 Kostas Patas1
Kostas Patas1 Anne Willing1†
Anne Willing1† Cüneyt Demiralay2†
Cüneyt Demiralay2† Jan Broder Engler1Andreea Lupu1,3Caren Ramien1
Jan Broder Engler1Andreea Lupu1,3Caren Ramien1 Tobias Schäfer4
Tobias Schäfer4 Christian Gach4Laura Stumm2Kenneth Chan5
Christian Gach4Laura Stumm2Kenneth Chan5 Marissa Vignali5
Marissa Vignali5 Petra C. Arck6
Petra C. Arck6 Manuel A. Friese1
Manuel A. Friese1 Ole Pless4Klaus Wiedemann2Agorastos Agorastos2†
Ole Pless4Klaus Wiedemann2Agorastos Agorastos2† Stefan M. Gold1,7*†
Stefan M. Gold1,7*†
- 1Institut für Neuroimmunologie und Multiple Sklerose (INIMS), Universitätsklinikum Hamburg-Eppendorf, Hamburg, Germany
- 2Klinik für Psychiatrie und Psychotherapie, Universitätsklinikum Hamburg-Eppendorf, Hamburg, Germany
- 3Immunomodulation Group, Cantacuzino National Research Institute, Bucharest, Romania
- 4Fraunhofer IME ScreeningPort, Hamburg, Germany
- 5Adaptive Biotechnologies, Seattle, WA, Unites States
- 6Experimentelle Feto-Maternale Medizin, Universitätsklinikum Hamburg-Eppendorf, Hamburg, Germany
- 7Charité – Universitätsmedizin Berlin, Humboldt Universität zu Berlin, Berlin Institute of Health (BIH), Klinik für Psychiatrie und Psychotherapie, Campus Benjamin Franklin (CBF), Berlin, Germany
While a link between inflammation and the development of neuropsychiatric disorders, including major depressive disorder (MDD) is supported by a growing body of evidence, little is known about the contribution of aberrant adaptive immunity in this context. Here, we conducted in-depth characterization of T cell phenotype and T cell receptor (TCR) repertoire in MDD. For this cross-sectional case–control study, we recruited antidepressant-free patients with MDD without any somatic or psychiatric comorbidities (n = 20), who were individually matched for sex, age, body mass index, and smoking status to a non-depressed control subject (n = 20). T cell phenotype and repertoire were interrogated using a combination of flow cytometry, gene expression analysis, and next generation sequencing. T cells from MDD patients showed significantly lower surface expression of the chemokine receptors CXCR3 and CCR6, which are known to be central to T cell differentiation and trafficking. In addition, we observed a shift within the CD4+ T cell compartment characterized by a higher frequency of CD4+CD25highCD127low/− cells and higher FOXP3 mRNA expression in purified CD4+ T cells obtained from patients with MDD. Finally, flow cytometry-based TCR Vβ repertoire analysis indicated a less diverse CD4+ T cell repertoire in MDD, which was corroborated by next generation sequencing of the TCR β chain CDR3 region. Overall, these results suggest that T cell phenotype and TCR utilization are skewed on several levels in patients with MDD. Our study identifies putative cellular and molecular signatures of dysregulated adaptive immunity and reinforces the notion that T cells are a pathophysiologically relevant cell population in this disorder.
Introduction
Major depressive disorder (MDD) affects approximately 15% of adults over their lifespan (1) and is among the top two leading causes of years lived with disability worldwide (2). The etiopathogenesis of MDD is multifactorial and many neurobiological systems have been implicated (3, 4). Despite advanced understanding of the pathophysiology, approximately 30% of patients do not respond—even after several treatment attempts—to current antidepressants, which mainly target monoaminergic neurotransmission (5, 6). This underlines the need for approaches that target mechanisms beyond monoamine modulation (7).
Recently, accumulating experimental animal and human data have highlighted the importance of aberrant immunity in the development of depression (8). Similarly, epidemiological data have linked infections and autoimmunity to the risk of the disorder (9, 10). Exploring immune alterations in MDD may, thus, have the potential to open new avenues for treatment, since the immune system could be more amendable by therapeutic modulation than targets in the central nervous system (CNS).
Much of the research on immune dysfunction in MDD has focused on activation of the innate immune system (8). However, intriguing new evidence suggests that the different branches of the immune system might be differentially affected in MDD with relative impairment of adaptive immune responses (11), although this has not yet been explored in a targeted way on a cellular level. Moreover, preclinical studies have implicated the presence of a co-evolutionary link between T cell responses and CNS function (12, 13) and it is now increasingly recognized that the T cell compartment regulates cognition and mood-related behaviors in experimental animals (14, 15). Along similar lines, aberrant T cell differentiation and attenuated reactivity to CNS-derived antigens were hypothesized to play a role in MDD pathophysiology (8, 16). In view of these putative “antigen selection” pressures (i.e., infections and skewed specificity), we hypothesized that peripheral blood T cells from MDD patients would exhibit an altered phenotype as well as a biased profile in the T cell receptor (TCR) repertoire compared to non-depressed controls.
Materials and Methods
Subjects
We enrolled antidepressant-free patients with MDD but no other psychiatric comorbidities as well as non-depressed volunteers, matched pairwise for sex, age, current smoking status (yes/no), and BMI. All participants were of European descent.
Inclusion criteria for MDD patients: (a) psychiatrist-confirmed diagnosis of MDD, single or recurrent, according to DSM-IV criteria; (b) a minimum score of 18 points on the Hamilton Rating Scale for Depression (HRSD); (c) age 18–65 years; (d) at least 8 weeks free of any psychiatric medication (e.g., antidepressants, antipsychotics, and mood stabilizers).
Inclusion criteria for non-depressed controls: (a) no current or lifetime mood disorder diagnosis and (b) a score ≤5 on the Quick Inventory of Depressive Symptoms-Self Report (QIDS-SR).
Exclusion criteria for all participants: (a) past or present self-reported diagnosis of a major medical condition, including chronic or acute inflammatory, metabolic, and neurological disorders; (b) Axes I or II comorbidities; (c) regular use of either prescribed, over-the-counter medication or illicit drugs; thus, any subject on anti-inflammatory drugs, cholesterol-reducing drugs, and other possibly immune-modifying agents was excluded (see Table S1 in Supplementary Material); (d) drinking of more than 100 g of alcohol per week; (e) basic blood laboratory test values deviating significantly from the normal range; (f) current adverse life events (e.g., divorce, loss of job, and illness in the family); (g) pregnancy or nursing; and (h) recent vaccination (within the past month). Hypothyroidism in euthyroid state through hormonal substitution and hypertension in normotensive state through antihypertensive medication did not represent exclusion criteria (see Table S1 in Supplementary Material).
Patients were recruited through our specialized depression out-patient clinic program at the Department of Psychiatry and Psychotherapy, University Medical Center Hamburg-Eppendorf. Non-depressed controls were recruited from the same geographical region through advertisements and from the staff of the University Medical Center Hamburg-Eppendorf. All participants provided written informed consent before enrollment in the study. This study has been approved by the appropriate Ethics Review Committee (Ethik-Kommission der Ärztekammer Hamburg, Ethikvotum PV4161 and PV4719).
Clinical Assessments
All subjects underwent detailed clinical assessments, including medical history, current medication, and psychiatric comorbidities. Diagnosis was established with the Structured Clinical Interview for the DSM-IV-TR Axis I Disorders (SCID-I) and depression severity was assessed using the HDRS by experienced board-certified psychiatrists (Cüneyt Demiralay and Agorastos Agorastos).
Peripheral Blood Mononuclear Cells (PBMC) Isolation and Biobanking
We obtained approximately 30 ml of blood in S-Monovette K3 EDTA tubes and 7 ml in S-Monovette Serum-Gel tubes (Sarstedt). All samples were collected in the morning (8:00 a.m.). PBMCs were then isolated using a Ficoll–Hypaque gradient as described (17), aliquoted in RPMI containing 10% DMSO and 25% FCS at 1 × 107 cells/ml, gradually cooled down to −80°C in a Mr. Frosty for 18 h and stored in liquid nitrogen until assayed. A small amount of whole blood (50 µl) was used for total and differential leukocyte counts using a Coulter Ac·T Diff hematology analyzer (Beckman Coulter). All other assays were performed using cryopreserved PBMCs or serum with one subject from each group run in parallel to control systematic variation in reagents used.
Cell Purification
For qRT-PCR and TCR sequencing analyses, CD4+ T cells were purified using magnetic beads (negative selection by BD IMag Human CD4 T Lymphocyte Enrichment Set, BD Biosciences) from PBMC aliquots thawed in cell separation buffer (1% human serum, 2 mM EDTA in PBS). In our hands, this method typically yields a cell purity of approximately 95% for CD4+ T cells as confirmed by flow cytometry.
Flow Cytometry
We used flow cytometry with hierarchical gating strategies adapted from established guidelines for analysis of human PBMCs and suitable for cryopreserved samples (18, 19). Our panel offered survey phenotyping of T cell subsets, including Tregs, as well as B cells and NK cells (see Figure S1 in Supplementary Material). The following fluorochrome-conjugated monoclonal antibodies were used: CD3 BV605 (OKT3, Biolegend), CD4 Alexa 700 (RPA-T4, Biolegend), CD8α V500 (SK1, BD Biosciences), CCR6 PE-Cy7 (G034E3, Biolegend), CXCR3 PE (G025H7, Biolegend), CD127 APC (A019D5, Biolegend), CD25 BV421 (M-A251, Biolegend), CD19 V500 (HIB19, BD Biosciences), CD56 PE-Cy7 (HCD56, Biolegend), CD20 PE (2H7, BD Biosciences), CD14 V450 (MFP9, BD Biosciences), and CD16 FITC (3G8, BD Biosciences). We also used the commercially available IOTest® Beta Mark Kit (Beckman Coulter) for quantitative determination of the human TCR Vβ repertoire in CD4+ and CD8+ cells [nomenclature from Ref. (20)].
Surface Staining
To exclude dead cells from further analyses, we used the LIVE/DEAD Fixable Near-IR Dead Cell Stain Kit (Life Technologies) before applying any surface and intracellular stainings. Up to 1 × 106 thawed PBMCs were washed with protein-free cold PBS (PAA Laboratories) at 485 g for 5 min at 4°C and then resuspended in 100 µl cold PBS containing the amine-reactive dye in 1:1,000 dilution. After light-protected incubation for 20 min at 4°C, cells were washed with cold PBS and subsequently stained for surface antigens, resuspended in 90 µl staining buffer (0.1% BSA, 0.02% NaN3 in PBS) and incubated with 0.1 µg/µl human IgG (Jackson ImmunoResearch) for 5 min at room temperature. Surface staining reactions were performed by light-protected incubation with 10 µl of Vβ-specific reagent mixture and/or 10 µl of 10× surface antibody cocktails for 30 min at 4°C. After washing with staining buffer at 485 g for 5 min at 4°C, cells were either resuspended in 250 µl staining buffer for acquisition or fixed for intracellular staining.
Intracellular Staining
Surface-stained cells were resuspended in 100 µl fixation buffer (Biolegend), incubated for 20 min at room temperature, washed twice with 1× permeabilization wash buffer (Biolegend) at 485 g for 5 min at 4°C and serially incubated with 0.1 µg/µl human IgG (5 min, at room temperature) and anti-CXCR3 antibodies for 30 min at room temperature. Cells were again washed twice with 1 ml permeabilization wash buffer and resuspended in 250 µl staining buffer for acquisition.
Data were acquired using a BD FACS LSR II flow cytometer and the FACS Diva v6.2 operating software. At least 1 × 105 live lymphocytes were acquired from case–control samples during the same session and using the same acquisition settings. Variability of instrument performance was normalized by use of Cytometer Setup and Tracking beads (BD Biosciences). Data analysis and plotting were performed using FlowJo v10.0.8 (Tree Star).
Serum Immunoassays for CXCL10 and CXCL11
CXCL10 and CXCL11 in sera were assayed with a multiplex bead-based immunoassay LEGENDplex (Biolegend) according to manufacturer’s instructions. For data acquisition and analysis, a BD FACS LSR II flow cytometer and the LEGENDplex v7.0 data analysis software were used, respectively.
Serum Radioimmunoassays for ACTH and Cortisol
Stress hormone levels (ACTH and cortisol) were measured in sera obtained at 8:00 a.m. with commercially available radioimmunoassays (IBL IRMA and ICN Biomedicals RIA, respectively), according to manufacturer’s instructions.
Reverse Transcription and Real-Time PCR
RNA was extracted from purified cell populations using RNeasy Plus Universal Mini Kit (Qiagen). 250–500 ng aliquots were used for cDNA synthesis by RevertAid H Minus First Strand cDNA Synthesis Kit (Thermo Scientific), followed by TaqMan assays (FOXP3: Hs01085834_m1, T-bet: Hs00203436_m1, GATA3: Hs00231122_m1, RORC: Hs01076122_m1) in ABI Prism 7900 HT Fast Real-Time PCR system (Applied Biosystems). All reactions were performed in triplicate. The expression levels of the genes of interest were calculated as 2−ΔCt relative to the geometric mean expression of three housekeeping genes (IPO8: Hs00183533_m1, TBP: Hs00427620_m1, RPL13A: Hs04194366_g1), shown to be stably expressed in primary human T cells (21).
Next Generation Sequencing of the TCRβ Repertoire
For sequencing of the CDR3 region in CD4+ T cell clones, we extracted total genomic DNA from negatively purified CD4+ T cells using the DNeasy Blood and Tissue Kit (Qiagen), according to manufacturer’s instructions. We then amplified and sequenced the CDR3 region of rearranged TCRβ genes, which were defined according to IMGT (22), using previously described immunosequencing protocols [Adaptive Biotechnologies (23)]. This provides an unbiased measurement of the frequencies of individual T cell clones. Raw sequence data were pre-processed by Adaptive Biotechnologies for PCR and sequencing error correction (23) and uploaded into the immunoSEQ® Analyzer. Post-analysis was conducted using VDJtools v1.0.7 (24). Sequencing data are available using the following URL: http://doi.org/10.21417/B7RG6H.
Statistical Analyses
All continuous variables are presented as median values with interquartile range, unless otherwise specified. Differences between patients and matched controls were tested for statistical significance using the paired Wilcoxon signed-rank test. For dichotomous variables, the McNemar’s test was used. Bivariate correlation analyses were conducted using Spearman’s rank correlation test. As a measure of Vβ repertoire skewing, we calculated the Gini-TCR index, as previously described (25). This index is a direct measure of TCR Vβ usage with higher values indicating higher clonality (i.e., less evenly distributed repertoire). All statistical analyses were performed using SPSS version 19 (IBM). A two-tailed p < 0.05 was considered significant and p < 0.10 was considered a trend. All graphs were made using GraphPad Prism v5.04, except for the pie charts in Figure 6, which were generated using the R package ggplot2.
Results
Descriptives
Clinical characteristics of the MDD and control groups can be found in Table 1. Patient level clinical characteristics including non-psychiatric medication are provided in Table S1 in Supplementary Material. Depression severity in the MDD group was moderate to severe. 18 MDD patients were antidepressant-naïve at the time of inclusion.
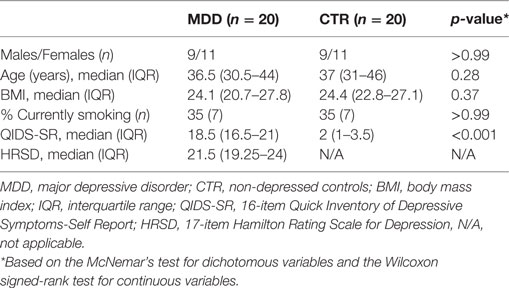
Table 1. Clinical characteristics.
Circulating Leukocyte Numbers and Frequencies
In order to obtain a general overview of immune cell composition in the peripheral blood, we first analyzed the absolute counts and frequencies of major leukocyte subsets. We observed no significant differences in absolute counts of circulating granulocytes, monocytes, and lymphocytes (Figure 1A), frequencies of major T cell subsets (CD4+ or CD8+) or B cells (Figures 1B,C). In line with recent studies (26–28), natural killer (NK) cells showed a trend toward lower frequency in MDD patients (p = 0.062; Figure 1B). Here, group differences were mainly driven by the putatively regulatory CD56highCD16− NK subset (p = 0.018; Figure 1D).
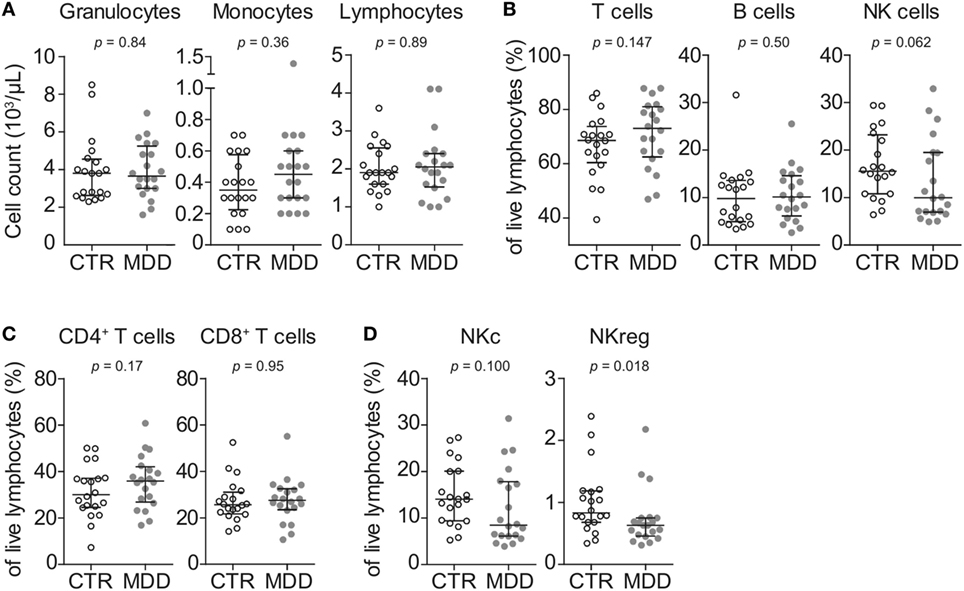
Figure 1. Peripheral blood counts and frequencies of major leukocyte subsets. (A) Absolute peripheral blood granulocyte, monocyte, and lymphocyte counts were obtained from major depressive disorder (MDD) patients and matched non-depressed controls (CTR) using a Coulter Ac·T Diff hematology analyzer (n = 40). (B) Frequencies of total CD3+ lymphocytes (T cells), CD3−CD56−CD19+ B cells and CD3−CD19−CD20−CD14−CD56+ natural killer (NK) cells were obtained by flow cytometric analysis of thawed peripheral blood mononuclear cells. (C) T cells were further discriminated into CD4+CD8− and CD8+CD4− subsets. (D) Among NK cells, CD56lowCD16+ cytotoxic (NKc) cells and CD56highCD16− regulatory (NKreg) cells were also identified. Graphs depict medians with interquartile ranges. For all comparisons, the Wilcoxon signed-rank test was used.
Chemokine Receptor Expression CXCR3 and CCR6
To identify possible shifts in T cell phenotype, we next analyzed the expression of two chemokine receptors that are characteristic of effector T cell differentiation (CXCR3 and CCR6) (18). T cells of MDD patients showed significant reductions in the surface expression of both CXCR3 (p = 0.001; Figures 2A,B) and CCR6 (p = 0.033; Figures 2C,D). This was seen in both CD4+ and CD8+ T cell subsets for CXCR3 (Figure 2B) and mainly in CD4+ T cells for CCR6 (Figure 2D). In 17 out of our 20 case–control pairs, CXCR3 expression was lower in the MDD subjects.
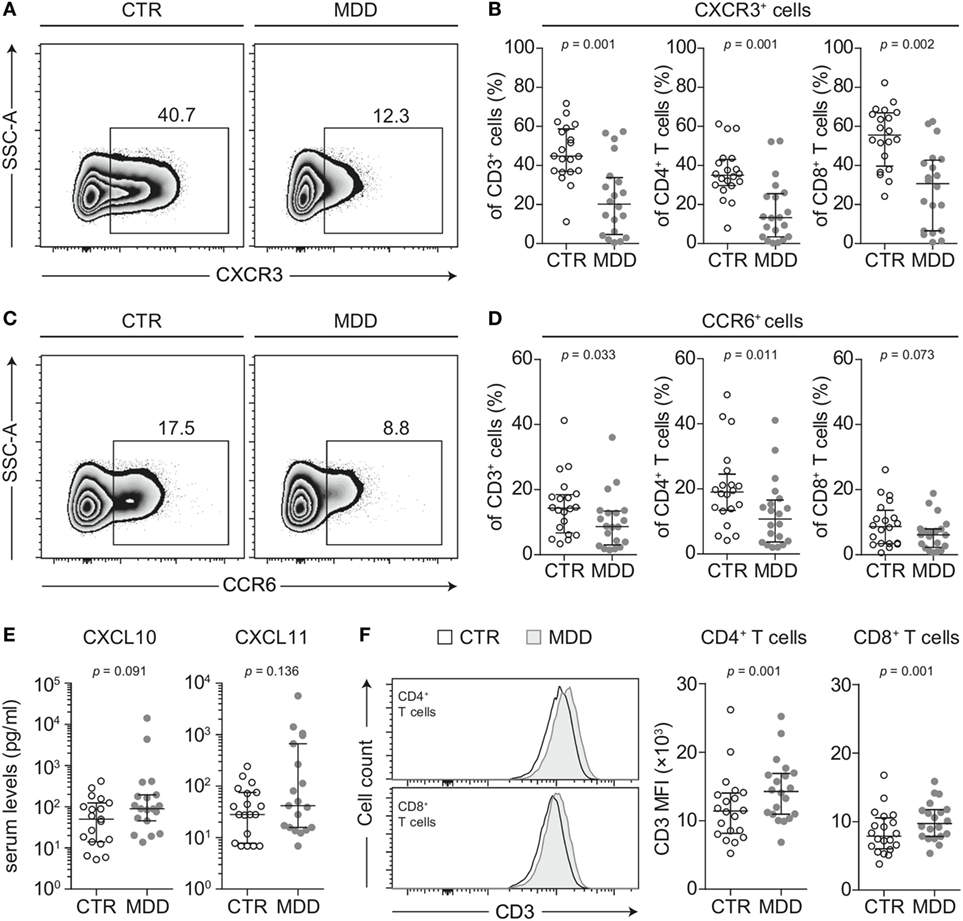
Figure 2. CXCR3 and CCR6 expression in T cells of major depressive disorder (MDD) patients and non-depressed controls. (A) CXCR3-expressing T cells were identified by flow cytometric analysis of peripheral blood mononuclear cells from MDD patients and matched non-depressed controls (CTR). Displayed values are frequencies of CXCR3+ T cells expressed as a percentage of live CD3+ lymphocytes from a representative case–control pair. (B) Percentages of CXCR3-expressing total T cells, CD4+, and CD8+ T cells were quantified in our cohort (n = 40). (C,D) Similar analyses were conducted for the surface expression of CCR6 on total T cells as well as on the CD4+ and CD8+ T cell subsets. (E) The CXCR3 ligands CXCL10 and CXCL11 were quantified in sera of MDD patients and matched controls using a cytometric bead array (n = 38). (F) Surface CD3 MFI levels were measured by flow cytometric analysis of CD4+ and CD8+ T cells from MDD patients and matched controls (n = 40). All graphs depict medians with interquartile ranges. For all comparisons, the Wilcoxon signed-rank test was used. SSC-A, side scatter-area; CXCR3, CXC-chemokine receptor type 3; CCR6, CC-chemokine receptor type 6; MFI, median fluorescence intensity.
Lower surface CXCR3 expression was not T cell-specific, as the percentage of CXCR3-expressing CD3− lymphocytes (non-T cells) was also found to be significantly lower in MDD patients (p = 0.001; Figure 3A). However, lower surface expression of CCR6 was confined to T cells, as we detected no difference in the frequency of CCR6-expressing CD3− lymphocytes between patients and controls (p = 0.60; Figure 3A).
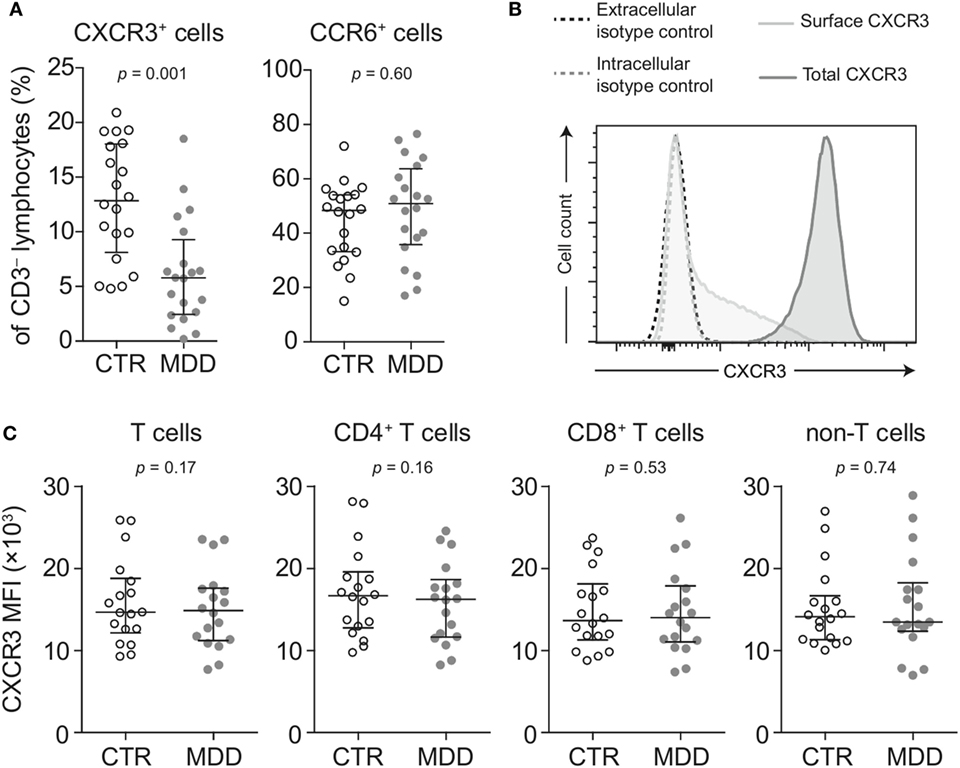
Figure 3. Surface and intracellular staining of CXCR3. (A) Percentages of CXCR3- and CCR6-expressing CD3− lymphocytes (non-T cells) were quantified in major depressive disorder (MDD) patients and matched non-depressed controls (CTR) (n = 40). (B) A representative plot shows fluorescence intensity of CXCR3 expression in intact (surface CXCR3; light gray-shaded curve) relative to fixed and permeabilized T cells (total cellular CXCR3; dark gray-shaded curve). Isotype-matched negative controls were used at the same concentration before fixation (black-dashed curve) and after fixation-permeabilization (gray-dashed curve) and showed no positive staining for CXCR3. (C) Total cellular CXCR3 MFI levels were measured by flow cytometric analysis of fixed and permeabilized peripheral blood mononuclear cells (PBMCs) from MDD patients and matched controls (n = 36). Stained PBMCs were gated on live CD3+ lymphocytes (T cells), CD4+ and CD8+ T cell subsets as well as CD3− lymphocytes (non-T cells). Graphs depict medians with interquartile ranges. For all comparisons, the Wilcoxon signed-rank test was used. CXCR3: CXC-chemokine receptor type 3; CCR6: CC-chemokine receptor type 6; MFI: median fluorescence intensity.
Next, we conducted several analyses to explore potential mechanisms that could underlie the extensive loss of surface CXCR3 expression in MDD. We found no differences in total cellular levels of CXCR3 (i.e., surface and intracellular combined) in T cells and non-T cells (Figures 3B,C), suggesting different receptor turnover at the cell membrane in MDD.
T cells rapidly internalize CXCR3 when exposed to CXCL10 and CXCL11 (29) and there is evidence to suggest that CXCL10 is elevated in MDD (30). In our sample, serum CXCL10 tended to be higher in MDD, although this difference reached only a trend (p = 0.091; Figure 2E). Descriptively, CXCL11 was also elevated but this did not reach statistical significance (Figure 2E). However, the intercorrelation between the two chemokines was robust (Spearman’s rho = 0.862, p < 0.001).
CD3 stimulation can be an alternative mechanism accounting for downregulation of both CXCR3 and CCR6 from the cell surface (31). In line with this hypothesis, we observed higher median fluorescence intensity of CD3 expression in T cells of MDD patients (p = 0.001; Figure 2F). It has been previously described that upregulation of surface CD3 is associated with prolonged TCR engagement (32), which provided further rationale for our hypothesis that MDD patients may bear TCR repertoires of biased clonal composition (see T cell repertoire analyses).
CD4+ T Cell Subsets and Transcription of Master Regulators of T Cell Differentiation
Shifts in CD4+ T cell subsets could indicate another aspect of skewed T cell differentiation in MDD (16). Indeed, we observed that MDD patients exhibited significantly higher frequency of CD25highCD127low/− T cells (p = 0.048; Figures 4A,B), a surface phenotype associated with regulatory T (Treg) cells in humans (19, 33). In addition, in a subset of 10 case–control pairs with sufficient biomaterial available, we observed increased mRNA levels of the Treg transcription factor FOXP3 (p = 0.007), while no significant changes were observed in the levels of mRNA for transcription factors linked to differentiation of the Th1 (T-bet), Th2 (GATA3), or Th17 (RORC) lineages (Figure 4C). Corroborating a relative shift toward a Treg-associated phenotype at the expense of Th1 differentiation, FOXP3 mRNA expression was significantly and positively correlated with CD25highCD127low/− frequency (Spearman’s rho = 0.583, p = 0.007; Figure 4D) but negatively correlated with the frequency of CXCR3-expressing CD4+ T cells (Spearman’s rho = − 0.555, p = 0.011). No correlation was found for CCR6-expressing CD4+ T cells (Spearman’s rho = −0.194, p = 0.41).
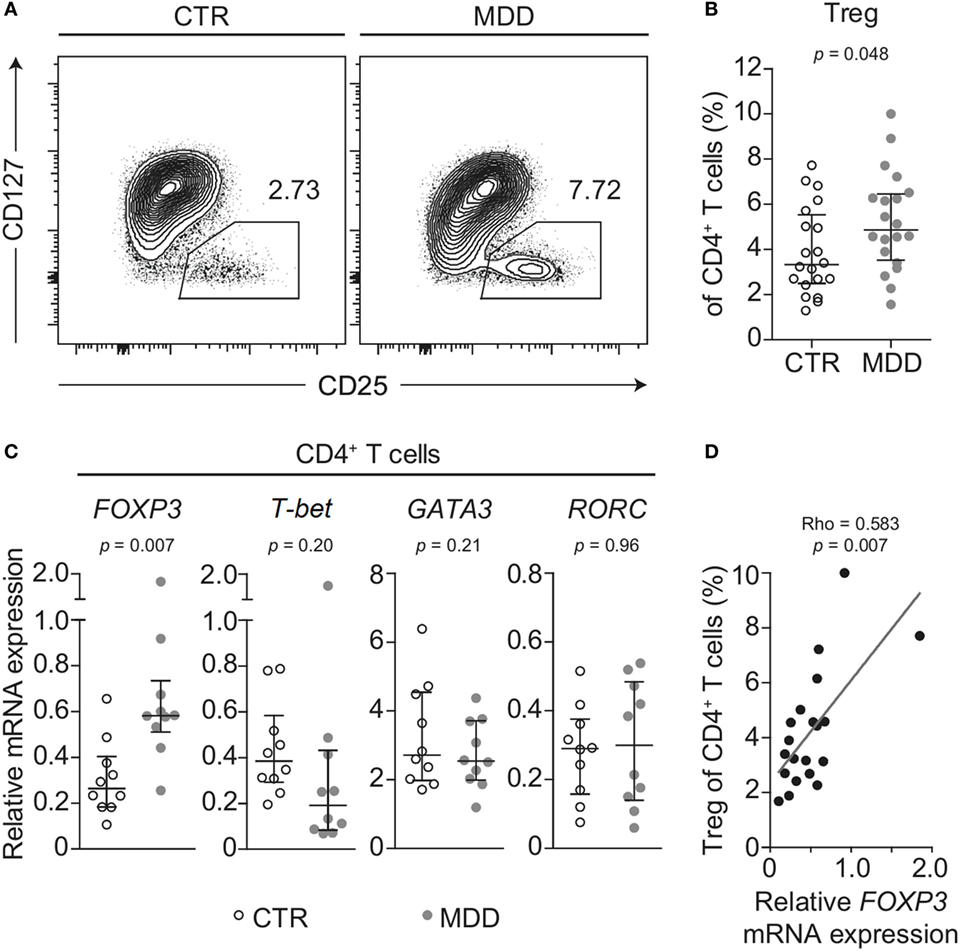
Figure 4. Regulatory T cells in major depressive disorder (MDD) patients and non-depressed controls. (A) Regulatory T cells (Tregs) were identified by flow cytometric analysis of peripheral blood mononuclear cells from MDD patients and matched non-depressed controls (CTR). Displayed values are frequencies of Tregs expressed as a percentage of live CD4+ T cells from a representative case–control pair. (B) Differences in Treg frequency are depicted for the entire cohort (n = 40). (C) Negatively selected CD4+ T cells from a subsample of patients and matched controls (n = 20) were analyzed for mRNA expression of the T helper-associated transcription factors Forkhead box P3 (FOXP3), T-box 21 (T-bet), GATA binding protein 3 (GATA3), and RAR related orphan receptor C (RORC), respectively. Expression was normalized to the geometric mean expression of three housekeeping genes (IPO8, TBP, RPL13A). (D) The correlation between the expression levels of the gene FOXP3 in purified CD4+ T cells and the frequency of Tregs expressed as a percentage of CD4+ T cells is plotted (n = 20). Graphs depict medians with interquartile ranges. For all comparisons, the Wilcoxon signed-rank test was used.
T Cell Repertoire Analysis
In order to explore whether a skewed T cell phenotype in MDD is paralleled by a biased TCR expression profile, we examined TCR Vβ family distribution by flow cytometry-based analysis of the entire cohort (n = 40). Overall, the T cell repertoire of MDD patients showed a trend for higher Gini-TCR indices in CD4+ cells (p = 0.057; Figure 5A), indicating that the CD4+ TCR repertoire might be less evenly distributed in MDD. By contrast, no evidence for skewing of the CD8+ T cell repertoire was observed (p = 0.36; data not shown).
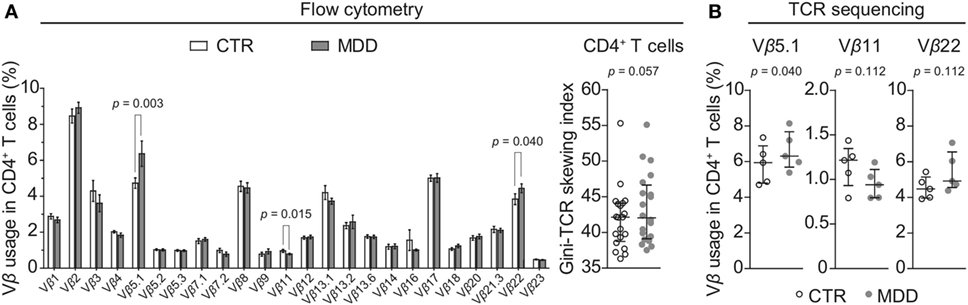
Figure 5. CD4+ T cell repertoire in major depressive disorder (MDD) patients and non-depressed controls. (A) T cell receptor (TCR) variable β chain (Vβ) family distribution analysis was performed by means of flow cytometric interrogation of CD4+ T cells from MDD patients and matched non-depressed controls (CTR) (n = 40). The resulting clonogram represents percentages (mean ± SEM) of the usage of 24 Vβ families. The Gini-TCR skewing index was next applied to the flow cytometric Vβ repertoire analysis (graph on right). Significant post hoc comparisons are denoted for the families Vβ 5.1, Vβ 11, and Vβ 22 (two-tailed, uncorrected p-values). (B) CD4+ T cells were negatively selected from a subsample of patients and matched controls (n = 10) and total genomic DNA was extracted for next generation sequencing of the TCRβ CDR3 repertoire. Usage of the families Vβ 5.1 (TCRBV05-01), Vβ 11 (TCRBV25-01), and Vβ 22 (TCRBV02-01) was then followed up (one-tailed planned comparisons). For all comparisons, the Wilcoxon signed-rank test was used.
Among CD4+ T cells, we found significantly higher usage of the Vβ families 5.1 and 22 (unadjusted p = 0.003 and p = 0.040, respectively; Figure 5A) and lower usage of Vβ 11 (unadjusted p = 0.015; Figure 5A). Next, we sought to confirm these observations by using next-generation sequencing in a subgroup of five case–control pairs, which were chosen in a manner blind to chemokine receptor expression and Gini-TCR index differences (see Table S1 in Supplementary Material). Sequencing of the beta chain CDR3 region in purified CD4+ T cells confirmed the increased usage of Vβ 5.1 (TCRBV05-01) in the MDD group as compared to the control group (p = 0.040, one-tailed planned comparison; Figure 5B). Differences regarding Vβ 11 (TCRBV25-01) and Vβ 22 (TCRBV02-01) showed the same pattern as the flow cytometry analysis in the whole cohort but did not reach statistical significance (Figure 5B). The top five Vβ 5.1 T cell clones for each subject, which were all “private” clones, are displayed in Figure 6.
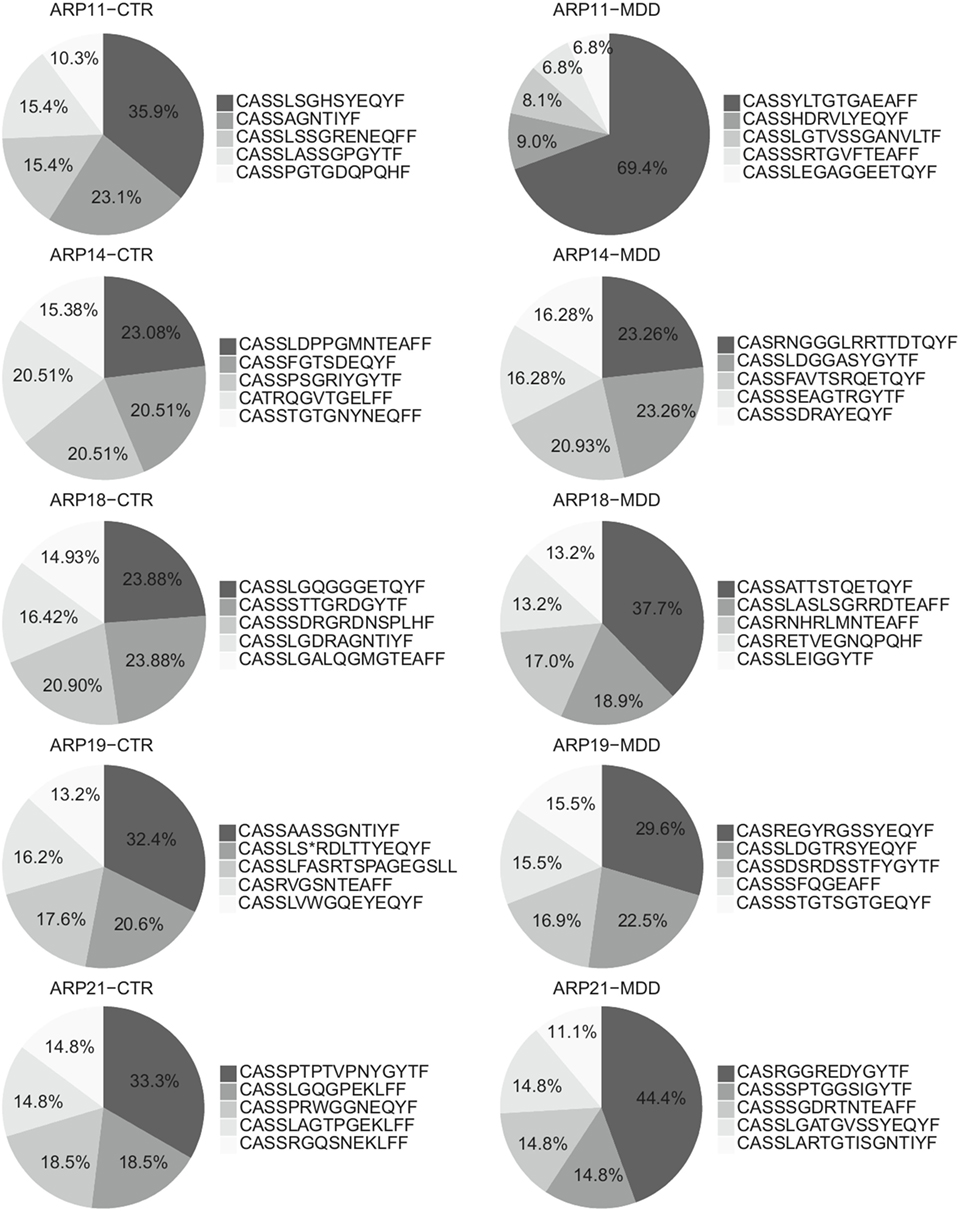
Figure 6. Top Vβ 5.1-expressing clones. Relative frequencies of top five clones expressing Vβ 5.1 (TCRBV05-01) and their CDR3 amino acid sequence in patients with major depressive disorder (MDD) and matched non-depressed controls (CTR) (n = 10).
Association between Immune and Neuroendocrine Variables
In order to explore whether T cell alterations are associated with neuroendocrine dysregulation in MDD patients (34), we conducted correlation analyses between cellular immune measures found to differ between the MDD and control groups and serum levels of stress hormones (ACTH and cortisol). We found no associations of ACTH or cortisol levels with our T cell phenotype and TCR repertoire outcomes (all p > 0.15, data not shown).
Discussion
Here, we provide converging evidence from several cellular and molecular approaches that antidepressant-free MDD patients display biased T cell phenotype and TCR repertoire as compared to closely matched, non-depressed controls. Peripheral blood T cells from MDD patients showed a shift in the CD4+ T compartment toward Treg cells, paralleled by lower surface expression of the T helper differentiation-related chemokine receptors CXCR3 and CCR6. In addition, a less diverse TCR utilization profile was seen within the CD4+ but not the CD8+ T cell subset.
Our cellular immune markers did not show any significant associations with circulating stress hormone levels (ACTH, cortisol); therefore, changes in T cell phenotype and TCR repertoire are unlikely to simply represent an epiphenomenon of neuroendocrine dysregulation in MDD (34). Furthermore, given the range of group differences in surface chemokine receptor expression as well as shifts in CD4+ T cell phenotype, our observations probably reflect a skewing in T cell differentiation, rather than implicating single molecules in MDD pathophysiology. Nevertheless, it is worthwhile to consider the potential functional implications of some of the candidate markers we identify in this study.
CXCR3 is a chemokine receptor that is highly expressed on effector CD4+ and CD8+ T cells and grants them entry into otherwise restricted sites of Th1-type inflammation and infection (35). Although we have not tested the functional consequences of lower surface CXCR3 in MDD directly, it has been previously shown that ligand-induced internalization of this receptor is associated with abolished migratory capacity of both CD4+ and CD8+ human T cells toward a cognate ligand (36). Furthermore, animal studies have shown that CXCR3 enhances the ability of T cells to safeguard against infections (37–39). Whether or not T cells from patients with MDD have a functional impairment with regard to their ability to respond to and clear infections should be, thus, investigated in future studies.
Intriguingly, animal models have shown that virus-induced “sickness-behavior,” which bears symptomatic and immunological similarities to MDD (40, 41), depends on the CXCL10-CXCR3 axis (42). Therefore, lower expression of CXCR3 as reported here might directly be involved in MDD pathophysiology. Along similar lines, a post mortem study showed that CXCR3+ T cells can be found in the human CNS, hinting at a potential role of these cells for maintenance and/or immune surveillance in the brain (43). Whether or not lower surface expression of CXCR3 in peripheral blood T cells might interfere with these functions, however, remains speculative. Ideally, this should further be explored in appropriate animal models, where immune cell trafficking into the CNS as well as peripheral tissues can be monitored.
Another functional implication of our findings could be overly suppressed host T cell responses in MDD owing to a bias toward Treg differentiation. It is worth noting that similar phenomena have been observed in chronic viral infections (44) as well as in animal models of neuropsychiatric disorders (45, 46).
Strengths of our study include the detailed clinical characterization and homogeneity of our patient cohort with moderate to severe MDD but no other psychiatric diagnoses or major somatic comorbidities. Further minimizing potential confounds, our study used close pairwise matching of cases and controls for variables that likely have an impact on immune function, such as sex, age, BMI, and smoking status. Moreover, all participants were non/mild drinkers and we only included currently unmedicated and mostly antidepressant-naïve MDD patients, using a clear cut-off in depression severity (HDRS score of 18 or more).
However, some limitations of our study have to be considered. First, no data were available for other clinical variables that may have influenced immune function, e.g., physical activity and nutrition (47). All MDD subjects were untreated with antidepressants and only four participants (n = 2 in the MDD group and n = 2 in the control group) were receiving non-psychiatric medications that were allowed in the study. Having said that, it is not possible to assess and control all potential additional factors that might be associated with MDD in cross-sectional case–control studies in humans. Thus, future longitudinal studies will be required to better understand the factors linked to the immune abnormalities observed herein. Second, the strict matching and the additional exclusion criteria applied increased the clinical homogeneity of the MDD group, but also led to a comparatively small study sample. This obviously has implications for the generalizability of our findings.
Beyond the advantages of more homogeneous patient groups in research studies, even in this small sample, we quite closely replicated previous immunological findings in MDD [e.g., elevated serum levels of CXCL10 (30)]. Furthermore, our results are in line with a recent whole blood transcriptomic analysis, which identified lower CCR6 transcripts in both antidepressant-treated and antidepressant-free MDD cohorts (48) and our findings on lower NK cell frequency are consistent with lower expression of NK-related genes in MDD (26). Thus, we are confident that our well-characterized cohort is representative of MDD patients.
Our results on higher Treg frequency are consistent with recent reports showing a higher percentage of CD25+CD127lowCCR4+ Tregs in antidepressant-free depressed patients (28) and a positive association between the frequency of CD25highCD127low Tregs and depressive symptoms in older adults following an acute stressor (49). However, our results are in conflict with other previous studies indicating lower frequency of Tregs in MDD patients (27, 50). One possible explanation for this discrepancy could be differences in the clinical characteristics of the study samples (medication status, age, BMI). In addition, methodological differences in Treg definition could also have contributed to these discrepancies so that functional analyses of Treg suppressive capacity will be needed in the future to more specifically determine the role of Tregs in MDD.
In summary, we provide converging evidence from molecular and cellular analyses for a skewed T cell phenotype and CD4+ T cell repertoire in antidepressant-free MDD patients. These findings from our hypothesis-driven study should be confirmed in larger studies and expanded by employing unbiased systems biology approaches. It is important to note that besides MDD, other psychiatric disorders such as schizophrenia have been linked to immune alterations. In schizophrenia, many of the known risk genes are involved in immune regulation (51) and data from animal models, clinical studies, and epidemiology support a role of the immune system in its pathobiology (c.f. (52) for a recent review). Moreover, meta-analyses have confirmed changes in lymphocyte subset counts and frequencies (53) and cytokine levels (54). However, it should be noted that both on a genetic level (55) as well as with regard to immunological parameters such as cytokine levels (56), there is considerable overlap between major psychiatric disorders, including MDD, schizophrenia, and bipolar disorder. Thus, more work is needed to determine if any immune markers are specific to a given disorder or maybe linked to a specific symptom domain observed across diagnostic categories. Ultimately, this may have the potential to open new avenues for research toward an immunotherapy for MDD.
Ethics Statement
This study has been approved by the appropriate Ethics Review Committee (Ethik-Kommission der Ärztekammer Hamburg, Ethikvotum PV4161 and PV4719). All participants provided written informed consent before enrolment in the study.
Author Contributions
Conception and design: KP and SMG. Execution of experiments: KP, AL, CR, TS, CG, KC, and MV. Acquisition of data: KP., CD, LS, and AA. Analysis of data: KP, AW, JE, and AA. Interpretation of data: KP, AW, PA, MF, OP, KW, AA, and SMG. Obtained funding: AA, OP, and SMG. Drafting of the manuscript: KP and SMG. Revision of the manuscript for important intellectual content: AW, CD, JE, AL, CR, TS, CG, LS, KC, MV, PA, MF, OP, KW, and AA.
Conflict of Interest Statement
KP, AW, CD, JE, AL, CR, CG, TS, LS, and KW have no potential conflicts of interest to disclose. PA has received research funding from the Deutsche Forschungsgemeinschaft (DFG). MF has received research funding from the Deutsche Forschungsgemeinschaft (DFG) and the Federal Ministry of Education and Research (BMBF). OP has received research funding from the Federal Ministry of Education and Research (BMBF). AA has received research funding from the Werner Otto Foundation. SG has received honoraria from Mylan GmbH and research funding from the Deutsche Forschungsgemeinschaft (DFG), the Federal Ministry of Education and Research (BMBF), the National MS Society (NMSS), the Werner Otto Foundation, and the European Commission. KC and MV have employment and equity ownership with Adaptive Biotechnologies.
Funding
This study was supported by a grant from the Werner Otto Stiftung Hamburg (grant no. WO15/88 to AA and SMG). The research leading to these results has received funding from the European Union Seventh Framework Program (FP7/2007-2013) under grant agreement n°[268381] (to SMG). Part of this work was supported by the Federal Ministry of Education and Research (BMBF, grant 161A130 to SMG and grant 16GW0082 to OP) and the Deutsche Forschungsgemeinschaft (KFO296, GO1357/8-1 to SMG). KP was in part supported by the Greek Scholarships Foundation (IKY, bequest in memory of Maria Zaoussi). AL was supported by an EFIS/Immunology Letters Short-Term Fellowship and the South East Europe Cooperation, University of Hamburg. SMG is supported by a Heisenberg Professorship from the Deutsche Forschungsgemeinschaft (DFG, GO1357/5-1 and 5-2). Data analysis provided by Adaptive Biotechnologies.
Supplementary Material
The Supplementary Material for this article can be found online at http://www.frontiersin.org/articles/10.3389/fimmu.2018.00291/full#supplementary-material.
References
1. Bromet E, Andrade LH, Hwang I, Sampson NA, Alonso J, de Girolamo G, et al. Cross-national epidemiology of DSM-IV major depressive episode. BMC Med (2011) 9:90. doi:10.1186/1741-7015-9-90
2. Vos T, Barber RM, Bell B, Bertozzi-Villa A, Biryukov S, Bolliger I, et al. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet (2015) 386(9995):743–800. doi:10.1016/S0140-6736(15)60692-4
3. Kupfer DJ, Frank E, Phillips ML. Major depressive disorder: new clinical, neurobiological, and treatment perspectives. Lancet (2012) 379(9820):1045–55. doi:10.1016/S0140-6736(11)60602-8
4. Otte C, Gold SM, Penninx BW, Pariante CM, Etkin A, Fava M, et al. Major depressive disorder. Nat Rev Dis Primers (2016) 2:16065. doi:10.1038/nrdp.2016.65
5. Rush AJ, Trivedi MH, Wisniewski SR, Nierenberg AA, Stewart JW, Warden D, et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D report. Am J Psychiatry (2006) 163(11):1905–17. doi:10.1176/ajp.2006.163.11.1905
6. Cipriani A, Furukawa TA, Salanti G, Geddes JR, Higgins JP, Churchill R, et al. Comparative efficacy and acceptability of 12 new-generation antidepressants: a multiple-treatments meta-analysis. Lancet (2009) 373(9665):746–58. doi:10.1016/S0140-6736(09)60046-5
7. Papakostas GI, Ionescu DF. Towards new mechanisms: an update on therapeutics for treatment-resistant major depressive disorder. Mol Psychiatry (2015) 20(10):1142–50. doi:10.1038/mp.2015.92
8. Miller AH, Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol (2016) 16(1):22–34. doi:10.1038/nri.2015.5
9. Andersson NW, Goodwin RD, Okkels N, Gustafsson LN, Taha F, Cole SW, et al. Depression and the risk of severe infections: prospective analyses on a nationwide representative sample. Int J Epidemiol (2016) 45(1):131–9. doi:10.1093/ije/dyv333
10. Benros ME, Waltoft BL, Nordentoft M, Ostergaard SD, Eaton WW, Krogh J, et al. Autoimmune diseases and severe infections as risk factors for mood disorders: a nationwide study. JAMA Psychiatry (2013) 70(8):812–20. doi:10.1001/jamapsychiatry.2013.1111
11. Leday GGR, Vertes PE, Richardson S, Greene JR, Regan T, Khan S, et al. Replicable and coupled changes in innate and adaptive immune gene expression in two case-control studies of blood microarrays in major depressive disorder. Biol Psychiatry (2017) 83(1):70–80. doi:10.1016/j.biopsych.2017.01.021
12. Kipnis J. Multifaceted interactions between adaptive immunity and the central nervous system. Science (2016) 353(6301):766–71. doi:10.1126/science.aag2638
13. Filiano AJ, Xu Y, Tustison NJ, Marsh RL, Baker W, Smirnov I, et al. Unexpected role of interferon-gamma in regulating neuronal connectivity and social behaviour. Nature (2016) 535(7612):425–9. doi:10.1038/nature18626
14. Herkenham M, Kigar SL. Contributions of the adaptive immune system to mood regulation: mechanisms and pathways of neuroimmune interactions. Prog Neuropsychopharmacol Biol Psychiatry (2016) 79(Pt A):49–57. doi:10.1016/j.pnpbp.2016.09.003
15. Kipnis J, Gadani S, Derecki NC. Pro-cognitive properties of T cells. Nat Rev Immunol (2012) 12(9):663–9. doi:10.1038/nri3280
16. Miller AH. Depression and immunity: a role for T cells? Brain Behav Immun (2010) 24(1):1–8. doi:10.1016/j.bbi.2009.09.009
17. Gold SM, Chalifoux S, Giesser BS, Voskuhl RR. Immune modulation and increased neurotrophic factor production in multiple sclerosis patients treated with testosterone. J Neuroinflammation (2008) 5:32. doi:10.1186/1742-2094-5-32
18. Maecker HT, McCoy JP, Nussenblatt R. Standardizing immunophenotyping for the human immunology project. Nat Rev Immunol (2012) 12(3):191–200. doi:10.1038/nri3158
19. Mahnke YD, Beddall MH, Roederer M. OMIP-015: human regulatory and activated T-cells without intracellular staining. Cytometry A (2013) 83(2):179–81. doi:10.1002/cyto.a.22230
20. Wei S, Charmley P, Robinson MA, Concannon P. The extent of the human germline T-cell receptor V beta gene segment repertoire. Immunogenetics (1994) 40(1):27–36. doi:10.1007/BF00163961
21. Ledderose C, Heyn J, Limbeck E, Kreth S. Selection of reliable reference genes for quantitative real-time PCR in human T cells and neutrophils. BMC Res Notes (2011) 4:427. doi:10.1186/1756-0500-4-427
22. Lefranc MP. IMGT, the international ImMunoGeneTics database. Nucleic Acids Res (2003) 31(1):307–10. doi:10.1093/nar/gkg085
23. Robins HS, Campregher PV, Srivastava SK, Wacher A, Turtle CJ, Kahsai O, et al. Comprehensive assessment of T-cell receptor beta-chain diversity in alphabeta T cells. Blood (2009) 114(19):4099–107. doi:10.1182/blood-2009-04-217604
24. Shugay M, Bagaev DV, Turchaninova MA, Bolotin DA, Britanova OV, Putintseva EV, et al. VDJtools: unifying post-analysis of T cell receptor repertoires. PLoS Comput Biol (2015) 11(11):e1004503. doi:10.1371/journal.pcbi.1004503
25. van der Geest KS, Abdulahad WH, Horst G, Lorencetti PG, Bijzet J, Arends S, et al. Quantifying distribution of flow cytometric TCR-Vbeta usage with economic statistics. PLoS One (2015) 10(4):e0125373. doi:10.1371/journal.pone.0125373
26. Jansen R, Penninx BW, Madar V, Xia K, Milaneschi Y, Hottenga JJ, et al. Gene expression in major depressive disorder. Mol Psychiatry (2016) 21(3):339–47. doi:10.1038/mp.2015.57
27. Grosse L, Hoogenboezem T, Ambree O, Bellingrath S, Jorgens S, de Wit HJ, et al. Deficiencies of the T and natural killer cell system in major depressive disorder: T regulatory cell defects are associated with inflammatory monocyte activation. Brain Behav Immun (2016) 54:38–44. doi:10.1016/j.bbi.2015.12.003
28. Suzuki H, Savitz J, Kent Teague T, Gandhapudi SK, Tan C, Misaki M, et al. Altered populations of natural killer cells, cytotoxic T lymphocytes, and regulatory T cells in major depressive disorder: association with sleep disturbance. Brain Behav Immun (2017) 66:193–200. doi:10.1016/j.bbi.2017.06.011
29. Meiser A, Mueller A, Wise EL, McDonagh EM, Petit SJ, Saran N, et al. The chemokine receptor CXCR3 is degraded following internalization and is replenished at the cell surface by de novo synthesis of receptor. J Immunol (2008) 180(10):6713–24. doi:10.4049/jimmunol.180.10.6713
30. Wong ML, Dong C, Maestre-Mesa J, Licinio J. Polymorphisms in inflammation-related genes are associated with susceptibility to major depression and antidepressant response. Mol Psychiatry (2008) 13(8):800–12. doi:10.1038/mp.2008.59
31. Sallusto F, Kremmer E, Palermo B, Hoy A, Ponath P, Qin S, et al. Switch in chemokine receptor expression upon TCR stimulation reveals novel homing potential for recently activated T cells. Eur J Immunol (1999) 29(6):2037–45. doi:10.1002/(SICI)1521-4141(199906)29:06<2037::AID-IMMU2037>3.0.CO;2-V
32. Schrum AG, Turka LA, Palmer E. Surface T-cell antigen receptor expression and availability for long-term antigenic signaling. Immunol Rev (2003) 196:7–24. doi:10.1046/j.1600-065X.2003.00083.x
33. Wingender G, Kronenberg M. OMIP-030: characterization of human T cell subsets via surface markers. Cytometry A (2015) 87(12):1067–9. doi:10.1002/cyto.a.22788
34. Stetler C, Miller GE. Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosom Med (2011) 73(2):114–26. doi:10.1097/PSY.0b013e31820ad12b
35. Groom JR, Luster AD. CXCR3 in T cell function. Exp Cell Res (2011) 317(5):620–31. doi:10.1016/j.yexcr.2010.12.017
36. Winter D, Moser J, Kriehuber E, Wiesner C, Knobler R, Trautinger F, et al. Down-modulation of CXCR3 surface expression and function in CD8+ T cells from cutaneous T cell lymphoma patients. J Immunol (2007) 179(6):4272–82. doi:10.4049/jimmunol.179.6.4272
37. Hickman HD, Reynoso GV, Ngudiankama BF, Cush SS, Gibbs J, Bennink JR, et al. CXCR3 chemokine receptor enables local CD8(+) T cell migration for the destruction of virus-infected cells. Immunity (2015) 42(3):524–37. doi:10.1016/j.immuni.2015.02.009
38. Sakai S, Kauffman KD, Schenkel JM, McBerry CC, Mayer-Barber KD, Masopust D, et al. Cutting edge: control of Mycobacterium tuberculosis infection by a subset of lung parenchyma-homing CD4 T cells. J Immunol (2014) 192(7):2965–9. doi:10.4049/jimmunol.1400019
39. Cohen SB, Maurer KJ, Egan CE, Oghumu S, Satoskar AR, Denkers EY. CXCR3-dependent CD4(+) T cells are required to activate inflammatory monocytes for defense against intestinal infection. PLoS Pathog (2013) 9(10):e1003706. doi:10.1371/journal.ppat.1003706
40. Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci (2008) 9(1):46–56. doi:10.1038/nrn2297
41. Maes M, Berk M, Goehler L, Song C, Anderson G, Galecki P, et al. Depression and sickness behavior are Janus-faced responses to shared inflammatory pathways. BMC Med (2012) 10:66. doi:10.1186/1741-7015-10-66
42. Blank T, Detje CN, Spiess A, Hagemeyer N, Brendecke SM, Wolfart J, et al. Brain endothelial- and epithelial-specific interferon receptor chain 1 drives virus-induced sickness behavior and cognitive impairment. Immunity (2016) 44(4):901–12. doi:10.1016/j.immuni.2016.04.005
43. Smolders J, Remmerswaal EB, Schuurman KG, Melief J, van Eden CG, van Lier RA, et al. Characteristics of differentiated CD8(+) and CD4 (+) T cells present in the human brain. Acta Neuropathol (2013) 126(4):525–35. doi:10.1007/s00401-013-1155-0
44. Li S, Gowans EJ, Chougnet C, Plebanski M, Dittmer U. Natural regulatory T cells and persistent viral infection. J Virol (2008) 82(1):21–30. doi:10.1128/JVI.01768-07
45. Baruch K, Deczkowska A, Rosenzweig N, Tsitsou-Kampeli A, Sharif AM, Matcovitch-Natan O, et al. PD-1 immune checkpoint blockade reduces pathology and improves memory in mouse models of Alzheimer’s disease. Nat Med (2016) 22(2):135–7. doi:10.1038/nm.4022
46. Cohen H, Ziv Y, Cardon M, Kaplan Z, Matar MA, Gidron Y, et al. Maladaptation to mental stress mitigated by the adaptive immune system via depletion of naturally occurring regulatory CD4+CD25+ cells. J Neurobiol (2006) 66(6):552–63. doi:10.1002/neu.20249
47. Irwin MR, Miller AH. Depressive disorders and immunity: 20 years of progress and discovery. Brain Behav Immun (2007) 21(4):374–83. doi:10.1016/j.bbi.2007.01.010
48. Powell TR, McGuffin P, D’Souza UM, Cohen-Woods S, Hosang GM, Martin C, et al. Putative transcriptomic biomarkers in the inflammatory cytokine pathway differentiate major depressive disorder patients from control subjects and bipolar disorder patients. PLoS One (2014) 9(3):e91076. doi:10.1371/journal.pone.0091076
49. Ronaldson A, Gazali AM, Zalli A, Kaiser F, Thompson SJ, Henderson B, et al. Increased percentages of regulatory T cells are associated with inflammatory and neuroendocrine responses to acute psychological stress and poorer health status in older men and women. Psychopharmacology (2016) 233(9):1661–8. doi:10.1007/s00213-015-3876-3
50. Grosse L, Carvalho LA, Birkenhager TK, Hoogendijk WJ, Kushner SA, Drexhage HA, et al. Circulating cytotoxic T cells and natural killer cells as potential predictors for antidepressant response in melancholic depression. Restoration of T regulatory cell populations after antidepressant therapy. Psychopharmacology (Berl) (2016) 233(9):1679–88. doi:10.1007/s00213-015-3943-9
51. Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature (2014) 511(7510):421–7. doi:10.1038/nature13595
52. Khandaker GM, Cousins L, Deakin J, Lennox BR, Yolken R, Jones PB. Inflammation and immunity in schizophrenia: implications for pathophysiology and treatment. Lancet Psychiatry (2015) 2(3):258–70. doi:10.1016/S2215-0366(14)00122-9
53. Miller BJ, Gassama B, Sebastian D, Buckley P, Mellor A. Meta-analysis of lymphocytes in schizophrenia: clinical status and antipsychotic effects. Biol Psychiatry (2013) 73(10):993–9. doi:10.1016/j.biopsych.2012.09.007
54. Miller BJ, Buckley P, Seabolt W, Mellor A, Kirkpatrick B. Meta-analysis of cytokine alterations in schizophrenia: clinical status and antipsychotic effects. Biol Psychiatry (2011) 70(7):663–71. doi:10.1016/j.biopsych.2011.04.013
55. Network, Pathway Analysis Subgroup of Psychiatric Genomics Consortium. Psychiatric genome-wide association study analyses implicate neuronal, immune and histone pathways. Nat Neurosci (2015) 18(2):199–209. doi:10.1038/nn.3922
Keywords: adaptive immunity, major depressive disorder, chemokine receptors, regulatory T cells, T cell receptor repertoire
Citation: Patas K, Willing A, Demiralay C, Engler JB, Lupu A, Ramien C, Schäfer T, Gach C, Stumm L, Chan K, Vignali M, Arck PC, Friese MA, Pless O, Wiedemann K, Agorastos A and Gold SM (2018) T Cell Phenotype and T Cell Receptor Repertoire in Patients with Major Depressive Disorder. Front. Immunol. 9:291. doi: 10.3389/fimmu.2018.00291
Received: 06 November 2017; Accepted: 01 February 2018;
Published: 20 February 2018
Edited by:
Jens Geginat, Istituto Nazionale Genetica Molecolare (INGM), ItalyReviewed by:
Mario (Mago) Clerici, Università degli Studi di Milano, ItalyRoberto Furlan, San Raffaele Hospital (IRCCS), Italy
Copyright: © 2018 Patas, Willing, Demiralay, Engler, Lupu, Ramien, Schäfer, Gach, Stumm, Chan, Vignali, Arck, Friese, Pless, Wiedemann, Agorastos and Gold. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stefan M. Gold, stefan.gold@charite.de
†These authors have contributed equally to this work.