 Jonathan Niño-Sánchez†
Jonathan Niño-Sánchez† Vega Tello†
Vega Tello† Virginia Casado-del Castillo
Virginia Casado-del Castillo Michael R. Thon
Michael R. Thon Ernesto P. Benito
Ernesto P. Benito José María Díaz-Mínguez*
José María Díaz-Mínguez*
- Departamento de Microbiología y Genética, Instituto Hispano-Luso de Investigaciones Agrarias, Universidad de Salamanca, Salamanca, Spain
The dynamics of root and hypocotyl colonization, and the gene expression patterns of several fungal virulence factors and plant defense factors have been analyzed and compared in the interaction of two Fusarium oxysporum f. sp. phaseoli strains displaying clear differences in virulence, with a susceptible common bean cultivar. The growth of the two strains on the root surface and the colonization of the root was quantitatively similar although the highly virulent (HV) strain was more efficient reaching the central root cylinder. The main differences between both strains were found in the temporal and spatial dynamics of crown root and hypocotyl colonization. The increase of fungal biomass in the crown root was considerably larger for the HV strain, which, after an initial stage of global colonization of both the vascular cylinder and the parenchymal cells, restricted its growth to the newly differentiated xylem vessels. The weakly virulent (WV) strain was a much slower and less efficient colonizer of the xylem vessels, showing also growth in the intercellular spaces of the parenchyma. Most of the virulence genes analyzed showed similar expression patterns in both strains, except SIX1, SIX6 and the gene encoding the transcription factor FTF1, which were highly upregulated in root crown and hypocotyl. The response induced in the infected plant showed interesting differences for both strains. The WV strain induced an early and strong transcription of the PR1 gene, involved in SAR response, while the HV strain preferentially induced the early expression of the ethylene responsive factor ERF2.
Introduction
Fusarium oxysporum Schlechtend.:Fr. is an anamorphic species complex (FOSC) with considerable morphological and physiological variation (O’Donnell et al., 2009). This fungus is ubiquitous in soils worldwide being able to grow saprophytically or colonizing plants. The pathogenic strains collectively may infect more than 100 different hosts, ranging from globally important crops such as cotton, tomato, banana, and several legumes to immunocompromised patients (Nucci and Anaissie, 2007; Michielse and Rep, 2009). Despite the broad host range at the species complex level, the individual isolates are able to infect only one or a few plant species. This allows for a classification into formae speciales based on host-specificity (Booth, 1971; Armstrong and Armstrong, 1975), making the F. oxysporum species complex a very attractive system for the study of the relationships between pathogenicity, virulence and host-specificity. F. oxysporum f. sp. phaseoli infects some species of the genus Phaseolus, mainly Phaseolus vulgaris and P. coccineus (de Vega-Bartol et al., 2011), and is one of the principal agents limiting dry bean production worldwide.
The cellular basis of the host-pathogen interaction in vascular wilt diseases was first described using transmission electron-microscopy (Bishop and Cooper, 1983). The use of laser scanning spectral confocal microscopy (CLSM) has improved our knowledge of the details concerning infection in multiple hosts and different races of the pathogen. The majority of studies show that invasion of the host by F. oxysporum strains expressing the green fluorescent protein (GFP) begins with the development of a hyphal network around root hairs and the upper part of taproot, followed by penetration and colonization of the root epidermis. Once the hyphae are inside the roots the development of disease symptoms and degree of colonization in resistant and susceptible cultivars show differences in several hosts. F. oxysporum f. sp. medicaginis colonizes the cortex and central cylinder of roots of resistant and susceptible cultivars in similar ways. In Medicago truncatula inoculation of a resistant cultivar resulted in the delay of symptom development and reduction of the disease severity with respect to the inoculation of a susceptible cultivar, but no correlation could be found with the degree of colonization of the root parenchyma and central cylinder (Ramírez-Suero et al., 2010). In watermelon, the pathogen can colonize the lateral roots in compatible interactions, but it is unable to penetrate the taproot when infecting a resistant cultivar (Lü et al., 2013). A more complex interaction has been recently reported in the infection of chickpea with F. oxysporum f. sp. ciceris. In compatible interactions the pathogen grows intercellularly in the root cortex, reaches the xylem and then advances upward through the xylem of the stem (Jiménez-Fernández et al., 2013). All the incompatible interactions were asymptomatic, but a race 0 strain was able to colonize the xylem vessels of root and stem of the resistant cultivar JG-62, while in another resistant cultivar, WR-315, it was halted in the intercellular spaces of the root cortex failing to reach the xylem. However, a race 5 strain progressed up to the hypocotyl in the same resistant cultivar WR-315 (Jiménez-Fernández et al., 2013). Therefore, colonization of xylem vessels does not guarantee a compatible interaction, at least in chickpea.
All the examples described above are based on the analysis of host-specificity (formae speciales) and cultivar specificity (races). Much less attention has been paid to interactions involving more virulent or less aggressive strains, even though virulence is often the only difference between a devastating and a tolerable disease. Previously, we characterized several groups of virulence in F. oxysporum f. sp. phaseoli and demonstrated that virulence is not correlated with race, because strains that belong to the same race may display different virulence on the same common bean cultivar (Alves-Santos et al., 2002a). However, virulence depends in part on the host, because equally virulent strains on P. vulgaris show differences in virulence on P. coccineus (de Vega-Bartol et al., 2011). We have also shown that some virulence factors are specific to highly virulent (HV) strains (Alves-Santos et al., 2002b) and specifically induced in planta (Ramos et al., 2007). In the only confocal microscopy study reported to date involving strains of F. oxysporum differing in virulence, we obtained evidence about the importance of the speed of xylem vessel colonization as a determinant of disease severity (García-Sánchez et al., 2010). This analysis left open some important questions related to the differences in colonization between HV and weakly virulent (WV) strains. For example, do HV strains develop a higher amount of fungal biomass inside the host than WV strains? What are the spatial milestones of colonization that differentiate both types of strains? Finally, if there is a differential expression of fungal virulence factors and plant defense factors, when does it happen?
In this work we addressed the above questions by determining the qualitative and quantitative differences on the spatial and temporal dynamics of colonization of a susceptible common bean cultivar infected by WV and HV strains of F. oxysporum f. sp. phaseoli, the gene expression patterns of several fungal virulence factors, and the host defense response involving expression of genes related to the salicylic acid (SA) defense induced response and the ethylene (ET)/jasmonic mediated response. Systemic acquired resistance (SAR) confers protection against a broad spectrum of microorganisms. SAR requires the signal molecule SA and is associated with accumulation of pathogenesis-related proteins (PRs; Durrant and Dong, 2004). Also, the cross-talk between ET and jasmonate (JA) signaling pathways determines the activation of defense responses against pathogens.
Materials and Methods
Fungal Strains and Culture Conditions
The F. oxysporum f. sp. phaseoli strains FOP-SP1 (HV) and FOP-SP4 (WV) used in this study have been described (Alves-Santos et al., 1999, 2002a). All strains were grown as previously described (Alves-Santos et al., 1999). Fungal cultures were established from frozen mycelia stored on 25% glycerol (v/v) at -80°C, and incubated at 25°C with continuous light for 1 week (solid media) or 5 days at 120–180 rpm (liquid cultures).
Development of GFP-Expressing Strains of F. oxysporum
Plasmid pRF-HU-GFP was constructed to express GFP in F. oxysporum f. sp. phaseoli. A 3.1 Kb fragment containing the sGFP gene under the control of the glyceraldehyde 3-phosphate deshydrogenase promoter of Aspergillus nidulans was excised from plasmid gGFP (Maor et al., 1998; Sukno et al., 2008) by digestion with XbaI and ClaI. This 3.1 Kb XbaI-ClaI fragment was blunt-ended using the Klenow fragment of T4 DNA polymerase and ligated into the binary vector pRF-HU (Frandsen et al., 2008) digested with PacI and also blunt-ended. Plasmid pRF-HU-PFTF1GFP was designed to express GFP under the control of the FTF1 promoter (PFTF1). The pair of primers B310User-5FTF1RBamHI (Table 1) was used to amplify a 670 bp DNA fragment containing the promoter of gene FTF1 using genomic DNA of strain FOP-SP1 as template. In parallel, primers 5GFPFBamHI and GFPUserR were used to amplify the coding region of sGFP using as template the plasmid gGFP. Both PCR amplification products were digested with BamHI (as recognition sequences were included in one of the primers of each pair) and then ligated using T4 DNA ligase. 100 μl of the ligation reaction were used as template to amplify the cassette PFTF1::GFP. The primers used to this end (B310User and GFPUserR) included the sequences needed to ligate the PCR product to plasmid pRF-HU digested with PacI and Nt.BbvCI USERTM enzimes (Frandsen et al., 2008). Both plasmids, pRF-HU-GFP and pRF-HU-PFTF1GFP, were used to genetically transform F. oxysporum f.sp. phaseoli by means of the A. tumefaciens-mediated transformation procedure as previously described (Mullins et al., 2001; Ramos et al., 2007).
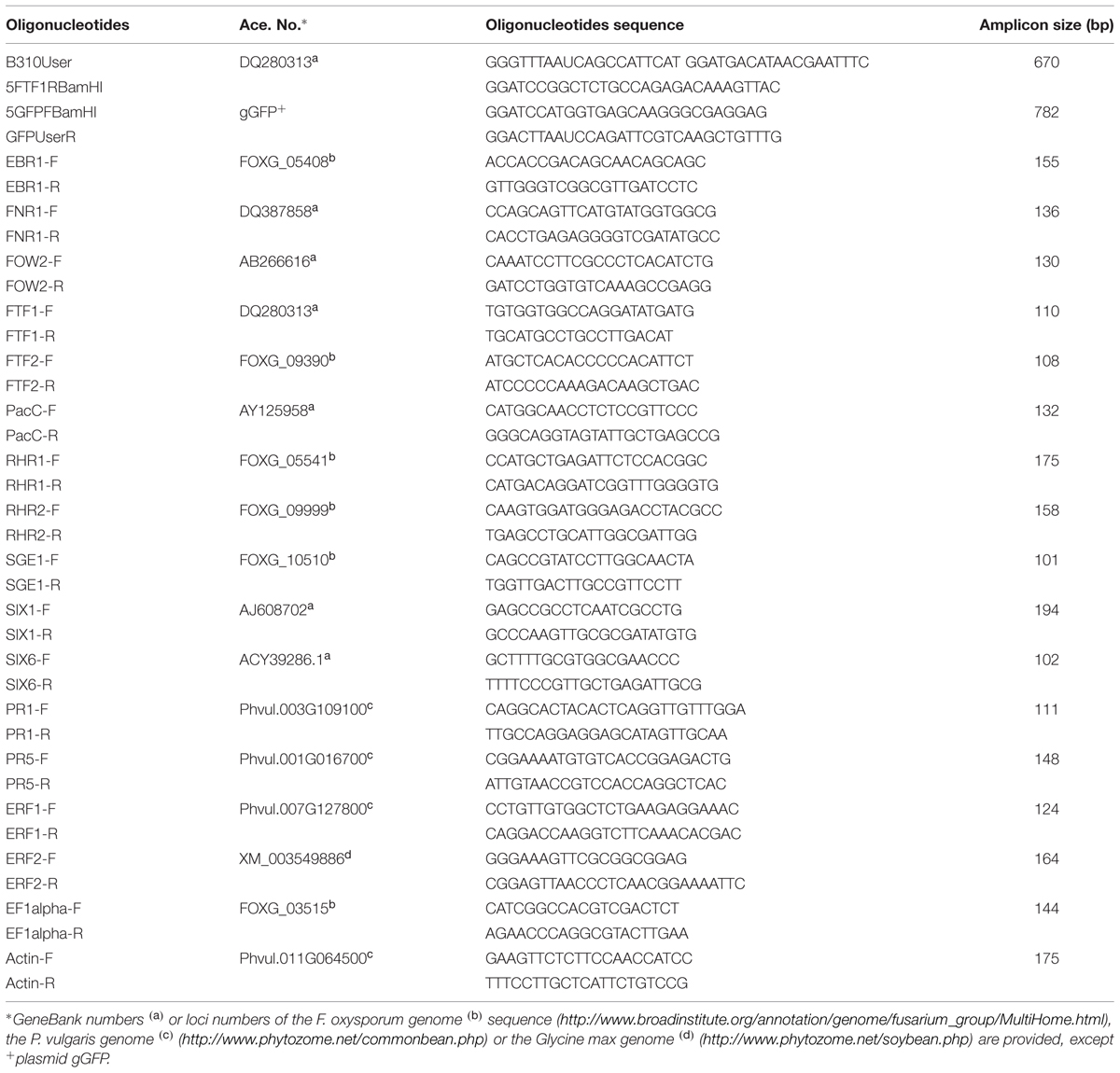
TABLE 1. Oligonucleotides used in this study.
Pathogenicity Tests
Inoculation of P. vulgaris L. cv Blanca Riñón with conidia from the F. oxysporum strains tested was mainly carried out as previously described (Alves-Santos et al., 1999). After inoculation, the plants were either transferred to pots and further incubated in a greenhouse for regular pathogenicity tests, or to 50 ml Falcon tubes filled with PGM (Plant Growth Medium) solution and covered with foil. These hydroponic cultures were incubated in an environment-controlled chamber at 25°C and relative humidity of 60–80% with a 16 h photoperiod fluorescent light. All the strains and transformants were simultaneously tested in each inoculation experiment to reduce variability in the plant response due to environmental conditions. Pathogenicity tests were repeated three times and 12 replica plants were included per treatment.
External symptoms of disease in plants grown in Falcon tubes could not be correlated with those previously recorded using the CIAT scale in plants grown in pots, as growth of the plants under the hydroponic conditions used was slower. However, distinctions could be made between plants inoculated with the HV or the WV virulent strains, both in the number of chlorotic leaves and the degree of necrosis in the vascular system. A new scale was designed according to the degree of external (chlorotic leaves) and internal (necrotic lesions) symptoms observed. 0: healthy plant with no symptoms of chlorosis or necrosis; 1: plant with no chlorotic leaves but light necrotic lesions; 2: plant with some clorotic leaves and necrotic lesions in the crown region and the xylem system; 3: plant with most of the leaves either chlorotic or dead, extensive necrosis of the crown region and necrosis of the vascular system; 4: dead plant.
Microscopic Examination by Means of Confocal Laser Microscopy
The plants inoculated with FOP-SP1 or FOP-SP4 and the controls treated with water were maintained in hydroponic cultures and examined each day for a period of 21 days after infection. For the examination of histological structure plant sections were fixed and embedded in paraffin as described (Jiménez et al., 2007), and then stained with toluidine blue 0.1% in potassium phosphate buffer 0.1 M pH 6.8. For CLSM analysis the plants were taken from the Falcon tubes and the roots washed in sterile distilled water. Plant sections or longitudinal samples 1 mm in thickness were manually sliced with a razor blade from the following tissues: root hairs, tap root zones (apex, intermediate, upper), crown and hypocotyl. Plant tissues were stained with a drop of propidium iodide (SIGMA) at a concentration of 10 mg/ml for 15 min and the excess propidium iodide was removed by washing with sterile distilled water. Then, samples were placed on microscope slides, submerged in glycerol 50% (v/v) and gently squashed with a cover glass. Microscopic analysis was performed using a laser scanning spectral confocal microscope (TCS2-SP2, Leica Microsystems, Germany). Excitation was provided by an argon laser (488 nm). Fluorescence of GFP was detected at 495–520 nm and plant tissue fluorescence (either propidium iodide stained and autofluorescence) at 595–680 nm.
The quantification of xylem vessel colonization was performed with the help of ImageJ. Data were obtained from at least six images per plant section showing growing hyphae of the HV strain or the WV strain. Parenchyma and xylem vessels were compared and GFP cell fluorescence was used to estimate the ratio of fungal colonization of similar areas in both tissues.
Nucleic Acid Manipulations
Genomic DNA was extracted from F. oxysporum mycelium according to the procedures previously described (Alves-Santos et al., 1999, 2002b; Ramos et al., 2007). RNA was extracted from P. vulgaris plants at different times after inoculation with F. oxysporum strains. Roots and hypocotyls were cut, immediately frozen at -80°C and ground in a mortar with pestle under liquid nitrogen. Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. RNA was treated with DNAase Turbo DNA-free (Ambion Inc., Austin, TX, USA), tested for integrity by running samples in agarose gels and quantified using a Nanodrop Spectrophotometer (Thermo Scientific, Waltham, MA, USA). Southern blots were carried out as described previously (Ramos et al., 2007). DNA probes were labeled with digoxigenin-dUTP (Roche Diagnostics) by the PCR method using the Biotools DNA polymerase (Biotools SA, Spain) and plasmid pFR-HU-GFP DNA as template. Prehybridization, hybridization, washings, and detection were performed using a chemiluminescent detection procedure using CDP-Star (Roche Diagnostics) according to the manufacturer’s recommendations.
Real-Time Quantitative Analysis of Gene Expression
Samples of plants inoculated with F. oxysporum were collected and frozen at -80°C. Total RNA was extracted as described before. cDNA was synthesized from 1 μg of total RNA in 20 μl (total volume) reactions using the PrimeScriptTM RT reagent kit (Takara Biosystems), and oligo (dT) as a primer, according to the manufacturer’s procedure.
The primer pairs used in RT-qPCR assays are shown in Table 1. Primer pairs used to analyze the expression of F. oxysporum genes were designed according to alignments performed using all the F. oxysporum genome sequences available at the Broad Institute database. They were verified in PCR experiments using genomic DNA from FOP-S1 and FOP-SP4 as template. All of them were able to amplify DNA fragments of the expected size in both strains, except the fragment corresponding to FTF1, as this gene is specific of HV strains. The amplicons were sequenced to verify their identities. Primer pairs used to analyze the expression of plant genes were designed on the basis of the Glycine max sequences. The amplicons obtained using common bean DNA as template were sequenced to confirm that they were homologues of the G. max genes. Before using a pair of primers in an expression analysis, its PCR amplification efficiency was calculated using six serial dilutions of cDNA as template. All the primer pairs tested showed efficiencies greater than 97%. qPCR was carried out in 10 μl reaction mixtures containing 1X KAPA SYBR Green qPCR Master Mix (Kapa Biosystems), 500 nM forward and reverse primers and 1 μl of the reverse transcription reaction. Amplifications were performed in a StepOnePlusTM Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) using the thermal profile recommended by the manufacturer (40 cycles of 95°C for 3 s and 60°C for 30 s). The presence of only one specific peak was checked in the melting curve (dissociation curve), which was run at 0.5°C increments every 5 s within a range of 55–95°C. The qPCR product size was checked by electrophoresis in 2.5% (w/v) agarose gels that were stained with ethidium bromide and visualized under UV light.
Two or three sets of samples from different infection assays were used (biological replicates). Two independent cDNA preparations per biological replicate were obtained, and three replicas of each cDNA were analyzed. The endogenous reference genes used were the F. oxysporum EF1a encoding a translation elongation factor and the common bean actin gene. The expression of both genes was found to remain constant under all the conditions used in the present study (data not shown). Relative expression levels were calculated using the comparative cycle threshold (Ct) method (Bustin, 2000; Pfaffl et al., 2002), using the formula RQ (relative quantity of transcript) = 2-ΔΔCt, where ΔCt = Ctspecific gene – Ctreference gene of any given sample, and ΔΔCt = ΔCt (as previously calculated) – ΔCt of 1 dpi FOP-SP4 inoculated samples, when specific fungal genes where analyzed, or ΔCt of mock inoculated plants, when plant genes were analyzed. All Ct calculations, integration of the amplification efficiency data and statistical analyses were performed using StepOneTM Software V2.2.2.
Fungal Biomass Quantification
The relative fungal DNA amount was quantified by means of qPCR. Six infected plants were randomly selected and collected for each condition assayed, and DNA obtained as previously described. For the detection and quantification of fungal DNA primers that generate fragments of the EF1α and FTF2 (Fusarium Transcription Factor 2) genes were used (Table 1). Both genes are found as single-copy sequences in the genome of F. oxysporum. A fragment of the actin gene from common bean, generated by using primers Actin-F and Actin-R (Table 1), was used as an endogenous gene to normalize differences in DNA template amounts. The qPCR reaction components and cycling conditions were as above described, except that 1 ng of DNA was used as template.
Fungal biomass in inoculated samples was estimated as the relative quantity of FOP-SP1 or FOP-SP4 DNA normalized to an endogenous control of P. vulgaris DNA. Standard curves for F. oxysporum and P. vulgaris were constructed based on the relationship of Ct values and known host and pathogen DNA concentrations. Genomic DNA isolated from fungal mycelia grown in culture and samples of roots, root crowns, and hypocotyls of non-inoculated plants were used to create the standard curves. Each sample was evaluated in triplicate and the assay was repeated two times for each of the three biological replicates. The analyses were performed using StepOneTM Software V2.2.2.
Results
Cytological Characterization of Colonization by Weakly Virulent and Highly Virulent Strains of F. oxysporum f. sp. phaseoli
Green fluorescent protein transformants of F. oxysporum f. sp. phaseoli were generated by transformation of strains FOP-SP1 and FOP-SP4 with plasmid pRF-HU-GFP. Both strains are wild types isolated in El Barco de Ávila (Spain) that share IGS-A haplotype, mating type (MAT1-2) and race (6). However, FOP-SP1 is a HV strain that belongs to VCG 167, while FOP-SP4 belongs to VCG 166 and behaves as WV when inoculated on common bean plants (Alves-Santos et al., 1999, 2002a). Several GFP expressing transformants were obtained for each strain and examined for mycelial growth, both in solid and liquid culture, sporulation and virulence. No significant differences could be observed in the rates of growth in PDA medium and rates of sporulation in PDB medium between the GFP transformants and their respective wild type strains. Infection assays were also performed to compare GFP expressing strains with the wild type strains by inoculation of cultivar Blanca Riñón, but no significant differences (P > 0.9 according to “Student’s” t-test) could be observed when the mean disease symptoms or the disease progression rate (García-Sánchez et al., 2010) were assessed (data not shown).
To compare the patterns of common bean colonization by WV and HV strains three different regions of inoculated common bean plants were chosen for confocal microscopy based on the different architecture of their vascular system: the tap root, the root crown and the hypocotyl (Figure 1). The initial stages of root colonization were similar in plants inoculated with WV (FOP-SP4) and HV (FOP-SP1) strains. Following root dip inoculation, a dense web of hyphae grew over the surface of the taproot and the lateral roots (Figures 2G–L). Both strains showed a marked preference for the colonization of interstitial regions, especially the junctions of lateral roots with the tap root (Figures 2A–F) which appear to serve as preferred penetration areas. Specialized penetration structures could not be observed for any of the strains.
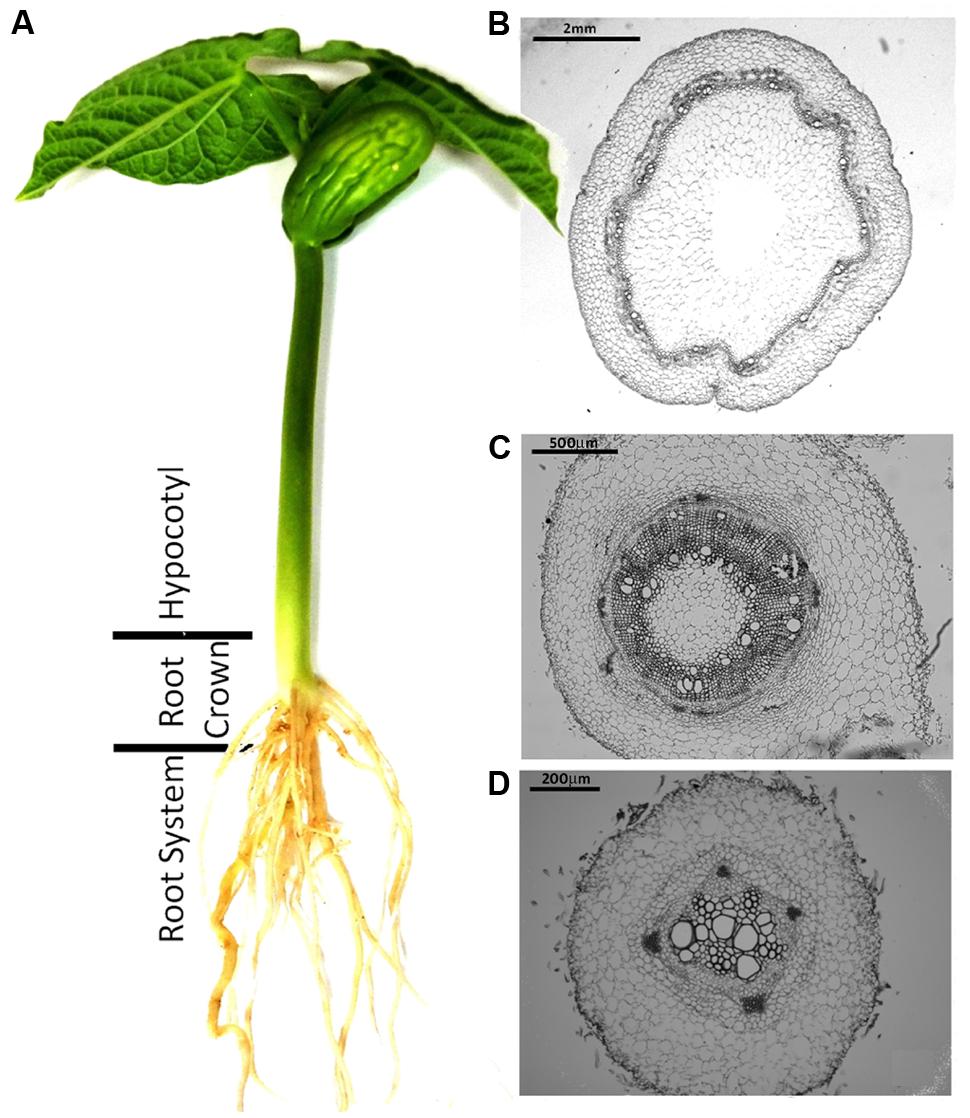
FIGURE 1. Common bean plant (Phaseolus vulgaris L.) and histological structure of the plant regions analyzed in the present study. (A) Common bean plant grown in hydroponic culture. (B–D) Cross sections of the three plant regions analyzed in the present study showing the definitive architecture of the xylem and phloem vessels in the hypocotyl (B), the primordial rings of xylem and phloem vessels in the root crown (C) and the undifferentiated central cylinder in the tap root (D). Plant sections were fixed, embedded in paraffin and stained with toluidine blue for microscopic examination.
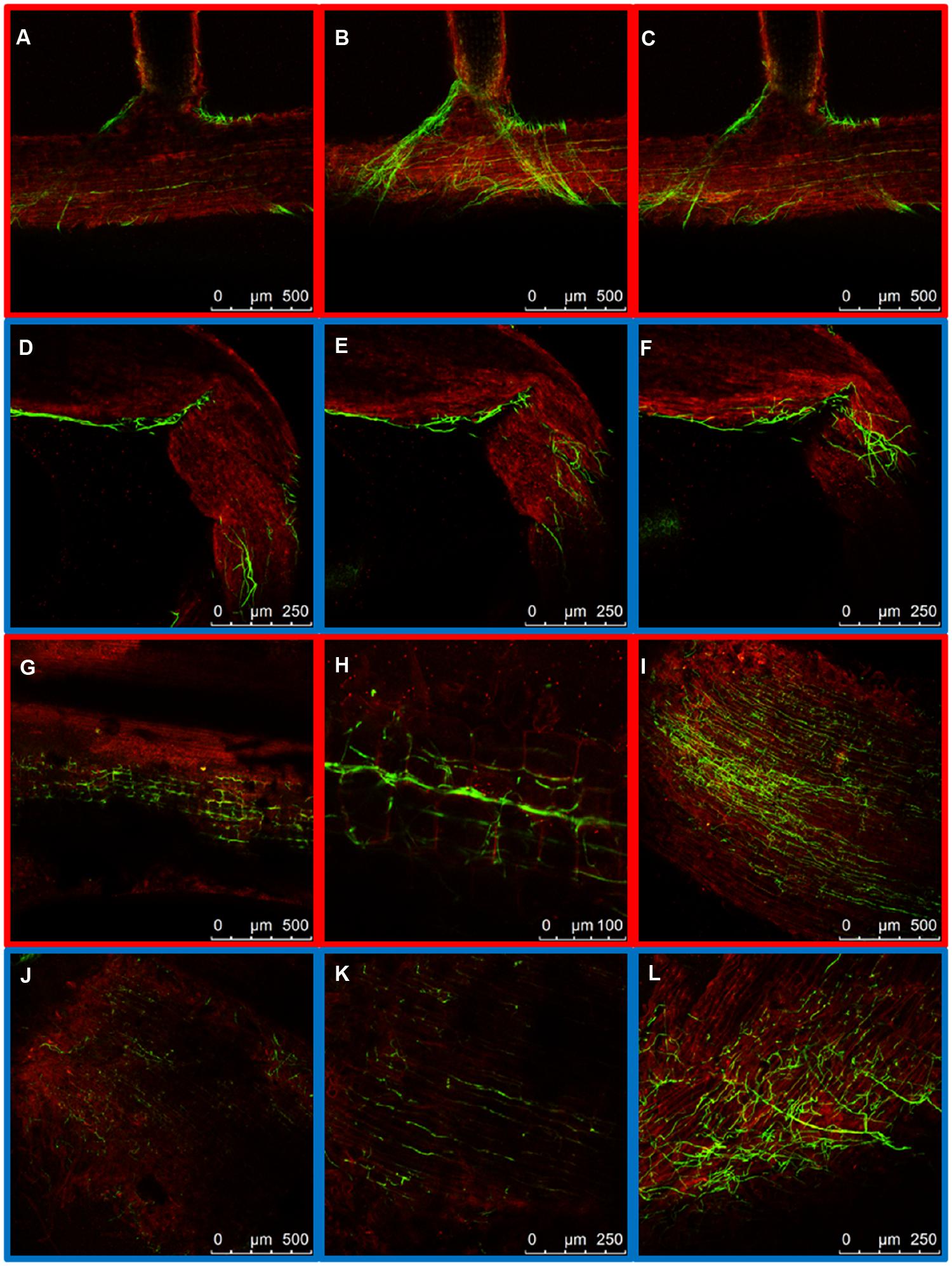
FIGURE 2. External colonization of the root system by highly virulent (HV; FOP-SP1) and weakly virulent (WV; FOP-SP4) Fusarium oxysporum f. sp. phaseoli strains. Colonization of the taproot and lateral roots of plants inoculated with the HV strain FOP-SP1 (red framed images) and the WV strain FOP-SP4 (blue framed images) was visualized by confocal laser scanning microscopy 1–3 days post inoculation (dpi). (A–C) FOP-SP1 mycelium growing in the junction of a lateral root with the taproot. (D–F) FOP-SP4 mycelium growing in an interstitial zone of a lateral root. Intermediate root zones showing epidermis colonization by FOP-SP1 hyphae (G–I) and FOP-SP4 hyphae (J–L).
At 7 days post inoculation (dpi) the taproots were almost completely colonized by both strains. However, a clear distinction in the main distribution of the mycelium could be observed. The WV strain showed extensive intercellular growth in the root cortex, mainly along the longitudinal axis of the root (Figure 3B), although some hyphae could also be seen inside the xylem vessels of the central root cylinder. In contrast, the hyphae of the HV strain quickly reached the central root xylem where they spread along the xylem vessels (Figure 3A). Comparison of cross sections of the taproot clearly demonstrates how the WV strain behaves mainly as a root cortex colonizer (Figures 3E,F) while the HV strains behaves as a fast vascular colonizer (Figures 3C,D).
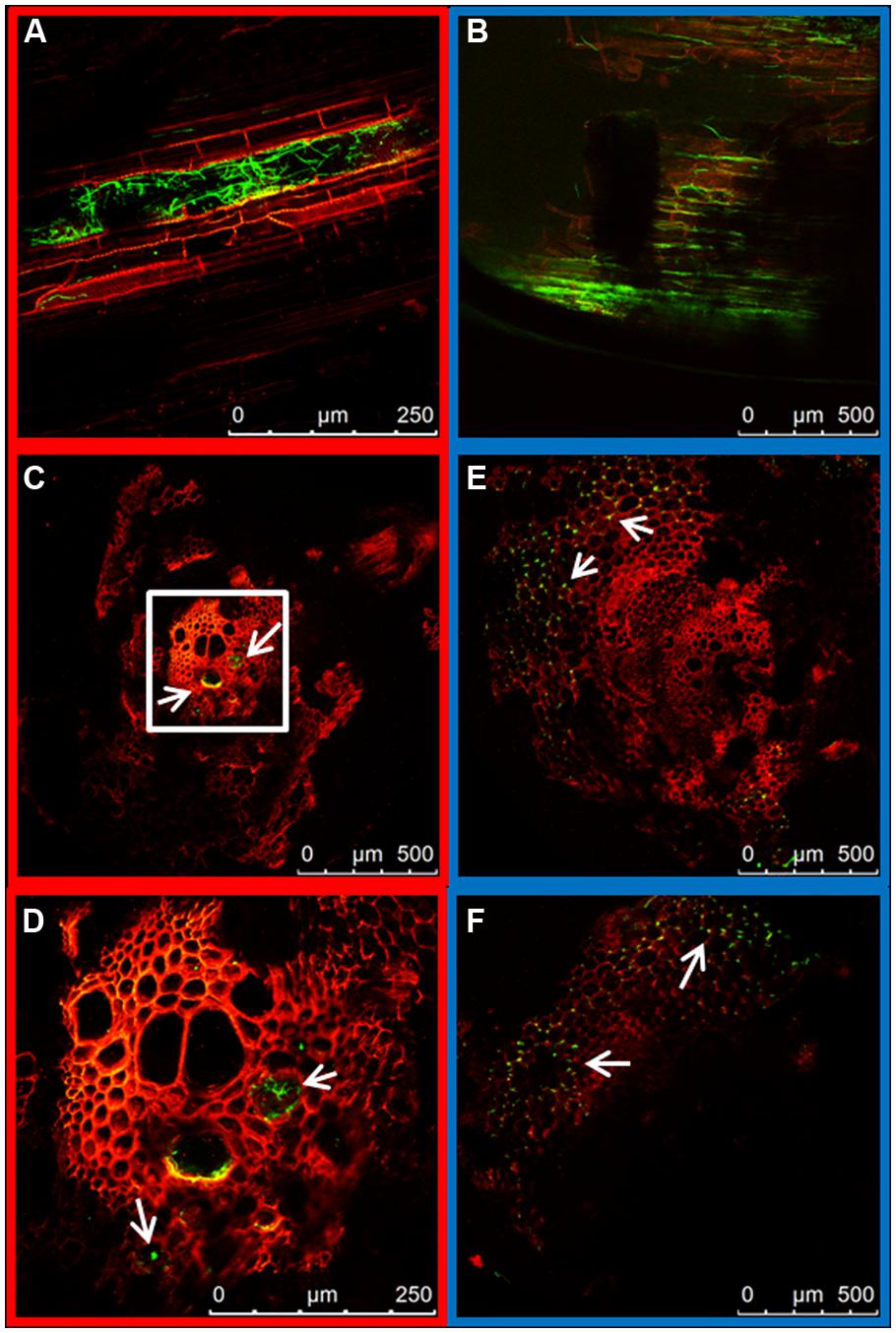
FIGURE 3. Internal colonization of the root system by HV (FOP-SP1) and WV (FOP-SP4) F. oxysporum f. sp. phaseoli strains. Mycelial growth of FOP-SP1 and FOP-SP4 inside the taproot was visualized in longitudinal sections (A,B) and cross sections (C–F) of infected plants 1–3 dpi. The arrows indicate hyphae of FOP-SP1 growing inside the vessels of the central cylinder of the taproot (red framed images) and hyphae of FOP-SP4 colonizing the cortex of the taproot (blue framed images).
Fungal mycelia could also be seen in the root crown at 7 dpi. Two differences could be observed between the two strains. First, root crowns of plants inoculated with the HV strain had more green fluorescent hyphae than those inoculated with the WV strain (Figures 4A,B respectively). Second, the asymmetrical distribution of hyphae in xylem vessels and the intercellular spaces of cortex, observed in the taproot, was maintained. The colonization of the vascular system by the HV strain was preferentially observed in adjacent vessels, indicating that hyphae can penetrate the vessels (Figures 4C,E). Mycelium of the WV strain was mostly detected around the cells of the cortex, although some hyphae could also be seen growing inside the xylem vessels (Figures 4D,F). Plants inoculated with the HV strain were rated 1 and those inoculated with the WV strain were rated 0.
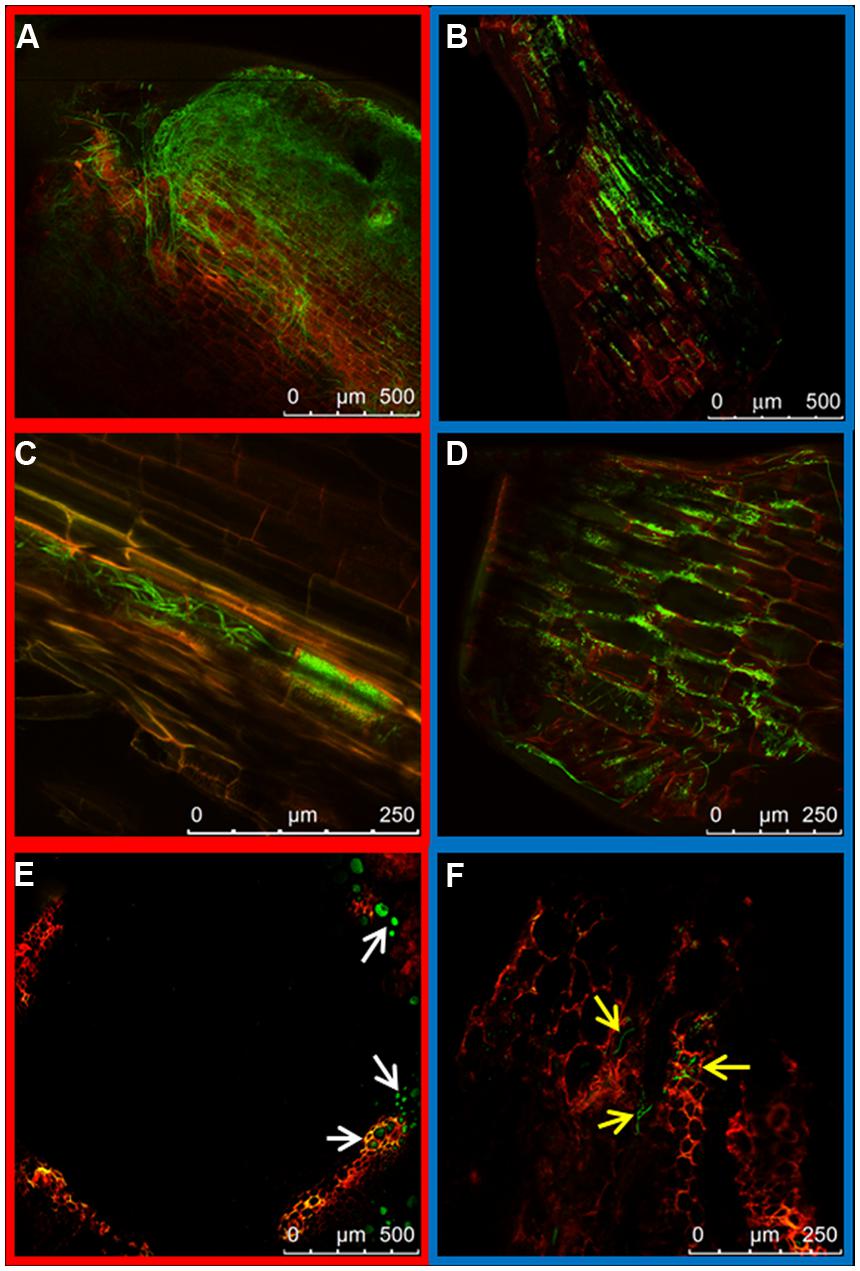
FIGURE 4. Colonization of the root crown by HV (FOP-SP1) and WV (FOP-SP4) F. oxysporum f. sp. phaseoli strains. The growth of mycelium of FOP-SP1 and FOP-SP4 in the root crown region was visualized in longitudinal sections (A–D) and cross sections (E,F) of plants at 5–7 dpi. The hyphae of FOP-SP1 heavily colonized the cortex (A) and the xylem vessels (C and white arrows in E) of the root crown. The growth of hyphae of FOP-SP4 in the root crown predominantly occurred along the interstitial spaces of cortical cells (B,D) although some hyphae also colonized the xylem vessels (yellow arrows in F).
Early observations of plant hypocotyls (10 dpi) showed hyphae of the HV strain colonizing the inside of the xylem vessels (Figure 5A) while the hyphae of the WV strain were preferentially seen along the walls of parenchymal cells in longitudinal sections (Figure 5B). Inoculated plants were rated 2 and 1, respectively. This observation and the difference in the ratings were confirmed in cross sections at 14 dpi (Figures 5C,E) when disease rates reached 3 for plants inoculated with the HV strain and 2 for plants inoculated with the WV strain. Although xylem vessels containing growing hyphae were observed for both strains (Figures 5D,F), the incidence (proportion of colonized vessels) was dramatically different, reaching 81.06% (±8.88) for the HV strain and 11.39% (±7.03) for the WV strain. Cross sections of plants 21 dpi showed massive amounts of hyphae of the HV strain growing almost exclusively inside the xylem vessels (Figures 5G,H), while most of the hyphae of the WV strain were around the parenchymal cells (Figures 5I,J). At this time, plants inoculated with the HV strain were almost dead and showed complete necrosis of the crown region and extensive necrosis of the xylem system of the hypocotyl (disease rate 4). Plants inoculated with the WV strain had some chlorosis, necrotic crown and some symptoms of necrosis in the xylem system of the hypocotyl (disease rate 3).
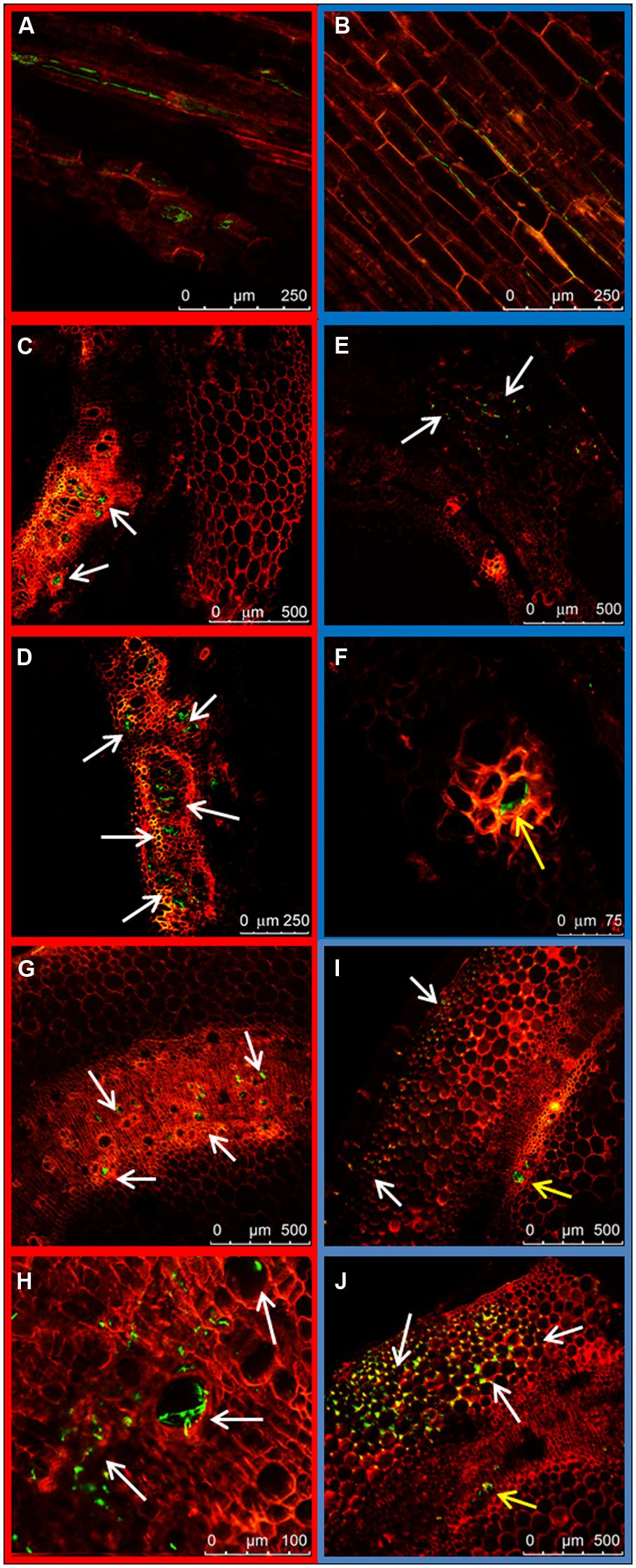
FIGURE 5. Colonization of the hypocotyl by HV (FOP-SP1) and WV (FOP-SP4) F. oxysporum f. sp. phaseoli strains. The growth of mycelium of FOP-SP1 (red framed images) and FOP-SP4 (blue framed images) was visualized in longitudinal sections (A,B) and cross sections (C–J). Hyphae of FOP-SP1 were detected growing inside the xylem vessels almost exclusively (arrows) at 10 dpi (A), 14 dpi (C,D), and 21 dpi (G,H). Hyphae of FOP-SP4 were mainly detected growing along the cells of cortex and parenchyma (white arrows), at 10 dpi (B), 14 dpi (E,F) and 21 dpi (I,J), although some hyphae were also seen inside the xylem vessels (yellow arrows).
Quantification of Fungal Growth
In order to determine whether the patterns of colonization showed by both strains were correlated to differences in fungal biomass, the growth of both strains of the pathogen inside the plant was evaluated by means of a qPCR assay. F. oxysporum DNA relative to that of common bean was measured by assaying the F. oxysporum EF1α and FTF2 genes of FOP-SP1 and FOP-SP4 strains, and the P. vulgaris actin gene.
The accumulation of fungal biomass in the root system was similar in plants inoculated either with the HV strain or the WV strain, and reached a maximum at 3 dpi (Figure 6). In contrast, The accumulation of mycelium both in the root crown region and the hypocotyl was higher in plants inoculated with the HV strain than in plants inoculated with the WV strain: 16 times more in root crown at 7 dpi and more than five times in hypocotyl at 21 dpi (Figure 6).
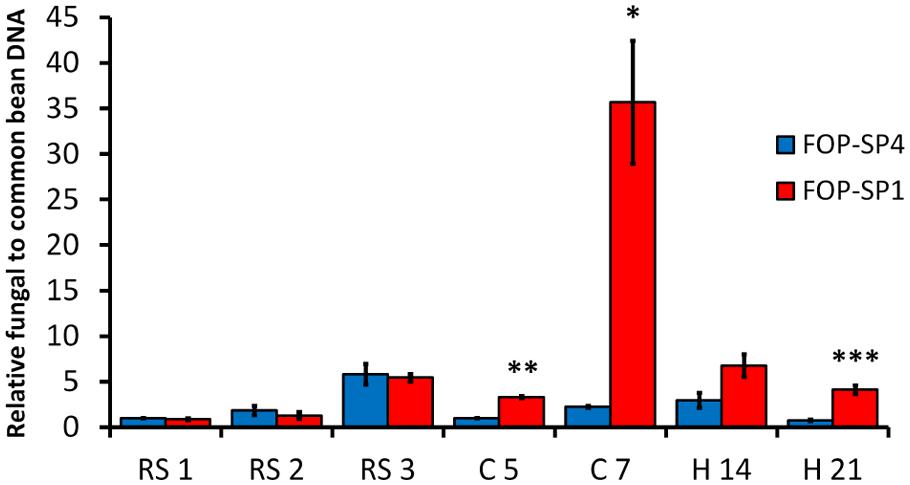
FIGURE 6. Quantification of fungal biomass in P. vulgaris plants colonized by HV and WV strains of F. oxysporum f. sp. phaseoli. FOP-SP4 DNA (blue bars) or FOP-SP1 DNA (red bars) relative to that of common bean was measured by assaying the fungal FTF2 gene and the plant actin gene by RT-qPCR using DNA extracted from the root system at 1, 2, and 3 dpi (RS1, RS2, and RS3), root crown at 5 and 7 dpi (C5 and C7) and hypocotyl at 14 and 21 dpi (H14 and H21). All measurements were referred to the value obtained for FOP-SP4 colonization at RS1 (arbitrary value of 1.0). Significative differences were observed for C5 (**p < 0.01), C7 (∗p < 0.05), and H21 (∗∗∗p < 0.001).
Taking together the confocal microscopy observations and the qPCR fungal biomass quantification results, two main differences emerge in the colonization pattern of HV and WV strains. First, the HV strain is a specialized vascular colonizer, while the WV strain, although also able to colonize the vascular system, is better at colonizing cortex and parenchyma. Second, the HV strain accumulates a significantly higher amount of mycelium in plant aerial regions than the WV strain.
Expression Analysis of Selected Fungal Virulence Genes
In order to understand the genetic basis of the differential colonization patterns displayed by HV and WV strains, we analyzed the in planta changes of expression of a selected group of genes already shown to be involved in virulence or suspected to be virulence factors (Table 2). RNA samples were purified from root (1, 2 and 3 dpi), crown (5 and 7 dpi) and hypocotyl (14 and 21 dpi) and used in RT-qPCR experiments to measure the expression of each gene analyzed (Figure 7). The most relevant changes of gene expression in root samples were the upregulation of RHR1 and EBR1 in both strains and the specific upregulation of SGE1 (fivefold increase) and FTF2 (threefold increase) in the WV strain. A drastic change in gene expression pattern related to the virulence of each strain was observed in crown at 5 dpi. With the exception of SGE1 and RHR2 all the genes analyzed showed upregulation (twofold or more) during the infection with the HV strain, reaching highest levels for FTF1, SIX1, and RHR1 (more than fivefold increase). Two days later (7 dpi) the same plant region showed significant upregulation of FNR1, PacC, FTF2 and RHR1 in the WV strain. Relatively high expression patterns (threefold or more) in hypocotyl colonized by the WV strain remained for RHR1 (14 and 21 dpi), PacC and FTF2 (21 dpi), and in the case of hypocotyl colonized by the HV strain, for PacC (21 dpi), FTF2 (14 and 21 dpi) and most significantly for FTF1 (14 dpi) and SIX1 and RHR1 (14 and 21 dpi).

TABLE 2. Fungal genes analyzed during colonization of Phaseolus vulgaris plants by weakly and highly virulent strains of F. oxysporum.
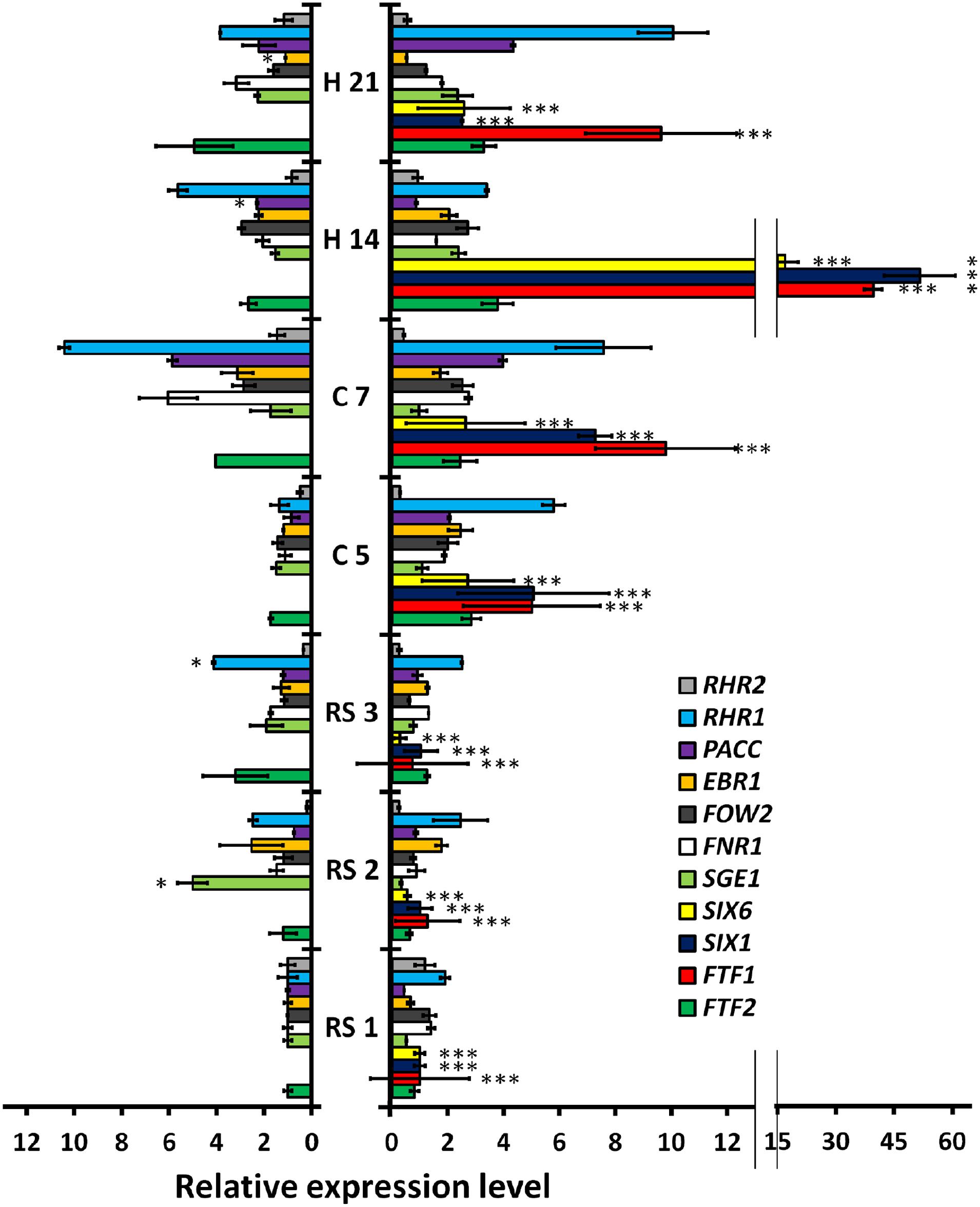
FIGURE 7. Quantitative reverse transcription polymerase chain reaction (RT-qPCR) analysis of time-course expression of eleven fungal genes in P. vulgaris plants colonized by HV and WV strains of F. oxysporum f. sp. phaseoli. The plant regions assayed and the time intervals after inoculation are indicated in the center of the figure: root system at 1, 2, and 3 dpi (RS1, RS2, ans RS3, respectively); root crown at 5 and 7 dpi (C5 and C7, respectively); and hypocotyl at 14 and 21 dpi (H14 and H21, respectively). The bars on the left show the results obtained in common bean plants colonized by the WV strain (FOP-SP4). The bars on the right show the corresponding results for plants colonized by the HV strain (FOP-SP1). The expression ratios were normalized by using the fungal EF1α gene as an endogenous control. An arbitrary value of 1.0 was denoted for the transcript level of each gene in plants colonized by the WV strain at RS1, except for the FTF1, SIX1, and SIX6 transcripts whose 1.0 values correspond to the transcript level of each gene in plants colonized by the HV strain at RS1. The results shown are the averages and their respective standard deviations obtained in three independent biological experiments. The expression level value differences between each pair of samples (the expression of the same gene in plants colonized by the HV strain or the WV strain) were tested using the t-test and indicated by (*p < 0.05) and (***p < 0.001).
Significant differences in the gene expression patterns between both strains were observed for FTF1, SIX1 and SIX6. The difference is obvious for FTF1, as it is specific to the HV strains, but both SIX1 and SIX6 are present in the genome of the WV strain although no expression could be detected at any stage of the infection process. Also, it is worth to note the upregulation of SGE1 in root tissues colonized by the WV strain at 2 dpi, and the upregulation of RHR1 in root crown and hypocotyl colonized by the HV strain at 5 and 21 dpi, respectively.
Expression Analysis of Selected Plant Defense Genes
We investigated the response of the pathogenesis related gene PR1 and two ET responsive factors (ERF1 and ERF2) in the plant tissues colonized either by the WV strain or the HV strain (Figure 8). Expression of PR5 could not be detected in a consistent way and was discarded for further analysis. PR1 showed an early downregulation in plant roots infected either by the WV strain or the HV strain, but 1 day later its expression was 50 times higher in roots infected by the WV strain than in roots infected by the HV strain. This difference was progressively attenuated and disappeared in infected root crown at 5 dpi. The expression of the ERF1 gene was similar in plants infected either by the WV strain or the HV strain. Contrary to the PR1 expression pattern, the induction of the ERF1 gene reached a maximum in roots at 1 dpi and then slowly decreased. ERF2 was also induced at early stages of infection but it was significantly overexpressed in roots infected by the HV strain. Therefore, two main findings must be highlighted. First, SAR response is repressed at the beginning of the infection while the ET-mediated response is activated in plants inoculated either with the HV or WV strains. Second, SAR response is delayed in plants infected by the HV strain with respect to those inoculated with the WV strain, while the ET responsive factor ERF2 is significantly induced.
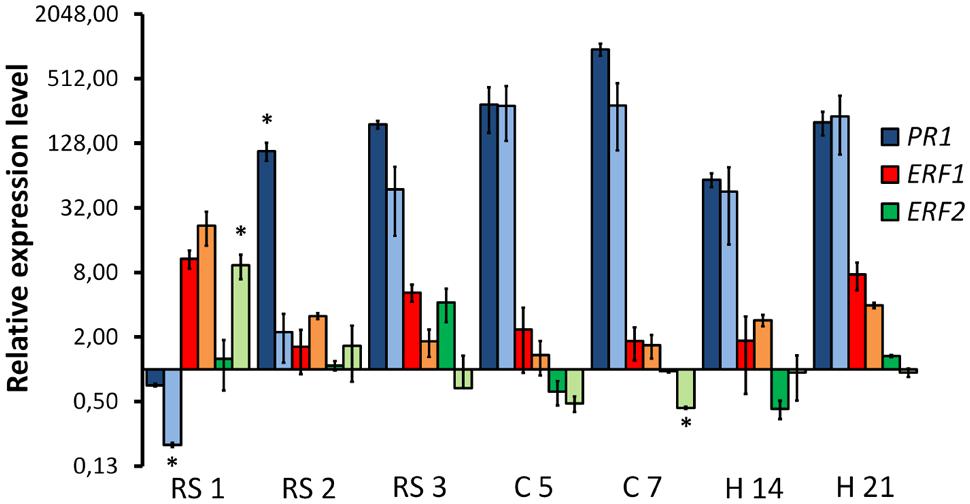
FIGURE 8. Quantitative reverse transcription polymerase chain reaction analysis of time-course expression of common bean genes involved in the defense response in plants colonized by HV and WV strains of F. oxysporum f. sp. phaseoli. The plant regions assayed and the time intervals after inoculation are indicated in the X axis: root system at 1, 2, and 3 dpi (RS1, RS2, and RS3, respectively); root crown at 5 and 7 dpi (C5 and C7, respectively); and hypocotyl at 14 and 21 dpi (H14 and H21, respectively). The relative expression measurements in the Y axis are indicated in a logarithmic scale. Dark bars for each color indicate expression of the corresponding gene in plants colonized by the WV strain, and light bars indicate expression of the corresponding gene in plants colonized by the HV strain. The expression ratios were normalized by using the common bean actin gene as endogenous control. The levels of expression for a pair of measurements (dark and light bars) were tested using the t-test and significative differences (p < 0.05) indicated by ∗.
FTF1 Expression in PTFT1::GFP Transformants
The expression profile of the gene encoding the virulence factor FTF1 above described is in contrast with former reports which showed maximum expression in plant stem 24 h post inoculation (Ramos et al., 2007). In order to confirm our present observations, we obtained transformants of strain FOP-SP1 with a construct that included the GFP gene under the control of the FTF1 promoter. The first observations of GFP-expressing mycelium in colonized plant tissues corresponded to the crown region at 7 dpi (Figure 9A) in accordance with a 10-fold overexpression of the gene. From this time GFP fluorescence was clearly visible inside the xylem vessels of hypocotyls at 10 dpi (Figure 9B) and 14 dpi (Figures 9C,D). Most of the xylem vessels in hypocotyls at 21 dpi were occluded by fungal mycelium (Figures 9E,F).
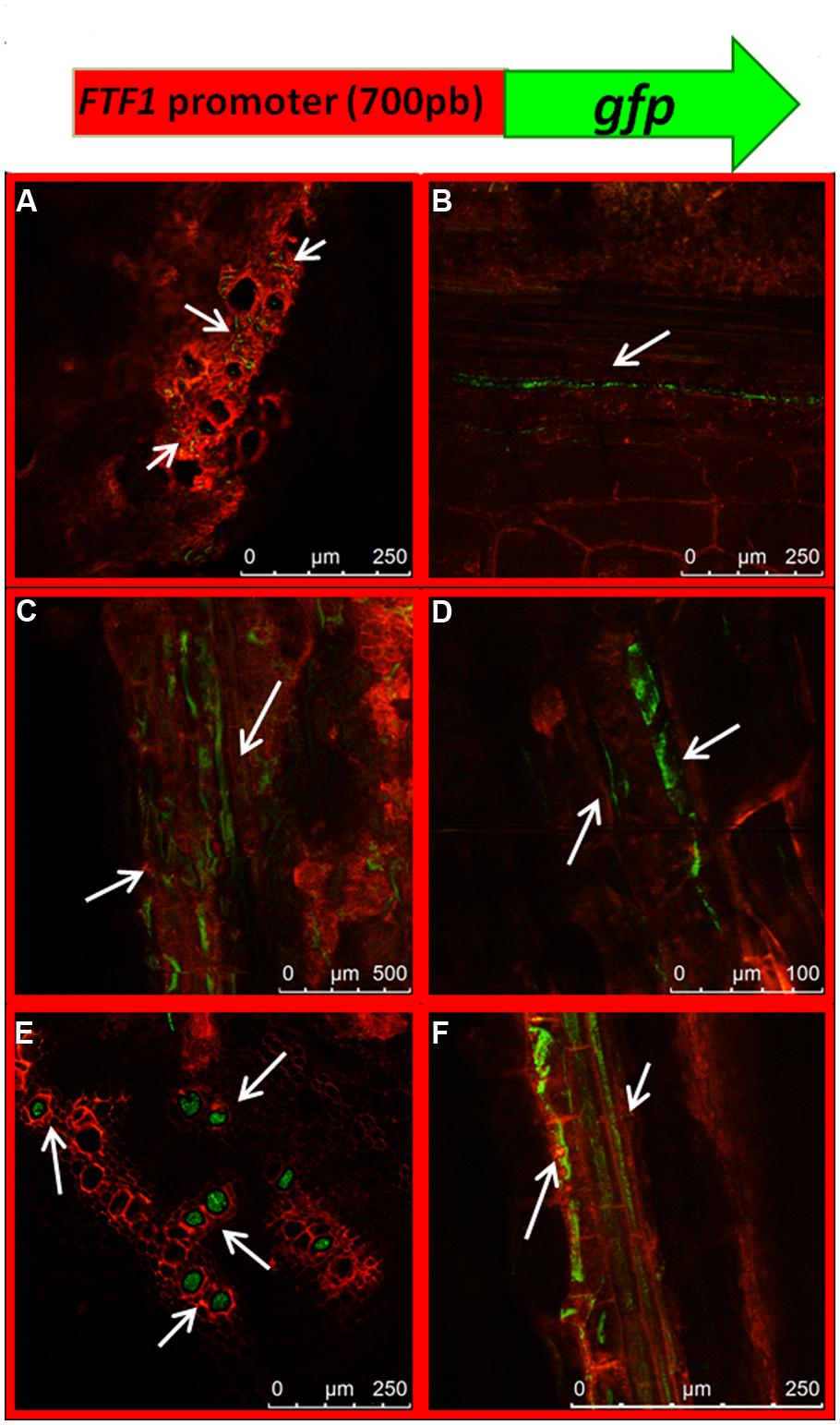
FIGURE 9. Dynamics of plant colonization by a FOP-SP1 transformant expressing the GFP gene under the control of the FTF1 promoter (PFTF1::GFP). The transgene expressed in the transformant PFTF1::GFP is schematically depicted at the top of the figure. Arrows indicate mycelial growth as visualized in cross sections (A,E) and longitudinal sections (B–D,F) of root crown at 7 dpi (A) and hypocotyl at 10 dpi (B), 14 dpi (C,D) and 21 dpi (E,F).
The expression pattern of FTF1 in the confocal microscopy analysis matches the expression profile obtained in the RT-qPCR experiments, and confirms that upregulation of FTF1 takes place in crown and hypocotyl from 7 dpi, under the present experimental conditions.
Discussion
The vast majority of microscopy analysis of plant colonization by F. oxysporum have been related to the details of particular interactions (Bishop and Cooper, 1983; Czymmek et al., 2007; Nahalkova et al., 2008; Li et al., 2011), or the differences in the colonization of susceptible and resistant cultivars (Bishop and Cooper, 1983; Zvirin et al., 2010; Jiménez-Fernández et al., 2013; Lü et al., 2013). Almost no attention has been paid to the study of differences in plant colonization based on the virulence of the infecting strains. Most likely this is because of the scarce availability of strains able to infect the same host while displaying a range of virulence, which to our knowledge has only been shown in F. oxysporum f. sp. phaseoli (Alves-Santos et al., 2002a) and F. oxysporum f. sp. dianthi (Gómez-Lama Cabanás et al., 2012).
Fusarium oxysporum f. sp. phaseoli strains may be classified as weakly and highly virulent against P. vulgaris (Alves-Santos et al., 2002a) or as weakly, highly and supervirulent against P. coccineus (de Vega-Bartol et al., 2011). In a former report (García-Sánchez et al., 2010) we described two main differences in the plant colonization pattern of a HV strain (FOP-SP2) and a WV strain (FOP-SP8), the latter being slower in the rate of xylem colonization and with a reduced capability to colonize xylem vessels. With the aim to gain deeper insight into host colonization features, we have analyzed two more strains (FOP-SP1 and FOP-SP4) and used a hydroponics system which allows for easier access to plant roots for sectioning and confocal microscopy visualization.
Penetration of host roots by F. oxysporum is assumed to occur by direct penetration or through wounds (Nelson, 1981). In the absence of wounds, most reports show that the preferred zone of ingress is the root apex (Sarrocco et al., 2007; Jiménez-Fernández et al., 2013) or roothairs (Lagopodi et al., 2002). We did not observe differences between the HV and the WV strains with regards to the ingress area. Both strains showed a preference for the junctions of the lateral roots with the taproot, where most likely they can take advantage of discontinuities in the plant epidermis. The main difference observed at this stage between the HV and WV strains was the ability of the HV strain to reach the root vascular cylinder and quickly progress toward the root crown. The specialized dissemination of the HV strain inside the root is not related to a better growth rate as the total amount of fungal biomass in the root is very similar for both strains. The only differential expression detected in fungal genes corresponds to SGE1 at 2 dpi and RHR1 at 3 dpi, both upregulated in the WV strain. The genes which expression is exclusive to the HV strain (FTF1, SIX1 and SIX6) do not show significant upregulation in root, indicating that their induction is not required for penetration nor for the colonization of this part of the plant. The absence of significant expression of FTF1 in roots is in agreement with previous reports (Ramos et al., 2007). In contrast with these results, some differences related to virulence are found in the expression of plant defense genes. PR1 is significantly downregulated during the first 24 h of infection by the HV strain, while the opposite occurs to the common bean homologue of ERF2, one of the genes involved in the ET/JA defense response (Fujimoto et al., 2000). Ethylene and JA biosyntheses are activated by a variety of forms of stress, including the biotic stress induced by pathogens, and both pathways converge in the transcriptional activation of ERF1 (Lorenzo et al., 2002) what makes it a good marker to detect the activation of both phytohormones. The homologue of ERF1 in common bean is overexpressed in plants infected either with the WV strain or the HV strain at 1 dpi. It is worth to note that this induction of ERF1 is accompanied by the already mentioned downregulation of PR1, which occurs not only in plants infected by the HV strain, but also when they are infected by the WV strain, although to a lesser extent. Diverse analysis have revealed that SA, ET and JA pathways influence the outcome of the disease produced by F. oxysporum in Arabidopsis (reviewed in Berrocal-Lobo and Molina, 2008). Our results indicate that higher PR1 gene and lower ERF genes expression during the first 2 days of root infection correlate with a higher rate of parenchyma colonization and a lower rate of vascular cylinder colonization (infection with the WV strain), and vice versa (infection with the HV strain).
Vascular pathogens are characterized by their ability to colonize the vascular system of the host. The collapse of the xylem vessels, caused by the accumulation of fungal mycelium and host defensive molecules, produces a drastic reduction in the transport of water to the aerial parts of the plant and finally its death. The architecture of the stem vascular system develops in the crown area where the primordial ring of xylem vessels is formed. We observed in this area the most drastic differences, either in the amount of fungal biomass and its distribution, between the strains analyzed. Expression of several genes in both strains at this time point seems to be important to sustain crown colonization, namely FTF2, FNR1, EBR1, PacC and RHR1. The upregulation of FTF2 actually begins in the root, in the case of the WV strain, and it is surprising as former analyses carried out by our group suggested this gene was not involved in host infection (unpublished results). FTF2 and EBR1 are members of different gene family expansions characteristic of F. oxysporum. Both genes appear as single copies in the genomes of several Fusarium spp., but the paralogues EBR2, EBR3, EBR4 and the copies of FTF1 are present only in F. oxysporum (de Vega-Bartol, unpublished; Jonkers et al., 2013). However, the relative importance of the paralogues and the single copy genes are different. EBR1 is the most abundant transcript of the EBR family during host infection and its deletion results in a reduction of virulence, while the rest of the paralogues do not seem to have a role in host colonization (Jonkers et al., 2013). In the case of FTF2, the most abundant transcripts of the family during common bean infection are produced by the copies of FTF1 and, although we have not yet analyzed a mutant deleted in FTF2, FTF1 is exclusive to the HV strains of F. oxysporum f. sp. phaseoli.
The upregulation of FNR1 and RHR1 seems to be related to the adaptation of fungal nutrition to a new compromising environment. FNR1 has been described to mediate the adaptation to nitrogen-poor conditions in planta (Divon et al., 2006), such as those potentially found inside the xylem vessels. The A. nidulans rhaR gene encodes a transcription factor that positively regulates the expression of both the enzymes that liberate L-rhamnose from complex plant substrates (alpha-L-rhamnosidases) as well as those involved in the assimilation of this sugar, and hence confers A. nidulans the ability to adapt its metabolism to the availability of L-rhamnose (Tamayo-Ramos et al., 2012; Pardo and Orejas, 2014). Interestingly, RNA-seq data obtained from the Aspergillus Genome database (AspGD) suggest that rhaR could be preferentially expressed under nitrogen limiting conditions. Therefore, it is conceivable that once the mycelium of F. oxysporum grows inside the vascular system, where the preferred sugars are absent, it must obtain other carbon sources from the cell wall components of the lignified xylem vessels. There is a paralogue of RHR1 in F. oxysporum, named RHR2, which does not show significant changes in expression during the colonization process, neither in the WV strain nor the HV strain.
PacC mediates regulation of gene expression by ambient pH in fungi (Peñalva et al., 2008). It has been shown in F. oxysporum f. sp. lycopersici that PacC transcript levels rise when the fungus is grown in alkaline conditions (Caracuel et al., 2003). Here we show that PacC transcript levels increase at 7 dpi, in coincidence with the beginning of the colonization of the hypocotyl xylem vessels. This suggests that ambient pH inside the vessels is alkaline and that genes expressed under alkaline conditions are required for their colonization by either HV or WV strains.
The highest levels of gene expression were detected in the hypocotyl at 14 dpi and correspond to FTF1, which is unique to the HV strains, and the effector encoding genes SIX1 and SIX6. At that time, observations of plants infected with the HV strain show that growth is restricted to the xylem vessels, thus indicating that FTF1, SIX1 and SIX6 are expressed during colonization of the vascular system. SIX1 and SIX6 are recognized as genuine effectors, as they contribute to the general virulence of F. oxysporum f. sp. lycopersici (Rep et al., 2004; Gawehns et al., 2014). Homologues of these genes have been found in other formae speciales like F. oxysporum f. sp. cubense (Niño-Sánchez, unpublished) and F. oxysporum f. sp. phaseoli (this work), although only the copies present in the HV strain are expressed during infection. It cannot be ruled out that the SIX1 and SIX6 alleles in the genome of the WV strain are not functional. Alternatively, the expression of SIX1 and SIX6 might need a transcription activator not present in the WV strain, such as FTF1. The expression of FTF1, SIX1 and SIX6, was detected at low levels in root 1 dpi, in accordance with the former results obtained for SIX1 during tomato infection (van der Does et al., 2008). Upregulation of the three genes begins in the root crown 5 dpi, matching the accumulation of mycelium in this area and the beginning of massive colonization of xylem vessels.
The classical view of plant colonization by fungal vascular wilt pathogens assumes that, after penetration, the fungi colonize the cortical cells and then hyphae migrate intercellularly toward the vascular parenchyma cells and invade xylem vessels. Once the xylem is reached, the mycelium is mostly confined inside the vessels, until the necrosis of host tissues allows for general colonization (Yadeta and Thomma, 2013). In this work we have shown that a high level of virulence correlates with a high ability to quickly infect and colonize the plant vascular system, with almost no colonization of the root cortex, and that WV strains are efficient colonizers of host tissues before they become necrotized. Also, our data suggest a correlation between virulence and the expression of some components exclusive of the genome of HV strains, such as the FTF1 transcription factor, and other components that, although not exclusive, are only expressed in HV strains, such as the effectors SIX1 and SIX6. FTF1 paralogs are part of the unique sequences found in the genome of F. oxysporum, designated as lineage specific (Ma et al., 2010). It has been proposed that F. oxysporum lineage specific sequences located in dispensable chromosomes are the basis of host specialization and could explain the polyphyletic origins of most formae speciales (Ma et al., 2010). Our results show that they also contribute to explain F. oxysporum virulence.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by grant AGL2012-39876-C02-01 from MINECO (Spain). JN-S and VC-dC are the recipients of fellowships AP2009-3559 and AP2010-2742, respectively, from MECD (Spain). VT held a postdoctoral position financed by grant AGL2012-39876-C02-01. We thank M. Orejas for valuable suggestions on the RHR genes, I. Martín for excellent advice on the preparation of plant samples for microscopic examination, R. Frandsen for advice on the use of the pRF-HU vectors, and S. Sukno for critical reading of the manuscript and many valuable suggestions.
References
Alves-Santos, F. A., Cordeiro-Rodrigues, L., Sayagués, J. M., Martín-Domínguez, R., García-Benavides, P., Crespo, M. C., et al. (2002a). Pathogenicity and race characterization of Fusarium oxysporum f. sp. phaseoli isolates from Spain and Greece. Plant Pathol. 51, 605–611. doi: 10.1046/j.1365-3059.2002.00745.x
Alves-Santos, F. M., Ramos, B., García-Sánchez, M. A., Eslava, A. P., and Díaz-Mínguez, J. M. (2002b). A DNA-based procedure for in planta detection of Fusarium oxysporum f. sp. phaseoli. Phytopathology 92, 237–244. doi: 10.1094/PHYTO.2002.92.3.237
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Alves-Santos, F. M., Benito, E. P., Eslava, A. P., and Díaz-Mínguez, J. M. (1999). Genetic diversity of Fusarium oxysporum strains from common bean fields in Spain. Appl. Environ. Microbiol. 65, 3335–3340.
Armstrong, G. M., and Armstrong, J. K. (1975). Reflections on the wilt fusaria. Annu. Rev. Phytopathol. 13, 95–103. doi: 10.1146/annurev.py.13.090175.000523
Berrocal-Lobo, M., and Molina, A. (2008). Arabidopsis defense response against Fusarium oxysporum. Trends Plant Sci. 13, 145–150. doi: 10.1016/j.tplants.2007.12.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bishop, C. D., and Cooper, R. M. (1983). An ultrastructural study of root invasion in three vascular wilt diseases. Physiol. Plant Pathol. 22, 15–27. doi: 10.1016/S0048-4059(83)81034-0
Bustin, S. A. (2000). Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 25, 169–193. doi: 10.1677/jme.0.0250169
Caracuel, Z., Roncero, M., Espeso, E., González-Verdejo, C., García-Maceira, F., and Di Pietro, A. (2003). The pH signalling transcription factor PacC controls virulence in the plant pathogen Fusarium oxysporum. Mol. Microbiol. 48, 765–779. doi: 10.1046/j.1365-2958.2003.03465.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Czymmek, K. J., Fogg, M., Powell, D. H., Sweigard, J., Park, S. Y., and Kang, S. (2007). In vivo time-lapse documentation using confocal and multi-photon microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium oxysporum. Fungal Genet. Biol. 44, 1011–1023. doi: 10.1016/j.fgb.2007.01.012
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Vega-Bartol, J. J., Martín-Dominguez, R., Ramos, B., García-Sánchez, M. A., and Díaz-Mínguez, J. M. (2011). New Virulence Groups in Fusarium oxysporum f. sp.phaseoli: the expression of the gene coding for the transcription factor ftf1 correlates with virulence. Phytopathology 101, 470–479. doi: 10.1094/PHYTO-09-10-0252
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Divon, H. H., Ziv, C., Davydov, O., Yarden, O., and Fluhr, R. (2006). The global nitrogen regulator, FNR1, regulates fungal nutrition-genes and fitness during Fusarium oxysporum pathogenesis. Mol. Plant Pathol. 7, 485–497. doi: 10.1111/j.1364-3703.2006.00354.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Durrant, W. E., and Dong, X. (2004). Systemic acquired resistance. Annu. Rev. Phytopathol. 42, 185–209. doi: 10.1146/annurev.phyto.42.040803.140421
Frandsen, R. J., Andersson, J. A., Kristensen, M. B., and Giese, H. (2008). Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 9:70. doi: 10.1186/1471-2199-9-70
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fujimoto, S. Y., Ohta, M., Usui, A., Shinshi, H., and Ohme-Takagi, M. (2000). Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 12, 393–404. doi: 10.1105/tpc.12.3.393
García-Sánchez, M., Martín-Rodrigues, N., Ramos, B., de Vega-Bartol, J. J., Perlin, M. H., and Díaz-Mínguez, J. M. (2010). fost12, the Fusarium oxysporum homolog of the transcription factor Ste12, is upregulated during plant infection and required for virulence. Fungal Genet. Biol. 47, 216–225. doi: 10.1016/j.fgb.2009.11.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gawehns, F., Houterman, P. M., Ichou, F. A., Michielse, C. B., Hijdra, M., Cornelissen, B. J. C., et al. (2014). The Fusarium oxysporum effector Six6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant Microbe Interact. 27, 336–348. doi: 10.1094/MPMI-11-13-0330-R
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gómez-Lama Cabanás, C., Valverde-Corredor, A., and Artés, E. (2012). Molecular analysis of Spanish populations of Fusarium oxysporum f. sp.dianthi demonstrates a high genetic diversity and identifies virulence groups in races 1 and 2 of the pathogen. Eur. J. Plant Pathol. 132, 561–576. doi: 10.1007/s10658-011-9901-4
Imazaki, L., Kurahashi, M., Lida, Y., and Tsuge, T. (2007). Fow2, a Zn(II)2Cys6-type transcription regulator, controls plant infection of the vascular wilt fungus Fusarium oxysporum. Mol. Microbiol. 63, 737–753. doi: 10.1111/j.1365-2958.2006.05554.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jiménez, T., Martín, I., Labrador, E., and Dopico, B. (2007). A chickpea Kunitz trypsin inhibitor is located in cell wall of elongating seedling organs and vascular tissue. Planta 226, 45–55. doi: 10.1007/s00425-006-0465-z
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jiménez-Fernández, D., Landa, B. B., Kang, S., Jiménez-Díaz, R. M., and Navas-Cortés, J. A. (2013). Quantitative and microscopic assessment of compatible and incompatible interactions between chickpea cultivars and Fusarium oxysporum f. sp. ciceris Races. PLoS ONE 8:e61360. doi: 10.1371/journal.pone.0061360
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jonkers, W., Xayamongkhon, H., Haas, M., Olivain, C., van der Does, H. C., Broz, K., et al. (2013). EBR1 genomic expansion and its role in virulence of Fusarium species. Environ. Microbiol. 16, 1982–2003. doi: 10.1111/1462-2920.12331
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lagopodi, A. L., Ram, A. F., Lamers, G. E., Punt, P. J., Van den Hondel, C. A., Lugtenberg, B. J., et al. (2002). Novel aspects of tomato root colonization and infection by Fusarium oxysporum f. sp. radicis-lycopersici revealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker. Mol. Plant Microbe Interact. 15, 172–179. doi: 10.1094/MPMI.2002.15.2.172
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, C., Chen, S., Zuo, C., Sun, Q., Ye, Q., Yi, G., et al. (2011). The use of GFP-transformed isolates to study infection of banana with Fusarium oxysporum f. sp.cubense race 4. Eur. J. Plant Pathol. 131, 327–340. doi: 10.1007/s10658-011-9811-5
Lorenzo, O., Piqueras, R., Sánchez-Serrano, J. J., and Solano, R. (2002). ETHYLENE RESPONSE FACTOR 1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 15, 165–178. doi: 10.1105/tpc.007468
Lü, G., Guo, S., Zhang, H., Geng, L., Martyn, R. D., and Xu, Y. (2013). Colonization of Fusarium wilt-resistant and susceptible watermelon roots by a green-fluorescent-protein-tagged isolate of Fusarium oxysporum f.sp. niveum. J. Phytopathol. 162, 228–237. doi: 10.1111/jph.12174
Ma, L. J., van der Does, H. C., Borkovich, K. A., Coleman, J. J., Daboussi, M. J., Di Pietro, A., et al. (2010). Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464, 367–373. doi: 10.1038/nature08850
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maor, R., Puyesky, M., Horwitz, B. A., and Sharon, A. (1998). Use of green fluorescent protein (GFP) for studying development and fungal-plant interaction in Cochliobolus heterostrophus. Mycol. Res. 102, 491–496. doi: 10.1017/S0953756297005789
Michielse, C. B., and Rep, M. (2009). Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 10, 311–324. doi: 10.1111/j.1364-3703.2009.00538.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mullins, E., Chen, X., Romaine, P., Raina, R., Geiser, D., and Kang, S. (2001). Agrobacterium-mediated transformation of Fusarium oxysporum: an efficient tool for insertional mutagenesis and gene transfer. Phytopathology 91, 173–180. doi: 10.1094/PHYTO.2001.91.2.173
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nahalkova, J., Fatehi, J., Olivain, C., and Alabouvette, C. (2008). Tomato root colonization by fluorescent-tagged pathogenic and protective strains of Fusarium oxysporum in hydroponic culture differs from root colonization in soil. FEMS Microbiol. Lett. 286, 152–157. doi: 10.1111/j.1574-6968.2008.01241.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nelson, P. E. (1981). “Life cycle and epidemiology of Fusarium oxysporum,” in Fungal wilt Diseases of Plants, eds M. E. Mace, A. A. Bell, and C. H. Beckman (New York: Academic Press), 51–80.
Nucci, M., and Anaissie, E. (2007). Fusarium infections in inmunocompromised patients. Clin. Microbiol. Rev. 20, 695–704. doi: 10.1128/CMR.00014-07
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
O’Donnell, K., Gueidan, C., Sink, S., Johnston, P., Crous, P., Glenn, A., et al. (2009). A two-locus DNA sequence database for typing plant and human pathogens within the Fusarium oxysporum species complex. Fungal Genet. Biol. 46, 936–948. doi: 10.1016/j.fgb.2009.08.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pardo, E., and Orejas, M. (2014). The Aspergillus nidulans Zn(II)2Cys6 transcription factor AN5673/RhaR mediates L-rhamnose utilization and the production of alpha-L-rhamnosidases. Microb. Cell Fact. 13, 161.
Peñalva, M. A., Tilburn, J., Bignell, E., and Arst, H. N. (2008). Ambient pH gene regulation in fungi: making connections. Trends Microbiol. 16, 291–300. doi: 10.1016/j.tim.2008.03.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pfaffl, M. W., Horgan, G. W., and Dempfle, L. (2002). Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 30, e36. doi: 10.1093/nar/30.9.e36
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ramírez-Suero, M., Khanshour, A., Martinez, Y., and Rickauer, M. (2010). A study on the susceptibility of the model legume plant Medicago truncatula to the soil-borne pathogen Fusarium oxysporum. Eur. J. Plant Pathol. 126, 517–530. doi: 10.1007/s10658-009-9560-x
Ramos, B. (2005). Análisis de factores de patogenicidad y virulencia en el hongo Fusarium oxysporum f. sp. phaseoli. Ph.D. thesis, Universidad de Salamanca, Salamanca, Spain.
Ramos, B., Alves-Santos, F. M., García-Sánchez, M. A., Martín-Rodrigues, N., Eslava, A. P., and Díaz-Mínguez, J. M. (2007). The gene coding for a new transcription factor (ftf1) of Fusarium oxysporum is only expressed during infection of common bean. Fungal Genet. Biol. 44, 864–876. doi: 10.1016/j.fgb.2007.03.003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rep, M., van der Does, H. C., Meijer, M., van Wijk, R., Houterman, P. M., Dekker, H. L., et al. (2004). A small, cysteine-rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I-3-mediated resistance in tomato. Mol. Microbiol. 53, 1373–1383. doi: 10.1111/j.1365-2958.2004.04177.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sarrocco, S., Falaschi, N., Vergara, M., Nicoletti, F., and Vannacci, G. (2007). Use of Fusarium oxysporum f. sp. dianthi transformed with marker genes to follow colonization of carnation roots. J. Plant Pathol. 89, 47–54.
Sukno, S., García, V., Shaw, B., and Thon, M. (2008). Root infection and systemic colonization of maize by Colletotrichum graminicola. Appl. Environ. Microbiol. 74, 823–832. doi: 10.1128/AEM.01165-07
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tamayo-Ramos, J. A., Flipphi, M., Pardo, E., Manzanares, P., and Orejas, M. (2012). L-Rhamnose induction of Aspergillus nidulans alpha-L-rhamnosidase genes is glucose repressed via a CreA-independent mechanism acting at the level of inducer uptake. Microb. Cell Fact. 11, 26. doi: 10.1186/1475-2859-11-26
van der Does, H. C., Duyvesteijn, R. G. E., Goltstein, P. M., van Schie, C. C. N., Manders, E. M. M., Cornelissen, B. J. C., et al. (2008). Expression of effector gene SIX1 of Fusarium oxysporum requires living plant cells. Fungal Genet. Biol. 45, 1257–1264. doi: 10.1016/j.fgb.2008.06.002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yadeta, K. E., and Thomma, B. P. H. J. (2013). The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 4:97. doi: 10.3389/fpls.2013.00097
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhao, C., Waalwijk, W., de Wit, P. J., van der Lee, T., and Tang, D. (2011). EBR1, a novel Zn2Cys6, transcription factor, affects virulence and apical dominance of the hyphal tip in Fusarium graminearum. Mol. Plant Microb. Interact. 24, 1407–1418. doi: 10.1094/MPMI-06-11-0158
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: Fusarium oxysporum, Phaseolus vulgaris, confocal microscopy, virulence, effector
Citation: Niño-Sánchez J, Tello V, Casado-del Castillo V, Thon MR, Benito EP and Díaz-Mínguez JM (2015) Gene expression patterns and dynamics of the colonization of common bean (Phaseolus vulgaris L.) by highly virulent and weakly virulent strains of Fusarium oxysporum. Front. Microbiol. 6:234. doi: 10.3389/fmicb.2015.00234
Received: 12 January 2015; Accepted: 10 March 2015;
Published online: 02 April 2015
Edited by:
Helio K. Takahashi, Universidade Federal de Sao Paulo, BrazilReviewed by:
Todd B. Reynolds, University of Tennessee, USAMaría Teresa González Jaén, Complutense University of Madrid, Spain
Copyright © 2015 Niño-Sánchez, Tello, Casado-del Castillo, Thon, Benito and Díaz-Mínguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José María Díaz-Mínguez, Departamento de Microbiología y Genética, Instituto Hispano-Luso de Investigaciones Agrarias, Universidad de Salamanca, C/Río Duero 12, Villamayor, 37185 Salamanca, Spain josediaz@usal.es
† These authors have contributed equally to this work.