 Sergio Rosales-Mendoza
Sergio Rosales-Mendoza Ricardo Nieto-Gómez
Ricardo Nieto-Gómez Carlos Angulo
Carlos Angulo- 1Laboratorio de Biofarmacéuticos Recombinantes, Facultad de Ciencias Químicas, Universidad Autónoma de San Luis Potosí, San Luis Potosí, San Luis Potosí, Mexico
- 2Grupo de Inmunología & Vacunología, Centro de Investigaciones Biológicas del Noroeste, SC., La Paz, Baja California Sur, Mexico
The Ebola virus (EBOV) epidemic indicated a great need for prophylactic and therapeutic strategies. The use of plants for the production of biopharmaceuticals is a concept being adopted by the pharmaceutical industry, with an enzyme for human use currently commercialized since 2012 and some plant-based vaccines close to being commercialized. Although plant-based antibodies against EBOV are under clinical evaluation, the development of plant-based vaccines against EBOV essentially remains an unexplored area. The current technologies for the production of plant-based vaccines include stable nuclear expression, transient expression mediated by viral vectors, and chloroplast expression. Specific perspectives on how these technologies can be applied for developing anti-EBOV vaccines are provided, including possibilities for the design of immunogens as well as the potential of the distinct expression modalities to produce the most relevant EBOV antigens in plants considering yields, posttranslational modifications, production time, and downstream processing.
Introduction
The last Zaire Ebola virus (EBOV) epidemic outbreak in Guinea, which began in December 2013, quickly spread and six West-African countries were greatly affected (Guinea, Liberia, Sierra Leone, Mali, Nigeria, and Senegal). There have also been reports of cases within health-care workers from the USA, Spain, and the United Kingdom. Fortunately, the overall case incidence has dropped, and no reports on confirmed cases during the last week of December 2015 were generated. Nonetheless, according to a report on December 27, 2015, there have been 25,637 confirmed, probable, or suspected cases of EBOV disease (EVD) in Guinea, Liberia, and Sierra Leone (Figure 1), with over 11,000 reported deaths, which surpasses all previous EBOV outbreaks combined (World Health Organization1). Therefore, the EBOV constitutes an imminent and serious threat to public health, as well as a potential bioterrorism agent (1). EBOV represents one of the three genera composed of the family Filoviridae (2). The EBOV genus comprises five species: (1) Sudan ebolavirus (SUDV), (2) Zaire ebolavirus (ZEBOV), (3) Côte d’Ivoire ebolavirus (also known as Ivory Coast ebolavirus or Tai Forest ebolavirus, TAFV), (4) Reston ebolavirus (RESTV), and (5) Bundibugyo ebolavirus. All of these species, with the exception of the RESTV, have shown to cause disease in human beings (3, 4). After an incubation period of 3–21 days, the EVD generally progresses quickly, with symptoms of fever, diarrhea, vomiting, systemic inflammatory response syndrome, organ dysfunction, and hemorrhagic manifestations that end in death (5).
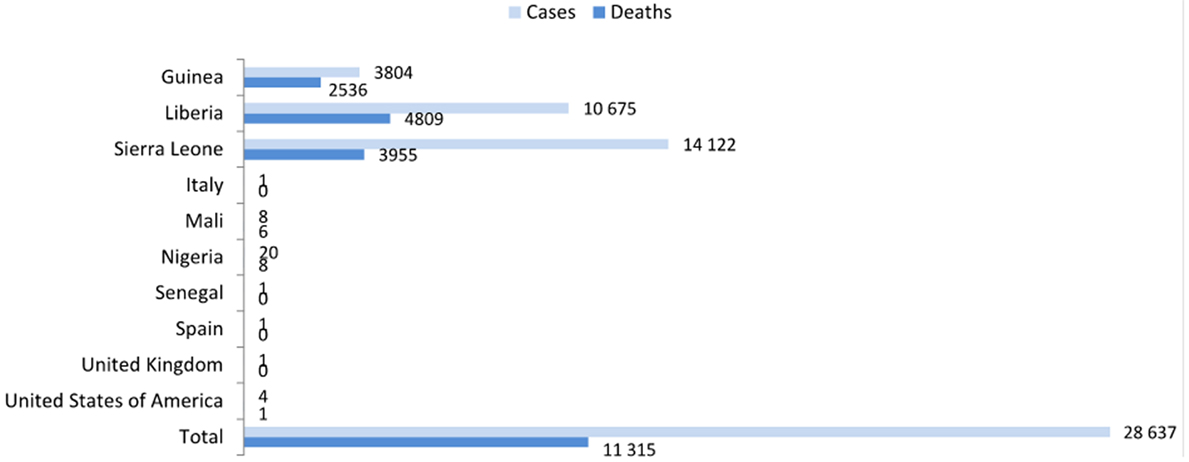
Figure 1. Confirmed, probable, and suspected EBOV disease cases worldwide (data up to 27 December 2015; report of December 30 from the World Health Organization, http://www.who.int/en/).
Despite the substantial efforts made to develop rational prophylactic and chemotherapeutic interventions, no licensed countermeasures are available for the treatment of EVD as of now. EBOV is introduced into the human population through close contact with bodily fluids of infected animals such as primates and fruit bats. EBOV then spreads through human-to-human transmission via direct contact (through broken skin or mucous membranes) with bodily fluids of infected people. Therefore, most efficient measures to control the EVD spread consist of the isolation of patients establishing strict barrier nursing procedures to protect health-care workers (5). Looking at this situation, the development of effective therapeutics for the prevention and treatment of EBOV infections is urgently needed. In the case of immunotherapies, achieving broad and long-lasting humoral immunity at the mucosa and systemic levels against many EBOV species as possible is a key goal (6). The most advanced immunotherapy against EVD is ZMapp (Mapp Biopharmaceutical, San Diego, CA, USA), a drug consisting of humanized monoclonal antibodies (mAbs) capable of neutralizing the EBOV. This treatment, based on passive immunity, has been successful in non-human primates (NHPs) and efforts for its licensing and introduction into the market are ongoing (7). ZMapp has already been used on a compassionate basis to treat a few patients of EVD; however, the clinical efficacy of this specific cocktail as a treatment of EVD in humans remains uncertain (8). Vaccination is the ideal approach to fight this disease since prophylaxis could be achieved through the administration of a minimum number of doses. Vaccinology offers a myriad of possibilities for the development of vaccines against EBOV, and according to the ClinicalTrials.gov database,2 47 studies of Ebola vaccine trials have been registered. One of the biggest challenges in achieving global vaccination is developing production platforms accessible to developing countries. For instance, protein subunit vaccines are obtained, distributed, and administered through processes requiring complex downstream steps, cold chain, and delivery systems that involve specialized personnel and equipment. All of these aspects hamper vaccination availability and usage in developing countries. Therefore, the next-generation platforms for vaccine production, distribution, and delivery have been proposed to develop low-cost and broad coverage vaccination strategies. In this context, plant-based platforms constitute an attractive technology with the following attributes: (i) since the use of sophisticated bioreactors and complex downstream processing are avoided, the cost of a plant-derived product is 10–50 times lower than products derived from the fermentation with Escherichia coli (9) and 140 times lower when compared to baculovirus-infected insect cells (10); (ii) high biosynthetic capacity derived from a machinery that performs folding, assembly, and glycosylation; (iii) the plant systems offer high safety in the sense that they are not hosts of human or animal pathogens, in contrast to mammalian-based production systems where the risk of contamination with viruses and prions exists. Moreover, many plant tissues and fruits are safe for human consumption and thus can be used as oral delivery vehicles for vaccines, thereby avoiding the purification and processing required for conventional injectable vaccines. Therefore, plant-made oral vaccines can be easily formulated with freeze-dried plant material, which not only increases antigen concentration but also produces a material stable at room temperature avoiding the cold chain maintenance required for other delivery systems (11). This perspective constitutes the ideal case for vaccine development, and it has been consolidated in recent years with the successful delivery of many vaccines and other biopharmaceuticals by the oral route in test animals (12–14). The technology of plant-based vaccines and the current advances have been recently reviewed by distinct groups (15–17). During the last years, clinical trials have been conducted to evaluate the immunogenicity and safety of influenza virus vaccines with positive outcomes (18–21), which has stimulated the interest of the pharmaceutical industry in these platforms (Table 1).
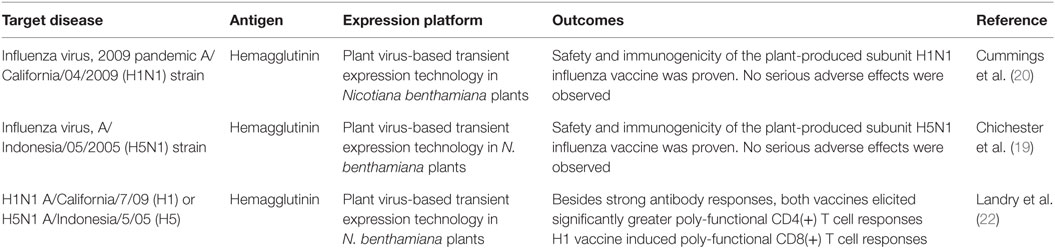
Table 1. Evaluations of plant-made vaccines in clinical trials reported over the last years.
In this review, the use of the technology of plant-based vaccines to develop attractive EBOV vaccines is placed in perspective. After describing the molecular approaches to express antigens in the plant cell, the relevant aspects of EBOV as well as conventional vaccines under development were are summarized; finally, the perspectives on how plant systems may lead to EBOV vaccines are identified and discussed.
Current Experimental Vaccines to Fight EBOV
While the precise mechanisms for immune protection against the EBOV infection are likely complex, it is noteworthy that vaccination against the EBOV surface glycoprotein (GP) is both necessary and sufficient for protection against virus infection as has been evidenced by several successful vaccination approaches (23, 24). This evidence suggests an important role of the GP in virus survival within the host. Several studies have pointed out that the humoral responses induced by these vaccines are strongly associated with protection (25–27), although some reports have clearly demonstrated that the cellular response aided in infection clearance as well (28).
The most advanced vaccines against EBOV are based on the viral GP that has demonstrated protection against EBOV in NHPs. It is important to point out that in 2002, the US Food and Drug Administration introduced the “animal rule” concept that aims to facilitate the licensing of vaccine or drug treatments against infection by the EBOV as well as other highly lethal human pathogens for which the efficacy evaluation in human beings would be unethical and field trials unreasonable (29). The application of the “animal rule” allows for the approval of any EBOV vaccine candidate based on efficacy testing in animal models, with defined immune correlates of protection, as well as Phase I and II clinical trials for safety and immunogenicity testing in human beings. Therefore, the development of animal models is critical for the evaluation and eventual approval of EBOV vaccine candidates. Promising animal models for investigating EBOV vaccines include Guinea pig (30), mouse (31), Syrian Golden hamster (32), marmoset (33), and ferret (34, 35). However, NHP models of EBOV infection, especially the model of cynomolgus macaques, have a stronger predictive value for human diseases and immune protection, and thus, it is the preferred model for EBOV vaccine development (24). Therefore, this review will focus on the most advanced vaccines that have been tested in NHPs and clinical trials. New promising vaccine candidates evaluated using other animal models will be mentioned briefly.
Overall, the candidate vaccines against the EBOV developed thus far can be divided into three categories: non-replicative expressing vector-based vaccines, replication-competent viral vector-based vaccines, and viral antigen-based vaccines (36). Most of the successful vaccines against the EVD rely on viral vectors in whose genome the EBOV GP gene was introduced (37). The vector-based vaccines have been evaluated in NHP and in clinical trials, whose outcomes are summarized in the following sections. Viruses used as vaccine vectors include vesicular stomatitis virus (VSV) (38), recombinant adenovirus replicons (39), recombinant parainfluenza virus (40), recombinant rabies virus (RABV) (41), and recombinant Venezuelan equine encephalitis virus (VEEV) (42). Protein-based vaccines such as virus-like particles (VLPs) have also demonstrated EVD protection in NHPs, but the characterization of most of the candidates has been performed in small animal models (43, 44). A general overview on the progress achieved for each type of vaccine is described in the following sections.
Non-Replicative Vector-Based Vaccines
Sullivan et al. (45) reported the first proof of concept on protection against EBOV infection by vaccination. The study revealed that priming with an EBOV GP DNA vaccine followed by boosting with a recombinant adenovirus-5 replicon expressing GP conferred complete protection against a lethal EBOV challenge in NHPs. Although promising and safe for human beings, the use of the most advanced adenovirus 5 replicon-based vaccine faces the problem of pre-existing immunity against the viral vector as well as a relatively low immunogenicity in human beings, since the antibody titers against GP were less than 300, while titers of 2,000 are associated with the protection of NHPs (46). An interesting alternative that may solve the problem derived from the pre-existing immunity against the vaccination vector consists in the use of Chimpanzee adenovirus-based vaccines (47).
Another encouraging example is a VEEV replicon that has been employed for EBOV vaccine development. VEEV replicons expressing EBOV GP and Sudan ebolavirus (SUDV) GP protected NHPs against a lethal EBOV as well as SUDV challenge when administered a single [1 × 1010 focus-forming units (FFU)] simultaneous intramuscular vaccination (42). However, similar to adenovirus replicons, the requirement of high vaccine doses and a pre-existing immunity to VEEV will likely be the major obstacles for human application of this kind of vaccine. A mutant form of the EBOV, without the VP30 gene that is required for virus replication, was evaluated in mice and guinea pigs, and it was shown to confer complete protection against a lethal EBOV challenge after two immunizations (48). Moreover, the efficacy of this new replication-defective viral vector-based vaccine was also confirmed to confer immunoprotection in NHPs when administered by the intraperitoneal route twice at 3-week intervals with 1 × 106 FFU of Ebola ΔVP30 virus. However, this approach raised concerns with respect to virulence reversion, and thus, a new version of the vaccine consisted of the virus inactivated with hydrogen peroxide was generated, which remained antigenic and protective in NHPs when administered intramuscularly (1 × 107 FFU) one or two times with a 4-week interval (49).
Two replication-incompetent vectored vaccines have reached Phase III clinical trials: human adenovirus serotype 26 (Ad26) expressing the Ebola virus Mayinga variant GP (Ad26.ZEBOV) and Modified Vaccinia Virus Ankara-Bavarian Nordic Filo-vector (MVA-BN-Filo). Remarkably, Ad26.ZEBOV and MVA-BN-Filo vaccines resulted in sustained elevation of specific immunity, and no vaccine-related serious adverse events were observed in Phase I clinical trial. In this evaluation, the vaccinated (i.m.) groups were (1) with MVA-BN-Filo as prime vaccine on day 1 boosted by Ad26.ZEBOV on day 29 or day 57; and (2) with a priming dose of Ad26.ZEBOV boosted by MVA-BN-Filo on day 29 or day 57 (50). Therefore, Phases II and III were pursued. Moreover, the Phase IV, named “Long-term Safety Follow-up of Participants Exposed to the Candidate Ebola Vaccines Ad26.ZEBOV and/or MVA-BN-Filo” is active but not open for participant recruitment yet.
Replication-Competent Viral Vector-Based Vaccines
This category includes rhabdovirus-based viral vectors, including the VSV and RABV, and paramyxovirus-based vectors such as recombinant human parainfluenza virus 3 (HPIV3) expressing EBOV GP separately or in combination with nucleoprotein (NP). The potential of this kind of vaccine platform was shown when the recombinant VSV expressing the GPs of ZEBOV (strain Mayinga) was generated using the infectious clone for the VSV Indiana serotype. A single intramuscular immunization, measured in plaque-forming units (PFU) of the virus particles, of cynomolgus macaques (1 × 107 PFU) demonstrated to protect NHPs against a lethal challenge (1 × 103 PFU) of ZEBOV (strain Kikwit) isolated from a patient from the 1995 EBOV outbreak in Kikwit (38). Similarly, Marzi et al. (51) found complete protection of NHPs against ZEBOV (strain Makona) following the administration of a single dose given as late as 7 days before challenge in VSV–EBOV GP vaccinated animals. Looking to explore practical delivery routes, effective protection of NHPs was observed when the vaccine was administered either orally or intranasally with the subsequent EBOV challenge (52, 53). These findings opened the path to explore mucosal vaccination as a feasible strategy in combating the EVD. Furthermore, this vaccine platform showed potential as an early treatment since it induced beneficial effects in NHPs infected with the EBOV (54) and in individuals who have experienced incidental exposure or high-risk occupational exposure to the EBOV such as a needle stick handling (55, 56). Interestingly, a single intramuscular immunization (1 × 107 PFU in the caudal thigh) of the full-length parent RABV vaccine expressing the EBOV GP also conferred complete protection in rhesus macaques after a challenge with 1,000 PFU of the EBOV (strain Mayinga). However, its potency was lower when compared to recombinant VSV-based vaccines (41), such as the attenuated vesiculovax recombinant VSV-based vaccines expressing the EBOV GP, which protects macaques from a lethal challenge after a single dose (57). Another vaccine platform uses a paramyxovirus-based vector, such as the recombinant HPIV3 expressing EBOV GP alone or in combination with NP. These vaccines were constructed by inserting a transcription cassette encoding the EBOV (Mayinga strain) GP gene between the HPIV3 P and M genes alone or in combination with a cassette encoding the NP inserted between the HPIV3 HN and L genes. Rhesus monkeys were protected against the EBOV infection after receiving two doses of 2 × 107 tissue culture infectious dose (TCID50) (days 0 and 28) of combined intranasal and intratracheal inoculation and an intraperitoneally challenge on day 67 (39 days following the second vaccine dose) with 1,000 PFU of the EBOV (Zaire species, Mayinga strain) (40). This study reinforces the practical feasibility of immunization against the EVD via the respiratory tract (58). However, since these vaccine platforms are replication competent, their side effects for human vaccination is a major concern and merits further research. Recently, Phase I and II clinical trials have been conducted, and the results showed that rVSV-ZEBOV is immunogenic but also mild to moderate reactogenic. rVSV-ZEBOV used at 1–5 × 107 PFU (Phase I) provoke fever (25%) and oligoarthritis (22%) in vaccinated volunteers (6). A reduced dose of 3 × 105 (Phase II) PFU decreases viremia and reactogenicity but also antibody response levels without reducing the risk of vaccine-induced side effects (59). Remarkably, a Phase III trial in Guinea highlighted that the rVSV-ZEBOV is highly efficacious when administered in a single 2 × 107 PFU dose (estimated vaccine efficacy of 100%) and safe in preventing the EVD, while the assessment of vaccine-derived adverse events revealed promising outcomes (2 serious adverse events in 5,837 vaccinees) (60, 61).
Viral Protein/DNA-Based Vaccines
Konduru et al. (62) provided the first proof of concept that a subunit vaccine based on purified GP could elicit protective immune responses against the EBOV. In their study, a ZEBOV GP-Fc fusion protein was constructed coding for the C-terminal end (1–637 aa) of the extracellular domain from the ZEBOV GP (Mayinga strain) and the crystallizable fragment (Fc) from human IgG1. The ZEBOV GP-Fc protein fusion was produced in transfected Chinese hamster ovary cells. C57BL/6 mice were intraperitoneally vaccinated (i.p.) with 100 μg of purified ZEBOVGP-Fc in complete Freund’s adjuvant and boosts (25 μg in incomplete Freund’s adjuvant) were administered at 21, 45, and 60 days post-priming. A 90% of protection in the vaccinated mice was achieved after a lethal challenge by i.p. injection with 1,000 PFU of mouse-adapted ZEBOV. Similar results were obtained by Phoolcharoen et al. (63) in which the GP was fused to a mAb that recognizes an epitope in the GP, resulting in the production of EBOV immune complexes (EICs). Remarkably, the EICs were produced in Nicotiana benthamiana plants by transient expression. The purified EICs were tested in mice, administered by the subcutaneous route four times on days 0, 21, 42, and 63, and the immunogenic properties determined. Although antigen–antibody immune complexes were efficiently processed and presented to immune effector cells, they found that co-delivery of the EIC with toll-like receptor (TLR) agonists elicited a more robust antibody response in mice than the EICs alone. Among the compounds tested, polyinosinic:polycytidylic acid (poly I:C, a TLR-3 agonist) was highly effective as an adjuvant. After vaccinating mice with EIC plus poly I:C, 80% of the animals were protected against a lethal challenge with live EBOV. These results are encouraging but further research is needed to optimize the immunogenicity of this vaccine and test its efficacy in NHP models with the subsequent determination of its safety in clinical trials. Another viral antigen-based vaccine strategy is the use of VLP that can direct the target antigen to antigen presenting cells, such as dendritic cells, stimulating antibody, and cellular immune responses. Interestingly, three i.m. immunizations at 42-day intervals with enveloped EBOV VLPs containing the EBOV GP, NP, and VP40 matrix protein, along with RIBI adjuvant, conferred protection to NHPs against a lethal challenge with EBOV, thus providing the first evidence that protective immunity can be elicited by non-viral vector-based vaccines in NHPs (43). Moreover, the versatility of VLPs should be noted that they can either be used as carriers of immune-stimulating molecules or enriched with chimeric EBOV GP carrying additional epitopes as an approach to enhance immune responses.
It is also important to note that the results on three DNA vaccines (INO-4201, -4202, and -4212) and one recombinant protein subunit vaccine (EBOV GP1,2 with Matrix-M) have not been published yet and probably will bring new perspectives in the race of developing new Ebola vaccines (64). DNA vaccines expressing the EBOV GP have also been tested in human beings during Phase I clinical trials with safe and immunogenic properties when applied under a scheme comprising three i.m. doses (2, 4, and 8 mg) on days 0, 28, and 56 (65) and an homologous boost (2 mg) at week 32 or after (66).
Perspectives for EBOV Vaccine Development
Despite the milestone of establishing durable protection against the EBOV, future developments are required to increase qualitative or quantitative resolution of the protective and non-protective humoral immune responses (67). Two encouraging vaccines based on GP have been evaluated under Phase I and Phase II clinical trials (Table 2) showing durable protection in the cynomolgus macaque model (47, 68, 69). Based on promising data from the initial clinical trials, gathered in the late 2014, the WHO in combination with the Health Ministry of Guinea, Médecins Sans Frontières from Epicentre, and The Norwegian Institute of Public Health launched a Phase III trial in Guinea on March 7, 2015. This trial tested the VSV–EBOV (VSVΔG-ZEBOV-GP) vaccine for efficacy and effectiveness in preventing the EVD (60). The results indicated that the vaccine is highly efficacious and safe, and likely effective in the population when delivered during an EVD outbreak via a ring vaccination strategy (60). In addition, the plan includes testing another advanced vaccine called ChAd3 (ChAd3-ZEBOV-GP; GSK). The follow-up study to compare the safety and efficacy of the ChAd3 Ebola Zaire and VSVΔG-ZEBOV-GP virus vaccines through Phase II/III clinical trials in volunteers from Liberia led to promising results upon the first 4 months, and serious adverse effects were not reported (24, 64, 70). ChAd3 is an example that the current EBOV vaccines require cell-based production and storage at low temperature, thereby creating obstacles in scalable manufacturing and shelf-life in developing countries (67).
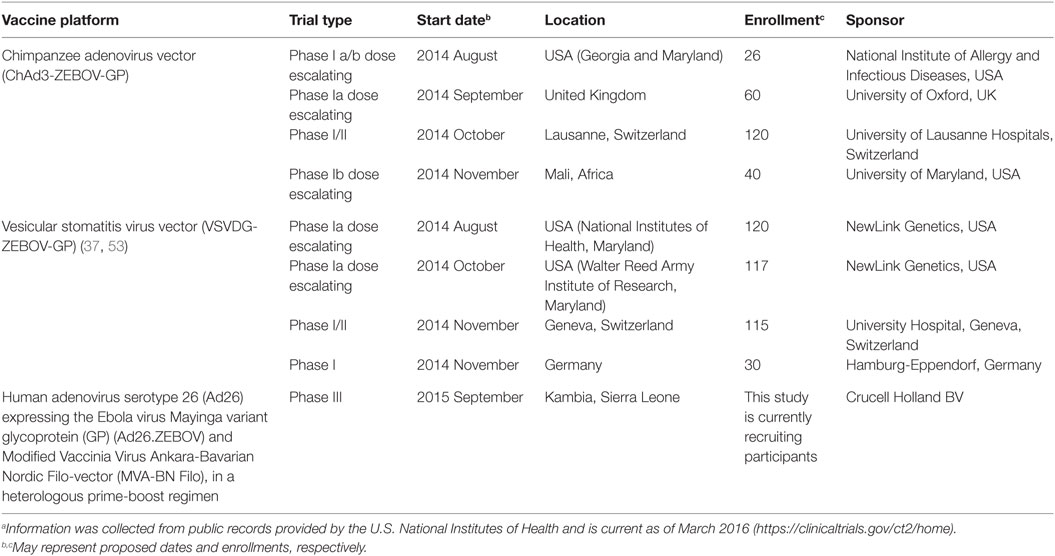
Table 2. Current EBOV Food and Drug Administration-approved vaccine trials.a
Overall, non-replicative vector-based vaccines face the problem of pre-existing immunity and/or the induction of anti-vector immune responses that may decrease their efficacy, while viral replication-competent vaccines face important human safety or adverse side effects concerns. By contrast, the vaccine strategies based on viral protein antigens are not affected by those issues. In this context, plant-made vaccines can be a reasonable alternative in the fight against the EVD.
How Could EBOV Plant-Based Vaccines be Developed?
The key steps involved in the development of plant-made vaccine prototypes include the following: design putative functional immunogens and develop genetically engineered plants expressing the antigen or establishing viral vector-based platforms for transient expression, estimate yields and antigenic properties of the target antigen, assess the immunogenic potential of the candidate vaccine in test animals in terms of protective immunity and safety, and perform clinical trials once preclinical studies have provided acceptable outcomes (Figure 2).

Figure 2. Results from the in silico epitope analysis of the African Zaire ebolavirus (ZEBOV) spike glycoprotein sequence (GenBank: AIE11809). Regions in red indicate the epitopes reported by Becquart et al. (71), based on reactivity with sera collected from human survivors as an indication of the induction of neutralizing humoral responses. Regions in yellow indicate the epitopes reported by Vaughan et al. (72) as EBOV-related B-cell epitopes found in the Immune Epitope Database. Regions in blue indicate conserved regions of ZEBOV for the African continent overlapping with the epitopes reported in both articles. Regions in green indicate matches of the conserved regions found in the bioinformatics analysis and the epitopes reported in the aforementioned articles.
Possibilities for the Design of Immunogens
A successful proof of concept on EBOV plant-based vaccines will include the design and production of full-length viral proteins such as GP, matrix viral protein (VP40), and NP antigens, as well as chimeric proteins carrying conserved protective epitopes capable of inducing anti-EBOV neutralizing antibodies. Examples of the latter approach include the following linear epitopes: EQHHRRTDN, VIKLDISEA, and LITNTIAGV (25). Hopefully, the current knowledge on the protective EBOV sequences as well as the technologies to produce heterologous proteins in plant cells will accelerate the development of plant-made vaccine candidates against EBOV.
Regarding epitope vaccines, Wilson et al. (25) reported a GP epitope that is conserved among all Ebola viruses demonstrating that a specific mAb was able to protect mice from a lethal EBOV infection. Subsequently, it was found that although some EBOV GP epitopes induce an antibody-dependent enhancement of EBOV infection, antibodies against other specific EBOV GP epitopes were required to control an EBOV infection (73). Another recent study demonstrated that a linkage region (aa 393–556) of the GP (called MFL) contains a furin cleavage site and an internal fusion loop responsible for important viral functions (74). This region was the major contributor to immunogenicity in terms of the induction of humoral immune responses and neutralizing antibodies against the EBOV (75). Interestingly, the study by Becquart et al. (71), using sera from infected patients, identified specific B-cell epitopes in four EBOV proteins [GP, NP, and matrix viral protein (VP40 and VP35)]. Among them, the specific immunodominant VP40 and GP epitopes were detected by IgG antibodies from asymptomatic individuals and symptomatic Gabonese EBOV infected survivors, respectively. These findings strongly suggest that an effective epitopic subunit vaccine should induce humoral IgG responses targeting specific GP and VP40 epitopes. One interesting approach in the design of an epitope-based vaccine capable of triggering protective immune responses is the use of immunoinformatic tools. In this regard, the potential of inducing both humoral and cell-mediated immunity by T and B cells against the EBOV epitopes was recently assessed by Khan et al. (76). From the complete proteomes of EBOVs, the amino acid sequences were retrieved using UniProt Knowledge Base and bioinformatic analyses were conducted to study antigenicity, solvent-accessible regions, surface accessibility, flexibility, MHC class-I-binding epitopes (cellular immune response), and B-cell-binding epitopes (antibody immune response) from those proteins. The in silico capability of each protein sequence to initiate an immune response allowed for the identification of the most promissory L protein comprise of 128 amino acids, which is also known as RNA-dependent RNA polymerase. This protein reached the highest antigenicity score in VaxiJen analysis among all the query proteins. The downstream bioinformatic analysis showed that the 9-mer epitope TLASIGTAF was the selected potential epitope-based vaccine candidate for inducing cytotoxic T cell immune responses by considering its overall epitope conservancy (76.60%), human population coverage (53–81%), and the affinity for highest number of MHC-I (HLA) molecules (n = 12). Similarly, the L protein was evaluated to identify B cell epitopes and the 9-mer epitope PEEQEQSAE (spanning region from 42 to 50 amino acids) of the L protein was the most potential B cell epitope to induce antibody-mediated immune responses. However, it should be considered that L protein is the last one expressed during viral replication, and thus, a vaccine targeting only this antigen may result in low efficacy. Therefore, vaccine design should contemplate a combination of L protein epitopes with those of early proteins, such as GP. Thus, the combination of experimental data with immunoinformatic prediction approaches opens up a new horizon to design effective multiepitopic vaccines able to induce protective antibody immune responses against the EBOV. In fact, the Immune Epitope Database and Analysis Resource3 has reported an integrative immunopredictive and experimental analysis for “functional epitopes.” These epitopes are identified using assays that demonstrate their potential to induce positive outcomes when virus neutralization assays or challenge experiments are performed. A high percentage of the selected epitopes were from the GP (55%) and NP (33%) proteins. The functional EBOV-related B cell epitopes were only found in these two proteins (72). On the other hand, an in silico analysis to identify EBOV conserved sequences among the EBOV variants, matching with the abovementioned functional analysis, has allowed the identification of a set of promising GP Zaire EBOV B-cell epitopes comprising the following sequence: NISGQSPARTSSDPE, NTPVYKLDISEATQVGQHHRRAD, and TAGNNNTHHQDTGEE SASSGKLGLITNTI AGVAGLITGGRRTR. These sequences are considered promising candidates for multiepitopic vaccine design (Figure 3).
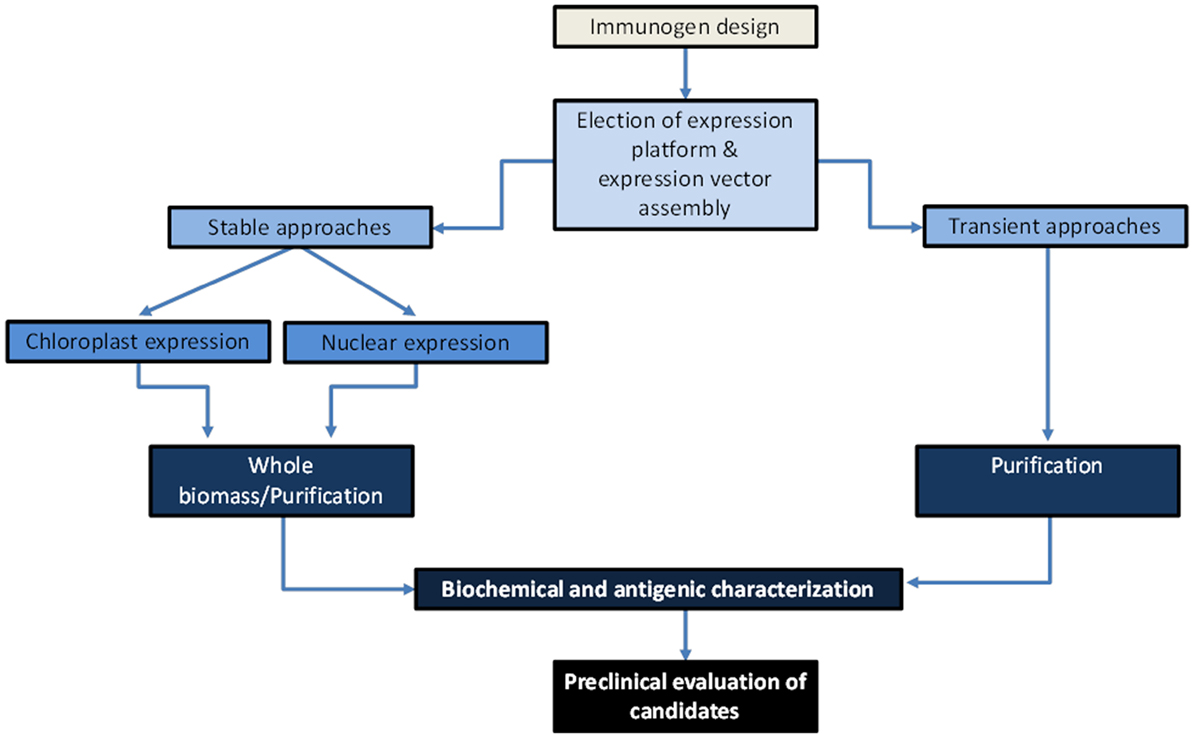
Figure 3. Scheme on the path for development of Ebola virus plant-based vaccine candidates. Antigens will be designed to serve as strong mucosal immunogens, and coding genes will be assembled into expression vectors elected according to the expression approach to be assessed. Antigen production can be achieved transiently through strategies of chimeric virus (first-generation vectors) or deconstructed virus (second-generation vectors, e.g., agroinfiltration with viral pro-vectors), or stably through a nuclear transformation approach (transformation mediated by Agrobacterium or physical methods) or chloroplast transformation approach (transformation mediated by physical methods). A subsequent characterization of the plant-made antigens will comprise estimating antigen yields and antigenic properties. During preclinical trials, it is envisioned that transient approaches will serve as a high productive platform that will render parenteral vaccines after a purification process, which are ideal as prime doses, while stable transformed lines from edible crops may serve as low-cost oral vaccines formulated with freeze-dried plant biomass.
Since vaccines administered through mucosal membranes, mainly by oral route, are the most convenient approach for mass vaccination, the developments in this direction are a priority. However, epitopes are not good immunogens per se and thus must be coupled to carrier proteins or adjuvant sequences that favor uptake and efficient antigen presentation. Antigen uptake at the mucosa can be aided by the use of transmucosal carriers, such as the B subunits from either the cholera toxin (CTB) or the enterotoxigenic E. coli heat-labile toxin (LTB). These proteins produce oligomeric structures that bind the GM1 ganglioside on the surface of gut epithelial cells, mediating the translocation into the submucosal compartment where the antigen can be processed by dendritic cells with the subsequent induction of adaptive immune responses (77). These properties enable both CTB and LTB to be highly immunogenic and serve as effective carrier proteins and adjuvants for unrelated coupled antigens (78, 79). Therefore, the designed chimeric proteins, through genetic fusion, are proposed as candidates that could result in immunogens capable of inducing strong anti-EBOV antibody responses using oral immunization under the plant-based vaccine concept. This idea is also supported by the proven oral immunogenic activity of CTB- and LTB-based chimeric antigens produced in plants (79, 80). Therefore, specific EBOV epitopes in the form of CTB- or LTB-based chimeras could serve as candidates to induce immunoprotective humoral responses against the EBOV. Other strategies might include the design of chimeras comprising target epitopes and cell penetrating peptides, such as those derived from the HIV-1 Tat protein or the Drosophila melanogaster Antennapedia homeodomain (penetratin), which increase the cellular uptake of large molecules (81, 82).
Another attractive possibility in developing plant-based EBOV vaccines could be based on VLPs. It is well established that plants can synthetize structural viral proteins that self assemble into VLPs. These structures are macromolecular complexes that typically are highly immunogenic due to their complexity. Two types of VLPs can be produced: those based only on structural viral proteins and those based on envelope viral proteins associated to a membrane layer from the plant cell. VLPs derived from bluetongue virus, Norwalk virus, influenza virus, Hepatitis B virus (HBV) (nucleocapsid antigen), human papilloma virus, and rotavirus have been successfully assembled in plants (83). These viral proteins have also been engineered to display unrelated epitopes and thus serve, as in the case of CTB and LTB, as immunogenic carriers. This strategy has been successfully applied in a number of cases (80, 84).
Another alternative for immunogen design consists of recombinant immune complexes (RICs). RICs rely on the production of self-polymerized chimeras, whose monomeric form is comprised of the antigen of interest fused to the heavy chain of a mAb against the same antigen of interest. This has been found to be an effective strategy to increase antigen accumulation in transgenic plants enhancing immunogenicity (85).
Expression and Delivery Possibilities
High antigen yields will constitute a key factor in the flowchart to define the viability of the vaccination approach. In particular when oral vaccine development is pursued, high doses of antigen are typically required. If this aspect is addressed, the ambitious goal of developing oral vaccines will be greatly favored. Oral vaccines constitute the most attractive immunization approach since they offer easier and safer administration as well as the possibility of inducing mucosal and systemic immune responses. Although low expression levels were a limitation in the initial attempts at exploring the viability of plant-based vaccines, it is envisioned that the current optimized expression platforms will allow the production of the targeted antigens at acceptable yields to reach the required level in the plant biomass that could reasonably constitute an oral dose (86).
Each expression modality possesses particular advantages but at the same time imposes some limitations. Therefore, the selection of the expression platform should follow a case-by-case analysis contemplating the nature of the elected antigen, the delivery approach, and the required time response (Table 3). For instance, viral vector-based systems offer high yields, efficient production of complex glycosylated proteins, and the shortest production time among the plant-based platforms. However, since these processes are based in Nicotiana species and agroinfiltration, parenteral vaccines can only be produced after an extensive purification process to obtain an antigen free of bacterial compounds and toxic plant metabolites (87). Therefore, VLP-based EBOV vaccines using the GP or VP40 antigens can be ideally produced in viral vector-based platforms as a quick response to epidemics, parenterally immunizing the population at risk. However, it should be considered that these vaccines will not result in low-cost formulations and will require sterile devices and trained personnel for administration. In fact, there is one approved patent covering the production of EBOV VLPs in plants (88). A report by Phoolcharoen et al. (89), where a geminiviral vector was used for expression of the EIC in leaves of N. benthamiana, illustrates the potential for producing functional EBOV antigens at convenient yields.
Table 3. Identified expression options for specific EBOV immunogens using the available plant expression technologies.
In the case of transplastomic approaches, the average yields are lower than those of the viral expression vector but still considered convenient. However, it should be considered that the time for generating transplastomic lines is very long and no complex posttranslational modifications, such as glycosylation, occur in this organelle (90). Therefore, this platform is ideal for the production of epitope-based vaccines where no complex antigens requiring glycosylation are targeted. One attractive avenue consists in the use of edible plant species for which chloroplast transformation has been established. This is the case of lettuce, which was used for the production of some vaccines (91–93).
By contrast, stable nuclear expression also offers high biosynthetic capacity and propagation of cells in bioreactors. The time for generating transformed lines depends on the species but is generally shorter than that required for transplastomic approaches. Yields are in general modest but can be optimized using several approaches such as organelle targeting and formation of protein bodies (94). Interestingly, several edible plant species can be efficiently transformed. For instance, lettuce can be transformed efficiently using Agrobacterium tumefaciens (95). Another interesting species is the carrot (Daucus carota), for which there are efficient protocols for Agrobacterium-mediated transformation (96, 97). This host, D. carota, is relevant considering that the first plant-made biopharmaceutical for human use introduced into the market (Taliglucerase), which is a glucocerebrosidase for Gaucher’s disease treatment, was expressed in carrot cell cultures. This fact implies that the production processes and the regulatory framework are already in place for this system (98). In fact, the company that developed this process is also working on validating the oral delivery of a recombinant product using carrot cells (99).
Considerations for Plant-Based EBOV Vaccines in Preclinical Evaluations
The antigenic and immunogenic properties of the target immunogens should be evaluated through molecular and immunological analyses. At the same time, these techniques will allow antigen quantification. For strategies based on LTB or CTB as carriers, proper folding and formation of pentameric structures produced in plants can be assessed by evaluating their interaction with the GM1 receptor in GM1–ELISA assays. Positive signals for this analysis imply that the chimeric protein is assembled into the pentameric form, and therefore, a proper uptake can be expected at the mucosa.
On the other hand, VLPs are usually detected via electron microscopy, which provides evidence of their successful assembly. VLPs have the ability to stimulate strong immune responses upon oral delivery. In fact, it is considered that the compact and highly ordered structures of VLPs can provide resistance to digestive proteases (100). It is worth mentioning that the antigenic proteins for the HBV are one of the most studied models for production of plant-derived VLPs (101). It has been widely demonstrated that HBV VLP carriers spontaneously assemble in plant cells, resulting in VLPs that preserve their structure (102).
In terms of posttranslational modifications, glycosylation is of particular relevance in the production of vaccines based on GP. It should be considered that distinct glycosylation processes occur in plants with respect to mammalian cells: complex type glycans in plants possess, unlike GPs in mammals, a β(1,2′)-xylose residue, and/or an α(1,3′)-fucose residue linked to the core glycan (103), and a second N-acetylglucosamine (GlcNAc) is enzymatically added to the mannose core, and lack of β(1,4′)-galactose- and sialic acid-containing complex type glycans as well as the bi-antennary N-glycans production found in mammals. However, these differences on glycosylation do not necessarily result in a non-functional or low quality product. In fact, in the case of vaccines, there is the possibility that differential glycans associated with the plant-derived antigen could enhance immunogenicity (104). Moreover, recent advances in the plant glycoengineering allow human-like glycomodification and optimization of the desired glycan structures for enhancing safety and functionality of recombinant vaccines (105).
Another important consideration is the use of rodent or large animal models mentioned above that will allow assessing the immunogenicity and immunoprotective potential of the plant-made EBOV vaccine candidates.
Two cases can be highlighted as examples of how the above mentioned methodologies have resulted in desirable vaccine prototypes: (i) a plant-based vaccine candidate against malaria has been produced in plants by using a transplastomic expression approach. Fusion proteins consisting of CTB along with the antigens malaria apical membrane antigen-1 (AMA1) or merozoite surface protein-1 were produced in lettuce and tobacco leaves; these candidates induced humoral responses and protective immunity against a cholera toxin challenge. Moreover, both oral and injectable vaccination with CTB-AMA-1 resulted in the blocking of the parasite from entering the erythrocytes (13). It should also be considered that LTB has been successfully produced in several crops, including corn and potato (106, 107). The potato-made LTB was used to conduct a pioneering Phase I clinical trial, showing its capacity to achieve seroconversion with no major adverse effects following an oral immunization scheme (106). (ii) A vaccine prototype against Mycobacterium tuberculosis has been developed following an approach based on RICs. The early secreted Ag85B and the latency-associated Acr antigen were expressed in tobacco plants as fusion proteins along with an anti-Acr antibody. Remarkably, Bacillus Calmette–Guérin (BCG)-immunized mice boosted intranasally with TB-RICs showed a significant reduction in M. tuberculosis lung infection in comparison with the group immunized only with BCG (108).
Based on the current evidence on the efficacy of several plant-based vaccines orally administered (16), it is proposed that plant-based formulations may result in strong immune responses that could provide immunoprotection against EBOV. It should also be considered that plant-based vaccines could be applied as oral boosters in prime-boost immunization approaches. This focus has proven useful in many plant-based vaccine prototypes, including those against Yersinia pestis, HBV, and M. tuberculosis (11, 108, 109). All of the aspects mentioned in this article are critical in defining the feasibility of performing evaluations of the plant-based vaccine candidates in clinical trials.
Concluding Remarks
There is an urgent need to develop efficacious vaccines against the EVD. Although preclinical trials are continuously reported for EBOV vaccine prototypes, efforts to develop low-cost vaccine production platforms should be contemplated. Plant-made vaccines offer the potential to address large-scale vaccine production at low cost, thereby facilitating the success of global vaccination programs, especially in developing and poorer countries where coverage is problematic mainly due to vaccine costs. Only one plant-made vaccine candidate has been developed against the EBOV thus far. Therefore, systematic efforts are required to expand this important research field. The path to address this objective will include (i) the design of protective antigens based on the current knowledge of the EBOV immunogenic determinants and on eficacious immunogenic carriers, preferably those that are highly effective in mucosal membranes; (ii) achieving sufficient antigen yields in edible plant biomass to establish models for oral immunization using minimally processed plant biomass; and (iii) validating the safety as well as the immunogenic and immunoprotective potential of plant-made vaccine candidates in test animals.
Each expression platform offers particular advantages, and the election should be based on the nature of the chosen antigen, the required time response, and desired delivery route. In conclusion, the continuing effort toward the development of plant-made vaccines prototypes could lead to important data to select approaches with the realistic goal of providing efficacious and cost-effective strategies to protect against the EVD. Thus, we encourage research in this direction to accelerate the fight against this deadly disease.
Author Contributions
SR-M designed the content of the review, wrote the plant sciences aspects, and corrected the full manuscript. RN-G performed the epitope analysis, made the figures and participated in the general writting of the manuscript. CA designed in part the content of the review, and wrote the immunology and virology aspects.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
The current investigations from the group are supported by CONACYT/México (grant 271182 to SR-M and grant 151818 to CA). Omar Gonzalez edited the English version of this manuscript.
Footnotes
References
1. Kosal ME. A new role for public health in bioterrorism deterrence. Front Public Health (2014) 2:278. doi:10.3389/fpubh.2014.00278
2. Jun SR, Leuze MR, Nookaew I, Uberbacher EC, Land M, Zhang Q, et al. Ebolavirus comparative genomics. FEMS Microbiol Rev (2015) 39:764–78. doi:10.1093/femsre/fuv031
3. Marsh GA, Haining J, Robinson R, Foord A, Yamada M, Barr JA. Ebola Reston virus infection of pigs: clinical significance and transmission potential. J Infect Dis (2011) 204:S804–9. doi:10.1093/infdis/jir300
4. Kuhn JH, Becker S, Ebihara H, Geisbert TW, Johnson KM, Kawaoka Y, et al. Proposal for a revised taxonomy of the family Filoviridae: classification, names of taxa and viruses, and virus abbreviations. Arch Virol (2010) 155:2083–103. doi:10.1007/s00705-010-0814-x
5. Lado M, Walker NF, Baker P, Haroon S, Brown CS, Youkee D, et al. Clinical features of patients isolated for suspected Ebola virus disease at Connaught Hospital, Freetown, Sierra Leone: a retrospective cohort study. Lancet Infect Dis (2015) 15:1024–33. doi:10.1016/S1473-3099(15)00137-1
6. Agnandji ST, Huttner A, Zinser ME, Njuguna P, Dahlke C, Fernandes JF, et al. Phase 1 trials of rVSV Ebola vaccine in Africa and Europe. N Engl J Med (2016) 374:1647–60. doi:10.1056/NEJMoa1502924
7. Qiu X, Wong G, Audet J, Bello A, Fernando L, Alimonti JB, et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature (2014) 514:47–53. doi:10.1038/nature13777
8. Tully CM, Lambe T, Gilbert SC, Hill AS. Emergency Ebola response: a new approach to the rapid design and development of vaccines against emerging diseases. Lancet Infect Dis (2015) 15:356–9. doi:10.1016/S1473-3099(14)71071-0
9. Avesani L, Bortesi L, Santi L, Falorni A, Pezzotti M. Plant-made pharmaceuticals for the prevention and treatment of autoimmune diseases: where are we? Expert Rev Vaccines (2010) 9:957–69. doi:10.1586/erv.10.82
10. Avesani L, Merlin M, Gecchele E, Capaldi S, Brozzetti A, Falorni A, et al. Comparative analysis of different biofactories for the production of a major diabetes autoantigen. Transgenic Res (2014) 23:281–91. doi:10.1007/s11248-013-9749-9
11. Alvarez ML, Pinyerd HL, Crisantes JD, Rigano MM, Pinkhasov J, Walmsley AM, et al. Plant-made subunit vaccine against pneumonic and bubonic plague is orally immunogenic in mice. Vaccine (2006) 24:2477–9. doi:10.1016/j.vaccine.2005.12.057
12. Su J, Zhu L, Sherman A, Wang X, Lin S, Kamesh A, et al. Low cost industrial production of coagulation factor IX bioencapsulated in lettuce cells for oral tolerance induction in hemophilia. Biomaterials (2015) 70:84–93. doi:10.1016/j.biomaterials.2015.08.004
13. Davoodi-Semiromi A, Schreiber M, Nalapalli S, Verma D, Singh ND, Banks RK, et al. Chloroplast-derived vaccine antigens confer dual immunity against cholera and malaria by oral or injectable delivery. Plant Biotechnol J (2010) 8:223–42. doi:10.1111/j.1467-7652.2009.00479.x
14. Monreal-Escalante E, Govea-Alonso DO, Hernández M, Cervantes J, Salazar-González JA, Romero-Maldonado A, et al. Towards the development of an oral vaccine against porcine cysticercosis: expression of the protective HP6/TSOL18 antigen in transgenic carrots cells. Planta (2016) 243:675–85. doi:10.1007/s00425-015-2431-0
15. Hernández M, Rosas G, Cervantes J, Fragoso G, Rosales-Mendoza S, Sciutto E. Transgenic plants: a 5-year update on oral antipathogen vaccine development. Expert Rev Vaccines (2014) 13:1523–36. doi:10.1586/14760584.2014.953064
16. Chan HT, Daniell H. Plant-made oral vaccines against human infectious diseases –are we there yet? Plant Biotechnol (2015) 13:1056–70. doi:10.1111/pbi.12471
17. Streatfield SJ, Kushnir N, Yusibov V. Plant-produced candidate countermeasures against emerging and reemerging infections and bioterror agents. Plant Biotechnol J (2015) 13:1136–59. doi:10.1111/pbi.12475
18. Ward BJ, Landry N, Trépanier S, Mercier G, Dargis M, Couture M. Human antibody response to N-glycans present on plant-made influenza virus-like particle (VLP) vaccines. Vaccine (2014) 32:6098–106. doi:10.1016/j.vaccine
19. Chichester JA, Jones RM, Green BJ, Stow M, Miao F, Moonsammy G, et al. Safety and immunogenicity of a plant-produced recombinant hemagglutinin-based influenza vaccine (HAI-05) derived from A/Indonesia/05/2005 (H5N1) influenza virus: a phase 1 randomized, double-blind, placebo-controlled, dose-escalation study in healthy adults. Viruses (2012) 4:3227–44. doi:10.3390/v4113227
20. Cummings JF, Guerrero ML, Moon JE, Waterman P, Nielsen RK, Jefferson S, et al. Safety and immunogenicity of a plant-produced recombinant monomer hemagglutinin-based influenza vaccine derived from influenza A (H1N1)pdm09 virus: a phase 1 dose-escalation study in healthy adults. Vaccine (2014) 32:2251–9. doi:10.1016/j.vaccine.2013.10.017
21. Bosch D, Schots A. Plant glycans: friend or foe in vaccine development? Expert Rev Vaccines (2010) 9:835–42. doi:10.1586/erv.10.83
22. Landry N, Pillet S, Favre D, Poulin JF, Trépanier S, Yassine-Diab B, et al. Influenza virus-like particle vaccines made in Nicotiana benthamiana elicit durable, poly-functional and cross-reactive T cell responses to influenza HA antigens. Clin Immunol (2014) 154(2):164–77. doi:10.1016/j.clim.2014.08.003
23. Pavot V. Ebola virus vaccines: where do we stand? Clin Immunol (2016) 173:44–9. doi:10.1016/j.clim.2016.10.016
24. Mire CE, Geisbert TW, Feldmann H, Marzi A. Ebola virus vaccines – reality or fiction? Expert Rev Vaccines (2016) 15:1421–30. doi:10.1080/14760584.2016.1178068
25. Wilson JA, Hevey M, Bakken R, Guest S, Bray M, Schmaljohn AL. Epitopes involved in antibody-mediated protection from Ebola virus. Science (2000) 287:1664–6. doi:10.1126/science.287.5458.1664
26. Wong G, Richardson JS, Pillet S, Patel A, Qiu X, Alimonti J, et al. Immune parameters correlate with protection against Ebola virus infection in rodents and nonhuman primates. Sci Transl Med (2012) 4:158ra146. doi:10.1126/scitranslmed
27. Marzi A, Engelmann F, Feldmann F, Haberthur K, Shupert WL, Brining D, et al. Antibodies are necessary for rVSV/ZEBOV-GP-mediated protection against lethal Ebola virus challenge in nonhuman primates. Proc Natl Acad Sci U S A (2013) 110:1893–8. doi:10.1073/pnas.1209591110
28. Sullivan NJ, Hensley L, Asiedu C, Geisbert TW, Stanley D, Johnson J, et al. CD8+ cellular immunity mediates rAd5 vaccine protection against Ebola virus infection of nonhuman primates. Nat Med (2011) 17:1128–31. doi:10.1038/nm.2447
29. Food and Drug Administration. (2014). Available from: http://www.fda.gov/BiologicsBloodVaccines/GuidanceComplianceRegulatoryInformation/default.htm
30. Connolly BM, Steele KE, Davis KJ, Geisbert TW, Kell WM, Jaax NK, et al. Pathogenesis of experimental Ebola virus infection in guinea pigs. J Infect Dis (1999) 179(Suppl 1):S203–17. doi:10.1086/514305
31. Bray M, Davis K, Geisbert T, Schmaljohn C, Huggins J. A mouse model for evaluation of prophylaxis and therapy of Ebola hemorrhagic fever. J Infect Dis (1998) 178:651–61. doi:10.1086/515386
32. Ebihara H, Zivcec M, Gardner D, Falzarano D, LaCasse R, Rosenke R, et al. A Syrian golden hamster model recapitulating Ebola hemorrhagic fever. J Infect Dis (2013) 207:306–18. doi:10.1093/infdis/jis626
33. Carrion R Jr, Ro Y, Hoosien K, Ticer A, Brasky K, de la Garza M, et al. A small nonhuman primate model for filovirus-induced disease. Virology (2011) 420:117–24. doi:10.1016/j.virol.2011.08.022
34. Cross RW, Mire CE, Borisevich V, Geisbert JB, Fenton KA, Geisbert TW. The domestic ferret (Mustela putorius furo) as a lethal infection model for 3 species of ebolavirus. J Infect Dis (2016) 214:565–9. doi:10.1093/infdis/jiw209
35. Kozak R, He S, Kroeker A, de La Vega MA, Audet J, Wong G, et al. Ferrets infected with Bundibugyo virus or Ebola virus recapitulate important aspects of human filovirus disease. J Virol (2016) 90:9209–23. doi:10.1128/JVI.01033-16
36. Ye L, Yang C. Development of vaccines for prevention of Ebola virus infection. Microbes Infect (2015) 17:98–108. doi:10.1016/j.micinf.2014.12.004
37. Hoenen T, Groseth A, Feldmann H. Current Ebola vaccines. Expert Opin Biol Ther (2012) 12:859–72. doi:10.1517/14712598.2012.685152
38. Jones SM, Feldmann H, Stroher U, Geisbert JB, Fernando L, Grolla A, et al. Live attenuated recombinant vaccine protects nonhuman primates against Ebola and Marburg viruses. Nat Med (2005) 11:786–90. doi:10.1038/nm1258
39. Sullivan N, Geisbert T, Geisbert J, Shedlock D, Xu L, Lamoreaux L, et al. Immune protection of nonhuman primates against Ebola virus with single low-dose adenovirus vectors encoding modified GPs. PLoS Med (2006) 3:e177. doi:10.1371/journal.pmed.0030177
40. Bukreyev A, Rollin PE, Tate MK, Yang L, Zaki SR, Shieh WJ, et al. Successful topical respiratory tract immunization of primates against Ebola virus. J Virol (2007) 81:6379–88. doi:10.1128/JVI.00105-07
41. Blaney JE, Marzi A, Willet M, Papaneri AB, Wirblich C, Feldmann F, et al. Antibody quality and protection from lethal Ebola virus challenge in nonhuman primates immunized with rabies virus based bivalent vaccine. PLoS Pathog (2013) 9:e1003389. doi:10.1371/journal.ppat.1003389
42. Herbert AS, Kuehne AI, Barth JF, Ortiz RA, Nichols DK, Zak SE, et al. Venezuelan equine encephalitis virus replicon particle vaccine protects nonhuman primates from intramuscular and aerosol challenge with Ebolavirus. J Virol (2013) 87:4952–64. doi:10.1128/JVI.03361-12
43. Warfield KL, Swenson DL, Olinger GG, Kalina WV, Aman MJ, Bavari S. Ebola virus-like particle-based vaccine protects nonhuman primates against lethal Ebola virus challenge. J Infect Dis (2007) 196(Suppl 2):S430–7. doi:10.1086/520583
44. Swenson DL, Wang D, Luo M, Warfield KL, Woraratanadharm J, Holman DH, et al. Vaccine to confer to nonhuman primates complete protection against multistrain Ebola and Marburg virus infections. Clin Vaccine Immunol (2008) 15:460–7. doi:10.1128/CVI.00431-07
45. Sullivan NJ, Sanchez A, Rollin PE, Yang ZY, Nabel GJ. Development of a preventive vaccine for Ebola virus infection in primates. Nature (2000) 408:605–9. doi:10.1038/35046108
46. Ledgerwood JE, Costner P, Desai N, Holman L, Enama ME, Yamshchikov G, et al. A replication defective recombinant Ad5 vaccine expressing Ebola virus GP is safe and immunogenic in healthy adults. Vaccine (2010) 29:304–13. doi:10.1016/j.vaccine.2010.10.037
47. Stanley DA, Honko AN, Asiedu C, Trefry JC, Lau-Kilby AW, Johnson JC, et al. Chimpanzee adenovirus vaccine generates acute and durable protective immunity against Ebola virus challenge. Nat Med (2014) 20:1126–9. doi:10.1038/nm.3702
48. Halfmann P, Ebihara H, Marzi A, Hatta Y, Watanabe S, Suresh M, et al. Replication-deficient Ebolavirus as a vaccine candidate. J Virol (2009) 83:3810–5. doi:10.1128/JVI.00074-09
49. Marzi A, Halfmann P, Hill-Batorski L, Feldmann F, Shupert WL, Neumann G, et al. Vaccines. An Ebola whole-virus vaccine is protective in nonhuman primates. Science (2015) 348:439–42. doi:10.1126/science.aaa4919
50. Milligan ID, Gibani MM, Sewell R, Clutterbuck EA, Campbell D, Plested E, et al. Safety and immunogenicity of novel adenovirus type 26- and modified vaccinia Ankara-vectored Ebola vaccines: a randomized clinical trial. JAMA (2016) 315:1610–23. doi:10.1001/jama.2016.4218
51. Marzi A, Robertson SJ, Haddock E, Feldmann F, Hanley PW, Scott DP, et al. Ebola vaccine. VSV-EBOV rapidly protects macaques against infection with the 2014/15 Ebola virus outbreak strain. Science (2015) 349:739–42. doi:10.1126/science.aab3920
52. Geisbert TW, Daddario-Dicaprio KM, Geisbert JB, Reed DS, Feldmann F, Grolla A, et al. Vesicular stomatitis virus-based vaccines protect nonhuman primates against aerosol challenge with Ebola and Marburg viruses. Vaccine (2008) 26:6894–900. doi:10.1016/j.vaccine.2008.09.082
53. Qiu X, Fernando L, Alimonti JB, Melito PL, Feldmann F, Dick D, et al. Mucosal immunization of cynomolgus macaques with the VSVDeltaG/ZEBOVGP vaccine stimulates strong Ebola GP-specific immune responses. PLoS One (2009) 4:e5547. doi:10.1371/journal.pone.0005547
54. Geisbert TW, Daddario-DiCaprio KM, Williams KJ, Geisbert JB, Leung A, Feldmann F, et al. Recombinant vesicular stomatitis virus vector mediates postexposure protection against Sudan Ebola hemorrhagic fever in nonhuman primates. J Virol (2008) 82:5664–8. doi:10.1128/JVI.00456-08
55. Lai L, Davey R, Beck A, Xu Y, Suffredini AF, Palmore T, et al. Emergency postexposure vaccination with vesicular stomatitis virus-vectored Ebola vaccine after needlestick. JAMA (2015) 313:1249–55. doi:10.1001/jama.2015
56. Günther S, Feldmann H, Geisbert TW, Hensley LE, Rollin PE, Nichol ST, et al. Management of accidental exposure to Ebola virus in the biosafety level 4 laboratory, Hamburg, Germany. J Infect Dis (2011) 204(Suppl 3):S785–90. doi:10.1093/infdis/jir298
57. Mire CE, Matassov D, Geisbert JB, Latham TE, Agans KN, Xu R, et al. Single dose attenuated Vesiculovax vaccines protect primates against Ebola Makona virus. Nature (2015) 520:688–91. doi:10.1038/nature14428
58. DiNapoli JM, Yang L, Samal SK, Murphy BR, Collins PL, Bukreyev A. Respiratory tract immunization of non-human primates with a Newcastle disease virus-vectored vaccine candidate against Ebola virus elicits a neutralizing antibody response. Vaccine (2010) 29:17–25. doi:10.1016/j.vaccine.2010.10.024
59. Huttner A, Dayer JA, Yerly S, Combescure C, Auderset F, Desmeules J, et al. The effect of dose on the safety and immunogenicity of the VSV Ebola candidate vaccine: a randomised double-blind, placebo-controlled phase 1/2 trial. Lancet Infect Dis (2015) 15:1156–66. doi:10.1016/S1473-3099(15)00154-1
60. Henao-Restrepo AM, Longini IM, Egger M, Dean NE, Edmunds WJ, Camacho A, et al. Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: interim results from the guinea ring vaccination cluster-randomised trial. Lancet (2015) 386:857–66. doi:10.1016/S0140-6736(15)61117-5
61. Henao-Restrepo AM, Camacho A, Longini IM, Watson CH, Edmunds WJ, Egger M, et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). Lancet (2017) 389:505–18. doi:10.1016/S0140-6736(16)32621-6
62. Konduru K, Bradfute SB, Jacques J, Manangeeswaran M, Nakamura S, Morshed S, et al. Ebola virus glycoprotein Fc fusion protein confers protection against lethal challenge in vaccinated mice. Vaccine (2011) 29:2968–77. doi:10.1016/j.vaccine.2011.01.113
63. Phoolcharoen W, Dye JM, Kilbourne J, Piensook K, Pratt WD, Arntzen CJ, et al. A nonreplicating subunit vaccine protects mice against lethal Ebola virus challenge. Proc Natl Acad Sci U S A (2011) 108:20695–700. doi:10.1073/pnas.1117715108
64. Martins KA, Jahrling PB, Bavari S, Kuhn JH. Ebola virus disease candidate vaccines under evaluation in clinical trials. Expert Rev Vaccines (2016) 15:1101–12. doi:10.1080/14760584.2016.1187566
65. Martin JE, Sullivan NJ, Enama ME, Gordon IJ, Roederer M, Koup RA, et al. A DNA vaccine for Ebola virus is safe and immunogenic in a phase I clinical trial. Clin Vaccine Immunol (2006) 13:1267–77. doi:10.1128/CVI.00162-06
66. Sarwar UN, Costner P, Enama ME, Berkowitz N, Hu Z, Hendel CS, et al. Safety and immunogenicity of DNA vaccines encoding Ebola virus and Marburgvirus wild-type glycoproteins in a phase I clinical trial. J Infect Dis (2015) 211:549–57. doi:10.1093/infdis/jiu511
67. Cooper CL, Bavari S. A race for an Ebola vaccine: promises and obstacles. Trends Microbiol (2015) 23:65–6. doi:10.1016/j.tim.2014.12.005
68. Mire CE, Geisbert JB, Agans KN, Satterfield BA, Versteeg KM, Fritz EA, et al. Durability of a vesicular stomatitis virus-based marburg virus vaccine in nonhuman primates. PLoS One (2014) 9:e94355. doi:10.1371/journal.pone.0094355
69. Wong G, Audet J, Fernando L, Fausther-Bovendo H, Alimonti JB, Kobinger GP, et al. Immunization with vesicular stomatitis virus vaccine expressing the Ebola glycoprotein provides sustained long-term protection in rodents. Vaccine (2014) 32:5722–9. doi:10.1016/j.vaccine.2014.08.028
70. Kennedy SB, Neaton JD, Lane HC, Kieh MWS, Massaquoi MBF, Touchette NA, et al. Implementation of an Ebola virus disease vaccine clinical trial during the Ebola epidemic in Liberia: design, procedures, and challenges. Clin Trials (2016) 13:49–56. doi:10.1177/1740774515621037
71. Becquart P, Mahlakõiv T, Nkoghe D, Leroy EM. Identification of continuous human B-cell epitopes in the VP35, VP40, nucleoprotein and glycoprotein of Ebola virus. PLoS One (2014) 9:e96360. doi:10.1371/journal.pone.0096360
72. Vaughan K, Ponomarenko J, Sette A. (2014). Ebola Virus Related Immune Epitope Data Curated in the IEDB. Available from: http://help.iedb.org/entries/51011785-Ebola-virus-related-immune-epitope-data-curated-in-the-IEDB
73. Takada A, Ebihara H, Feldmann H, Geisbert TW, Kawaoka Y. Epitopes required for antibody-dependent enhancement of Ebola virus infection. J Infect Dis (2007) 196(Suppl 2):S347–56. doi:10.1086/520581
74. Lee JE, Saphire EO. Ebola virus glycoprotein structure and mechanism of entry. Future Virol (2009) 4:621–35. doi:10.2217/fvl.09.56
75. Wang Y, Liu Z, Dai Q. A highly immunogenic fragment derived from Zaire Ebola virus glycoprotein elicits effective neutralizing antibody. Virus Res (2014) 189:254–61. doi:10.1016/j.virusres.2014.06.001
76. Khan MA, Hossain MU, Rakib-Uz-Zaman SM, Morshed MN. Epitope-based peptide vaccine design and target site depiction against Ebola viruses: an immunoinformatics study. Scand J Immunol (2015) 82:25–34. doi:10.1111/sji.12302
77. Spangler BD. Structure of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol Rev (1992) 56:622–47.
78. Guan C, Ji J, Jin C, Wang G, Li X, Guan W. Expression of cholera toxin B subunit-lumbrokinase in edible sunflower seeds – the use of transmucosal carrier to enhance its fusion protein’s effect on protection of rats and mice against thrombosis. Biotechnol Prog (2014) 30:1029–39. doi:10.1002/btpr.1963
79. Rosales-Mendoza S, Soria-Guerra RE, Moreno-Fierros L, Govea-Alonso DO, Herrera-Díaz A, Korban SS, et al. Immunogenicity of nuclear-encoded LTB:ST fusion protein from Escherichia coli expressed in tobacco plants. Plant Cell Rep (2011) 30:1145–52. doi:10.1007/s00299-011-1023-0
80. Yu J, Langridge WH. A plant-based multicomponent vaccine protects mice from enteric diseases. Nat Biotechnol (2001) 19:548–52. doi:10.1038/89297
81. Frankel AD, Pabo CO. Cellular uptake of the tat protein from human immunodeficiency virus. Cell (1988) 55:1189–93. doi:10.1016/0092-8674(88)90263-2
82. Derossi D, Joliot AH, Chassaing G, Prochiantz A. The third helix of the Antennapedia homeodomain translocates through biological membranes. J Biol Chem (1994) 269:10444–50.
83. Scotti N, Rybicki EP. Virus-like particles produced in plants as potential vaccines. Expert Rev Vaccines (2013) 12:211–24. doi:10.1586/erv.12.147
84. Marusic C, Rizza P, Lattanzi L, Mancini C, Spada M, Belardelli F, et al. Chimeric plant virus particles as immunogens for inducing murine and human immune responses against human immunodeficiency virus type 1. J Virol (2001) 75:8434–9. doi:10.1128/JVI.75.18.8434-8439.2001
85. Chargelegue D, Drake P, Obregon P, Prada A, Fairweather N, Ma JK. Highly immunogenic and protective recombinant vaccine candidate expressed in transgenic plants. Infect Immun (2005) 73:5915–22. doi:10.1128/IAI.73.9.5915-5922.2005
86. Komarova TV, Baschieri S, Donini M, Marusic C, Benvenuto E, Dorokhov YL. Transient expression systems for plant-derived biopharmaceuticals. Expert Rev Vaccines (2010) 9:859–76. doi:10.1586/erv.10.85
87. Thuenemann EC, Meyers AE, Verwey J, Rybicki EP, Lomonossoff GP. A method for rapid production of heteromultimeric protein complexes in plants: assembly of protective bluetongue virus-like particles. Plant Biotechnol J (2013) 11(7):839–46. doi:10.1111/pbi.12076
88. D’aoust M, Couture M, Lavoie P, Vezina L. Virus Like Particle Production in Plants. U.S. Patent No 20,130,344,100 (2013).
89. Phoolcharoen W, Bhoo SH, Lai H, Ma J, Arntzen CJ, Chen Q, et al. Expression of an immunogenic Ebola immune complex in Nicotiana benthamiana. Plant Biotechnol J (2011) 9:807–16. doi:10.1111/j.1467-7652.2011.00593.x
90. Salazar-González JA, Monreal-Escalante E, Herrera Díaz A, Koop HU, Rosales-Mendoza S. Plastid-based expression strategies. In: Mendoza SR, editor. Genetically Engineered Plants as a Source of Vaccines Against Wide Spread Diseases – An Integrated View. New York, NY: Springer (2014). p. 61–78.
91. Lelivelt CL, van Dun KM, de Snoo CB, McCabe MS, Hogg BV, Nugent JM. Plastid transformation in lettuce (Lactuca sativa L.) by polyethylene glycol treatment of protoplasts. Methods Mol Biol (2014) 1132:317–30. doi:10.1007/978-1-62703-995-620
92. Ruhlman T, Ahangari R, Devine A, Samsam M, Daniell H. Expression of cholera toxin B-proinsulin fusion protein in lettuce and tobacco chloroplasts – oral administration protects against development of insulitis in non-obese diabetic mice. Plant Biotechnol J (2007) 5:495–510. doi:10.1111/j.1467-7652.2007.00259.x
93. Lakshmi PS, Verma D, Yang X, Lloyd B, Daniell H. Low cost tuberculosis vaccine antigens in capsules: expression in chloroplasts, bio-encapsulation, stability and functional evaluation in vitro. PLoS One (2013) 8:e54708. doi:10.1371/journal.pone.0054708
94. Alvarez ML, Topal E, Martin F, Cardineau GA. Higher accumulation of F1-V fusion recombinant protein in plants after induction of protein body formation. Plant Mol Biol (2010) 72:75–89. doi:10.1007/s11103-009-9552-4
95. Curtis IS. Lettuce (Lactuca sativa L.). Methods Mol Biol (2006) 343:449–58. doi:10.1385/1-59745-130-4:449
96. Rosales-Mendoza S, Soria-Guerra RE, de Jesús Olivera-Flores MT, López-Revilla R, Argüello-Astorga GR, Jimémez-Bremont JF, et al. Expression of Escherichia coli heat-labile enterotoxin b subunit (LTB) in carrot (Daucus carota L.). Plant Cell Rep (2007) 26:969–76. doi:10.1007/s00299-007-0310-2
97. Balestrazzi A, Carbonera D, Cella R. Transformation of Daucus carota hypocotyls mediated by Agrobacterium tumefaciens. J Genet Breed (1991) 45:135–40.
98. Pastores GM, Petakov M, Giraldo P, Rosenbaum H, Szer J, Deegan PB, et al. A phase 3, multicenter, open-label, switchover trial to assess the safety and efficacy of taliglucerase alfa, a plant cell-expressed recombinant human glucocerebrosidase, in adult and pediatric patients with Gaucher disease previously treated with imiglucerase. Blood Cells Mol Dis (2014) 53:253–60. doi:10.1016/j.bcmd.2014.05.004
99. Shaaltiel Y, Gingis-Velitski S, Tzaban S, Fiks N, Tekoah Y, Aviezer D. Plant-based oral delivery of β-glucocerebrosidase as an enzyme replacement therapy for Gaucher’s disease. Plant Biotechnol J (2015) 13:1033–40. doi:10.1111/pbi.12366
100. Smith ML, Richter L, Arntzen CJ, Shuler ML, Mason HS. Structural characterization of plant-derived hepatitis B surface antigen employed in oral immunization studies. Vaccine (2003) 21:4011–21. doi:10.1016/S0264-410X(03)00268-8
101. Meador LR, Mor TS. Meeting report VLPNPV: session 5: plant based technology. Hum Vaccin Immunother (2014) 10:3068–73. doi:10.4161/21645515.2014.979693
102. Huang Z, Elkin G, Maloney BJ, Beuhner N, Arntzen CJ, Thanavala Y, et al. Virus-like particle expression and assembly in plants: hepatitis B and Norwalk viruses. Vaccine (2005) 23:1851–8. doi:10.3748/wjg.v9.i5.996
103. Lerouge P, Bardor M, Pagny S, Gomord V, Faye L. N-glycosylation of recombinant pharmaceutical glycoproteins produced in transgenic plants: towards an humanisation of plant N-glycans. Curr Pharm Biotechnol (2000) 1:347–54. doi:10.2174/1389201003378843
104. Hamorsky KT, Kouokam JC, Jurkiewicz JM, Nelson B, Moore LJ, Husk AS, et al. N-glycosylation of cholera toxin B subunit in Nicotiana benthamiana: impacts on host stress response, production yield and vaccine potential. Sci Rep (2015) 5:8003. doi:10.1038/srep08003
105. Kim HS, Jeon JH, Lee KJ, Ko K. N-glycosylation modification of plant-derived virus-like particles: an application in vaccines. Biomed Res Int (2014) 2014:249519. doi:10.1155/2014/249519
106. Tacket CO, Mason HS, Losonsky G, Clements JD, Levine MM, Arntzen CJ. Immunogenicity in humans of a recombinant bacterial antigen delivered in a transgenic potato. Nat Med (1998) 4(5):607–9. doi:10.1038/nm0598-607
107. Chikwamba RK, Scott MP, Mejía LB, Mason HS, Wang K. Localization of a bacterial protein in starch granules of transgenic maize kernels. Proc Natl Acad Sci U S A (2003) 100:11127–32. doi:10.1073/pnas.1836901100
108. Pepponi I, Diogo GR, Stylianou E, van Dolleweerd CJ, Drake PM, Paul MJ, et al. Plant-derived recombinant immune complexes as self-adjuvanting TB immunogens for mucosal boosting of BCG. Plant Biotechnol J (2014) 12:840–50. doi:10.1111/pbi.12185
109. Pniewski T, Kapusta J, Bociag P, Wojciechowicz J, Kostrzak A, Gdula M, et al. Low-dose oral immunization with lyophilized tissue of herbicide-resistant lettuce expressing hepatitis B surface antigen for prototype plant-derived vaccine tablet formulation. J Appl Genet (2011) 52:125–36. doi:10.1007/s13353-010-0001-5
Keywords: Ebola virus, mucosal immunization, low-cost vaccine, global vaccination, molecular pharming, glycoprotein antigen, VP antigen
Citation: Rosales-Mendoza S, Nieto-Gómez R and Angulo C (2017) A Perspective on the Development of Plant-Made Vaccines in the Fight against Ebola Virus. Front. Immunol. 8:252. doi: 10.3389/fimmu.2017.00252
Received: 27 October 2016; Accepted: 20 February 2017;
Published: 10 March 2017
Edited by:
Daniela Santoro Rosa, Federal University of São Paulo, BrazilReviewed by:
Tomasz Pniewski, Institute of Plant Genetics (PAN), PolandChad Edward Mire, University of Texas Medical Branch, USA
Copyright: © 2017 Rosales-Mendoza, Nieto-Gómez and Angulo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio Rosales-Mendoza, rosales.s@fcq.uaslp.mx;
Carlos Angulo, eangulo@cibnor.mx