Corrigendum: Impact of Resistance to Fluconazole on Virulence and Morphological Aspects of Cryptococcus neoformans and Cryptococcus gattii Isolates
 Suélen A. Rossi1
Suélen A. Rossi1 Nuria Trevijano-Contador2
Nuria Trevijano-Contador2 Liliana Scorzoni1
Liliana Scorzoni1 Ana Cecilia Mesa-Arango3
Ana Cecilia Mesa-Arango3 Haroldo C. de Oliveira1Karin Werther4Tânia de Freitas Raso4
Haroldo C. de Oliveira1Karin Werther4Tânia de Freitas Raso4 Maria J. S. Mendes-Giannini1
Maria J. S. Mendes-Giannini1 Oscar Zaragoza2
Oscar Zaragoza2 Ana M. Fusco-Almeida1*
Ana M. Fusco-Almeida1*- 1Faculdade de Ciências Farmacêuticas, UNESP – Universidade Estadual Paulista, Campus Araraquara, Departamento de Análises Clíncas, São Paulo, Brazil
- 2Centro Nacional de Microbiologia, Unidad de Micologia, Instituto de Salud Carlos III, Majadahonda, Madrid, Spain
- 3Dermatology Research Group, Universidad de Antioquia, Medellín, Colombia
- 4Faculdade de Medicina Veterinária e Zootecnia, USP – Universidade de São Paulo, Departamento de Patologia, São Paulo, Brazil
Cryptococcus sp. are responsible for around 1 million cases of meningitis every year. Fluconazole (FLU) is commonly used in the treatment of cryptococcosis, mainly in immunocompromised patients and the resistance is usually reported after long periods of treatment. In this study, the morphological characterization and virulence profile of FLU-susceptible and FLU-resistant clinical and environmental isolates of C. neoformans and C. gattii were performed both in vitro and in vivo using the Galleria mellonella model. FLU-susceptible isolates from C. neoformans were significantly more virulent than the FLU-resistant isolates. FLU-susceptible C. gattii isolates showed a different virulence profile from C. neoformans isolates where only the environmental isolate, CL, was more virulent compared with the resistant isolates. Cell morphology and capsule size were analyzed and the FLU-resistant isolates did not change significantly compared with the most sensitive isolates. Growth at 37°C was also evaluated and in both species, the resistant isolates showed a reduced growth at this temperature, indicating that FLU resistance can affect their growth. Based on the results obtained is possible suggest that FLU resistance can influence the morphology of the isolates and consequently changed the virulence profiles. The most evident results were observed for C. neoformans showing that the adaptation of isolates to antifungal selective pressure influenced the loss of virulence.
Introduction
Cryptococcus neoformans and Cryptococcus gattii are the main etiologic agents of cryptococcosis. This infection occurs after inhalation of basidiospores, which are found in the environment (Martinez and Casadevall, 2006; Giles et al., 2009; Negroni, 2012). The clinical manifestations usually occur in pulmonary and cerebral forms and may ultimately progress to the most severe of the disease, meningoencephalitis (Chau et al., 2010; Kessler et al., 2010; Negroni, 2012). Globally, approximately one million cases of cryptococcal meningitis occur each year, the majority of which occur in sub-Saharan Africa (Park et al., 2009).
Cryptococcus neoformans has a worldwide distribution and mainly affects immunocompromised patients, causing high morbidity and mortality in these individuals (Li and Mody, 2010). C. gattii infections have historically been prevalent in regions with tropical and subtropical climates; however, outbreaks in humans and animals in temperate countries have been observed, which demonstrates that the fungus can adapt to new environments (Kidd et al., 2004; Byrnes and Heitman, 2009; Hagen et al., 2012). C. gattii is traditionally associated with immunocompetent individuals, but immunocompromised patients are also at risk of infection (Jobbins et al., 2010; Harris et al., 2011).
Some authors have reported that the sensitivity to azoles and virulence profiles can vary depending on the geographic region that the yeast is isolated, whether the isolate is clinical or environmental and on the molecular fungus type (Bovers et al., 2008a; Espinel-Ingroff et al., 2012a,b).
Currently, the Cryptococcus species complex is comprises in two species, C. neoformans serotypes A, D, and AD and C. gattii serotypes B and C (Boekhout et al., 1997, 2001; Kwon-Chung and Varma, 2006). C. neoformans consists of two varieties, C. neoformans variety grubii (serotype A) and C. neoformans variety neoformans (serotype D; Franzot et al., 1999). However, in the last years, several studies using molecular techniques have questioned the currently used of two species concept in the Cryptococcus species complex (Boekhout et al., 2001; Kidd et al., 2004; Bovers et al., 2008b; Meyer et al., 2009; Hagen et al., 2010, 2015). C. neoformans var. grubii is represented by AFLP1/VNI and two minor genotypes AFLP1A/VNII (VNB) and AFLP1B/VNII (Barreto de Oliveira et al., 2004; Litvintseva et al., 2006). C. neoformans var. neoformans is represented by AFLP2/VNIV and the hybrid (AD) by AFLP3/VNIII genotype (Trilles et al., 2003; Meyer et al., 2009). C. gattii is divided into five genotypes, AFLP4/VGI, AFLP6/VGII, AFLP7, and AFLP10/VGIV (serotypes B and C) and AFLP5/VGIII (Fraser et al., 2005; Hagen et al., 2010). Recently, Hagen et al. (2015) through phylogenetic and genotyping analysis, proposed to recognize the current C. neoformans var. grubii and C. neoformans var. neoformans as two separate species and, C. gattii in five separated species.
The increase in invasive fungal infections and the development of resistance mechanisms by some fungal species is of great concern given that antifungal treatment is usually aggressive, toxic, and inefficient (Spinello, 2013). Although the detection of resistance to antifungal agents is difficult, increased rates of fungal infections by resistant isolates and changes in breakpoints have been reported in patients exposed to long-term therapies (Fera et al., 2009; Wilke, 2011).
Virulence factors are mechanisms that allow the fungus to cause damage to the host (Casadevall and Pirofski, 1999, 2003). Many phenotypes have been specifically correlated with the virulence of C. neoformans, such as capsule production, melanin formation, and protein secretion. Furthermore, cell characteristics such as the cell wall and morphogenesis play an important role in host-fungus interactions (Alspaugh, 2014).
Murine animal models have been used in several studies to evaluate virulence and the efficacy of antifungal agents against different fungal species (Okawa et al., 2008); however, there is currently a particular interest in developing alternative models for studying microbial virulence (Mylonakis and Aballay, 2005; Chrisman et al., 2011; Muhammed et al., 2012). These models have provided considerable knowledge on different aspects of microbial infection (Fedhila et al., 2010) and were developed to present numerous advantages over mammalian models, such as low cost, ease of use, potential to conduct large-scale studies, and compliance with global trends regarding animal welfare and bioethics issues (Jacobsen, 2014).
Galleria mellonella is a lepidopteran that has been successfully used as a model to study the virulence of pathogenic fungi such as C. neoformans and C. gattii (Mylonakis et al., 2005; García-Rodas et al., 2011; Firacative et al., 2014), Candida albicans (Brennan et al., 2002; Rueda et al., 2014), and Aspergillus fumigatus (Cheema and Christians, 2011; Gomez-Lopez et al., 2014). Moreover, it has also been used to investigate the efficacy of antifungal therapy (Cowen et al., 2009; Mesa-Arango et al., 2012; Scorzoni et al., 2013).
It is known that the adaptation capacity of C. albicans to host environment through virulence factors can be modified in the presence or absence of a drug (Schulz et al., 2011). In bacteria, the development of antibiotic resistance mechanisms can generate a cost in adaptation (Levin et al., 1997; Andersson and Levin, 1999), especially by reducing the rate of bacterial growth (Andersson and Hughes, 2010). The impact of antifungal acquired resistance on virulence in some microorganisms, such as A. fumigatus, is still not well-understood. However, it is known that acquisition of resistance by specific mechanisms may result in phenotypic changes, such as, germination rate, hyphal growth, or growth rate of the resistant isolates. These changes, in turn, may result in a significant loss of virulence (Gomez-Lopez et al., 2014). These hypotheses show to us the importance further studies to better understand the relationship between the resistance to antifungal agents and the adaptation of the fungus to the host environment. Thus, in this study, we evaluated the impact of the development of resistance to fluconazole (FLU) by isolates of C. neoformans and C. gattii both on virulence and on morphological changes in vitro and in vivo using the G. mellonella model.
Materials and Methods
Strains and Growth Conditions
Three sequential clinical isolates of C. neoformans var. grubii (AFLP1VNI) were collected and classified according to the in vitro susceptibility to FLU as follows: resistant (30R), susceptible dose-dependent (27SDD), and susceptible (26S). Isolates were recovered from an HIV-positive patient with a history of relapses associated with treatment failure and were characterized by molecular typing and susceptibility to antifungal therapy. Additionally, the isolates were analyzed by RAPD (random amplified polymorphic DNA) and showed identical genetic profiles (data not shown). In addition, two environmental C. gattii isolates (isolated from a psittacine bird; AFLP6/VGII) were evaluated, one with reduced susceptibility to FLU (118R; Raso et al., 2004) and one with susceptibility to FLU (CL).The strains C. neoformans ATCC 90112 (AFLP1/VN1; The American Type Culture Collection) and C. gattii ATCC 56990 (AFLP4/VGI) were used. All species were obtained from the collection of the Laboratory of Clinical Mycology, Department of Clinical Analysis, Faculty of Pharmaceutical Sciences, UNESP, Araraquara.
Antifungal Susceptibility
To confirm the phenotype obtained initially, antifungal susceptibility tests were conducted for all isolates using FLU and amphotericin B (Amb), according to M27-A3 document from the Clinical and Laboratory Standards Institute. The range of antifungal concentrations used was 128–0.125 μg/ml for FLU and 16–0.0321 μg/ml for Amb.
Cryptococcus sp. Growth Curve at Different Temperatures
For each assay, 104 cells/mL were inoculated in 100 mL of Sabouraud broth. One milliliter of these cultures was collected after 3, 6, 9, 12, 24, 48, and 72 h and the optical density was determined at 600 nm using a spectrophotometer (Thermo Scientific Genesis). The Sadouraud broth was used as control in order to normalize readings.
Cryptococcus sp. In Vitro Capsule Induction
For in vitro morphological analysis, we induced capsule growth in all isolates as described by Zaragoza and Casadevall (2004). First, Cryptococcus sp. were cultivated in Sabouraud broth overnight at 30°C. Next, the inocula were concentrated in PBS (1 mL) and 100 μl were transferred to 10% Sabouraud buffered at pH 7.3 with 50 mM MOPS (3-(N-Morpholino) propanesulfonic acid, 4-Morpholinepropanesulfonic acid) and incubated for 24 h at 37°C with shaking at 150 rpm. Yeast cells were then suspended in India ink and photographed using a Leica DMI3000B microscope. Cell size was measured using Adobe Photoshop CS3 software. Statistics were performed by ANOVA with Tukey’s post-test.
Galleria mellonella Rearing and Larvae Manipulation
For all experiments, we used G. mellonella in the larval stage (Alcotán, Valencia, Spain). The larvae were fed with wax and pollen and maintained at 25°C (Ramarao et al., 2012) until reaching 0.2–0.3 g. Before infection, larvae pro-legs were cleaned with 70% ethanol and yeast suspensions were injected into larvae pro-legs using 10 μl Hamilton syringes (Hamilton, USA). Before all experiments, larvae were incubated at 30 or 37°C overnight and protected from light. Each experiment was performed in triplicate.
Survival Assay
Galleria mellonella larvae were infected with 106, 5 × 106, 107, 2.5 × 107, and 5 × 107 cells per larva and larval death was monitored over a period of 8 days. Lack of movement was the criterion used to define larval death. Larvae were incubated at 30 or 37°C. PBS-injected larvae and groups inoculated with C. neoformans ATCC 90112 of and C. gattii ATCC 56990 were used as controls. Statistics were analyzed using the Log-rank (Mantel–Cox) test.
Hemolymph Melanization
The melanization assay was performed as described by Scorzoni et al. (2013). Briefly, 10 larvae were infected with 107 cells/larvae and incubated at 37°C. The hemolymph was collected after 1 and 5 h of infection and 10 μl of the hemolymph was diluted 1:10 in cold PBS and centrifuged at 10,000 rpm for 5 min. The supernatants were transferred into a 96-well plate and read at 405 nm in a spectrophotometer (Thermo Scientific Genesis). PBS-injected larvae were used as a control. Statistics were performed by ANOVA with Tukey’s post-test.
Cryptococcus sp. Cell and Capsule Size Alterations After Infection in G. mellonella
Larvae were infected with 106 cells per larva and incubated at 37°C. Immediately after infection as well as 1 and 3 days post-infection, larvae were homogenized in 100 mm pore nylon filters (Falcon, BD, USA) with 1 mL of PBS. The resulting fluid was collected and centrifuged. The cell pellet was washed (1x) and suspended in 300 μl of PBS. Subsequently, an aliquot of the cell sample was stained with India ink and visualized using a Leica DMI 3000B microscope. Adobe Photoshop CS3 software was used to measure the cells. Statistics were performed by ANOVA with Tukey’s post-test.
Cryptococcus sp. Phagocytosis Assays in G. mellonella
Isolates of Cryptococcus sp. were first stained with 10 μg/ml of calcofluor white (Sigma, St. Louis, MO, USA) for 30 min at 37°C. Then, larvae were infected with 106 cells and phagocytosis was analyzed after 3 h of incubation at 37°C. Hemolymph was collected in Eppendorf tubes and diluted 1:2 in cold PBS. Phagocytosis was quantified visually using a Leica DMI 3000B microscope. Five larvae from each experimental situation were used. One hundred hemocytes from each larva were counted and the percentage of hemocytes containing Cryptococcus sp. internalized was calculated. Statistics were performed by ANOVA with Tukey’s post-test.
Results
Susceptibility Testing against Fluconazole and Amphotericin B
Antifungal susceptibility testing results showed that the C. neoformans isolate 30R and the C. gattii isolate 118R were resistant to FLU with MIC values of 64 mg/L. C. neoformans 27SDD (susceptible-dose dependent) showed a MIC value of 16 mg/L and ATCC 90112 and 26S were characterized as susceptible to FLU with MICs of 1 and 2 mg/L, respectively. C. gattii isolates CL and ATCC 56990 showed MICs of 4 mg/L. All isolates were susceptible to AmB with MIC concentrations ranging between 0.125 and 0.0625 mg/L. Throughout the work, all isolates were cultured in Sabouraud without adding FLU and even after several transfers, it became clear that resistance remained.
Growth Curve at Different Temperatures
The assay was carried out at 30 and 37°C. C. neoformans and C. gattii showed better growth at 30°C than at 37°C. All C. neoformans isolates showed slower growth at 37°C, especially FLU-resistant strain 30R. When the growth curves of C. gattii were analyzed, it was found that isolate ATCC 56990 was unable to grow at 37°C and isolate 118R showed much lower growth compared with the susceptible isolate CL at 37°C (Figure 1).
FIGURE 1
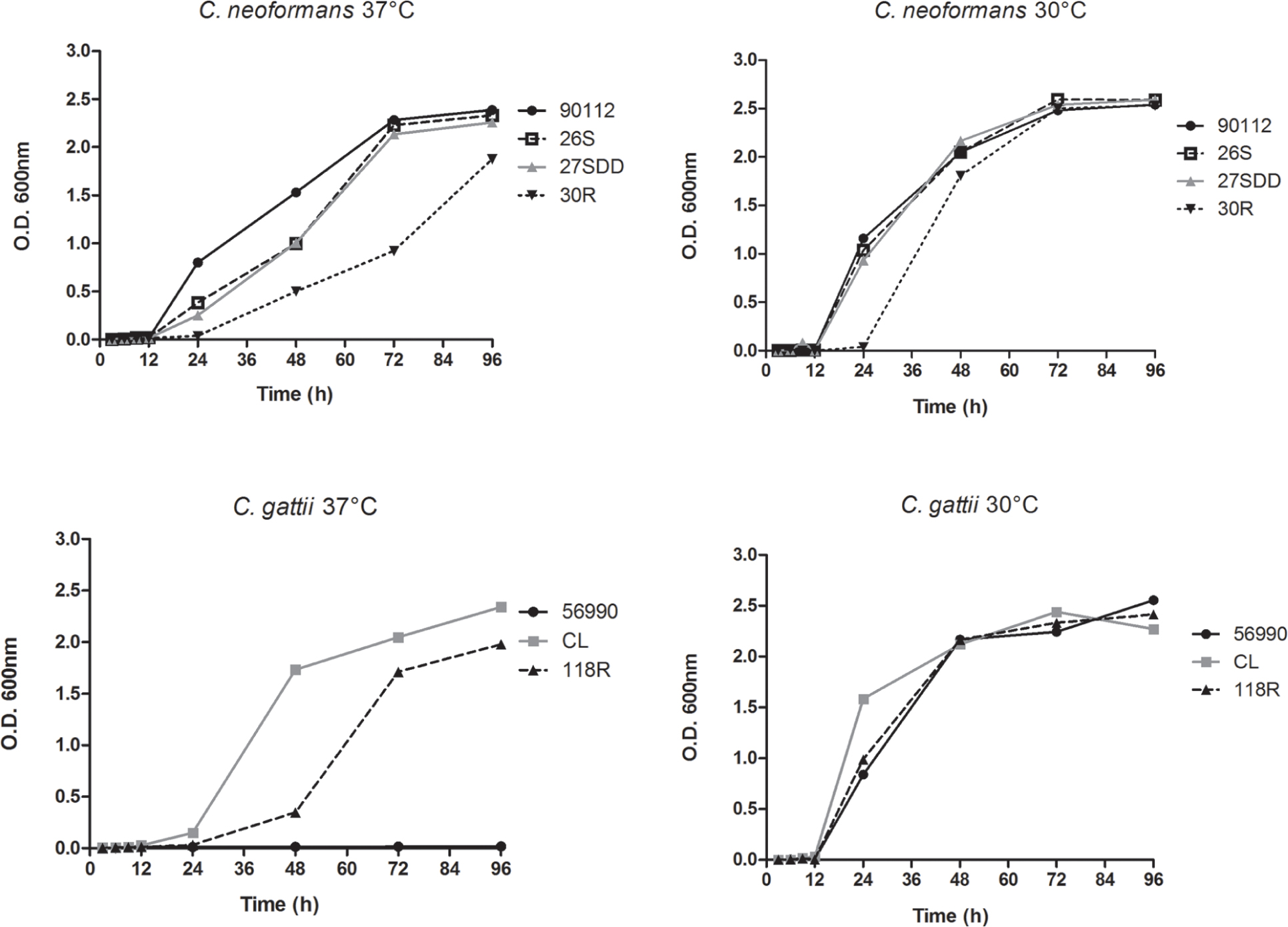
FIGURE 1. Sequential growth curve of Cryptococcus neoformans isolates, C. gattii isolates and the ATCC strains 90112 and 56990 at 30 and 37°C.
Cryptococcus sp. Virulence at 30 and 37°C
Sequential clinical isolates of C. neoformans (26S, 27SDD, and 30R) have different virulence profiles. The thermotolerance is a very important virulence factor in Cryptococcus sp., therefor the virulence test was performed at 30 and 37°C. This difference was more evident between the FLU-susceptible (26S) and FLU-resistant (30R) isolates at 37°C (Figure 2A). At 30°C, the C. neoformans isolates were less virulent compared with activity at 37°C, but the virulence profiles were maintained between them with being 26S more virulent and killing 100% of the larvae at day 6 of infection (Figure 2B). The C. neoformans strain ATCC 90112 was also used in this assay and its virulence profile was very similar to the profile observed for the FLU-susceptible isolate 26S. The virulence of the C. gattii isolates at both temperatures was lower when compared to the C. neoformans isolates (Figures 2C,D). The strain C. gattii ATCC 56990 did not kill any larvae when infection was conducted at 37° (2C). The FLU-resistant isolate 118R killed 40% of the larvae, while FLU-susceptible isolate CL killed 90% of the larvae at day 8 of infection at 37°C (Figure 2C). At 30°C, we found a different virulence profile for C. gattii, when compared to the assay performed at 37°C.
FIGURE 2
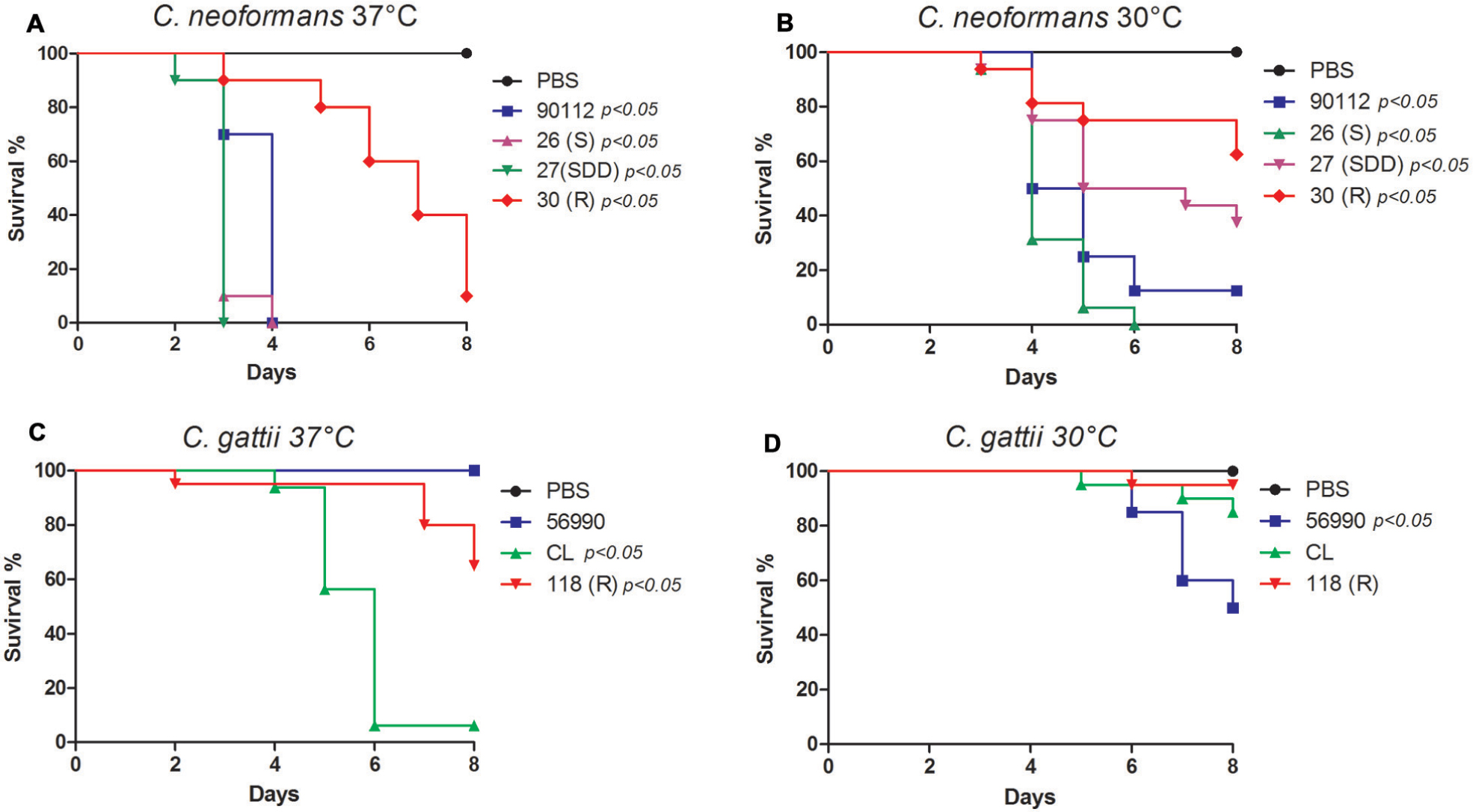
FIGURE 2. Virulence assay in Galleria mellonella. (A) Sequential clinical isolates of C. neoformans, 26S, 27SDD, and 30R incubated at 37°C; (B) Sequential clinical isolates of C. neoformans, 26S, 27SDD, and 30R incubated at 30°C; (C) Environmental isolates of C. gattii 118R and CL incubated at 37°C; (D) Environmental isolates of C. gattii 118R and CL incubated at 30°C.
To confirm the reduced virulence profile of the C. gattii isolates, the experiment was repeated with four additional isolates from the same species. These isolates were provided by the Mycology Reference Laboratory at the Institute of Health Carlos III of Madrid, Spain, and were identified as CL 5004 (AFLP4/VGI), CL 5010 (AFLP4/VGI), NIH191 (AFLP5/VGIII), and NIH198 (AFLP5/VGIII). Again, all C. gattii isolates were less virulent than C. neoformans (Supplementary Figure S1).
We also performed a survival curve with different inoculum concentrations (1 × 106, 5 × 106, 1 × 107, 2.5 × 107, and 5 × 107) at 37°C. This test was performed with isolates that were less virulent with standard inoculum of the 106 cells per larva in G. mellonella and was observed that the low virulence remained even when the inoculum was increased up to 5 × 107 cells per larvae (Supplementary Figure S2).
Hemolymph Melanization
During the virulence assays in G. mellonella, we observed significant melanization of larvae after infection with some isolates. Because of these findings, quantification of melanization in the hemolymph of the larvae was performed to verify this phenomenon. The results showed that the infection of larvae with all C. gattii isolates led to a significant melanin production after 5 h of infection (Figure 3D). However, as shown in Figure 3B, only the ATCC 90112 and 27SDD isolate of C. neoformans produced significant melanization after infection. In Figures 3A,C is observed that after 1 h of infection there was no significant melanization for most of the isolates, except for ATCC 56990 strain.
FIGURE 3
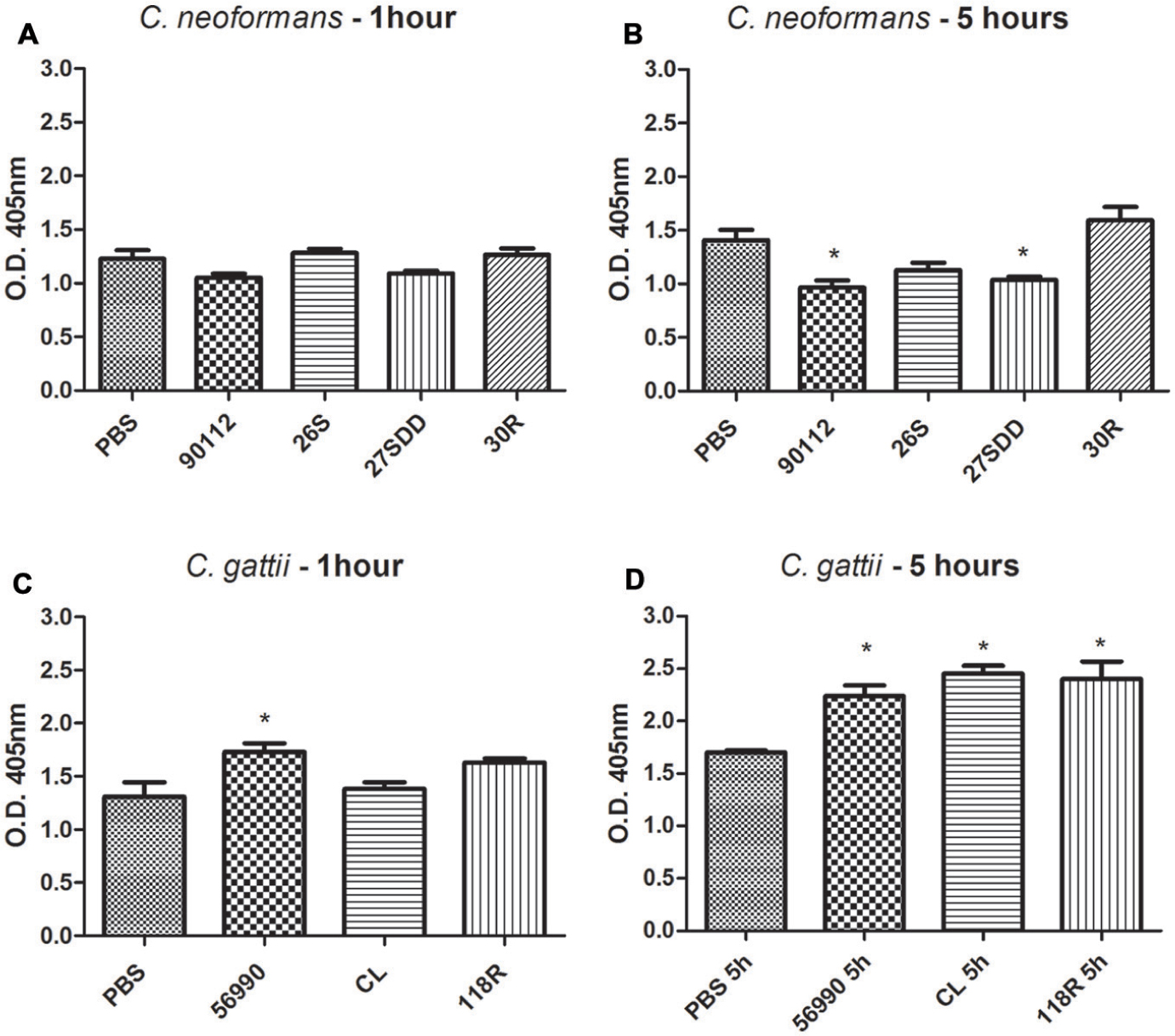
FIGURE 3. Quantification of melanization of C. gattii and C. neoformans isolates after inoculation in G. mellonella. The quantification was measured at different times, 1 and 5 h. (A) C. neoformans 1 h; (B) C. neoformans 5 h; (C) C. gattii 1 h; and (D) C. gattii 5 h. ∗p < 0.05.
In Vitro Capsule Induction at 37°C
We investigated whether resistance to FLU was associated with cell and capsule increase. After 24 h of incubation in capsule-inducing medium, all sequential clinical isolates of C. neoformans increased capsule size compared to the cells incubated in Sabouraud (p < 0.05, Figure 4A). When comparing the FLU-resistant isolate (30R) with the other isolates, the changes in 30R capsule size were more discrete compared to the ATCC 90112 and 27SDD isolates (p < 0.05).
FIGURE 4
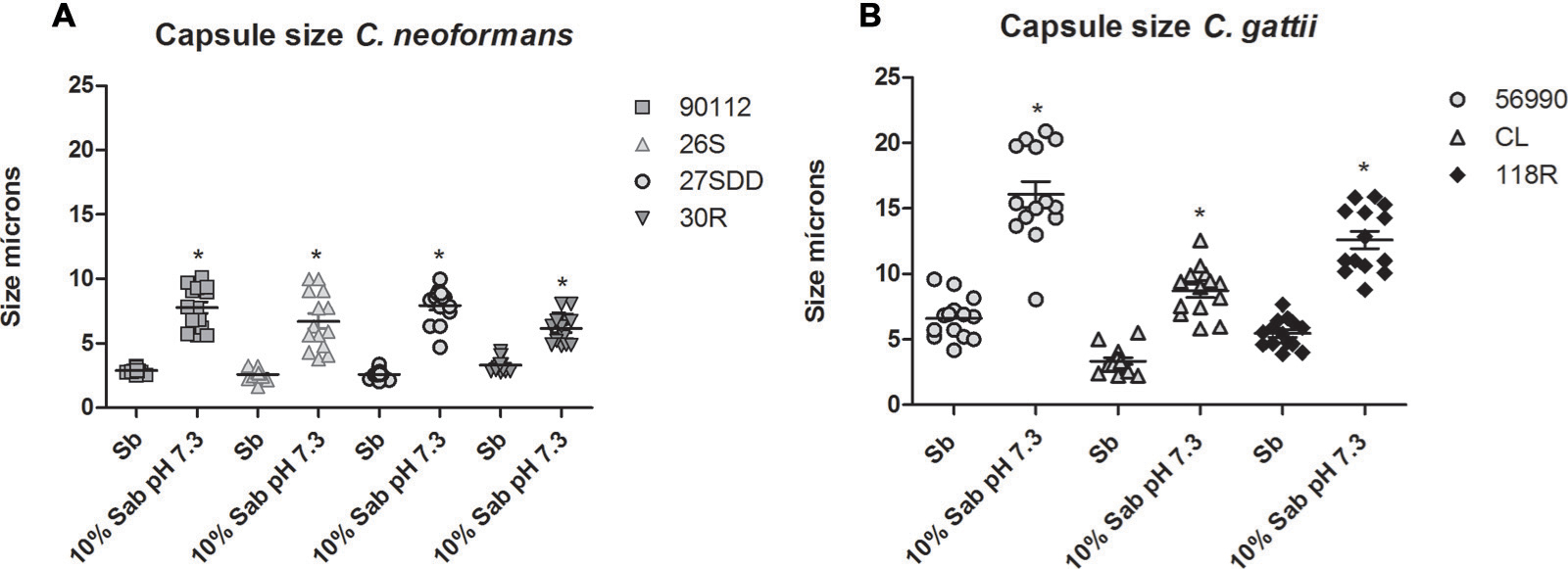
FIGURE 4. In vitro capsule induction of clinical isolates of C. neoformans and C. gattii environmental isolates. (A) ATCC 90112 and clinical isolates of C. neoformans 26S, 27SDD, and 30R; (B) ATCC 56990 and environmental isolates of C. gattii CL and 118R. ∗p < 0.05.
For C. gattii isolates, we observed that the cells of the environmental isolates and of ATCC 56990 had greater capsule induction (p < 0.05, Figure 4B) after incubation in capsule-inducing medium. The FLU-resistant 118R isolate achieved a remarkable increase in capsule size and showed a less significant change in capsule size only when compared with the strain ATCC 56990 (p < 0.05).
Morphology of Cells in G. mellonella
The total cell size and capsule size were measured before infection (Day 0) as well as 24 h (Day 1) and 72 h (Day 3) after infection in G. mellonella. A progressive increase in cell size was observed in C. neoformans isolates across the specified time intervals (D1 and D3) when compared with Day 0 (D0; p < 0.05), with the exception of the 27SDD isolate, which showed no significant changes after infection (Figures 5A and 7A–D). When the isolates were analyzed individually, it was found that the FLU-susceptible isolate 26S and the ATCC 90112 strain also showed a significant increase between D1 and D3 (p < 0.05). For C. gattii isolates, a significant increase in the cell size of ATCC 56990 and CL was observed at D1 and D3 in comparison to D0 (Figures 5B and 7E–G). The largest increase was observed in C. gattii strain ATCC 56990, where some cells reached 40 μm at D3 (Figure 7E). FLU-susceptible CL isolate achieved significant growth. The FLU-resistant 118R isolate only showed a significant increase at D3, however, no cells larger than 20 μ were observed (p < 0.05; Figure 7G).
FIGURE 5
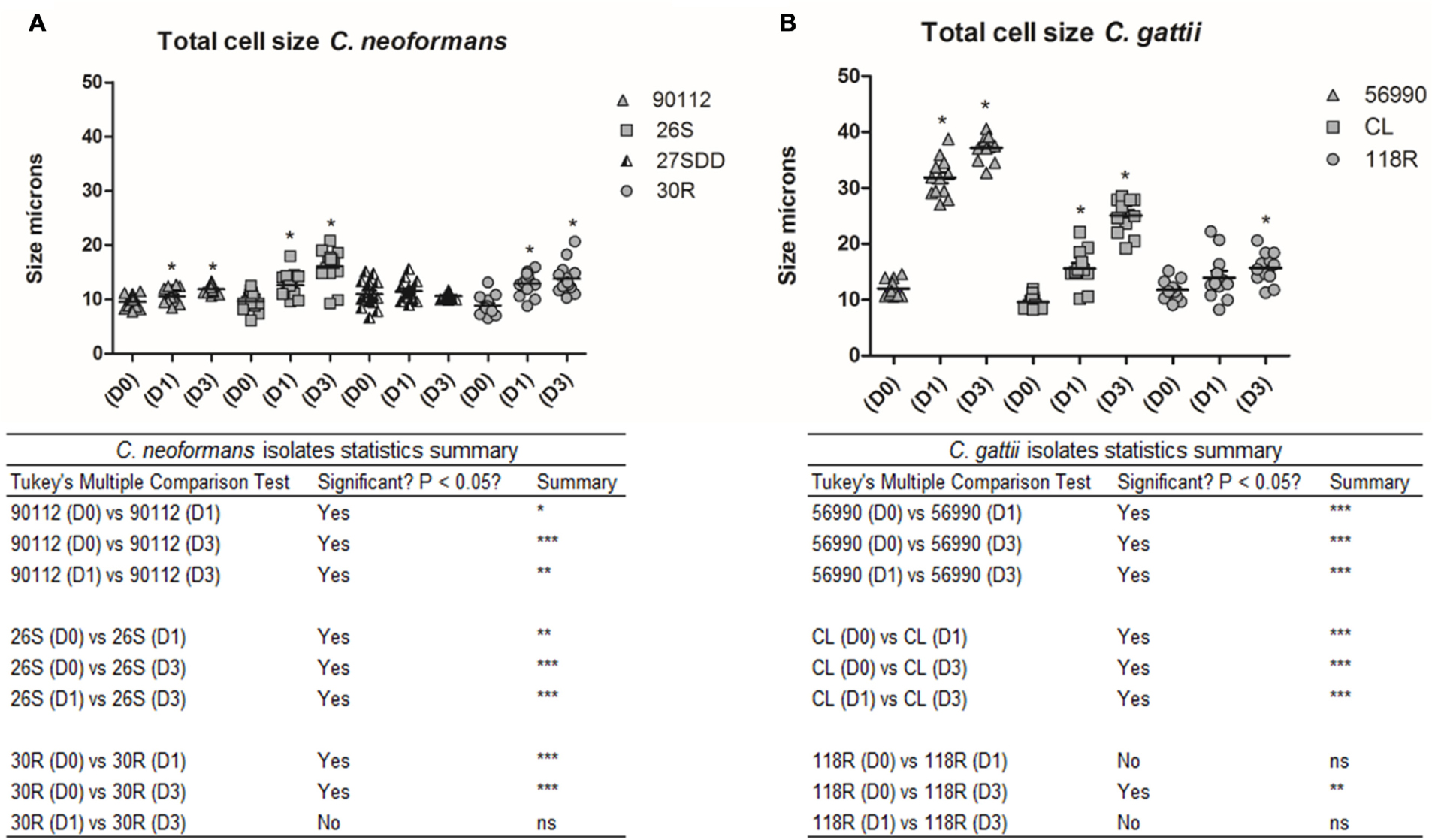
FIGURE 5. Total cell size in microns for C. neoformans and C. gattii isolates during infection in G. mellonella. The cells were recovered from G. mellonella at different times (D1 and D3) and controls were grown in Sabouraud (D0). (A) ATCC 90112 and sequential clinical isolates 26S, 27SDD, and 30R of C. neoformans; (B) ATCC 56990 and isolates CL and 118R of C. gattii. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
We also measured the capsule size of the different isolates. As shown in Figures 6A and 7A–D, all C. neoformans isolates presented a significant increase in capsule size after infection when compared with D0 (p < 0.05) but only 26S showed a significant and gradual increase. C. gattii isolates had a much greater capsule size than C. neoformans isolates (see Figures 6B and 7E–G). The cells from C. gattii ATCC 56990 strain had a greater change in size after infection in G. mellonella. Most cells of this strain reached a capsule size ranging between 20 and 30 μ (p < 0.05). The capsule of environmental FLU-susceptible CL isolate presented a gradual increase at all-time intervals (p < 0.05) and the 118R isolate had a significant increase only at D1 (p < 0.05).
FIGURE 6
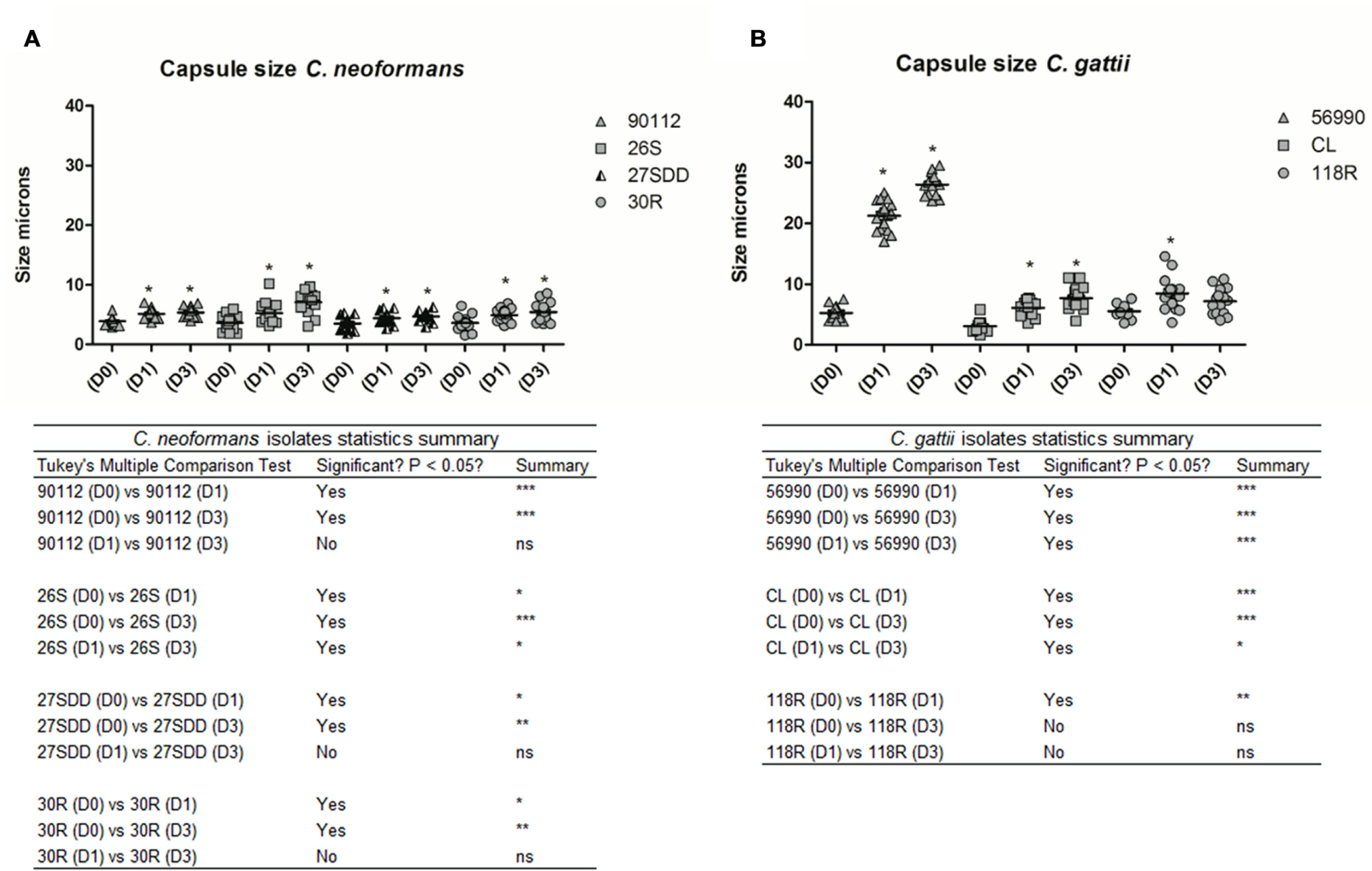
FIGURE 6. Capsule size in microns of isolates of C. neoformans and C. gattii after infection in G. mellonella. The cells were recovered from G. mellonella at different times (D1 and D3) and controls were grown in Sabouraud (D0). (A) ATCC 90112 and isolates 26S, 30R, and 27SDD of C. neoformans; (B) ATCC 56990 and isolates CL and 118R of C. gattii. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
FIGURE 7

FIGURE 7. Microscopy of C. neoformans and C. gattii isolates with India ink stain before (D0) and after infection in G. mellonella (D1, 24 h) and (D3, 72 h); (A) C. neoformans ATCC 90112; (B) C. neoformans isolate 26S; (C) C. neoformans isolate 27SDD; (D) C. neoformans isolate 30R; (E) C. gattii ATCC 56990; (F) C. gattii isolate CL; and (G) C. gattii isolate 118R.
In Vivo Phagocytosis Assay
To better elucidate the mechanisms of virulence of the selected isolates, we also performed a phagocytosis assay in G. mellonella. This assay showed that C. neoformans 30R isolate was phagocytosed to a lower degree compared with the ATCC 90112 (p < 0.05) strain. Among the other sequential clinical isolates, 26S, 27SDD and 30R, there was no significant difference observed in phagocytosis (Figure 8A). The C. gattii 118R isolate was the strain better phagocytosed, with almost a 40% phagocytosis rate compared to the CL and ATCC 56990 strains after infection in G. mellonella (see Figure 8B).
FIGURE 8
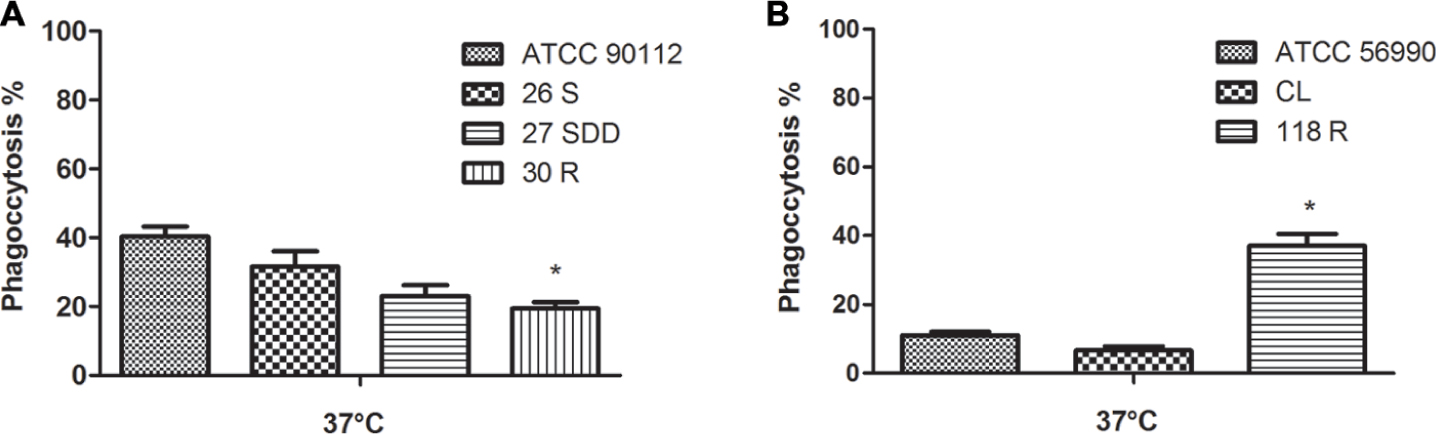
FIGURE 8. Phagocytosis index after infection in G. mellonella. (A) ATCC 90112 and isolates 26S, 27SDD, and 30R of C. neoformans; (B) ATCC 56990 and isolates 118R and CL of C. gattii. ∗p < 0.05.
Discussion
Fluconazole achieves good penetration into the central nervous system (CNS) and has minimal side effects; for this reason, FLU is commonly used in the treatment of cryptococcosis, mainly in immunocompromised patients (Brandt et al., 2001). However, in some strains, repeated exposure or prolonged treatment with azoles can lead to the development of resistance (Sanglard, 2002; Bicanic et al., 2009; Fera et al., 2009).
It is known that the development of azole resistance in fungi may occur through overexpression or mutations in the ERG11 gene, which encodes the enzyme lanosterol 14-α demethylase (ERG11p; Sanglard et al., 1998; Cowen et al., 2000; Rodero et al., 2003; Sheng et al., 2009; Oliveira Carvalho et al., 2013), and/or by overexpression of plasma membrane proteins that pump the drug out of the cells (Posteraro et al., 2003; Sanguinetti et al., 2006; Sanglard et al., 2009). In addition, the phenomenon of heteroresistance to FLU, an adaptive mechanism of drug tolerance, has been previously described in C. neoformans and C. gattii species (Sionov et al., 2009, 2010; Varma and Kwon-Chung, 2010).
In addition to the eight major molecular types identified from molecular techniques (C. neoformans, VNI, VNII, VNIII, VNIV, and C. gattii, VGI, VGII, VGIII e VGIV), VGII has received increased interest by different authors in recent years due to the appearance of the subtypes, VGIIa, VGIIb e VGIIc (Byrnes et al., 2010; Chong et al., 2010; Firacative et al., 2014), and the discovery that susceptibility to FLU varies depending on the molecular type. Other epidemiological studies have also found that, unlike isolates from Europe and North America, yeast species originating from Africa, the Middle East, Asia-Pacific, and parts of Latin America shows variable sensitivity to FLU (Pfaller et al., 2009). The relationship between FLU resistance and virulence in Cryptococcus sp. has been investigated but remains poorly understood. Sionov et al. (2009) found that strains of C. neoformans expressing FLU heteroressistance in vitro (MIC ≥ 32 μg/ml) were more virulent in mice when compared with more sensitive strains (MIC ≤ 8 μg/ml). In vivo data have also demonstrated that a strain of C. neoformans with overexpression of the AFR1 gene had significantly increased virulence concurrently with resistance (Sanguinetti et al., 2006). In the present study, clinical isolates of C. neoformans developed resistance to FLU possibly during patient therapy and virulence profiles were inversely proportional to the resistance, suggesting that adaptation to the selective pressure of the drug can lead to a decrease in virulence. We also found that the virulence of the C. neoformans 30R isolate is dependent on the concentration of inoculum, which was not observed in the isolate of C. gattii 118R.
Some studies show that environmental isolates of C. gattii have a low susceptibility to FLU (Chowdhary et al., 2011), however, the influence of FLU resistance on virulence in this species is not known. Varma and Kwon-Chung (2010) reported that C. gattii isolates showed heteroresistance to FLU, but the virulence was similar to those observed in isolates of C. neoformans. The C. gattii isolates in our study presented a low virulence profile in G. mellonella, regardless of sensitivity to FLU.
Confirming our results, recently, Santos et al. (2014), through the induction of FLU resistance in C. gattii isolates, observed that after the resistance induction, the virulence was diminished even in vivo as in vitro experiments. This low virulence profile was associated with the low activity of laccase and urease, a decrease in diameter and poor capsule formation (Santos et al., 2014).
Currently, it is known that morphological changes can also be considered as a virulence factor because the pathogen can evade immune recognition of the host and spread infection (Coelho et al., 2014). Nosanchuk et al. (1999) presented results showing the influence of Amb and FLU in cell morphology in C. neoformans. Using scanning electron microscopy, they found that cells cultured in medium with Amb or FLU in inhibitory concentrations showed a modified capsular appearance and were lower smaller when compared with cells cultured without these antifungals. Other studies have demonstrated that the occurrence of pseudo hyphae in C. neoformans increases its resistance to the environment but decreases its virulence in vivo (Magditch et al., 2012). Another type of morphological change is the appearance of C. neoformans giant cells, and several studies have demonstrated that this phenomenon occurs both in vitro and in vivo (Okagaki et al., 2010; Zaragoza et al., 2010; García-Rodas et al., 2011). In our results, we observed that both changes in cell size and in capsule size were detected in the majority of isolates, especially after in vivo infection. However, the increase in cell size of the C. gattii isolates (ATCC 56990 and CL) did not change the virulence, probably because the increased size of these cells was decisive for faster recognition of the immune system of G. mellonella, and the infection was contained. The FLU-susceptible C. neoformans isolate 26S was more virulent in G. mellonella, probably because of capsule increase after in vivo infection and efficient growth at 37°C. Another example of the adaptation of the fungus to high drug concentrations leading to a less virulent in vivo profile is paradoxical growth. This phenomenon is the ability of C. albicans cells to grow at high equinocandin concentrations while still fully susceptible to intermediate concentrations (Stevens et al., 2004). A study by Rueda et al. (2014) showed that this phenomenon can be an adjustment mechanism of fungus to high concentrations of caspofungin, leading to morphological changes and the rapid recognition by the immune system of the larvae. The cells maintained in high concentrations of caspofungin, influenced directly in the virulence in G. mellonella.
The growth at host body temperature is a requirement for virulence (Perfect, 2006), and in C. neoformans, the thermotolerance may have a disproportionate importance over any other virulence factor. When comparing C. neoformans with other species, many factors involved in virulence, such as the capsule and the production of laccase, make these species non-pathogenic because they cannot grow at the temperature of the host (Petter et al., 2001). Garcia-Solache et al. (2013) found that non-pathogenic strains of Cryptococcus were temperature-intolerant, and greater virulence in G. mellonella depended on cell concentration, whereas for C. neoformans var. grubii, the virulence increased at higher temperatures, showing that the rapid replication at 32°C can be responsible for increased virulence. The increased virulence of C. neoformans var. grubii at higher temperatures than the optimal growth temperature, could be a consequence of an increase in the stress response of the fungus, making it better able to survive in hostile conditions (Brown et al., 2007). Analyzing the growth curves of the isolates of C. neoformans showed that at 37°C, the FLU-resistant isolate grew less when compared to FLU-susceptible isolate, suggesting that, the resistance to FLU can somehow affect the growth at 37°C these isolates. In C. gattii the same was observed, but only in relation to isolate CL (sensitive FLU), because the ATCC 56990 strain was unable to grow at this temperature. The lack of growth at 37°C combined with the significant increase of cells upon infection in G. mellonella, may explain the virulence profile presented by the ATCC56990 strain, and low phagocytosis index. Further studies are necessary to confirm this phenotype.
Its known that genotypic differences can influence not only the virulence pattern of isolated, but in the sensitivity to azoles and clinical manifestations developed by pathogens. The phenotype observed in strain ATCC 56990 (AFLP4/VGI) can be related to genotype, however, more studies are needed to confirm this hypothesis, once differences in growth patterns, virulence and sensitivity are not linked only to genotypes and involves multiple factors. In recent study by Firacative et al. (2014) they observed that the virulence is not specifically related to a large molecular type of C. gattii, but with individual attributes.
Correlations between C. gattii molecular type and virulence pattern have been performed (Kidd et al., 2004; D’Souza et al., 2011). However, evaluations of virulence profile, in a global manner, cannot produce consistent results, suggesting that the molecular type is not crucial to the virulence of the strain (Chen et al., 2014).
The size of infectious particles can also influence virulence in G. mellonella. External particles are recognized by hemocytes, and at this stage, the result of the cellular immune response depends on the size of the particles. Small targets are phagocytosed while large targets are encapsulated and attacked by hemocytes (Lavine and Strand, 2002). Melanin plays an important role in the defense of G. mellonella and other invertebrate organisms. The humoral response in G. mellonella is involved in the production of various molecules with antimicrobial properties, including phenol oxidase enzyme (Eisenman et al., 2014). Melanization is a reaction catalyzed by the enzyme phenol oxidase, and it encapsulates foreign particles surrounding G. mellonella (Bidla et al., 2009). In our study, we observed that larvae melanization occurs when infected with C. gattii isolates in the first 24 h and did not occur after infection with C. neoformans isolates. These results are in agreement with the results found by Trevijano-Contador et al. (2015). These results suggest that recognition of isolated C. gattii cells by the immune system was more efficient than C. neoformans, probably due to cell size, increasing the survival of larvae. Scorzoni et al. (2013) found that melanization after infection with C. krusei was significant in G. mellonella and that the degree of the melanization was dependent on inoculum concentration and not on cell viability, indicating that melanization is a non-specific process that depends on the presence of foreign particles.
Our results also demonstrated that FLU-suceptible isolates of C. neoformans were more phagocytized after infection in G. mellonella and were still more virulent when these were compared with the FLU resistant isolates. These data suggest that our isolates can multiply within hemocytes, continuing infection after internalization because sensitive strains possess this ability (Santos et al., 2014). It seems that C. neoformans does not cause a reduction in the number of hemocytes in the first 2 h after infection (Mylonakis et al., 2005; García-Rodas et al., 2011), and this phenomenon may be related to the fact that this fungus is a facultative intracellular pathogen and can survive in phagocytic cells without affecting their viability (Feldmesser et al., 2001; García-Rodas and Zaragoza, 2012). C. neoformans behaves as a facultative intracellular pathogen in mammals and in other non-mammalian hosts (Diamond and Bennett, 1973; Steenbergen et al., 2001; Alvarez and Casadevall, 2006; Ma et al., 2006; Qin et al., 2011), and recently, Trevijano-Contador et al. (2015) demonstrated that the fungus is also able to multiply within hemocytes, suggesting that this ability may be another virulence factor. The FLU sensitive isolates of C. gattii were less phagocytosed when compared to the same drug resistant isolate, probably due to the increase of the capsule and the cell body as observed after infection in G. mellonella.
Fluconazole and other azole antifungal agents are widely used to prevent and treat infections caused by C. neoformans and C. gattii. Their proven efficacy and safety combined with their excellent pharmacokinetic profiles make these agents extremely important in the management of cryptococcal infections (Sanguinetti et al., 2006), but the development of resistance mechanisms and the low susceptibility of some isolates may make treatment difficult to maintain, mainly in countries where the mortality rates for these etiologic agents are high.
There are clinical differences between infections of C. neoformans and C. gattii, and treatment for longer periods in patients affected by C. gattii species led better results (Marr, 2011; Ngamskulrungroj et al., 2012; Chen et al., 2013). Therefore, understanding the mechanisms responsible for these phenotypic and clinical differences is extremely important because the expression of different virulence factors associated with each species is still poorly understood. Virulence studies conducted mainly with C. neoformans are essential, but it is difficult to extrapolate results from these tightly controlled studies to clinical isolates where multiple virulence determinants are expressed in various quantities in a coordinated and potentially host-dependent manner (Clancy et al., 2006). Cryptococcosis remains a difficult management issue, with little new drug development or recent definitive studies (Perfect et al., 2010).
How FLU resistance influences virulence is a complex issue that needs further study. However, the data obtained in this study clearly demonstrate that the adaptation of the fungus to the stress produced by the drug leads to loss of virulence, and morphological changes are involved in the production of this phenotype.
Author Contributions
SR, OZ, and AF-A conceived and designed the experiments. SR, NT-C, LS, AM-A, and HdO performed the experiments. SR, LS, HdO, OZ, MM-G, and AF-A analyzed the data. SR, NT-C, LS, AM-A, HdO, KW, TdFR, MM-G, OZ, and AF-A drafted the manuscript. All authors read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the Programa Institucional de Doutorado Sanduíche no Exterior from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (PDSE-CAPES – 462 1909-13-4), Rede Nacional de Métodos Alternativos – Conselho Nacional Científico e Tecnológico (RENAMA-CNPq 403586/2012-7) and Programa de Apoio ao Desenvolvimento Científico da Faculdade de Ciências Farmacêuticas da Unesp (PADC/FCF). OZ is funded by grant SAF2011-25140 by the Spanish Ministry for Economics and Competitivity.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00153
References
Alspaugh, J. A. (2014). Virulence mechanisms and Cryptococcus neoformans pathogenesis. Fungal Genet. Biol. 78, 55–58. doi: 10.1016/j.fgb.2014.09.004
Alvarez, M., and Casadevall, A. (2006). Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 16, 2161–2165. doi: 10.1016/j.cub.2006.09.061
Andersson, D. I., and Hughes, D. (2010). Antibiotic resistance and its cost: is it possible to reverse resistance? Nat. Rev. Microbiol. 8, 260–271. doi: 10.1038/nrmicro2319
Andersson, D. I., and Levin, B. R. (1999). The biological cost of antibiotic resistance. Curr. Opin. Microbiol. 2, 489–493. doi: 10.1016/S1369-5274(99)00005-3
Barreto de Oliveira, M. T., Boekhout, T., Theelen, B., Hagen, F., Baroni, F. A., Lazera, M. S., et al. (2004). Cryptococcus neoformans shows a remarkable genotypic diversity in Brazil. J. Clin. Microbiol. 42, 1356–1359. doi: 10.1128/JCM.42.3.1356-1359.2004
Bicanic, T., Meintjes, G., Rebe, K., Williams, A., Loyse, A., Wood, R., et al. (2009). Immune reconstitution inflammatory syndrome in HIV-associated cryptococcal meningitis: a prospective study. J. Acquir. Immune Defic. Syndr. 51, 130–134. doi: 10.1097/QAI.0b013e3181a56f2e
Bidla, G., Hauling, T., Dushay, M. S., and Theopold, U. (2009). Activation of insect phenoloxidase after injury: endogenous versus foreign elicitors. J. Innate Immun. 1, 301–308. doi: 10.1159/000168009
Boekhout, T., Theelen, B., Diaz, M., Fell, J. W., Hop, W. C., Abeln, E. C., et al. (2001). Hybrid genotypes in the pathogenic yeast Cryptococcus neoformans. Microbiology 147, 891–907. doi: 10.1099/00221287-147-4-891
Boekhout, T., Van Belkum, A., Leenders, A. C., Verbrugh, H. A., Mukamurangwa, P., Swinne, D., et al. (1997). Molecular typing of Cryptococcus neoformans: taxonomic and epidemiological aspects. Int. J. Syst. Bacteriol. 47, 432–442. doi: 10.1099/00207713-47-2-432
Bovers, M., Hagen, F., and Boekhout, T. (2008a). Diversity of the Cryptococcus neoformans-Cryptococcus gattii species complex. Rev. Iberoam. Micol. 25, S4–S12. doi: 10.1016/S1130-1406(08)70019-6
Bovers, M., Hagen, F., Kuramae, E. E., and Boekhout, T. (2008b). Six monophyletic lineages identified within Cryptococcus neoformans and Cryptococcus gattii by multi-locus sequence typing. Fungal Genet. Biol. 45, 400–421. doi: 10.1016/j.fgb.2007.12.004
Brandt, M. E., Pfaller, M. A., Hajjeh, R. A., Hamill, R. J., Pappas, P. G., Reingold, A. L., et al. (2001). Trends in antifungal drug susceptibility of Cryptococcus neoformans isolates in the United States: 1992 to 1994 and 1996 to 1998. Antimicrob. Agents Chemother. 45, 3065–3069. doi: 10.1128/AAC.45.11.3065-3069.2001
Brennan, M., Thomas, D. Y., Whiteway, M., and Kavanagh, K. (2002). Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med. Microbiol. 34, 153–157. doi: 10.1111/j.1574-695X.2002.tb00617.x
Brown, S. M., Campbell, L. T., and Lodge, J. K. (2007). Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 10, 320–325. doi: 10.1016/j.mib.2007.05.014
Byrnes, E. J., and Heitman, J. (2009). Cryptococcus gattii outbreak expands into the Northwestern United States with fatal consequences. F1000 Biol. Rep. 1, 62. doi: 10.3410/B1-62
Byrnes, E. J., Li, W., Lewit, Y., Ma, H., Voelz, K., Ren, P., et al. (2010). Emergence and pathogenicity of highly virulent Cryptococcus gattii genotypes in the northwest United States. PLoS Pathog. 6:e1000850. doi: 10.1371/journal.ppat.1000850
Casadevall, A., and Pirofski, L. A. (1999). Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect. Immun. 67, 3703–3713.
Casadevall, A., and Pirofski, L. A. (2003). Microbial virulence results from the interaction between host and microorganism. Trends Microbiol. 11, 157–158; author reply 158–159. doi: 10.1016/S0966-842X(03)00008-8
Chau, T. T., Mai, N. H., Phu, N. H., Nghia, H. D., Chuong, L. V., Sinh, D. X., et al. (2010). A prospective descriptive study of cryptococcal meningitis in HIV uninfected patients in Vietnam – high prevalence of Cryptococcus neoformans var grubii in the absence of underlying disease. BMC Infect. Dis. 10:199. doi: 10.1186/1471-2334-10-199
Cheema, M. S., and Christians, J. K. (2011). Virulence in an insect model differs between mating types in Aspergillus fumigatus. Med. Mycol. 49, 202–207. doi: 10.3109/13693786.2010.512301
Chen, S. C., Korman, T. M., Slavin, M. A., Marriott, D., Byth, K., Bak, N., et al. (2013). Antifungal therapy and management of complications of cryptococcosis due to Cryptococcus gattii. Clin. Infect. Dis. 57, 543–551. doi: 10.1093/cid/cit341
Chen, S. C., Meyer, W., and Sorrell, T. C. (2014). Cryptococcus gattii infections. Clin. Microbiol. Rev. 27, 980–1024. doi: 10.1128/CMR.00126-13
Chong, H. S., Dagg, R., Malik, R., Chen, S., and Carter, D. (2010). In vitro susceptibility of the yeast pathogen Cryptococcus to fluconazole and other azoles varies with molecular genotype. J. Clin. Microbiol. 48, 4115–4120. doi: 10.1128/JCM.01271-10
Chowdhary, A., Randhawa, H. S., Sundar, G., Kathuria, S., Prakash, A., Khan, Z., et al. (2011). In vitro antifungal susceptibility profiles and genotypes of 308 clinical and environmental isolates of Cryptococcus neoformans var. grubii and Cryptococcus gattii serotype B from north-western India. J. Med. Microbiol. 60, 961–967. doi: 10.1099/jmm.0.029025-0
Chrisman, C. J., Albuquerque, P., Guimaraes, A. J., Nieves, E., and Casadevall, A. (2011). Phospholipids trigger Cryptococcus neoformans capsular enlargement during interactions with amoebae and macrophages. PLoS Pathog. 7:e1002047. doi: 10.1371/journal.ppat.1002047
Clancy, C. J., Nguyen, M. H., Alandoerffer, R., Cheng, S., Iczkowski, K., Richardson, M., et al. (2006). Cryptococcus neoformans var. grubii isolates recovered from persons with AIDS demonstrate a wide range of virulence during murine meningoencephalitis that correlates with the expression of certain virulence factors. Microbiology 152, 2247–2255. doi: 10.1099/mic.0.28798-0
Coelho, C., Bocca, A. L., and Casadevall, A. (2014). The tools for virulence of Cryptococcus neoformans. Adv. Appl. Microbiol. 87, 1–41. doi: 10.1016/B978-0-12-800261-2.00001-3
Cowen, L. E., Sanglard, D., Calabrese, D., Sirjusingh, C., Anderson, J. B., and Kohn, L. M. (2000). Evolution of drug resistance in experimental populations of Candida albicans. J. Bacteriol. 182, 1515–1522. doi: 10.1128/JB.182.6.1515-1522.2000
Cowen, L. E., Singh, S. D., Köhler, J. R., Collins, C., Zaas, A. K., Schell, W. A., et al. (2009). Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. U.S.A. 106, 2818–2823. doi: 10.1073/pnas.0813394106
Diamond, R. D., and Bennett, J. E. (1973). Growth of Cryptococcus neoformans within human macrophages in vitro. Infect. Immun. 7, 231–236.
D’Souza, C. A., Kronstad, J. W., Taylor, G., Warren, R., Yuen, M., Hu, G., et al. (2011). Genome variation in Cryptococcus gattii, an emerging pathogen of immunocompetent hosts. MBio 2, e00342-10. doi: 10.1128/mBio.00342-10
Eisenman, H. C., Duong, R., Chan, H., Tsue, R., and Mcclelland, E. E. (2014). Reduced virulence of melanized Cryptococcus neoformans in Galleria mellonella. Virulence 5, 611–618. doi: 10.4161/viru.29234
Espinel-Ingroff, A., Aller, A. I., Canton, E., Castañón-Olivares, L. R., Chowdhary, A., Cordoba, S., et al. (2012a). Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voriconazole. Antimicrob. Agents Chemother. 56, 5898–5906. doi: 10.1128/AAC.01115-12
Espinel-Ingroff, A., Chowdhary, A., Cuenca-Estrella, M., Fothergill, A., Fuller, J., Hagen, F., et al. (2012b). Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for amphotericin B and flucytosine. Antimicrob. Agents Chemother. 56, 3107–3113. doi: 10.1128/AAC.06252-11
Fedhila, S., Buisson, C., Dussurget, O., Serror, P., Glomski, I. J., Liehl, P., et al. (2010). Comparative analysis of the virulence of invertebrate and mammalian pathogenic bacteria in the oral insect infection model Galleria mellonella. J. Invertebr. Pathol. 103, 24–29. doi: 10.1016/j.jip.2009.09.005
Feldmesser, M., Tucker, S., and Casadevall, A. (2001). Intracellular parasitism of macrophages by Cryptococcus neoformans. Trends Microbiol. 9, 273–278. doi: 10.1016/S0966-842X(01)02035-2
Fera, M. T., La Camera, E., and De Sarro, A. (2009). New triazoles and echinocandins: mode of action, in vitro activity and mechanisms of resistance. Expert Rev. Anti Infect. Ther. 7, 981–998. doi: 10.1586/eri.09.67
Firacative, C., Duan, S., and Meyer, W. (2014). Galleria mellonella model identifies highly virulent strains among all major molecular types of Cryptococcus gattii. PLoS ONE 9:e105076. doi: 10.1371/journal.pone.0105076
Franzot, S. P., Salkin, I. F., and Casadevall, A. (1999). Cryptococcus neoformans var. grubii: separate varietal status for Cryptococcus neoformans serotype A isolates. J. Clin. Microbiol. 37, 838–840.
Fraser, J. A., Giles, S. S., Wenink, E. C., Geunes-Boyer, S. G., Wright, J. R., Diezmann, S., et al. (2005). Same-sex mating and the origin of the Vancouver Island Cryptococcus gattii outbreak. Nature 437, 1360–1364. doi: 10.1038/nature04220
García-Rodas, R., Casadevall, A., Rodríguez-Tudela, J. L., Cuenca-Estrella, M., and Zaragoza, O. (2011). Cryptococcus neoformans capsular enlargement and cellular gigantism during Galleria mellonella infection. PLoS ONE 6:e24485. doi: 10.1371/journal.pone.0024485
García-Rodas, R., and Zaragoza, O. (2012). Catch me if you can: phagocytosis and killing avoidance by Cryptococcus neoformans. FEMS Immunol. Med. Microbiol. 64, 147–161. doi: 10.1111/j.1574-695X.2011.00871.x
Garcia-Solache, M. A., Izquierdo-Garcia, D., Smith, C., Bergman, A., and Casadevall, A. (2013). Fungal virulence in a lepidopteran model is an emergent property with deterministic features. MBio 4, e00100-13. doi: 10.1128/mBio.00100-13
Giles, S. S., Dagenais, T. R., Botts, M. R., Keller, N. P., and Hull, C. M. (2009). Elucidating the pathogenesis of spores from the human fungal pathogen Cryptococcus neoformans. Infect. Immun. 77, 3491–3500. doi: 10.1128/IAI.00334-09
Gomez-Lopez, A., Forastiero, A., Cendejas-Bueno, E., Gregson, L., Mellado, E., Howard, S. J., et al. (2014). An invertebrate model to evaluate virulence in Aspergillus fumigatus: the role of azole resistance. Med. Mycol. 52, 311–319. doi: 10.1093/mmy/myt022
Hagen, F., Colom, M. F., Swinne, D., Tintelnot, K., Iatta, R., Montagna, M. T., et al. (2012). Autochthonous and dormant Cryptococcus gattii infections in Europe. Emerg. Infect. Dis. 18, 1618–1624. doi: 10.3201/eid1810.120068
Hagen, F., Illnait-Zaragozi, M. T., Bartlett, K. H., Swinne, D., Geertsen, E., Klaassen, C. H., et al. (2010). In vitro antifungal susceptibilities and amplified fragment length polymorphism genotyping of a worldwide collection of 350 clinical, veterinary, and environmental Cryptococcus gattii isolates. Antimicrob. Agents Chemother. 54, 5139–5145. doi: 10.1128/AAC.00746-10
Hagen, F., Khayhan, K., Theelen, B., Kolecka, A., Polacheck, I., Sionov, E., et al. (2015). Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet. Biol. 78, 16–48. doi: 10.1016/j.fgb.2015.02.009
Harris, J. R., Lockhart, S. R., Debess, E., Marsden-Haug, N., Goldoft, M., Wohrle, R., et al. (2011). Cryptococcus gattii in the United States: clinical aspects of infection with an emerging pathogen. Clin. Infect. Dis. 53, 1188–1195. doi: 10.1093/cid/cir723
Jacobsen, I. D. (2014). Galleria mellonella as a model host to study virulence of Candida. Virulence 5, 237–239. doi: 10.4161/viru.27434
Jobbins, S. E., Hill, C. J., D’souza-Basseal, J. M., Padula, M. P., Herbert, B. R., and Krockenberger, M. B. (2010). Immunoproteomic approach to elucidating the pathogenesis of cryptococcosis caused by Cryptococcus gattii. J. Proteome Res. 9, 3832–3841. doi: 10.1021/pr100028t
Kessler, A. T., Al Kharrat, T., and Kourtis, A. P. (2010). Cryptococcus neoformans as a cause of bronchiolitis obliterans organizing pneumonia. J. Infect. Chemother. 16, 206–209. doi: 10.1007/s10156-010-0039-7
Kidd, S. E., Hagen, F., Tscharke, R. L., Huynh, M., Bartlett, K. H., Fyfe, M., et al. (2004). A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. U.S.A. 101, 17258–17263. doi: 10.1073/pnas.0402981101
Kwon-Chung, K. J., and Varma, A. (2006). Do major species concepts support one, two or more species within Cryptococcus neoformans? FEMS Yeast Res. 6, 574–587. doi: 10.1111/j.1567-1364.2006.00088.x
Lavine, M. D., and Strand, M. R. (2002). Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 32, 1295–1309. doi: 10.1016/S0965-1748(02)00092-9
Levin, B. R., Lipsitch, M., Perrot, V., Schrag, S., Antia, R., Simonsen, L., et al. (1997). The population genetics of antibiotic resistance. Clin. Infect. Dis. 24(Suppl. 1), S9–S16. doi: 10.1093/clinids/24.Supplement_1.S9
Li, S. S., and Mody, C. H. (2010). Cryptococcus. Proc. Am. Thorac. Soc. 7, 186–196. doi: 10.1513/pats.200907-063AL
Litvintseva, A. P., Thakur, R., Vilgalys, R., and Mitchell, T. G. (2006). Multilocus sequence typing reveals three genetic subpopulations of Cryptococcus neoformans var. grubii (serotype A), including a unique population in Botswana. Genetics 172, 2223–2238. doi: 10.1534/genetics.105.046672
Ma, H., Croudace, J. E., Lammas, D. A., and May, R. C. (2006). Expulsion of live pathogenic yeast by macrophages. Curr. Biol. 16, 2156–2160. doi: 10.1016/j.cub.2006.09.032
Magditch, D. A., Liu, T. B., Xue, C., and Idnurm, A. (2012). DNA mutations mediate microevolution between host-adapted forms of the pathogenic fungus Cryptococcus neoformans. PLoS Pathog. 8:e1002936. doi: 10.1371/journal.ppat.1002936
Marr, K. A. (2011). Cryptococcus gattii: the tip of the iceberg. Clin. Infect. Dis. 53, 1196–1198. doi: 10.1093/cid/cir738
Martinez, L. R., and Casadevall, A. (2006). Susceptibility of Cryptococcus neoformans biofilms to antifungal agents in vitro. Antimicrob. Agents Chemother. 50, 1021–1033. doi: 10.1128/AAC.50.3.1021-1033.2006
Mesa-Arango, A. C., Forastiero, A., Bernal-Martínez, L., Cuenca-Estrella, M., Mellado, E., and Zaragoza, O. (2012). The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol. 51, 461–472. doi: 10.3109/13693786.2012.737031
Meyer, W., Aanensen, D. M., Boekhout, T., Cogliati, M., Diaz, M. R., Esposto, M. C., et al. (2009). Consensus multi-locus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med. Mycol. 47, 561–570. doi: 10.1080/13693780902953886
Muhammed, M., Fuchs, B. B., Wu, M. P., Breger, J., Coleman, J. J., and Mylonakis, E. (2012). The role of mycelium production and a MAPK-mediated immune response in the C. elegans-Fusarium model system. Med. Mycol. 50, 488–496. doi: 10.3109/13693786.2011.648217
Mylonakis, E., and Aballay, A. (2005). Worms and flies as genetically tractable animal models to study host-pathogen interactions. Infect. Immun. 73, 3833–3841. doi: 10.1128/IAI.73.7.3833-3841.2005
Mylonakis, E., Moreno, R., El Khoury, J. B., Idnurm, A., Heitman, J., Calderwood, S. B., et al. (2005). Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 73, 3842–3850. doi: 10.1128/IAI.73.7.3842-3850.2005
Negroni, R. (2012). Cryptococcosis. Clin. Dermatol. 30, 599–609. doi: 10.1016/j.clindermatol.2012.01.005
Ngamskulrungroj, P., Chang, Y., Sionov, E., and Kwon-Chung, K. J. (2012). The primary target organ of Cryptococcus gattii is different from that of Cryptococcus neoformans in a murine model. MBio 3, e00103-12. doi: 10.1128/mBio.00103-12
Nosanchuk, J. D., Cleare, W., Franzot, S. P., and Casadevall, A. (1999). Amphotericin B and fluconazole affect cellular charge, macrophage phagocytosis, and cellular morphology of Cryptococcus neoformans at subinhibitory concentrations. Antimicrob. Agents Chemother. 43, 233–239.
Okagaki, L. H., Strain, A. K., Nielsen, J. N., Charlier, C., Baltes, N. J., Chrétien, F., et al. (2010). Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 6:e1000953. doi: 10.1371/journal.ppat.1000953
Okawa, Y., Miyauchi, M., and Kobayashi, H. (2008). Comparison of pathogenicity of various Candida tropicalis strains. Biol. Pharm. Bull. 31, 1507–1510. doi: 10.1248/bpb.31.1507
Oliveira Carvalho, V., Okay, T. S., Melhem, M. S., Walderez Szeszs, M., and Del Negro, G. M. (2013). The new mutation L321F in Candida albicans ERG11 gene may be associated with fluconazole resistance. Rev. Iberoam. Micol. 30, 209–212. doi: 10.1016/j.riam.2013.01.001
Park, B. J., Wannemuehler, K. A., Marston, B. J., Govender, N., Pappas, P. G., and Chiller, T. M. (2009). Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 23, 525–530. doi: 10.1097/QAD.0b013e328322ffac
Perfect, J. R. (2006). Cryptococcus neoformans: the yeast that likes it hot. FEMS Yeast Res. 6, 463–468. doi: 10.1111/j.1567-1364.2006.00051.x
Perfect, J. R., Dismukes, W. E., Dromer, F., Goldman, D. L., Graybill, J. R., Hamill, R. J., et al. (2010). Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 50, 291–322. doi: 10.1086/649858
Petter, R., Kang, B. S., Boekhout, T., Davis, B. J., and Kwon-Chung, K. J. (2001). A survey of heterobasidiomycetous yeasts for the presence of the genes homologous to virulence factors of Filobasidiella neoformans, CNLAC1 and CAP59. Microbiology 147, 2029–2036. doi: 10.1099/00221287-147-8-2029
Pfaller, M. A., Diekema, D. J., Gibbs, D. L., Newell, V. A., Bijie, H., Dzierzanowska, D., et al. (2009). Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: 10.5-year analysis of susceptibilities of noncandidal yeast species to fluconazole and voriconazole determined by CLSI standardized disk diffusion testing. J. Clin. Microbiol. 47, 117–123. doi: 10.1128/JCM.01747-08
Posteraro, B., Sanguinetti, M., Sanglard, D., La Sorda, M., Boccia, S., Romano, L., et al. (2003). Identification and characterization of a Cryptococcus neoformans ATP binding cassette (ABC) transporter-encoding gene, CnAFR1, involved in the resistance to fluconazole. Mol. Microbiol. 47, 357–371. doi: 10.1046/j.1365-2958.2003.03281.x
Qin, Q. M., Luo, J., Lin, X., Pei, J., Li, L., Ficht, T. A., et al. (2011). Functional analysis of host factors that mediate the intracellular lifestyle of Cryptococcus neoformans. PLoS Pathog. 7:e1002078. doi: 10.1371/journal.ppat.1002078
Ramarao, N., Nielsen-Leroux, C., and Lereclus, D. (2012). The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. J. Vis. Exp. 70, e4392. doi: 10.3791/4392
Raso, T. F., Werther, K., Miranda, E. T., and Mendes-Giannini, M. J. (2004). Cryptococcosis outbreak in psittacine birds in Brazil. Med. Mycol. 42, 355–362. doi: 10.1080/13693780410001712061
Rodero, L., Mellado, E., Rodriguez, A. C., Salve, A., Guelfand, L., Cahn, P., et al. (2003). G484S amino acid substitution in lanosterol 14-alpha demethylase (ERG11) is related to fluconazole resistance in a recurrent Cryptococcus neoformans clinical isolate. Antimicrob. Agents Chemother. 47, 3653–3656. doi: 10.1128/AAC.47.11.3653-3656.2003
Rueda, C., Cuenca-Estrella, M., and Zaragoza, O. (2014). Paradoxical growth of Candida albicans in the presence of caspofungin is associated with multiple cell wall rearrangements and decreased virulence. Antimicrob. Agents Chemother. 58, 1071–1083. doi: 10.1128/AAC.00946-13
Sanglard, D. (2002). Resistance of human fungal pathogens to antifungal drugs. Curr. Opin. Microbiol. 5, 379–385. doi: 10.1016/S1369-5274(02)00344-2
Sanglard, D., Coste, A., and Ferrari, S. (2009). Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res. 9, 1029–1050. doi: 10.1111/j.1567-1364.2009.00578.x
Sanglard, D., Ischer, F., Koymans, L., and Bille, J. (1998). Amino acid substitutions in the cytochrome P-450 lanosterol 14alpha-demethylase (CYP51A1) from azole-resistant Candida albicans clinical isolates contribute to resistance to azole antifungal agents. Antimicrob. Agents Chemother. 42, 241–253.
Sanguinetti, M., Posteraro, B., La Sorda, M., Torelli, R., Fiori, B., Santangelo, R., et al. (2006). Role of AFR1, an ABC transporter-encoding gene, in the in vivo response to fluconazole and virulence of Cryptococcus neoformans. Infect. Immun. 74, 1352–1359. doi: 10.1128/IAI.74.2.1352-1359.2006
Santos, J. R., Holanda, R. A., Frases, S., Bravim, M., Araujo, G. E. S., Santos, P. C., et al. (2014). Fluconazole alters the polysaccharide capsule of Cryptococcus gattii and leads to distinct behaviors in murine Cryptococcosis. PLoS ONE 9:e112669. doi: 10.1371/journal.pone.0112669
Schulz, B., Weber, K., Schmidt, A., Borg-Von Zepelin, M., and Ruhnke, M. (2011). Difference in virulence between fluconazole-susceptible and fluconazole-resistant Candida albicans in a mouse model. Mycoses 54, e522–e530. doi: 10.1111/j.1439-0507.2010.01970.x
Scorzoni, L., De Lucas, M. P., Mesa-Arango, A. C., Fusco-Almeida, A. M., Lozano, E., Cuenca-Estrella, M., et al. (2013). Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 8:e60047. doi: 10.1371/journal.pone.0060047
Sheng, C., Miao, Z., Ji, H., Yao, J., Wang, W., Che, X., et al. (2009). Three-dimensional model of lanosterol 14 alpha-demethylase from Cryptococcus neoformans: active-site characterization and insights into azole binding. Antimicrob. Agents Chemother. 53, 3487–3495. doi: 10.1128/AAC.01630-08
Sionov, E., Chang, Y. C., Garraffo, H. M., and Kwon-Chung, K. J. (2009). Heteroresistance to fluconazole in Cryptococcus neoformans is intrinsic and associated with virulence. Antimicrob. Agents Chemother. 53, 2804–2815. doi: 10.1128/AAC.00295-09
Sionov, E., Lee, H., Chang, Y. C., and Kwon-Chung, K. J. (2010). Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 6:e1000848. doi: 10.1371/journal.ppat.1000848
Spinello, A. (2013). New insights into HIV/AIDS-associated Cryptococcosis. ISRN AIDS 2013, 471363. doi: 10.1155/2013/471363
Steenbergen, J. N., Shuman, H. A., and Casadevall, A. (2001). Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl. Acad. Sci. U.S.A. 98, 15245–15250. doi: 10.1073/pnas.261418798
Stevens, D. A., Espiritu, M., and Parmar, R. (2004). Paradoxical effect of caspofungin: reduced activity against Candida albicans at high drug concentrations. Antimicrob. Agents Chemother. 48, 3407–3411. doi: 10.1128/AAC.48.9.3407-3411.2004
Trevijano-Contador, N., Herrero-Fernández, I., García-Barbazán, I., Scorzoni, L., Rueda, C., Rossi, S. A., et al. (2015). Cryptococcus neoformans induces antimicrobial responses and behaves as a facultative intracellular pathogen in the non mammalian model Galleria mellonella. Virulence 6, 66–74. doi: 10.4161/21505594.2014.986412
Trilles, L., Lazéra, M., Wanke, B., Theelen, B., and Boekhout, T. (2003). Genetic characterization of environmental isolates of the Cryptococcus neoformans species complex from Brazil. Med. Mycol. 41, 383–390. doi: 10.1080/1369378031000137206
Varma, A., and Kwon-Chung, K. J. (2010). Heteroresistance of Cryptococcus gattii to fluconazole. Antimicrob. Agents Chemother. 54, 2303–2311. doi: 10.1128/AAC.00153-10
Wilke, M. (2011). Treatment and prophylaxis of invasive candidiasis with anidulafungin, caspofungin and micafungin and its impact on use and costs: review of the literature. Eur. J. Med. Res. 16, 180–186. doi: 10.1186/2047-783X-16-4-180
Zaragoza, O., and Casadevall, A. (2004). Experimental modulation of capsule size in Cryptococcus neoformans. Biol. Proced. Online 6, 10–15. doi: 10.1251/bpo68
Keywords: Cryptococcus sp., resistance, fluconazole, virulence, Galleria mellonella
Citation: Rossi SA, Trevijano-Contador N, Scorzoni L, Mesa-Arango AC, de Oliveira HC, Werther K, de Freitas Raso T, Mendes-Giannini MJS, Zaragoza O and Fusco-Almeida AM (2016) Impact of Resistance to Fluconazole on Virulence and Morphological Aspects of Cryptococcus neoformans and Cryptococcus gattii Isolates. Front. Microbiol. 7:153. doi: 10.3389/fmicb.2016.00153
Received: 09 November 2015; Accepted: 29 January 2016;
Published: 16 February 2016.
Edited by:
Marcio L. Rodrigues, Oswaldo Cruz Foundation, BrazilReviewed by:
Ferry Hagen, Canisius Wilhelmina Hospital, NetherlandsTaissa Vieira Machado Vila, Federal University of Rio de Janeiro, Brazil
Copyright © 2016 Rossi, Trevijano-Contador, Scorzoni, Mesa-Arango, de Oliveira, Werther, de Freitas Raso, Mendes-Giannini, Zaragoza and Fusco-Almeida. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. Fusco-Almeida, almeidaf@fcfar.unesp.br