 Akifumi Yamashita
Akifumi Yamashita Tsuyoshi Sekizuka
Tsuyoshi Sekizuka Makoto Kuroda
Makoto Kuroda- Pathogen Genomics Center, National Institute of Infectious Diseases, Tokyo, Japan
Although next-generation sequencing (NGS) technology provides a comprehensive means with which to identify potential pathogens from clinical specimens, simple and user-friendly bioinformatics pipelines are expected to obtain the entire viral genome sequence, subsequently providing traceability, based on extensive molecular phylogenetic analyses. We have developed a web-based integrated NGS analysis tool for the viral genome (virus genome-targeted assembly pipeline: VirusTAP), which includes extensive sequence subtraction of host- or bacteria-related NGS reads prior to de novo assembly, leading to the prompt and accurate assembly of viral genome sequences from metagenomic NGS reads. The VirusTAP web site is at https://gph.niid.go.jp/cgi-bin/virustap/index.cgi/.
Introduction
As next-generation sequencing (NGS) technology is becoming a more common means with which to detect pathogens from patients or clinical samples, the need for simple and effective bioinformatics pipelines for the analysis of NGS reads has increased in parallel. One of the major difficulties in this process is the correct de novo assembly of viral genomes from crude metagenomic deep sequencing reads, including large amounts of bacteria and human related sequencing reads. Such read contaminations often force the server to overload during de novo assembly and might cause mis-assembly of the resultant contigs. Pre-filtering by host-mapping subtraction could lead to efficient de novo assembly, allowing the rapid and accurate procurement of a complete viral genome sequence. In addition to the accuracy of de novo assembly, the exclusion of human-related sequences can circumvent conflicting ethical issues by avoiding analyzing the personal genetic information of patients.
To facilitate the steps that require computational resources and skill, we constructed a web-based integrated NGS analysis tool for the viral genome (virus genome-targeted assembly pipeline: VirusTAP) by performing the following informatics steps: (1) quality trimming and adaptor removal, (2) read subtraction of host- and bacteria-related sequences by a read-mapping method, and (3) de novo assembly using multiple combinations of recently advanced assemblers. On the VirusTAP website, users can complete viral genome assemblies from raw NGS reads just by clicking several selections. VirusTAP is freely available for academic users from the following website: https://gph.niid.go.jp/cgi-bin/virustap/index.cgi.
Materials and Methods
Clinical Specimens and NGS Short Reads for Viral Genome Assembly
Sample NGS short reads were prepared using stool specimens from a patient with rotavirus gastroenteritis and analyzed as described previously (Mizukoshi et al., 2014). In brief, total RNA was prepared from patient feces, followed by RNA-seq library preparation using a ScriptSeq v2 RNA-seq library preparation kit (Epicentre, Madison, WI, USA). Deep sequencing was performed to obtain 120-mer paired-end (PE) short reads with a MiSeq Reagent kit v2 (Illumina, San Diego, CA, USA). The sample short-read sequences have been deposited in the DNA Data Bank of Japan (DDBJ; accession number: DRA004165).
The study protocol was approved by the institutional medical ethics committee of the National Institute of Infectious Diseases in Japan (Approval No. 576), and it was conducted according to the Declaration of Helsinki Principles.
NGS Read Processing
VirusTAP accepts single- and PE reads (#1 in Figure 1). Read quality trimming was performed using the skewer (Jiang et al., 2014) or fastq-mcf program in the ea-utils package1, with an additional trimming filter for unreliable sequences after a user specified quality score (#2 in Figure 1). Host read subtraction by read-mapping was performed with the bwasw program (version 0.7.9a-r786; Li and Durbin, 2009) against ribosomal RNAs (16, 18, 23, 28, 5S and internal transcribed spacers rRNA were retrieved from the following ftp site2), bacterial genome sequences3 and the latest host organism genome sequences4 (#3–1 in Figure 1).
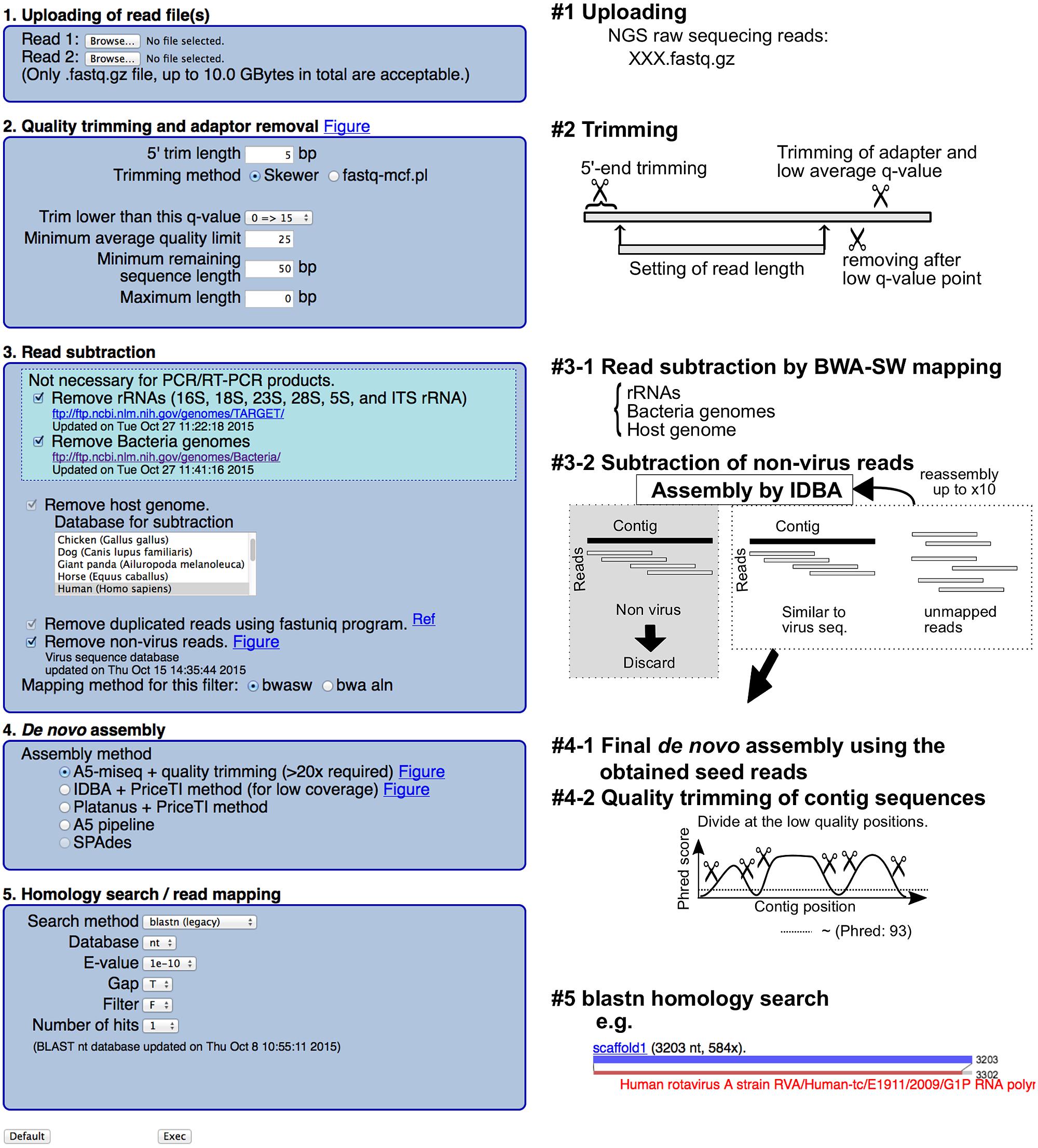
FIGURE 1. Schematic representation of the VirusTAP procedures.
To further remove residual non-virus sequence reads, the above subtracted PE reads are assembled with IDBA-UD (Peng et al., 2012), followed by the filtering of non-virus PE reads with megablast (v. 2.2.26) (Altschul et al., 1997) and RAPSearch2 (v. 2.16) (Zhao et al., 2012) search against the customized viral nucleotide/protein sequences from the NCBI nt/nr database based on virus taxonomy, excluding bacteriophages5 (#3–2 in Figure 1). No significant e-value less than either 1e – 10 for megablast or e – 30 for RAPSearch2 was determined for non-virus contigs, and residual PE reads were further subtracted for the following process. This non-virus filtering step will be repeated up to 10 times or until non-virus contigs disappear. After removing the non-virus sequencing reads, broken PE reads are also removed for the following de novo assembly.
The de novo assembly pipeline can be selected from the following four pipelines (#4–1 in Figure 1): A5-miseq (Coil et al., 2015), Platanus (version 1.2.1) (Kajitani et al., 2014) with PriceTI (Ruby et al., 2013), and IDBA-UD (Peng et al., 2012) with PriceTI (Ruby et al., 2013). PriceTI (v. 1.2) (Ruby et al., 2013) performs PE iterative contig extension from pre-assembly contigs by Platanus or IDBA-UD.
A5-miseq is one of the most recommended de novo assemblers for general NGS PE reads because assemble accuracy for the resultant scaffolds can be checked using the resulting “XXX.scaffolds.fastq” file based on the read-mapping score. The contigs were divided at the nucleotide position, where the Phred score is less than 93 and the resulting sequences shorter than 100 bp are removed. The final scaffolds were subjected to bwasw read mapping and a megablast homology search against the NCBI nt database; the alignment results are visualized at the web page.
Results and Discussion
Basic VirusTAP Procedure
Briefly, a schematic representation of VirusTAP is shown in Figure 1. The VirusTAP process starts with the quality trimming, followed by host- and bacteria-related read subtraction. The read subtraction is performed in the following two steps: first, read-mapping with the bwasw program to rRNA sequences, bacterial genome sequences, and selected virus-host genomes, such as human, mouse, or monkey, was performed; second, non-virus filtering against a virus nucleotide/protein database, excluding bacteriophages, can be selected for further subtraction to extract possible virus-related NGS reads. The above-described highly extensive non-virus read subtraction facilitates effective de novo assembly and prompt analysis, achieving both time savings and accuracy.
VirusTAP Performance Comparison with Direct Metagenomic Assembly Methods
Using metagenomic RNA-seq short reads obtained from stool specimens from a patient with rotavirus gastroenteritis (Mizukoshi et al., 2014), the performance of VirusTAP was compared with that of other de novo assembly methods without subtraction treatment. For instance, direct metagenomic assembly without subtraction using a CLC genome workbench v.8.5.1 (QIAGEN, Aarhus Denmark) or A5-miseq produced 23,937 or 13,365 scaffolds, respectively; VirusTAP significantly reduced the results to 15 scaffolds (Figure 2A step 4).

FIGURE 2. Performance comparison of VirusTAP with non-subtraction methods. (A) Sample metagenomic RNA-seq reads obtained from the rotavirus gastroenteritis patient were analyzed by VirusTAP and non-subtraction methods. (B) Pair-wise alignment view of the most similar hit for each scaffold. Eleven scaffolds showed similarity to eleven segments of rotavirus genome sequences. NCBI Web blast search (BLASTN or BLASTX) can be directly performed for each scaffold by clicking the button to reconfirm the similarity. (C) Visualization of the read-mapping result in the scaffold 4. Viral quasispecies and read coverage on each scaffold can be confirmed by the read-mapping viewer using the bam file in the downloaded results package.
Those direct metagenomic assembly methods generated unexpectedly abundant scaffolds related to multiple organisms at step 4 in Figure 2A. In particular, over 80% of scaffolds were composed of bacterial sequences (19,525 and 11,266 scaffolds by CLC genome and A5-miseq, respectively; Figure 2A), suggesting that subsequent homology searches and taxonomy classifications are very laborious and time-consuming procedures to identify the final viral genome sequence. Indeed, the entire VirusTAP process for these sample reads required ∼17 min to obtain 11 segments of the rotavirus genome sequence, whereas the other two methods required more than 10 h, because of the time-consuming de novo assembly and homology search for all abundant scaffolds (Figure 2A).
In addition to the stool sample, human serum specimen of Dengue fever patient was investigated by RNA-seq, followed by VirusTAP analysis. Since bacterial contamination is not common in serum, plasma, spinal fluid, and urine specimens, VirusTAP efficiently removed the more than 90% of host-related sequence in total reads to obtain possible virus sequence reads (data not shown). Only human genome subtraction could be sufficient for the serum specimen, but fecal or pharyngeal specimens contain unfavorable bacterial reads for virus genome assembly. This study demonstrated that rather laborious sample such as bacteria rich stool sample can be acceptable to determine a virus genome by VirusTAP, it includes non-virus filtering (see step #3–2 in Figure 1) to facilitate the virus genome assembly.
The results of the homology search of pair-wise alignment for each scaffold can be visualized on the web (Figure 2B) or by downloading the full-results package, although VirusTAP provides the results against the latest NCBI nt database. Thus, users can reconfirm the obtained scaffolds by NCBI BLASTN or BLASTX web search with a direct web link (Figure 2B). The downloaded results package includes the bwa-mapping “sorted.bam” file, which can be imported into the reads-mapping viewer to visualize nucleotide polymorphisms, insertion-deletion and read coverage depth. In this sample study, a T/C heterogeneous mixture was identified at the 1,729 nt position of scaffold 4 (Figure 2C), indicating a possible quasispecies with a valine or alanine residue at the 578 aa position of the VP4 protein. VP4 is a spike protein at the outer capsid and has receptor binding activities and functions in membrane penetration (Trask et al., 2012), suggesting that the quasispecies could emerge during rotavirus gastroenteritis, contributing to the antigenic drift during rotavirus infection.
Author Contributions
TS and MK performed the metagenomic sequencing and statistical and bioinformatics analyses. AY and MK participated in the design of the study, performed the statistical analysis, and drafted the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by a grant for Research on Emerging and Re-emerging Infectious Diseases (H25 Shinko-Ippan-015) from the Ministry of Health, Labour and Welfare, Japan, and was also supported by the Research Program on Emerging and Re-emerging Infectious Diseases (15fk0108011h0003 and 15fm0108022h0001) from the Japan Agency for Medical Research and Development, AMED. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We would like to thank Dr. Kei Haga for helpful suggestions.
Footnotes
- ^ http://code.google.com/p/ea-utils/
- ^ ftp://ftp.ncbi.nlm.nih.gov/genomes/TARGET/
- ^ ftp://ftp.ncbi.nlm.nih.gov/genomes/all/
- ^ ftp://ftp.ncbi.nlm.nih.gov/genomes/
- ^ ftp://ftp.ncbi.nih.gov/pub/taxonomy/
References
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Coil, D., Jospin, G., and Darling, A. E. (2015). A5-miseq: an updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 31, 587–589. doi: 10.1093/bioinformatics/btu661
Jiang, H., Lei, R., Ding, S. W., and Zhu, S. (2014). Skewer: a fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinformatics 15:182. doi: 10.1186/1471-2105-15-182
Kajitani, R., Toshimoto, K., Noguchi, H., Toyoda, A., Ogura, Y., Okuno, M., et al. (2014). Efficient de novo assembly of highly heterozygous genomes from whole-genome shotgun short reads. Genome Res. 24, 1384–1395. doi: 10.1101/gr.170720.113
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Mizukoshi, F., Kuroda, M., Tsukagoshi, H., Sekizuka, T., Funatogawa, K., Morita, Y., et al. (2014). A food-borne outbreak of gastroenteritis due to genotype G1P[8] rotavirus among adolescents in Japan. Microbiol. Immunol. 58, 536–539. doi: 10.1111/1348-0421.12176
Peng, Y., Leung, H. C., Yiu, S. M., and Chin, F. Y. (2012). IDBA-UD: a de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 28, 1420–1428. doi: 10.1093/bioinformatics/bts174
Ruby, J. G., Bellare, P., and Derisi, J. L. (2013). PRICE: software for the targeted assembly of components of (Meta) genomic sequence data. G3 (Bethesda) 3, 865–880. doi: 10.1534/g3.113.005967
Trask, S. D., Ogden, K. M., and Patton, J. T. (2012). Interactions among capsid proteins orchestrate rotavirus particle functions. Curr. Opin. Virol. 2, 373–379. doi: 10.1016/j.coviro.2012.04.005
Keywords: NGS, viral genome, de novo assembly, web service, host genome subtraction
Citation: Yamashita A, Sekizuka T and Kuroda M (2016) VirusTAP: Viral Genome-Targeted Assembly Pipeline. Front. Microbiol. 7:32. doi: 10.3389/fmicb.2016.00032
Received: 24 November 2015; Accepted: 11 January 2016;
Published: 02 February 2016.
Edited by:
Akihide Ryo, Yokohama City University, JapanReviewed by:
Anita Schürch, University Medical Center Utrecht, NetherlandsYoshinori Ito, Nagoya University Graduate School of Medicine, Japan
Osvaldo Zagordi, University of Zurich, Switzerland
Copyright © 2016 Yamashita, Sekizuka and Kuroda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Makoto Kuroda, makokuro@niid.go.jp