 Tae Chul Moon
Tae Chul Moon A. Dean Befus
A. Dean Befus Marianna Kulka
Marianna Kulka- 1Pulmonary Research Group, Department of Medicine, University of Alberta, Edmonton, AB, Canada
- 2National Institute for Nanotechnology, National Research Council, Edmonton, AB, Canada
Mast cells (MC) are widely distributed throughout the body and are common at mucosal surfaces, a major host–environment interface. MC are functionally and phenotypically heterogeneous depending on the microenvironment in which they mature. Although MC have been classically viewed as effector cells of IgE-mediated allergic diseases, they are also recognized as important in host defense, innate and acquired immunity, homeostatic responses, and immunoregulation. MC activation can induce release of pre-formed mediators such as histamine from their granules, as well as release of de novo synthesized lipid mediators, cytokines, and chemokines that play diverse roles, not only in allergic reactions but also in numerous physiological and pathophysiological responses. Indeed, MC release their mediators in a discriminating and chronological manner, depending upon the stimuli involved and their signaling cascades (e.g., IgE-mediated or Toll-like receptor-mediated). However, the precise mechanisms underlying differential mediator release in response to these stimuli are poorly known. This review summarizes our knowledge of MC mediators and will focus on what is known about the discriminatory release of these mediators dependent upon diverse stimuli, MC phenotypes, and species of origin, as well as on the intracellular synthesis, storage, and secretory processes involved.
Introduction
Human mast cells (MC) are often characterized by their ability to release a variety of important mediators with a diversity of biological activities (1). The regulated release of peptides, amines, lipids, and even some gases depends on several molecular pathways: a prominent one of which releases large dense core vesicles (granules) through regulated exocytosis (degranulation); other pathways depend upon de novo production of mediators and complex vesicle trafficking and recycling, including constitutive secretion, exosomal and endosomal pathways; and other secretory pathways that are not dependent upon vesicles or membrane-bound moieties [e.g., gases such as nitric oxide by diffusion (2), lipid mediators from lipid bodies]. Although research is providing important new insights, we understand remarkably little about how the mediators are sorted into these secretory pathways and differentially released (Tables 1 and 2). Unanswered questions include: how are these pathways similar/dissimilar; how are mediators sorted into various compartments (e.g., progranules, granules, lysosomes, secretory vesicles, and exosomes); which stimuli activate these secretory pathways, and which proteins are involved; how do MC selectively release different cargo given different stimuli?

Table 1. Mediators stored in human mast cell granules and their sorting mechanisms.
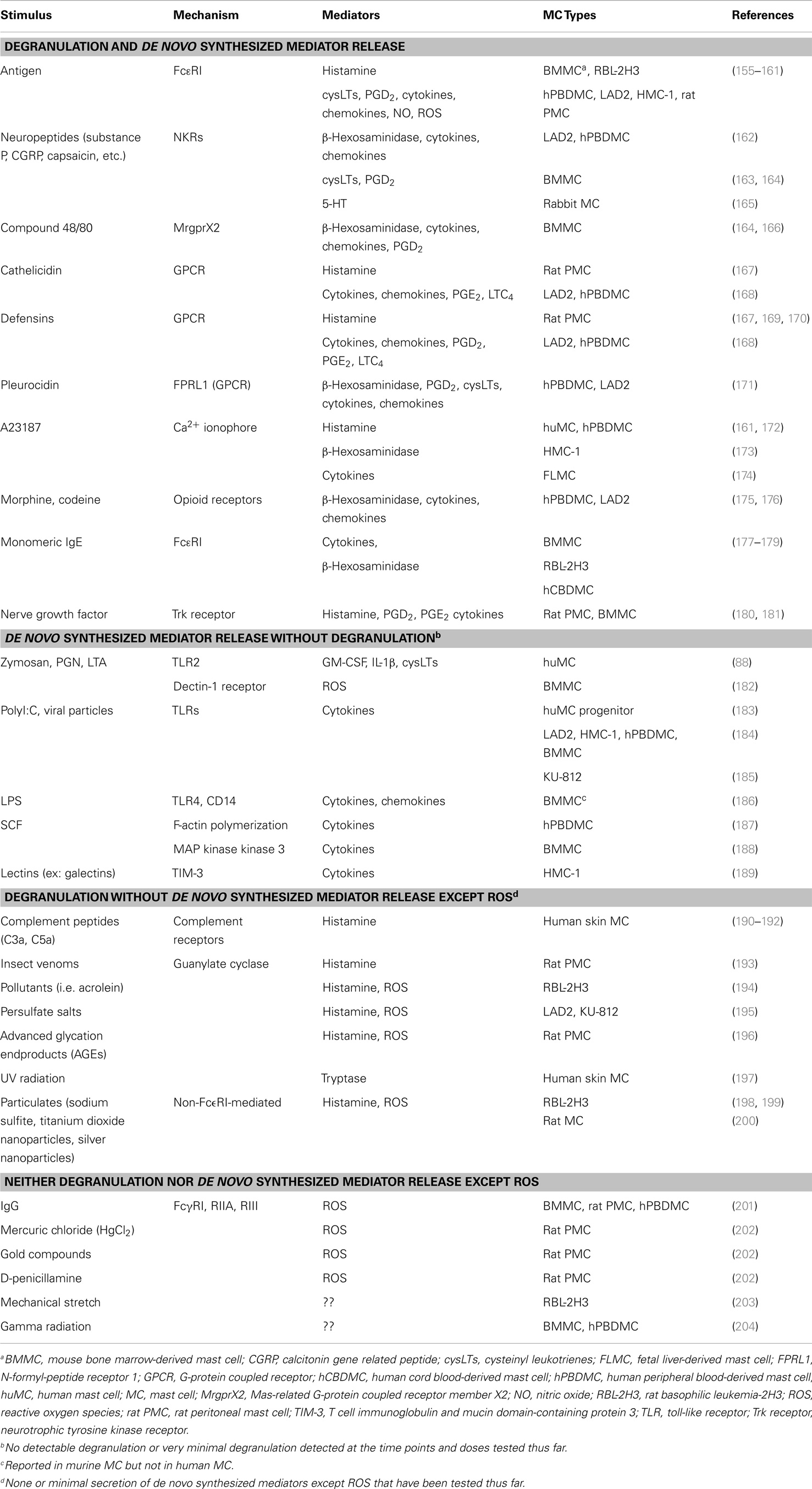
Table 2. Stimuli-selective mediator release from mast cells (some representative examples).
Constitutive exocytosis occurs in the absence of discernable stimuli for trafficking of secretory vesicles to the plasma membrane and can occur throughout the lifetime of a cell (3). Regulated exocytosis occurs after a clearly defined stimulus, either through changes in the extracellular environment [temperature (4, 5), pH (6), radiation (7), or osmolarity (8)] or ligation of a cell surface receptor (9). The pathways that control constitutive and regulated exocytosis have been extensively studied using powerful tools in high-resolution microscopy, molecular biology and animal model systems, and some of the molecules involved have been identified.
The terms degranulation, secretion, and exocytosis are often used interchangeably but have subtle variations in meaning. Degranulation refers to the loss of or release of granules and is most often associated with MC and basophils, both of which are characterized by their large intracellular granules. Secretion involves the release of a substance from one place of containment to another, i.e., from a cell to its extracellular environment or a gland to the skin’s surface. Excretion is the elimination of a waste material from a cell or organ. Exocytosis is a process of cellular secretion or excretion in which substances contained in vesicles are discharged from the cell by fusion of the vesicular membrane with the outer cell membrane (10–12). MC exhibit all forms of these release events but MC are perhaps best known for their rapid secretion of granules (degranulation) that contain large stores of pre-formed mediators (9).
This review identifies our current understanding of the biogenesis of various mediator compartments, and the mechanisms of sorting and release of mediators from these compartments (Figure 1). We present some new postulates about exocytosis that may be particularly relevant to the MC, a highly specialized secretory cell (13). We also refer the readers some excellent recent articles for more details on various aspects of this subject (9, 14–19).
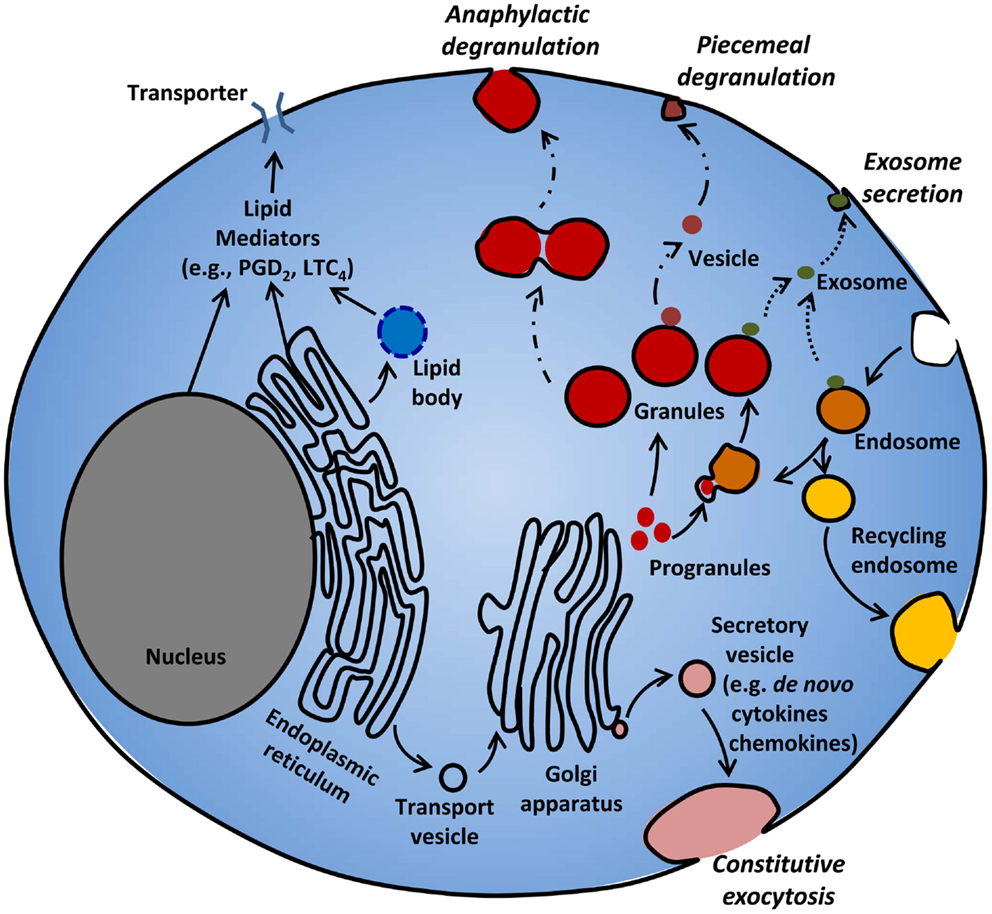
Figure 1. Mediator release from MC. MC release various mediators from different compartments following different stimuli. MC rapidly release pre-stored granule contents by piecemeal or anaphylactic degranulation. Immature progranules and mature granules can fuse with endosomes, and store lysosomal proteins. Some mediators can be released from granules and endosomes through exosomal secretion. Lipid mediators such as PGD2 and LTC4 are synthesized in lipid bodies, nuclear and ER membranes, and released through active transporters. De novo synthesized cytokines and chemokines packaged in secretory vesicles are released through constitutive exocytosis.
Pre-Stored Mediator Release from MC Granules
Mediators Stored in MC Granules
Mast cells are morphologically characterized by numerous, electron dense cytoplasmic granules which contain biogenic amines [histamine, serotonin] (20); several serine and other proteases {e.g., tryptase-α, -βI, -βII, -βIII, -γ [protease, serine S1 family member (PRSS) 31], -δ, chymase-1, cathepsin G, granzyme B, and carboxypeptidase A3} (21–31); lysosomal enzymes [β-glucuronidase (20), β-hexosaminidase (20), arylsulfatase (20)]; some cytokines [TNF (32), bFGF (33), IL-4 (34), and SCF (35)]; and proteoglycans [heparin (36, 37), chondroitin sulfates (36)] (Table 1). MC are able to overcome the large thermodynamic hurdle of storing high concentrations of these mediators in their granules by trapping them in an anionic gel matrix composed of chondroitin sulfates and heparin (38).
Subtypes of human MC are distinguished by the presence or absence of different serine proteases in their granules (i.e., tryptase+/chymase−: MCT, tryptase+/chymase+: MCTC, and tryptase−/chymase+: MCC). MC activation has typically been measured by monitoring the release of granule mediators (degranulation), with a particular focus on histamine, β-hexosaminidase, or tryptase (39, 40). Pre-stored mediator release through MC degranulation can be an early and rapid event following stimulation, resulting in the release of large portions of stored histamine within 15–90 s. This release of pre-formed mediators enables not only rapid anaphylactic reactions and allergic responses but also initiates recruitment of leukocytes to sites of pathogen invasion, activation of innate immune processes, and inflammatory responses (1). Other longer term responses associated with granule-derived mediators include wound healing and tissue remodeling processes through multiple communications with other cells (e.g., fibroblast proliferation and extracellular matrix production by histamine and MC proteases) (41).
MC Granule Heterogeneity and Biogenesis
Heterogeneity
Mast cells granules, also called secretory lysosomes, contain both lysosomal proteins such as acid hydrolases, e.g., β-hexosaminidase, as well as mediators such as histamine, and can secrete both together. MC also contain traditional lysosomes that can release enzymes such as β-hexosaminidase independently of histamine (42). Raposo et al. (43) distinguished three types of granules in mouse bone marrow-derived MC (BMMC) based on their contents of MHC class II, the lysosomal marker β-hexosaminidase, lysosomal membrane protein (LAMP)-1, LAMP-2 and mannose-6-phosphate receptors (M6PR), and the biogenic amine, serotonin: type I granules contain MHC class II, β-hexosaminidase, LAMP-1, LAMP-2, and M6PR but not serotonin (perhaps a classical lysosome); type II granules contain MHC class II, β-hexosaminidase, LAMP-1, LAMP-2, M6PR, and serotonin (perhaps a late secretory lysosome); type III granules contain β-hexosaminidase and serotonin but not MHC class II (Table 3) (42, 43). Baram et al. proposed a model wherein type II granules are generated by fusion of type I and type III granules, which contain lysosomal proteins and secretory amines, respectively (42). However, there has been little experimental follow-up of this postulate and there is evidence that MC have more diverse types of granules than depicted by this model (Figure 2). Indeed, the relationship between this classification of granules and observations that serotonin and cathepsin D vs. histamine and TNF exist in distinct granule populations (see below) in mouse MC is unclear (44). It is likely that MC granules are more heterogeneous than the three types shown above (Figure 2; Table 3) and that this heterogeneity may depend on the tissue of residence and the species, health status, and even age of the individual (1, 45).
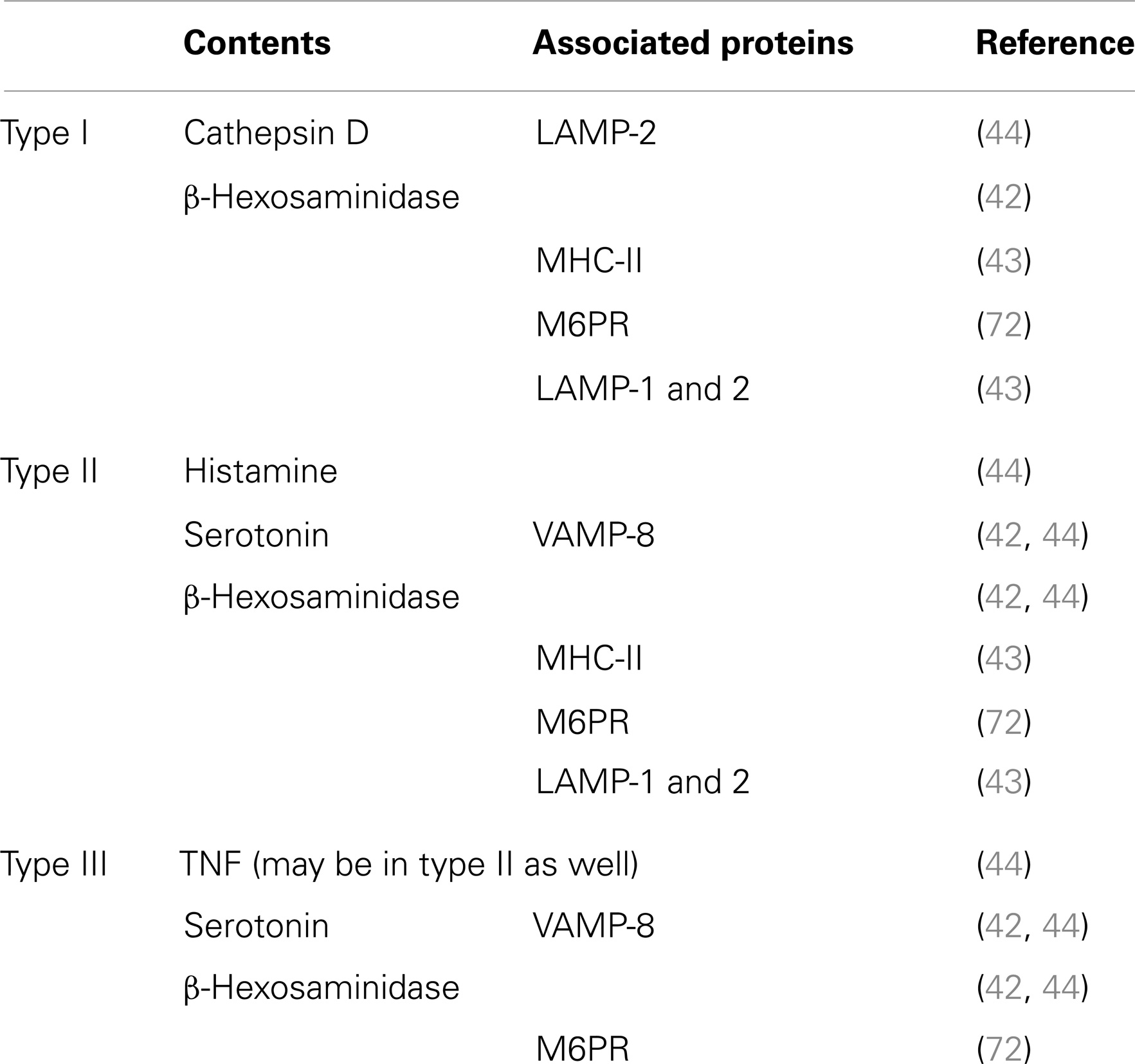
Table 3. Mast cell secretory granule subsets.
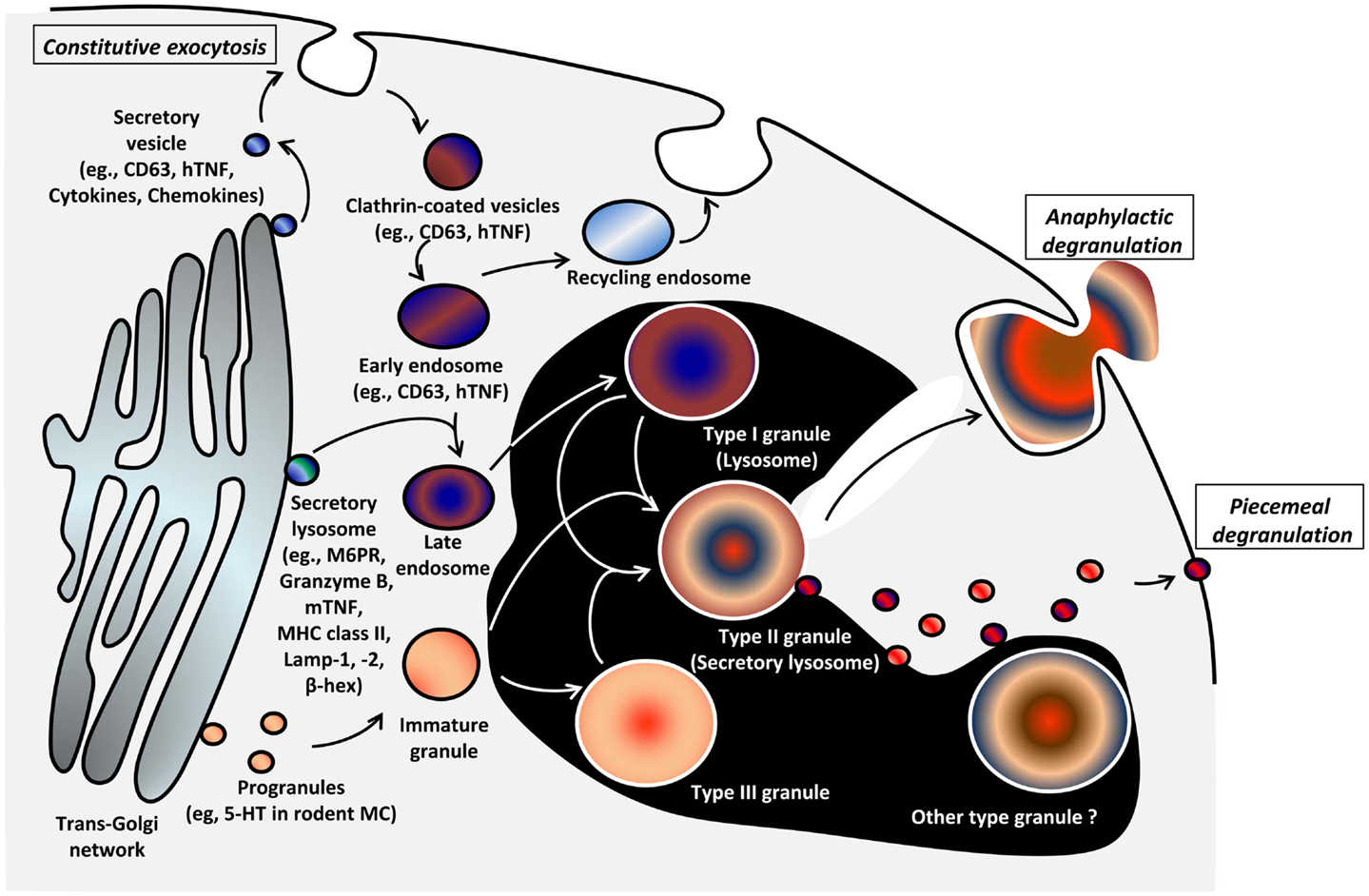
Figure 2. Model of genesis of MC secretory lysosomes (granules) and their heterogeneity/plasticity [adapted from Raposo et al. (43)]. Type I granules and type III granules are formed from lysosomal/endosomal pathway and by unit granule fusion from the trans-Golgi region, respectively. Secretory lysosomes that bud from trans-Golgi network contain MHC class II molecules, mannose-6-phosphate receptor (M6PR), and the lysosomal markers LAMP-1, -2, and β-hexosaminidase. It is postulated that post-endosomal, type II secretory lysosomes arise through the fusion of Type I and III granules. The relationship of this model to observations of heterogeneity of secretory lysosomes with regard to histamine or 5-HT content and VAMP-8 expression is unclear and there likely exists more granule heterogeneity/plasticity than three types (44). The mechanism of genesis of granule types is poorly understood (black area).
Biogenesis
The biogenesis of MC granules involves regulated fusion of what are called unit granules (small fusogenic granules) (46). These early unit granules buds from the trans-Golgi region and fuse to generate progranules in a region delimited by the outermost Golgi cisternae, rough endoplasmic reticulum (ER), and mature granules in the cytoplasm. The volumes of progranules are multiples of unit granules (i.e., volume of progranule created by three unit granules is three times unit granule volume). Progranules leave this zone as immature granules and become mature through a fusion process with other immature or mature granules. A process called “condensation” reduces the granule volume and organizes the contents, generating various sizes of mature granules (15). In addition to the homotypic fusion, which is postulated to form type III granules, immature granules or type III granules are also able to fuse to endosomes or lysosomes (Type I granules) in what might be the pathway that forms type II granules (secretory lysosomes) as Baram et al. proposed (see above and Figure 2) (42, 43). However, MC granules are likely more heterogeneous than the three types postulated, and our understanding of the later phase of granule biogenesis is thus depicted inside a black area in Figure 2.
Molecules involved in MC granule biogenesis
Several proteins involved in MC granule biogenesis and maturation have been identified (Table 4).
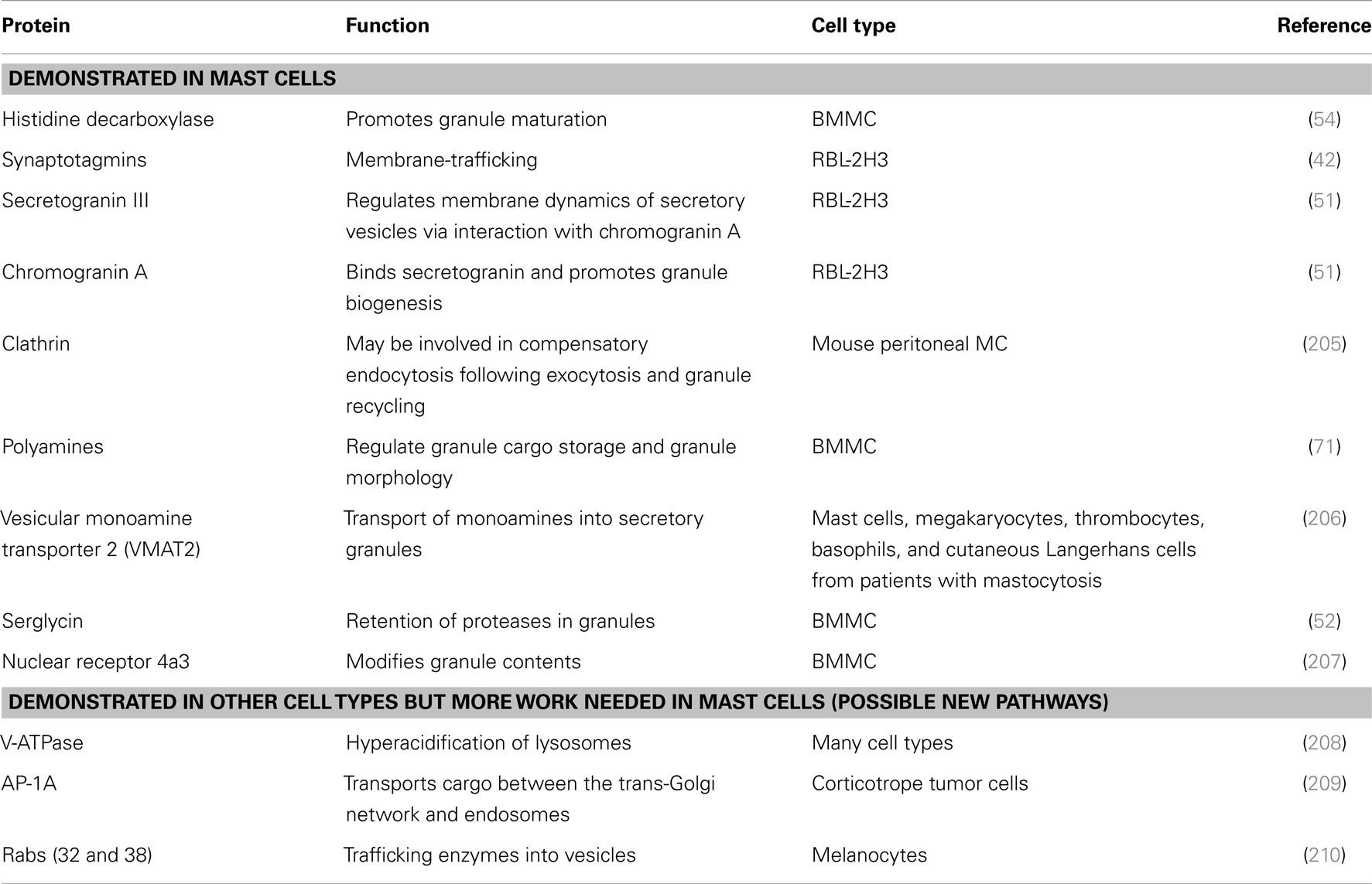
Table 4. Molecules that are (or may be) involved in mast cell granule biogenesis and homeostasis.
Rab GTPases. Rab3d and Rab5 play roles in the fusion of immature granules. MC from Rab3d knockout mice have granules that are larger than in MC from wild-type mice (47), while knockdown of endogenous Rab5 or expression of constitutively negative mutants of Rab5 significantly reduces the size of granules and increases their number (48). Moreover, Rab5 plays a role not only in homotypic granule fusion (type III granule biogenesis) but also in granule/endosome heterotypic fusion (type II granule biogenesis), and vesicle associated membrane protein (VAMP)-8 is involved in Rab5-mediated fusion of granules.
Lysosomal trafficking regulator. Chediak–Higashi syndrome, a mutation of the lysosomal trafficking regulator (LYST) causes the formation of giant granules in many cells including MC, and can be studied using the orthologous lyst-deficient beige (Lystbg/Lystbg) mouse. In MC and pancreatic acinar cells of beige mice, there is giant granule formation, presumably the result of disordered fusion of granules, suggesting Chediak–Higashi syndrome (CHS)/Lyst plays a role in controlling granule fusion (15, 49).
Synaptotagmins. The synaptotagmins (Syts) are membrane-trafficking proteins with at least 15 members in mammals. The RBL-2H3 MC line expresses Syt II, III, V, and IX (42) and RBL-2H3 treated with antisense to Syt III showed enlarged granule size, impairment in granule maturation, and formation and delivery of internalized transferrin to the perinuclear endocytic recycling compartment involved in a slow recycling pathway (50).
Granin family. The granin family of proteins, first described in neuroendocrine cells, are also important in MC granule biogenesis and maturation. Secretogranin III, for example, is present in MC granules, depleted during MC degranulation and over-expression of this protein causes an expansion of the secretory vesicle compartment (51). Although the biologic role of this family of proteins is not well understood, they are involved in cholesterol sequestration, interact with chaperones of granule proteins, serve as precursors of granule cargo and function as calcium buffering proteins, making them potentially intriguing players in the life history of MC granules. Is it possible that some of their many functions are necessary to control progranule fusion and thereby aid their maturation of secretory lysosomes/granules? Could these proteins somehow regulate the core components of the fusion machinery and thereby determine which progranules fuse together? Further experimentation is required in this area.
Histamine and proteoglycans. Some granule mediators themselves such as histamine and the core proteoglycans such as serglycin are also important components of the granule maturation process. For example, BMMC from serglycin knockout mice have functional secretory granules but they are defective in dense core formation (52). Furthermore, these serglycin−/− BMMC are resistant to apoptosis associated with reduced release of proteases and defective caspase-3 activation (53). In addition, the lack of histamine or the enzymes that control its synthesis significantly alters the morphology and contents of granules. Peritoneal MC from histidine carboxylase knockout mice show abnormal granule morphology and contain fewer proteases and heparin (54). This is mainly due to the down-regulation of genes encoding granule proteases and enzymes involved in heparin biosynthesis. Interestingly, agonists of the H4 histamine receptor and exogenous application of histamine restored granule maturation. Therefore, histamine likely influences early steps in granule maturation but has little role in maintaining the integrity of fully formed mature granules since depletion of histamine from mature MC had no obvious effect on granule structure. It would be interesting to examine the effect of histamine depletion on immature MC (i.e., from CD34+ progenitors) as they progress through the MC differentiation process.
Adaptor-protein family. Components of our models of MC granule biogenesis can be extrapolated from other cells with complex secretory granule processes, such as pituitary lactotropes, pancreatic β-cells, and transfected tumor cells. In these cells, progranules in the trans cisterna of the Golgi are covered with clathrin coats, which contain the adaptor-protein (AP) family of proteins that can bind cytosolic tails of transmembrane protein cargo, facilitating their entry into budding vesicles (55). Immature secretory granules are not responsive to secretagogues and it appears that one of the essential roles of the AP proteins is to facilitate their maturation (56). Could APs perform a similar function in MC? Clearly, additional studies are needed to further understand the genesis, heterogeneity, and plasticity of MC granules and uncover potential therapeutic opportunities within such knowledge; a frontier.
Mechanisms of Sorting and Storage of Pre-Formed Mediators in MC Granules
The most unique and igneous feature of MC granules is their ability to store large concentrations of mediators in a small space for long periods. In theory, collecting a high concentration of such highly charged mediators in a membrane-enclosed space would require a large amount of osmotic work and create a thermodynamic disadvantage. However, MC trap the mediators in an anionic gel matrix composed mainly of heparin and chondroitin sulfate, which confers a huge thermodynamic advantage (38, 57). Upon cell activation, the polymer gel phase undergoes a transition and swells to release the mediators (58). This efficient solution is not-surprisingly conserved among living organisms and even phytoplankton use a similar packaging and degranulation process (59). Current understanding of sorting and storage mechanisms of pre-formed mediators are reviewed below and in Table 1 but it is still poorly understood.
Proteoglycans
Proteoglycans, a core component of MC granules are heavily glycosylated proteins, consisting of a core protein and glycosaminoglycan side chains that are covalently attached to the core through glycosidic bonds. In MC, serglycin is a dominant core protein, and heparin and chondroitin sulfate are dominant glycosaminoglycans that can be used to distinguish some MC subpopulations (1). Sulfation of glycosaminoglycans imparts a negative charge on the proteoglycan, which is an important mechanism that helps to retain proteases and biogenic amines in MC granules (60). An ion exchange mechanism with the charged glycosaminoglycans is thought to be significant in mediator release from granules (61, 62). Moreover, as outlined above, proteoglycans are important in granule composition and maturation. However, details of the sorting mechanism of proteoglycans and other mediators into MC granules are poorly understood. Studies with rat pancreatic acinar cells provide clues and suggest that glycosaminoglycan side chains are necessary for proteoglycan sorting into granules, as deletion of serine–glycine repeat region of the serglycin core protein and treatment with p-nitrophenyl-β-D-xylopyranoside, an alternate substrate for glycosaminoglycan side chain attachment, prevented sorting into granules and lead to accumulation of proteoglycans in Golgi (63).
MC proteases
Mast cell proteases are synthesized in the ER, modified in the Golgi complex, and sorted into progranules that bud from the trans-Golgi. Retention of MC proteases in granules depends on the serglycin proteoglycan in murine MC, as absence of mouse MC protease (mMCP)-5 (chymase) and carboxypeptidase, and reduction of mMCP-6 (tryptase) occurs in the granules of MC from serglycin deficient mice despite normal expression of protease mRNA and granule formation (52). Although limited information is available with other kinds of proteases, serglycin proteoglycan is involved in certain protease retention in MC granules. However, the mechanisms underlying the trafficking of MC proteases into the granule need to be elucidated and confirmed in human MC, although the storage of heparin and chondroitin sulfate proteoglycan in human MC granules has been shown (36).
Biogenic amines
Histamine and serotonin are biogenic amines stored in MC granules. There is evidence that transport of biogenic amines from cytosol into the MC granules occurs in a vesicular monoamine transporter 2 (VMAT2)-dependent manner (64, 65). Moreover, retention of biogenic amines and release from the granule is serglycin proteoglycan dependent (66). Whether retention of both MC proteases and biogenic amines are directly dependent on serglycin proteoglycan or involve only the glycosaminoglycan side chains, e.g., heparin, with their electrostatic charges (67–70), needs to be elucidated and extended to studies with human MC. Some polyamines (such as putrescine, spermidine, and spermine) are required for granule homeostasis and possibly aid in the native conformation and packaging of other granule molecules such as histamines and proteases (71).
Lysosomal enzymes
Many lysosomal enzymes (Table 1) are found in MC granules but the detail mechanisms of their sorting, trafficking, storage, and secretion are poorly understood. It is postulated that they are transported into type II MC granules when granules and endosomes fuse (Figure 2). Clearly, lysosomal enzymes can be found in both type II granules, as well as classical lysosomes (type I granules), and secreted from both compartments. Using MC from serglycin knockout mice, it was shown that storage and release of β-hexosaminidase is independent of serglycin (52).
Cytokines
Among the large number of cytokines and chemokines released after MC activation, TNF (32, 72), bFGF (33), IL-4 (34), and SCF (35) are known to be pre-stored in MC granules, and can be released by regulated exocytosis, as well as synthesized following MC activation and released through constitutive exocytosis (Figure 1) (73, 74). Many other cytokines and chemokines appear not to be stored [e.g., GM-CSF (75)], but are newly synthesized following MC activation and are secreted by constitutive exocytosis over the course of several hours/days (discussed below) (9, 76). For storage in the granule, there appears to be a different trafficking mechanism for TNF in rodent and human MC. In rodent MC, sorting of TNF from ER to granules occurs via a brefeldin A- and monensin-sensitive route, utilizing a M6PR-dependent pathway and N-linked glycosylation of asparagine at N86 (Figure 2) (72). By contrast in human MC, TNF does not utilize this pathway as the N-linked glycosylation motif NSS of rodent TNF is replaced by the RTP motif (32). By transfecting and chasing fluorescence-tagged TNF into human MC lines (HMC-1 and LAD2), Olszewski et al. showed that in human MC, TNF traffics to the plasma membrane transiently, but then is stored in MC granules by endocytosis (Figure 2) (32). These observations emphasize that evidence acquired from studies of rodent MC must be validated for human MC, as has been shown for several other examples of species differences among MC (1). Apart from TNF, the mechanisms of trafficking of bFGF, IL-4, and SCF, another cytokines stored in human MC granules have not been studied, and we still do not fully understand how these cytokines are sorted in granules and which secretion pathway(s) initiates their release. Based on the binding affinity of bFGF for heparin (77), the retention mechanism of bFGF in the granule is likely to be heparin dependent, although this needs to be confirmed. Immunohistochemistry has shown that IL-4 but not IL-5 are stored in MC secretory granules in the lung parenchyma and nasal mucosa of patients with active allergic rhinitis (34). Although the amount of IL-4 in the granules increases after FcεRI-mediated activation, it is unclear whether the majority of IL-4 released extracellularly is due to degranulation or constitutive exocytosis.
Mechanisms of Secretion of Pre-Formed Mediators from MC Granules
Two types of degranulation have been described for MC: piecemeal degranulation (PMD) and anaphylactic degranulation (AND) (Figures 1 and 2). Both PMD and AND occur in vivo, ex vivo, and in vitro in MC in human (78–82), mouse (83), and rat (84). PMD is selective release of portions of the granule contents, without granule-to-granule and/or granule-to-plasma membrane fusions. PMD in MC has been identified in numerous settings, ranging from chronic psychosocial stress (85) to estradiol (86), CCL2 (87) and TLR stimulation (88), and interactions with CD4+/CD25+ regulatory T cells (89). The granule morphology is relatively well retained following PMD, although ultrastructural changes are evident (79). It has been proposed that the mechanism of PMD involves the budding of vesicles containing selected mediators from granules and their transport to the plasma membrane, fusion, and mediator release (Figures 1 and 2) (90). Little is known about the molecular machinery involved in these processes.
In contrast to PMD, AND is the explosive release of granule contents or entire granules to the outside of cells after granule-to-granule and/or granule-to-plasma membrane fusions (Figures 1 and 2). Ultrastructural studies show that AND starts with granule swelling and matrix alteration after appropriate stimulation (e.g., FcεRI-crosslinking). Granule-to-granule membrane fusions, degranulation channel formation, and pore formation occur, followed by granule matrix extrusion (81). Granule-to-granule and/or granule-to-plasma membrane fusions in AND are mediated by soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNAREs) (91). In human intestinal MC, protein expression of soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP)-23, syntaxin (STX)-1B, STX-2, STX-3, STX-4, VAMP-2, VAMP-3, VAMP-7, VAMP-8, and STX-6 have been reported (92). However, only VAMP-7 and VAMP-8 were found to translocate to the plasma membrane and interact with SNAP-23 or STX-4 upon activation. Moreover, inhibition of SNAP-23, STX-4, VAMP-7, or VAMP-8, but not VAMP-2 or VAMP-3, reduced histamine release mediated by FcεRI-crosslinking (92). Therefore, VAMP-7, VAMP-8, SNAP-23, and STX-4 are important SNARE molecules in human intestinal MC granule fusion and exocytosis.
Molecules involved in MC degranulation
Several proteins involved in MC degranulation were listed in Table 5.

Table 5. Molecules involved in mast cell degranulation.
Mammalian uncoordinated-18 proteins. The functions of SNARE proteins are regulated by several accessory proteins, but our knowledge is incomplete and at least in part, the information is controversial (Table 5). Munc 13-4 was shown to be a target of Rab27a and Munc 13-4-transduced RBL-2H3 release more histamine compare to the mock-transduced cells after IgE/Ag stimulation (93). In rodent MC, mammalian uncoordinated-18 (Munc-18)-2, located in the granule membrane, interacts with STX-3 and plays a role in granule-to-granule as well as granule-to-plasma membrane fusion (94, 95) whereas, Munc-18-3, located in the plasma membrane, also interacts with STX-4 (94). Following activation of RBL-2H3, another protein, complexin II translocates from the cytosol to the plasma membrane and interacts with a SNARE complex. Although translocation of complexin II to the plasma membrane did not induce membrane fusion, the reduction of degranulation after knockdown of this protein suggests that complexin II is a positive regulator of MC degranulation (96). The fusion of the SNAREs with the plasma membrane has been examined using transmission and freeze-fracture electron microscopy and biophysical modeling. About 30–60 s after activation, unetchable circular impressions about 80–100 nm in diameter were found on the E face (intracellular face) of the plasma membrane (97). These impressions are not permanent but are postulated to form the fusion sites for the granules directly preceding degranulation. Under certain conditions, these fusion sites can form rosettes and the coupling of this structure with the plasma membrane may then form a cup-shaped structure called a porosome (98). Due to the nanometer size of these SNARE docking sites, the MC degranulation complex has been called a nano-machine (97).
Vesicle associated membrane proteins. A study using BMMC from VAMP-8 deficient mice showed reduced serotonin, cathepsin D, and β-hexosaminidase release, but normal histamine and TNF release following IgE-mediated or PMA/ionomycin stimulation (44). By contrast, transfection of VAMP-8 in RBL-2H-3 did not affect the calcium ionophore/12-O-tetradecanoyl-13-acetate- or IgE-mediated release of fluorescent-labeled neuropeptide Y, which is stored in the same granules as serotonin and β-hexosaminidase (48). These conflicting data may be the result of different experimental systems (former using deficient mouse of VAMP-8 and latter using over-expression), or the fact that the latter study examined mediator release indirectly with fluorescent-labeled neuropeptide Y. Although further study is required, VAMP-8 is likely involved in granule biogenesis and degranulation of subsets of MC granules, which contain serotonin, cathepsin D, and β-hexosaminidase. Moreover, this suggests that there is heterogeneity of MC granules (Figure 2), and that distinct mechanisms are involved in mediator release between subsets of granules.
Synaptotagmins. Synaptotagmin (Syt) II depresses Ca2+-triggered secretion of β-hexosaminidase and MHC class II release (42), but increases cathepsin D release in RBL-2H3 and mouse BMMC (99). Moreover, in Syt II knockout mice there is a marked deficiency in degranulation and an impaired passive cutaneous anaphylaxis response (62). In RBL-2H3, Syt IX can regulate protein export from the endocytic recycling compartment to the plasma membrane and play a role in sorting proteins of secretory granules (100). Much remains to be learned about these proteins and MC function.
Rab GTPases. In addition to their role in granulogenesis mentioned above, the Rab family of GTPases is also involved in MC degranulation. Over-expression of Rab3a in RBL-2H3 showed no (101), or inhibitory effects (102) on FcεRI-mediated β-hexosaminidase release, while over-expression of Rab3d demonstrated that it translocates from the granule to the plasma membrane (103), and inhibits degranulation (101). Recently, it was established that Rab27a is located in histamine-containing granules in RBL-2H3 and that over-expression of constitutively active Rab27a reduced FcεRI-mediated histamine secretion (93). Munc 13-4 was found to be a target of Rab27a, and the Rab27a-Munc 13-4 complex was required for docking of granules to the plasma membrane and release of granule contents in RBL-2H3 cells (104). Moreover, Rab27a regulates cortical actin stability with its effectors melanophilin (Mlph) and myosin, as well as Rab27a/b/Munc13-4-dependent granule exocytosis (105). Rab27b knockout mice exhibited reduced passive cutaneous anaphylaxis and defects in FcεRI-mediated β-hexosaminidase release from BMMC (106). Although Rab5 is involved in MC granule biogenesis (see above), a transfection study showed that Rab5 is not involved in granule mediator release (48).
Rho GTPases. Among Rho GTPases, Rac and Cdc42 play a positive role in RBL-2H3 degranulation by regulating IP3 production, upstream of Ca2+ influx and interacting with PLCγ1 (107, 108). More recently, the roles of Rac1 and Rac2, which have a 92% sequence identity, in MC degranulation were dissected using knockout mice (109). In BMMC from Rac2 knockout mice, FcεRI-, but not Ca2+ ionophore-mediated β-hexosaminidase release was defective because of a decrease in Ca2+ flux without changing F-actin remodeling and membrane ruffling, which are regulated by Rac1 (109).
Others. The cytoskeleton and the microtubule network is an essential component of the degranulation process in MC. Proteins such as DOCK5 (110), MARCKS (111), and myosin VI (112) regulate the progress of secretory granules through the cytoskeletal network and allow them to dock with the plasma membrane. When these proteins are disrupted, the degranulation process does not occur normally. Some of these pathways remain unexplored in MC, yet we know that some microtubule events facilitate granule fusion and are essential to degranulation. For example, myosin Va forms a complex with Rab27a and Mlph thereby regulating cortical F-actin stability upstream of Rab27a/b/Munc13-4-dependent granule exocytosis (105).
Although calcium flux is unequivocally an essential feature of the degranulation process [recently reviewed by Fahrner et al. (113) and Ashmole and Bradding (114)], other ion exchange complexes have also been implicated, such as potassium and chloride channels that facilitate and regulate Ca2+ signaling. Certainly, disruption of membrane potentials using cationic liposomes can impair Ca2+ flux and suppress the function of SNAP-23 and STX-4 (115). It has been suggested that inhibitors of these ion exchange pathways may be useful in the treatment of inflammatory diseases that are mediated by MC activation and degranulation (116).
Calcium flux is essential for degranulation but its role may be more extensive than first postulated and some theories have questioned the long-standing belief that granule membranes must fuse with the plasma membrane to facilitate exocytosis. There are new theories of exocytosis that have not yet been examined in MC, including the theory of porocytosis, or secretion without membrane fusion, in which Ca2+ ions form salt bridges among adjacent lipid molecules through which mediators would move according to mass action (13). This quantal secretion theory has been postulated to be important in neuromuscular junctions and the central nervous system and it offers an intriguing process for mechanisms underlying constitutive exocytosis. However, this mathematical model has not been validated experimentally in neurons or other secretory cells. Autophagy, an evolutionarily conserved bulk degradation system that facilitates the clearance of intracellular molecules, has also been shown to be an important regulator of MC exocytosis. A recent study by Ushio et al. has shown that proteins that normally control autophagy may also facilitate the fusion of small secretory vesicles and facilitate their fusion with the plasma membrane (117). In fact, many new regulatory pathways have been connected to degranulation and exocytosis. However, we need more sophisticated model system to validate these new theories. Given that most molecules known to be involved in MC degranulation have been studied in rodent models, their roles in human MC must be examined and potential distinctions between MC phenotypes in different species should be investigated.
Secretion of Exosomes
Exosomes are membrane-bound vesicles of ~30–100 nm that appear to bud from the internal surface of multivesicular bodies in the endosomal compartment of many cell types including MC (Figure 1) (118). They are important in cell–cell communications and a breadth of physiological and pathophysiological responses, notably antigen presentation and host defenses. The contents of exosomes include a richness of lipids such as ceramide, cholesterol, phosphatidylserine, and sphingomyelin; a great diversity of proteins (200–400) including MHC class II, phospholipases, heat shock proteins, co-stimulatory molecules (CD40, CD40L, and CD86), adhesion molecules, kinases, tetraspanins, cytoskeletal proteins, chaperones, aldolase A, TNF, FcεRI chains, processed peptides from antigens; and a plethora of mRNA (>1800) and microRNA (>100) species (119–122). Exosomes transfer diverse cargo and functional capacity among cells; for example, mRNAs and microRNAs in MC exosomes can be transferred between human and mouse MC or other cells and control gene expression in recipient cells (119, 120). Carroll-Portillo et al. emphasized the potential functional significance of IgE-antigen and antigenic peptide–MHC class II MC–T cell–Dendritic cell interactions, as well as MC uptake of antigen-crosslinked receptor (16).
Mast cell exosomes can be secreted by both constitutive or regulated exocytosis (Figure 1) (16, 43) and the composition of protein constituents differs depending upon which pathway is employed (16). Knowledge is advancing rapidly about the molecular bases of exosome biogenesis, regulation of exosome loading, and the secretory pathways involved. The reader is referred to recent reviews of this intriguing and rapidly evolving subject (118, 123).
Cytokine/Chemokine Secretion from Compartments Other than Secretory Lysosomes
In addition to regulated secretion of a limited repertoire of cytokines from stores in granules (above) through pathways of PMD or AND, MC secrete a diversity of cytokines and chemokines by other pathways [diagrammed by Lorentz et al. (14) in their Figure 3], including: constitutive exocytosis, better known in macrophages [e.g., Ref. (3, 9), etc.], and exosomal secretion (Figure 1). Frank et al. showed that involvement of SNAREs in chemokine release of human intestinal MC using their specific neutralizing antibodies. They showed that CXCL8, CCL2, CCL3, and CCL4 release after IgE/α-IgE stimulation were abrogated by inhibition of STX-3 or SNAP-23, but not by inhibition of STX-2 or VAMP-3. Moreover, inhibition of different SNARE subsets selectively reduce chemokine release (i.e., STX-4 or VAMP-8 inhibition selectively reduce CXCL8, and STX-6 inhibition reduces CXCL8 and CCL2) (124). However, other study using VAMP-8−/− BMMC showed that VAMP-8 does not affect cytokine/chemokine secretion (125). Therefore, there has been insufficient exploration of the molecular elements involved in these pathways in MC; careful analyses are needed in comparison to other cells such as macrophages, as well as among various MC subsets and between human and other MC.
Lipid Mediator Release
Activated MC release an abundance of arachidonic acid metabolites, notably leukotriene (LT) C4, prostaglandin (PG) D2, and platelet activating factor (PAF) (126–128). These lipid mediators have bronchoconstricting and vasoactive properties, but also participate in host defense, inflammation, and allergic diseases through diverse activities such as effector cell trafficking, antigen presentation, immune cell activation, and fibrosis (129–131). Eicosanoids, lipid mediators derived from arachidonic acid, are de novo synthesized and released immediately from activated cells rather than stored, and exert autocrine and paracrine functions. However, recent observations of intracellular localization of the PGD2 receptor, DP2/CRTh2, suggest intracrine functions of this mediator (132, 133). Eicosanoid synthesis can occur at several sites in the cell, including the ER (134), nuclear membrane (134, 135), phagosomes (136), and cytoplasmic lipid bodies (Figure 1) (81, 82, 137, 138), and the site of synthesis likely depends on the cell type, as well as the nature of the stimulation [see review in Ref. (139)]. Whether the enzymes and other proteins needed for eicosanoid synthesis are de novo synthesized following cell activation, or are dispersed in the cell and then translocated to a site of synthesis following activation, are under debate. In either case, how these proteins are targeted to a specific intracellular compartment of eicosanoid synthesis is unknown.
Lipid Bodies
Lipid bodies also referred to as lipid droplets are osmiophilic organelles surrounded by a monolayer of phospholipids and containing lipids with a unique fatty acid composition. Lipid bodies are composed of a core rich in neutral lipids and a variety of proteins depending upon the cell type and the conditions of stimulation (Figure 3) (137, 138, 140). These organelles are one of the major sites of eicosanoid generation. They are inducible during inflammatory processes and increase in size and number in several types of leukocytes, including MC (141–144). The numbers of lipid bodies in the cytoplasm of MC differ among MC phenotypes. For example, lipid bodies are rare in human skin MC in comparison to numbers in lung and gut MC (81, 82). In addition to eicosanoid synthesis, a few cytokines and chemokines are also found in lipid bodies of eosinophils (TNF), neutrophils (TNF), macrophages (TNF), and MC (TNF and bFGF) (33, 142). Much remains to be elucidated about the kinds of cytokines stored in lipid bodies, functions of these cytokines, and the mechanisms of their release.
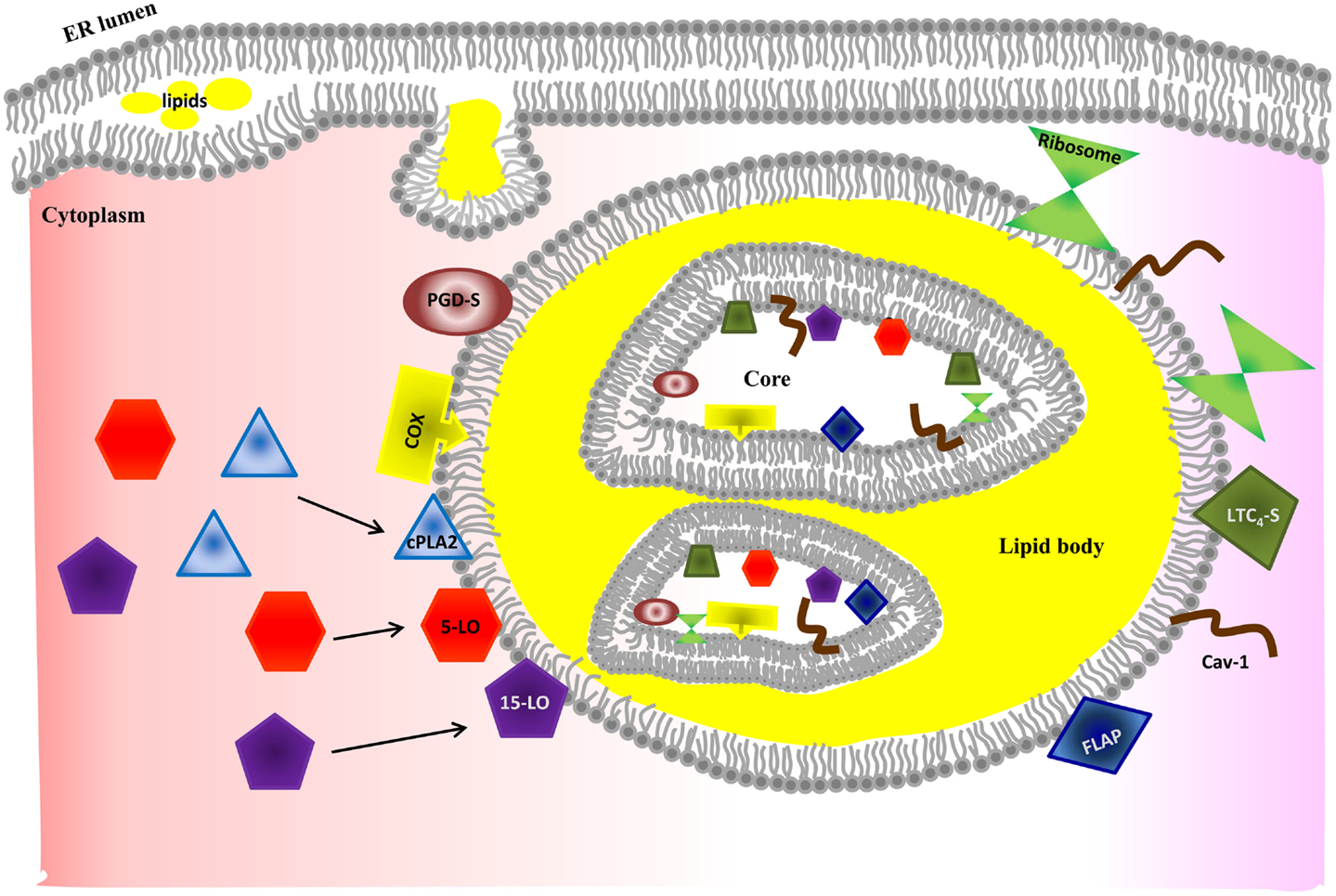
Figure 3. Model of lipid body biogenesis and structure. Neutral lipids synthesized in the ER accumulate between bilayer of ER membrane and bud off as a lipid body. Lipid bodies have phospholipid monolayer on their outside. However, lipid bodies contain a bilayer core structure inside, which provides a hydrophilic area. The bilayer core can be created by incorporation of multiple loops of ER membrane and explains how ER membrane proteins (e.g., caveolin-1 and ribosome) are incorporated into lipid bodies. However, the exact mechanism of formation of the bilayer structure is poorly understood. Enzymes required for eicosanoid production have been found in both outer membrane and core of lipid bodies. Increased intracellular Ca2+ after MC stimulation induces activation and translocation of cPLA2, 5-LO, and 15-LO to the lipid body membrane for eicosanoid synthesis. Further studies are required to unveil how MC control synthesis and secretion of arachidonic acid metabolites in a stimulus-specific fashion.
Lipid Body Biogenesis
Although the details of the biogenesis of lipid bodies in MC are poorly understood, they bud from the ER membrane where neutral lipids are synthesized (Figure 3) (145). Lipid bodies can also increase in size by fusion and this process occurs in a rapid regulated way (146).
Mechanisms of Synthesis and Secretion of Lipid Mediators
Eicosanoids are made by oxidation of 20-carbon fatty acids. After activation, MC synthesize PGD2 and LTC4 from sequential enzymatic reactions of arachidonic acid metabolism of cyclooxygenase and lipoxygenase pathways, respectively. PAF also can be synthesized from lysophosphatidylcholine by PAF-acetyltransferase. Both arachidonic acid and lysophosphatidylcholine can be generated from membrane phospholipid (1-O-alkyl-2-arachidonoyl-sn-glycero-3-phosphocholine) by phospholipase A2. Recently, studies on pro-resolving lipid mediators, such as resolvins, protectins, and maresins from eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) have been advanced in macrophages and our understanding of their physiological, pathophysiological functions in resolution of inflammation are improved, but little is known their synthesis in MC (147).
Because eicosanoids synthesized intracellularly are negatively charged at physiological pH (148), they diffuse poorly across membranes. Thus, their secretion involves active transport through either organic anion transporters of the ATP-binding cassette type C family, also known as multidrug resistance-related proteins (149–152), or organic anion transporters of the solute carrier superfamily (153, 154). However, little is known of these mechanisms in MC and further study is needed to identify the mechanisms and their regulation.
Conclusion
Over several decades, there have been significant advances in our knowledge of MC biology that have transformed our understanding of this multifaceted immune cell from an effector cell in allergic inflammation to a major player in innate and acquired immunity. Because of the pivotal role of MC in allergic and other inflammatory reactions, therapeutic strategies to disrupt the action of MC mediators have been developed and used widely [e.g., anti-histamines, anti-IgE (omalizumab), and Cys-LT1 receptor antagonist (montelukast)]. However, there remain large gaps in our knowledge about intracellular trafficking of MC mediators, particularly in the selective mechanisms of storage and secretion that are often dependent on the specific stimuli involved. Our knowledge of the biogenesis of MC cargos, the heterogeneity of granules, and the molecules involved in trafficking pathways of mediator secretion are rapidly evolving and likely to be a productive in helping to unravel the complexities of MC biology. A greater understanding of mediator specific trafficking pathways will provide opportunities to develop novel therapeutic targets for the treatment of increasingly wide-spread allergic and other inflammatory diseases.
Author Contributions
Tae Chul Moon, A. Dean Befus, and Marianna Kulka equally contributed to writing this paper.
Conflict of Interest Statement
The Guest Associate Editor Paige Lacy declares that, despite having collaborated with authors Tae Chul Moon and A. Dean Befus, the review process was handled objectively and no conflict of interest exists. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the Allergy Genes and Environment (AllerGen) Network of Centres of Excellence, Canada, the Canadian Institutes of Health Research (CIHR), the National Sciences and Engineering Research Council (NSERC), and the National Institute for Nanotechnology (NINT), Canada.
Abbreviations
AND, anaphylactic degranulation; BMMC, bone marrow-derived mast cell; ER, endoplasmic reticulum; LAMP, lysosomal membrane protein; MC, mast cell(s); M6PR, mannose-6-phosphate receptor; PMD, piecemeal degranulation; SNAP, soluble N-ethylmaleimide-sensitive factor attachment protein; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; STX, syntaxin; Syt, synaptotagmin; VAMP, vesicle associated membrane protein.
References
1. Moon TC, St Laurent CD, Morris KE, Marcet C, Yoshimura T, Sekar Y, et al. Advances in mast cell biology: new understanding of heterogeneity and function. Mucosal Immunol (2010) 3(2):111–28. doi: 10.1038/mi.2009.136
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
2. Lancaster JR Jr. A tutorial on the diffusibility and reactivity of free nitric oxide. Nitric Oxide (1997) 1(1):18–30. doi:10.1006/niox.1996.0112
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
3. Lacy P, Stow JL. Cytokine release from innate immune cells: association with diverse membrane trafficking pathways. Blood (2011) 118(1):9–18. doi:10.1182/blood-2010-08-265892
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
4. Mori S, Saino T, Satoh Y. Effect of low temperatures on compound 48/80-induced intracellular Ca2+ changes and exocytosis of rat peritoneal mast cells. Arch Histol Cytol (2000) 63(3):261–70. doi:10.1679/aohc.63.261
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
5. Schick B, Austen KF. Rat serosal mast cell degranulation mediated by chymase, an endogenous secretory granule protease: active site-dependent initiation at 1 degree C. J Immunol (1986) 136(10):3812–8.
6. Alfonso A, Cabado AG, Vieytes MR, Botana LM. Calcium-pH crosstalks in rat mast cells: cytosolic alkalinization, but not intracellular calcium release, is a sufficient signal for degranulation. Br J Pharmacol (2000) 130(8):1809–16. doi:10.1038/sj.bjp.0703490
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
7. Muller K, Meineke V. Radiation-induced mast cell mediators differentially modulate chemokine release from dermal fibroblasts. J Dermatol Sci (2011) 61(3):199–205. doi:10.1016/j.jdermsci.2011.01.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
8. Gulliksson M, Palmberg L, Nilsson G, Ahlstedt S, Kumlin M. Release of prostaglandin D2 and leukotriene C4 in response to hyperosmolar stimulation of mast cells. Allergy (2006) 61(12):1473–9. doi:10.1111/j.1398-9995.2006.01213.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
9. Wernersson S, Pejler G. Mast cell secretory granules: armed for battle. Nat Rev Immunol (2014) 14(7):478–94. doi:10.1038/nri3690
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
10. Chicoine E, Esteve PO, Robledo O, Van Themsche C, Potworowski EF, St-Pierre Y. Evidence for the role of promoter methylation in the regulation of MMP-9 gene expression. Biochem Biophys Res Commun (2002) 297(4):765–72. doi:10.1016/S0006-291X(02)02283-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
11. Burgoyne RD, Morgan A. Secretory granule exocytosis. Physiol Rev (2003) 83(2):581–632. doi:10.1152/physrev.00031.2002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
13. Silver RB, Pappas GD. Secretion without membrane fusion: porocytosis. Anat Rec B New Anat (2005) 282(1):18–37. doi:10.1002/ar.b.20050
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
14. Lorentz A, Baumann A, Vitte J, Blank U. The SNARE machinery in mast cell secretion. Front Immunol (2012) 3:143. doi:10.3389/fimmu.2012.00143
15. Hammel I, Lagunoff D, Galli SJ. Regulation of secretory granule size by the precise generation and fusion of unit granules. J Cell Mol Med (2010) 14(7):1904–16. doi:10.1111/j.1582-4934.2010.01071.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
16. Carroll-Portillo A, Surviladze Z, Cambi A, Lidke DS, Wilson BS. Mast cell synapses and exosomes: membrane contacts for information exchange. Front Immunol (2012) 3:46. doi:10.3389/fimmu.2012.00046
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
17. Galli SJ, Tsai M. IgE and mast cells in allergic disease. Nat Med (2012) 18(5):693–704. doi:10.1038/nm.2755
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
18. Benado A, Nasagi-Atiya Y, Sagi-Eisenberg R. Protein trafficking in immune cells. Immunobiology (2009) 214(6):403–21. doi:10.1016/j.imbio.2008.11.011
19. Stow JL, Murray RZ. Intracellular trafficking and secretion of inflammatory cytokines. Cytokine Growth Factor Rev (2013) 24(3):227–39. doi:10.1016/j.cytogfr.2013.04.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
20. Schwartz LB, Lewis RA, Seldin D, Austen KF. Acid hydrolases and tryptase from secretory granules of dispersed human lung mast cells. J Immunol (1981) 126(4):1290–4.
21. Miller JS, Westin EH, Schwartz LB. Cloning and characterization of complementary DNA for human tryptase. J Clin Invest (1989) 84(4):1188–95. doi:10.1172/JCI114284
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
22. Miller JS, Moxley G, Schwartz LB. Cloning and characterization of a second complementary DNA for human tryptase. J Clin Invest (1990) 86(3):864–70. doi:10.1172/JCI114786
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
23. Vanderslice P, Ballinger SM, Tam EK, Goldstein SM, Craik CS, Caughey GH. Human mast cell tryptase: multiple cDNAs and genes reveal a multigene serine protease family. Proc Natl Acad Sci U S A (1990) 87(10):3811–5. doi:10.1073/pnas.87.10.3811
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
24. Wong GW, Tang Y, Feyfant E, Sali A, Li L, Li Y, et al. Identification of a new member of the tryptase family of mouse and human mast cell proteases which possesses a novel COOH-terminal hydrophobic extension. J Biol Chem (1999) 274(43):30784–93. doi:10.1074/jbc.274.43.30784
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
25. Caughey GH, Raymond WW, Blount JL, Hau LW, Pallaoro M, Wolters PJ, et al. Characterization of human gamma-tryptases, novel members of the chromosome 16p mast cell tryptase and prostasin gene families. J Immunol (2000) 164(12):6566–75. doi:10.4049/jimmunol.164.12.6566
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
26. Wang HW, McNeil HP, Husain A, Liu K, Tedla N, Thomas PS, et al. Delta tryptase is expressed in multiple human tissues, and a recombinant form has proteolytic activity. J Immunol (2002) 169(9):5145–52. doi:10.4049/jimmunol.169.9.5145
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
27. Wong GW, Foster PS, Yasuda S, Qi JC, Mahalingam S, Mellor EA, et al. Biochemical and functional characterization of human transmembrane tryptase (TMT)/tryptase gamma. TMT is an exocytosed mast cell protease that induces airway hyperresponsiveness in vivo via an interleukin-13/interleukin-4 receptor alpha/signal transducer and activator of transcription (STAT) 6-dependent pathway. J Biol Chem (2002) 277(44):41906–15. doi:10.1074/jbc.M205868200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
28. Schwartz LB, Irani AM, Roller K, Castells MC, Schechter NM. Quantitation of histamine, tryptase, and chymase in dispersed human T and TC mast cells. J Immunol (1987) 138(8):2611–5.
29. Pejler G, Knight SD, Henningsson F, Wernersson S. Novel insights into the biological function of mast cell carboxypeptidase A. Trends Immunol (2009) 30(8):401–8. doi:10.1016/j.it.2009.04.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
30. Schechter NM, Wang ZM, Blacher RW, Lessin SR, Lazarus GS, Rubin H. Determination of the primary structures of human skin chymase and cathepsin G from cutaneous mast cells of urticaria pigmentosa lesions. J Immunol (1994) 152(8):4062–9.
31. Strik MC, de Koning PJ, Kleijmeer MJ, Bladergroen BA, Wolbink AM, Griffith JM, et al. Human mast cells produce and release the cytotoxic lymphocyte associated protease granzyme B upon activation. Mol Immunol (2007) 44(14):3462–72. doi:10.1016/j.molimm.2007.03.024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
32. Olszewski MB, Groot AJ, Dastych J, Knol EF. TNF trafficking to human mast cell granules: mature chain-dependent endocytosis. J Immunol (2007) 178(9):5701–9. doi:10.4049/jimmunol.178.9.5701
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
33. Dvorak AM, Morgan ES, Weller PF. Ultrastructural immunolocalization of basic fibroblast growth factor to lipid bodies and secretory granules in human mast cells. Histochem J (2001) 33(7):397–402. doi:10.1023/A:1013771827069
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
34. Wilson SJ, Shute JK, Holgate ST, Howarth PH, Bradding P. Localization of interleukin (IL) -4 but not IL-5 to human mast cell secretory granules by immunoelectron microscopy. Clin Exp Allergy (2000) 30(4):493–500. doi:10.1046/j.1365-2222.2000.00756.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
35. de Paulis A, Minopoli G, Arbustini E, de Crescenzo G, Dal Piaz F, Pucci P, et al. Stem cell factor is localized in, released from, and cleaved by human mast cells. J Immunol (1999) 163(5):2799–808.
36. Stevens RL, Fox CC, Lichtenstein LM, Austen KF. Identification of chondroitin sulfate E proteoglycans and heparin proteoglycans in the secretory granules of human lung mast cells. Proc Natl Acad Sci U S A (1988) 85(7):2284–7. doi:10.1073/pnas.85.7.2284
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
37. Metcalfe DD, Lewis RA, Silbert JE, Rosenberg RD, Wasserman SI, Austen KF. Isolation and characterization of heparin from human lung. J Clin Invest (1979) 64(6):1537–43. doi:10.1172/JCI109613
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
38. Uvnas B. Recent observations on mechanisms of storage and release of mast cell histamine. Applicability to other biogenic amines. Agents Actions Suppl (1992) 36:23–33.
39. Hogan AD, Schwartz LB. Markers of mast cell degranulation. Methods (1997) 13(1):43–52. doi:10.1006/meth.1997.0494
40. Hohman RJ, Dreskin SC. Measuring degranulation of mast cells. Curr Protoc Immunol (2001) Chapter 7:Unit726. doi:10.1002/0471142735.im0726s08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
41. Douaiher J, Succar J, Lancerotto L, Gurish MF, Orgill DP, Hamilton MJ, et al. Development of mast cells and importance of their tryptase and chymase serine proteases in inflammation and wound healing. Adv Immunol (2014) 122:211–52. doi:10.1016/B978-0-12-800267-4.00006-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
42. Baram D, Adachi R, Medalia O, Tuvim M, Dickey BF, Mekori YA, et al. Synaptotagmin II negatively regulates Ca2+-triggered exocytosis of lysosomes in mast cells. J Exp Med (1999) 189(10):1649–58. doi:10.1084/jem.189.10.1649
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
43. Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, Desaymard C. Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol Biol Cell (1997) 8(12):2631–45. doi:10.1091/mbc.8.12.2631
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
44. Puri N, Roche PA. Mast cells possess distinct secretory granule subsets whose exocytosis is regulated by different SNARE isoforms. Proc Natl Acad Sci U S A (2008) 105(7):2580–5. doi:10.1073/pnas.0707854105
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
45. Gunin AG, Kornilova NK, Vasilieva OV, Petrov VV. Age-related changes in proliferation, the numbers of mast cells, eosinophils, and CD45-positive cells in human dermis. J Gerontol A Biol Sci Med Sci (2011) 66(4):385–92. doi:10.1093/gerona/glq205
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
46. Hammel I, Dvorak AM, Peters SP, Schulman ES, Dvorak HF, Lichtenstein LM, et al. Differences in the volume distributions of human lung mast cell granules and lipid bodies: evidence that the size of these organelles is regulated by distinct mechanisms. J Cell Biol (1985) 100(5):1488–92. doi:10.1083/jcb.100.5.1488
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
47. Riedel D, Antonin W, Fernandez-Chacon R, Alvarez de Toledo G, Jo T, Geppert M, et al. Rab3D is not required for exocrine exocytosis but for maintenance of normally sized secretory granules. Mol Cell Biol (2002) 22(18):6487–97. doi:10.1128/MCB.22.18.6487-6497.2002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
48. Azouz NP, Zur N, Efergan A, Ohbayashi N, Fukuda M, Amihai D, et al. Rab5 is a novel regulator of mast cell secretory granules: impact on size, cargo, and exocytosis. J Immunol (2014) 192(9):4043–53. doi:10.4049/jimmunol.1302196
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
49. Durchfort N, Verhoef S, Vaughn MB, Shrestha R, Adam D, Kaplan J, et al. The enlarged lysosomes in beige j cells result from decreased lysosome fission and not increased lysosome fusion. Traffic (2012) 13(1):108–19. doi:10.1111/j.1600-0854.2011.01300.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
50. Grimberg E, Peng Z, Hammel I, Sagi-Eisenberg R. Synaptotagmin III is a critical factor for the formation of the perinuclear endocytic recycling compartment and determination of secretory granules size. J Cell Sci (2003) 116(Pt 1):145–54. doi:10.1242/jcs.00186
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
51. Prasad P, Yanagihara AA, Small-Howard AL, Turner H, Stokes AJ. Secretogranin III directs secretory vesicle biogenesis in mast cells in a manner dependent upon interaction with chromogranin A. J Immunol (2008) 181(7):5024–34. doi:10.4049/jimmunol.181.7.5024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
52. Henningsson F, Hergeth S, Cortelius R, Abrink M, Pejler G. A role for serglycin proteoglycan in granular retention and processing of mast cell secretory granule components. FEBS J (2006) 273(21):4901–12. doi:10.1111/j.1742-4658.2006.05489.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
53. Melo FR, Waern I, Ronnberg E, Abrink M, Lee DM, Schlenner SM, et al. A role for serglycin proteoglycan in mast cell apoptosis induced by a secretory granule-mediated pathway. J Biol Chem (2011) 286(7):5423–33. doi:10.1074/jbc.M110.176461
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
54. Nakazawa S, Sakanaka M, Furuta K, Natsuhara M, Takano H, Tsuchiya S, et al. Histamine synthesis is required for granule maturation in murine mast cells. Eur J Immunol (2014) 44(1):204–14. doi:10.1002/eji.201343838
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
55. Doray B, Kornfeld S. Gamma subunit of the AP-1 adaptor complex binds clathrin: implications for cooperative binding in coated vesicle assembly. Mol Biol Cell (2001) 12(7):1925–35. doi:10.1091/mbc.12.7.1925
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
56. Bonnemaison ML, Eipper BA, Mains RE. Role of adaptor proteins in secretory granule biogenesis and maturation. Front Endocrinol (2013) 4:101. doi:10.3389/fendo.2013.00101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
57. Lagunoff D. Analysis of dye binding sites in mast cell granules. Biochemistry (1974) 13(19):3982–6. doi:10.1021/bi00716a025
58. Nitzany E, Hammel I, Meilijson I. Quantal basis of vesicle growth and information content, a unified approach. J Theor Biol (2010) 266(1):202–9. doi:10.1016/j.jtbi.2010.06.019
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
59. Chin WC, Orellana MV, Quesada I, Verdugo P. Secretion in unicellular marine phytoplankton: demonstration of regulated exocytosis in Phaeocystis globosa. Plant Cell Physiol (2004) 45(5):535–42. doi:10.1093/pcp/pch062
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
60. Ronnberg E, Melo FR, Pejler G. Mast cell proteoglycans. J Histochem Cytochem (2012) 60(12):950–62. doi:10.1369/0022155412458927
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
61. Ikeya M, Yamanoue K, Mochizuki Y, Konishi H, Tadokoro S, Tanaka M, et al. Orai-2 is localized on secretory granules and regulates antigen-evoked Ca2+ mobilization and exocytosis in mast cells. Biochem Biophys Res Commun (2014) 451(1):62–7. doi:10.1016/j.bbrc.2014.07.060
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
62. Melicoff E, Sansores-Garcia L, Gomez A, Moreira DC, Datta P, Thakur P, et al. Synaptotagmin-2 controls regulated exocytosis but not other secretory responses of mast cells. J Biol Chem (2009) 284(29):19445–51. doi:10.1074/jbc.M109.002550
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
63. Biederbick A, Licht A, Kleene R. Serglycin proteoglycan is sorted into zymogen granules of rat pancreatic acinar cells. Eur J Cell Biol (2003) 82(1):19–29. doi:10.1078/0171-9335-00287
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
64. Merickel A, Edwards RH. Transport of histamine by vesicular monoamine transporter-2. Neuropharmacology (1995) 34(11):1543–7. doi:10.1016/0028-3908(95)00148-Y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
65. Travis ER, Wang YM, Michael DJ, Caron MG, Wightman RM. Differential quantal release of histamine and 5-hydroxytryptamine from mast cells of vesicular monoamine transporter 2 knockout mice. Proc Natl Acad Sci U S A (2000) 97(1):162–7. doi:10.1073/pnas.97.1.162
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
66. Ringvall M, Ronnberg E, Wernersson S, Duelli A, Henningsson F, Abrink M, et al. Serotonin and histamine storage in mast cell secretory granules is dependent on serglycin proteoglycan. J Allergy Clin Immunol (2008) 121(4):1020–6. doi:10.1016/j.jaci.2007.11.031
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
67. Humphries DE, Wong GW, Friend DS, Gurish MF, Qiu WT, Huang C, et al. Heparin is essential for the storage of specific granule proteases in mast cells. Nature (1999) 400(6746):769–72. doi:10.1038/23481
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
68. Uvnas B, Aborg CH, Bergendorff A. Storage of histamine in mast cells. Evidence for an ionic binding of histamine to protein carboxyls in the granule heparin-protein complex. Acta Physiol Scand Suppl (1970) 336:1–26.
69. Rabenstein DL, Bratt P, Peng J. Quantitative characterization of the binding of histamine by heparin. Biochemistry (1998) 37(40):14121–7. doi:10.1021/bi980625y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
70. Bergendorff A, Uvnas B. Storage of 5-hydroxytryptamine in rat mast cells. Evidence for an ionic binding to carboxyl groups in a granule heparin-protein complex. Acta Physiol Scand (1972) 84(3):320–31. doi:10.1111/j.1748-1716.1972.tb05183.x
71. Garcia-Faroldi G, Rodriguez CE, Urdiales JL, Perez-Pomares JM, Davila JC, Pejler G, et al. Polyamines are present in mast cell secretory granules and are important for granule homeostasis. PLoS One (2010) 5(11):11. doi:10.1371/journal.pone.0015071
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
72. Olszewski MB, Trzaska D, Knol EF, Adamczewska V, Dastych J. Efficient sorting of TNF-alpha to rodent mast cell granules is dependent on N-linked glycosylation. Eur J Immunol (2006) 36(4):997–1008. doi:10.1002/eji.200535323
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
73. Benyon RC, Bissonnette EY, Befus AD. Tumor necrosis factor-alpha dependent cytotoxicity of human skin mast cells is enhanced by anti-IgE antibodies. J Immunol (1991) 147(7):2253–8.
74. Gordon JR, Galli SJ. Mast cells as a source of both preformed and immunologically inducible TNF-alpha/cachectin. Nature (1990) 346(6281):274–6. doi:10.1038/346274a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
75. Okayama Y, Kobayashi H, Ashman LK, Dobashi K, Nakazawa T, Holgate ST, et al. Human lung mast cells are enriched in the capacity to produce granulocyte-macrophage colony-stimulating factor in response to IgE-dependent stimulation. Eur J Immunol (1998) 28(2):708–15. doi:10.1002/(SICI)1521-4141(199802)28:02<708::AID-IMMU708>3.0.CO;2-A
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
76. Voehringer D. Protective and pathological roles of mast cells and basophils. Nat Rev Immunol (2013) 13(5):362–75. doi:10.1038/nri3427
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
77. Moscatelli D. High and low affinity binding sites for basic fibroblast growth factor on cultured cells: absence of a role for low affinity binding in the stimulation of plasminogen activator production by bovine capillary endothelial cells. J Cell Physiol (1987) 131(1):123–30. doi:10.1002/jcp.1041310118
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
78. Dvorak AM, McLeod RS, Onderdonk A, Monahan-Earley RA, Cullen JB, Antonioli DA, et al. Ultrastructural evidence for piecemeal and anaphylactic degranulation of human gut mucosal mast cells in vivo. Int Arch Allergy Immunol (1992) 99(1):74–83. doi:10.1159/000236338
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
79. Dvorak AM, Morgan ES. Diamine oxidase-gold enzyme-affinity ultrastructural demonstration that human gut mucosal mast cells secrete histamine by piecemeal degranulation in vivo. J Allergy Clin Immunol (1997) 99(6 Pt 1):812–20. doi:10.1016/S0091-6749(97)80016-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
80. Ebihara N, Watanabe Y, Murakami A. The ultramicrostructure of secretory granules of mast cells in giant papillae of vernal keratoconjunctivitis patients. Nihon Ganka Gakkai Zasshi (2008) 112(7):581–9.
81. Dvorak AM, Massey W, Warner J, Kissell S, Kagey-Sobotka A, Lichtenstein LM. IgE-mediated anaphylactic degranulation of isolated human skin mast cells. Blood (1991) 77(3):569–78.
82. Dvorak AM, Kissell S. Granule changes of human skin mast cells characteristic of piecemeal degranulation and associated with recovery during wound healing in situ. J Leukoc Biol (1991) 49(2):197–210.
83. Dvorak AM, Tepper RI, Weller PF, Morgan ES, Estrella P, Monahan-Earley RA, et al. Piecemeal degranulation of mast cells in the inflammatory eyelid lesions of interleukin-4 transgenic mice. Evidence of mast cell histamine release in vivo by diamine oxidase-gold enzyme-affinity ultrastructural cytochemistry. Blood (1994) 83(12):3600–12.
84. Chyczewski L, Debek W, Chyczewska E, Debek K, Bankowski E. Morphology of lung mast cells in rats treated with bleomycin. Exp Toxicol Pathol (1996) 48(6):515–7. doi:10.1016/S0940-2993(96)80070-4
85. Vicario M, Guilarte M, Alonso C, Yang P, Martinez C, Ramos L, et al. Chronological assessment of mast cell-mediated gut dysfunction and mucosal inflammation in a rat model of chronic psychosocial stress. Brain Behav Immun (2010) 24(7):1166–75. doi:10.1016/j.bbi.2010.06.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
86. Yan XJ, Feng CC, Liu Q, Zhang LY, Dong X, Liu ZL, et al. Vagal afferents mediate antinociception of estrogen in a rat model of visceral pain: the involvement of intestinal mucosal mast cells and 5-hydroxytryptamine 3 signaling. J Pain (2014) 15(2):204–17. doi:10.1016/j.jpain.2013.10.012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
87. Iwamoto S, Asada Y, Ebihara N, Hori K, Okayama Y, Kashiwakura J, et al. Interaction between conjunctival epithelial cells and mast cells induces CCL2 expression and piecemeal degranulation in mast cells. Invest Ophthalmol Vis Sci (2013) 54(4):2465–73. doi:10.1167/iovs.12-10664
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
88. McCurdy JD, Olynych TJ, Maher LH, Marshall JS. Cutting edge: distinct Toll-like receptor 2 activators selectively induce different classes of mediator production from human mast cells. J Immunol (2003) 170(4):1625–9. doi:10.4049/jimmunol.170.4.1625
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
89. Frossi B, D’Inca F, Crivellato E, Sibilano R, Gri G, Mongillo M, et al. Single-cell dynamics of mast cell-CD4+ CD25+ regulatory T cell interactions. Eur J Immunol (2011) 41(7):1872–82. doi:10.1002/eji.201041300
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
90. Crivellato E, Nico B, Gallo VP, Ribatti D. Cell secretion mediated by granule-associated vesicle transport: a glimpse at evolution. Anat Rec (2010) 293(7):1115–24. doi:10.1002/ar.21146
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
91. Woska JR Jr., Gillespie ME. SNARE complex-mediated degranulation in mast cells. J Cell Mol Med (2012) 16(4):649–56. doi:10.1111/j.1582-4934.2011.01443.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
92. Sander LE, Frank SP, Bolat S, Blank U, Galli T, Bigalke H, et al. Vesicle associated membrane protein (VAMP)-7 and VAMP-8, but not VAMP-2 or VAMP-3, are required for activation-induced degranulation of mature human mast cells. Eur J Immunol (2008) 38(3):855–63. doi:10.1002/eji.200737634
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
93. Goishi K, Mizuno K, Nakanishi H, Sasaki T. Involvement of Rab27 in antigen-induced histamine release from rat basophilic leukemia 2H3 cells. Biochem Biophys Res Commun (2004) 324(1):294–301. doi:10.1016/j.bbrc.2004.09.050
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
94. Martin-Verdeaux S, Pombo I, Iannascoli B, Roa M, Varin-Blank N, Rivera J, et al. Evidence of a role for Munc18-2 and microtubules in mast cell granule exocytosis. J Cell Sci (2003) 116(Pt 2):325–34. doi:10.1242/jcs.00216
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
95. Tadokoro S, Kurimoto T, Nakanishi M, Hirashima N. Munc18-2 regulates exocytotic membrane fusion positively interacting with syntaxin-3 in RBL-2H3 cells. Mol Immunol (2007) 44(13):3427–33. doi:10.1016/j.molimm.2007.02.013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
96. Tadokoro S, Nakanishi M, Hirashima N. Complexin II facilitates exocytotic release in mast cells by enhancing Ca2+ sensitivity of the fusion process. J Cell Sci (2005) 118(Pt 10):2239–46. doi:10.1242/jcs.02338
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
97. Hammel I, Meilijson I. The stealthy nano-machine behind mast cell granule size distribution. Mol Immunol (2015) 63(1):45–54. doi:10.1016/j.molimm.2014.02.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
98. Jena BP. Secretory vesicles transiently dock and fuse at the porosome to discharge contents during cell secretion. Cell Biol Int (2010) 34(1):3–12. doi:10.1042/CBI20090161
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
99. Baram D, Peng Z, Medalia O, Mekori YA, Sagi-Eisenberg R. Synaptotagmin II negatively regulates MHC class II presentation by mast cells. Mol Immunol (2002) 38(16-18):1347–52. doi:10.1016/S0161-5890(02)00086-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
100. Haberman Y, Ziv I, Gorzalczany Y, Hirschberg K, Mittleman L, Fukuda M, et al. Synaptotagmin (Syt) IX is an essential determinant for protein sorting to secretory granules in mast cells. Blood (2007) 109(8):3385–92. doi:10.1182/blood-2006-07-033126
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
101. Roa M, Paumet F, Le Mao J, David B, Blank U. Involvement of the ras-like GTPase rab3d in RBL-2H3 mast cell exocytosis following stimulation via high affinity IgE receptors (Fc epsilonRI). J Immunol (1997) 159(6):2815–23.
102. Smith J, Thompson N, Thompson J, Armstrong J, Hayes B, Crofts A, et al. Rat basophilic leukaemia (RBL) cells overexpressing Rab3a have a reversible block in antigen-stimulated exocytosis. Biochem J (1997) 323(Pt 2):321–8.
103. Tuvim MJ, Adachi R, Chocano JF, Moore RH, Lampert RM, Zera E, et al. Rab3D, a small GTPase, is localized on mast cell secretory granules and translocates to the plasma membrane upon exocytosis. Am J Respir Cell Mol Biol (1999) 20(1):79–89. doi:10.1165/ajrcmb.20.1.3279
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
104. Elstak ED, Neeft M, Nehme NT, Voortman J, Cheung M, Goodarzifard M, et al. The munc13-4-rab27 complex is specifically required for tethering secretory lysosomes at the plasma membrane. Blood (2011) 118(6):1570–8. doi:10.1182/blood-2011-02-339523
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
105. Singh RK, Mizuno K, Wasmeier C, Wavre-Shapton ST, Recchi C, Catz SD, et al. Distinct and opposing roles for Rab27a/Mlph/MyoVa and Rab27b/Munc13-4 in mast cell secretion. FEBS J (2013) 280(3):892–903. doi:10.1111/febs.12081
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
106. Mizuno K, Tolmachova T, Ushakov DS, Romao M, Abrink M, Ferenczi MA, et al. Rab27b regulates mast cell granule dynamics and secretion. Traffic (2007) 8(7):883–92. doi:10.1111/j.1600-0854.2007.00571.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
107. Hong-Geller E, Cerione RA. Cdc42 and Rac stimulate exocytosis of secretory granules by activating the IP3/calcium pathway in RBL-2H3 mast cells. J Cell Biol (2000) 148(3):481–94. doi:10.1083/jcb.148.3.481
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
108. Hong-Geller E, Holowka D, Siraganian RP, Baird B, Cerione RA. Activated Cdc42/Rac reconstitutes Fcepsilon RI-mediated Ca2+ mobilization and degranulation in mutant RBL mast cells. Proc Natl Acad Sci U S A (2001) 98(3):1154–9. doi:10.1073/pnas.98.3.1154
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
109. Baier A, Ndoh VN, Lacy P, Eitzen G. Rac1 and Rac2 control distinct events during antigen-stimulated mast cell exocytosis. J Leukoc Biol (2014) 95:763–74. doi:10.1189/jlb.0513281
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
110. Ogawa K, Tanaka Y, Uruno T, Duan X, Harada Y, Sanematsu F, et al. DOCK5 functions as a key signaling adaptor that links FcepsilonRI signals to microtubule dynamics during mast cell degranulation. J Exp Med (2014) 211(7):1407–19. doi:10.1084/jem.20131926
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
111. Haddock BJ, Zhu Y, Doyle SP, Abdullah LH, Davis CW. Role of MARCKS in regulated secretion from mast cells and airway goblet cells. Am J Physiol Lung Cell Mol Physiol (2014) 306(10):L925–36. doi:10.1152/ajplung.00213.2013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
112. Bond LM, Peden AA, Kendrick-Jones J, Sellers JR, Buss F. Myosin VI and its binding partner optineurin are involved in secretory vesicle fusion at the plasma membrane. Mol Biol Cell (2011) 22(1):54–65. doi:10.1091/mbc.E10-06-0553
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
113. Fahrner M, Derler I, Jardin I, Romanin C. The STIM1/Orai signaling machinery. Channels (2013) 7(5):330–43. doi:10.4161/chan.26742
114. Ashmole I, Bradding P. Ion channels regulating mast cell biology. Clin Exp Allergy (2013) 43(5):491–502. doi:10.1111/cea.12043
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
115. Inoh Y, Tadokoro S, Tanabe H, Inoue M, Hirashima N, Nakanishi M, et al. Inhibitory effects of a cationic liposome on allergic reaction mediated by mast cell activation. Biochem Pharmacol (2013) 86(12):1731–8. doi:10.1016/j.bcp.2013.09.023
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
116. Ma HT, Beaven MA. Regulators of Ca(2+) signaling in mast cells: potential targets for treatment of mast cell-related diseases? Adv Exp Med Biol (2011) 716:62–90. doi:10.1007/978-1-4419-9533-9_5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
117. Ushio H, Ueno T, Kojima Y, Komatsu M, Tanaka S, Yamamoto A, et al. Crucial role for autophagy in degranulation of mast cells. J Allergy Clin Immunol (2011) 127(5):1267–76e6. doi:10.1016/j.jaci.2010.12.1078
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
118. Kowal J, Tkach M, Thery C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol (2014) 29C:116–25. doi:10.1016/j.ceb.2014.05.004
119. Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol (2007) 9(6):654–9. doi:10.1038/ncb1596
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
120. Ekstrom K, Valadi H, Sjostrand M, Malmhall C, Bossios A, Eldh M, et al. Characterization of mRNA and microRNA in human mast cell-derived exosomes and their transfer to other mast cells and blood CD34 progenitor cells. J Extracell Vesicles (2012) 1:18389. doi:10.3402/jev.v1i0.18389
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
121. Skokos D, Botros HG, Demeure C, Morin J, Peronet R, Birkenmeier G, et al. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J Immunol (2003) 170(6):3037–45. doi:10.4049/jimmunol.170.6.3037
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
122. Merluzzi S, Betto E, Ceccaroni AA, Magris R, Giunta M, Mion F. Mast cells, basophils and B cell connection network. Mol Immunol (2015) 63(1):94–103. doi:10.1016/j.molimm.2014.02.016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
123. Villarroya-Beltri C, Baixauli F, Gutierrez-Vazquez C, Sanchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. Semin Cancer Biol (2014) 28C:3–13. doi:10.1016/j.semcancer.2014.04.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
124. Frank SP, Thon KP, Bischoff SC, Lorentz A. SNAP-23 and syntaxin-3 are required for chemokine release by mature human mast cells. Mol Immunol (2011) 49(1-2):353–8. doi:10.1016/j.molimm.2011.09.011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
125. Tiwari N, Wang CC, Brochetta C, Ke G, Vita F, Qi Z, et al. VAMP-8 segregates mast cell-preformed mediator exocytosis from cytokine trafficking pathways. Blood (2008) 111(7):3665–74. doi:10.1182/blood-2007-07-103309
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
126. Hogaboam CM, Donigi-Gale D, Shoupe TS, Bissonnette EY, Befus AD, Wallace JL. Platelet-activating factor synthesis by peritoneal mast cells and its inhibition by two quinoline-based compounds. Br J Pharmacol (1992) 105(1):87–92. doi:10.1111/j.1476-5381.1992.tb14215.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
127. Mencia-Huerta JM, Lewis RA, Razin E, Austen KF. Antigen-initiated release of platelet-activating factor (PAF-acether) from mouse bone marrow-derived mast cells sensitized with monoclonal IgE. J Immunol (1983) 131(6):2958–64.
128. Schleimer RP, Davidson DA, Lichtenstein LM, Adkinson NF Jr. Selective inhibition of arachidonic acid metabolite release from human lung tissue by antiinflammatory steroids. J Immunol (1986) 136(8):3006–11.
129. Boyce JA. Eicosanoid mediators of mast cells: receptors, regulation of synthesis, and pathobiologic implications. Chem Immunol Allergy (2005) 87:59–79. doi:10.1159/000087571
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
130. Peters-Golden M, Canetti C, Mancuso P, Coffey MJ. Leukotrienes: underappreciated mediators of innate immune responses. J Immunol (2005) 174(2):589–94. doi:10.4049/jimmunol.174.2.589
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
131. Joo M, Sadikot RT. PGD synthase and PGD2 in immune response. Mediators Inflamm (2012) 2012:503128. doi:10.1155/2012/503128
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
132. Moon TC, Campos-Alberto E, Yoshimura T, Bredo G, Rieger AM, Puttagunta L, et al. Expression of DP2 (CRTh2), a prostaglandin D2 receptor, in human mast cells. PLoS One (2014) 9(9):e108595. doi:10.1371/journal.pone.0108595
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
133. Binda C, Genier S, Cartier A, Larrivee JF, Stankova J, Young JC, et al. A G protein-coupled receptor and the intracellular synthase of its agonist functionally cooperate. J Cell Biol (2014) 204(3):377–93. doi:10.1083/jcb.201304015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
134. Brock TG, McNish RW, Peters-Golden M. Capacity for repeatable leukotriene generation after transient stimulation of mast cells and macrophages. Biochem J (1998) 329(Pt 3):519–25.
135. Brock TG, McNish RW, Peters-Golden M. Translocation and leukotriene synthetic capacity of nuclear 5-lipoxygenase in rat basophilic leukemia cells and alveolar macrophages. J Biol Chem (1995) 270(37):21652–8. doi:10.1074/jbc.270.37.21652
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
136. Balestrieri B, Hsu VW, Gilbert H, Leslie CC, Han WK, Bonventre JV, et al. Group V secretory phospholipase A2 translocates to the phagosome after zymosan stimulation of mouse peritoneal macrophages and regulates phagocytosis. J Biol Chem (2006) 281(10):6691–8. doi:10.1074/jbc.M508314200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
137. Dvorak AM, Dvorak HF, Peters SP, Shulman ES, MacGlashan DW Jr., Pyne K, et al. Lipid bodies: cytoplasmic organelles important to arachidonate metabolism in macrophages and mast cells. J Immunol (1983) 131(6):2965–76.
138. Pacheco P, Bozza FA, Gomes RN, Bozza M, Weller PF, Castro-Faria-Neto HC, et al. Lipopolysaccharide-induced leukocyte lipid body formation in vivo: innate immunity elicited intracellular Loci involved in eicosanoid metabolism. J Immunol (2002) 169(11):6498–506. doi:10.4049/jimmunol.169.11.6498
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
139. Bozza PT, Bakker-Abreu I, Navarro-Xavier RA, Bandeira-Melo C. Lipid body function in eicosanoid synthesis: an update. Prostaglandins Leukot Essent Fatty Acids (2011) 85(5):205–13. doi:10.1016/j.plefa.2011.04.020
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
140. Thiam AR, Farese RV Jr., Walther TC. The biophysics and cell biology of lipid droplets. Nat Rev Mol Cell Biol (2013) 14(12):775–86. doi:10.1038/nrm3699
141. Bozza PT, Yu W, Cassara J, Weller PF. Pathways for eosinophil lipid body induction: differing signal transduction in cells from normal and hypereosinophilic subjects. J Leukoc Biol (1998) 64(4):563–9.
142. Beil WJ, Weller PF, Peppercorn MA, Galli SJ, Dvorak AM. Ultrastructural immunogold localization of subcellular sites of TNF-alpha in colonic Crohn’s disease. J Leukoc Biol (1995) 58(3):284–98.
143. Reginato AJ, Schumacher HR, Allan DA, Rabinowitz JL. Acute monoarthritis associated with lipid liquid crystals. Ann Rheum Dis (1985) 44(8):537–43. doi:10.1136/ard.44.8.537
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
144. Dichlberger A, Kovanen PT, Schneider WJ. Mast cells: from lipid droplets to lipid mediators. Clin Sci (2013) 125(3):121–30. doi:10.1042/CS20120602
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
145. Wilfling F, Haas JT, Walther TC, Farese RV Jr. Lipid droplet biogenesis. Curr Opin Cell Biol (2014) 29C:39–45. doi:10.1016/j.ceb.2014.03.008
146. Bozza PT, Bandeira-Melo C. Mechanisms of leukocyte lipid body formation and function in inflammation. Mem Inst Oswaldo Cruz (2005) 100(Suppl 1):113–20. doi:10.1590/S0074-02762005000900020
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
147. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature (2014) 510(7503):92–101. doi:10.1038/nature13479
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
148. Chan BS, Satriano JA, Pucci M, Schuster VL. Mechanism of prostaglandin E2 transport across the plasma membrane of HeLa cells and Xenopus oocytes expressing the prostaglandin transporter "PGT". J Biol Chem (1998) 273(12):6689–97. doi:10.1074/jbc.273.12.6689
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
149. Jedlitschky G, Keppler D. Transport of leukotriene C4 and structurally related conjugates. Vitam Horm (2002) 64:153–84. doi:10.1016/S0083-6729(02)64005-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
150. Rius M, Hummel-Eisenbeiss J, Keppler D. ATP-dependent transport of leukotrienes B4 and C4 by the multidrug resistance protein ABCC4 (MRP4). J Pharmacol Exp Ther (2008) 324(1):86–94. doi:10.1124/jpet.107.131342
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
151. Reid G, Wielinga P, Zelcer N, van der Heijden I, Kuil A, de Haas M, et al. The human multidrug resistance protein MRP4 functions as a prostaglandin efflux transporter and is inhibited by nonsteroidal antiinflammatory drugs. Proc Natl Acad Sci U S A (2003) 100(16):9244–9. doi:10.1073/pnas.1033060100
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
152. Lin ZP, Zhu YL, Johnson DR, Rice KP, Nottoli T, Hains BC, et al. Disruption of cAMP and prostaglandin E2 transport by multidrug resistance protein 4 deficiency alters cAMP-mediated signaling and nociceptive response. Mol Pharmacol (2008) 73(1):243–51. doi:10.1124/mol.107.039594
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
153. Adachi H, Suzuki T, Abe M, Asano N, Mizutamari H, Tanemoto M, et al. Molecular characterization of human and rat organic anion transporter OATP-D. Am J Physiol Renal Physiol (2003) 285(6):F1188–97. doi:10.1152/ajprenal.00402.2002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
154. Nishio T, Adachi H, Nakagomi R, Tokui T, Sato E, Tanemoto M, et al. Molecular identification of a rat novel organic anion transporter moat1, which transports prostaglandin D2, leukotriene C4, and taurocholate. Biochem Biophys Res Commun (2000) 275(3):831–8. doi:10.1006/bbrc.2000.3377
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
155. Burd PR, Rogers HW, Gordon JR, Martin CA, Jayaraman S, Wilson SD, et al. Interleukin 3-dependent and -independent mast cells stimulated with IgE and antigen express multiple cytokines. J Exp Med (1989) 170(1):245–57. doi:10.1084/jem.170.1.245
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
156. Lewis RA, Soter NA, Diamond PT, Austen KF, Oates JA, Roberts LJ II. Prostaglandin D2 generation after activation of rat and human mast cells with anti-IgE. J Immunol (1982) 129(4):1627–31.
157. MacDonald AJ, Pick J, Bissonnette EY, Befus AD. Rat mucosal mast cells: the cultured bone marrow-derived mast cell is biochemically and functionally analogous to its counterpart in vivo. Immunology (1998) 93(4):533–9. doi:10.1046/j.1365-2567.1998.00465.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
158. Gilchrist M, McCauley SD, Befus AD. Expression, localization, and regulation of NOS in human mast cell lines: effects on leukotriene production. Blood (2004) 104(2):462–9. doi:10.1182/blood-2003-08-2990
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
159. Moon TC, Yoshimura T, Parsons T, Befus AD. Microenvironmental regulation of inducible nitric oxide synthase expression and nitric oxide production in mouse bone marrow-derived mast cells. J Leukoc Biol (2012) 91(4):581–90. doi:10.1189/jlb.0811436
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
160. Swindle EJ, Metcalfe DD, Coleman JW. Rodent and human mast cells produce functionally significant intracellular reactive oxygen species but not nitric oxide. J Biol Chem (2004) 279(47):48751–9. doi:10.1074/jbc.M409738200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
161. Wang XS, Sze Wing S, Kwok Ho Y, Lau HY. Functional characterization of human mast cells cultured from adult peripheral blood. Int Immunopharmacol (2006) 6(5):839–47. doi:10.1016/j.intimp.2005.12.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
162. Kulka M, Sheen CH, Tancowny BP, Grammer LC, Schleimer RP. Neuropeptides activate human mast cell degranulation and chemokine production. Immunology (2008) 123(3):398–410. doi:10.1111/j.1365-2567.2007.02705.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
163. Karimi K, Kool M, Nijkamp FP, Redegeld FA. Substance P can stimulate prostaglandin D(2) and leukotriene C(4) generation without granule exocytosis in murine mast cells. Eur J Pharmacol (2004) 489(1-2):49–54. doi:10.1016/j.ejphar.2004.03.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
164. Ogasawara T, Murakami M, Suzuki-Nishimura T, Uchida MK, Kudo I. Mouse bone marrow-derived mast cells undergo exocytosis, prostanoid generation, and cytokine expression in response to G protein-activating polybasic compounds after coculture with fibroblasts in the presence of c-kit ligand. J Immunol (1997) 158(1):393–404.
165. Reynier-Rebuffel AM, Mathiau P, Callebert J, Dimitriadou V, Farjaudon N, Kacem K, et al. Substance P, calcitonin gene-related peptide, and capsaicin release serotonin from cerebrovascular mast cells. Am J Physiol (1994) 267(5 Pt 2):R1421–9.
166. McNeil BD, Pundir P, Han L, Meeker S, Undem BJ, Kulka M, et al. Identification of the receptor for mast cell basic secretagogues and its critical role in pseudo-allergic drug reactions. Nature (2014) (in press).
167. Niyonsaba F, Someya A, Hirata M, Ogawa H, Nagaoka I. Evaluation of the effects of peptide antibiotics human beta-defensins-1/-2 and LL-37 on histamine release and prostaglandin D(2) production from mast cells. Eur J Immunol (2001) 31(4):1066–75. doi:10.1002/1521-4141(200104)31:4<1066::AID-IMMU1066>3.0.CO;2-#
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
168. Niyonsaba F, Ushio H, Hara M, Yokoi H, Tominaga M, Takamori K, et al. Antimicrobial peptides human beta-defensins and cathelicidin LL-37 induce the secretion of a pruritogenic cytokine IL-31 by human mast cells. J Immunol (2010) 184(7):3526–34. doi:10.4049/jimmunol.0900712
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
169. Befus AD, Mowat C, Gilchrist M, Hu J, Solomon S, Bateman A. Neutrophil defensins induce histamine secretion from mast cells: mechanisms of action. J Immunol (1999) 163(2):947–53.
170. Subramanian H, Gupta K, Lee D, Bayir AK, Ahn H, Ali H. Beta-defensins activate human mast cells via Mas-related gene X2. J Immunol (2013) 191(1):345–52. doi:10.4049/jimmunol.1300023
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
171. Pundir P, Catalli A, Leggiadro C, Douglas SE, Kulka M. Pleurocidin, a novel antimicrobial peptide, induces human mast cell activation through the FPRL1 receptor. Mucosal Immunol (2014) 7(1):177–87. doi:10.1038/mi.2013.37
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
172. Behrendt H, Rosenkranz U, Schmutzler W. Ultrastructure of isolated human mast cells during histamine release induced by ionophore A 23187. Int Arch Allergy Appl Immunol (1978) 56(2):188–92. doi:10.1159/000232022
173. Balletta A, Lorenz D, Rummel A, Gerhard R, Bigalke H, Wegner F. Human mast cell line-1 (HMC-1) cells exhibit a membrane capacitance increase when dialysed with high free-Ca(2+) and GTPgammaS containing intracellular solution. Eur J Pharmacol (2013) 720(1-3):227–36. doi:10.1016/j.ejphar.2013.10.022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
174. Hosokawa J, Suzuki K, Nakagomi D, Tamachi T, Takatori H, Suto A, et al. Role of calcium ionophore A23187-induced activation of IkappaB kinase 2 in mast cells. Int Arch Allergy Immunol (2013) 161(Suppl 2):37–43. doi:10.1159/000350357
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
175. Sheen CH, Schleimer RP, Kulka M. Codeine induces human mast cell chemokine and cytokine production: involvement of G-protein activation. Allergy (2007) 62(5):532–8. doi:10.1111/j.1398-9995.2007.01345.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
176. Shalit M, Rubinow A, Granit L, Bar-Sela S. Codeine-induced mast cell degranulation in human skin: effect of calcium channel blockers. Ann Allergy (1987) 59(6):461–3.
177. Kalesnikoff J, Huber M, Lam V, Damen JE, Zhang J, Siraganian RP, et al. Monomeric IgE stimulates signaling pathways in mast cells that lead to cytokine production and cell survival. Immunity (2001) 14(6):801–11. doi:10.1016/S1074-7613(01)00159-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
178. Pandey V, Mihara S, Fensome-Green A, Bolsover S, Cockcroft S. Monomeric IgE stimulates NFAT translocation into the nucleus, a rise in cytosol Ca2+, degranulation, and membrane ruffling in the cultured rat basophilic leukemia-2H3 mast cell line. J Immunol (2004) 172(7):4048–58. doi:10.4049/jimmunol.172.7.4048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
179. Matsuda K, Piliponsky AM, Iikura M, Nakae S, Wang EW, Dutta SM, et al. Monomeric IgE enhances human mast cell chemokine production: IL-4 augments and dexamethasone suppresses the response. J Allergy Clin Immunol (2005) 116(6):1357–63. doi:10.1016/j.jaci.2005.08.042
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
180. Marshall JS, Gomi K, Blennerhassett MG, Bienenstock J. Nerve growth factor modifies the expression of inflammatory cytokines by mast cells via a prostanoid-dependent mechanism. J Immunol (1999) 162(7):4271–6.
181. Tomioka M, Stead RH, Nielsen L, Coughlin MD, Bienenstock J. Nerve growth factor enhances antigen and other secretagogue-induced histamine release from rat peritoneal mast cells in the absence of phosphatidylserine. J Allergy Clin Immunol (1988) 82(4):599–607. doi:10.1016/0091-6749(88)90971-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
182. Yang Z, Marshall JS. Zymosan treatment of mouse mast cells enhances dectin-1 expression and induces dectin-1-dependent reactive oxygen species (ROS) generation. Immunobiology (2009) 214(4):321–30. doi:10.1016/j.imbio.2008.09.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
183. Sundstrom JB, Hair GA, Ansari AA, Secor WE, Gilfillan AM, Metcalfe DD, et al. IgE-FcepsilonRI interactions determine HIV coreceptor usage and susceptibility to infection during ontogeny of mast cells. J Immunol (2009) 182(10):6401–9. doi:10.4049/jimmunol.0801481
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
184. Kulka M, Alexopoulou L, Flavell RA, Metcalfe DD. Activation of mast cells by double-stranded RNA: evidence for activation through Toll-like receptor 3. J Allergy Clin Immunol (2004) 114(1):174–82. doi:10.1016/j.jaci.2004.03.049
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
185. King CA, Marshall JS, Alshurafa H, Anderson R. Release of vasoactive cytokines by antibody-enhanced dengue virus infection of a human mast cell/basophil line. J Virol (2000) 74(15):7146–50. doi:10.1128/JVI.74.15.7146-7150.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
186. McCurdy JD, Lin TJ, Marshall JS. Toll-like receptor 4-mediated activation of murine mast cells. J Leukoc Biol (2001) 70(6):977–84. doi:10.1152/physrev.00031.2002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
187. Smrz D, Bandara G, Beaven MA, Metcalfe DD, Gilfillan AM. Prevention of F-actin assembly switches the response to SCF from chemotaxis to degranulation in human mast cells. Eur J Immunol (2013) 43(7):1873–82. doi:10.1002/eji.201243214
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
188. MacNeil AJ, Junkins RD, Wu Z, Lin TJ. Stem cell factor induces AP-1-dependent mast cell IL-6 production via MAPK kinase 3 activity. J Leukoc Biol (2014) 95(6):903–15. doi:10.1189/jlb.0713401
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
189. Kojima R, Ohno T, Iikura M, Niki T, Hirashima M, Iwaya K, et al. Galectin-9 enhances cytokine secretion, but suppresses survival and degranulation, in human mast cell line. PLoS One (2014) 9(1):e86106. doi:10.1371/journal.pone.0086106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
190. Erdei A, Andrasfalvy M, Peterfy H, Toth G, Pecht I. Regulation of mast cell activation by complement-derived peptides. Immunol Lett (2004) 92(1-2):39–42. doi:10.1016/j.imlet.2003.11.019
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
191. Hartmann K, Henz BM, Kruger-Krasagakes S, Kohl J, Burger R, Guhl S, et al. C3a and C5a stimulate chemotaxis of human mast cells. Blood (1997) 89(8):2863–70.
192. El-Lati SG, Dahinden CA, Church MK. Complement peptides C3a- and C5a-induced mediator release from dissociated human skin mast cells. J Invest Dermatol (1994) 102(5):803–6. doi:10.1111/1523-1747.ep12378589
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
193. Mendes MA, Palma MS. Two new bradykinin-related peptides from the venom of the social wasp Protopolybia exigua (Saussure). Peptides (2006) 27(11):2632–9. doi:10.1016/j.peptides.2006.05.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
194. Hochman DJ, Collaco CR, Brooks EG. Acrolein induction of oxidative stress and degranulation in mast cells. Environ Toxicol (2014) 29(8):908–15. doi:10.1002/tox.21818
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
195. Pignatti P, Frossi B, Pala G, Negri S, Oman H, Perfetti L, et al. Oxidative activity of ammonium persulfate salt on mast cells and basophils: implication in hairdressers’ asthma. Int Arch Allergy Immunol (2013) 160(4):409–19. doi:10.1159/000343020
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
196. Sick E, Brehin S, Andre P, Coupin G, Landry Y, Takeda K, et al. Advanced glycation end products (AGEs) activate mast cells. Br J Pharmacol (2010) 161(2):442–55. doi:10.1111/j.1476-5381.2010.00905.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
197. Iddamalgoda A, Le QT, Ito K, Tanaka K, Kojima H, Kido H. Mast cell tryptase and photoaging: possible involvement in the degradation of extra cellular matrix and basement membrane proteins. Arch Dermatol Res (2008) 300(Suppl 1):S69–76. doi:10.1007/s00403-007-0806-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
198. Collaco CR, Hochman DJ, Goldblum RM, Brooks EG. Effect of sodium sulfite on mast cell degranulation and oxidant stress. Ann Allergy Asthma Immunol (2006) 96(4):550–6. doi:10.1016/S1081-1206(10)63549-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
199. Yang W, Lee S, Lee J, Bae Y, Kim D. Silver nanoparticle-induced degranulation observed with quantitative phase microscopy. J Biomed Opt (2010) 15(4):045005. doi:10.1117/1.3470104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
200. Wang Y, Chen Z, Ba T, Pu J, Chen T, Song Y, et al. Susceptibility of young and adult rats to the oral toxicity of titanium dioxide nanoparticles. Small (2013) 9(9-10):1742–52. doi:10.1002/smll.201201185
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
201. Swindle EJ, Coleman JW, DeLeo FR, Metcalfe DD. FcepsilonRI- and Fcgamma receptor-mediated production of reactive oxygen species by mast cells is lipoxygenase- and cyclooxygenase-dependent and NADPH oxidase-independent. J Immunol (2007) 179(10):7059–71. doi:10.4049/jimmunol.179.10.7059
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
202. Wolfreys K, Oliveira DB. Alterations in intracellular reactive oxygen species generation and redox potential modulate mast cell function. Eur J Immunol (1997) 27(1):297–306. doi:10.1002/eji.1830270143
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
203. Komiyama H, Miyake K, Asai K, Mizuno K, Shimada T. Cyclical mechanical stretch enhances degranulation and IL-4 secretion in RBL-2H3 mast cells. Cell Biochem Funct (2014) 32(1):70–6. doi:10.1002/cbf.2973
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
204. Soule BP, Brown JM, Kushnir-Sukhov NM, Simone NL, Mitchell JB, Metcalfe DD. Effects of gamma radiation on FcepsilonRI and TLR-mediated mast cell activation. J Immunol (2007) 179(5):3276–86. doi:10.4049/jimmunol.179.5.3276
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
205. Cabeza JM, Acosta J, Ales E. Mechanisms of granule membrane recapture following exocytosis in intact mast cells. J Biol Chem (2013) 288(28):20293–305. doi:10.1074/jbc.M113.459065
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
206. Anlauf M, Schafer MK, Schwark T, von Wurmb-Schwark N, Brand V, Sipos B, et al. Vesicular monoamine transporter 2 (VMAT2) expression in hematopoietic cells and in patients with systemic mastocytosis. J Histochem Cytochem (2006) 54(2):201–13. doi:10.1369/jhc.5A6739.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
207. Garcia-Faroldi G, Melo FR, Bruemmer D, Conneely OM, Pejler G, Lundequist A. Nuclear receptor 4a3 (nr4a3) regulates murine mast cell responses and granule content. PLoS One (2014) 9(2):2. doi:10.1371/journal.pone.0089311
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
208. Borges R, Dominguez N, Estevez-Herrera J, Pereda D, Machado JD. Vesicular Ca2+ mediates granule motion and exocytosis. Cell Calcium (2012) 51(3-4):338–41. doi:10.1016/j.ceca.2011.12.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
209. Bonnemaison M, Back N, Lin Y, Bonifacino JS, Mains R, Eipper B. AP-1A controls secretory granule biogenesis and trafficking of membrane secretory granule proteins. Traffic (2014) 15(10):1099–121. doi:10.1111/tra.12194
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
210. Bultema JJ, Di Pietro SM. Cell type-specific Rab32 and Rab38 cooperate with the ubiquitous lysosome biogenesis machinery to synthesize specialized lysosome-related organelles. Small GTPases (2013) 4(1):16–21. doi:10.4161/sgtp.22349
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: granule, lipid body, exosome, lysosome, exocytosis, secretion
Citation: Moon TC, Befus AD and Kulka M (2014) Mast cell mediators: their differential release and the secretory pathways involved. Front. Immunol. 5:569. doi: 10.3389/fimmu.2014.00569
Received: 17 September 2014; Accepted: 23 October 2014;
Published online: 14 November 2014.
Edited by:
Paige Lacy, University of Alberta, CanadaReviewed by:
Frank A. Redegeld, Utrecht University, NetherlandsAxel Lorentz, University of Hohenheim, Germany
Copyright: © 2014 Moon, Befus and Kulka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marianna Kulka, National Institute for Nanotechnology, National Research Council, 11421 Saskatchewan Drive, Edmonton, AB T6G 2M9, Canada e-mail: marianna.kulka@nrc-cnrc.gc.ca