 Sergei V. Bukin1*
Sergei V. Bukin1* Olga N. Pavlova1
Olga N. Pavlova1 Andrei Y. Manakov2Elena A. Kostyreva3Svetlana M. Chernitsyna1Elena V. Mamaeva1Tatyana V. Pogodaeva4Tamara I. Zemskaya1
Andrei Y. Manakov2Elena A. Kostyreva3Svetlana M. Chernitsyna1Elena V. Mamaeva1Tatyana V. Pogodaeva4Tamara I. Zemskaya1
- 1Laboratory of Hydrocarbon Microbiology, Limnological Institute, Russian Academy of Science, Irkutsk, Russia
- 2Laboratory of Clathrate Compounds, Nikolaev Institute of Inorganic Chemistry, Russian Academy of Science, Novosibirsk, Russia
- 3Laboratory of Petroleum Geochemistry, Trofimuk Institute of Petroleum Geology and Geophysics, Russian Academy of Science, Novosibirsk, Russia
- 4Laboratory of Hydrochemistry and Atmosphere Chemistry, Limnological Institute, Russian Academy of Science, Irkutsk, Russia
The ability to compare the composition and metabolic potential of microbial communities inhabiting the subsurface sediment in geographically distinct locations is one of the keys to understanding the evolution and function of the subsurface biosphere. Prospective areas for study of the subsurface biosphere are the sites of hydrocarbon discharges on the bottom of the Lake Baikal rift, where ascending fluxes of gas-saturated fluids and oil from deep layers of bottom sediments seep into near-surface sediment. The samples of surface sediments collected in the area of the Posolskaya Bank methane seep were cultured for 17 months under thermobaric conditions (80°C, 5 MPa) with the addition of complementary organic substrate, and a different composition for the gas phase. After incubation, the presence of intact cells of microorganisms, organic matter transformation and the formation of oil biomarkers was confirmed in the samples, with the addition of Baikal diatom alga Synedra acus detritus, and gas mixture CH4:H2:CO2. Taxonomic assignment of the 16S rRNA sequence data indicates that the predominant sequences in the enrichment were Sphingomonas (55.3%), Solirubrobacter (27.5%) and Arthrobacter (16.6%). At the same time, in heat-killed sediment and in sediment without any additional substrates, which were cultivated in a CH4 atmosphere, no geochemical changes were detected, nor the presence of intact cells and 16S rRNA sequences of Bacteria and Archaea. This data may suggest that the decomposition of organic matter under culturing conditions could be performed by microorganisms from low-temperature sediment layers. One possible explanation of this phenomenon is migration of the representatives of the deep thermophilic community through fault zones in the near surface sediment layers, together with gas-bearing fluids.
Introduction
The dark energy biosphere was hypothesized as a suite of habitats or ecosystems that are physically located in environments that exist in permanent darkness, and was proposed in the late 20th century (Gold, 1992). The first data on microbial activity, the total number and diversity of cultured microorganisms in the deep layers of marine sediments and continental crust (Sinclair and Ghiorse, 1989; Cragg et al., 1990; Parkes et al., 1990, 1994; Kieft et al., 1995), obtained by deep drilling, indicated that subsurface biosphere ecosystems are one of the largest microbiological habitats on Earth. To date, Bacteria and Archaea have been identified in ocean sediments at depths of up to 2500 m below sea floor (Roussel et al., 2008; Ciobanu et al., 2014; Inagaki et al., 2015). The direct measurement of cell activity indicates that the microorganisms are metabolically active in deep sediments aged >16 million years (Schippers et al., 2005).
The areas with the greatest abundance and activity of microorganisms in the subsurface biosphere are associated with high concentrations of organic matter and/or the inorganic electron donors and acceptors (D’Hondt et al., 2004; Parkes et al., 2014). An example of such “hotspots” is the tectonically active areas, where the flows of gasses and fluids ascending from deep sediments or the basaltic layer of the Earth’s crust determine the additional supply of carbon and energy (Cowen et al., 2003; Parkes et al., 2005; Engelen et al., 2008; Boetius and Wenzhöfer, 2013). Fluid flows, rich in H2, acetate, and methane, as well as other hydrocarbons formed in the deep sediments as a result of thermogenic activation and the degradation of kerogen (Horsfield et al., 2006; Parkes et al., 2007), may transport the representatives of the deep thermophilic community, like submarine hydrothermal vents (Hubert et al., 2009).
In addition to marine ecosystems, rift lakes, in particular Lake Baikal in Russia, are the prospective places for studying the subsurface biosphere (Biddle et al., 2012). Lake Baikal is the oldest (more than 25 million years) and deepest (maximum depth 1642 m) freshwater lake in the world. Located in a tectonically active zone, the lake has discharges of oil and gas-bearing fluids, mud volcanoes, and gas-hydrate deposits (Kuzmin et al., 2000; Kontorovich et al., 2007; Khlystov et al., 2013). Catagenesis biomarkers in the discharged oil and the isotopic composition of gas indicate that, in some areas of the lake, oil and gas migrate from depths of up to 7 km to the floor of the lake, where the temperature is close to 4°C (Khlystov et al., 2007; Kontorovich et al., 2007). The abnormally high concentrations of sulfates, bicarbonates, chlorides, and the ions of alkali and alkali-earth metals composing the pore waters of surface sediments in discharge areas may be due to the highly mineralized deep fluids passing through them (Granina et al., 2001). Ascending fluid flows also explain the presence, of the valves of ancient diatoms of the genus Tertiarius occurring in areas with calm sedimentation at depths of 300 m, in the upper layers of the bottom sediments of mud volcanoes (Bradbury et al., 1994; Klerkx et al., 2003). Similarly, in areas of Lake Baikal with deep-water discharges, the members of the deep thermophile community transported by the ascending fluid flows can enter the cold surface horizons of bottom sediments.
This study mainly aimed to ascertain possible existence of microorganisms, carrying out the transformation of organic matter under high temperatures typical for the deep horizons of the Earth’s crust, in the near-surface sediments of Lake Baikal. For this reason, we have cultured natural samples of sediments adjacent to the zone of tectonic fault under anaerobic conditions at temperature of 80°C and pressure of 5 MPa. We have also analyzed the microbial community using high-resolution pyrosequencing of the 16S rRNA gene fragments and characterized the processes of transformation of organic matter using gas chromatography mass spectrometry (GC-MS).
Materials and Methods
Site Description and Sampling
This study deals with the microbial community of bottom sediments from the Lake Baikal area of the methane seep Posolskaya Bank (Southern Basin, 52°02′10″ N, 105°50′36″ E, water depth 500 m; Figure 1A). The Posolskaya Bank is a pronounced underwater elevation bordering Southern and Central Basins of the lake, and is a part of the delta flank area of the Selenga River (Bezrukova et al., 2005; Figure 1B). The geological data (Klerkx et al., 2006; Naudts et al., 2012) as well as the analysis of the carbon isotopic composition of the discharged gas (Kalmychkov et al., 2006) show that some gas-saturated fluid flows come from deep layers below the stability zone of gas hydrates.
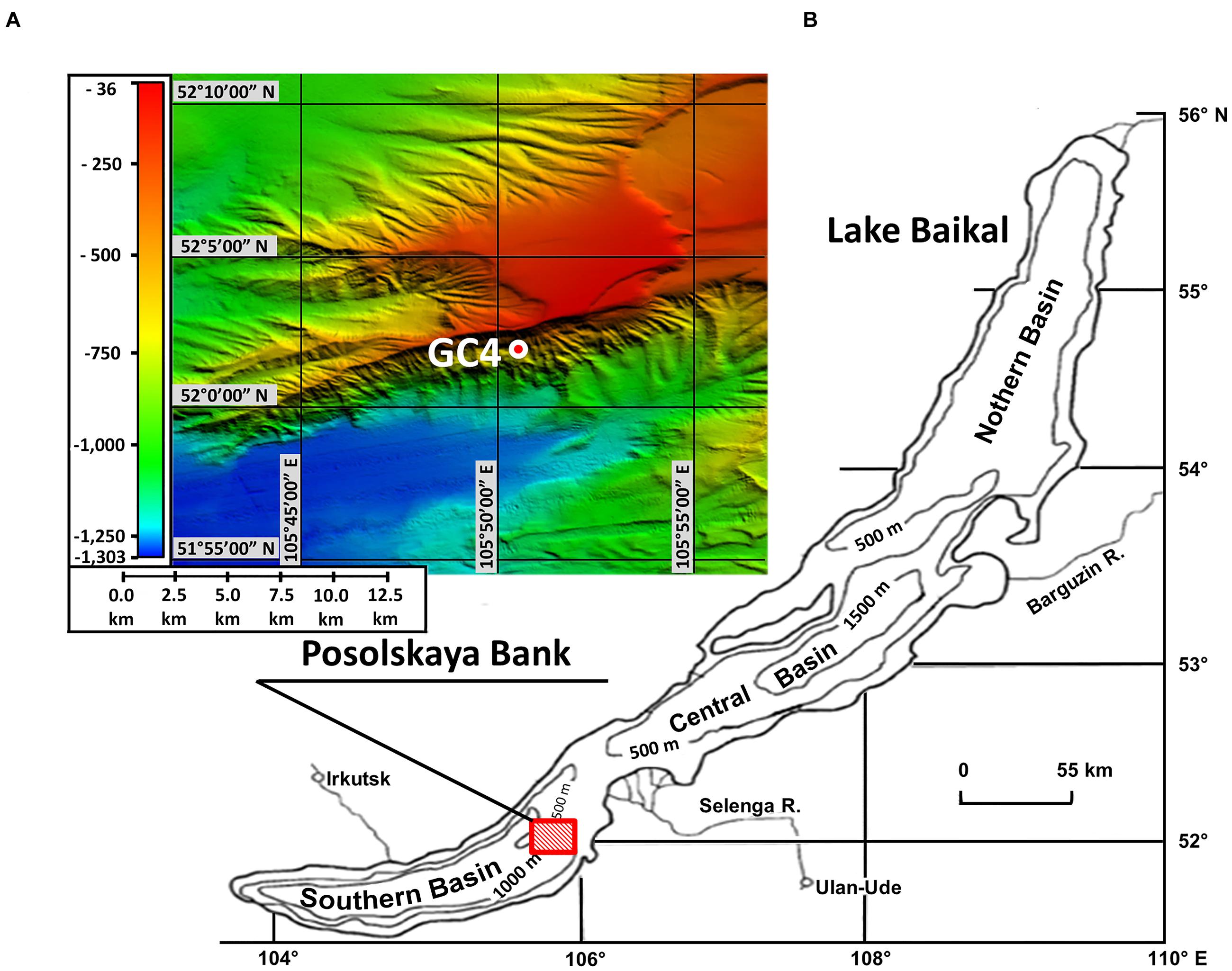
FIGURE 1. Location of sampling area. (A) Bathymetric map. Locations of GC4 core sample examined in this study are indicated with red-white circle. The map was created by Oleg M. Khlystov and Andrei V. Khabuev within the project No. 23.8 of the Program of RAS Presidium and the Fonds Wetenschappelijk Onderzoek project No. 1.5.198.09. (B) Location of underwater elevation Posolskaya Bank on the chart of Lake Baikal.
Bottom sediments were obtained during the expedition in June 2012 using the gravity corer (Ver-2012-06-GC4) from board of the research vessel (RV) G.Y. Vereschagin. After lifting the core on board, the samples of sediments for chemical analysis of pore waters, DNA extraction and experimental procedures (about 600 g) were collected. Sediment samples were taken aseptically from central part of the core with sterile 60-ml syringes (luer end removed). Chemical analysis of pore waters and DNA extraction were carried out on-board immediately after sampling. Sediments selected for the experiment from the 60–100 cm blf (cm below lake floor) layer adjoining the gas-hydrate interlayers were homogenized, divided into three parts (200 g each) and stored into liquid nitrogen (-196°C) during delivery to the laboratory.
Chemical Analysis of Pore Water
Pore water extractions were made from 14 samples (100 g each) collected all along the core. Pore water was extracted through the 20 min sediment centrifugation (initially at 10000 rev/min and subsequently at 14000 rev/min). The anion and cation contents in water were determined according to the methods described previously (Zemskaya et al., 2010).
Experimental Cultivation
Bottom sediments were cultured in three experimental autoclaves (Supplementary Figure S1) at 80°C and 5 MPa under different conditions of gas phase composition and type of nutrient substrate added.
In autoclave #1, the sediment sample was supplemented with detritus of S. acus from the pure culture obtained in a photobioreactor at the Department of Cell Ultrastructure of the Limnological Institute (Shishlyannikov et al., 2011). The culture purity was confirmed by phase-contrast and epifluorescence microscopy, plating on different media (LB, FPA/10, DA) and by PCR for amplification of a bacterial 16S rRNA gene fragment as described (Shishlyannikov et al., 2011). Diatom cells were collected on sterile polycarbonate membrane with a pore size of 1 μm (Millipore, USA), placed in sterile 1.5 ml microtubes (Axygen, USA) and stored at -80°C before experiment. The gas phase in the autoclave #1 consisted of the gas mixture CH4:H2:CO2 (50%:40%:10%). Autoclave #2 contained only the sediment without additional substrates and the sediments from autoclave #3 were pre-sterilized by autoclaving at 135°C for 40 min, and served as a negative control in order to confirm that microorganisms were involved in all studied processes. The gas phase in the autoclaves # 2 and #3 consisted of CH4.
The experiment procedure was as follows: sediment samples (200 g each) were defrosted at room temperature. Sample #1 was homogenized with 5 g of the detritus of S. acus. Sample (#3) was pre-sterilized by autoclaving. Further, 100 g of the each sample were stored into liquid nitrogen and then used for total organic carbon (TOC) analysis prior to the experiment; other halves of the samples (100 g) were placed in 170 ml glass beakers and saturated with sterile Baikal water in the proportion of approximately 1/4 of the sediment volume (in order to avoid drying during cultivation). The beakers were covered with Teflon lids and placed in 400 ml experimental steel autoclaves. All components were sterilized by autoclaving (135°C for 40 min). In order to avoid contamination, all handling was carried out in biosafety cabinet (BMB-II-Laminar-S-1,5, Lamsystems, Russia). The autoclaves were flushed with gas mixture CH4:H2:CO2 or CH4, increasing the pressure up to 4.2 MPa (pressure control was carried out in autoclaves integrated manometers). All gasses were sterilized by filtration. The autoclaves were placed in thermostats at a temperature 80°C where, as a result of the heating, the pressure in the autoclaves was increased up to 5 MPa.
In the autoclaves, the sediments were cultured for 17 months (the maximum period that was technically possible). At the end of the experiment, sediments from each autoclave were homogenized; 95 g of the each sample were stored into liquid nitrogen and then used for molecular genetic studies (20 g) and GC–MS analysis (75 g); 5 g of the each sample were suspended in sterile Milli-Q water and used for microscopic analysis.
Total Organic Carbon
Content and composition of TOC were detected in the samples of bottom sediments before (considering the organic substrates added) and at the end of the experiment. Preparation of the samples, identification of the content of organic carbon (Corg, %), isolation of soluble (bitumoid) organic matter and GC–MS analyses were performed as described previously (Pavlova et al., 2016). All measurements were triplicated.
Microscopy
To detect the viable microorganisms in the samples, we used light and epifluorescence microscopy. For the latter, the smears prepared on glass were dehydrated in a series of solutions with rising concentrations of ethanol (50%→70%→96%, 3 min each); then they were stained with the solutions of 4′,6-diamidino-2-phenylindole (DAPI) or 3-N,3-N,6-N,6-N-tetramethylacridine-3,6-diamine (Acridine orange, AO) and observed using the microscope Axio Imager M1 (ZEISS, Germany).
DNA Extraction
Homogenized natural sediments (10 g) and 10 g each of the three sediments samples subjected to cultivation under thermobaric conditions were used for DNA extraction. DNA was extracted according to phenol-chloroform-isoamyl alcohol method (Sambrook et al., 1989) in a modified version (Shubenkova et al., 2005). Four independent DNA extractions were carried out for each sample. In order to avoid contamination, all handling was carried out in DNA/RNA UV-cleaner box (Biosan, Latvia), using ultraviolet-treated (60 min) plasticware and micropipettes. In addition, a negative control (DNA extraction with sterile TE-buffer) was prepared for each independent DNA extractions, to ensure that no contamination with exogenous amplifiable DNA occurred during the different stages of sample treatment. Concentration and quality of extracted DNA were measured with a spectrophotometer SmartSpec Plus (Bio-Rad, USA).
454 Pyrosequencing and Phylogenetic Analysis
For all sediment samples, pyrosequencing was carried out using libraries of amplicons obtained by PCR with primers 341F (Muyzer et al., 1993) - 785R (Lee et al., 1993) for the V3–V4 region of the bacterial 16S rRNA gene and primers A2Fa (Reysenbach and Pace, 1995) – A519R (Sørensen and Teske, 2006) for the V1–V3 region of archaeal 16S rRNA gene. Three independent PCR mixtures were pooled for each sample to decrease PCR bias. Negative controls (reaction mixture without DNA) were included in each set of PCR reactions. PCR conditions are described in Supplementary Table S1. When direct positive PCR amplifications failed [PCR products were not visible on a 1% (w/v), agarose gel stained with ethidium bromide; for example – experimental sediment samples #2, 3; negative controls of DNA extractions], DNA extracts were additionally amplified with a nested PCR using several primer combinations (Supplementary Table S2).
Sequencing was performed using the genome sequencer GS FLX 454 (Roche, USA) with the Titanium series reagents according to the manufacturer’s recommendations. The obtained readings were analyzed using the software package Mothur 1.31.2 (Schloss et al., 2009). The sequencing errors were removed by the PyroNoise algorithm (Quince et al., 2011) selecting the sequences with the size of more than 150 bp (more than 200 bp for Archaea) and length of homopolymer tracts less than 6 bp which were aligned by archaeal and bacterial 16S rRNA sequences from the SILVA1 database. The NAST algorithm with a k-mer length of 8 bp was employed for sequence alignment. Chimeric sequences were identified by the UCHIME algorithm (Edgar et al., 2011) with standard parameters.
For taxonomic analysis, the obtained readings were classified by the SILVA2 taxonomy at a confidence threshold of 80%. The sequences with 97% similarity (genetic distance 0.03) were grouped into operational taxonomic units (OTU). Sequence clustering was based on unweighted pair group method with arithmetic mean analysis of genetic distance. Rarefaction curves, Good’s coverage, ACE and Chao1 (richness) indices and the inverse Simpson index (diversity) were calculated based on the identified OTUs using the Mothur software package. The OTU0.03 sequences were compared with the known 16S rRNA sequences from NCBI database using the BLASTN3 program with default parameters.
Nucleotide sequences of the 16S rRNA were deposited in NCBI Short Read Archive (SRAid: SRR2912888, SRR2912890, SRR2146995).
Results
Geochemical Characteristics of Sediments Used in the Experiment
The sediment layers (60–100 cm blf) used in the experiment were recovered friable gray aleuropelite silt with many cracks caused by degassing. At depths of 80 cm blf and below, there were gas hydrates forming small and massive layers (Figure 2A). The total number of ions in pore waters (123.06–142.83 mg l-1) was slightly higher than the values in sediments at the reference sites which inherited the chemical composition of the lake water column (Pogodaeva et al., 2007; Figure 2B). We registered high concentrations of acetate ions (up to 22.3 mg l-1) and ammonium ions (up to 22.3 mg l-1) compared to the reference sites. Concentration of sulfate ions (≤0.6 mg l-1) in the layer at 60–100 cm blf was insignificant in comparison with the values in upper sediment layers. Methane concentration in the studied core exceeded 90 μmol dm-3 of raw silt even in the upper layers of sediment (Pimenov et al., 2014).
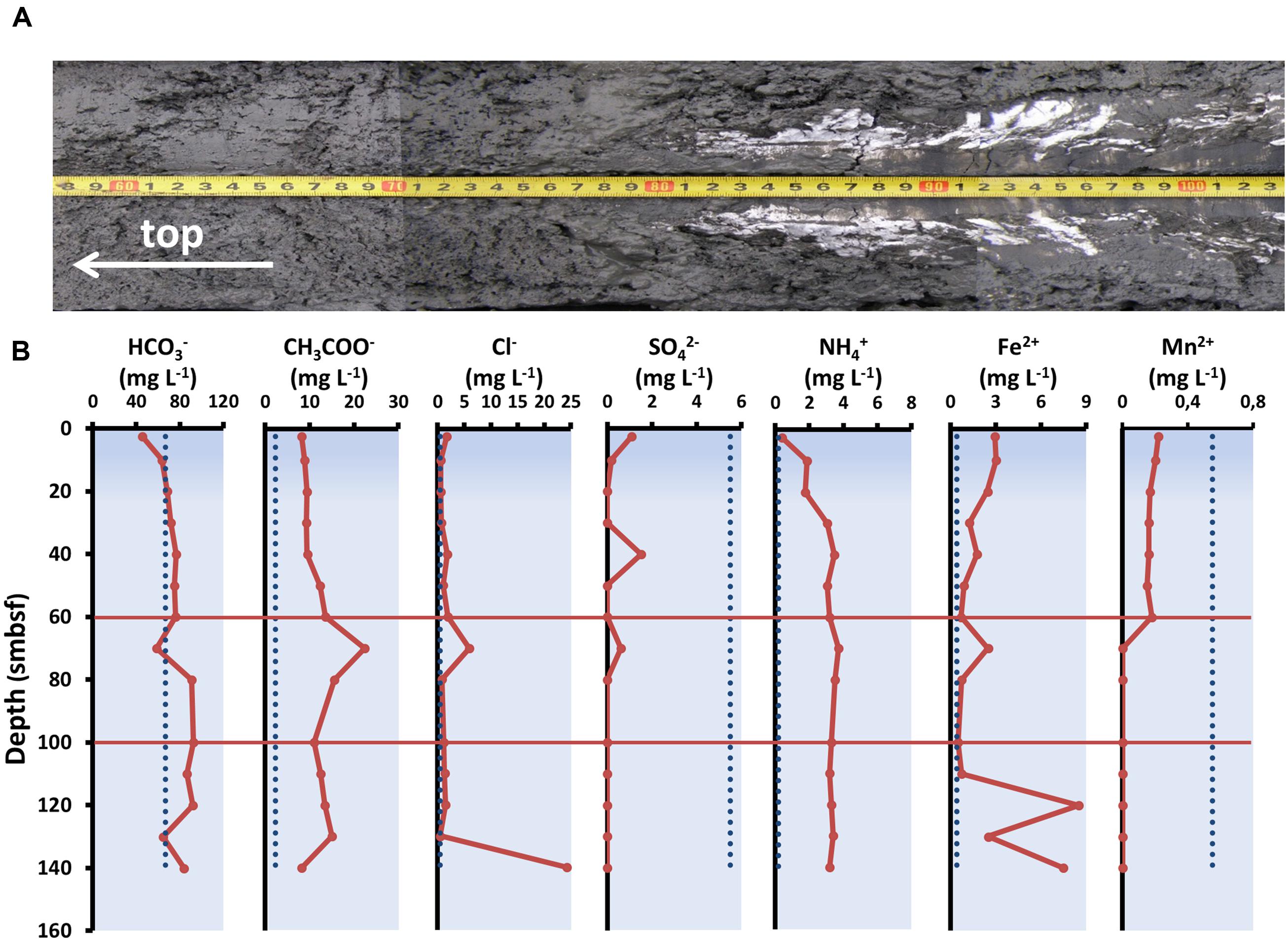
FIGURE 2. Physico-chemical parameters of sediment used in the experiment. (A) Structure of bottom sediment. (B) Concentrations of individual ions in the pore waters of bottom sediments from the core studied. The dashed lines indicate average ion concentrations in water column of Lake Baikal (according to Falkner et al., 1991; Namsaraev and Zemskaya, 2000).
Phylogenetic Composition of the Microbial Community in Samples of Bottom Sediments before the Experiment
Total 16S rRNA datasets for the samples from the horizons referred to the studied layer of bottom sediments include 7093 sequences assigned to bacterial 16S rRNA gene and 4902 to the archaeal gene. Phylogenetic analysis of the communities indicated that the members of the phyla Chloroflexi (23.6%), candidate phylum Atribacteria [OP9/JS1 (Dodsworth et al., 2013; Nobu et al., 2015)] (11.6%), Deinococcus-Thermus (11.4%), Acidobacteria (7.8%), OP8 (6.7%), WS3 (5.5%), TM7 (4.8%), Planctomycetes (4.7%) and Firmicutes (3.5%) dominated Bacteria (Figure 3A). “Rare taxons” comprising less than 3% of the sequences included the phyla Actinobacteria, Bacteroidetes, Caldiserica, Cyanobacteria, Lentisphaerae, Nitrospirae, Proteobacteria (α, β, γ, and δ), Verrucomicrobia.
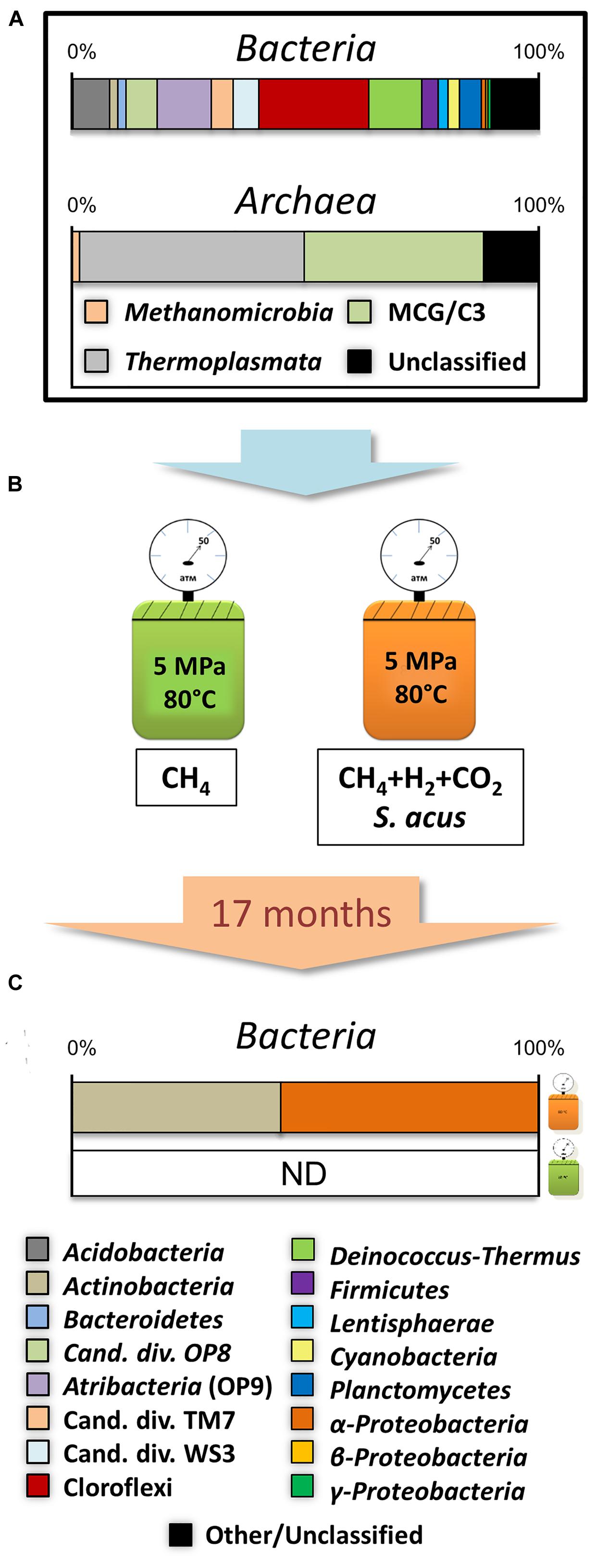
FIGURE 3. Experimental scheme. (A) Total composition of natural microbial community from individual layers of bottom sediments. (B) Culturing conditions. (C) Community composition of samples obtained after 17 months of culturing of bottom sediments under thermophilic conditions.
Within the Archaea domain, all classified sequences were distributed between the phylum Bathyarchaeota [Miscellaneous Crenarchaeotic Group, MCG (Evans et al., 2015)] – (32.2%) and the Thermoplasmata (47.9%) and Methanomicrobia (1.71%) classes representing the phylum Euryarchaeota (Figure 3A).
Results of Microscopy of the Samples after Culturing
After 17 months of thermobaric culturing in the autoclaves, DAPI and AO staining indicated the single cells of microorganisms only in the samples of bottom sediments added with the detritus of the algae S. acus. Microorganisms associated with the sediment particles were mostly baculiform (Figures 4A–D).
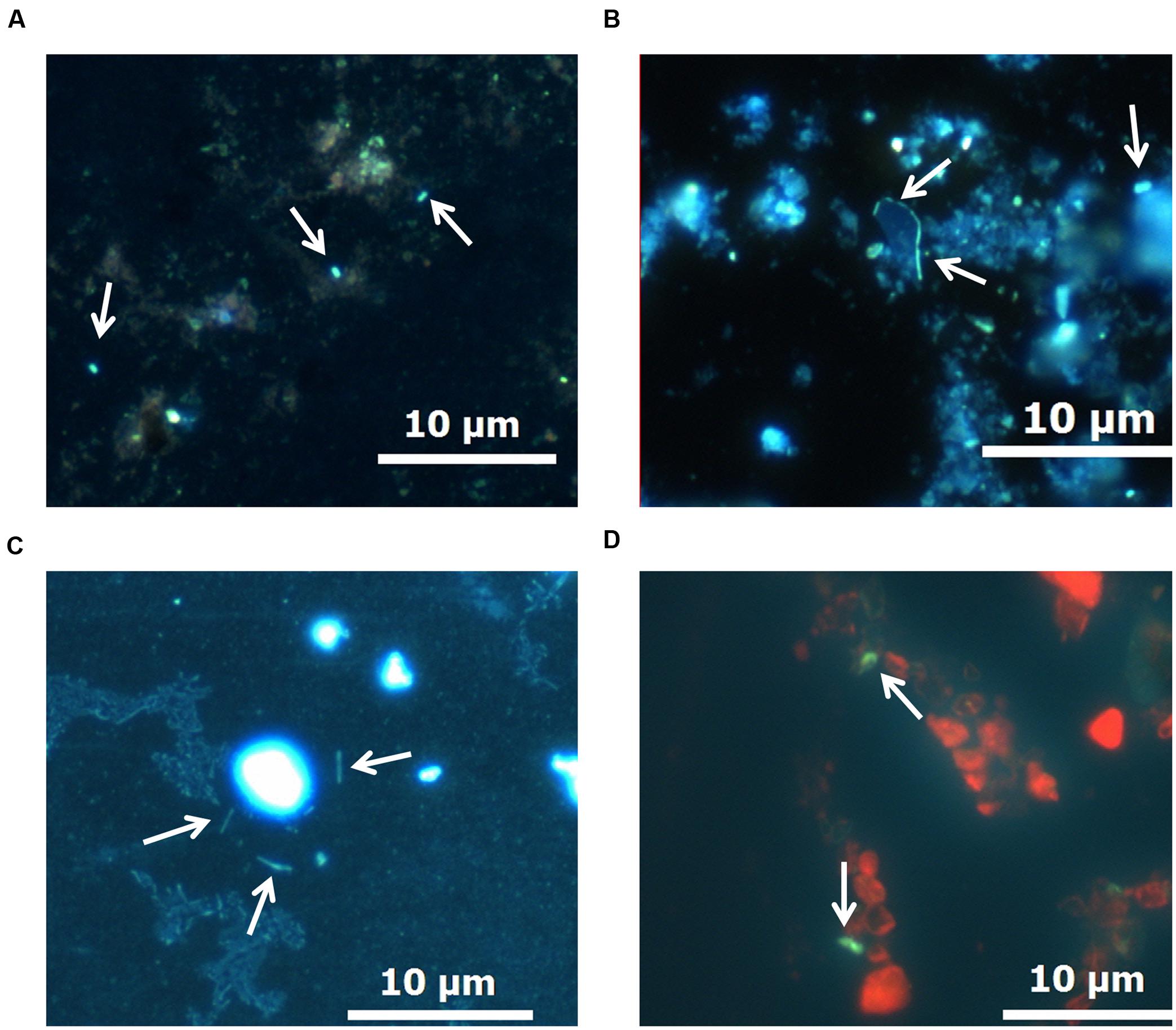
FIGURE 4. Morphology of microorganisms in bottom sediments after the experiment. (A–C) DAPI staining. (D) Acridine orange (AO) staining.
Results of Analysis of the Microbial Community from Bottom Sediments after 17 Months of Culturing
After the experiment, positive PCR amplifications were obtained only for DNA extracted from sample #1. A library of amplicons with primers on Bacteria was obtained for this sample. No positive PCR amplifications with primers on Archaea were obtained at that time. The DNA which was extracted from sediment samples #2 and #3 was not high-molecular, and all direct and nested PCR amplifications for these samples failed.
We obtained a library of amplification products of the 16S rRNA gene fragment comprising 13072 nucleotide sequences with an average length of 275 bp. The sequences were grouped into 41 OTU0.03. The rarefaction curves in Supplementary Figure S2, as well as the values of the ACE and Chao1 indices (Supplementary Table S3), indicate that the resulting sequencing volume is sufficient for the characterization of community diversity at the genus level (0.03 genetic distance).
Taxonomic classification showed that 99.4% of all sequences belonged to microorganisms of the genera Arthrobacter (16.6%) and Solirubrobacter (27.5%) associated with the phylum Actinobacteria (44.5%), and to the genus Sphingomonas (55.3%) representing the α-Proteobacteria class (55.3%; Figures 3B,C). At the same time, 0.2% of the sequences shared the phyla Bacteroidetes, Cyanobacteria, Planctomycetes, Verrucomicrobia and γ, δ- Proteobacteria classes, and were similar to those detected in natural sediment samples.
The comparison of the resultant sequences with the NCBI database using the BLAST protocol indicated that of the 16S rRNA sequences, the most similar to those obtained after cultivation, represented microorganisms found in the soil, water, and bottom sediments of freshwater and marine basins, as well as in groundwater and hydrocarbon contaminated ecosystems (Table 1).
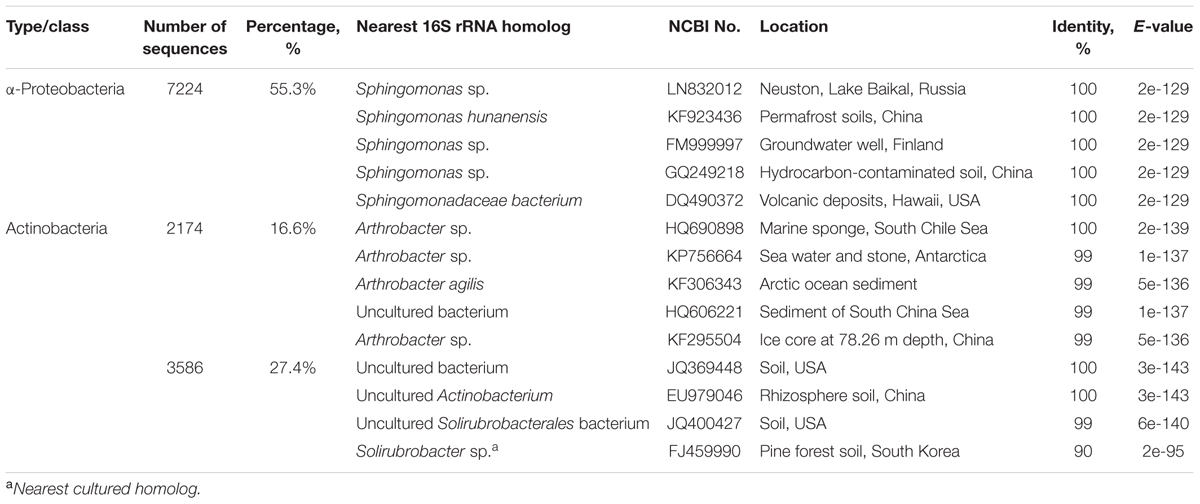
TABLE 1. The bacteria from the GenBank database most closely related, according to 16S rRNA gene sequences identified from bottom sediments after 17 months of culturing under thermophilic conditions.
Transformation of Organic Matter in the Experiment
Changes in the composition of organic matter were observed only in the sample from the autoclave #1. The TOC content of the sediment decreased during the experiment from 0.77 to 0.66%. Chloroform dissoluble bitumoid from the sediment before and after the experiment was only 0.02%; thus it was impossible to identify asphaltenes. Saturated hydrocarbons in the bitumoid before the experiment were 19.6% and asphalt-resinous compounds were 80.4%. After the experiment, the content of the latter in the sample increased to 91.2%. Before the experiment, the sample lacked aromatic fraction, whereas after the experiment it was 4.4%. Bitumoid coefficients (the ratio of the extract yield to the TOC content) in the samples before and after the experiment were 2.68 and 3.48, respectively. The ratio of acyclic isoprenoids, pristane/phytane, increased from 0.53 to 0.63. The carbon preference index (CPI), calculated according to the (1), decreased from 5.1 to 3.
The distribution curve of normal alkanes in the sample before the experiment (Supplementary Figure S3) was saw-toothed and asymmetric, with a maximum at C31. The n-C27/n–C17 ratio was 10. The distribution parameters of normal alkanes in the sample changed after the experiment (Supplementary Figure S3). The n-C27/n–C17 increased to 15.4.
Before and after the experiment, ethyl-cholestanes dominated the cyclic hydrocarbon biomarkers of C27–C30 steranes in the saturated fraction of the sample (37.3 and 39.2%, respectively). C27–C35 hopanes dominated terpanes (53.0% before and after the experiment). In this fraction, we also recorded an increase abundance of such biomarkers as gammacerane and retene after the experiment (Supplementary Figures S4 and S5).
Discussion
The natural microbial community of the bottom sediments studied was represented by the taxa which are mainly widespread in freshwater ecosystems (Newton et al., 2011). However, the uncultured members of Atribacteria and Chloroflexi, which were abundant in the studied sample, are not typical of the communities of sediments in Lake Baikal (Zemskaya et al., 2015). In Baikal, as in many other freshwater lakes (Winters et al., 2014; Zhang et al., 2014; Ding et al., 2015), the most abundant phyla are Proteobacteria and Actinobacteria. Previously, communities where the taxa Atribacteria, Chloroflexi, and Bathyarchaeota were most abundant was identified only in the subsurface hydrate-associated sediments of the Saint Petersburg methane seep in the Central Baikal Basin (Kadnikov et al., 2011). Members of the three phyla were registered in the bottom sediments of freshwater lakes (Zhang et al., 2014; Ding et al., 2015), but they were mostly characteristic of the various specific deep marine sub-seafloor locations, including seeps and gas-hydrate sites (Orcutt et al., 2011; Parkes et al., 2014; Ruff et al., 2015). Chloroflexi are mainly found in bottom sediments enriched with organic matter: whereas the representatives of the candidate phylum Atribacteria form the basis of the communities from the hydrate-associated sediments; Bathyarchaeota contains a huge number of diverse phylogenetic lineages and is ubiquitous (Inagaki et al., 2006; Webster et al., 2006; Orcutt et al., 2011; Parkes et al., 2014). Chloroflexi and Atribacteria show heterotrophic metabolisms and are involved in various stages of the destruction of detritus or organic matter originating from other sources (Webster et al., 2006; Kubo et al., 2012; Dodsworth et al., 2013; Hug et al., 2013). Metabolic reconstruction of the two Bathyarchaeota genomes revealed the presence of genes that are diagnostic of the capacity to perform methylotrophic methanogenesis or anaerobic oxidation of methane (AOM) and also use lactate, peptides, monosaccharides, and pyruvate for energy production (Evans et al., 2015). Another interesting group of Archaea, which sequences were abundant in natural sediment, is Thermoplasmata. Although most cultured members of the Thermoplasmata are primarily found in oxic and/or hot environments, sequences of the uncultured representatives of this group (uncultured Thermoplasmatales in our case) are abundant in cold anoxic sediments (Lloyd et al., 2013). Network analysis showed that Thermoplasmata recurrently co-occur with Bathyarchaeota in sediment ecosystems, suggesting a potentially relevant trophic connection between the two clades (Fillol et al., 2015).
The abundance of Atribacteria, Chloroflexi, Bathyarchaeota, and Thermoplasmata in the sediments analyzed may thus be due to active processes of destruction of detrital material in the studied area of gas discharge. This correlates with the high acetate and ammonium concentrations in the core, which may be a result of both the local active anaerobic decomposition of organic matter (and/or nitrate reduction) and transportation with deep fluid flow (Krylov et al., 2008; Sass and Parkes, 2011; Boetius and Wenzhöfer, 2013). Functions of primary and secondary fermenting bacteria in the studied community can represent other abundant taxa, namely: Acidobacteria, Deinococcus-Thermus, Firmicutes, OP8, WS3, TM7. Representatives of Acidobacteria can also use iron as an electron acceptor for anaerobic respiration (Ward et al., 2009), whereas Planctomycetes are capable of the anaerobic oxidation of ammonium (Fuerst and Sagulenko, 2011). A small number of methanogenic Methanomicrobia is consistent with data suggesting that the methane in the studied area mostly comes from the underlying layers (Klerkx et al., 2006; Naudts et al., 2012).
Taxonomic assignment of the 16S rRNA sequence data indicates that the predominant sequences after cultivation were Actinobacteria and Proteobacteria, which were initially considered rare taxa. At that time, no Archaea were detected. Significant changes in the composition and diversity of the microbial community from natural sediments that occurred in the course of the exposure under the experimental conditions may show that the abundant taxa are likely to be indigenous to the cold near-surface bottom sediments and not able to survive at high temperature. Intact cells of microorganisms were found after experiment, however. Also, in the course of the culturing, the CPI value also decreased from 5.1 to 3, indicating that the transformation degree of the hydrocarbon component in organic matter reached 41%. Hydrocarbon molecules with an odd number of carbon atoms are known to prevail in organic matter from sediments not exposed to diagenetic and catagenetic transformations. The values of the odd/even ratio (CPI in different modifications) are therefore 5–6. During the thermal transformation of organic matter, the number of odd and even hydrocarbon molecules equalizes. The CPI value approaches one and does not change in subsequent late-catagenetic transformations (after the main stage of oil formation; Mukhopadhyay et al., 1979). In this case (culturing at 80°C), however, the observed changes probably resulted from the biological degradation processes, since these values on the one hand approach the threshold for living organisms, but on the other hand are the lower boundary of the values characteristic of metacatagenesis and ensure the transformation of organic matter free of the influence of physical factors. In nature, the temperature conditions of the thermal decomposition of organic matter (catagenesis) range from 85 to several 100°C (Schobert, 2013). So, under the experimental conditions, the microorganisms from the cold near-surface bottom sediments should thus survive at high temperature and cause deep destruction of organic matter, respiring very slowly or anaerobically.
Most of the phylotypes identified after the experiment were related to heterotrophs of the genera Arthrobacter, Solirubrobacter, and Sphingomonas. Bacteria of these genera are capable of utilizing a wide range of organic substrates, including aromatic hydrocarbons, chlorinated and other toxic compounds (Daane et al., 2002; Choi et al., 2004; Story et al., 2004; Nordin et al., 2005) and play an important role in the microbial communities of oil-contaminated soils (Peng et al., 2015). Cultivated strains of Arthrobacter, Solirubrobacter, and Sphingomonas are typical inhabitants of mesophilic soil environments, however. Furthermore, the ability to grow under anaerobic conditions due to nitrate reduction or fermentation was previously identified only in some Arthrobacter species (Eschbach et al., 2003). It is therefore unlikely that these microorganisms, as representatives of the indigenous microbial community of the cold near-surface bottom sediments, were able to survive under anaerobic hyperthermophilic conditions. One possible explanation for their abundance is that these microorganisms are one of the representatives of the microbial community of deep hot sediment layers which can migrate to the near surface sediment layers through fault zones, together with gas-bearing fluids. Both positive heat-flow anomalies near the venting sites and a negative heat flow anomaly zone to the south of the venting sites were registered within the Posolskaya Bank structure, which may indicate a convective fluid loop (Klerkx et al., 2006; Naudts et al., 2012). Early, some representatives of Arthrobacter, Solirubrobacter, and Sphingomonas were enriched from deep biosphere (Fredrickson et al., 1995; Balkwill et al., 1997; Crocker et al., 2000; Chang et al., 2007; Kobayashi et al., 2008), including thermophilic conditions (Ciobanu et al., 2014). Cultivation-independent molecular biological techniques also confirmed the presence of these bacteria in the deep biosphere (Inagaki et al., 2006; Mason et al., 2010; Breuker et al., 2011; Purkamo et al., 2013). According to one of the hypotheses, the widespread representatives of the phyla Actinobacteria and Proteobacteria may be explained by their flexible metabolism, allowing them to adapt to environmental changes that would have been maintained during the progressive burial of sediments (Ciobanu et al., 2014). In fact, many OTUs from ~40 to 60°C sediment associated with lignite coal beds at ~1.5 to 2.5 km below the seafloor in the Pacific Ocean deeper layers showed high sequence similarity to those from soil environments (Actinobacteria, Proteobacteria, and Firmicutes, etc.; Inagaki et al., 2015). Nevertheless, we cannot assert that representatives of Arthrobacter, Solirubrobacter, and Sphingomonas performed transformation of organic matter in the experiment, because they could simply have remained intact during the culturing. At that time, those thermophilic microorganisms which performed decomposition of organic matter may have lost viability as a result of exhaustion of the energy and/or carbon sources during long time cultivation. In previous studies, after culturing the microbial community of near-surface sediments of Lake Baikal under thermobaric conditions within a shorter period (11 months), thermophilic microorganisms were enriched (Pavlova et al., 2016).
The signs of microbial activity after 17 months of culturing in bottom sediments, supplemented with the S. acus detritus, can be explained by relatively high concentrations of organic matter and presence of CO2 and H2. The alga could be used as a substrate for fermenting bacteria, at that time CO2 could be utilized as an electron acceptor for autotrophic H2 utilization, such as hydrogenotrophic methanogenesis and acetogenesis. Previously, it was shown that homo-acetogenesis are activated during cultivation of bituminous coal and sand under anaerobic, CO2-rich conditions at 40 MPa and 40°C (Ohtomo et al., 2013). Also, authors decelerate that no methanogens were activated during their experiment (Ohtomo et al., 2013). That is consistent with the fact that no methanogenic Archaea were detected after our experiment. In autoclave #1 the microorganisms could also use substrates formed in the course of low-temperature activation of organic matter. It was reported that the warming of sediments during burial can activate both buried organic and inorganic compounds and stimulates both H2 production and prokaryotic activity, while sterile controls showed negligible H2 formation and other geochemical changes, even at temperatures as high as 100°C (e.g., basalt incubation for 130 days; Parkes et al., 2011).
Another distinctive feature is an increased abundance of the pentacyclic oil biomarker, gammacerane. Gammacerane is recognized as a precursor of gammacerane, a polycyclic hydrocarbon detected in sedimentary rocks dating as far back as the late Proterozoic (~850 Mya). Gammacerane is usually regarded as marking the sediment accumulation in saline environments (Sinninghe Damsté et al., 1995; Peters et al., 2005). In our case, the initial substrate was a freshwater source, as indicated by the numerical value of the isoprenoid ratios pristine/phytane. According to the data obtained in Vaz dos Santos Neto et al. (1998), gammacerane has δ13C values compatible with derivation from ciliates feeding on algae, cyanobacteria and another microorganisms growing under variable paleoenvironmental conditions. Gammacerane is also a major biomarker in many lacustrine oils and bitumens, including the Green River marl and oils from China, where the organic matter originated mainly from algae and bacteria (Peters et al., 2005). In our previous studies on culturing the microbial community of near-surface sediments from the cold methane seep Goloustnoe (Southern Basin), we obtained comparable data. The sediments after the experiment showed the formation of the polycyclic aromatic hydrocarbon, retene. Our experimental data confirms that retene can be generated in Lake Baikal during the process of biomass destruction of diatoms performed by a microbial community from deep bottom sediments (Pavlova et al., 2016). The experiments showed that microbial communities from bottom sediments of Lake Baikal with different chemical characteristics, but sufficient content of organic matter, under certain temperatures, are capable of destroying organic substrates followed by the formation of such biomarkers as retene and gammacerane.
Author Contributions
TZ, OP, and SB conceived the study. OP and SB collected the samples. AM and SB carried out experimental cultivation. EK performed GC-MS analysis. TP carried out chemical analysis. SB performed all data analysis with support from EM and SC. SB wrote the manuscript with input from all authors.
Funding
This work was supported by framework of the state task No 0345–2014–0009 “Geobiochemical studies of the methane cycles…”, No 0345–2015–0030 “The study of structural and functional characteristics of biological communities…” The work was supported by Interdisciplinary Integration Project SB RAS No. 82 and in part the Russian Foundation for Basic Research grant No. 16-04-00181.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Oleg M. Khlystov, Andrei V. Khabuev and the crew of RV G.Y. Vereschagin for assistance in sampling and map creating. The authors are also grateful staff of the Limnological Institute Darya P. Petrova, Yuri P. Galachyants, Maria V. Bashenkhaeva, and Anna V. Lomakina for assistance in 16S rRNA gene amplicon sequencing and analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00690
Footnotes
- ^ http://www.mothur.org/wiki/Silva_reference_files
- ^ http://www.arb-silva.de
- ^ http://blast.ncbi.nlm.nih.gov
References
Balkwill, D. L., Drake, G. R., Reeves, R. H., Fredricson, J. K., White, D. C., Ringelberg, D. B., et al. (1997). Taxonomic study of aromatic-degrading bacteria from deep-terrestrial-subsurface sediments and description of Sphingomonas aromaticivorans sp. nov., Sphingomonas subterranea sp. nov., and Sphingomonas stygia sp. nov. Int. J. Syst. Evol. Microbiol. 47, 191–201. doi: 10.1099/00207713-47-1-191
Bezrukova, E., Bukharov, A., Bychinsky, V., Fedenya, S., Gelety, V., Goreglyad, A., et al. (2005). A new quaternary record of regional tectonic, sedimentation and paleoclimate changes from drill core BDP-99 at Posolskaya Bank. Lake Baikal. Quat. Int. 136, 105–121. doi: 10.1016/j.quaint.2004.11.012
Biddle, J. F., Sylvan, J. B., Brazelton, W. J., Tully, B. J., Edwards, K. J., Moyer, C. L., et al. (2012). Prospects for the study of evolution in the deep biosphere. Front. Microbiol. 2:285. doi: 10.3389/fmicb.2011.00285
Boetius, A., and Wenzhöfer, F. (2013). Seafloor oxygen consumption fueled by methane from cold seeps. Nat. Geosci. 6, 725–734. doi: 10.1038/ngeo1926
Bradbury, J. P., Bezrukova, Y. V., Chernyaeva, G. P., Colman, S. M., Khursevich, G., King, J. W., et al. (1994). A synthesis of post-glacial diatom records from Lake Baikal. J. Paleolimnol. 10, 213–252. doi: 10.1007/bf00684034
Breuker, A., Köweker, G., Blazejak, A., and Schippers, A. (2011). The deep biosphere in terrestrial sediments in the chesapeake Bay area, Virginia, USA. Front. Microbiol. 2:156. doi: 10.3389/fmicb.2011.00156
Chang, H. W., Bae, J. W., Nam, Y. D., Kwon, H. Y., Park, J. R., Shin, K. S., et al. (2007). Arthrobacter subterraneus sp. nov., isolated from deep subsurface water of the South Coast of Korea. J. Microbiol. Biotechnol. 17, 1875–1879.
Choi, K. K., Park, C. W., Kim, S. Y., Lyoo, W. S., Lee, S. H., and Lee, J. W. (2004). Polyvinyl alcohol degradation by Microbacterium barkeri KCCM 10507 and Paeniblacillus amylolyticus KCCM 10508 in dyeing wastewater. J. Microbiol. Biotechnol. 14, 1009–1013.
Ciobanu, M.-C., Burgaud, G., Dufresne, A., Breuker, A., Rédou, V., Ben Maamar, S., et al. (2014). Microorganisms persist at record depths in the subseafloor of the Canterbury Basin. ISME J. 8, 2352–2352. doi: 10.1038/ismej.2014.110
Cowen, J. P., Giovannoni, S. J., Kenig, F., Johnson, H. P., Butterfield, D., Rappé, M. S., et al. (2003). Fluids from aging ocean crust that support microbial life. Science 299, 120–123. doi: 10.1126/science.1075653
Cragg, B. A., Parkes, R. J., Fry, J. C., Herbert, R. A., Wimpenny, J. W. T., and Getliff, J. M. (1990). Bacterial biomass and activity profiles within deep sediment layers. Proc. Oce. Drill. Prog. 112, 607–619. doi: 10.2973/odp.proc.sr.112.161.1990
Crocker, F. H., Fredrickson, J. K., White, D. C., Ringelberg, D. B., and Balkwill, D. L. (2000). Phylogenetic and physiological diversity of Arthrobacter strains isolated from unconsolidated subsurface sediments. Microbiology 146, 1295–1310. doi: 10.1099/00221287-146-6-1295
Daane, L. L., Harjono, I., Barns, S. M., Launen, L. A., Palleron, N. J., and Häggblom, M. M. (2002). PAH-degradation by Paenibacillus spp. and description of Paenibacillus naphthalenovorans sp. nov., a naphthalene-degrading bacterium from the rhizosphere of salt marsh plants. Int. J. Syst. Evol. Microbiol. 52, 131–139. doi: 10.1099/00207713-52-1-131
D’Hondt, S., Jørgensen, B. B., Miller, D. J., Batzke, A., Blake, R., Cragg, B. A., et al. (2004). Distributions of microbial activities in deep subseafloor sediments. Science 306, 2216–2221. doi: 10.1126/science.1101155
Ding, X., Peng, X.-J., Jin, B.-S., Xiao, M., Chen, J.-K., Li, B., et al. (2015). Spatial distribution of bacterial communities driven by multiple environmental factors in a beach wetland of the largest freshwater lake in China. Front. Microbiol. 6:129. doi: 10.3389/fmicb.2015.00129
Dodsworth, J. A., Blainey, P. C., Murugapiran, S. K., Swingley, W. D., Ross, C. A., Tringe, S. G., et al. (2013). Single-cell and metagenomic analyses indicate a fermentative and saccharolytic lifestyle for members of the OP9 lineage. Nat. Comms. 4:1854. doi: 10.1038/ncomms2884
dos Santos Neto, E., Hayes, J. M., and Takaki, T. (1998). Isotopic biogeochemistry of the Neocomian lacustrine and Upper Aptian marine-evaporitic sediments of the Potiguar Basin, Northeastern Brazil. Org. Geochem. 28, 361–381. doi: 10.1016/s0146-6380(98)00007-2
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Engelen, B., Ziegelmüller, K., Wolf, L., Köpke, B., Gittel, A., Cypionka, H., et al. (2008). Fluids from the oceanic crust support microbial activities within the deep biosphere. Geomicrobiol. J. 25, 56–66. doi: 10.1080/01490450701829006
Eschbach, M., Möbitz, H., Rompf, A., and Jahn, D. (2003). Members of the genus Arthrobacter grow anaerobically using nitrate ammonification and fermentative processes: anaerobic adaptation of aerobic bacteria abundant in soil. FEMS Microbiol. Lett. 223, 227–230. doi: 10.1016/s0378-1097(03)00383-5
Evans, P. N., Parks, D. H., Chadwick, G. L., Robbins, S. J., Orphan, V. J., Golding, S. D., et al. (2015). Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics. Science 350, 434–438. doi: 10.1126/science.aac7745
Falkner, K. K., Measures, C. I., Herbelin, S. E., Edmond, J. M., and Weiss, R. F. (1991). The major and minor element geochemistry of Lake Baikal. Limnol. Oceanogr. 36, 413–423. doi: 10.4319/lo.1991.36.3.0413
Fillol, M., Auguet, J.-C., Casamayor, E. O., and Borrego, C. M. (2015). Insights in the ecology and evolutionary history of the Miscellaneous Crenarchaeotic Group lineage. ISME J. 10, 665–677. doi: 10.1038/ismej.2015.143
Fredrickson, J. K., McKinley, J. P., Nierzwicki-Bauer, S. A., White, D. C., Ringelberg, D. B., Rawson, S. A., et al. (1995). Microbial community structure and biogeochemistry of Miocene subsurface sediments: implications for long-term microbial survival. Mol. Ecol. 4, 619–626. doi: 10.1111/j.1365-294x.1995.tb00262.x
Fuerst, J. A., and Sagulenko, E. (2011). Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function. Nat. Rev. Micro. 9, 403–413. doi: 10.1038/nrmicro2578
Gold, T. (1992). The deep, hot biosphere. Proc. Natl. Acad. Sci. U.S.A. 89, 6045–6049. doi: 10.1073/pnas.89.13.6045
Granina, L. Z., Callender, E., Lomonosov, I. S., Mats, V. D., and Golobokova, L. P. (2001). Anomalies in the composition of Baikal pore waters. Rus. Geol. Geoph. 42, 362–372.
Horsfield, B., Schenk, H. J., Zink, K., Ondrak, R., Dieckmann, V., Kallmeyer, J., et al. (2006). Living microbial ecosystems within the active zone of catagenesis: Implications for feeding the deep biosphere. Earth Planet. Sci. Lett. 246, 55–69. doi: 10.1016/j.epsl.2006.03.040
Hubert, C., Loy, A., Nickel, M., Arnosti, C., Baranyi, C., Bruchert, V., et al. (2009). A constant flux of diverse thermophilic bacteria into the cold arctic seabed. Science 325, 1541–1544. doi: 10.1126/science.1174012
Hug, L. A., Castelle, C. J., Wrighton, K. C., Thomas, B. C., Sharon, I., Frischkorn, K. R., et al. (2013). Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling. Microbiome 1:22. doi: 10.1186/2049-2618-1-22
Inagaki, F., Hinrichs, K.-U., Kubo, Y., Bowles, M. W., Heuer, V. B., Hong, W.-L., et al. (2015). Exploring deep microbial life in coal-bearing sediment down to 2.5 km below the ocean floor. Science 349, 420–424. doi: 10.1126/science.aaa6882
Inagaki, F., Nunoura, T., Nakagawa, S., Teske, A., Lever, M., Lauer, A., et al. (2006). Biogeographical distribution and diversity of microbes in methane hydrate-bearing deep marine sediments on the Pacific Ocean Margin. Proc. Natl. Acad. Sci. U.S.A. 103, 2815–2820. doi: 10.1073/pnas.0511033103
Kadnikov, V. V., Mardanov, A. V., Beletsky, A. V., Shubenkova, O. V., Pogodaeva, T. V., Zemskaya, T. I., et al. (2011). Microbial community structure in methane hydrate-bearing sediments of freshwater Lake Baikal. FEMS Microbiol. Ecol. 79, 348–358. doi: 10.1111/j.1574-6941.2011.01221.x
Kalmychkov, G. V., Egorov, A. V., Kuzmin, M. I., and Khlystov, O. M. (2006). Genetic types of methane from Lake Baikal. Dokl. Earth Sci. 411, 1462–1465. doi: 10.1134/s1028334x06090285
Khlystov, O. M., De Batist, M., Shoji, H., Hachikubo, A., Nishio, S., and Naudts, L. (2013). Gas hydrate of lake baikal: discovery and varieties. J. Asian Earth Sci. 62, 162–166. doi: 10.1016/j.jseaes.2012.03.009
Khlystov, O. M., Gorshkov, A. G., Egorov, A. V., Zemskaya, T. I., Granin, N. G., Kalmychkov, G. V., et al. (2007). Oil in the lake of world heritage. Dokl. Earth Sci. 415, 682–685. doi: 10.1134/s1028334x07050042
Kieft, T. L., Fredrickson, J. K., McKinley, J. P., Bjornstad, B. N., Rawson, S. A., Phelps, T. J., et al. (1995). Microbiological comparisons within and across contiguous lacustrine, paleosol, and fluvial subsurface sediments. Appl. Environ. Microbiol. 61, 749–757.
Klerkx, J., De Batist, M., Poort, J., Hus, R., Van Rensbergen, P., Khlystov, O. M., et al. (2006). Tectonically controlled methane escape in Lake Baikal. Advances in the geological storage of carbon dioxide. NATO Sci. Ser. IV. Earth Environ. Sci. 65, 203–219. doi: 10.1007/1-4020-4471-2_17
Klerkx, J., Zemskaya, T. I., Matveeva, T. V., Khlystov, O. M., Namsaraev, B. B., Dagurova, O. P., et al. (2003). Methane hydrates in surface layer of deep-water sediments in Lake Baikal (in Russian). Dokl. Earth Sci. 393, 822–826.
Kobayashi, T., Koide, O., Mori, K., Shimamura, S., Matsuura, T., Miura, T., et al. (2008). Phylogenetic and enzymatic diversity of deep subseafloor aerobic microorganisms in organics- and methane-rich sediments off Shimokita Peninsula. Extremophiles 12, 519–527. doi: 10.1007/s00792-008-0157-7
Kontorovich, A. E., Kashirtsev, V. A., Moskvin, V. I., Burshtein, L. M., Zemskaya, T. I., Kostyreva, E. A., et al. (2007). Petroleum potential of Baikal deposits. Rus. Geol. Geoph. 48, 1046–1053. doi: 10.1016/j.rgg.2007.11.004
Krylov, A., Khlystov, O., Zemskaya, T., Minami, H., Hachikubo, A., Nunokawa, Y., et al. (2008). First discovery and formation process of authigenic siderite from gas hydrate–bearing mud volcanoes in fresh water: Lake Baikal, eastern Siberia. Geophys. Res. Lett. 35:5. doi: 10.1029/2007gl032917
Kubo, K., Lloyd, K. G., Biddle, J. F., Amann, R., Teske, A., and Knittel, K. (2012). Archaea of the Miscellaneous Crenarchaeotal Group are abundant, diverse and widespread in marine sediments. ISME J. 6, 1949–1965. doi: 10.1038/ismej.2012.37
Kuzmin, M. I., Geletiy, V. F., Kalmychkov, G. V., Kuznetsov, F. A., Larionov, E. G., and Manakov, A. (2000). The first discovery of the gas hydrates in the sediments of the Lake Baikal. Ann. N. Y. Acad. Sci. 912, 112–115. doi: 10.1111/j.1749-6632.2000.tb06764.x
Lee, S., Malone, C., and Kemp, P. (1993). Use of multiple 16S rRNA-targeted fluorescent probes to increase signal strength and measure cellular RNA from natural planktonic bacteria. Mar. Ecol. Prog. Ser. 101, 193–201. doi: 10.3354/meps101193
Lloyd, K. G., Schreiber, L., Petersen, D. G., Kjeldsen, K. U., Lever, M. A., Steen, A. D., et al. (2013). Predominant archaea in marine sediments degrade detrital proteins. Nature 496, 215–218. doi: 10.1038/nature12033
Mason, O. U., Nakagawa, T., Rosner, M., Van Nostrand, J. D., Zhou, J., Maruyama, A., et al. (2010). First investigation of the microbiology of the deepest layer of ocean crust. PLoS ONE 5:e15399. doi: 10.1371/journal.pone.0015399
Mukhopadhyay, P. K., Hagemann, H. W., Hollerbach, A., and Welte, D. H. (1979). The relation between organic geochemical and petrological parameters of coal in Indian coal basins. Energ. Source 4, 313–328. doi: 10.1080/00908317908908067
Muyzer, G., De Waal, E. C., and Uitterlinden, A. G. (1993). Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 59, 695–700.
Namsaraev, B. B., and Zemskaya, T. I. (2000). Microbiological Processes of Carbon Circulation in Lake Baikal Bottom Sediments. Novosibirsk: SB RAS Publication, Filial.
Naudts, L., Khlystov, O., Granin, N., Chensky, A., Poort, J., and De Batist, M. (2012). Stratigraphic and structural control on the distribution of gas hydrates and active gas seeps on the Posolsky Bank, Lake Baikal. Geo Mar. Lett. 32, 395–406. doi: 10.1007/s00367-012-0286-y
Newton, R. J., Jones, S. E., Eiler, A., McMahon, K. D., and Bertilsson, S. (2011). A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 75, 14–49. doi: 10.1128/mmbr.00028-10
Nobu, M. K., Dodsworth, J. A., Murugapiran, S. K., Rinke, C., Gies, E. A., Webster, G., et al. (2015). Phylogeny and physiology of candidate phylum “Atribacteria” (OP9/JS1) inferred from cultivation-independent genomics. ISME J 10, 273–286. doi: 10.1038/ismej.2015.97
Nordin, K., Unell, M., and Jansson, J. K. (2005). Novel 4-Chlorophenol degradation gene cluster and degradation route via hydroxyquinol in Arthrobacter chlorophenolicus A6. Appl. Environ. Microbiol. 71, 6538–6544. doi: 10.1128/aem.71.11.6538-6544.2005
Ohtomo, Y., Ijiri, A., Ikegawa, Y., and Inagaki, F. (2013). Biological CO2 conversion to acetate in subsurface coal-sand formation using a high-pressure reactor system. Front. Microbiol. 4:361. doi: 10.3389/fmicb.2013.00361
Orcutt, B. N., Sylvan, J. B., Knab, N. J., and Edwards, K. J. (2011). Microbial ecology of the dark ocean above, at, and below the seafloor. Microbiol. Mol. Biol. Rev. 75:361. doi: 10.1128/mmbr.00039-10
Parkes, R. J., Cragg, B., Roussel, E., Webster, G., Weightman, A., and Sass, H. (2014). A review of prokaryotic populations and processes in sub-seafloor sediments, including biosphere: geosphere interactions. Mar. Geol. 352, 409–425. doi: 10.1016/j.margeo.2014.02.009
Parkes, R. J., Cragg, B. A., Bale, S. J., Getliff, J. M., Goodman, K., Rochelle, P. A., et al. (1994). Deep bacterial biosphere in Pacific Ocean sediments. Nature 371, 410–413. doi: 10.1038/371410a0
Parkes, R. J., Cragg, B. A., Fry, J. C., Herbert, R. A., and Wimpenny, J. W. T. (1990). Bacterial biomass and activity in deep sediment layers from the Peru margin. Philos. Trans. R. Soc. London A. 331, 139–153. doi: 10.1098/rsta.1990.0061
Parkes, R. J., Linnane, C. D., Webster, G., Sass, H., Weightman, A. J., Hornibrook, E. R. C., et al. (2011). Prokaryotes stimulate mineral H2 formation for the deep biosphere and subsequent thermogenic activity. Geology 39, 219–222. doi: 10.1130/g31598.1
Parkes, R. J., Webster, G., Cragg, B. A., Weightman, A. J., Newberry, C. J., Ferdelman, T. G., et al. (2005). Deep sub-seafloor prokaryotes stimulated at interfaces over geological time. Nature 436, 390–394. doi: 10.1038/nature03796
Parkes, R. J., Wellsbury, P., Mather, I. D., Cobb, S. J., Cragg, B. A., Hornibrook, E. R. C., et al. (2007). Temperature activation of organic matter and minerals during burial has the potential to sustain the deep biosphere over geological timescales. Org. Geochem. 38, 845–852. doi: 10.1016/j.orggeochem.2006.12.011
Pavlova, O. N., Zemskaya, T. I., Lomakina, A. V., Shubenkova, O. V., Manakov, A. Y., Moskvin, V. I., et al. (2016). Transformation of organic matter by microbial community in sediments of Lake Baikal under experimental thermobaric conditions of protocatagenesis. Geomicrobiol. J. (in press). doi: 10.1080/01490451.2015.1069910
Peng, M., Zi, X., and Wang, Q. (2015). Bacterial community diversity of oil-contaminated soils assessed by high throughput sequencing of 16S rRNA genes. Int. J. Environ. Res. Public Health 12, 12002–12015. doi: 10.3390/ijerph121012002
Peters, K. E., Walters, C. C., and Moldowan, J. M. (2005). The Biomarker Guide: Biomarkers and Isotopes in Petroleum Systems and Earth History. Cambridge: Cambridge University Press.
Pimenov, N. V., Zakharova, E. E., Bryukhanov, A. L., Korneeva, V. A., Kuznetsov, B. B., Tourova, T. P., et al. (2014). Activity and structure of the sulfate-reducing bacterial community in the sediments of the southern part of Lake Baikal. Microbiology 83, 47–55. doi: 10.1134/s0026261714020167
Pogodaeva, T. V., Zemskaya, T. I., Golobokova, L. P., Khlystov, O. M., Minami, H., and Sakagami, H. (2007). Chemical composition of pore waters of bottom sediments in different Baikal basins. Rus. Geol. Geoph. 48, 886–900. doi: 10.1016/j.rgg.2007.02.012
Purkamo, L., Bomberg, M., Nyyssönen, M., Kukkonen, I., Ahonen, L., Kietäväinen, R., et al. (2013). Dissecting the deep biosphere: retrieving authentic microbial communities from packer-isolated deep crystalline bedrock fracture zones. FEMS Microbiol. Ecol. 85, 324–337. doi: 10.1111/1574-6941.12126
Quince, C., Lanzen, A., Davenport, R. J., and Turnbaugh, P. J. (2011). Removing noise from pyrosequenced amplicons. BMC Bioinformation 12:38. doi: 10.1186/1471-2105-12-38
Reysenbach, A.-L., and Pace, N. R. (1995). “Reliable amplification of hyperthermophilic archaeal 16S rRNA genes by the polymerase chain reaction,” in Archaea: A Laboratory Manual, Thermophiles, eds F. T. Robb and A. R. Place (New York, NY: Cold Spring Harbour Laboratory Press), 101–107.
Roussel, E. G., Bonavita, M.-A. C., Querellou, J., Cragg, B. A., Webster, G., Prieur, D., et al. (2008). Extending the sub-sea-floor biosphere. Science 320, 1046–1046. doi: 10.1126/science.1154545
Ruff, S. E., Biddle, J. F., Teske, A. P., Knittel, K., Boetius, A., and Ramette, A. (2015). Global dispersion and local diversification of the methane seep microbiome. Proc. Natl. Acad. Sci. U.S.A. 112, 4015–4020. doi: 10.1073/pnas.1421865112
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning. A Laboratory Manual, 2nd Edn. New York, NY: Cold Spring Harbor.
Sass, H., and Parkes, R. J. (2011). “Sub-seafloor sediments: an extreme but globally significant prokaryotic habitat (taxonomy, diversity, ecology),” in Extremophiles Handbook, eds K. Horikoshi, G. Antranikian, A. T. Bull, F. T. Robb, and K. O. Stetter (Tokyo, Japan: Springer), 1016–1036.
Schippers, A., Neretin, L. N., Kallmeyer, J., Ferdelman, T. G., Cragg, B. A., Parkes, R. J., et al. (2005). Prokaryotic cells of the deep sub-seafloor biosphere identified as living bacteria. Nature 433, 861–864. doi: 10.1038/nature03302
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/aem.01541-09
Schobert, H. (2013). Chemistry of Fossil Fuels and Biofuels. New York, NY: Cambridge University Press. doi: 10.1017/cbo9780511844188
Shishlyannikov, S. M., Zakharova, Y. R., Volokitina, N. A., Mikhailov, I. S., Petrova, D. P., and Likhoshway, Y. V. (2011). A procedure for establishing an axenic culture of the diatom Synedra acus subsp. radians (Kütz.) Skabibitsch. from Lake Baikal. Limnol. Oceanogr. Methods 9, 478–484. doi: 10.4319/lom.2011.9.478
Shubenkova, O. V., Zemskaya, T. I., Chernitsyna, S. M., Khlystov, O. M., and Triboi, T. I. (2005). The first results of an investigation into the phylogenetic diversity of microorganisms in Southern Baikal sediments in the region of subsurface discharge of methane hydrates. Microbiology 74, 314–320. doi: 10.1007/s11021-005-0069-9
Sinclair, J. L., and Ghiorse, W. C. (1989). Distribution of aerobic bacteria, protozoa, algae and fungi in deep subsurface sediments. Geomicrobiol. J. 7, 15–31. doi: 10.1080/01490458909377847
Sinninghe Damsté, J. S., Kenig, F., Koopmans, M. P., Köster, J., Schouten, S., Hayes, J. M., et al. (1995). Evidence for gammacerane as an indicator of water column stratification. Geochim. Cosmochim. Acta 59, 1895–1900. doi: 10.1016/0016-7037(95)00073-9
Sørensen, K. B., and Teske, A. (2006). Stratified communities of active archaea in deep marine subsurface sediments. Appl. Environ. Microbiol. 72, 4596–4603. doi: 10.1128/aem.00562-06
Story, S. P., Kline, E. L., Hughes, T. A., Riley, M. B., and Hayasaka, S. S. (2004). Degradation of aromatic hydrocarbons by Sphingomonas paucimobilis strain EPA505. Arch. Environ. Con. Tox. 47, 168–176. doi: 10.1007/s00244-004-3069-2
Ward, N. L., Challacombe, J. F., Janssen, P. H., Henrissat, B., Coutinho, P. M., Wu, M., et al. (2009). Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 75, 2046–2056. doi: 10.1128/aem.02294-08
Webster, G., Parkes, R. J., Cragg, B. A., Newberry, C. J., Weightman, A. J., and Fry, J. C. (2006). Prokaryotic community composition and biogeochemical processes in deep subseafloor sediments from the Peru Margin. FEMS Microbiol. Ecol. 58, 65–85. doi: 10.1111/j.1574-6941.2006.00147.x
Winters, A. D., Marsh, T. L., Brenden, T. O., and Faisal, M. (2014). Molecular characterization of bacterial communities associated with sediments in the Laurentian Great Lakes. J. Great Lakes Res. 40, 640–645. doi: 10.1016/j.jglr.2014.04.008
Zemskaya, T., Lomakina, A., Mamaeva, E., Zakharenko, A., Pogodaeva, T., Petrova, D., et al. (2015). Bacterial communities in sediments of Lake Baikal from areas with oil and gas discharge. Aquat. Microb. Ecol. 76, 95–109. doi: 10.3354/ame01773
Zemskaya, T. I., Pogodaeva, T. V., Shubenkova, O. V., Chernitsina, S. M., Dagurova, O. P., Buryukhaev, S. P., et al. (2010). Geochemical and microbiological characteristics of sediments near the Malenky mud volcano (Lake Baikal, Russia), with evidence of Archaea intermediate between the marine anaerobic methanotrophs ANME-2 and ANME-3. Geo Mar. Lett. 30, 411–425. doi: 10.1007/s00367-010-0199-6
Keywords: subsurface biosphere, Lake Baikal, microbial community, methane, gammacerene
Citation: Bukin SV, Pavlova ON, Manakov AY, Kostyreva EA, Chernitsyna SM, Mamaeva EV, Pogodaeva TV and Zemskaya TI (2016) The Ability of Microbial Community of Lake Baikal Bottom Sediments Associated with Gas Discharge to Carry Out the Transformation of Organic Matter under Thermobaric Conditions. Front. Microbiol. 7:690. doi: 10.3389/fmicb.2016.00690
Received: 20 January 2016; Accepted: 26 April 2016;
Published: 10 May 2016.
Edited by:
Baolei Jia, Chung-Ang University, South KoreaReviewed by:
Amy Michele Grunden, North Carolina State University, USATrinity L. Hamilton, University of Cincinnati, USA
Copyright © 2016 Bukin, Pavlova, Manakov, Kostyreva, Chernitsyna, Mamaeva, Pogodaeva and Zemskaya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergei V. Bukin, sergeibukin@lin.irk.ru