 Sabine Delannoy1
Sabine Delannoy1 Byron D. Chaves2Sarah A. Ison2Hattie E. Webb2
Byron D. Chaves2Sarah A. Ison2Hattie E. Webb2 Lothar Beutin3José Delaval4Isabelle Billet5
Lothar Beutin3José Delaval4Isabelle Billet5 Patrick Fach1*
Patrick Fach1*- 1Food Safety Laboratory, Université Paris-Est, Anses (French Agency for Food, Environmental and Occupational Health and Safety), Platform IdentyPath, Maisons-Alfort, France
- 2Department of Animal and Food Sciences, Texas Tech University, Lubbock, TX, USA
- 3Division of Microbial Toxins, National Reference Laboratory for Escherichia coli, Federal Institute for Risk Assessment, Berlin, Germany
- 4Laboratoire de Touraine, (LDA37) Conseil Départemental, Tours, France
- 5SAS Charal Groupe Bigard, Cholet, France
Current methods for screening Enterohemorrhagic Escherichia coli (EHEC) O157 and non-O157 in beef enrichments typically rely on the molecular detection of stx, eae, and serogroup-specific wzx or wzy gene fragments. As these genetic markers can also be found in some non-EHEC strains, a number of “false positive” results are obtained. Here, we explore the suitability of five novel molecular markers, espK, espV, ureD, Z2098, and CRISPRO26:H11 as candidates for a more accurate screening of EHEC strains of greater clinical significance in industrialized countries. Of the 1739 beef enrichments tested, 180 were positive for both stx and eae genes. Ninety (50%) of these tested negative for espK, espV, ureD, and Z2098, but 12 out of these negative samples were positive for the CRISPRO26:H11 gene marker specific for a newly emerging virulent EHEC O26:H11 French clone. We show that screening for stx, eae, espK, and espV, in association with the CRISPRO26:H11 marker is a better approach to narrow down the EHEC screening step in beef enrichments. The number of potentially positive samples was reduced by 48.88% by means of this alternative strategy compared to the European and American reference methods, thus substantially improving the discriminatory power of EHEC screening systems. This approach is in line with the EFSA (European Food Safety Authority) opinion on pathogenic STEC published in 2013.
Introduction
Shiga toxin-producing Escherichia coli (STEC) are important zoonotic pathogens comprising more than 400 serotypes (Beutin and Fach, 2014). A fraction of these serotypes are able to cause bloody diarrhea and may progress to hemolytic uremic syndrome (HUS). This subset of STECs is termed Enterohemorrhagic E. coli (EHEC) (Beutin and Fach, 2014). STEC O157:H7 has been the first pathogenic E. coli whose presence in foodstuffs was regulated. Today, non-O157 STEC infections have increased greatly, sometimes accounting for up to 70% of notified STEC infections (Brooks et al., 2005; Johnson et al., 2006; Gould et al., 2013; EFSA, 2014). Consequently, regulations in the US and in the EU have evolved to include some non-O157 STEC serogroups together with serotype O157:H7.
Successful implementation of these regulations in the food industry across the world requires effective detection methods that are both specific and sensitive. Detection of non-O157 STEC in foods is particularly challenging because these bacteria lack phenotypic characteristics that distinguish them invariably from the large number of non-STEC flora that share the same habitat. Additionally, they may be present in very low numbers, with a heterogeneous distribution in the food matrices. The environment of the food matrix may also trigger stress responses from the STEC strains and induce latent physiological states that further complicate detection (Wang et al., 2013).
In the absence of a clear definition of virulent STEC strains, the ISO/CEN 13136:2012 Technical Specification (ISO, 2012) and US MLG5B.05 (USDA-FSIS, 2014) use a stepwise approach, comprising an initial screening step for virulence genes (Shiga toxin genes, stx, and intimin gene, eae), followed by testing of O-serogroup specific gene markers. Because the stx and eae genes can be independently present in a number of non-pathogenic strains of E. coli and other Enterobacteriaceae, the first screening step generates numerous signals from samples that do not necessarily contain a true EHEC strain. These stx/eae positive enrichments must then be subjected to a second screening targeting the O-group gene markers. As testing is completed on enrichment broths that contain a mixture of different cells, the different target gene signals (i.e., stx, eae, and the O-group markers) may arise from different individual non-pathogenic strains. Lastly it is necessary to perform an isolation step to confirm the presence of the different markers in a single isolate by PCR.
We have previously shown that combining the detection of espK with either espV, ureD, or Z2098 is a highly sensitive and specific approach for identifying the top seven clinically important EHEC serotypes in industrialized countries (Delannoy et al., 2013a). These markers were shown to be preferentially associated with E. coli strains carrying stx and eae genes, known as typical EHEC, and could be used in conjunction with stx/eae screening to better identify samples that may be more likely to contain a true EHEC.
Recently, a new clone of STEC O26:H11 harboring stx2a and strongly associated with HUS has emerged (Bielaszewska et al., 2013). Characterization of O26:H11 stx2 circulating in France (Delannoy et al., 2015a,b) demonstrated that some stx2a or stx2d positive strains do not have any of the espK, espV, ureD, or Z2098 markers. Hence, such clones would evade a first detection step solely based on stx/eae and a combination of those genetic markers. Therefore, we identified a CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) sequence specific for this O26:H11 EHEC French clone (Delannoy et al., 2015a). Detection of this CRISPR sequence would advantageously be combined with stx and eae in the first screening step to identify this new O26:H11 clone. Furthermore, we have previously developed CRISPR-based real-time PCR assays able to detect the top seven EHEC serotypes and the German O104:H4 STEC clone responsible for a very large European STEC outbreak in 2011 (Delannoy et al., 2012a,b; Miko et al., 2013). These CRISPR PCR assays proved highly sensitive and specific when tested on a large collection of E. coli strains comprising various E. coli pathogroups (Delannoy et al., 2012a,b). Such CRISPR markers may substitute O group testing in a more targeted second step.
Different approaches have been tested to refine the detection systems for EHEC. Some involve detection of ecf1, a plasmid gene highly associated with E. coli strains that are positive for stx, eae, and ehxA (Livezey et al., 2015); while others involve detection of specific eae-subtypes (Bugarel et al., 2010). The top seven EHEC serotypes are exclusively associated with certain eae-subtypes (Oswald et al., 2000; Bugarel et al., 2010; Madic et al., 2010). Indeed, O157:H7 and O145:H28 serotypes are associated with the eae-gamma subtype; O26:H11 are associated with the eae-beta subtype; O103:H2, O121:H19, and O45:H2 are associated with the eae-epsilon subtype; and O111:H8 are associated with the eae-theta subtype. Real-time PCR targeting these eae-subtypes were previously developed and tested in raw milk cheese (Madic et al., 2011) and cattle feces (Bibbal et al., 2014). These could also be used as targets in a more targeted second step.
We attempted to develop an alternative real-time PCR-based approach to improve the detection of the clinically important EHEC by reducing the number of potential positive samples that require further confirmation of the O-antigen markers. We also aimed at reducing the number of samples for which isolation is attempted, as the isolation step is laborious, time-consuming, and not always successful (Wang et al., 2013). The objective of this project was to evaluate the discriminatory power of these various genetic markers to predict the presence of the top seven EHEC serogroups compared to that of the strategy proposed by the ISO/CEN TS13136:2012 (ISO, 2012) and MLG5B.05 (USDA-FSIS, 2014).
Materials and Methods
E. coli Control Strains
E. coli control strains used for this study (Table 1) comprised a panel of ATCC E. coli strains (n = 42) and E. coli reference strains derived from the BfR and Anses collections (n = 13). The origin and characteristics of the E. coli strains from BfR and Anses have been previously described (Delannoy et al., 2012a, 2013a). Cultivation of bacteria and preparation of DNA was performed as previously described (Delannoy et al., 2012a, 2013a).

Table 1. PCR-screening of the genetic markers in STEC isolates used as reference.
Beef Samples, Enrichment, and DNA Extraction
A set of 1739 beef samples composed of ground beef and carcasses were collected from routine screening using the GeneDisc array (Pall GeneDisc, Bruz, France) at the Veterinary Departmental Laboratory of Touraine, France during a 1-year period as well as in meat production plants. For this study, sampling was biased to get greater numbers of DNA samples positive for stx alone (n = 306), positive for stx and eae (n = 180), positive for eae alone (n = 200), and negative for both stx and eae (n = 1053). This sampling scheme does not represent the prevalence of STEC and EHEC in French beef. All samples were incubated in buffered peptone water (BioMerieux, Marcy l'étolie, France) for 18–24 h at 37°C. After enrichment, DNA was extracted from 1 ml of enriched sample using the InstaGene matrix (Bio-Rad Laboratories, Marnes-La Coquette, France) following manufacturer's instruction and DNA was stored at −20°C until use. When samples were found positive for stx, eae, and rfbEO157, isolation of strains was attempted by local laboratories for confirmation of EHEC O157:H7. Following the recommendation of the French ministry of agriculture the appropriate sanitary measures were taken in positive cases of EHEC O157:H7. Unfortunately, because this study was performed several months after the samples were collected, the original samples were not conserved to attempt isolation from presumptive positives with the alternate methods described in this study.
High-Throughput Real-Time PCR
A LightCycler® 1536 (Roche, Meylan, France) was used to perform high throughput real-time PCR amplifications as described previously (Delannoy et al., 2012a), except that 1 μl of sample DNA was used in each reaction for a final reaction volume of 2 μl. The thermal profile was modified as follows: 95°C for 1 min, followed by 45 cycles of 95°C for 0 s, and 58°C for 30 s. All ramp rates were set to 2°C/s. E. coli gene targets used for the real-time PCR amplification and all primers and probes that have previously been described are reported in Table 2. An inhibition control (IC) was performed on each sample to check for potential inhibition of the PCR reaction due to intrinsic characteristics of the sample. The IC is a recombinant pBluescript IISK+ plasmid containing the dsb gene from Ehrlichia canis (Michelet et al., 2014). The plasmid was added to each sample at a concentration of approximately 0.3 pg/μl. Primers and probe specific for the E. canis dsb gene were used to detect the IC (Michelet et al., 2014).

Table 2. Primer and probe sequences used in this study.
Results
Presence of stx1, stx2, eae, espK, espV, Z2098, and ureD in E. coli Strains
The presence of stx1, stx2, eae, espK, espV, Z2098, and ureD was tested by PCR in a panel of E. coli strains obtained from culture collections (Table 1). All E. coli strains used in this study were positive for the stx and eae genes and therefore can be considered as typical EHEC strains. Strains were associated with the following eae variants: eae-beta (O26:H11, O26:HND, O103:H11, O5, O118:H16, O123:H11), eae-gamma (O145:H28, O145:HND, O157:H7, O55:H7), eae-epsilon (O121:H19, O45:H2, O103:H2, O103:H11), and eae-theta (O111:H8, O111:HND, O103:H25). The eae subtype of strain CB10528 (O172:H25) could not be determined. Distribution of the genetic markers espK, espV, Z2098, and ureD in the 55 EHEC strains is shown in Table 1. Overall, the genetic markers investigated were detected in most of the EHEC strains examined. With the exception of the new O26:H11 stx2-positive strain (CB14699) and one strain of serotype O157:H7 (CDC-C984), all of the strains were positive for espK. The espV gene was not detected in two E. coli O26:H11 strains (ATCC2196 and CB14699), in O45:H2, O118:H16, O5, or in O123:H11 isolates. The Z2098 gene marker tested negative in only a few strains: the new O26:H11 stx2 positive clone (strain CB14699) and in the O55:H7 strains. The ureD gene was absent in the new O26:H11 stx2 positive clone (strain CB14699) and in the O55:H7 strains. It was also absent from a few O103:H2 isolates (strains PMK5, CB12062, CB12092). In summary, all of the strains were positive for one or more of the genes espK, espV, Z2098, and ureD, with the exception of the new O26:H11 stx2 positive clone (CB14699).
Screening Beef Enrichments for stx1, stx2, eae, espK, espV, Z2098, and ureD
A set of 1739 beef samples was screened for the presence of stx1, stx2, eae, espK, espV, Z2098, and ureD. The stx genes were detected in 27.95% of the samples (486/1739). The eae gene was detected in 21.85% of the samples (380/1739). The two genes were simultaneously present in 10.35% of the samples (180/1739). The genes espK and/or Z2098 were detected in 7.42% of the samples (129/1739) (Figure 1A), while espK and/or espV were found in 130 samples (7.48%) (Figure 1B) and espK and/or ureD was recorded in 145 samples (8.34%) (Figure 1C).
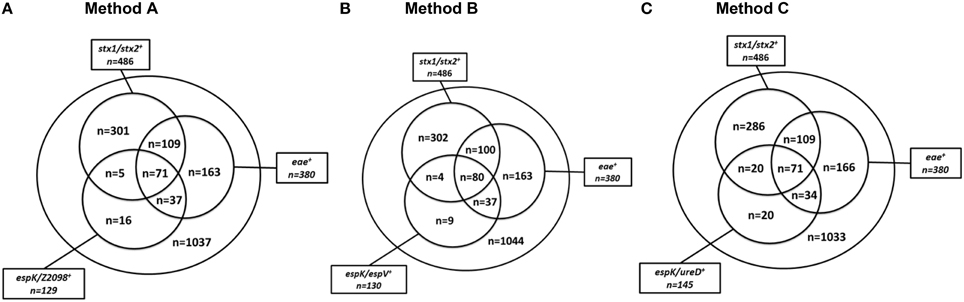
Figure 1. Distribution of the genetic markers stx, eae, espK, espV, and Z2098 among 1739 beef samples. (A) (stx1/stx2, eae, and espK/Z2098) is Method A, (B) (stx1/stx2, eae, and espK/espV) is Method B, (C) (stx1/stx2, eae, and espK/ureD) is Method C. stx1/stx2+ for samples giving a positive result for stx1 and/or stx2, eae+ for samples giving a positive result for eae, espK/Z2098+ for samples giving a positive result for espK and/or Z2098, espK/espV+ for samples giving a positive result for espK and/or espV, espK/ureD+ for samples giving a positive result for espK and/or ureD.
By using the stx and eae genes for screening beef samples, following the ISO/CEN TS13136 and MLG5B.05 methods, 180 samples (10.35%) were recorded as stx/eae positive and should, therefore, be subjected to a second screening step for EHEC-serogroups. Pre-screening of stx/eae positive samples for espK with either Z2098 (alternate method A), or ureD (alternate method C), provided a 60% reduction of the number of samples that should be submitted to a second screening targeting the O-group gene markers (n = 71). Using the alternate method B (stx/eae/espK/espV), 80 of the 1739 samples (4.6%) needed to be submitted to a further screening for serogroup determination, which represents a reduction by 55% of the number of samples subjected to a second screening.
Figure 2 shows the comparison of the alternative methods A–C with the 180 beef samples that tested positive for both the stx and eae genes. A total of 90 stx and eae positive beef samples tested negative for espK, espV, Z2098, and ureD (sector 8, Figure 2). However, the inclusion of the CRISPRO26:H11 PCR revealed 12 espK, espV, Z2098, and ureD negative samples that were positive for both the new CRISPRO26:H11 clone and eae-beta, the variant of the intimin gene carried by EHEC O26:H11 (see below).
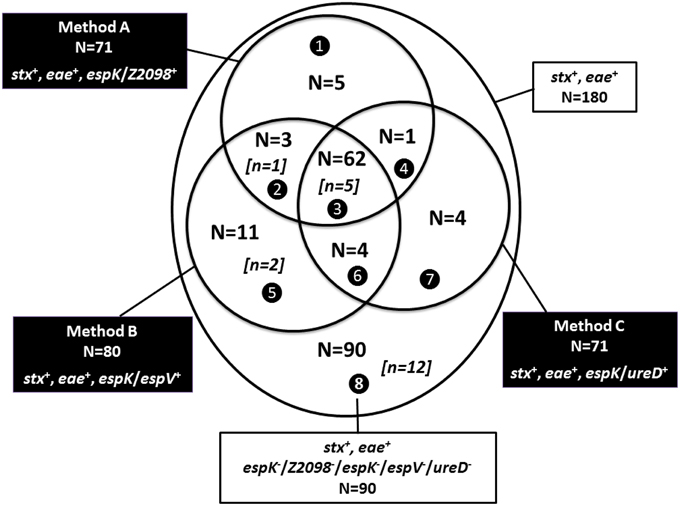
Figure 2. Comparison of Methods A–C on 180 beef samples that tested positive for both stx and eae genes. stx+ for samples giving a positive result for stx1 and/or stx2; eae+ for samples giving a positive result for eae; espK/Z2098+ for samples giving a positive result for espK and/or Z2098; espK/espV+ for samples giving a positive result for espK and/or espV; espK/ureD+ for samples giving a positive result for espK and/or ureD. Sector ❶ (stx+, eae+, Z2098+), sector ❷ (stx+, eae+, Z2098+, espV+), sector ❸ (stx+, eae+, espK/Z2098+, espK/espV+, espK/ureD+), sector ❹ (stx+, eae+, Z2098+, ureD+), sector ❺ (stx+, eae+, espV+), sector ❻ (stx+, eae+, espV+, ureD+), sector ❼ (stx+, eae+, ureD+) and sector ❽ (stx+, eae+). [n = x] is the number of samples that tested positive by the CRISPRO26:H11 PCR assay detecting the new O26 clone, N = x is the total number of samples per sector.
Screening Beef Samples for stx1, stx2, eae, and CRISPRO26:H11
The 180 beef samples that are positive for stx and eae were also tested by the CRISPRO26:H11 PCR test (SP_O26-E, as described in Delannoy et al., 2015a), that detects the new EHEC O26:H11 French clone (stx2 and eae positive, espK, espV, Z2098, ureD negative). Among the 180 samples tested, 20 stx/eae positive samples were also found to be positive for SP_O26-E and should therefore be submitted to a further screening for serogroup determination. Interestingly, most of them (16/20) were found positive for stx2, 2 samples were positive for stx1 only and 2 had an unknown stx subtype. Twelve of these 20 were also negative for espK, espV, Z2098, and ureD (Figure 2, sector 8). Finally, when combining the use of CRISPRO26:H11 (SP_O26-E PCR test) with alternate method A, 85 samples should be submitted to a second screening targeting the O-group gene markers. When using alternate methods B and C with the CRISPRO26:H11 PCR test, 92 and 86 samples should be submitted to the second step, respectively.
Screening Beef Samples for stx1, stx2, eae, and the Top Seven EHEC Serogroups
As recommended in the ISO/TS 13136 (EU) and MLG5B.05 (US) reference methods, the 180 stx and eae positive samples were tested for the top seven EHEC serogroups. Among these, 115 samples were positive for at least one of the top seven US regulated EHEC serogroups and 99 were positive for at least one of the top five EHEC serogroups screened by the European ISO/TS 13136 method (data not shown). The most frequently found serogroup was O103 (n = 71), followed by O26 (n = 45), O121 (n = 38), O157 (n = 18), O45 (n = 14), O145 (n = 6), and O111 (n = 1). Interestingly, 30 samples were positive for 2 serogroups, 11 were positive for 3 serogroups, and 7 for more than 4 serogroups. In final, 41.74% (48/115) of the beef samples tested positive for more than one O-group marker.
Screening Beef Samples for stx1, stx2, eae, and the eae Subtypes
The 180 stx and eae positive beef samples were further tested for the eae subtypes gamma, beta, epsilon and theta, which are associated with one or more of the top seven EHEC serogroups. Among these 180 stx and eae positive samples, 135 tested positive for at least one of the four eae-subtypes: gamma, beta, epsilon and theta (data not shown). The most frequently detected eae-subtypes were eae-beta (n = 94) and eae-theta (n = 65), followed by eae-epsilon (n = 15), and eae-gamma (n = 5). Trying to correlate the eae subtype with the serogroup, we identified 51 beef samples for which at least one serogroup was associated with the corresponding related eae-subtype. Among these 51 samples, 6 were positive with multiple serogroups.
Comparison of Alternative Methods A–C for Screening Beef Samples
We identified 62 samples that were recorded positive with the three alternative methods A–C (sector 3, Figure 2) and therefore must be submitted to a second screening targeting the O-group gene markers. These samples are strongly suspected to contain typical EHEC and five of these are also suspected to contain the new CRISPRO26:H11 clone. In addition we found 28 samples (sectors 1, 2, 4, 5, 6, 7 from Figure 2) that were positive by one or two alternative methods only. PCR results obtained for these 28 samples for eae subtypes and top seven O-groups were as follows: only one sample among the 5 samples of sector 1 (Figure 2) was positive for the association of O26 and eae-beta, but it tested negative for the different CRISPRO26:H11 assays targeting EHEC O26:H11 [SP_O26-C, SP_O26-D, and SP_O26-E, as described in Delannoy et al., 2012a, 2015a (data not shown)]. The only sample from sector 4 (Figure 2) and each of the four samples from sectors 6 and 7 (Figure 2) were found negative by the association of the top seven serogroups and the corresponding eae-subtypes. From sector 2 (Figure 2), only one out of three samples was found positive for the new CRISPRO26:H11 clone (this sample tested positive by PCR for O26, eae-beta, SP_O26-E; and was also stx2 positive which is consistent with the new clone). The two other samples were not suspected as “presumptive positive” based on the eae-subtypes and top seven EHEC serotypes determination. In sector 5 (Figure 2), out of eleven samples three were found “presumptive positive” for EHEC O26 (among them two were suspected to be positive for the new O26:H11 clone). Finally, “presumptive positive” samples were recorded in sectors 2, 3, and 5 which lead to consider the alternate method B as the best one among the other alternate methods for screening EHEC strains. In order to complete the screening of EHEC and not to exclude the new virulent O26 clone, the alternate method B should include the screening of the new CRISPRO26:H11 assay.
Discussion
A STEC seropathotype classification has been based upon the serotype association with human epidemics, bloody diarrhea, and HUS, and has been developed as a tool to assess the clinical and public health risks associated with non-O157 EHEC and STEC strains (Karmali et al., 2003). This approach has been of considerable value in defining pathogenic STEC serotypes of importance in cases of human infection (EFSA, 2007; Coombes et al., 2011); however it does not resolve the underlying problem with strains that have not yet been fully serotyped. Furthermore, classification based solely on the presence of seropathotype is inadequate with illnesses linked to STEC serotypes other than O157:H7 that are on the rise worldwide, indicating that some of these organisms may be emerging pathogens. In 2013, the Panel on Biological Hazards (BIOHAZ) of the European Food Safety Authority (EFSA) published a Scientific Opinion on “VTEC-seropathotype and scientific criteria regarding pathogenicity assessment” (EFSA, 2013). This document has focused attention on the applicability of the Karmali seropathotype concept. The document does not provide a scientific definition of a pathogenic STEC but states that the seropathotype classification of Karmali et al. (2003) does not define pathogenic STEC nor does it provide an exhaustive list of pathogenic serotypes. It is not possible to fully define human pathogenic STEC or identify factors for STEC that absolutely predict the potential to cause human disease, but strains positive for Shiga-toxin (in particular the stx2 genes) and eae (intimin production) genes are associated with a higher risk of more severe illness than other virulence factor combinations (EFSA, 2013). Severe disease, and particularly HUS, is linked to certain serotypes and strains and this link must be the result of particular genetic factors or combinations of factors that have to be determined. A new molecular classification scheme has been proposed by the EFSA Panel on Biological Hazards (BIOHAZ) that relies more on virulence factors than seropathotypes. It is proposed that STEC serogroups O157, O26, O103, O145, O111, and O104 in combination with stx and eae or stx and both aaiC (secreted protein of EAEC) and aggR (plasmid-encoded regulator) genes should be considered as presenting a potentially higher risk for bloody diarrhea and HUS, such strains are categorized in group I (EFSA, 2013). For any other serogroups in combination with the same genes, the potential risk is regarded as high for diarrhea, but currently unknown for HUS, such strains are categorized in group II. The inclusion of aaiC and aggR genes in the proposed molecular approach is due to the O104:H4 outbreak, which was caused by a highly virulent strain (Frank et al., 2011). This appears to be an exceptional event (Prager et al., 2014) and future surveillance will provide data that may be used to review the inclusion of these virulence factors but a recent study showed that French cattle are not a reservoir of the highly virulent enteroaggregative Shiga toxin-producing E. coli of serotype O104:H4 (Auvray et al., 2012).
Following the EFSA opinion, several laboratories have attempted to develop detection and identification methods for strains of groups I and II, and although substantial progress has been made, a practical method of pathogenic STEC detection has yet to be validated. Molecular methods for screening EHEC O157 and non-O157 in beef products rely currently on the molecular detection of stx, eae, and the top five or top seven EHEC serogroups in mixed bacterial enrichments as described in the ISO/TS 13136 (EU) and MLG5B.05 (US) reference methods (ISO, 2012; USDA-FSIS, 2014), followed by attempted isolation of the correct strain. These approaches bear the disadvantage that food samples that carry mixtures of stx-negative E. coli carrying eae genes together with eae-negative STEC falsely indicate the presence of EHEC when analyzed for the genes mentioned above (Beutin et al., 2009; Fratamico and Bagi, 2012; Wasilenko et al., 2014). Hence, screening food enrichment broths for stx and eae genes may cause needless disruption and costs for food producers when the risk of low-virulence STEC is overestimated. The erroneous identification of cucumbers as the source in the outbreak in Germany in 2011 cost European fruit and vegetable producers approximately € 812 million (Commission of the European Communities., 2011). Thus, new DNA targets that unambiguously identify typical EHEC strains (stx-positive and eae-positive E. coli strains) in complex samples are a desirable goal. This new panel of genes might include novel genetic markers recently identified as highly associated to pathogenic STEC strains (Delannoy et al., 2013a,b). Thus, the associations of espK with either espV, ureD, or Z2098 were found to be the best combinations for more specific and sensitive detection of the top seven EHEC strains, allowing detection of 99.3–100% of these strains. In addition, detection of 93.7% of the typical EHEC strains belonging to other serotypes than the top seven offered the possibility for identifying new emerging typical EHEC strains (Delannoy et al., 2013a,b). Conversely, these different combinations of genetic markers were very rarely associated with STEC (1.6–3.6%) and with non-pathogenic E. coli (1.1–3.4%). The objective of the present study was to refine the EHEC screening systems for testing beef samples via the incorporation of these additional gene targets espK, espV, ureD, and Z2098 in the detection scheme. In addition to these four targets, we included a CRISPRO26:H11 target in the detection scheme that has been designed for detecting a new clone of STEC O26:H11 harboring stx2 only and strongly associated with HUS in France (Delannoy et al., 2015a,b). This new EHEC O26:H11 French clone is positive for eae (eae-beta subtype) but does not contain any of the above markers espK, espV, ureD, or Z2098. The fact that the new French clone is missing espK, espV, ureD, and Z2098 does not mean that these genes are not required EHEC virulence factors. We have previously demonstrated that they are significantly associated with EHEC strains and not the other pathotypes. It does suggest however that the new French clone may harbor a different set of virulence genes (just like atypical EHEC lack eae), which can be investigated through additional genomic studies. In the meantime, although CRISPR sequences are not virulence factors per se, we have demonstrated that certain specific spacers are associated with EHEC strains from the top7 serotypes and thus can provide specific and sensitive detection of the top 7 EHEC strains (Delannoy et al., 2012a,b).
To validate the pertinence of this new approach we screened 1739 beef samples and collected 180 samples that tested positive for both stx and eae, and must be subjected to a second screening step for serogroup determination according to the ISO/TS 13136 (EU) and MLG5B.05 (US) reference methods (ISO, 2012; USDA-FSIS, 2014). Among these 180 samples, 135 tested positive for at least one of the four eae-subtypes gamma, beta, epsilon and theta, which are related to typical EHEC and in particular to those of the top seven serogroups (Bugarel et al., 2010). Introduction of the eae-subtypes in the screening step provided a reduction by 25% of the number of stx/eae positive samples that should be subjected to a further screening for serogroup determination. A more significant refinement of the first EHEC screening step was achieved by including espK, espV, ureD, Z2098, and CRISPRO26:H11 target genes in the detection scheme. Thus, a reduction by 52.78–52.22% of the number of samples subjected to a further screening for serogroup determination was obtained by using respectively the alternate method A (stx/eae/espK/Z2098) or method C (stx/eae/espK/ureD) in combination with the CRISPRO26:H11 PCR assay detecting the new O26 clone. A reduction by 48.88% of the number of “presumptive positive” samples was obtained using the alternate method B (stx/eae/espK/espV) in association with the CRISPRO26:H11 PCR assay. Given the additional information on the association of the top seven serogroups and the eae-subtypes, we determined the last approach, i.e., method B (stx/eae/espK/espV) in association with the CRISPRO26:H11 PCR assay, as the best approach to narrow down the EHEC screening step in beef samples. Using such an approach, 92 samples must be subjected to a further screening for serogroup determination vs. 180 with the conventional stx/eae approach used in the ISO/TS 13136 (EU) and MLG5B.05 (US) reference methods (ISO, 2012; USDA-FSIS, 2014). This constitutes a significant reduction (almost 50%) of the number of samples subjected to a second screening targeting the O-group gene markers. Moreover, this approach is in line with the EFSA opinion that has identified STEC strains of groups I and II as presenting the potential higher risk for diarrhea and HUS (EFSA, 2013). Identification of additional gene markers, i.e., espK, espV, and CRISPRO26:H11 to better distinguish typical EHEC from other E. coli pathogroups would substantially enhance the power of EHEC test systems providing a significant reduction of “presumptive positive” in beef samples. Such a new approach would provide to the agroindustry a novel method for tracking EHEC in food samples. This work should be considered with interest to draw up the outline of a future standard that will follow the recommendations of EFSA.
Author Contributions
Conceived and designed the experiments: SD, PF. Performed the experiments: SD, BC, SI, HW, JD, IB. Analyzed the data: SD, BC, PF. Contributed reagents/materials/analysis tools: SD, LB, JD, IB, PF. Wrote the paper: SD, PF. Critical revision of the paper for important intellectual content: SD, BC, SI, HW, LB, JD, IB, PF.
Funding
The project was partially financed by the French “joint ministerial program of R&D against CBRNE risks” (Grant number C17609-2).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Auvray, F., Dilasser, F., Bibbal, D., Kérourédan, M., Oswald, E., and Brugère, H. (2012). French cattle is not a reservoir of the highly virulent enteroaggregative Shiga toxin-producing Escherichia coli of serotype O104:H4. Vet Microbiol. 158, 443–445. doi: 10.1016/j.vetmic.2012.02.029
Beutin, L, Fach, P. (2014). Detection of shiga toxin-producing Escherichia coli from nonhuman sources and strain typing. Microbiol. Spectr. 2:EHEC-0001-2013. doi: 10.1128/microbiolspec.EHEC-0001-2013
Beutin, L., Jahn, S., and Fach, P. (2009). Evaluation of the ‘GeneDisc’ real-time PCR system for detection of enterohaemorrhagic Escherichia coli (EHEC) O26, O103, O111, O145 and O157 strains according to their virulence markers and their O- and H-antigen-associated genes. J. Appl. Microbiol. 106, 1122–1132. doi: 10.1111/j.1365-2672.2008.04076.x
Bibbal, D., Loukiadis, E., Kérourédan, M., Peytavin de Garam, C., Ferré, F., Cartier, P., et al. (2014). Intimin gene (eae) subtype-based real-time PCR strategy for specific detection of Shiga toxin-producing Escherichia coli serotypes O157:H7, O26:H11, O103:H2, O111:H8, and O145:H28 in cattle feces. Appl. Environ. Microbiol. 80, 1177–1184. doi: 10.1128/AEM.03161-13
Bielaszewska, M., Mellmann, A., Bletz, S., Zhang, W., Köck, R., Kossow, A., et al. (2013). Enterohemorrhagic Escherichia coli O26:H11/H-: a new virulent clone emerges in Europe. Clin. Infect. Dis. 56, 1373–1381. doi: 10.1093/cid/cit055
Brooks, J. T., Sowers, E. G., Wells, J. G., Greene, K. D., Griffin, P. M., Hoekstra, R. M., et al. (2005). Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983-2002. J. Infect. Dis. 192, 1422–1429. doi: 10.1086/466536
Bugarel, M., Beutin, L., and Fach, P. (2010). Low-density macroarray targeting non-locus of enterocyte effacement effectors (nle genes) and major virulence factors of Shiga toxin-producing Escherichia coli (STEC): a new approach for molecular risk assessment of STEC isolates. Appl. Environ. Microbiol. 76, 203–211. doi: 10.1128/AEM.01921-09
Commission of the European Communities (2011). Commission Staff Working Document on Lessons Learned from the 2011 Outbreak of Shiga Toxin-Producing Escherichia coli (STEC) O104:H4 in Sprouted Seeds. SANCO/13004/2011, 1–23. Available online at http://ec.europa.eu/food/food/biosafety/salmonella/docs/cswd_lessons_learned_en.pdf (Accessed on August 8, 2015).
Coombes, B. K., Gilmour, M. W., and Goodman, C. D. (2011). The evolution of virulence in non-o157 shiga toxin-producing Escherichia coli. Front. Microbiol. 2:90. doi: 10.3389/fmicb.2011.00090
Delannoy, S., Beutin, L., Burgos, Y., and Fach, P. (2012b). Specific detection of enteroaggregative hemorrhagic Escherichia coli O104:H4 strains by use of the CRISPR locus as a target for a diagnostic real-time PCR. J. Clin. Microbiol. 50, 3485–3492. doi: 10.1128/JCM.01656-12
Delannoy, S., Beutin, L., and Fach, P. (2012a). Use of clustered regularly interspaced short palindromic repeat sequence polymorphisms for specific detection of enterohemorrhagic Escherichia coli strains of serotypes O26:H11, O45:H2, O103:H2, O111:H8, O121:H19, O145:H28, and O157:H7 by real-time PCR. J. Clin. Microbiol. 50, 4035–4040. doi: 10.1128/JCM.02097-12
Delannoy, S., Beutin, L., and Fach, P. (2013a). Discrimination of enterohemorrhagic Escherichia coli (EHEC) from non-EHEC strains based on detection of various combinations of type III effector genes. J. Clin. Microbiol. 51, 3257–3262. doi: 10.1128/JCM.01471-13
Delannoy, S., Beutin, L., and Fach, P. (2013b). Towards a molecular definition of enterohemorrhagic Escherichia coli (EHEC): detection of genes located on O island 57 as markers to distinguish EHEC from closely related enteropathogenic E. coli strains. J. Clin. Microbiol. 51, 1083–1088. doi: 10.1128/JCM.02864-12
Delannoy, S., Mariani-Kurkdjian, P., Bonacorsi, S., Liguori, S., and Fach, P. (2015a). Characteristics of emerging human-pathogenic Escherichia coli O26:H11 strains isolated in France between 2010 and 2013 and carrying the stx2d gene only. J. Clin. Microbiol. 53, 486–492. doi: 10.1128/JCM.02290-14
Delannoy, S., Mariani-Kurkdjian, P., Bonacorsi, S., Liguori, S., Ison, S. A., and Fach, P. (2015b). Draft genome sequences of human-pathogenic Escherichia coli O26:H11 strains carrying the stx2 gene only and circulating in France. Genome Announc. 3:e00852-15. doi: 10.1128/genomeA.00852-15
EFSA (European Food Safety Authority) (2007). Scientific Opinion of the Panel on Biological Hazards (BIOHAZ) on monitoring of verotoxigenic Escherichia coli (VTEC) and identification of human pathogenic VTEC types. EFSA J. 579, 1–61. doi: 10.2903/j.efsa.2007.579
EFSA (European Food Safety Authority) (2013). Scientific Opinion of the Panel on Biological Hazards (BIOHAZ) on VTEC-seropathotype and scientific criteria regarding pathogenicity assessment. EFSA J. 11, 3138, 1–106. doi: 10.2903/j.efsa.2013.3138
EFSA (European Food Safety Authority, European Centre for Disease Prevention Control). (2014). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2012. EFSA J. 12, 3547, 312. doi: 10.2903/j.efsa.2014.3547
Frank, C., Werber, D., Cramer, J. P., Askar, M., Faber, M., an der Heiden, M., et al. (2011). Epidemic profile of Shiga-toxin-producing Escherichia coli O104:H4 outbreak in Germany. N. Engl. J. Med. 365, 1771–1780. doi: 10.1056/NEJMoa1106483
Fratamico, P. M., and Bagi, L. K. (2012). Detection of Shiga toxin-producing Escherichia coli in ground beef using the GeneDisc real-time PCR system. Front. Cell Infect. Microbiol. 2:152. doi: 10.3389/fcimb.2012.00152
Fratamico, P. M., DebRoy, C., Miyamoto, T., and Liu, Y. (2009). PCR detection of enterohemorrhagic Escherichia coli O145 in food by targeting genes in the E. coli O145 O-antigen gene cluster and the shiga toxin 1 and shiga toxin 2 genes. Foodborne Pathog Dis. 6, 605–611. doi: 10.1089/fpd.2008.0254
Gould, L. H., Mody, R. K., Ong, K. L., Clogher, P., Cronquist, A. B., Garman, K. N., et al. (2013). Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000–2010: epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 10, 453–460. doi: 10.1089/fpd.2012.1401
ISO (International Organization for Standardization) (2012). ISO/TS 13136:2012, Microbiology of Food and Animal Feed—Real-Time Polymerase Chain Reaction (PCR) –Based Method for the Detection of Food-Borne Pathogens—Horizontal Method for the Detection of Shiga Toxin-Producing Escherichia coli (STEC) and the Determination of O157, O111, O26, O103, and O145 Serogroups. Available online at: http://www.iso.org/iso/iso_catalogue/catalogue_tc/catalogue_detail.htm?cs number=53328. (Accessed on August 8, 2015).
Johnson, K. E., Thorpe, C. M., and Sears, C. L. (2006). The emerging clinical importance of non-O157 Shiga toxin-producing Escherichia coli. Clin. Infect. Dis. 43, 1587–1595. doi: 10.1086/509573
Karmali, M. A., Mascarenhas, M., Shen, S., Ziebell, K., Johnson, S., Reid-Smith, R., et al. (2003). Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 41, 4930–4940. doi: 10.1128/JCM.41.11.4930-4940.2003
Livezey, K. W, Groschel, B., and Becker, M. M. (2015). Use of the ecf1 gene to detect Shiga toxin-producing Escherichia coli in beef samples. J. Food Prot. 78, 675–684. doi: 10.4315/0362-028X.JFP-14-417
Madic, J., Peytavin de Garam, C., Vingadassalon, N., Oswald, E., Fach, P., Jamet, E., et al. (2010). Simplex and multiplex real-time PCR assays for the detection of flagellar (H-antigen) fliC alleles and intimin (eae) variants associated with enterohaemorrhagic Escherichia coli (EHEC) serotypes O26:H11, O103:H2, O111:H8, O145:H28 and O157:H7. J. Appl. Microbiol. 109, 1696–1705. doi: 10.1111/j.1365-2672.2010.04798.x
Madic, J., Vingadassalon, N., Peytavin de Garam, C. P., Marault, M., Scheutz, F., Brugère, H., et al. (2011). Detection of Shiga toxin-producing Escherichia coli serotypes O26:H11, O103:H2, O111:H8, O145:H28, and O157:H7 in raw-milk cheeses by using multiplex real-time PCR. Appl. Environ. Microbiol. 77, 2035–2041. doi: 10.1128/AEM.02089-10
Michelet, L., Delannoy, S., Devillers, E., Umhang, G., Aspan, A., Juremalm, M., et al. (2014). High-throughput screening of tick-borne pathogens in Europe. Front. Cell Infect. Microbiol. 4:103. doi: 10.3389/fcimb.2014.00103
Miko, A., Delannoy, S., Fach, P., Strockbine, N. A., Lindstedt, B. A., Mariani-Kurkdjian, P., et al. (2013). Genotypes and virulence characteristics of Shiga toxin-producing Escherichia coli O104 strains from different origins and sources. Int. J. Med. Microbiol. 303, 410–421. doi: 10.1016/j.ijmm.2013.05.006
Nielsen, E. M., and Andersen, M. T. (2003). Detection and characterization of verocytotoxin-producing Escherichia coli by automated 5′ nuclease PCR assay. J. Clin. Microbiol. 41, 2884–2893. doi: 10.1128/JCM.41.7.2884-2893.2003
Oswald, E., Schmidt, H., Morabito, S., Karch, H., Marchès, O., and Caprioli, A. (2000). Typing of intimin genes in human and animal enterohemorrhagic and enteropathogenic Escherichia coli: characterization of a new intimin variant. Infect. Immun. 68, 64–71. doi: 10.1128/IAI.68.1.64-71.2000
Perelle, S., Dilasser, F., Grout, J., and Fach, P. (2004). Detection by 5′-nuclease PCR of Shiga-toxin producing Escherichia coli O26, O55, O91, O103, O111, O113, O145 and O157:H7, associated with the world's most frequent clinical cases. Mol. Cell Probes. 18, 185–192. doi: 10.1016/j.mcp.2003.12.004
Perelle, S., Dilasser, F., Grout, J., and Fach, P. (2005). Detection of Escherichia coli serogroup O103 by real-time polymerase chain reaction. J. Appl. Microbiol. 98, 1162–1168. doi: 10.1111/j.1365-2672.2005.02545.x
Prager, R., Lang, C., Aurass, P., Fruth, A., Tietze, E., and Flieger, A. (2014). Two novel EHEC/EAEC hybrid strains isolated from human infections. PLoS ONE 9:e95379. doi: 10.1371/journal.pone.0095379
USDA-FSIS (2014). Detection and Isolation of non-O157 Shiga Toxin-Producing Escherichia coli (STEC) from Meat Products and Carcass and Environmental Sponges. In Microbiological Laboratory Guidelines chapter 5B.05. Available online at http://www.fsis.usda.gov/wps/wcm/connect/7ffc02b5-3d33-4a79-b50c-81f208893204/MLG-5B.pdf?MOD=AJPERES (Accessed on December 10, 15).
Wang, F., Yang, Q., Kase, J. A., Meng, J., Clotilde, L. M., Lin, A., et al. (2013). Current trends in detecting non-O157 Shiga toxin-producing Escherichia coli in food. Foodborne Pathog. Dis. 10, 665–677. doi: 10.1089/fpd.2012.1448
Wasilenko, J. L., Fratamico, P. M., Sommers, C., DeMarco, D. R., Varkey, S., Rhoden, K., et al. (2014). Detection of Shiga toxin-producing Escherichia coli (STEC) O157:H7, O26, O45, O103, O111, O121, and O145, and Salmonella in retail raw ground beef using the DuPont™ BAX® system. Front. Cell Infect. Microbiol. 4:81. doi: 10.3389/fcimb.2014.00081
Keywords: STEC, O157, non-O157, espK, espV, ureD, Z2098, CRISPR
Citation: Delannoy S, Chaves BD, Ison SA, Webb HE, Beutin L, Delaval J, Billet I and Fach P (2016) Revisiting the STEC Testing Approach: Using espK and espV to Make Enterohemorrhagic Escherichia coli (EHEC) Detection More Reliable in Beef. Front. Microbiol. 7:1. doi: 10.3389/fmicb.2016.00001
Received: 30 October 2015; Accepted: 05 January 2016;
Published: 22 January 2016.
Edited by:
Pina Fratamico, United States Department of Agriculture, Agricultural Research Service, USAReviewed by:
James L. Smith, United States Department of Agriculture, USAGeorge Carl Paoli, United States Department of Agriculture, USA
Copyright © 2016 Delannoy, Chaves, Ison, Webb, Beutin, Delaval, Billet and Fach. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patrick Fach, patrick.fach@anses.fr