Sediment Composition and Facies of Coral Reef Islands in the Spermonde Archipelago, Indonesia
 Alexander Janßen1,2*
Alexander Janßen1,2*  André Wizemann2
André Wizemann2  André Klicpera2 Dewi Y. Satari3 Hildegard Westphal2,4
André Klicpera2 Dewi Y. Satari3 Hildegard Westphal2,4  Thomas Mann2
Thomas Mann2- 1Center for Marine Environmental Sciences, Bremen, Germany
- 2Leibniz Centre for Tropical Marine Research, Bremen, Germany
- 3Research and Development Center for Marine, Coastal, and Small Islands, Hasanuddin University, Makassar, Indonesia
- 4Faculty of Geosciences, University of Bremen, Bremen, Germany
Sedimentological and geomorphological characteristics of coral reef islands are strongly related to past and recent boundary conditions such as the hydrodynamic regime, wind directions, sea-level fluctuations, and the ecological footprint of the surrounding reef complexes. Alterations in the physical, chemical, and biological boundary controls may affect the stability of reef islands. Additionally, these factors are of importance in the context of future climate change. Such alterations through time may well be documented within the sedimentary record of reef islands and a better knowledge on its effects could help to improve our understanding of island responses to future changes of the status quo. However, detailed studies on the sedimentology and geomorphology of reef islands from southwest Sulawesi, Indonesia, are still rare. Here we report on the sedimentary composition and related facies zonation of four uninhabited coral reef islands in the Spermonde Archipelago. Sediment samples from onshore- and reef-flat environments were analyzed in regard to their grain size, component assemblages and facies distribution. Our results show that the analyzed island sediments are characterized by medium- to coarse-grained sand fractions and are well to poorly sorted. Across all islands examined, the surface sediment is predominately composed of materials identified as scleractinian coral and coralline red algae fragments, with minor additions from bivalves, gastropods and foraminifers. Importantly, statistical analyses of the variations in the percentage of these components allow for a clear sedimentary distinction of the four study sites into three outer shelf islands, situated closer to the open marine Makassar Strait, and one inner shelf island. On the inner shelf island, additional subsurface sedimentological analyses indicate a potential shift in major sediment contributors through time, preserved as coral-dominated accumulations within the subsurface samples, and coralline red algae-dominated deposits on top. These findings highlight the practical use of detailed sedimentological studies for the reconstruction of environmental changes in the Spermonde Archipelago.
Introduction
Coral reef islands are low-lying accumulations of largely unconsolidated carbonate sediments. They are typically situated on top of lithified reef substrates and surrounded by a living reef crest. In contrast to continental coastlines in higher latitudes, tropical beaches and reef islands are predominately composed of carbonate sediments supplied by these adjacent reefs. Coral reef islands form in open ocean settings, such as atolls, or in shallow marine environments, surrounded by barrier reefs. The formation and evolution of reef islands is strongly connected to the surrounding reef systems, as these are the source of sediments that accumulate to form an island (Stoddart and Steers, 1977; Yamano et al., 2005). Today most tropical reef islands are strongly affected by human induced climate change and rising sea levels which makes the knowledge about reef island building processes a valuable tool for predicting future stability of these landforms.
Reef island sediments generally consist of a variety of skeletal fragments, shells and hard-parts of reef-forming and reef-associated calcifying organisms. Sediments therefore either represent direct remains from, e.g., corals, coralline red algae, green algae (Halimeda), foraminifers, molluscs, and bryozoans, or derive from the activity of bioeroders such as boring sponges, parrotfish, and sea urchins (Perry et al., 2011; Kench et al., 2012). During transport, the hydrodynamic behavior of these biologically derived sediments is influenced by changes in grain size and morphology due to post-mortem abrasion and breakdown (Yamano et al., 2000; Perry et al., 2011; Ford and Kench, 2012). The decrease in grain size toward a suitable fraction for sandy reef island sediments is time dependent and therefore, a time lag exists between potential ecological shifts of the surrounding reef community composition and island morphological responses (Morgan and Kench, 2016). However, as species presence and abundance in the surrounding reef ecosystem control the type and initial grain size of the sediments as well as the rate of sediment supply, ecological transitions are a key influence on future island resilience (Perry et al., 2011).
In order to better understand ecological variability through time and its effects on sedimentation patterns, the sedimentary facies of four uninhabited reef islands in the Spermonde Archipelago has been investigated. Based on sedimentological analysis and interpretations of the facies association, we show that the composition of island sediments is determined by their geographic position on the archipelago (i.e., proximal vs. distal). Additionally, the reefal ecology of uninhabited islands also is influenced by nearby local sources of nutrients, as well as leisure and building activities taking place on neighboring inhabited islands. This may result in increased seawater turbidity and eutrophication affecting the reef system. This study provides implications for the usefulness of sedimentological studies in terms of reconstructions of the environmental and anthropogenic history of islands in the Spermonde Archipelago and similar settings.
Study Site
The Spermonde Shelf is located between 4 and 6°S in the southern Makassar Strait, west of Makassar in SW Sulawesi (Figure 1A). It covers an area of about 16,000 km2 with a westward increasing water depth down to 60 m below mean sea level (mbsl) where the shelf reaches a physical barrier (Figure 1B). These barrier reefs form an area of more shallow water, 15–20 mbsl with some islands, shoals and islets on top, separating the Spermonde platform from the deeper Makassar Strait (Renema and Troelstra, 2001). A couple of east-west-trending channels incise the shelf and additionally cut through the barrier (Wilson and Vecsei, 2005).
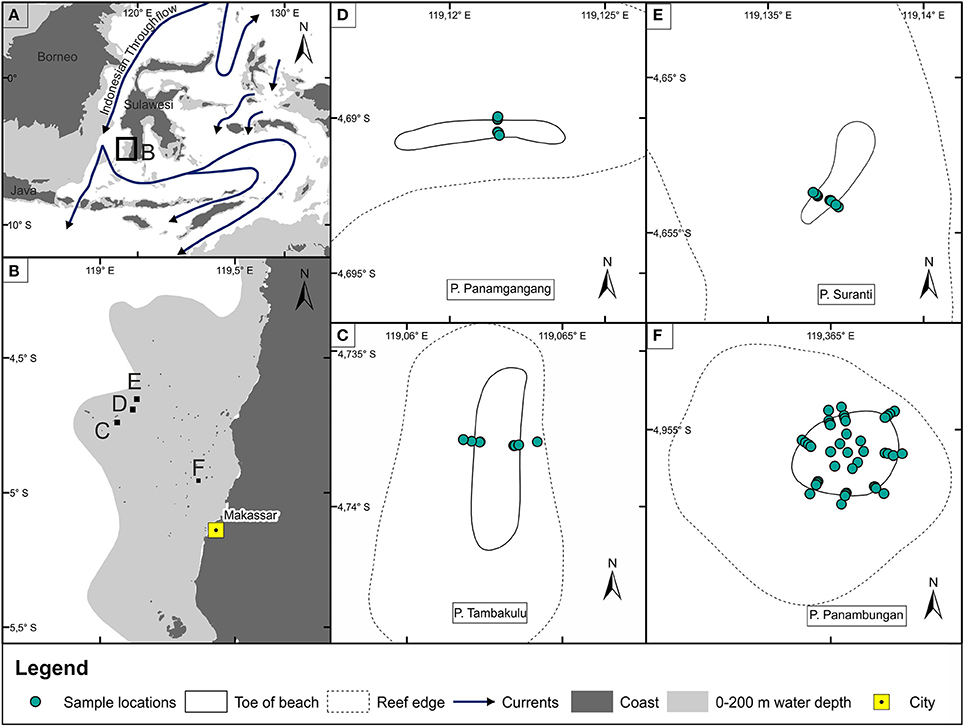
Figure 1. Overview map of (A) Southeast Asia with the major ocean currents (adopted from Ding et al., 2013). (B) Spermonde Archipelago showing the locations of the four investigated islands. (C–F) Study islands Pulau Tambakulu, Pulau Panamgangang, Pulau Suranti and Pulau Panambungan and locations of the surface sediment samples (green points).
The main characterizing feature of the Spermonde Archipelago is the occurrence of some 120 reef islands and sand cays. This study presents the results of a detailed sedimentological study on four of them. These four islands are Pulau Tambakulu (Figure 1C), Pulau Panamgangang (Figure 1D), Pulau Suranti (Figure 1E), and Pulau Panambungan (Figure 1F). P. Panamgangang, P. Suranti, and P. Tambakulu are situated along the outer rim of the Spermonde Shelf, while P. Panambungan is located in the inner shelf setting, about 19 km off Makassar harbor (Figure 1B).
Within the Makassar Strait, the Indonesian Through-Flow (Figure 1A) occurs and acts as an important heat exchange between the Pacific and the Indian ocean (Ding et al., 2013). This current shows a high temporal variability, due to the monsoonal changes in mean wind directions (Figure 2, Mayer and Damm, 2012). The NW monsoon during boreal winter, especially in December and January, accelerates the surface flow to the south. In turn, a direct SE monsoon wind direction slows down the southward flow. However, the rimmed shelf structure of the Spermonde Archipelago protects the inner shelf from stronger hydrodynamic energy settings (Renema and Troelstra, 2001).
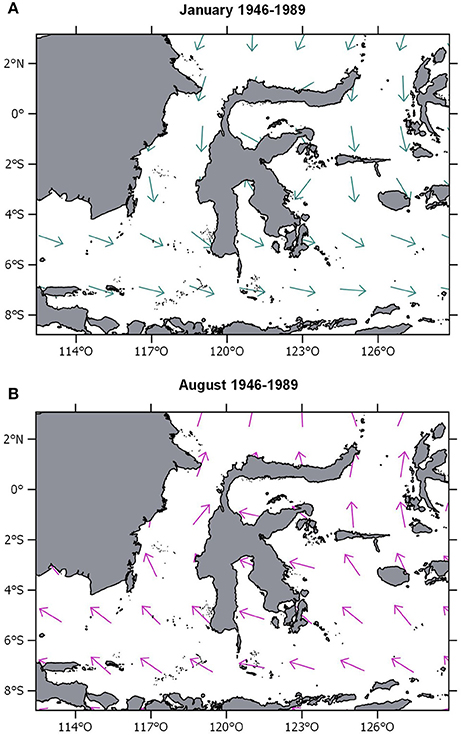
Figure 2. (A) Map of Sulawesi with monsoon winds toward east-southeast during wet season (mean wind direction January 1946–1989). (B) Map of Sulawesi with monsoon winds toward north-northwest during dry season (mean wind direction August 1946–1989). Wind data derived from COADS Monthly Climatology dataset (Woodruff et al., 1998).
Important for the geomorphology of reef islands in the Spermonde are monsoonal wind patterns (Wijsman-Best et al., 1982). The climate in the Spermonde Archipelago is tropical and varies little during the seasons. Only the seasonal change in rainfall has an impact on the regional climate. From June to October the dry monsoon prevails, while from November to March the rainy monsoon predominates. The annual precipitation varies between 2,100 mm in the rainy and around 200 mm in the dry season (Moore, 1995). The wind direction is east-southeast during dry monsoon, and north-northwest, during rainy monsoon (Manjunatha et al., 2015). During the NW monsoon the shelf experiences stronger waves (Wilson and Vecsei, 2005). During the dry season, the south-eastern winds reaching the Spermonde are weaker due to the fact that the shelf area is sheltered by the Sulawesi mainland and oceanic swell is smaller (Hoeksema, 1990).
Fluvial influence onto the Spermonde Shelf is related to the discharge of the two major rivers Maros in the north, and Jene Berang in the southern part of the shelf (Renema and Troelstra, 2001). Discharged river water contains volcanic erosion products from the hinterland and sewage from Makassar (Renema et al., 2001). The fluvial influence in terms of sedimentation on the shelf is largely restricted to a zone not extending 4 km from the coast (Storm, 1989).
The Spermonde Shelf is affected by two different tidal regimes, the semidiurnal and the diurnal tide (Ray et al., 2005). Along the Spermonde Shelf the semidiurnal tide has an amplitude of about 10–30 cm and propagates northwest through Banda and Flores seas, and the Strait of Makassar (see Figure 1, Ray et al., 2005). In contrast, the diurnal tide passes southwards through the Makassar Strait, with an amplitude of about 25–30 cm at the Spermonde shelf and a maximum springtide of 1.5 m measured at the harbor of Makassar (Ray et al., 2005; Meteo365.com Ltd., 2015).
Besides natural forcing, similar to other seascapes across Indonesia, anthropogenic activities steadily have increased in the Spermonde Archipelago. Today, most islands located on the mid and inner Spermonde shelf are populated (Burke et al., 2012). Inhabited islands potentially influence and alter reef ecosystems and frameworks of nearby uninhabited islands.
Materials and Methods
Sixty-nine surface sediment samples were collected in October-November 2012 and 2013 for further analysis on the study islands P. Suranti (n = 9), P. Panamgangang (n = 10), P. Tambakulu (n = 10), and P. Panambungan (n = 40; Figure 1). The exact sample positions were recorded using a Garmin Montana 650 handheld GPS (see Supplementary Material). Samples were taken along transects across the islands, from the reef flat close to the toe of the beach to the top of the island. Bulk sediments samples of ~200 g each were carefully collected from the top ~5 cm of the substrate with a hand-shovel (cf., Wizemann et al., 2015). On P. Panambungan, additionally 42 subsurface sediment samples were extracted along four vertical sections [one section in the center of the island (CS) and three sections at the periphery (S1–S3)] down to 1.25 m below surface (Figure 3, CS: n = 9, S1: n = 14, S2: n = 10, S3: n = 9). Geomorphological information on P. Panambungan was obtained by surveys along three transects using an automatic level. In Figure 3, all elevations are displayed in relation to present-day mean sea level (see Mann et al., 2016 for details).
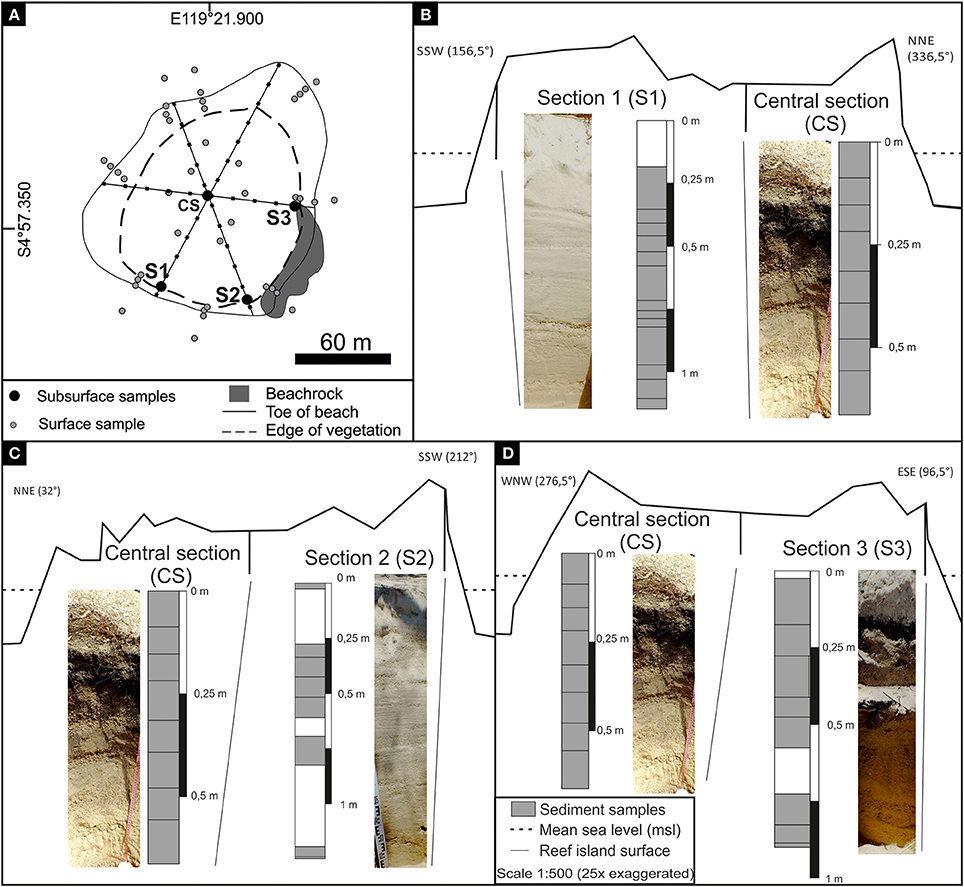
Figure 3. (A) Map of P. Panambungan indicating surface and subsurface sampling locations. The subsurface sampling location Central section (CS) is situated in the center of the island. Transects display the geomorphology of P. Panambungan. (B) One transect crosses the island from SSW to NNE, with subsurface sampling location Section 1 (S1) and CS. (C) The NNE-SSW trending transect includes subsurface sampling location CS and Section 2 (S2). (D) In WNW-ESE trending transect subsurface sampling locations Section 3 (S3) and CS are covered. Subsurface sampling locations are shown with corresponding sampling site image and a profile bar, with gray areas indicating sampling depths.
Each sediment sample was washed with distillated water to dissolve salt enrichments, and oven-dried for 48 h at 40°C. To sieve the samples, each charge was split into a working- and an archive-half. The working sample was then sieved into the fractions >2,000, 2,000–1,000, 1,000–500, 500–250, 250–125, 125–63, and <63 μm, according to the Wentworth (1922) classification. The dry sieving was done using a vibratory sieving machine (Retsch AS 200) with an amplitude of 2.00 mm for 3 min sieving time. The results of the weighting process were added to a cumulative grain size distribution following Wentworth (1922). The statistical evaluations were processed using the G2SD package in R (Fournier et al., 2014).
Components for all fractions >125 μm were analyzed and, where possible, determined. For each sample, components were counted using a binocular microscope (Leica S6E) and a picking tray. Grains with diameters <125 μm were not investigated, due to the limited identification potential of such small skeletal fragments. Unidentifiable carbonate fragments were counted as “bioclasts.” For each fraction, at least 100 individual carbonate components were identified (i.e., excluding bioclasts). In the >2,000 μm fraction, when the minimum number of 100 grains was not reached, all identified grains were counted (i.e., including bioclasts). Results of the component analyses have been statistically examined for zonal patterns using the cluster package in R (Maechler et al., 2016).
Results
Grain Size Characteristics of Surface Sediments
The 69 surface sediment samples show a grain size distribution from gravel to fine sand. Within this grain size range, sandy gravel (>30% gravel) and gravelly sand (5–30% gravel) are the most abundant textural groups. Less frequent grain size groups are gravel (>80% gravel) and slightly gravelly sand (<5% gravel). In general, the analyzed surface sediment samples are well to poorly sorted (Table 1). In the following the sediment characteristics of the surface samples for each location (see Figure 1) are described in detail.
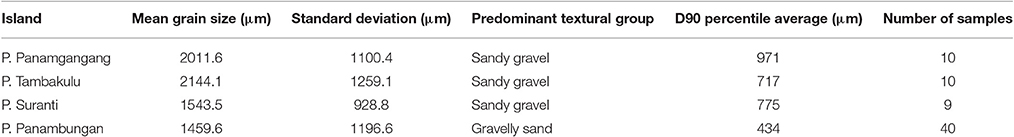
Table 1. Mean grain size distribution of surface sediments on P. Suranti, P. Tambakulu, P. Panamgangang, and P. Panambungan.
P. Panamgangang
P. Panamgangang (Figure 1F) is covered by very coarse sand, with moderately well to moderate sorting (50% of the analyzed samples). Ninety percent of the analyzed grains are coarser than 971 μm on average (see D90 value in Table 1 and Figure 4). Very fine-grained, poorly to well-sorted sediment is found in 30% of the samples, whereas coarse sediment with moderately well and poor sorting characterizes another 20%.
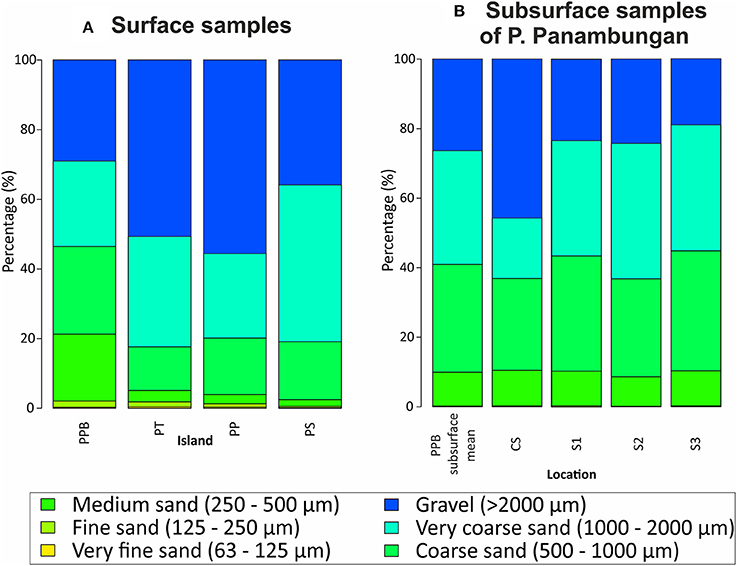
Figure 4. (A) Mean grain size distribution of surface sediment samples from the four study islands P. Panambungan (PPB), P. Tambakulu (PT), P. Panamgangang (PP) and P. Suranti (PS). (B) Mean grain size distribution of subsurface sediment samples (CS, T1–T3) from P. Panambungan.
P. Tambakulu
P. Tambakulu comprises coarse sandy sediment, with 90% of grains that are coarser than 734 μm on average (see D90 value in Table 1 and Figure 4). Very fine gravel and very coarse sand are the main sediment fractions of the surface samples from P. Tambakulu. The sediment is moderately to poorly sorted and is found in eight of ten samples, four each grain size group. Coarse sand is found in two sample locations with poorly and well-sorted specimens.
P. Suranti
On P. Suranti, 72% of the samples are classified within the group of very coarse sand, with 90% of grains coarser than 775 μm on average (see D90 value in Table 1 and Figure 4). Sorting ranges from moderately well to poor, with one well-sorted sample.
P. Panambungan
P. Panambungan (Figure 1F) is predominantly covered by coarse and very coarse sand, with 90% of grains that are coarser than 434 μm on average (see D90 value in Table 1 and Figure 4). The sediments show a moderate to poor sorting. Medium sand and fine gravel fractions occur in seven sample locations on P. Panambungan. Both the medium sand and fine gravel fractions are moderately to moderately well-sorted.
Grain Size Characteristics of Subsurface Sediments of P. Panambungan
Subsurface sediment from P. Panambungan shows similar characteristics as the surface sediment samples, with respect to the grain size distribution. Coarse sand with moderate to moderate well-sorting is the major fraction (45% of the samples). In the following the sediment characteristics of the subsurface samples for each section are described in detail.
Central Section (CS)
Sampling location CS is located in the center of P. Panambungan and shows a down-pit grain-size variability from gravel to slightly gravelly sand. The bottom layer is characterized by gravelly sand, followed upwards by slightly gravelly sand, sandy gravel, and gravelly sand. At 33 and 65 cm below surface CS shows very fine poorly sorted gravel (cf. Table 2 and Figure 3). In a sample, 12 cm below the surface, a relatively coarse layer occurs that is texturally classified as gravel. Poorly sediment sorting dominates in the layers of gravelly sand and sandy gravel (Figure 4).
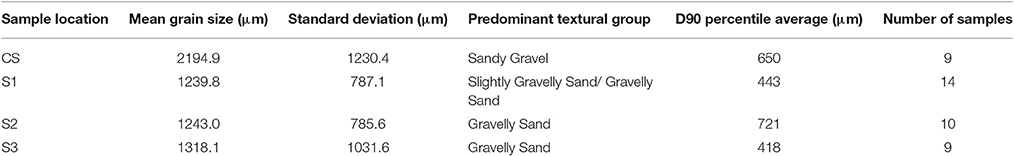
Table 2. Average grain size distribution from subsurface sampling location at of P. Panambungan.
Section 1 (S1)
Slightly gravelly sand and gravelly sand are composing 70% of the subsurface samples in S1. Sandy gravel occurs in 30% of the samples (Figures 3, 4). At the base of S1, moderately sorted gravelly sand occurs, followed by a layer of moderately well-sorted, slightly gravelly sand. Upwards, poorly sorted sandy gravel is covered by moderately sorted, slightly gravelly sand and poorly sorted sandy gravel. This is followed by poorly sorted gravel and moderately well-sorted, slightly gravelly sand interrupted by moderately sorted gravelly sand. Toward the top of the profile, poorly sorted gravelly sand is covered by moderately sorted gravelly sand and moderately well-sorted slightly gravelly sand (Figure 4).
Section 2 (S2)
In samples from sediment sampling location S2, gravelly sand is the most abundant textural group comprising 60% of the samples (Table 2 and Figures 3, 4). The basis of S2 is represented by poorly sorted gravelly sand and covered by a moderately sorted, slightly gravelly sand layer (Figure 4). At 67–55 cm below surface S2 is composed of well-sorted very coarse sand (cf., Table 2 and Figure 3).
Section 3 (S3)
Sediment samples from subsurface location S3 show a relatively uniform grain size distribution of gravelly sand (Table 2 and Figures 3, 4). At 77–75 cm below surface S3 shows very fine poorly sorted gravel (cf. Table 2 and Figure 3) Generally, sandy gravel and gravelly sand is present throughout the entire sampling depth with changing layer thicknesses. This co-occurs with a moderate to poor sorting.
Composition of Surface Sediments
The main component of the surface sediments, across all investigated islands are fragments of coralline algae, followed by coral remains which exhibit however a high variation in their abundance (Figure 5). In the following the mean sediment composition of the surface samples for each reef island are described in detail.
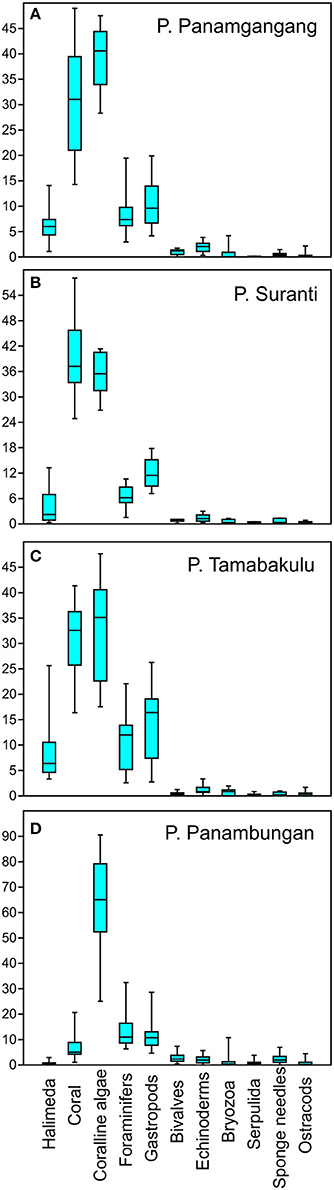
Figure 5. Box- and Whisker plots indicating the mean percentage distribution of surface sediment components from the four study islands (A) P. Panamgangang, (B) P. Tambakulu, (C) P. Suranti, and (D) P. Panambungan.
P. Panamgangang
The surface sediments of P. Panamgangang are on average composed of 29.7% of coral, 39.5% of coralline algae, 11.0% of gastropods, 8.6% of foraminifer fragments, 6.5% of Halimeda segments, and 1.0% of bivalves. Minor constituents are echinoderms with 1.9%, bryozoans with 0.8%, serpulid tubes with 0.1%, sponge needles with 0.5% and ostracods with 0.4%.
P. Suranti
39.5% of the average surface sedimentary composition on P. Suranti originates from coral, followed by 35.2% of coralline algae, 12.1% of gastropods, and 6.4% of foraminifer remains (Figure 5). Halimeda contribute with an average of 4.0% to the surface sediments and bivalves 0.8%. Echinoderms contribute with 1.4%, bryozoans with 0.5%, serpulid tubes with 0.2%, sponge needles with 0.6%, and ostracods with 0.3% contribute to the sediment.
P. Tambakulu
Surface sediments from P. Tambakulu are composed of 30.2% of coral, 33.8% of coralline algae, 13.4% of gastropod, and 10.7% of foraminifer shells and fragments. Furthermore, Halimeda contribute by an average of 8.6%, and bivalves with 0.4% to the sediment composition. Minor sediment constituents are echinoderms with 1.1%, bryozoans with 0.7%, serpulid tubes with 0.2%, sponge needles with 0.4%, and ostracods with 0.5%.
P. Panambungan
Surface sediments at P. Panambungan are mainly composed of coralline algae on the surface with 60.1% of the mean surface sediments being coralline algae fragments. 12.4% of the components are foraminifer shells, 10.8% are gastropods and 6.7% are coral fragments. Bivalve shell fragments contribute to the surface sediment by an average of 2.7%. Halimeda segments contribute by a mean of 0.7% to the surface sediment. Echinoderms contribute with 1.9%, bryozoans with 1.2%, serpulid tubes with 0.7%, sponge needles with 2.2%, and ostracods with 0.7% to the sediment.
Composition of Subsurface Sediments of P. Panambungan
Mean subsurface sediment composition on P. Panambungan is coral-dominated, yet with differences depending on the sampling locality (Figure 6). Samples from the central sample location CS are characterized by mean values of 80.2% of coral fragments. This is followed by gastropod shells (9.6% mean) and fragments from coralline algae (5.6% mean). Sampling location S1 is composed by an average of 70.9% of coral remains, 12.8% coralline algae, and 8.6% gastropods. Similar to the subsurface sediment composition of S1, the sediment samples of S2 exhibit mean contents of 47.7% coral, 36.2% of coralline algae, and 8.4% of gastropod shells. S3 is composed, on average, of 43.6% coral, 38.8% coralline algae, and 8.7% gastropod shells. Other components such as echinoderms, bryozoans, serpulid tubes, sponge needles, and ostracods contribute <4% to sediment.
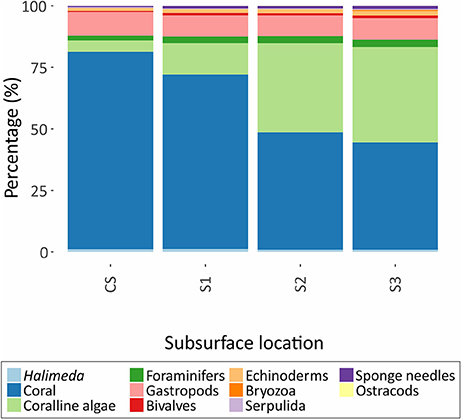
Figure 6. Bar chart indicating the mean percentage of subsurface sediment composition from the different sampling localities on P. Panambungan.
Discussion
Grain Size Distribution of Surface Sediments
Grain size analysis of surface sediment samples reveals homogeneity across the investigated reef islands in the Spermonde Archipelago. General trends in grain size distributions from the individual islands are difficult to identify, however a distinction between the proximal (mid-shelf) island P. Panambungan and the distal (outer shelf) islands, P. Panamgangang, P. Suranti, P. Tambakulu is possible. While the distal islands show sandy gravel as predominant textural group, the proximal island is covered by slightly finer gravelly sand (Table 1). The majority of grains on the distal islands is above 700 μm in size. In contrast, P. Panambungan shows a mean D90 value of 434 μm. These differences in mean grain size of surface sediments may be explained by a higher wave energy during the NW monsoon period from December to January (Mayer and Damm, 2012), and a stronger hydrodynamic setting influenced by the Indonesian Through-Flow at the Spermonde shelf rim (Renema and Troelstra, 2001).
Component Assemblages of Surface Sediments
A zonation of the Spermonde reef island in inner and outer shelf sites is also supported by the component analysis of surface sediments. The analyzed reef islands in the Spermonde clearly group into one inner (P. Panambungan) and three outer (P. Panamgangang, P. Suranti, P. Tambakulu) islands with respect to their surface sedimentary composition (Figures 8, 9).
The hierarchical clustering results indicate two major groups. Cluster 1 is solely composed of samples from P. Panambungan and Cluster 2 contains samples from P. Panamgangang, P. Suranti, P. Tambakulu, except six from P. Panambungan (Figures 7, 8). The setting of two distinct clustering centers in the k-means cluster package of R results in a similar expression, illustrating that the two groups describe either P. Panambungan or the outer Spermonde reef islands P. Panamgangang, P. Suranti, P. Tambakulu (Figure 8). Based on the statistical analysis the reef islands differ in their surface sediment composition, mainly in the contribution of coral and coralline algae (Figure 9). The outer rim reef islands are coral-dominated based on the surface sediment composition. Additionally, the outer islands exhibit a relatively higher abundance of Halimeda, foraminifers and gastropods. In contrast, the sediment composition of the inner island (P. Panambungan) is coralline algae-dominated (Figure 9).
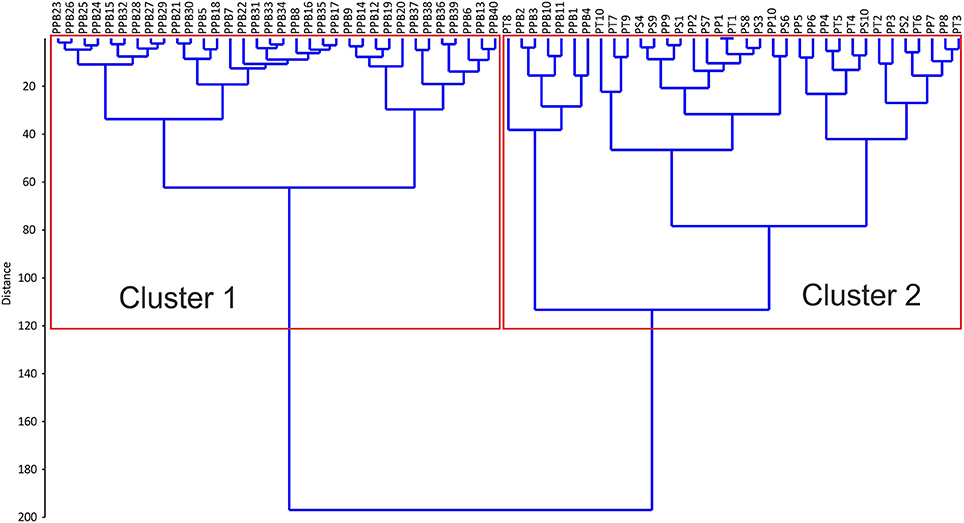
Figure 7. Hierarchical cluster results of surface sediment samples, based on their skeletal constituent composition (Ward's method Euclidean distance).
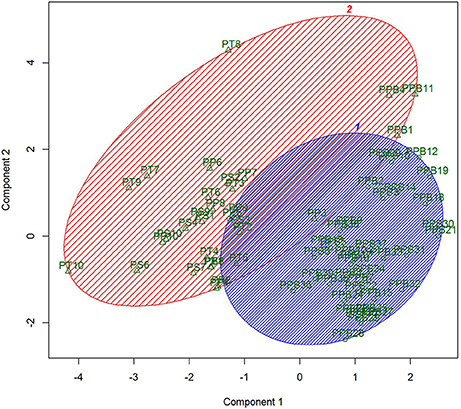
Figure 8. Spermonde reef island sediment samples, k-means cluster results suggesting two clusters. One cluster mainly composed of the samples from P. Panambungan (PPB) and a second cluster mainly composed of the samples from P. Panamgangang (PP), P. Suranti (PS) and P. Tambakulu (PT). The two components explain 62.85% of the point variability.
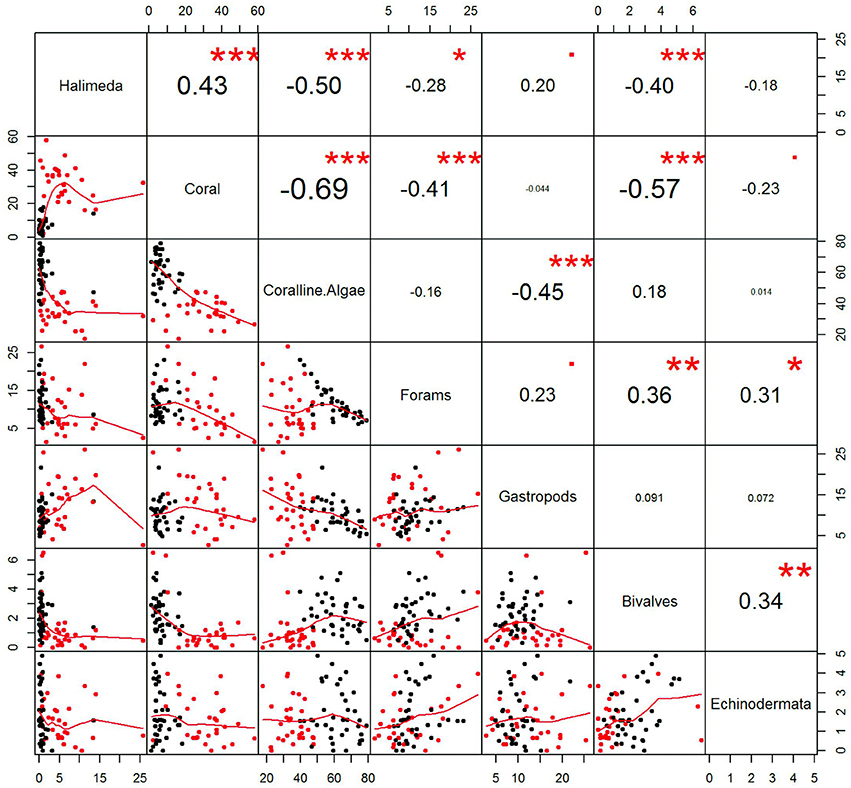
Figure 9. Statistical comparison of the main surface sediment components from the four study islands P. Panamgangang, P. Tambakulu, and P. Suranti and P. Panambungan. Dots in red indicate cluster 1, while dots in black indicate cluster 2. Based on previous statistical analysis (k-means). On the right side, corresponding correlation coefficients is shown, with the highest correlation between coral and coralline algae fragments followed by corals to bivalves.
Grain Size Distribution and Component Assemblages of Subsurface Sediments on P. Panambungan
The D90 grain size distribution of the subsurface sediments from P. Panambungan alternates roughly between 400 and 700 μm (Table 2). Multiple horizontal layers of variable grain size indicate that sediments became deposited on top of the island as a result of overwash deposition (Woodroffe et al., 1999; Barry et al., 2008). This mode of sedimentation is known from other reef island settings such as the Maldives (Kench et al., 2006).
An outstanding feature on P. Panambungan is the observed shift in the sections from a coral-dominated (base) to a coralline algae-dominated (top) facies. Therefore, and from a sedimentological point of view, the reef community at P. Panambungan likely is in a transitional state.
Facies Patterns—Geographical Variability or Phase Shifts in Reef Community Composition?
Previous studies report an on-offshore gradient in faunal assemblages across the Spermonde Archipelago (Cleary et al., 2005; Becking et al., 2006; Cleary and Renema, 2007; Hoeksema, 2012; Sawall et al., 2012). This may explain why P. Panambungan differs in its surface sediment composition in comparison to the outer reefs islands. Variations in reef ecology are a product of distance from the coast due to river discharge impacts, shelf depth, and seasonality (Polónia et al., 2015). Thus, several variables influence the reef community composition across the Spermonde Archipelago, including water quality and habitat-limiting factors such as water depth, dissolved organic matter, and nutrient availability (Polónia et al., 2015). The latter factors here are important as they change with distance to riverine input and result in a zonation of the Spermonde Archipelago (Figure 10). Riverine influence on the Spermonde shelf is most prominent nearshore and during the rainy monsoon (Polónia et al., 2015). Thus, this affects the natural reef community differently across the shelf (Plass-Johnson et al., 2016).
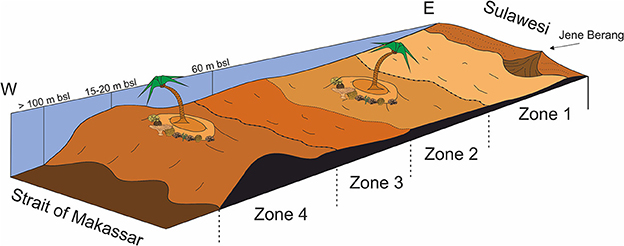
Figure 10. Schematic profile through the Spermonde Shelf following the zonation suggested by De Klerk (1982) and Troelstra et al. (1996). Nearshore factors controlling the spatial zonation are anthropogenic activities and river discharge. With increasing distance to the mainland, factors such as water depth, salinity, nutrient availability, and ocean currents become more important.
However, the coral-dominated subsurface composition of P. Panambungan opposes the hypothesis that differences in surface sediment composition purely result from the islands distance to the mainland. The change in sediment composition from subsurface to surface (i.e., from coral to algal) clearly indicates a shift of the reef community. Shifts in coral reef assemblage have been reported from other coral reefs, e.g., the Caribbean or in the Commonwealth of the Northern Mariana Islands (Alvarez-Filip et al., 2013; Inoue et al., 2013; Enochs et al., 2015). Such shifts in reef community may result in lower calcification rates of corals and a loss of coral framework rugosity (Alvarez-Filip et al., 2013). Following these evidences from different study sites all over the globe, a shift in reef community composition around P. Panambungan is conceivable.
Shifts in reef community from hard reef-building corals to soft non-reef building corals or macroalgal communities often occur due to eutrophication and higher sediment load, which increase seawater turbidity (Gardner, 2003; Alvarez-Filip et al., 2013; Inoue et al., 2013; Enochs et al., 2015; Plass-Johnson et al., 2016). Here anthropogenic influences likely derive from highly populated neighboring islands that act as direct stressors on the reefs, from overfishing, untreated sewage disposal resulting in algae overgrowth, and nutrient flushing (Lessios, 1988; Hughes and Connell, 1999; Bruno et al., 2003; Voss and Richardson, 2006; Perry et al., 2008, 2011; Yoshioka et al., 2016; Ziegler et al., 2016). Perry (1996) directly correlates changes in the sediment composition to a shift in reef community composition. The observed change on P. Panambungan from subsurface coral-dominated to surface coralline algae-dominated sediments thus likely is the result of a shift in reef community structure.
Nevertheless, further sedimentological studies are necessary in order to validate in how far the apparent change of main contributors to island sediments is related to a change in the surrounding reef community structure. Future studies could ideally focus on analyzing depth-profiles of the sedimentary composition and grain size distribution on outer shelf reef islands within the Spermonde. This will help to delimit if the islands further away from the mainland have experienced comparable shifts in sediment composition through time as it is observed on P. Panambungan. Furthermore, determining the exact timeframe of such reef community phase-shifts across islands is highly desirable. This may provide evidence if phase shifts have a local cause (i.e., anthropogenic activities) or have been triggered by major environmental events in the Spermonde Archipelago.
Conclusion
Here we report the results from the first detailed sedimentological study in the Spermonde Archipelago. Findings indicate that the geographic variability (i.e., proximal vs. distal) of the study islands within the Spermonde is reflected in both, the surface grain size distributions, which allow for cautious interpretations of the related hydrodynamic setting, and the surface sediment composition. Surface sediments of the mid-shelf island P. Panambungan are coralline algae-dominated, while the outer shelf islands P. Panamgangang, P. Suranti, P. Tambakulu represent a different reef faunal assemblage with higher abundances of Halimeda, foraminifers, gastropods and, in general, a coral-dominated facies.
Subsurface component assemblages on P. Panambungan are coral-dominated, similar to the surface facies of the outer shelf islands P. Panamgangang, P. Suranti, P. Tambakulu. The shift from coral-dominated sediments to a coralline-algae dominated facies, especially in the central sampling location (CS) on the inner shelf island P. Panambungan is of particular interest. A shift in sediment facies through time (i.e., from lower stratigraphic layers to upper stratigraphic layers) may well be related to a phase shift in the surrounding reef community composition that could also affect reef island stability in the medium-term (Perry et al., 2011).
Reef ecosystems are highly sensitive to alterations of their boundary conditions (such as nutrient levels, salinity, and turbidity). Abrupt changes in those may cause an ecological shift in the reef systems. As the spatial diversity of sediments at P. Panambungan likely is a response to altered environmental boundary conditions caused by anthropogenic activities, this is of major concern for reef management in the Spermonde. Beyond, Indonesia encompasses thousands of populated reef islands that morphologically rely on reef-derived carbonate sediments (Burke et al., 2012). Knowledge and awareness on the controlling factors of reef island alteration patterns thus are of high importance to sustain their future habitability.
Author Contributions
HW and TM designed the research project with substantial contributions by AJ and AW. TM and AK conducted fieldwork with substantial logistical support by DYS. AJ performed research and data analysis, prepared figures and wrote the first draft of the manuscript. AJ, AW, AK, HW and TM discussed and revisited the initial draft. All authors contributed to, and approved the final version of the manuscript to be published.
Funding
This study was funded by the Leibniz Centre for Tropical Marine Research (ZMT) in Bremen, Germany with support by the VolkswagenStiftung, Germany and its funding line “Key issues for Research and Society” as well as the Institutional Strategy of the University of Bremen, funded by the German Excellence Initiative.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study is based on the Master thesis of Janßen (2015). Sebastian Flotow is thanked for support in preparing the sediment samples. We gratefully acknowledge the support of Jamaluddin Jompa and Muhammad Lukman (MaCSI - UNHAS) before and during the fieldwork. Many thanks also to the Indonesian Ministry for Research and Technology (RISTEK). Fieldwork for this study was conducted under the Research Permit No. 399/SIP/ FRP/SM/X/2012 and Working Permit No. 2C11JD5741-L.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/article/10.3389/fmars.2017.00144/full#supplementary-material
References
Alvarez-Filip, L., Carricart-Ganivet, J. P., Horta-Puga, G., and Iglesias-Prieto, R. (2013). Shifts in coral-assemblage composition do not ensure persistence of reef functionality. Sci. Rep. 3:3486. doi: 10.1038/srep03486
Barry, S. J., Cowell, P. J., and Woodroffe, C. D. (2008). Growth-limiting size of atoll-islets: morphodynamics in nature. Mar. Geol. 247, 159–177. doi: 10.1016/j.margeo.2007.09.004
Becking, L. E., Cleary, D. F. R., Voogd, N. J., Renema, W., Beer, M., Soest, R. W. M., et al. (2006). Beta diversity of tropical marine benthic assemblages in the Spermonde Archipelago, Indonesia. Mar. Ecol. 27, 76–88. doi: 10.1111/j.1439-0485.2005.00051.x
Bruno, J. F., Petes, L. E., Drew Harvell, C., and Hettinger, A. (2003). Nutrient enrichment can increase the severity of coral diseases. Ecol. Lett. 6, 1056–1061. doi: 10.1046/j.1461-0248.2003.00544.x
Burke, L., Reytar, K., Spalding, M., Perry, A., Knight, M., Kushner, B., et al. (2012). Reefs at Risk: Revisited in the Coral Triangle. Washington, DC: World Resources Institute.
Cleary, D. F. R., and Renema, W. (2007). Relating species traits of Foraminifera to environmental variables in the Spermonde Archipelago, Indonesia. Mar. Ecol. Prog. Ser. 334, 73–82. doi: 10.3354/meps334073
Cleary, D. F. R., Becking, L. E., de Voogd, N. J., Renema, W., de Beer, M., van Soest, R. W. M., et al. (2005). Variation in the diversity and composition of benthic taxa as a function of distance offshore, depth and exposure in the Spermonde Archipelago, Indonesia. Estuar. Coast. Shelf Sci. 65, 557–570. doi: 10.1016/j.ecss.2005.06.025
De Klerk, L. G. (1982). Zeespiegel Riffen en Kustflakten in Zuitwest Sulawesi, Indonesia. Utrecht: Geography Studies.
Ding, X., Bassinot, F., Guichard, F., and Fang, N. Q. (2013). Indonesian Throughflow and monsoon activity records in the Timor Sea since the last glacial maximum. Mar. Micropaleontol. 101, 115–126. doi: 10.1016/j.marmicro.2013.02.003
Enochs, I. C., Manzello, D. P., Donham, E. M., Kolodziej, G., Okano, R., Johnston, L., et al. (2015). Shift from coral to macroalgae dominance on a volcanically acidified reef. Nat. Clim. Chang. 5, 1–9. doi: 10.1038/nclimate2758
Ford, M. R., and Kench, P. S. (2012). The durability of bioclastic sediments and implications for coral reef deposit formation. Sedimentology 59, 830–842. doi: 10.1111/j.1365-3091.2011.01281.x
Fournier, J., Gallon, R. K., and Paris, R. (2014). G2Sd: a new R package for the statistical analysis of unconsolidated sediments. Géomorphol. Reli. Process. Environ. 14, 73–78. doi: 10.4000/geomorphologie.10513
Gardner, T. A. (2003). Long-term region-wide declines in caribbean corals. Science 301, 958–960. doi: 10.1126/science.1086050
Hoeksema, B. W. (1990). Systematics and Ecology of Mushroom Corals (Scleractinia: Fungiidae). PhD Thesis, Leiden University, Leiden.
Hoeksema, B. W. (2012). Distribution patterns of mushroom corals (Scleractinia : Fungiidae) across the Spermonde Shelf, South Sulawesi. Raffles Bull. Zool. 60, 183–212.
Hughes, T. P., and Connell, J. H. (1999). Multiple stressors on coral reefs: a long -term perspective. Limnol. Oceanogr. 44, 932–940. doi: 10.4319/lo.1999.44.3_part_2.0932
Inoue, S., Kayanne, H., Yamamoto, S., and Kurihara, H. (2013). Spatial community shift from hard to soft corals in acidified water. Nat. Clim. Chang. 3, 1–5. doi: 10.1038/nclimate1855
Janßen, A. (2015). Sedimentary Facies of an Uninhabited Reef Island in the Spermonde Archipelago. Master's thesis, SW Sulawesi, Indonesia: University of Bremen.
Kench, P. S., Brander, R. W., Parnell, K. E., and McLean, R. F. (2006). Wave energy gradients across a Maldivian atoll: implications for island geomorphology. Geomorphology 81, 1–17. doi: 10.1016/j.geomorph.2006.03.003
Kench, P. S., Smithers, S. G., and McLean, R. F. (2012). Rapid reef island formation and stability over an emerging reef flat: Bewick Cay, northern Great Barrier Reef, Australia. Geology 40, 347–350. doi: 10.1130/G32816.1
Lessios, H. A. (1988). Mass mortality of Diadema antillarum in the Caribbean: what have we learned? Annu. Rev. Ecol. Syst. 19, 371–393. doi: 10.1146/annurev.es.19.110188.002103
Maechler, M., Rousseeuw, P., Struyf, A., Hubert, M., and Hornik, K. (2016). Cluster: Cluster Analysis Basics and Extensions. CRAN, R package cluster.
Manjunatha, B. R., Krishna, K. M., and Aswini, A. (2015). Anomalies of the sea surface temperature in the indonesian throughflow regions: a need for further investigation. Open Oceanogr. J. 8, 2–8. doi: 10.2174/1874252101408010002
Mann, T., Rovere, A., Schöne, T., Klicpera, A., Stocchi, P., Lukman, M., et al. (2016). The magnitude of a mid-Holocene sea-level highstand in the Strait of Makassar. Geomorphology 257, 155–163. doi: 10.1016/j.geomorph.2015.12.023
Mayer, B., and Damm, P. E. (2012). The Makassar Strait throughflow and its jet. J. Geophys. Res. 117, C07020. doi: 10.1029/2011JC007809
Meteo365.com Ltd. (2015). Tide Times for Makassar. Available online at: http://www.tide-forecast.com/locations/Ujung-Pandang-Sulawesi-Indonesia/tides/latest
Moore, M. D. (1995). Proxy Records of the Indonesian Low and the El Ni{tilde n}o-Southern Oscillation (ENSO) from Stable Isotope Measurements of Indonesian Reef Corals. Ph. D. Thesis, University of California, Berkeley, CA.
Morgan, K. M., and Kench, P. S. (2016). Reef to island sediment connections on a Maldivian carbonate platform: using benthic ecology and biosedimentary depositional facies to examine island-building potential. Earth Surf. Process. Landforms 41, 1815–1825. doi: 10.1002/esp.3946
Perry, C. T. (1996). The rapid response of reef sediments to changes in community composition; implications for time averaging and sediment accumulation. J. Sediment. Res. 66, 459. doi: 10.1306/D4268374-2B26-11D7-8648000102C1865D
Perry, C. T., Kench, P. S., Smithers, S. G., Riegl, B., Yamano, H., and O'Leary, M. J. (2011). Implications of reef ecosystem change for the stability and maintenance of coral reef islands. Glob. Chang. Biol. 17, 3679–3696. doi: 10.1111/j.1365-2486.2011.02523.x
Perry, C. T., Spencer, T., and Kench, P. S. (2008). Carbonate budgets and reef production states: a geomorphic perspective on the ecological phase-shift concept. Coral Reefs 27, 853–866. doi: 10.1007/s00338-008-0418-z
Plass-Johnson, J. G., Heiden, J. P., Abu, N., Lukman, M., and Teichberg, M. (2016). Experimental analysis of the effects of consumer exclusion on recruitment and succession of a coral reef system along a water quality gradient in the Spermonde Archipelago, Indonesia. Coral Reefs 35, 229–243. doi: 10.1007/s00338-015-1369-9
Polónia, A. R. M., Cleary, D. F. R., de Voogd, N. J., Renema, W., Hoeksema, B. W., Martins, A., et al. (2015). Habitat and water quality variables as predictors of community composition in an Indonesian coral reef: a multi-taxon study in the Spermonde Archipelago. Sci. Total Environ. 537, 139–151. doi: 10.1016/j.scitotenv.2015.07.102
Ray, R., Egbert, G., and Erofeeva, S. (2005). A brief overview of tides in the Indonesian Seas. Oceanography 18, 74–79. doi: 10.5670/oceanog.2005.07
Renema, W., and Troelstra, S. R. (2001). Larger foraminifera distribution on a mesotrophic carbonate shelf in SW Sulawesi (Indonesia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 175, 125–146. doi: 10.1016/S0031-0182(01)00389-3
Renema, W., Hoeksema, B. W., and Van Hinte, J. E. (2001). Larger benthic foraminifera and their distribution patterns on the Spermonde shelf, South Sulawesi. Zool. Verh. 334, 115–150.
Sawall, Y., Richter, C., Ramette, A., Wieczorek, S., Todd, C., Costerton, J., et al. (2012). Effects of eutrophication, seasonality and macrofouling on the diversity of bacterial biofilms in equatorial coral reefs. PLoS ONE 7:e39951. doi: 10.1371/journal.pone.0039951
Stoddart, D. R., and Steers, A. (1977). “The nature and orgin of coral reef islands,” in Biology and Geology of Coral Reefs, Vol. 4, eds A. O. Jones and R. Endean (New York, NY: Academic Press), 59–105.
Storm, C. (1989). Influences of River Systems and Coastal Hydrodynamics on the Water Quality during One Monsoonal Period: A Hydrological Survey of a Tropical Coastal Area in the Spermonde Archipelago. SW Sulawesi, Indonesia, rapport Geopro. Univ. Utr.
Troelstra, S. R., Jonkers, H. M., and De Rijk, S. (1996). Larger foraminifera from the spermonde archipelago (Sulawesi, Indonesia). Scr. Geol. 113, 93–120.
Voss, J. D., and Richardson, L. L. (2006). Nutrient enrichment enhances black band disease progression in corals. Coral Reefs 25, 569–576. doi: 10.1007/s00338-006-0131-8
Wentworth, C. K. (1922). A scale of grade and class terms for clastic sediments. J. Geol. 30, 377–392. doi: 10.1086/622910
Wijsman-Best, M., Moll, H., and De Klerk, L. G. (1982). “Present status of the coral reefs in the Spermonde Archipelago (South Sulawesi, Indonesia),” in Proc. 4th Int. Coral Reef Symp. Mar. Sci. Center, Univ. Philipp. Vol. 1, 263–267. Manila. Available online at: http://www.sciencedirect.com/science/article/pii/S0012825204000959 (Accessed February 26, 2014).
Wilson, M. E. J., and Vecsei, A. (2005). The apparent paradox of abundant foramol facies in low latitudes: their environmental significance and effect on platform development. Earth Sci. Rev. 69, 133–168. doi: 10.1016/j.earscirev.2004.08.003
Wizemann, A., Mann, T., Klicpera, A., and Westphal, H. (2015). Microstructural analyses of sedimentary Halimeda segments from the Spermonde Archipelago (SW Sulawesi, Indonesia): a new indicator for sediment transport in tropical reef islands? Facies 61:4. doi: 10.1007/s10347-015-0429-5
Woodroffe, C. D., McLean, R. F., Smithers, S. G., and Lawson, E. M. (1999). Atoll reef-island formation and response to sea-level change: West Island, Cocos (Keeling) Islands. Mar. Geol. 160, 85–104. doi: 10.1016/S0025-3227(99)00009-2
Woodruff, S. D., Diaz, H. F., Elms, J. D., and Worley, S. J. (1998). COADS Release 2 data and metadata enhancements for improvements of marine surface flux fields. Phys. Chem. Earth 23, 517–526. doi: 10.1016/S0079-1946(98)00064-0
Yamano, H., Kayanne, H., and Chikamori, M. (2005). An overview of the nature and dynamics of reef islands. Glob. Environ. Res. 9, 9–20.
Yamano, H., Miyajima, T., and Koike, I. (2000). Importance of foraminifera for the formation and maintenance of a coral sand cay: Green Island, Australia. Coral Reefs 19, 51–58. doi: 10.1007/s003380050226
Yoshioka, R. M., Kim, C. J. S., Tracy, A. M., Most, R., and Harvell, C. D. (2016). Linking sewage pollution and water quality to spatial patterns of Porites lobata growth anomalies in Puako, Hawaii. Mar. Pollut. Bull. 104, 313–321. doi: 10.1016/j.marpolbul.2016.01.002
Keywords: carbonate sediments, grain size, skeletal constituents, facies zonation, reef island evolution
Citation: Janßen A, Wizemann A, Klicpera A, Satari DY, Westphal H and Mann T (2017) Sediment Composition and Facies of Coral Reef Islands in the Spermonde Archipelago, Indonesia. Front. Mar. Sci. 4:144. doi: 10.3389/fmars.2017.00144
Received: 22 November 2016; Accepted: 28 April 2017;
Published: 17 May 2017.
Edited by:
Marc Kochzius, Vrije Universiteit Brussel, BelgiumReviewed by:
Katherine Dafforn, University of New South Wales, AustraliaRafael Sarda, Consejo Superior de Investigaciones Científicas, Spain
Copyright © 2017 Janßen, Wizemann, Klicpera, Satari, Westphal and Mann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander Janßen, ajanssen@marum.de