, while only pairs with high problem-solving performance may have been able to efficiently face an increased brood size if solvers better cope with stress. On the other hand, pairs with high problem-solving performance may have been able to face reproductive effort more efficiently than pairs with low problem-solving performance when brood size was reduced or unchanged, i.e., when the level of stress was low to moderate, but may not have been able to do so when the brood size was increased if solvers are not able to use their cognitive abilities adequately when stressed. Here, the ability of pairs with high problem-solving performance to achieve higher reproductive success than pairs with lower performance did not depend on the level of reproductive effort imposed through the manipulation, but depended on year. Taken together, these results provide support for the hypothesis that problem-solving performance may causally influence reproductive success in our study population, depending on environmental conditions.
At this stage, the origin of the difference in the relation between problem-solving performance and final number of fledglings between the two experimental years remains however unclear and deserves further studies. The absence of a significant difference between solvers and non-solvers in the final number of fledged young in 2013 was nonetheless not simply due to a lower sample size, but to a lower biological effect of problem-solving performance on reproductive success in that year. Although we cannot test this hypothesis because the experiment was performed in 2 years only, between-year variations in the environmental conditions might explain the difference between years observed. Table 1 shows that birds laid eggs earlier in 2012 than in 2013 (which was an extremely late year for forest passerines in most parts of Europe), suggesting a difference in the environment between the 2 years because great tits synchronized their reproduction according to caterpillar (i.e., main food resource for nestlings) development (Naef-Daenzer and Keller, 1999). Moreover, nestlings were heavier in 2012 than in 2013, suggesting that the environmental conditions in 2013 might have been harsher than in 2012. In our study site, food availability during nestling rearing is the main environmental factor affecting reproductive success (nest predation is very low on Gotland due to the absence of mustelid species; Doligez and Clobert, 2003). The higher success in 2012 compared to 2013 suggests that environmental conditions were more favorable in 2012, but we have no direct measure of food availability in these 2 years. The observed difference between years in the link between pair problem-solving performance and reproductive success could therefore be due to higher costs paid by solving pairs in harsher conditions, but higher benefits in more standard conditions, possibly due to a higher ability to exploit the environment when provisioning nestlings. Although the link between pair problem-solving ability and provisioning rate was observed in both years, other variables related to parental care and thus influencing fledgling number might vary between solvers and non-solvers according to environmental variation (Récapet et al., 2016). Hence, the links between parental cognitive ability, food provisioning and fitness may be context-dependent and vary according to the environmental conditions. Further work is however needed at this stage to identify such traits. Exploring the relative role of environmental vs. individual quality would require performing the manipulation over many years to meet a larger range of environmental conditions.
Research in humans has shown that two types of motivation can affect the expression of any cognitive ability: intrinsic and extrinsic motivation (Sternberg, 1985). While the extrinsic motivation is generally defined as the process by which an expected reward (or punishment) drives a behavior, its intensity and direction, the intrinsic motivation originates from individual traits (e.g., curiosity, interest, perseverance) and does not require reinforcement (Sternberg, 1985). These two types of motivation are likely to also affect the performance to cognitive tests in non-human species too (Hull, 1933; Sol et al., 2012; Byrne, 2013). Sex, social status or satiety can affect an individual's motivational state that, in turn, can directly impact both performance on food-motivated tasks and reproductive success. Because the basic level of motivation is bound to differ between individuals, for example due to differences in metabolic rate and condition (e.g., total energy reserve, Clancey and Byers, 2014), controlling for the effect of such motivation in correlative studies may be difficult. Relying on non-food-motivated tasks to design problem-solving tests, such as a species' aversion to a particular color (Keagy et al., 2009) or parents' drive to provision their young (Cauchard et al., 2013), may thus provide interesting alternatives to food-motivated tasks to measure problem-solving performance while minimizing the effect of extrinsic motivation. Indeed, even if females were more likely to solve the task than males, reflecting a potential sex-bias linked to parental investment in intrinsic motivation to solve the task (i.e., males may be less motivated to invest and solve our task because of extra pair paternity frequently occurring in this species; Lubjuhn et al., 1999; Griffith et al., 2002; Doligez, pers. obs for the study population), this is unlikely to affect the link between problem-solving performance and provisioning rate in our study: both male and female solving status related to provisioning rate. Our task may have been intrinsically more motivating for females than males, because the reward (i.e., access to nestlings) was directly related to parental care (which has already been observed in another population of great tits: Preiszner et al., 2017). Fortuitously, such sex difference can easily be taken into account by modeling the effects of sex when studying problem solving performance. To better understand the role of intrinsic and extrinsic in problem-solving performance, further work is required based on experiments explicitly designed to quantify motivation sources, although this may again prove challenging in the field.
An important caveat is nevertheless that we cannot exclude that a third factor, thus far unidentified, independently influenced both problem-solving performance and reproductive success. The deleterious effect of oxidative stress along aging, for example, can negatively affect both cognitive performance and reproductive success (Finkel and Holbrook, 2000; Fukui et al., 2002; Bize et al., 2008; Monaghan et al., 2009), leading to a positive correlation between these two variables. A direct experimental manipulation of problem-solving performance would be needed to confirm the causal link to reproductive success, but this may involve procedures such as manipulating conditions during development to affect the ontogeny of cognitive abilities in future recruits, which may prove difficult to implement in the field. Further work is therefore needed to fully confirm that problem-solving performance shapes reproductive success. To better understand the evolutionary potential of innovation in the wild, a more comprehensive work is needed to examine its link with long term fitness (i.e., lifetime reproductive success) consequences and as well as its heritability level in natural populations, two questions that remain largely unexplored so far (Quinn et al., 2016).
The final aim of this study was to investigate provisioning as a possible mechanism underlying a potential causal link between problem-solving performance and reproductive success. Our results showed that, whatever the level of reproductive effort imposed, pairs with at least one solver consistently outperformed pairs with non-solvers on terms of food provisioning rate, which did led to greater number of fledgling in one of the two study years. Thus, the ability to innovate might allow parents to provision their young more efficiently, either by (i) choosing and/or securing a higher quality breeding territory, or (ii) finding and/or selecting more and/or higher quality preys (Cole et al., 2012). Accordingly, solver pairs fledged more nestlings than pairs with at least one non-solver parent, without having to trade-offs nestling numbers against quality, as mean nestling body mass was not lower for solver pairs. Previous studies in great tits also showed clear positive links of food provisioning with brood size, but not with mean nestling body mass (Mutzel et al., 2013), supporting the hypothesis that solvers were able to raise more nestlings, at least in some years, due to their greater capacity to provision their brood. Further investigations, with a particular attention to the importance of territory quality and/or provisioning efficiency, are needed to improve our understanding of the behavioral mechanisms underlying a potential causal link between problem-solving performance and reproductive success. By exploring the role of motivation on problem-solving performance and differences in provisioning behavior in relation to problem-solving performance in the wild, our study constitutes nevertheless a first key step toward a mechanistic understanding of the consequences of innovation ability for individual fitness in the wild.
Author Contributions
LC and BD designed the study. LC, BA, NB, PB, and BD planned the research. LC and ML performed fieldwork. LC and PB analyzed data and all the co-authors contributed to write the paper.
Funding
This work was supported by a NSERC grant to BA, a PICS grant from the CNRS (INEE, n° 31520) to BD, a PhD writing up grant from the FESP (UdM) and a scholarship from the Biological Sciences Department (UdM) to LC and a mobility grant ERASMUS to ML. We are also grateful to the ABS, the BOU, the BES, the Frank M. Chapman from AMNH and the Fred Cooke from the SCO for awards and research grants attributed to LC.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Gotland'owners and Lars Gustafsson for access to the site, all the field assistants, and in particular Courcoux-Caro U., for their help with fieldwork and video analyses.
References
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67:48. doi: 10.18637/jss.v067.i01
Bize, P., Devevey, G., Monaghan, P., Doligez, B., and Christe, P. (2008). Fecundity and survival in relation to resistance to oxidative stress in a free-living bird. Ecology 89, 2584–2593. doi: 10.1890/07-1135.1
Bókony, V., Lendvai, Á. Z., Vágási, C. I., Pǎtraş, L., Pap, P. L., Németh, J., et al. (2013). Necessity or capacity? Physiological state predicts problem-solving performance in house sparrows. Behav. Ecol. 25, 124–135. doi: 10.1093/beheco/art094
Cauchard, L., Boogert, N. J., Lefebvre, L., Dubois, F., and Doligez, B. (2013). Problem-solving performance is correlated with reproductive success in a wild bird population. Anim. Behav. 85, 19–26. doi: 10.1016/j.anbehav.2012.10.005
Clancey, E., and Byers, J. A. (2014). The definition and measurement of individual condition in evolutionary studies. Ethology 120, 845–854. doi: 10.1111/eth.12272
Cole, E. F., Morand-Ferron, J., Hinks, A. E., and Quinn, J. L. (2012). Cognitive ability influences reproductive life history variation in the wild. Curr. Biol. 22, 1808–1812. doi: 10.1016/j.cub.2012.07.051
Doligez, B., and Clobert, J. (2003). Clutch size reduction as a response to increased nest preation rate in the collared flycatcher. Ecology 84, 2582–2588. doi: 10.1890/02-3116
Duckworth, R. A., and Badyaev, A. V. (2007). Coupling of dispersal and aggression facilitates the rapid range expansion of a passerine bird. Proc. Natl. Acad. Sci. U.S.A. 104, 15017–15022. doi: 10.1073/pnas.0706174104
Edmunds, N. B., Laberge, F., and McCann, K. S. (2016). A role for brain size and cognition in food webs. Ecol. Lett. 19, 948–955. doi: 10.1111/ele.12633
Finkel, T., and Holbrook, N. J. (2000). Oxidants, oxidative stress and the biology of ageing. Nature 408, 239–247. doi: 10.1038/35041687
Fukui, K., OMOI, N. O., Hayasaka, T., Shinnkai, T., Suzuki, S., Abe, K., et al. (2002). Cognitive impairment of rats caused by oxidative stress and aging, and its prevention by vitamin E. Ann. N.Y. Acad. Sci. 959, 275–284. doi: 10.1111/j.1749-6632.2002.tb02099.x
Garcia-Navas, V., and Sanz, J. J. (2010). Flexibility in the foraging behavior of blue tits in response to short-term manipulations of brood size. Ethology 116, 744–754. doi: 10.1111/j.1439-0310.2010.01788.x
Griffin, A. S., and Guez, D. (2014). Innovation and problem solving: a review of common mechanisms. Behav. Proces. 109, 121–134. doi: 10.1016/j.beproc.2014.08.027
Griffith, S. C., Owens, I. P. F., and Thuman, K. A. (2002). Extra pair paternity in birds: a review of interspecific variation and adaptive function. Mol. Ecol. 11, 2195–2212. doi: 10.1046/j.1365-294X.2002.01613.x
Hull, C. L. (1933). Differential habituation to internal stimuli in the albino rat. J. Comp. Psychol. 16, 255–273. doi: 10.1037/h0071710
Isden, J., Panayi, C., Dingle, C., and Madden, J. (2013). Performance in cognitive and problem-solving tasks in male spotted bowerbirds does not correlate with mating success. Anim. Behav. 86, 829–838. doi: 10.1016/j.anbehav.2013.07.024
Keagy, J., Savard, J. F., and Borgia, G. (2009). Male satin bowerbird problem-solving ability predicts mating success. Anim. Behav. 78, 809–817. doi: 10.1016/j.anbehav.2009.07.011
Kozlovsky, D., Branch, C., and Pravosudov, V. (2015). Problem-solving ability and response to novelty in mountain chickadees (Poecile gambeli) from different elevations. Behav. Ecol. Sociobiol. 69, 635–643. doi: 10.1007/s00265-015-1874-4
Laland, K. N., and Reader, S. M. (1999). Foraging innovation in the guppy. Anim. Behav. 57, 331–340. doi: 10.1006/anbe.1998.0967
Linden, M., Gustafsson, L., and Part, T. (1992). Selection on fledging mass in the collared flycatcher and the great tit. Ecology 73, 336–343. doi: 10.2307/1938745
Loepelt, J., Shaw, R. C., and Burns, K. C. (2016). Can you teach an old parrot new tricks? Cognitive development in wild kaka (Nestor meridionalis). Proc. R. Soc. B 283:1832. doi: 10.1098/rspb.2015.3056
Lubjuhn, T., Strohbach, S., Brun, J., Gerken, T., and Epplen, J. T. (1999). Extra-pair paternity in great tits (Parus major)—A long term study. Behaviour 136, 1157–1172. doi: 10.1163/156853999501810
Mateos-Gonzalez, F., Quesada, J., and Senar, J. C. (2011). Sexy birds are superior at solving a foraging problem. Biol. Lett. 7, 668–669. doi: 10.1098/rsbl.2011.0163
Monaghan, P., Metcalfe, N. B., and Torres, R. (2009). Oxidative stress as a mediator of life history trade-offs: mechanisms, measurements and interpretation. Ecol. Lett. 12, 75–92. doi: 10.1111/j.1461-0248.2008.01258.x
Mutzel, A., Dingemanse, N. J., Araya-Ajoy, Y. G., and Kempenaers, B. (2013). Parental provisioning behaviour plays a key role in linking personality with reproductive success. Proc. R. Soc. B 280:1764. doi: 10.1098/rspb.2013.1019
Naef-Daenzer, B., and Keller, L. F. (1999). The foraging performance of great and blue tits (Parus major and P-caerulens) in relation to caterpillar development, and its consequences for nestling growth and fledging weight. J. Anim. Ecol. 68, 708–718. doi: 10.1046/j.1365-2656.1999.00318.x
Neuenschwander, S., Brinkhof, M. W. G., Kölliker, M., and Richner, H. (2003). Brood size, sibling competition, and the cost of begging in great tits (Parus major). Behav. Ecol. 14, 457–462. doi: 10.1093/beheco/arg025
Overington, S. E., Cauchard, L., Morand-Ferron, J., and Lefebvre, L. (2009). Innovation in groups: does the proximity of others facilitate or inhibit performance? Behaviour 146, 1543–1564. doi: 10.1163/156853909X450131
Pagani-Núñez, E., and Senar, J. C. (2013). One hour of sampling is enough: great tit parus major parents feed their nestlings consistently across time. Acta Ornithol. 48, 194–200. doi: 10.3161/000164513X678847
Preiszner, B., Papp, S., Pipoly, I., Seress, G., Vincze, E., Liker, A., et al. (2017). Problem-solving performance and reproductive success of great tits in urban and forest habitats. Anim. Cogn. 20, 53–63. doi: 10.1007/s10071-016-1008-z
Quinn, J. L., Cole, E. F., Reed, T. E., and Morand-Ferron, J. (2016). Environmental and genetic determinants of innovativeness in a natural population of birds. Philos. Trans. R. Soc. B 371:1690. doi: 10.1098/rstb.2015.0184
Reader, S. M., and Laland, K. N. (2003). Animal Innovation: An Introduction: Oxford: Oxford University Press.
Récapet, C., Daniel, G., Taroni, J., Bize, P., and Doligez, B. (2016). Food supplementation mitigates dispersal-dependent differences in nest defence in a passerine bird. Biol. Lett. 12:20160097. doi: 10.1098/rsbl.2016.0097
Sanz, J. J., and Tinbergen, J. M. (1999). Energy expenditure, nestling age, and brood size: an experimental study of parental behavior in the great tit Parus major. Behav. Ecol. 10, 598–606. doi: 10.1093/beheco/10.5.598
Sol, D., Griffin, A. S., and Bartomeus, I. (2012). Consumer and motor innovation in the common myna: the role of motivation and emotional responses. Anim. Behav. 83, 179–188. doi: 10.1016/j.anbehav.2011.10.024
Sol, D., Griffin, A. S., Bartomeus, I., and Boyce, H. (2011). Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 6:19535. doi: 10.1371/journal.pone.0019535
Sternberg, R. J. (1985). Beyond IQ: A Triarchic Theory of Human Intelligence. Cambridge: Cambridge University Press.
Sutter, M., and Kawecki, T. J. (2009). Influence of learning on range expansion and adaptation to novel habitats. J. Evol. Biol. 22, 2201–2214. doi: 10.1111/j.1420-9101.2009.01836.x
Taylor, A. H., Hunt, G. R., and Gray, R. D. (2012). Context-dependent tool use in New Caledonian crows. Biol. Lett. 8, 205–207. doi: 10.1098/rsbl.2011.0782
Keywords: brood size manipulation, motivation, Parus major, problem-solving performance, provisioning rate, reproductive success
Citation: Cauchard L, Angers B, Boogert NJ, Lenarth M, Bize P and Doligez B (2017) An Experimental Test of a Causal Link between Problem-Solving Performance and Reproductive Success in Wild Great Tits. Front. Ecol. Evol. 5:107. doi: 10.3389/fevo.2017.00107
Received: 29 June 2017; Accepted: 25 August 2017;
Published: 08 September 2017.
Edited by:
Deseada Parejo, University of Extremadura, SpainReviewed by:
Wiebke Schuett, University of Hamburg, GermanyDaniela Campobello, University of Palermo, Italy
Copyright © 2017 Cauchard, Angers, Boogert, Lenarth, Bize and Doligez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laure Cauchard, laure.cauchard@umontreal.ca
†Shared senior authorship.