 Ana Lokmer1*
Ana Lokmer1* M. Anouk Goedknegt2
M. Anouk Goedknegt2 David W. Thieltges2
David W. Thieltges2 Dario Fiorentino1Sven Kuenzel3
Dario Fiorentino1Sven Kuenzel3 John F. Baines3,4
John F. Baines3,4 K. Mathias Wegner1
K. Mathias Wegner1- 1Coastal Ecology, Wadden Sea Station Sylt, Alfred Wegener Institute - Helmholtz Centre for Polar and Marine Research, List auf Sylt, Germany
- 2Department of Coastal Systems, Royal Netherlands Institute for Sea Research, Utrecht University, Texel, Netherlands
- 3Max Planck Institute for Evolutionary Biology, Plön, Germany
- 4Institute for Experimental Medicine, Christian-Albrechts-Universität zu Kiel, Kiel, Germany
Unveiling the factors and processes that shape the dynamics of host associated microbial communities (microbiota) under natural conditions is an important part of understanding and predicting an organism's response to a changing environment. The microbiota is shaped by host (i.e., genetic) factors as well as by the biotic and abiotic environment. Studying natural variation of microbial community composition in multiple host genetic backgrounds across spatial as well as temporal scales represents a means to untangle this complex interplay. Here, we combined a spatially-stratified with a longitudinal sampling scheme within differentiated host genetic backgrounds by reciprocally transplanting Pacific oysters between two sites in the Wadden Sea (Sylt and Texel). To further differentiate contingent site from host genetic effects, we repeatedly sampled the same individuals over a summer season to examine structure, diversity and dynamics of individual hemolymph microbiota following experimental removal of resident microbiota by antibiotic treatment. While a large proportion of microbiome variation could be attributed to immediate environmental conditions, we observed persistent effects of antibiotic treatment and translocation suggesting that hemolymph microbial community dynamics is subject to within-microbiome interactions and host population specific factors. In addition, the analysis of spatial variation revealed that the within-site microenvironmental heterogeneity resulted in high small-scale variability, as opposed to large-scale (between-site) stability. Similarly, considerable within-individual temporal variability was in contrast with the overall temporal stability at the site level. Overall, our longitudinal, spatially-stratified sampling design revealed that variation in hemolymph microbiota is strongly influenced by site and immediate environmental conditions, whereas internal microbiome dynamics and oyster-related factors add to their long-term stability. The combination of small and large scale resolution of spatial and temporal observations therefore represents a crucial but underused tool to study host-associated microbiome dynamics.
Introduction
Assessing the temporal and spatial stability of microbial communities is vital for understanding and predicting their response to disturbances (Shade et al., 2012) and thus their functioning in a changing environment. This requires knowledge of the underlying disturbance-free community dynamics (Hunt and Ward, 2015). More precisely, it is crucial to identify regular (e.g., daily, seasonal) patterns, normal range of variation in community dynamics, as well as the processes and factors affecting community assembly and structure (Mutshinda et al., 2009; Costello et al., 2012; Nemergut et al., 2013) to establish a baseline against which to measure disturbance effects (Stenuit and Agathos, 2015).
Although controlled repeated-measures experiments in the laboratory (e.g., Shade et al., 2011; Berga et al., 2012; Lokmer and Wegner, 2015) are indispensable for a mechanistic understanding of how environmental factors affect microbial community dynamics, such results may not directly translate to natural conditions, which represent a blend of abiotic and biotic disturbances varying in their intensity, predictability, spatial scale and duration (Bender et al., 1984; Sousa, 1984; Paine et al., 1998; Berga et al., 2012). Studying natural temporal variability of microbial communities represents a valuable complement to controlled experiments, as it provides an opportunity to estimate the effects of known environmental factors and disturbances, as well as to uncover yet unknown and potentially important determinants of community structure and dynamics (Shade et al., 2013; David et al., 2014; Faust et al., 2015). So far, longitudinal studies and time series have helped to elucidate the dynamics of free-living microbial communities ranging from marine sediments (Gobet et al., 2012) and coastal ocean (Gilbert et al., 2012) to soil (Kato et al., 2015) and freshwater habitats (Peura et al., 2015). Similar studies regarding host-associated microbiota are almost exclusively limited to humans, (e.g., Caporaso et al., 2011; David et al., 2014; DiGiulio et al., 2015), and a handful of model organisms (e.g., Fink et al., 2013; Marino et al., 2014). Whereas temporal patterns have been studied in non-model organisms, the focus remains on the population level and the sampling resolution usually coincides with significant host/environment-related shifts: developmental (e.g., Trabal et al., 2012; Hroncova et al., 2015), seasonal (e.g., Zurel et al., 2011; Bjork et al., 2013; Ransome et al., 2014) or abiotic disturbances (e.g., Vega Thurber et al., 2009; Wegner et al., 2013; Tracy et al., 2015). Longitudinal, individual-based, repeated-measure studies remain scarce (but see Pratte et al., 2015; Glasl et al., 2016).
Analogous to longitudinal studies, examining spatial variability and biogeographical patterns over multiple spatial scales, especially if combined with knowledge of environmental gradients, can shed light on relative importance of stochastic and deterministic processes for shaping microbial communities (Green and Bohannan, 2006; Nakaoka et al., 2006; Caruso et al., 2011; Hanson et al., 2012; Borer et al., 2013). One example is the distance-decay relationship, i.e., decreasing similarity between communities with increasing distance, a universal biogeographical pattern that arises through spatially-correlated environmental conditions or through dispersal limitation and has been demonstrated for microbial communities in both marine and terrestrial habitats (Bell, 2010; Martiny et al., 2011; Zinger et al., 2014; Nguyen and Landfald, 2015). Martiny et al. (2011) found that dispersal limitation affected community similarity within salt marshes, whereas environmental factors played more prominent role at regional or continental scale. Conversely, dispersal-related effects in arctic heathland soils were apparent only at larger scales (Hill et al., 2015), illustrating the importance of a particular context for interpretation of the observed patterns.
Spatially stratified sampling strategies can reveal drivers behind the structure and dynamics of free-living microbial communities (e.g., Caruso et al., 2011; Ristova et al., 2015) as well as of host-associated microbiota (Mihaljevic, 2012). In addition to environmental factors (Moro et al., 2011), examining spatial variation can disentangle geographic influences from those of host life stage (Hroncova et al., 2015), genotype and/or diet (Sudakaran et al., 2012; Yatsunenko et al., 2012; Linnenbrink et al., 2013). However, spatial and geographic information has so far primarily served to delineate core microbiomes (e.g., King et al., 2012; Wong et al., 2013; Dishaw et al., 2014) or to differentiate between the microbiomes of closely related species (Zouache et al., 2011; Phillips et al., 2012). This applies especially to marine hosts (e.g., Morrow et al., 2012; Reveillaud et al., 2014; Trabal Fernandez et al., 2014). Studies considering aspects of within-species spatial variation are less common and focused on large-scale differences between environmentally distinct sites (e.g., Trabal et al., 2012; Pierce et al., 2016; Ziegler et al., 2016). Although marine sedentary organisms offer a good opportunity to examine factors and processes shaping dynamics of their associated microbiota over multiple spatial scales using spatially nested designs, this has not been done yet. Including a temporal component into such studies would further improve our understanding of natural microbial community dynamics (e.g., Fortunato et al., 2012; Ransome et al., 2014; Pierce et al., 2016) and thus refine the reference framework for evaluating disturbance effects.
The Pacific oyster (Crassostrea gigas) is such a sedentary organism, highly suitable for the combined estimation of spatial and temporal patterns of microbiome assembly. However, site-specific differences in host-associated microbial communities cannot be separated from host factors by studying natural spatial variability only, as hosts at different sites can be adapted or acclimated to their abiotic and biotic environment (Wendling and Wegner, 2015) or differ due to historical reasons, e.g., invasion histories (Moehler et al., 2011). In contrast to vertebrates, microbiota of most other organisms are closely related to environmental microbial communities (Ley et al., 2008) and translocation experiments with algae (Campbell et al., 2015), and sponges (Burgsdorf et al., 2014) indicate that site is a major determinant of microbiome composition. However, similar experiments with oysters suggest that the influence of site and host factors differs between the tissues (Lokmer et al., 2016). Altogether, combining translocation with a survey of spatial and temporal variation represents a relatively simple means to improve our understanding of the dynamics and function of host-associated microbiota in marine sedentary organisms.
One important function of microbiota that directly contributes to host fitness is defense against pathogens (McFall-Ngai et al., 2013). For example, some of the bacteria inhabiting the oyster hemolymph (a tissue with immune function analogous to vertebrate blood) produce antimicrobial compounds, thus preventing colonization by external pathogens and disease (Defer et al., 2013; Desriac et al., 2014). Hemolymph microbiota can also play part in oyster interactions with abiotic environment (i.e., temperature) by quick (hours to days) adjustments in community composition (Lokmer and Wegner, 2015; Lokmer et al., 2016). Despite the openness of oyster circulatory system and high oyster filtration activity, some bacteria such as Vibrio spp. persist in the hemolymph in the absence of an environmental source population (e.g., if held in sterile seawater) over a range of environmental conditions and could thus be considered resident (Vasconcelos and Lee, 1972; Lokmer et al., 2016). Presence of other, transient bacteria is strictly dependent on the external source community and thus reflects immediate environmental conditions (Lokmer et al., 2016). Dynamics of resident and transient components of the hemolymph microbiome are thus likely shaped by different processes and factors. However, despite its significance for oyster fitness, our knowledge about the variability and dynamics of hemolymph microbiota under natural conditions is almost exclusively limited to a subset of cultivable and potentially pathogenic bacteria, mostly of the genus Vibrio (Garnier et al., 2007; Wendling et al., 2014; Lemire et al., 2015).
In order to examine how site and host genotype affect diversity, composition and dynamics of oyster hemolymph microbiota, we performed a reciprocal translocation experiment with two genetically differentiated oyster populations from two sites in the Wadden Sea (Texel, Netherlands and Sylt, Germany, Moehler et al., 2011), and repeatedly sampled hemolymph from the same individuals over one summer season (Figure 1). Prior to the field deployment, we administered antibiotics to half of the oysters in order to remove a large portion of resident microbiota and account for priority effects. In addition, our field deployment (Figure 1) allowed us to examine spatial variation of complete and resident hemolymph microbiota over small (< 1 m) and medium scales (101–102 m, within site). With such spatially and temporally stratified design we can now try to disentangle the relative contribution of different processes (immigration, within-microbiome interactions), and factors (host genetics, geography, environmental conditions) that shape the oyster hemolymph microbiota under natural conditions.
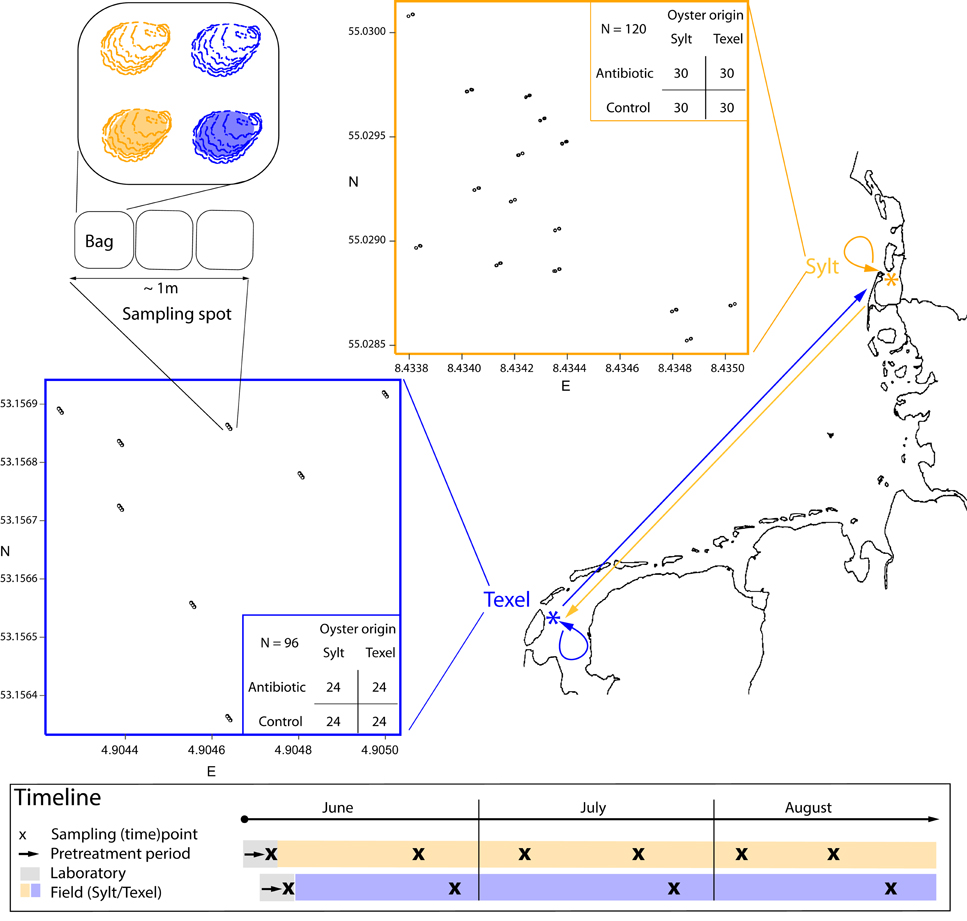
Figure 1. Experimental design and timeline. The spatially stratified sampling design is described on the map. The experimental sites (orange = Sylt, blue = Texel) are marked by an asterisk (*). The arrows on the map link the oyster collection sites with the deployment sites (i.e., half of the oysters were returned to their collection site while the other half was transplanted). The inserted plots show within-site deployment scheme with all sampling spots. Within each sampling spot there are two (Sylt) or three (Texel) bags with four oysters each. Each oyster in the bag belongs to one of the four treatment groups (i.e., antibiotic treatment/control × Sylt/Texel oyster origin). Note that the colors (orange = Sylt, blue = Texel) can denote site or the oyster origin, depending on the context. Tables show the total and the per treatment sample sizes for each site. The box below the map shows the sampling timeline for both sites.
Materials and Methods
The experimental setup and the experiment timeline are shown in Figure 1. The pretreatment, the sampling of the hemolymph and the seawater as well as wet-lab procedures are described in detail in (Lokmer et al., 2016) and therefore only briefly outlined here.
Oyster Collection, Laboratory Pretreatment and Sampling
Oysters from the northern and southern Wadden Sea populations (Moehler et al., 2011) were collected at intertidal mixed oyster/mussel beds at Oddewatt, Sylt, Germany (55° 1′ N, 8° 26′ E) and at de Cocksdorp, Texel, Netherlands (53° 0′ N, 4° 54′ E), respectively. After the removal of epibionts (mainly barnacles) by scrubbing, half of the animals were transported from Sylt to Texel and vice-versa (Figure 1). The laboratory pretreatment was then conducted at the AWI Wadden Sea Station Sylt for the Sylt experiment (i.e., for the oysters later deployed on Sylt) and at the NIOZ Texel for the Texel experiment (i.e., for the oysters later deployed on Texel). For the pretreatment, oysters from each population were divided into two groups: the control group was kept in local 0.2 μm filtered (sterile) seawater, whereas a mix of antibiotics with different mode of action (ampicillin, tetracycline, gentamicin, and kanamycine, Sigma-Aldrich, Hamburg, Germany, final concentration 400 μg/l seawater) was added to the second one to remove as wide range of resident bacteria as possible. After 4 days and prior to the field deployment, hemolymph samples for the analysis of the associated microbial communities were taken with a syringe.
Field Deployment and Sampling
Mesh bags with four oysters each (one per treatment combination: origin × antibiotic) were deployed in groups of two (Sylt) or three (Texel) and fixed with iron rods at original sites of collection on Sylt or Texel (Figure 1). In this way, we could estimate how the spatial scale - within the bag groups/sampling spots (< 1 m) and between the sampling spots (101–102 m) - affects hemolymph microbiota. Hemolymph and seawater samples for the analysis of microbiota were taken directly in the field, placed on ice and immediately frozen upon return to the laboratory. Sampling was performed biweekly on Sylt and monthly on Texel. This, along with some other differences between Sylt and Texel (total number of oysters: 120 on Sylt, 96 on Texel; experiment duration: 1 June—23 August 2012 on Sylt, 14 June—24 August 2012 on Texel) was due to logistic reasons.
DNA Extraction, PCR and Sequence Quality Control and Preprocessing
DNA was extracted from 200 ± 20 μl of hemolymph with Wizard SV 96 Genomic DNA Purification System (Promega, Manheim Germany) and from the rententate obtained by filtering (0.2 μl) of 100 ml seawater with the DNeasy Blood and Tissue Kit, Qiagen, Hilden, Germany. To check for bacterial contamination of reagents, we included additional blank extractions.
PCR (25 μl, 30 cycles, 1 min annealing at 55°C) of the 16S rRNA gene V1-V2 regions, including positive and negative (ddH2O) controls, was performed with equal concentrations of uniquely barcoded 27f and 338r PCR primers (Wang et al., 2015), using 0.5 unit of Phusion Hot Start II High-Fidelity DNA Polymerase per reaction. Equal amounts (estimated by gel electrophoresis and determined fluorometrically) of PCR products were mixed together, purified and sequenced on a Illumina MiSeq platform at the Max Planck Institute for Evolutionary Biology in Plön, Germany.
Sequence quality control and preprocessing was performed as described in Mothur MiSeq SOP (Schloss et al., 2009; Kozich et al., 2013). We defined OTUs (Operational Taxonomic Units) based on a 97% identity threshold. Based on rarefaction curves (not shown), we decided to subsample the dataset to 10,000 reads per sample (6 samples with less than 10,000 were also included, Supplementary Table S1). Due to some oyster mortality, the final dataset comprised of 713 samples in total: 10 seawater, and 703 hemolymph (166 laboratory and 537 field).
Raw demultiplexed reads are deposited at European Nucleotide Archive under the study accession number PRJEB9624.
Statistical Analysis
Statistical analyses were performed in R (R Core Team, 2013). The analysis of α-diversity was based on the Shannon's H index, calculated from the complete subsampled dataset (10,000 reads per sample). We first tested for differences between the seawater and oysters within a site and then between the oysters at the two sites using Asymptotic Wilcoxon Mann-Whitney Rank Sum Test (Wilcoxon RS Test). To assess the effects of oyster origin and antibiotic treatment on α-diversity, as well as its temporal dynamics, we fitted a separate linear mixed model for each site with oyster origin, antibiotic treatment, time and their interactions as fixed factors, and with oyster as a random factor. To test if bag and sampling spot influenced α-diversity, we fitted an additional model for each site excluding the laboratory samples, with oyster, bag and sampling spot as random factors. The models were fitted and tested using the packages lme4 (Bates et al., 2014), lmerTest (Kuznetsova et al., 2014), and MuMIn (Bartoń, 2014).
In our previous association network analysis of hemolymph microbiota (Lokmer et al., 2016), we identified a cluster consisting of the OTUs abundant in the seawater that could be defined as transient. We analyzed all the hemolymph samples in this study in the same way using the igraph package (Csardi and Nepusz, 2006) and again found this transient OTU assemblage (Supplementary Figure S1). To examine how transient microbiota affect β-diversity and distance-decay relationship, we performed the analyses on the complete dataset and excluding the transient OTUs.
For β-diversity, we removed low abundance OTUs (< 10 sequences in the sample) to reduce the dataset complexity (Gobet et al., 2010). The analysis was based on Bray-Curtis distances calculated from hellinger-transformed OTU tables. We used non-metric multidimensional scaling (NMDS) to visualize the overall variability in the dataset (including the seawater samples), and large-scale temporal variability (between sampling points) of hemolymph microbiota. We then analyzed hemolymph communities by constrained analysis of principal coordinates (CAP, Anderson and Willis, 2003), which takes into account only the variability associated with tested predictors, and by Permanova (non-parametric permutational multivariate analysis of variance, Anderson, 2001), using the functions capscale resp. adonis, both implemented in the vegan package (Oksanen et al., 2013). In order to examine how oyster origin, antibiotic treatment and distance affected the β-diversity throughout the summer, we analyzed the hemolymph communities separately at each of the four time-points: before deployment, and once in June, July and August. Although the sampling on Sylt and Texel was not simultaneous, the time difference was at most 10 days and the samples were analyzed together. Two additional sampling points on Sylt were analyzed as well. Variability explained by distance was partialled out prior to plotting CAP results in order to more clearly represent the effects of experimental treatments within sites. To explicitly identify the taxa (from phylum to genus level) contributing to the observed variability, we calculated multivariate generalized (negative binomial) mixed models (GLMs) for each date and for the whole dataset using the mvabund package (Wang et al., 2012).
In order to assess bacterial turnover at a large spatiotemporal scale, we calculated average Bray-Curtis distances between the composite communities at different sampling dates (i.e., all samples from a site at a given date were combined into a single sample) as well as the average individual dissimilarity within sampling dates. To estimate how autocorrelation within oyster individuals influenced community structure and dynamics, we compared the average Bray-Curtis distances between all the samples from the same oyster and between the corresponding number of randomly chosen samples from different oysters. To examine within-individual temporal dynamics, we calculated bacterial turnover within oysters as a proportion of OTUs shared between the initial and subsequent sampling points (Gobet et al., 2012).
The distance-decay relationship was analyzed as described in (Martiny et al., 2011). Briefly, we used 1- Bray-Curtis distance as a measure of similarity and calculated all pairwise distances between the samples from the same sampling date. We then calculated linear models for distance-decay relationship including all samples, as well as for within-spot (up to 1 m) and between-spots (tens of meters) distance ranges separately. In order to estimate how this relationship was affected by transient OTUs, we performed the analysis excluding the seawater OTUs and compared the resulting slopes to the original ones.
Temperature is an important determinant of oyster hemolymph microbiota (Lokmer and Wegner, 2015). The mean temperature experienced by the oysters throughout the experiment was estimated from the Sylt seawater temperature time-series (courtesy of Tatyana Romanova, Wadden Sea Station Sylt, Germany) and from NIOZ Jetty, Texel, Netherlands (van Aken, 2008). On Sylt, on-spot fine-scale temperature measurements were taken during sampling to compare microenvironmental and overall temperature variability.
Results
Hemolymph Microbiota under Laboratory and Field Conditions
During the pre-treatment, the oysters were kept in the laboratory. As laboratory conditions differ substantially from those in the oyster natural environment, we first examined their effect on the hemolymph microbiota. Whereas we found no systematic difference between α-diversity in the laboratory and in the field (Figure 2), NMDS ordination revealed that the laboratory conditions consistently affected hemolymph community composition at both sites, resulting in a clear separation of laboratory and field samples along the first NMDS axis (Figure 3). Laboratory samples were characterized by higher relative abundances of Fusobacteria, ε- and γ-Proteobacteria (mainly Arcobacter and Vibrionaceae), whereas α-Proteobacteria, Tenericutes and an unclassified bacterium related to Spirochaetes were more common in the field (Supplementary Figures S2–S4, Supplementary Table S2). In addition, laboratory communities were clearly separated from the seawater samples, which formed a small group within the cluster of field hemolymph communities (Figure 3). Permanova confirmed these results, showing that 13.9% of the compositional variability could be explained by sample type (hemolymph or seawater) for laboratory communities [F(1, 710) = 115.241, p = 0.001] and only 0.5% for the ones in the field [F(1, 710) = 4.503, p = 0.001]. This pattern reflects the absence of seawater OTUs in laboratory conditions (i.e., 0.2 μm filtered seawater) thus providing support for their transient character (Supplementary Figure S1), while indicating that these OTUs represent a common and important component of hemolymph microbiota under field conditions. Despite the resemblance between the seawater and hemolymph microbiota in the field, significantly higher heterogeneity of oyster-associated communities [Levene's test for homogeneity of multivariate variances, average distance to median: oyster = 0.588, seawater = 0.407, F(1, 546) = 55.9, p < 10−6, effect size = 0.147, Figure 3], implies that the hemolymph microbiota are not a simple reflection of the microbiota in the surrounding environment, but are shaped by other factors as well.
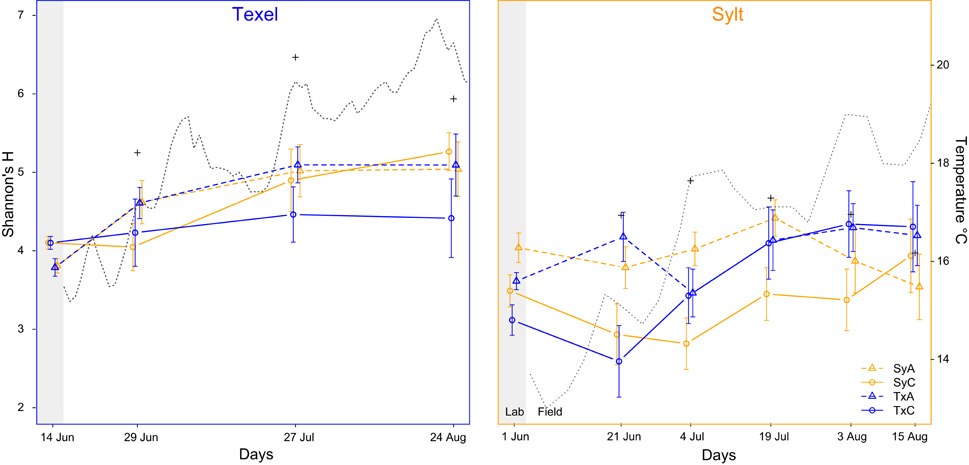
Figure 2. α-diversity of hemolymph and seawater (+) microbiota on Texel (left, blue frame), and Sylt (right, orange frame) throughout the sampling period. The dotted line represents mean daily temperature at the sites. Legend explanation: Sy and Tx refer to the oyster origin, A and C to antibiotic treatment and control, respectively.
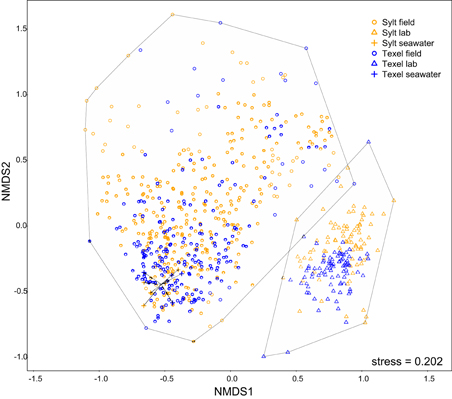
Figure 3. NMDS plot (Bray-Curtis) depicting variability within the complete dataset (all seawater and hemolymph samples). Hulls enclose laboratory and field samples, respectively.
Effects of Experimental Treatments (Translocation and Antibiotics) on Diversity, Composition, and Dynamics of Hemolymph Microbiota
To examine the temporal dynamics of α-diversity and quantify the effects of oyster origin and antibiotic treatment, we analyzed each site separately. On Sylt, a strong initial effect of antibiotic treatment persisted for at least 3 weeks (Figure 2A, main effect of treatment and time x treatment interaction of the Sylt model in Table 1). The initial sorting by antibiotic treatment was reversed toward the beginning of July, when diversity in control translocated oysters increased to match the diversity in their antibiotic-treated counterparts (Figure 2A, origin × treatment × time interaction of Sylt model in Table 1). In local Sylt oysters, the difference between the treatment and control remained visible until late July, when the diversity decreased in the antibiotic-treated animals. On Texel, the initial reduction of community diversity following antibiotic treatment was reversed over time in the field (Figure 2B, time × treatment interaction of the Texel model in Table 1). Although Figure 2B indicates a similar trend for Texel as observed on Sylt (i.e., the tendency of translocated oysters to group according to origin), the interactions between origin, antibiotic treatment and time were not significant in the Texel model.
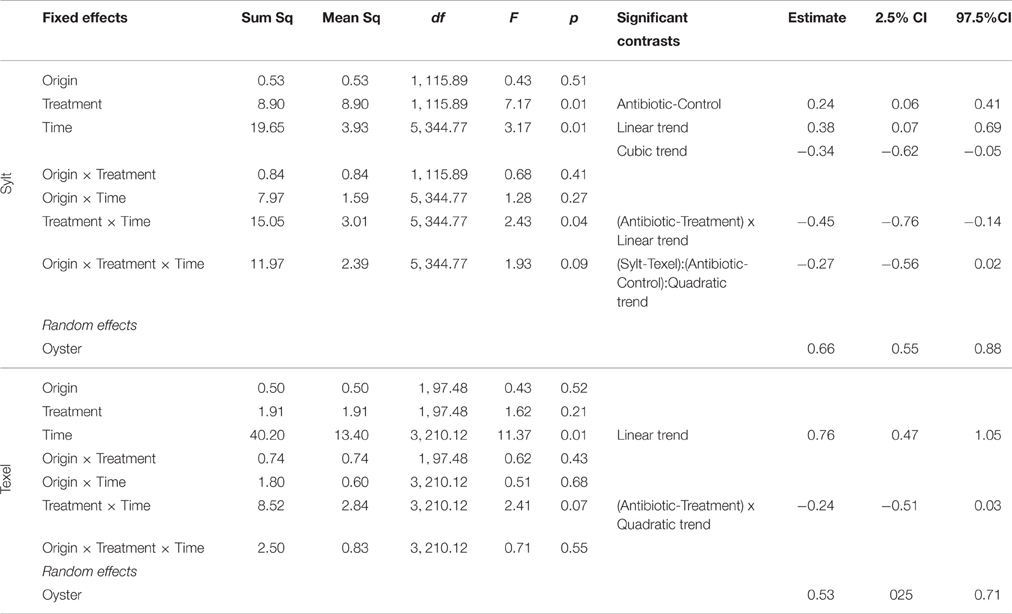
Table 1. Linear mixed models for Shannon's H of hemolymph microbiota on Sylt (dAIC = −6.57, logLik = −673.70 (df = 26), R2 marginal = 0.12, R2 conditional = 0.34), and Texel (dAIC = −21.92, logLik = −472.13 (df = 18), R2 marginal = 0.12, R2 conditional = 0.29) during the summer of 2012.
To explicitly quantify the effects of antibiotics and oyster origin on hemolymph microbial community composition, we analyzed each sampling point (Figure 1) separately. We observed strong initial effects of oyster origin and antibiotic treatment at both locations (Figure 4A, Table 2). Despite the tendency of the samples to separate according to origin along the first CAP axis and according to antibiotic treatment along the second CAP axis, both Figure 4A and significant two- and three-way interactions between the main factors in the Permanova (Table 2) and multivariate GLMs (Supplementary Table S3) imply that the effects of our treatments depended at least partially on the initial community composition and conditions. Two weeks after deployment, the signature of oyster origin was still apparent, but it disappeared soon afterwards (Table 2, Figures 4B–F). Similarly to α-diversity, the effect of antibiotic treatment persisted for a longer time (i.e., until the end of July). However, the variability explained by oyster origin and antibiotic treatment was generally small (1–2%, Table 2), indicating that the hemolymph community structure was largely determined by other factors (e.g., individual and/or microenvironmental variability). As expected, the exclusion of transient OTUs had virtually no influence on the variability explained by experimental treatments, since their presence and abundance should depend on immediate environmental conditions only (Table 2).
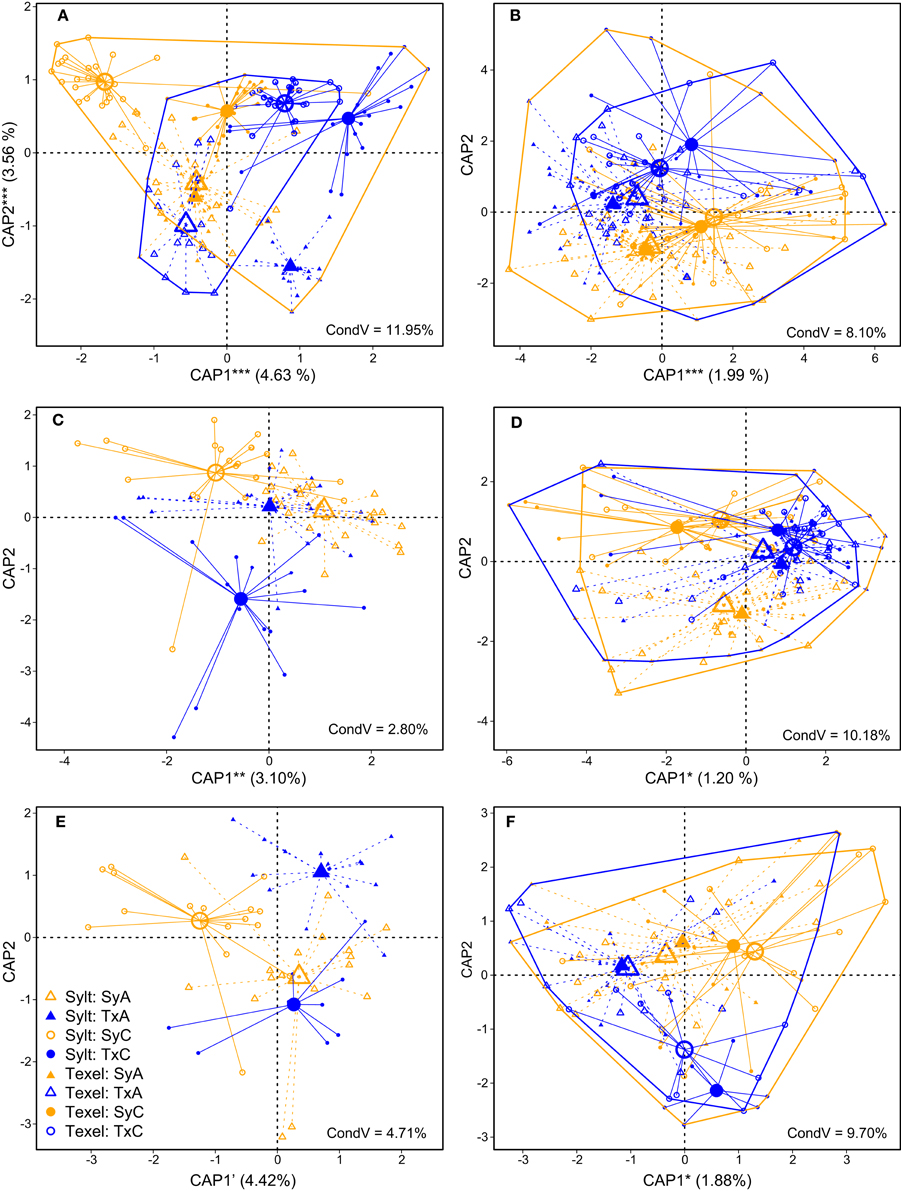
Figure 4. Constrained analysis of principal coordinates (CAP) of β-diversity of hemolymph communities on Sylt and Texel showing the effects of oyster origin and antibiotic treatment after partialling out the effect of distance (variation explained by distance is given in the plots, “CondV”). (A) Pre-deployment, (B) June, (C) July Sylt only, (D) July, (E) August Sylt only, (F) August. The percentages in parentheses represent the variability explained by significant axes. ***p < 0.001, **p < 0.01, *p < 0.05, 'p < 0.1. Hulls enclose all samples from the same site (Sylt = orange, Texel = blue).
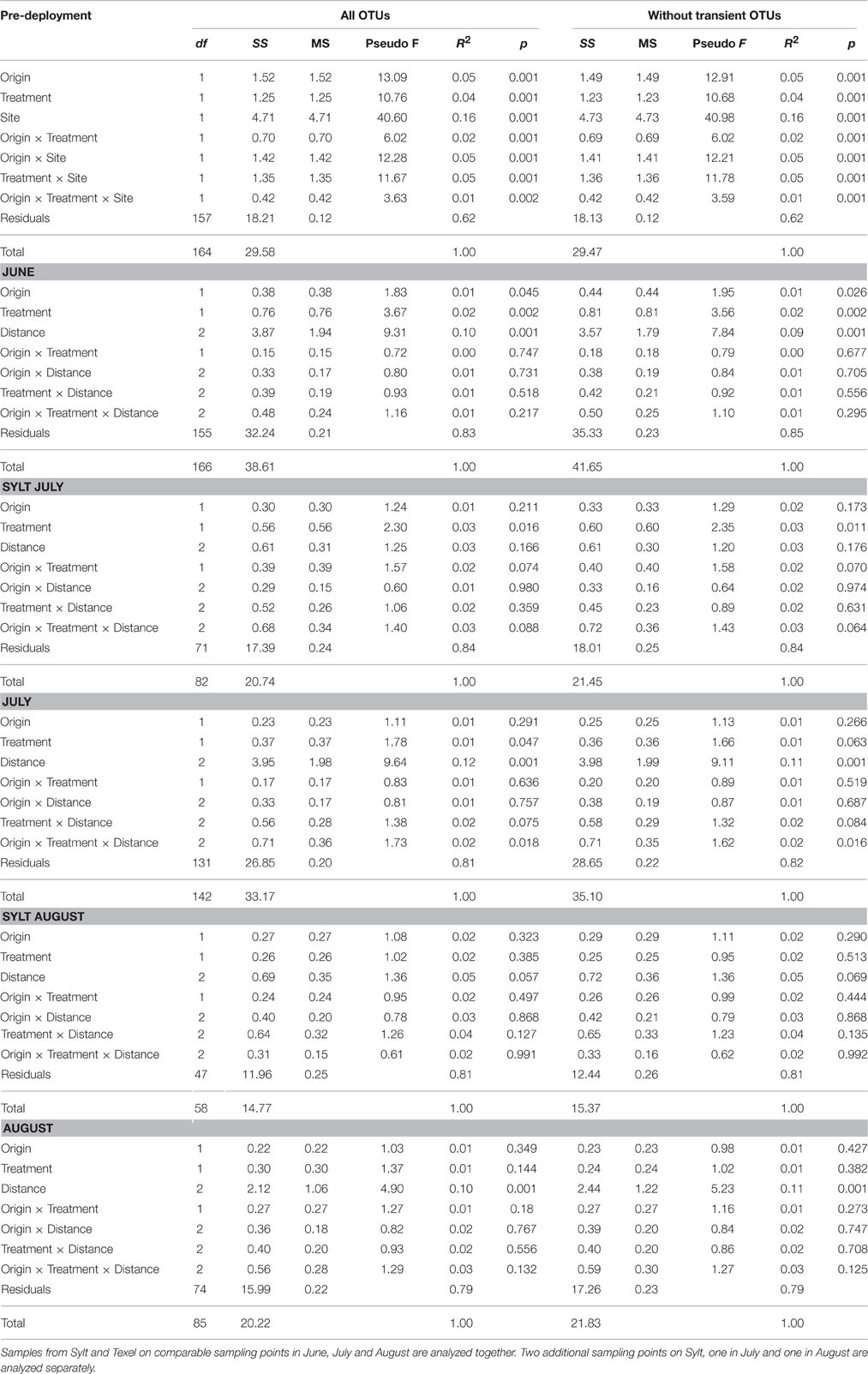
Table 2. Permanova (adonis) showing the effects of oyster origin, antibiotic treatment and distance on hemolymph communities at monthly sampling time points during the experiment.
Spatial Patterns and Dynamics of Hemolymph Microbiota Across Scales
At a large scale, α-diversity was higher on Texel than on Sylt, in the hemolymph (Shannon's H median: Texel = 4.422 (N = 289), Sylt = 3.816 (N = 414); Wilcox RS Test: Z = −7.032, p < 0.001, effect size = −0.265) as well as in the seawater (Shannon's H median: Texel = 6.186 (N = 4), Sylt = 4.468 (N = 6); Wilcox RS Test: Z = −2.559, p = 0.011, effect size = −0.810). Within-site analysis revealed lower diversity in the hemolymph compared to the seawater on Texel (Wilcoxon RS Test: Z = −2.335, p = 0.020, effect size = −0.127), and no differences on Sylt (Wilcoxon RS Test: Z = −0.759, p = 0.448). Similarly, we found a positive correlation between the seawater temperature and the diversity of the hemolymph microbiota on Texel (Kendall's τ = 0.140 ± 0.032, p = 0.01), but not on Sylt (p = 0.388). These discrepancies between the two sites suggest that α-diversity may be influenced by different biotic and abiotic factors at each site. To test for fine-scale spatial influence, we included the bag and the sampling spot as random factors in the α-diversity field-only models (Supplementary Table S4). However, we found no evidence that the spatial proximity resulted in more similar diversity values (Supplementary Table S4, random effects for spot and bag).
Regarding β-diversity, a relatively high amount of variability was explained by distance in all except Sylt-only models (Table 2 and conditional variability in Figure 4), indicating that this distance-related variability primarily reflected the differences between the sites. Multivariate GLMs (Supplementary Tables S5–S8) confirmed the considerable influence of site on community composition. However, the majority of significantly differing taxa were not very abundant (except for the unclassified Spirochaetes-related bacteria; compare univariate significant scores in Supplementary Tables S5–S8 with the taxa depicted in Supplementary Figures S2–S4, where only the taxa with >0.1 relative abundance in at least one oyster group are shown). These rare taxa may thus represent transient microbiota that reflect site-specific environmental conditions, as consistently higher abundance of Cyanobacteria on Sylt or Oceanospirillaceae on Texel might suggest (Supplementary Tables S5–S7).
Despite the differences between Texel and Sylt, the average dissimilarity between the individuals within a site largely exceeded overall dissimilarity between the sites (pairwise Bray-Curtis distance - mean ± SD - between individuals within a site on the same sampling date: Sylt = 0.779 ± 0.105, Texel = 0.751 ± 0.110; between Sylt and Texel composite communities on the same date = 0.490 ± 0.047). This high between-individual variability might suggest the important role of factors such as oyster genotype, physiology, condition and microenvironmental heterogeneity.
Although the community composition varied widely at each spatial scale, we detected a negative correlation between community similarity and geographic distance (Figure 5, overall distance-decay slope: b = −0.024, p < 0.001). The relationship was significantly stronger over small (up to 1 m, within spot: b = −0.102, p < 0.001) and intermediate (between spots, up to 186 m: b = −0.113, p < 0.001) spatial scales. Exclusion of transient OTUs affected neither overall (Figure 6, b = −0.027, p < 0.001) nor the small-scale distance-decay relationship (b = −0.082, p < 0.001). On the other hand, it flattened the distance-decay slope at intermediate spatial scale (Figure 6, b = −0.034, p < 0.001), suggesting that the transient OTUs could indeed reflect the higher probability of adjacent spots to experience similar environmental conditions during immersion. However, the same analysis performed for each month separately revealed that the distance-decay relationship varied over time (Supplementary Figure S4). The overall results were driven by patterns observed in June and July, whereas in August both large- and intermediate-scale slopes were steeper if transient OTUs were excluded. In fact, in August we found no significant distance-decay relationship at intermediate scale with the transient OTUs included (p = 0.065) or at the small scale regardless of the microbial community portion considered (complete: p = 0.167, without transient: p = 0.169).
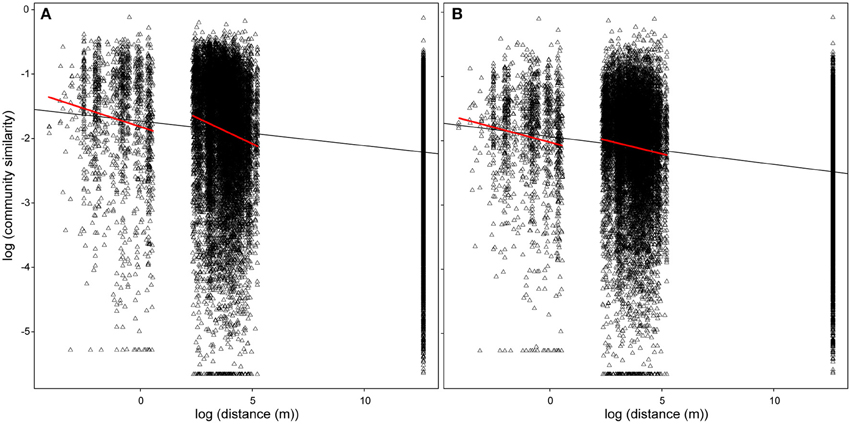
Figure 5. Distance-decay relationship including (A) all OTUs and (B) resident (excluding seawater) OTUs. The lines represent linear models fitted for the given spatial scale.
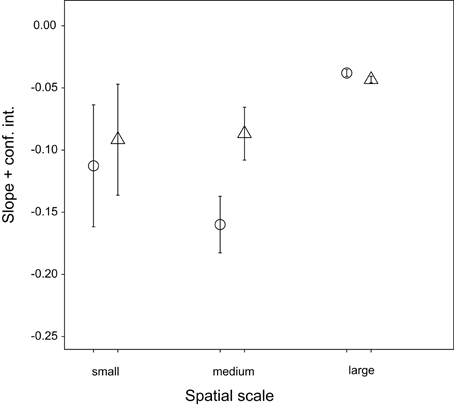
Figure 6. Effect of transient OTUs on slope of distance-decay relationship on small, medium and overall spatial scale. ○ = all OTUs, △ = without transient.
Temporal Dynamics of Hemolymph Microbiota: Sites and Individuals
Overall, α-diversity increased over the course of the experiment, with a clearer and more pronounced effect on Texel (Figure 2, Table 1, main effect of time in both Sylt and Texel model). Including the oyster individual as a random effect into the α-diversity models substantially increased the amount of explained variability, as illustrated by the difference between marginal (fixed effects only) and conditional (including random factors) R2 in the legend to Table 1. Thus, the factors like oyster genotype, but also historical contingency might play an important role in shaping the within-diversity of hemolymph microbiota.
Similar to spatial patterns, the analysis of large-scale temporal dynamics of β-diversity revealed relatively high degree of stability over the examined period (mean ± SD Bray-Curtis distance between composite communities on individual sampling dates: Sylt = 0.425 ± 0.060, Texel = 0.447 ± 0.068), with NMDS plot showing a slight tendency of hemolymph communities sampled in August to cluster separately from the rest at both sites (Supplementary Figure S5). However, the August samples were indistinguishable from the June and July samples down to the genus level (Supplementary Figure S3), indicating that the shift occurred at the OTU level. This overall temporal stability likely resulted from relatively stable overall environmental conditions throughout the sampling period (Supplementary Figure S6). On the other hand, temporal variability at the individual scale was substantial (mean ± SD Bray-Curtis distance = 0.818 ± 0.140), probably reflecting highly dynamic microenvironmental conditions experienced by the oysters at short timescales. Still, albeit high, the within-oyster temporal variability was smaller than the among-oyster variation (mean ± SD Bray-Curtis distance among oysters = 0.876 ± 0.105, Wilcox RS test: Z = −9.283, p < 10−6), implying that the oyster-related factors such as genotype as well as internal community dynamics and priority effects shape the composition of the hemolymph microbiota. In addition, high but constant within-individual turnover rate between consecutive sampling times suggests non-directional fluctuations in community composition during the sampling period, i.e., repeated occurrence of at least some taxa (Figure 7).
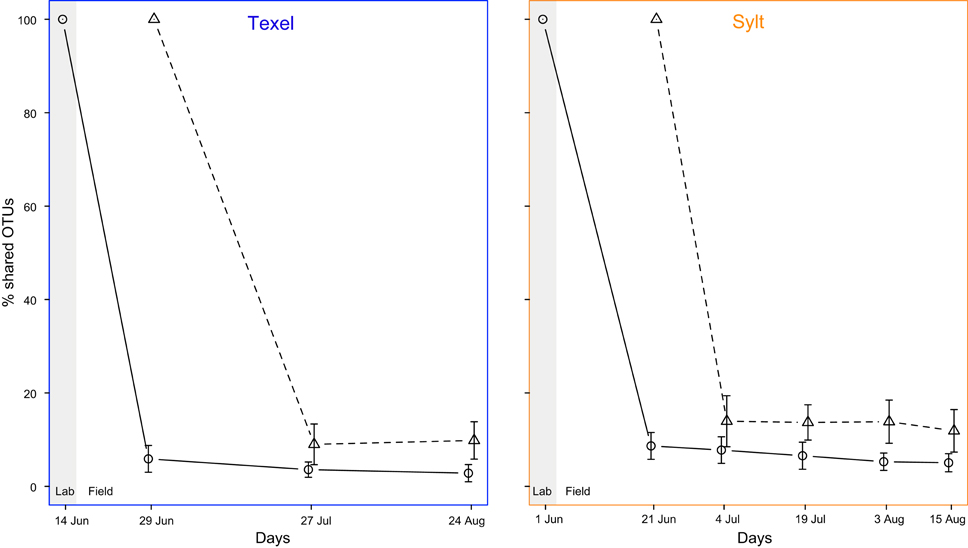
Figure 7. Bacterial community turnover, i.e., the percentage of OTUs shared between the samples from the same individual at the initial and the subsequent sampling points (○ = all samples, △ = laboratory samples excluded).
Discussion
Animal fitness is inextricably linked to the stability of its associated microbiota. An essential prerequisite for assessing the stability is a thorough understanding of factors and processes shaping the microbial community structure and dynamics. To identify these and their relative importance under natural conditions can be challenging, especially in a highly variable environment, such as the intertidal. Here, we combined experimental manipulation with a field survey of temporal and spatial patterns in Pacific oyster hemolymph microbiota over multiple scales to improve our understanding of their dynamics in complex natural environments. Our observations revealed high small-scale spatial variability in the field. This, together with pronounced differences between microbial community composition in the laboratory and in the field, implies a quick response of the hemolymph microbiota to large shifts in environmental conditions. Although environmentally-dependent acquisition/loss of transient microbiota indicate an important role of colonization by exogenous microbes, the persistent effects of translocation and antibiotic treatment together with recognizable individual temporal dynamics suggest that the community structure and dynamics are also influenced by host-related factors as well as by biotic interactions within the microbiome.
Resident and Transient Hemolymph Microbiota under Laboratory and Natural Conditions
Although common in oyster hemolymph under field conditions, seawater OTUs were virtually absent from the hemolymph-associated communities in the laboratory, where the oysters were kept in the filtered seawater. Dependence on the environmental source population implies that these OTUs could be considered transient (Vasconcelos and Lee, 1972). As such, they are expected to strongly depend on the immediate environmental conditions, in this case immersion during the tidal cycle in the field (Lokmer et al., 2016) and thus should not (and did not) reflect the long-term effects of our experimental treatments (Table 2, compare the variability explained by the treatments including/excluding transient microbiota). In addition, the strong effect of transient microbiota on the distance-decay slope over microenvironmentally variable scales, i.e., between the sampling spots (Figure 5), provides further support for the link between the immersion and the dynamics of transient bacteria. Explicitly, whereas the abiotic conditions experienced by oysters are very similar within the sampling spots (< 1 m), the factors such as immersion time and tidal currents likely differ between the spots in a distance-dependent manner, subsequently affecting the dynamics of the transient microbiota and the distance-decay slope.
Whereas our results clearly support the classification of seawater OTUs as transient, it is less clear which bacteria should be considered resident. For example, an unindentified bacterium close to Spirochaetes and a Tenericutes OTU were both abundant in the field and rare in the laboratory, indicating they might be transient. However, these bacteria have been previously found in Pacific oysters in Tasmania (Fernandez-Piquer et al., 2012), suggesting their affinity to form associations with oysters. Both Spirochaetes and Tenericutes are commonly isolated from various oyster tissues (Prieur et al., 1990; Green and Barnes, 2010; Husmann et al., 2010; King et al., 2012; Trabal et al., 2012; Wegner et al., 2013; Trabal Fernandez et al., 2014; Lokmer et al., 2016). However, Spirochaetes and Tenericutes are rare in the hemolymph in the laboratory (Lokmer and Wegner, 2015; Lokmer et al., 2016), where high abundance of Tenericutes is mainly linked to stress or even mortality (Lokmer and Wegner, 2015). Increased abundance of Tenericutes in the field observed here could thus have been due to a secondary infection of the injection site on the adductor muscle caused by repeated hemolymph sampling (Ayling et al., 2011). On the other hand, potential benefits that Tenericutes and Spirochaetes provide to their hosts would likely be nutrition-related (Prieur et al., 1990; Tanaka et al., 2004; Fraune and Zimmer, 2008) and the decrease in their abundance in the laboratory might have been linked to starvation, as oysters were not fed during the pretreatment period (Green and Barnes, 2010). Similarly, high abundance of Vibrio and Arcobacter commonly found in the laboratory (Lokmer and Wegner, 2015; Lokmer et al., 2016) could result from stationary conditions and represent a pre-disease state (Petton et al., 2015a). However, these bacteria are also commonly isolated in the field (Garnier et al., 2007; Wendling et al., 2014; Lokmer et al., 2016) and may play role in pathogen defense and acclimation (Defer et al., 2013; Lokmer and Wegner, 2015). Due to geographical (Petton et al., 2015b) and seasonal (Wegner et al., 2013; Wendling et al., 2014; Pierce et al., 2016) differences in dynamics of these potential hemolymph residents, further studies addressing large-scale spatial and temporal variation of hemolymph microbiota, their function as well as the factors affecting their interactions with oysters are needed (Pruzzo et al., 2005; Aagesen et al., 2013).
Variation of Oyster Microbiota Related to Origin and Antibiotic Treatment
Weeks-lasting effects of antibiotic treatment and oyster origin imply gradual turnover of resident microbiota and demonstrate the existence of internal community dynamics and importance of historical contingencies (Nemergut et al., 2013). Although antibiotics can have long-term negative effects on diversity in some cases (Stein et al., 2013), they increase diversity in others, probably due to the increased invasion susceptibility of the affected communities (Shea et al., 2004; Robinson et al., 2010). In relatively stable and isolated environments, such as mammalian gut, antibiotic-induced changes may induce permanent shifts resulting in alternative stable states (Stein et al., 2013). Oyster hemolymph, on the other hand, is a highly variable habitat closely connected with the external environment, and thus the establishment of such stable states in the associated microbial community is highly unlikely.
Genetic differentiation between oysters from Texel and Sylt (Moehler et al., 2011) could have contributed to the observed differences in β-diversity since oyster microbiota can assemble according to host genotype (Wegner et al., 2013). In addition, many Vibrio spp. are pathogenic and oyster populations rapidly adapt to their local Vibrio species (Rosa et al., 2012; Wendling and Wegner, 2015), suggesting that at least parts of hemolymph microbiota are affected by host genotype. However, the gradually decreasing difference between translocated and local oysters at both sites implies that the divergence in community composition on the host origin level was in the long run mainly affected by site, a common pattern found in marine sedentary animals (Burgsdorf et al., 2014; Lear et al., 2014; Campbell et al., 2015). Gradual turnover of resident bacteria following translocation has previously been demonstrated for Vibrio spp. populations in the oyster hemolymph (Wendling et al., 2014). Interestingly, the diversity of hemolymph microbial communities in control translocated oysters did not initially differ from their local counterparts, but increased during July at both sites, matching the timespan reported for the Vibrio spp. turnover (Wendling et al., 2014). This effect was only marginally significant, but it is tempting to speculate that this trend might have been linked to inability of translocated-oyster immune system to control the resident microbiota acquired from the new environment, resulting in more bacteria evading the oyster immune defenses and establishing in the hemolymph.
Close contact of the hemolymph with the external environment might dampen genotype-specific community assembly as opposed to other tissues more shielded from the environment (Wegner et al., 2013; Lokmer et al., 2016) as well as prevent the evolution of specialist hemolymph symbionts (Preheim et al., 2011). Nevertheless, the seawater and the coastal sediments are characterized by seasonally recurring bacterial populations (Gilbert et al., 2012; Gobet et al., 2012), likely resulting in predictable encounters between the oysters, their resident bacteria and external microbiota. Unpredictable disturbances to any component of that system, such as translocation or antibiotic treatment here, might influence the community structure and dynamics and subsequently affect oyster fitness (Lokmer et al., 2016).
Hemolymph Microbiota across Temporal and Spatial Scales
At coarse temporal and spatial resolution, the hemolymph microbiota appeared relatively stable throughout the sampling period and we found little variation in dominant taxa (Supplemetary Figures S2, S3, Supplementary Tables S2, S5–S7). Previous studies have demonstrated a strong effect of temperature on the hemolymph microbiota (Lokmer and Wegner, 2015), but have also shown that the community structure in the natural conditions exhibits a seasonal pattern and does not respond to quick temperature shifts (Wendling et al., 2014). In addition, we can show that hemolymph microbiota are also affected by the immediate external microbial environment (Lokmer et al., 2016). Therefore, high large-scale spatiotemporal stability can be explained by stable mean temperature throughout the sampling period at both sites (Figure 2, Supplementary Figure S6) as well as by seasonally stable microbial communities in oyster surroundings, namely in the sediments and the seawater (Campbell et al., 2011; Gilbert et al., 2012; Gobet et al., 2012).
Nevertheless, we detected differences, albeit rather small and mostly constrained to less abundant phylotypes (Supplementary Tables S5–S7), between the sites in both diversity and composition. Apart from dispersal limitation, regarding primarily OTUs within the dominant lineages, these differences might have been related to environmental factors such as smaller sediment grain size at Texel, which could affect the structure of associated microbial communities (Jackson and Weeks, 2008; Legg et al., 2012) in the environment and also result in higher number of suspended particles in the seawater with consequences for the oyster filtering activity (Riisgard, 1988; Frechette et al., 2016) and microbiota.
However, it is important to remember that, although the 16S rRNA gene and its fragments represent an important tool for understanding of microbial communities, they lack resolution power. In addition, taxonomy is only partially consistent with ecology (Koeppel and Wu, 2012) and allows solely for distinction between broad habitat types (Schmidt et al., 2014). 16S rRNA defined OTUs may consist of variety of ecotypes, and, in case of host-associated bacteria, they can significantly differ in crucial traits such as virulence (Koeppel and Wu, 2013; Lemire et al., 2015; Wendling and Wegner, 2015). Moreover, closely related bacteria exhibit adaptation at very small spatial scales (Belotte et al., 2003). Therefore, although the communities at both sites and throughout the summer appear similar through the lens of 16S rDNA based taxonomy, they can actually consist of ecologically different bacteria with important consequences for hemolymph microbiota dynamics and their oyster hosts.
Interestingly, in August we observed both a slight shift in community composition as well as a change of the distance-decay relationship. It remains unclear whether there is a link between the both, but the observed changes might have been related to spawning, which occurred during August at both sites (according to observations of spat size in autumn). Spawning represents a stressful period in the oyster lifecycle, increasing the susceptibility to pathogens and affecting the composition of associated Vibrio communities (Wendling et al., 2014) and likely of other oyster-associated bacterial populations.
In contrast to large-scale stability, the within-individual temporal variability and the between-individual variability at small spatial scale were high, likely reflecting microenvironmental spatiotemporal heterogeneity. Virtual lack of directionality in large-scale dynamics in combination with this high small-scale variability suggests that the latter could be related to extreme but periodic environmental fluctuations in the intertidal. Namely, the quick response of the hemolymph microbiota to such fluctuations (Lokmer and Wegner, 2015; Lokmer et al., 2016) may result in pronounced but predictable (cyclic) dynamics, as bacteria disappear from the hemolymph or fall below the detection limit (Caporaso et al., 2012; Shade et al., 2013, 2014), and re-colonize the oyster or increase in abundance when the conditions are right again. High but constant turnover rate (Figure 6) indeed suggests that bacterial populations may disappear or become very rare, but reappear at a later stage (Gobet et al., 2012). In addition to strong influence of the environment, factors such as oyster genotype, physiology and health condition are also likely to affect the structure of the hemolymph microbiome (Wegner et al., 2013; Lokmer and Wegner, 2015), accounting for the pronounced differences between the oysters and at the same time for the consistent temporal dynamics within individuals (i.e., large portion of α-diversity variability explained by individual).
Overall, our results confirm that temporal (Shade et al., 2013) and spatial (Martiny et al., 2011; Borer et al., 2013; O'Brien et al., 2016) scale strongly affect inference about community stability and dynamics (Faust et al., 2015). High perceived temporal stability of microbiota associated with the subtidal sessile marine invertebrates (Erwin et al., 2012; Bjork et al., 2013; Pita et al., 2013; Hardoim and Costa, 2014) is often related be low temporal and/or taxonomic resolution. While the analysis of composite communities reveals influences of large-scale environmental and host factors (e.g., site, season, tissue type), focusing on small-scale individual dynamics is necessary for deciphering host-microbiota interactions and thus for understanding their role for animal survival.
Conclusion
Our study aimed to examine the dynamics of Pacific oyster hemolymph microbiota under natural conditions. By switching the focus between large- and small-scale temporal and spatial variation, we identified potentially important factors and processes shaping the hemolymph microbiome. High small-scale variability (within-site or within-individual) likely reflects microenvironmental heterogeneity as well as host genetic differences, with the range of variability determined by large-scale mean abiotic conditions and internal microbiome interactions.
As drivers of host-associated community dynamics are numerous and act in a scale-dependent manner, the appropriate scale for investigations depends on the questions that one aims to address. Spatially stratified sampling designs and the analysis of individual and population temporal dynamics provide useful hints for choosing the adequate resolution. In this way one can also design experiments that will more closely mimic characteristics of the natural environment crucial for dynamics and assembly of host-associated microbiota and thus contribute to elucidating their role for animal fitness.
Author Contributions
AL planned and conducted the experiment, collected and analyzed the data and wrote the manuscript. AG and DT collected the data and critically revised the manuscript. DF took part in the analysis and interpretation of the data and critically revised the manuscript, SK collected the data, JB critically revised the manuscript, KW planned the experiment and wrote the manuscript.
Funding
This study was supported by the DFG (Deutsche Forschungsgemeinschaft) Emmy Noether Programme (We4641/1-3), Excellence Cluster 306 “Inflammation at Interfaces,” the Netherlands Organization for Scientific Research (NWO) and the German Federal Ministry of Education and Research (BMBF, NWO-ZKO project 839.11.002) and the International Max Research School for Evolutionary Biology. All procedures were performed according to national and European law and experiments were approved by the local authorities.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Mirjana Markovic, Jennifer Welsh, Anne-Karin Schuster and Cátia Carreira for their help with hemolymph sampling at the site on Texel, Silke Vollbrecht und Franziska Schade for assisting with laboratory work on Sylt, Tobias Mayr and Kaibil Escobar Wolf for cleaning the oysters and, Katja Cloppenborg-Schmidt at the Evolutionary Genomics group at Kiel University for assistance with the preparation of MiSeq libraries, Tatyana Romanova for Sylt temperature data and WaLTER for the NIOZ Jetty data.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01367
References
Aagesen, A. M., Phuvasate, S., Su, Y. C., and Hase, C. C. (2013). Persistence of Vibrio parahaemolyticus in the Pacific oyster, Crassostrea gigas, is a multifactorial process involving pili and flagella but not type III secretion systems or phase variation. Appl. Environ. Microbiol. 79, 3303–3305. doi: 10.1128/AEM.00314-13
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1046/j.1442-9993.2001.01070.x
Anderson, M. J., and Willis, T. J. (2003). Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84, 511–525. doi: 10.1890/0012-9658(2003)084[0511:CAOPCA]2.0.CO;2
Ayling, R. D., Bashiruddin, S., Davison, N. J., Foster, G., Dagleish, M. P., and Nicholas, R. A. (2011). The occurrence of mycoplasma phocicerebrale, mycoplasma phocidae, and mycoplasma phocirhinis in grey and common seals (Halichoerus grypus and Phoca vitulina) in the United Kingdom. J. Wildl. Dis. 47, 471–475. doi: 10.7589/0090-3558-47.2.471
Bates, D. M. M Bolker, B., and Walker, S. (2014). lme4: Linear Mixed-Effects Models Using Eigen and S4. Submitted to Journal of Statistical Software.
Bell, T. (2010). Experimental tests of the bacterial distance-decay relationship. ISME J. 4, 1357–1365. doi: 10.1038/ismej.2010.77
Belotte, D., Curien, J. B., Maclean, R. C., and Bell, G. (2003). An experimental test of local adaptation in soil bacteria. Evolution 57, 27–36. doi: 10.1111/j.0014-3820.2003.tb00213.x
Bender, E. A., Case, T. J., and Gilpin, M. E. (1984). Perturbation experiments in community ecology - theory and practice. Ecology 65, 1–13. doi: 10.2307/1939452
Berga, M., Székely, A. J., and Langenheder, S. (2012). Effects of disturbance intensity and frequency on bacterial community composition and function. PLoS ONE 7:e36959. doi: 10.1371/journal.pone.0036959
Björk, J. R., Díez-Vives, C., Coma, R., Ribes, M., and Montoya, J. M. (2013). Specificity and temporal dynamics of complex bacteria–sponge symbiotic interactions. Ecology 94, 2781–2791. doi: 10.1890/13-0557.1
Borer, E. T., Kinkel, L. L., May, G., and Seabloom, E. W. (2013). The world within: Quantifying the determinants and outcomes of a host's microbiome. Basic Appl. Ecol. 14, 533–539. doi: 10.1016/j.baae.2013.08.009
Burgsdorf, I., Erwin, P. M., López-Legentil, S., Cerrano, C., Haber, M., Frenk, S., and et al. (2014). Biogeography rather than association with cyanobacteria structures symbiotic microbial communities in the marine sponge Petrosia ficiformis. Front. Microbiol. 5:529. doi: 10.3389/fmicb.2014.00529
Campbell, A. H., Marzinelli, E. M., Gelber, J., and Steinberg, P. D. (2015). Spatial variability of microbial assemblages associated with a dominant habitat-forming seaweed. Front. Microbiol. 6:230. doi: 10.3389/fmicb.2015.00230
Campbell, B. J., Yu, L., Heidelberg, J. F., and Kirchman, D. L. (2011). Activity of abundant and rare bacteria in a coastal ocean. Proc. Natl. Acad. Sci. U.S.A. 108, 12776–12781. doi: 10.1073/pnas.1101405108
Caporaso, J. G., Lauber, C. L., Costello, E. K., Berg-Lyons, D., Gonzalez, A., Stombaugh, J., and et al. (2011). Moving pictures of the human microbiome. Genome Biol. 12:R50. doi: 10.1186/gb-2011-12-5-r50
Caporaso, J. G., Paszkiewicz, K., Field, D., Knight, R., and Gilbert, J. A. (2012). The western english channel contains a persistent microbial seed bank. ISME J. 6, 1089–1093. doi: 10.1038/ismej.2011.162
Caruso, T., Chan, Y., Lacap, D. C., Lau, M. C., McKay, C. P., and Pointing, S. B. (2011). Stochastic and deterministic processes interact in the assembly of desert microbial communities on a global scale. ISME J. 5, 1406–1413. doi: 10.1038/ismej.2011.21
Costello, E. K., Stagaman, K., Dethlefsen, L., Bohannan, B. J. M., and Relman, D. A. (2012). The application of ecological theory toward an understanding of the human microbiome. Science 336, 1255–1262. doi: 10.1126/science.1224203
Csardi, G., and Nepusz, T. (2006). The igraph software package for complex network research. Int. J. Complex Syst. 1695.
David, L. A., Materna, A. C., Friedman, J., Campos-Baptista, M. I., Blackburn, M. C., Perrotta, A., and et al. (2014). Host lifestyle affects human microbiota on daily timescales. Genome Biol. 15:R89. doi: 10.1186/gb-2014-15-7-r89
Defer, D., Desriac, F., Henry, J., Bourgougnon, N., Baudy-Floc'h, M., Brillet, B., and et al. (2013). Antimicrobial peptides in oyster hemolymph: the bacterial connection. Fish Shellfish Immunol. 34, 1439–1447. doi: 10.1016/j.fsi.2013.03.357
Desriac, F., Le Chevalier, P., Brillet, B., Leguerinel, I., Thuillier, B., Paillard, C., and et al. (2014). Exploring the hologenome concept in marine bivalvia: haemolymph microbiota as a pertinent source of probiotics for aquaculture. FEMS Microbiol. Lett. 350, 107–116. doi: 10.1111/1574-6968.12308
DiGiulio, D. B., Callahan, B. J., McMurdie, P. J., Costello, E. K., Lyell, D. J., Robaczewska, A., and et al. (2015). Temporal and spatial variation of the human microbiota during pregnancy. Proc. Natl. Acad. Sci. U.S.A. 112, 11060–11065. doi: 10.1073/pnas.1502875112
Dishaw, L. J., Flores-Torres, J., Lax, S., Gemayel, K., Leigh, B., Melillo, D., and et al. (2014). The gut of geographically disparate Ciona intestinalis harbors a core microbiota. PLoS ONE 9:e93386. doi: 10.1371/journal.pone.0093386
Erwin, P. M., Pita, L., Lopez-Legentil, S., and Turon, X. (2012). Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 78, 7358–7368. doi: 10.1128/AEM.02035-12
Faust, K., Lahti, L., Gonze, D., de Vos, W. M., and Raes, J. (2015). Metagenomics meets time series analysis: unraveling microbial community dynamics. Curr. Opin. Microbiol. 25, 56–66. doi: 10.1016/j.mib.2015.04.004
Fernandez-Piquer, J., Bowman, J. P., Ross, T., and Tamplin, M. L. (2012). Molecular analysis of the bacterial communities in the live Pacific oyster (Crassostrea gigas) and the influence of postharvest temperature on its structure. J. Appl. Microbiol. 112, 1134–1143. doi: 10.1111/j.1365-2672.2012.05287.x
Fink, C., Staubach, F., Kuenzel, S., Baines, J. F., and Roeder, T. (2013). Noninvasive analysis of microbiome dynamics in the fruit fly Drosophila melanogaster. Appl. Environ. Microbiol. 79, 6984–6988. doi: 10.1128/AEM.01903-13
Fortunato, C. S., Herfort, L., Zuber, P., Baptista, A. M., and Crump, B. C. (2012). Spatial variability overwhelms seasonal patterns in bacterioplankton communities across a river to ocean gradient. ISME J. 6, 554–563. doi: 10.1038/ismej.2011.135
Fraune, S., and Zimmer, M. (2008). Host-specificity of environmentally transmitted Mycoplasma-like isopod symbionts. Environ. Microbiol. 10, 2497–2504. doi: 10.1111/j.1462-2920.2008.01672.x
Frechette, M., Urquiza, J. M., Daigle, G., and Rioux-Gagnon, D. (2016). Clearance rate regulation in mussels: adding the effect of seston level to a model of internal state-based regulation. J. Exp. Mar. Biol. Ecol. 475, 1–10. doi: 10.1016/j.jembe.2015.11.003
Garnier, M., Labreuche, Y., Garcia, C., Robert, M., and Nicolas, J. L. (2007). Evidence for the involvement of pathogenic bacteria in summer mortalities of the Pacific oyster Crassostrea gigas. Microb. Ecol. 53, 187–196. doi: 10.1007/s00248-006-9061-9
Gilbert, J. A., Steele, J. A., Caporaso, J. G., Steinbrück, L., Reeder, J., Temperton, B., and et al. (2012). Defining seasonal marine microbial community dynamics. ISME J. 6, 298–308. doi: 10.1038/ismej.2011.107
Glasl, B., Herndl, G. J., and Frade, P. R. (2016). The microbiome of coral surface mucus has a key role in mediating holobiont health and survival upon disturbance. ISME J. 10, 2280–2292. doi: 10.1038/ismej.2016.9
Gobet, A., Böer, S. I., Huse, S. M., van Beusekom, J. E., Quince, C., Sogin, M. L., and et al. (2012). Diversity and dynamics of rare and of resident bacterial populations in coastal sands. ISME J. 6, 542–553. doi: 10.1038/ismej.2011.132
Gobet, A., Quince, C., and Ramette, A. (2010). Multivariate Cutoff Level Analysis (MultiCoLA) of large community data sets. Nucleic Acids Res. 38, e155. doi: 10.1093/nar/gkq545
Green, J., and Bohannan, B. J. M. (2006). Spatial scaling of microbial biodiversity. Trends Ecol. Evol. 21, 501–507. doi: 10.1016/j.tree.2006.06.012
Green, T. J., and Barnes, A. C. (2010). Bacterial diversity of the digestive gland of Sydney rock oysters, Saccostrea glomerata infected with the paramyxean parasite, Marteilia sydneyi. J. Appl. Microbiol. 109, 613–622. doi: 10.1016/J.Tree.2006.06.012
Hanson, C. A., Fuhrman, J. A., Horner-Devine, M. C., and Martiny, J. B. (2012). Beyond biogeographic patterns: processes shaping the microbial landscape. Nat. Rev. Microbiol. 10, 497–506. doi: 10.1038/nrmicro2795
Hardoim, C. C., and Costa, R. (2014). Temporal dynamics of prokaryotic communities in the marine sponge Sarcotragus spinosulus. Mol. Ecol. 23, 3097–3112. doi: 10.1111/mec.12789
Hill, R., Saetnan, E. R., Scullion, J., Gwynn-Jones, D., Ostle, N., and Edwards, A. (2015). Temporal and spatial influences incur reconfiguration of Arctic heathland soil bacterial community structure. Environ. Microbiol. 18, 1942–1953. doi: 10.1111/1462-2920.13017
Hroncova, Z., Havlik, J., Killer, J., Doskocil, I., Tyl, J., Kamler, M., and et al. (2015). Variation in honey bee gut microbial diversity affected by ontogenetic stage, age and geographic location. PLoS ONE 10:e0118707. doi: 10.1371/journal.pone.0118707
Hunt, D. E., and Ward, C. S. (2015). A network-based approach to disturbance transmission through microbial interactions. Front. Microbiol. 6:1182. doi: 10.3389/fmicb.2015.01182
Husmann, G., Gerdts, G., and Wichels, A. (2010). Spirochetes in Crystalline Styles of Marine Bivalves: Group-Specific Pcr Detection and 16s Rrna Sequence Analysis. J. Shellfish Res. 29, 1069–1075. doi: 10.2983/035.029.0409
Jackson, C. R., and Weeks, A. Q. (2008). Influence of particle size on bacterial community structure in aquatic sediments as revealed by 16S rRNA gene sequence analysis. Appl. Environ. Microbiol. 74, 5237–5240. doi: 10.1128/AEM.00923-08
Kato, H., Mori, H., Maruyama, F., Toyoda, A., Oshima, K., Endo, R., and et al. (2015). Time-series metagenomic analysis reveals robustness of soil microbiome against chemical disturbance. DNA Res. 22, 413–424. doi: 10.1093/dnares/dsv023
King, G. M., Judd, C., Kuske, C. R., and Smith, C. (2012). Analysis of stomach and gut microbiomes of the eastern oyster (Crassostrea virginica) from coastal Louisiana, USA. PLoS ONE 7:e51475. doi: 10.1371/journal.pone.0051475
Koeppel, A. F., and Wu, M. (2012). Lineage-dependent ecological coherence in bacteria. FEMS Microbiol. Ecol. 81, 574–582. doi: 10.1111/j.1574-6941.2012.01387.x
Koeppel, A. F., and Wu, M. (2013). Surprisingly extensive mixed phylogenetic and ecological signals among bacterial Operational Taxonomic Units. Nucleic Acids Res. 41, 5175–5188. doi: 10.1093/nar/gkt241
Kozich, J. J., Westcott, S. L., Baxter, N. T., Highlander, S. K., and Schloss, P. D. (2013). Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 79, 5112–5120. doi: 10.1128/AEM.01043-13
Kuznetsova, A., Brockhoff, P., and Christensen, R. (2014). lmerTest: Tests for Random and Fixed Effects for Linear Mixed Effect Models (lmer Objects of lme4 Package). R package version 2.0–11.
Lear, G., Bellamy, J., Case, B. S., Lee, J. E., and Buckley, H. L. (2014). Fine-scale spatial patterns in bacterial community composition and function within freshwater ponds. Isme J. 8, 1715–1726. doi: 10.1038/ismej.2014.21
Legg, T. M., Zheng, Y., Simone, B., Radloff, K. A., Mladenov, N., González, A., and et al. (2012). Carbon, metals, and grain size correlate with bacterial community structure in sediments of a high arsenic aquifer. Front. Microbiol. 3:82. doi: 10.3389/fmicb.2012.00082
Lemire, A., Goudenege, D., Versigny, T., Petton, B., Calteau, A., Labreuche, Y., and et al. (2015). Populations, not clones, are the unit of vibrio pathogenesis in naturally infected oysters. ISME J. 9, 1523–1531. doi: 10.1038/ismej.2014.233
Ley, R. E., Lozupone, C. A., Hamady, M., Knight, R., and Gordon, J. I. (2008). Worlds within worlds: evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 6, 776–788. doi: 10.1038/nrmicro1978
Linnenbrink, M., Wang, J., Hardouin, E. A., Künzel, S., Metzler, D., and Baines, J. F. (2013). The role of biogeography in shaping diversity of the intestinal microbiota in house mice. Mol. Ecol. 22, 1904–1916. doi: 10.1111/mec.12206
Lokmer, A., Kuenzel, S., Baines, J. F., and Wegner, K. M. (2016). The role of tissue-specific microbiota in initial establishment success of Pacific oysters. Environ. Microbiol. 18, 970–987. doi: 10.1111/1462-2920.13163
Lokmer, A., and Wegner, K. M. (2015). Hemolymph microbiome of Pacific oysters in response to temperature, temperature stress and infection. ISME J. 9, 670–682. doi: 10.1038/ismej.2014.160
Marino, S., Baxter, N. T., Huffnagle, G. B., Petrosino, J. F., and Schloss, P. D. (2014). Mathematical modeling of primary succession of murine intestinal microbiota. Proc. Natl. Acad. Sci. U.S.A. 111, 439–444. doi: 10.1073/pnas.1311322111
Martiny, J. B. H., Eisen, J. A., Penn, K., Allison, S. D., and Horner-Devine, M. C. (2011). Drivers of bacterial beta-diversity depend on spatial scale. Proc. Natl. Acad. Sci. U.S.A. 108, 7850–7854. doi: 10.1073/pnas.1016308108
McFall-Ngai, M., Hadfield, M. G., Bosch, T. C., Carey, H. V., Domazet-Lošo, T., Douglas, A. E., and et al. (2013). Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. U.S.A. 110, 3229–3236. doi: 10.1073/pnas.1218525110
Mihaljevic, J. R. (2012). Linking metacommunity theory and symbiont evolutionary ecology. Trends Ecol. Evol. 27, 323–329. doi: 10.1016/j.tree.2012.01.011
Moehler, J., Wegner, K. M., Reise, K., and Jacobsen, S. (2011). Invasion genetics of Pacific oyster Crassostrea gigas shaped by aquaculture stocking practices. J. Sea Res. 66, 256–262. doi: 10.1016/j.seares.2011.08.004
Moro, C. V., Thioulouse, J., Chauve, C., and Zenner, L. (2011). Diversity, geographic distribution, and habitat-specific variations of microbiota in natural populations of the chicken mite, Dermanyssus gallinae. J. Med. Entomol. 48, 788–796. doi: 10.1603/ME10113
Morrow, K. M., Moss, A. G., Chadwick, N. E., and Liles, M. R. (2012). Bacterial associates of two Caribbean coral species reveal species-specific distribution and geographic variability. Appl. Environ. Microbiol. 78, 6438–6449. doi: 10.1128/AEM.01162-12
Mutshinda, C. M., O'Hara, R. B., and Woiwod, I. P. (2009). What drives community dynamics? Proc. Biol. Sci. 276, 2923–2929. doi: 10.1098/rspb.2009.0523
Nakaoka, M., Ito, N., Yamamoto, T., Okuda, T., and Noda, T. (2006). Similarity of rocky intertidal assemblages along the Pacific coast of Japan: effects of spatial scales and geographic distance. Ecol. Res. 21, 425–435. doi: 10.1007/s11284-005-0138-6
Nemergut, D. R., Schmidt, S. K., Fukami, T., O'Neill, S. P., Bilinski, T. M., Stanish, L. F., and et al. (2013). Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 77, 342–356. doi: 10.1128/MMBR.00051-12
Nguyen, T. T., and Landfald, B. (2015). Polar front associated variation in prokaryotic community structure in Arctic shelf seafloor. Front. Microbiol. 6:17. doi: 10.3389/fmicb.2015.00017
O'Brien, S. L., Gibbons, S. M., Owens, S. M., Hampton-Marcell, J., Johnston, E. R., Jastrow, J. D., and et al. (2016). Spatial scale drives patterns in soil bacterial diversity. Environ. Microbiol. 18, 2039–2051. doi: 10.1111/1462-2920.13231
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., and et al. (2013). vegan: Community Ecology Package. R package version 2.0-7 ed.
Paine, R. T., Tegner, M. J., and Johnson, E. A. (1998). Compounded perturbations yield ecological surprises. Ecosystems 1, 535–545. doi: 10.1007/s100219900049
Petton, B., Boudry, P., Alunno-Bruscia, M., and Pernet, F. (2015a). Factors influencing disease-induced mortality of Pacific oysters Crassostrea gigas. Aquaculture. Environ. Interact. 6, 205–222. doi: 10.3354/aei00125
Petton, B., Bruto, M., James, A., Labreuche, Y., Alunno-Bruscia, M., and Le Roux, F. (2015b). Crassostrea gigas mortality in France: the usual suspect, a herpes virus, may not be the killer in this polymicrobial opportunistic disease. Front. Microbiol. 6:686. doi: 10.3389/fmicb.2015.00686
Peura, S., Bertilsson, S., Jones, R. I., and Eiler, A. (2015). Resistant microbial co-occurrence patterns inferred by network topology. Appl. Environ. Microbiol. 81, 2090–2097. doi: 10.1128/AEM.03660-14
Phillips, C. D., Phelan, G., Dowd, S. E., McDonough, M. M., Ferguson, A. W., Delton Hanson, J., and et al. (2012). Microbiome analysis among bats describes influences of host phylogeny, life history, physiology and geography. Mol. Ecol. 21, 2617–2627. doi: 10.1111/j.1365-294X.2012.05568.x
Pierce, M. L., Ward, J. E., Holohan, B. A., Zhao, X. W., and Hicks, R. E. (2016). The influence of site and season on the gut and pallial fluid microbial communities of the eastern oyster, Crassostrea virginica (Bivalvia, Ostreidae): community-level physiological profiling and genetic structure. Hydrobiologia 765, 97–113. doi: 10.1007/s10750-015-2405-z
Pita, L., Erwin, P. M., Turon, X., and Lopez-Legentil, S. (2013). Till death do us part: stable sponge-bacteria associations under thermal and food shortage stresses. PLoS ONE 8:e80307. doi: 10.1371/journal.pone.0080307
Pratte, Z. A., Richardson, L. L., and Mills, D. K. (2015). Microbiota shifts in the surface mucopolysaccharide layer of corals transferred from natural to aquaria settings. J. Invertebr. Pathol. 125, 42–44. doi: 10.1016/j.jip.2014.12.009
Preheim, S. P., Boucher, Y., Wildschutte, H., David, L. A., Veneziano, D., Alm, E. J., and et al. (2011). Metapopulation structure of Vibrionaceae among coastal marine invertebrates. Environ. Microbiol. 13, 265–275. doi: 10.1111/j.1462-2920.2010.02328.x
Prieur, D., Nicolas, J. L., Plusquellec, A., and Vigneulle, M. (1990). Interactions between Bivalve Mollusks and Bacteria in the Marine-Environment. Oceanogr. Mar. Biol. 28, 277–352.
Pruzzo, C., Gallo, G., and Canesi, L. (2005). Persistence of vibrios in marine bivalves: the role of interactions with haemolymph components. Environ. Microbiol. 7, 761–772. doi: 10.1111/j.1462-2920.2005.00792.x
Ransome, E., Rowley, S. J., Thomas, S., Tait, K., and Munn, C. B. (2014). Disturbance to conserved bacterial communities in the cold-water gorgonian coral Eunicella verrucosa. FEMS Microbiol. Ecol. 90, 404–416. doi: 10.1111/1574-6941.12398
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing).
Reveillaud, J., Maignien, L., Murat Eren, A., Huber, J. A., Apprill, A., Sogin, M. L., and et al. (2014). Host-specificity among abundant and rare taxa in the sponge microbiome. ISME J. 8, 1198–1209. doi: 10.1038/ismej.2013.227
Riisgard, H. U. (1988). Efficiency of particle retention and filtration-rate in 6 Species of Northeast American Bivalves. Mar. Ecol. Prog. Ser. 45, 217–223. doi: 10.3354/meps045217
Ristova, P. P., Wenzhöfer, F., Ramette, A., Felden, J., and Boetius, A. (2015). Spatial scales of bacterial community diversity at cold seeps (Eastern Mediterranean Sea). ISME J. 9, 1306–1318. doi: 10.1038/ismej.2014.217
Robinson, C. J., Schloss, P., Ramos, Y., Raffa, K., and Handelsman, J. (2010). Robustness of the bacterial community in the cabbage white butterfly larval midgut. Microb. Ecol. 59, 199–211. doi: 10.1007/s00248-009-9595-8
Rosa, R. D., de Lorgeril, J., Tailliez, P., Bruno, R., Piquemal, D., and Bachère, E. (2012). A hemocyte gene expression signature correlated with predictive capacity of oysters to survive Vibrio infections. BMC Genomics 13:252. doi: 10.1186/1471-2164-13-252
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., and et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Schmidt, T. S., Matias Rodrigues, J. F., and von Mering, C. (2014). Ecological consistency of SSU rRNA-based operational taxonomic units at a global scale. PLoS Comput. Biol. 10:e1003594. doi: 10.1371/journal.pcbi.1003594
Shade, A., Caporaso, J. G., Handelsman, J., Knight, R., and Fierer, N. (2013). A meta-analysis of changes in bacterial and archaeal communities with time. ISME J. 7, 1493–1506. doi: 10.1038/ismej.2013.54
Shade, A., Jones, S. E., Caporaso, J. G., Handelsman, J., Knight, R., Fierer, N., and et al. (2014). Conditionally rare taxa disproportionately contribute to temporal changes in microbial diversity. MBio 5, e01371–e01314. doi: 10.1128/mBio.01371-14
Shade, A., Peter, H., Allison, S. D., Baho, D. L., Berga, M., Bürgmann, H., and et al. (2012). Fundamentals of microbial community resistance and resilience. Front. Microbiol. 3:417. doi: 10.3389/fmicb.2012.00417
Shade, A., Read, J. S., Welkie, D. G., Kratz, T. K., Wu, C. H., and McMahon, K. D. (2011). Resistance, resilience and recovery: aquatic bacterial dynamics after water column disturbance. Environ. Microbiol. 13, 2752–2767. doi: 10.1111/j.1462-2920.2011.02546.x
Shea, K., Roxburgh, S. H., and Rauschert, E. S. J. (2004). Moving from pattern to process: coexistence mechanisms under intermediate disturbance regimes. Ecol. Lett. 7, 491–508. doi: 10.1111/j.1461-0248.2004.00600.x
Sousa, W. P. (1984). The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 15, 353–391. doi: 10.1146/annurev.es.15.110184.002033
Stein, R. R., Bucci, V., Toussaint, N. C., Buffie, C. G., Rätsch, G., Pamer, E. G., and et al. (2013). Ecological modeling from time-series inference: insight into dynamics and stability of intestinal microbiota. PLoS Comput. Biol. 9:e1003388. doi: 10.1371/journal.pcbi.1003388
Stenuit, B., and Agathos, S. N. (2015). Deciphering microbial community robustness through synthetic ecology and molecular systems synecology. Curr. Opin. Biotechnol. 33, 305–317. doi: 10.1016/j.copbio.2015.03.012
Sudakaran, S., Salem, H., Kost, C., and Kaltenpoth, M. (2012). Geographical and ecological stability of the symbiotic mid-gut microbiota in European firebugs, Pyrrhocoris apterus (Hemiptera, Pyrrhocoridae). Mol. Ecol. 21, 6134–6151. doi: 10.1111/mec.12027
Tanaka, R., Ootsubo, M., Sawabe, T., Ezura, Y., and Tajima, K. (2004). Biodiversity and in situ abundance of gut microflora of abalone (Haliotis discus hannai) determined by culture-independent techniques. Aquaculture 241, 453–463. doi: 10.1016/j.aquaculture.2004.08.032
Trabal, N., Mazón-Suástegui, J. M., Vázquez-Juárez, R., Asencio-Valle, F., Morales-Bojórquez, E., and Romero, J. (2012). Molecular analysis of bacterial microbiota associated with oysters (Crassostrea gigas and Crassostrea corteziensis) in different growth phases at two cultivation sites. Microb. Ecol. 64, 555–569. doi: 10.1007/s00248-012-0039-5
Trabal Fernández, N., Mazón-Suástegui, J. M., Vázquez-Juárez, R., Ascencio-Valle, F., and Romero, J. (2014). Changes in the composition and diversity of the bacterial microbiota associated with oysters (Crassostrea corteziensis, Crassostrea gigas and Crassostrea sikamea) during commercial production. FEMS Microbiol. Ecol. 88, 69–83. doi: 10.1111/1574-6941.12270
Tracy, A. M., Koren, O., Douglas, N., Weil, E., and Harvell, C. D. (2015). Persistent shifts in Caribbean coral microbiota are linked to the 2010 warm thermal anomaly. Environ. Microbiol. Rep. 7, 471–479. doi: 10.1111/1758-2229.12274
van Aken, H. (2008). Variability of the water temperature in the western Wadden Sea on tidal to centennial time scales. J. Sea Res. 60, 227–334. doi: 10.1016/j.seares.2008.09.001
Vasconcelos, G. J., and Lee, J. S. (1972). Microbial flora of Pacific oysters (Crassostrea gigas) subjected to ultraviolet-irradiated seawater. Appl. Microbiol. 23, 11–16.
Vega Thurber, R., Willner-Hall, D., Rodriguez-Mueller, B., Desnues, C., Edwards, R. A., Angly, F., and et al. (2009). Metagenomic analysis of stressed coral holobionts. Environ. Microbiol. 11, 2148–2163. doi: 10.1111/j.1462-2920.2009.01935.x
Wang, J., Kalyan, S., Steck, N., Turner, L. M., Harr, B., Künzel, S., and et al. (2015). Analysis of intestinal microbiota in hybrid house mice reveals evolutionary divergence in a vertebrate hologenome. Nat. Commun. 6, 6440. doi: 10.1038/ncomms7440
Wang, Y., Naumann, U., Wright, S. T., and Warton, D. I. (2012). mvabund- an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 3, 471–474. doi: 10.1111/j.2041-210X.2012.00190.x
Wegner, K. M., Volkenborn, N., Peter, H., and Eiler, A. (2013). Disturbance induced decoupling between host genetics and composition of the associated microbiome. BMC Microbiol. 13:252. doi: 10.1186/1471-2180-13-252
Wendling, C. C., Batista, F. M., and Wegner, K. M. (2014). Persistence, seasonal dynamics and pathogenic potential of Vibrio communities from Pacific oyster hemolymph. PLoS ONE 9:e94256. doi: 10.1371/journal.pone.0094256
Wendling, C. C., and Wegner, K. M. (2015). Adaptation to enemy shifts: rapid resistance evolution to local Vibrio spp. in invasive Pacific oysters. Proc. Biol. Sci. 282:20142244. doi: 10.1098/rspb.2014.2244
Wong, A. C., Chaston, J. M., and Douglas, A. E. (2013). The inconstant gut microbiota of Drosophila species revealed by 16S rRNA gene analysis. ISME J. 7, 1922–1932. doi: 10.1038/ismej.2013.86
Yatsunenko, T., Rey, F. E., Manary, M. J., Trehan, I., Dominguez-Bello, M. G., Contreras, M., and et al. (2012). Human gut microbiome viewed across age and geography. Nature 486, 222–227. doi: 10.1038/nature11053
Ziegler, M., Roik, A., Porter, A., Zubier, K., Mudarris, M. S., Ormond, R., and et al. (2016). Coral microbial community dynamics in response to anthropogenic impacts near a major city in the central Red Sea. Mar. Pollut. Bull. 105, 629–640. doi: 10.1016/j.marpolbul.2015.12.045
Zinger, L., Boetius, A., and Ramette, A. (2014). Bacterial taxa-area and distance-decay relationships in marine environments. Mol. Ecol. 23, 954–964. doi: 10.1111/mec.12640
Zouache, K., Raharimalala, F. N., Raquin, V., Tran-Van, V., Raveloson, L. H., Ravelonandro, P., and et al. (2011). Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti, from different geographic regions of Madagascar. FEMS Microbiol. Ecol. 75, 377–389. doi: 10.1111/j.1574-6941.2010.01012.x
Keywords: host-associated communities, Crassostrea gigas, distance-decay relationship, spatiotemporal patterns, spatiotemporal dynamics, marine invertebrate microbiota, amplicon analysis
Citation: Lokmer A, Goedknegt MA, Thieltges DW, Fiorentino D, Kuenzel S, Baines JF and Wegner KM (2016) Spatial and Temporal Dynamics of Pacific Oyster Hemolymph Microbiota across Multiple Scales. Front. Microbiol. 7:1367. doi: 10.3389/fmicb.2016.01367
Received: 27 April 2016; Accepted: 18 August 2016;
Published: 31 August 2016.
Edited by:
Christine Moissl-Eichinger, Medical University of Graz, AustriaReviewed by:
Colleen A. Burge, University of Maryland, Baltimore Country, USAElisabeth Margaretha Bik, Stanford University, USA
Copyright © 2016 Lokmer, Goedknegt, Thieltges, Fiorentino, Kuenzel, Baines and Wegner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Lokmer, alokmer@awi.de