 Beatrix Tettmann1
Beatrix Tettmann1 Christine Niewerth2
Christine Niewerth2 Frank Kirschhöfer1Anke Neidig1
Frank Kirschhöfer1Anke Neidig1 Andreas Dötsch1
Andreas Dötsch1 Gerald Brenner-Weiss1
Gerald Brenner-Weiss1 Susanne Fetzner2†
Susanne Fetzner2† Joerg Overhage1*†
Joerg Overhage1*†- 1Karlsruhe Institute of Technology, Institute of Functional Interfaces, Karlsruhe, Germany
- 2Institute for Molecular Microbiology and Biotechnology, University of Münster, Münster, Germany
The 2-alkyl-3-hydroxy-4(1H)-quinolone 2,4-dioxygenase HodC was previously described to cleave the Pseudomonas quinolone signal, PQS, which is exclusively used in the complex quorum sensing (QS) system of Pseudomonas aeruginosa, an opportunistic pathogen employing QS to regulate virulence and biofilm development. Degradation of PQS by exogenous addition of HodC to planktonic cells of P. aeruginosa attenuated production of virulence factors, and reduced virulence in planta. However, proteolytic cleavage reduced the efficacy of HodC. Here, we identified the secreted protease LasB of P. aeruginosa to be responsible for HodC degradation. In static biofilms of the P. aeruginosa PA14 lasB::Tn mutant, the catalytic activity of HodC led to an increase in viable biomass in newly formed but also in established biofilms, and reduced the expression of genes involved in iron metabolism and siderophore production, such as pvdS, pvdL, pvdA, and pvdQ. This is likely due to an increase in the levels of bioavailable iron by degradation of PQS, which is able to sequester iron from the surrounding environment. Thus, HodC, despite its ability to quench the production of virulence factors, is contraindicated for combating P. aeruginosa biofilms.
Introduction
Pseudomonas aeruginosa is one of the most important opportunistic human pathogens, causing a variety of life-threatening infections in immunocompromised patients (Gellatly and Hancock, 2013). Moreover, P. aeruginosa is the dominant and most significant pathogen in patients suffering from cystic fibrosis, causing very difficult to treat pulmonary infections (Hutchison and Govan, 1999; Rajan and Saiman, 2002). It has a number of intrinsic resistance mechanisms including multidrug resistance efflux systems, low outer membrane permeability, and β-lactamases, produces a large arsenal of virulence factors, and moreover forms robust biofilms (Gellatly and Hancock, 2013).
Like many other pathogenic bacteria, P. aeruginosa co-ordinates group behavior, such as the synthesis of many virulence factors and biofilm development, via cell-to-cell communication or quorum sensing (QS) (Williams et al., 2007). The sophisticated QS network of P. aeruginosa comprises several interconnected signaling circuits, with the Las and Rhl systems producing and responding to N-3-oxo-dodecanoyl homoserine lactone and N-butanoyl homoserine lactone, respectively, and the PQS circuit using specific 2-n-alkyl-4(1H)-quinolone (AQ) signals. 2-Heptyl-3-hydroxy-4(1H)-quinolone, the “Pseudomonas quinolone signal” (PQS), is the major AQ signal in P. aeruginosa (Pesci et al., 1999). Both PQS and its biosynthetic precursor HHQ (2-heptyl-4(1H)-quinolone) act as coinducers of the transcriptional regulator PqsR (MvfR) (Déziel et al., 2005; Wade et al., 2005; Xiao et al., 2006; Diggle et al., 2007). PQS, besides its role as a QS signal molecule, modulates membrane properties (Mashburn and Whiteley, 2005), acts as ferric iron chelator (Bredenbruch et al., 2006; Diggle et al., 2007) and pro-oxidant (Häussler and Becker, 2008), and exerts pro-apoptotic and host immune modulatory activities (Hooi et al., 2004; Skindersoe et al., 2009; Hänsch et al., 2014).
PQS has been reported to promote biofilm development (Diggle et al., 2003), and pqsA and pqsC mutants, which are unable to produce AQs, are poor biofilm producers (Müsken et al., 2010). Because PQS did not induce biofilm formation in mutants deficient of the signal transduction histidine kinase RetS or the sensor/response regulator protein GacS, it has been suggested that enhancement of biofilm by PQS is at least partially dependent on the RetS-GacAS-Rsm system (Guo et al., 2014). GacA positively controls the small regulatory RNA RsmZ, which acts by sequestering the RsmA protein (Sonnleitner and Haas, 2011). RsmA directly and via modulating cyclic di-GMP levels controls diverse functions related to the P. aeruginosa switch between planktonic and biofilm lifestyles (Frangipani et al., 2014).
Because PQS signaling is involved in control of virulence factor production as well as biofilm maturation, interference with this QS system has been discussed as an attractive anti-virulence strategy (Lesic et al., 2007; Pustelny et al., 2009; Storz et al., 2012; Lu et al., 2014). Possible targets to interfere with QS circuits are the enzymes involved in signal biosynthesis, the signal receptor, or the signal itself. With respect to the latter, it seems that enzyme-catalyzed modification or degradation of bacterial signal molecules is wide-spread in nature. However, the majority of the quorum quenching enzymes identified to date are lactonases or acylases hydrolyzing N-acylhomoserine lactones, while only few enzymes have been described that are active toward other signal molecules (Fetzner, 2015). Among these, the heterocyclic-ring-cleaving enzyme “Hod” (1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase) from Arthrobacter sp. Rue61a, despite its preference for 2-alkyl-3-hydroxy-4(1H)-quinolones with short alkyl substituents, is capable of cleaving PQS to form carbon monoxide and N-octanoylanthranilic acid. Exogenous addition of the enzyme to P. aeruginosa planktonic cultures resulted in significant down-regulation of the expression of key virulence factors and in reduction of P. aeruginosa pathogenesis in a plant infection model, highlighting the potential of quenching virulence through the enzymatic degradation of PQS (Pustelny et al., 2009). However, Pustelny et al. (2009) also observed that cleavage of Hod by extracellular proteases of P. aeruginosa reduced its efficiency as a quorum-quenching agent. P. aeruginosa secretes multiple proteases which degrade both soluble and structural host proteins and thus contribute to its pathogenicity. Among them, especially the metalloproteinases LasA, LasB, AprA and the serine protease PrpL (protease IV) have been correlated with virulence (Caballero et al., 2001).
In this study, we investigated the effect of HodC on biofilms of P. aeruginosa PA14 strains deficient in the production of the protease LasB, which was found to be liable for proteolytic degradation of the enzyme. The presence of exogenous, catalytically active HodC led to increased biomass in newly formed but also established biofilms, and down-regulated a set of genes which are under control of the ferric uptake regulator (Fur). Degradation of the iron chelator PQS is likely accompanied by an increase in readily bioavailable iron which is responsible for a gain in biofilm biomass.
Materials and Methods
Bacterial Strains, Media, and Culture Conditions
The bacterial strains used in this study are listed in Table 1. All mutant strains were confirmed by PCR (data not shown). Growth was routinely performed in lysogeny broth (LB) or BM2 minimal medium (Overhage et al., 2008) at 37°C with shaking at 170 rpm unless otherwise indicated. When required, gentamicin was used at a final concentration of 30 μg/ml for P. aeruginosa transposon mutants and ampicillin and kanamycin at final concentrations of 100 and 30 μg/ml, respectively, for recombinant Escherichia coli.

Table 1. Bacterial strains used in this study.
For recording growth curves, bacterial cultures were grown overnight in LB and diluted in fresh LB to obtain starting optical densities of OD600nm = 0.1. Aliquots of these dilutions were apportioned in 96-well microtiter plates (100 μl per well). Growth was recorded using a TECAN Infinite® 200 PRO plate reader (Tecan, Maennedorf, Switzerland) under shaking conditions. Two independent experiments were performed with three replicates for each strain or condition.
HodC Purification and Activity Assay
Since wild-type Hod protein is prone to oxidative dimerization by the formation of an intermolecular disulfide bridge, Hod protein carrying a substitution of Cys69 by serine, termed HodC, was used. HodC shows the same catalytic activity as Hod (Frerichs-Deeken et al., 2004). HodC-H251A protein, termed iHodC, which is virtually inactive due to its inability to initiate catalysis by deprotonating the organic substrate (Frerichs-Deeken et al., 2004), was used in control experiments. Purification of the recombinant His6-tagged HodC proteins from recombinant E. coli strains (Table 1) was performed as described by Beermann et al. (2007). For storage at −80°C, 10% glycerol (vol/vol) was added to the protein stock solutions. To control the activity of HodC protein before and after an experiment, its catalytic activity was determined spectrophotometrically by measuring 3-hydroxy-2-methyl-4(1H)-quinolone consumption as described previously (Frerichs-Deeken et al., 2004). 1 Unit of enzyme activity is defined as the amount of HodC catalyzing the conversion of 1 μmol substrate per minute under the conditions of the assay. Concentrations of HodC were determined by absorption measurements using an extinction coefficient (ε280nm) of 1.937 ml mg−1 cm−1 (Beermann et al., 2007).
Enzyme Stability Assay
Stability of HodC against different proteases produced by P. aeruginosa was monitored by measuring the enzyme activity in the presence of stationary phase culture supernatants of P. aeruginosa PA14, and of the PA14 mutants lasB::Tn, lasA::Tn, prpL::Tn, and aprA::Tn, respectively. Bacterial strains were grown in LB medium overnight at 37°C and 160 rpm. These cultures were used to inoculate 15 ml LB medium to an OD600 nm of 0.05, and cells were further grown for 10 h at 37°C and vigorous shaking. Afterwards, cells were pelleted by centrifugation (9000 × g, 10 min, 4°C) and the culture supernatants were collected and filter sterilized. For analysis, 100 μl/ml culture supernatant was incubated in sodium phosphate buffer pH 8.0 with 0.75 μg/ml HodC at 37°C, and enzyme activity was measured at different time points.
Chemicals
The PQS cleavage product, N-octanoylanthranilic acid, was synthesized according to the method described by Wells et al. (1952) with some modifications. Briefly, octanoyl chloride (Sigma Aldrich, Taufkirchen, Germany) was added dropwise to a solution of methyl anthranilate (Sigma Aldrich, Taufkirchen, Germany) dissolved in ethyl acetate at 0°C while stirring. Following heating to 50°C for 5 min the solution was stirred over night at room temperature resulting in a clear solution. Subsequently, this solution was successively washed with water, 1 M sodium hydroxide, 1 M hydrochloric acid and finally with brine. After drying with Na2SO4 the organic solvent was removed by evaporation leading to a crude oil of methyl N-octanoylanthranilate which was used without further purification. Hydrolysis of the acylated anthranilate was carried out with 0.5 M sodium hydroxide solved in ethanol. The reaction mixture was refluxed for 3 h. After cooling, the solution was acidified with 4 M hydrochloric acid and extracted two times with n-hexane. Evaporation yielded crude N-octanoylanthranilic acid which was recrystallized in n-hexane. ESI-TOF/MS for C15H22NO3+ ([M + H]+): calculated m/z = 264.159; found m/z = 264.114.
CORM-2, a carbon monoxide releasing molecule, and PQS were purchased from Sigma Aldrich (Taufkirchen, Germany).
Attachment, Biofilm Formation, and Extracellular DNA
Rapid attachment of bacterial cells to a surface was analyzed as described previously (Yeung et al., 2011). Briefly, overnight cultures grown in LB-medium were washed and diluted in LB medium to an OD600 nm of 1.0. Aliquots (100 μl) of this suspension were used to inoculate each well of a microtiter plate. Cells were allowed to adhere for 60 min at 37°C prior to staining with crystal violet. All experiments were done in triplicates with 6 individual repeats per measurement (n = 18).
The abiotic solid surface assay was used to measure biofilm formation according to the method described by Friedman and Kolter (2004) with the following modifications. Overnight cultures were diluted 1:100 in fresh LB medium, inoculated into 96-well microtiter plates and incubated for 24 h at 37°C without shaking to allow bacterial adherence and biofilm formation. After incubation the biofilm cells were stained using 0.1% (w/v) crystal violet and the absorbance was measured at 595 nm using a TECAN Infinite® 200 PRO microtiter plate reader.
For the determination of colony forming units (CFU), biofilms were grown in glass bottom petri dishes as described above in the presence or absence of 70 U/ml HodC. After 24 h of incubation, planktonic cells were removed by gentle washing. The remaining, adherent cells were scraped off the petri dish surface using cell scrapers, transferred into a new test tube and vortexed vigorously for 10–30 s. Several dilutions were prepared and the CFU were determined by the drop plate method (Herigstad et al., 2001).
Extracellular DNA in biofilm cultures was determined according to the method described previously (Yang et al., 2007). Briefly, overnight cultures were diluted 1:100 in fresh LB medium supplemented with 0.05 mM propidium iodide and biofilms were grown in microtiter plates at 37°C under static conditions. After 24 h of incubation, the absorbance of propidium iodide was measured at 490 nm using a MultisKan MS photometer (Labsystems, Bradenton, USA) and cell density at 595 nm using a TECAN Infinite® 200 PRO.
Fluorescence Microscopy
For microscopic analyses, overnight cultures of P. aeruginosa PA14 lasB::Tn were diluted 1:100 in LB, HodC was added at a final concentration of 70 U/ml, and the suspensions were used to inoculate glass bottom petri dishes (MatTek, Ashland, USA). After 24 h of incubation at 37°C under static conditions, planktonic cells were removed from the medium by gently washing with LB medium and the attached viable biofilm cells were stained using 5-cyano-2,3-ditolyl tetrazolium chloride (CTC) for 3 h in the dark as described previously (Li et al., 2013). Fluorescence microscopy was carried out using an Axioplan 2 imaging system (Carl Zeiss, Oberkochem, Germany) with appropriate filter sets.
Transcriptome Analysis
RNA for transcriptome analysis was isolated from biofilm cells of the P. aeruginosa PA14 lasB::Tn strain, grown for 24 h at 37°C under static conditions in microtiter plates in LB medium in the presence or absence of 70 U/ml HodC or iHodC, respectively. The biofilm cells of 40 wells for each condition were pooled, total RNA was extracted using RNeasy Midi columns (QIAGEN), and DNase treatment of isolated RNA samples were performed as described previously (Breidenstein et al., 2012). Depletion of rRNA was accomplished with the MICROBExpress™ bacterial mRNA enrichment kit (life technologies) according to the manufacturer's protocol. RNA-Seq was performed as described previously (Tettmann et al., 2014). Briefly, sequencing libraries were generated from 50 ng of rRNA depleted RNA samples following the Truseq RNA protocol (Illumina). Paired end reads (2 × 50 nucleotides) were obtained with a Hiseq1000 using SBS v3 kits (Illumina). Cluster detection and base calling were performed using RTAv1.13, and quality of reads was assessed with CASAVA v1.8.1 (Illumina). The reads were mapped against the genome of P. aeruginosa PA14 (accession number: NC_008463) using bowtie2 (Langmead and Salzberg, 2012). The genomic annotation of P. aeruginosa PA14 was downloaded from the Pseudomonas Genome database (www.pseudomonas.com) (Winsor et al., 2009). Gene expression was determined by counting for each gene the number of reads that overlapped with the annotation location using HTSeq (Anders et al., 2015). Differential expression was calculated using the R package DESeq2 (Love et al., 2014), and genes were assumed to be differentially expressed, if the fold-change was at least two-fold (±) and the P-value less than 0.05. Complete expression data is deposited at the Sequence Read Archive NCBI under the accession number SRP046054.
Results
HodC is Stable in Cultures of P. aeruginosa PA14 lasB::Tn
The 2-alkyl-3-hydroxy-4(1H)-quinolone 2,4-dioxygenase HodC was previously shown to be able to quench PQS signaling in P. aeruginosa, leading to decreased production of several virulence factors, however, HodC activity in P. aeruginosa cultures was significantly reduced over time by proteolytic cleavage (Pustelny et al., 2009). In order to identify the extracellular protease(s) of P. aeruginosa PA14 involved in inactivation of HodC, its activity was determined in the presence of cell-free culture supernatants of P. aeruginosa PA14 and supernatants of the four protease deficient mutants lasB::Tn (inactivation of elastase LasB), lasA::Tn (LasA), prpL::Tn (protease IV), and aprA::Tn (alkaline metalloproteinase). While supernatants of P. aeruginosa PA14 as well as of the lasA, prpL, and aprA mutant strains strongly decreased HodC activity already after 10 min of incubation, no change in activity was observed for supernatants of P. aeruginosa PA14 lasB::Tn in comparison to the LB control (Figure 1). Therefore, the lasB mutant was chosen for all following experiments to ensure high HodC activity even during prolonged incubation periods.
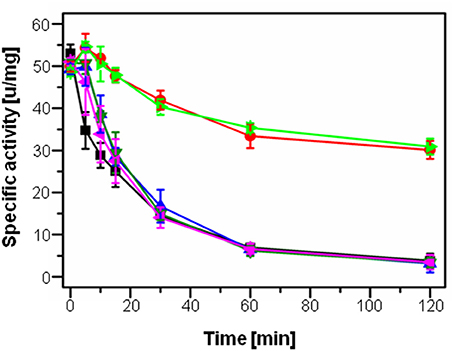
Figure 1. Specific activity of HodC upon incubation with culture supernatants of P. aeruginosa strains. 100 μl/ml sterile LB medium (light green), or 100 μl/ml culture supernatant of P. aeruginosa PA14 (blue), the lasA::Tn mutant (black), the aprA::Tn mutant (dark green), the prpL::Tn mutant (pink), and the lasB::Tn mutant (red) in sodium phosphate buffer (pH 8.0) were incubated with 0.75 μg/ml HodC at 37°C, and enzyme activity was measured at different time points. Error bars indicate standard deviations from three independent experiments.
HodC Activity Stimulates an Increase in Biomass in Newly Formed but Also in Established Biofilms
Since PQS is important for biofilm formation (Diggle et al., 2003), we analyzed the effect of HodC on P. aeruginosa biofilm development. Addition of HodC to static biofilm cultures of P. aeruginosa PA14 resulted in a moderate increase in biofilm biomass after 24 h of incubation, whereas the lasB::Tn mutant exhibited a much stronger, five-fold increase in biofilm biomass in the presence of HodC (Figure 2A). This enhancement in biofilm formation increased with increasing concentrations of the enzyme in the culture media (Figure 2B). In contrast, addition of inactive HodC (iHodC) to biofilm cultures of both P. aeruginosa PA14 wild-type and the lasB mutant did not have an impact on biofilm development (Figure 2A). Remarkably, HodC addition even influenced established biofilms, as we observed an almost two-fold increase of biofilm biomass of pre-grown biofilms in the presence of the enzyme (Figure 3).
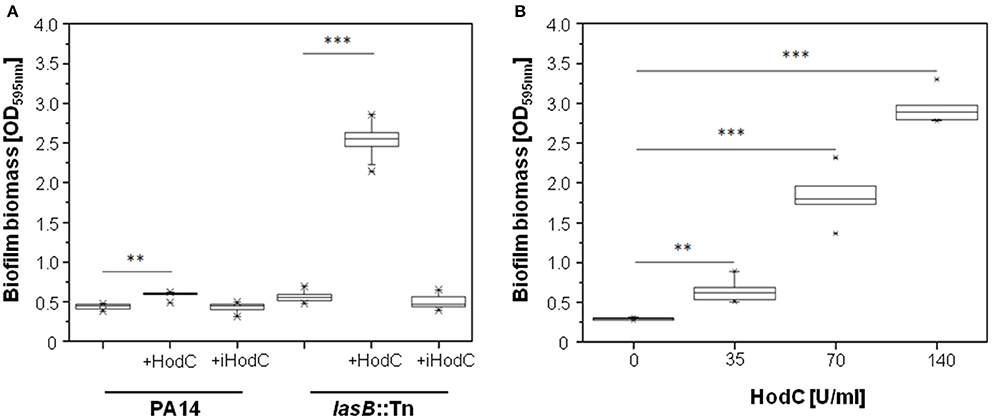
Figure 2. Effect of HodC on biofilm formation of P. aeruginosa. (A) Effect of HodC (70 U/ml) and iHodC (at the same protein concentration) on wild-type P. aeruginosa PA14 and P. aeruginosa PA14 lasB::Tn. (B) Biofilm formation in response to different concentrations of HodC. (A,B) Overnight cultures (in LB) were diluted (1:100) in LB in 96-well microtiter plates, and supplemented with HodC protein as indicated. After incubation for 24 h at 37°C, cells were stained with 0.1% (w/v) crystal violet and quantified by measuring OD595nm. Experiments were done in triplicates with 6 individual repeats per measurement (n = 18). Statistical analyses were performed with the Mann-Whitney U-test. **p ≤ 0.01; ***p ≤ 0.001. The highest and lowest outliers are indicated by “X.”
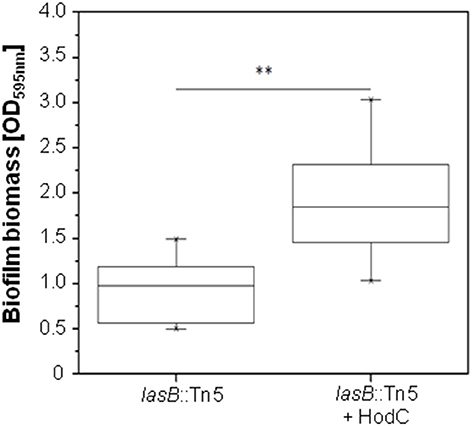
Figure 3. Effect of HodC on pre-grown biofilms of P. aeruginosa PA14 lasB::Tn. Overnight cultures (in LB) were diluted (1:100) in LB in 96-well microtiter plates, and incubated for 24 h at 37°C. After removal of planktonic cells and a washing step with LB, the biofilm was covered with fresh LB without or with HodC (70 U/ml) and incubated for another 24 h at 37°C. Biofilm cells were stained with 0.1% (w/v) crystal violet and quantified by measuring OD595nm. Experiments were done in triplicates with 6 individual repeats per measurement (n = 18). Statistical analyses were performed with the Mann-Whitney U-test. **p ≤ 0.01. The highest and lowest outliers are indicated by “X.”
To analyze whether the observed enhancement in biofilm mass was indeed due to an increase in viable cells, we evaluated biofilms of P. aeruginosa PA14 lasB::Tn on glass bottom petri dishes using fluorescence microscopy in combination with CTC staining, which allows the visualization of metabolically active cells. The biofilms exhibited a much higher proportion of viable cells when grown in the presence of HodC (Figure 4). Moreover, determination of CFUs revealed 9.2 × 108 (± 3 × 108) CFU/ml and 22.5 × 108 (± 2.7 × 108) CFU/ml for untreated and HodC-treated biofilms, respectively, corresponding to a 2.4-fold increase in viable biofilm cells in the presence of HodC. However, HodC (70 U/ml) did not affect planktonic growth of P. aeruginosa PA14 in LB (data not shown). Since initial adherence of bacterial cells strongly impacts biofilm development, we investigated whether HodC affects biofilm growth already at the very initial phase of cell attachment. To this end, attachment of P. aeruginosa PA14 lasB::Tn was determined after 1 h of incubation with or without HodC. Quantification of surface-attached cells by crystal violet staining revealed that HodC addition did not alter initial cell adhesion of P. aeruginosa (Supplementary Figure 1A). In addition, quantification of extracellular DNA, which is an important factor in biofilm formation, revealed no differences in eDNA of biofilms grown in the presence of HodC compared to the iHodC control biofilms (Supplementary Figure 1B). Taken together, the results indicate that the enhanced biofilm in presence of HodC is due to the increase in viable biomass.
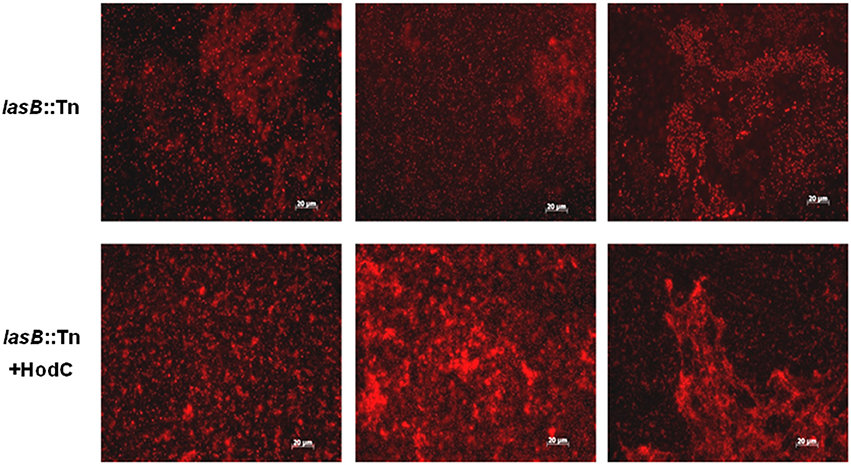
Figure 4. Biofilms of P. aeruginosa PA14 lasB::Tn in presence and absence of HodC, stained with CTC. Overnight cultures were diluted (10−2) in LB, supplemented with HodC at a final concentration of 70 U/ml, and the suspensions were used to inoculate glass bottom petri dishes (MatTek, Ashland, USA). After 24 h of incubation at 37°C under static conditions, planktonic cells were removed by washing with LB medium and the attached viable biofilm cells were stained using 5-cyano-2,3-ditolyl tetrazolium chloride (CTC). Fluorescence microscopy was carried out using an Axioplan 2 imaging system with appropriate filter sets. Experiments were performed in triplicate and representative images are shown.
The PQS Degradation Products Carbon Monoxide and N-octanoylanthranilic Acid Do Not Influence Biofilm Development
In order to identify the mechanisms underlying the stimulation in biofilm formation by HodC, we first analyzed whether N-octanoylanthranilic acid and carbon monoxide, the products of HodC-catalyzed PQS cleavage, have an effect on biofilm development of the P. aeruginosa PA14 lasB::Tn strain. In agreement with an earlier study by Murray et al. (2012), no significant increase in biofilm formation was observed in the presence of 10–20 μM of the CO-releasing molecule CORM-2, but biofilm formation was decreased strongly at higher concentrations of CORM-2 (Figure 5A). Given that supernatants of overnight cultures of P. aeruginosa PA14 contain between 5 and 25 μM PQS (Diggle et al., 2003; Lépine et al., 2003), we analyzed the impact of N-octanoylanthranilic acid on biofilm biomass at concentrations of up to 30 μM. The data (Figure 5B) indicated that this PQS cleavage product has also no influence on biofilm development. Overall, these results indicated that the two metabolites formed by HodC-mediated PQS cleavage are not responsible for the observed biofilm phenotype.
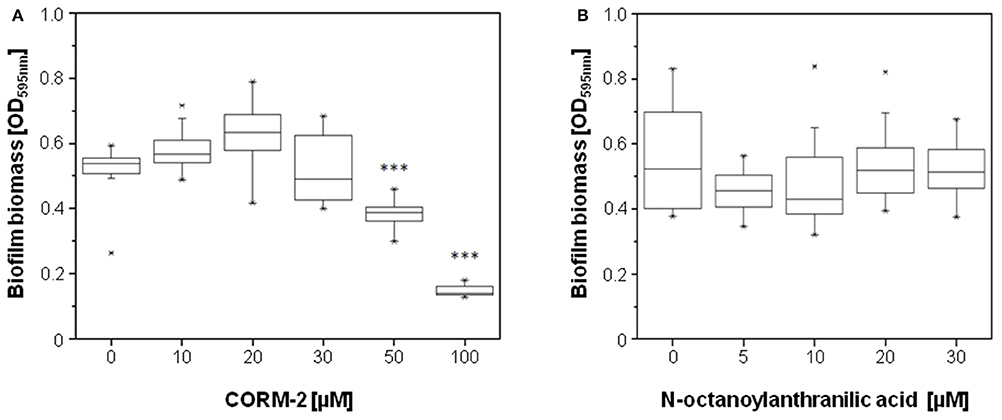
Figure 5. Effect of the products of HodC-catalyzed PQS cleavage on biofilm formation of P. aeruginosa PA14 lasB::Tn. Overnight cultures (in LB) were diluted (10−2) (A) in BM2 medium (without casamino acids), or (B) in LB, and supplemented with the either the CO-releasing molecule CORM-2 (A), or with N-octanoylanthranilic acid (B) as indicated. After incubation for 24 h at 37°C, cells were stained with 0.1% (w/v) crystal violet and quantified by measuring OD595nm. Experiments were done in triplicates with 6 individual repeats per measurement (n = 18). Statistical analyses were performed with the Mann-Whitney U-test. The highest and lowest outliers are indicated by “X.” (A) ***p ≤ 0.001, in relation to biofilms of P. aeruginosa PA14 lasB::Tn as formed in absence of compounds.
HodC Activity in Biofilm Cultures Results in Down-Regulation of Fur-Controlled Genes, Likely Due to Increased Iron Availability
To get an insight into the molecular basis for the observed biofilm phenotype, transcriptome analyses were performed using Illumina sequencing, and the expression profile of P. aeruginosa PA14 lasB::Tn biofilms treated with HodC was compared to that of untreated control biofilm cells. This analysis identified a total set of 38 genes that were differentially regulated in the presence of HodC (Table 2). Most of the 34 genes that were down-regulated are directly or indirectly under control of the ferric uptake regulator Fur (Ochsner et al., 2002). Among them are the operon comprising the fumarate hydratase (fumC) and superoxide dismutase (sodM) genes (Hassett et al., 1997), and genes coding for the regulation and biosynthesis of the siderophore pyoverdine (pvdS, pvdL, pvdA, pvdQ) and for proteins involved in heme uptake and degradation (phuS, phuR, hemO). Additional transcriptome analyses for cells incubated with iHodC did not reveal any significant differences in gene expression except a 3.1-fold upregulation of the glpD gene, which is coding for glycerol-3-phosphate dehydrogenase, in comparison to the untreated control biofilms (data not shown). This glpD upregulation was comparable to the expression observed in response to HodC (Table 2) and is most likely due to the presence of glycerol in the enzyme stock solutions. Thus, the observed transcriptome data demonstrate that the changes in gene expression are entirely depending on the catalytic activity of HodC.
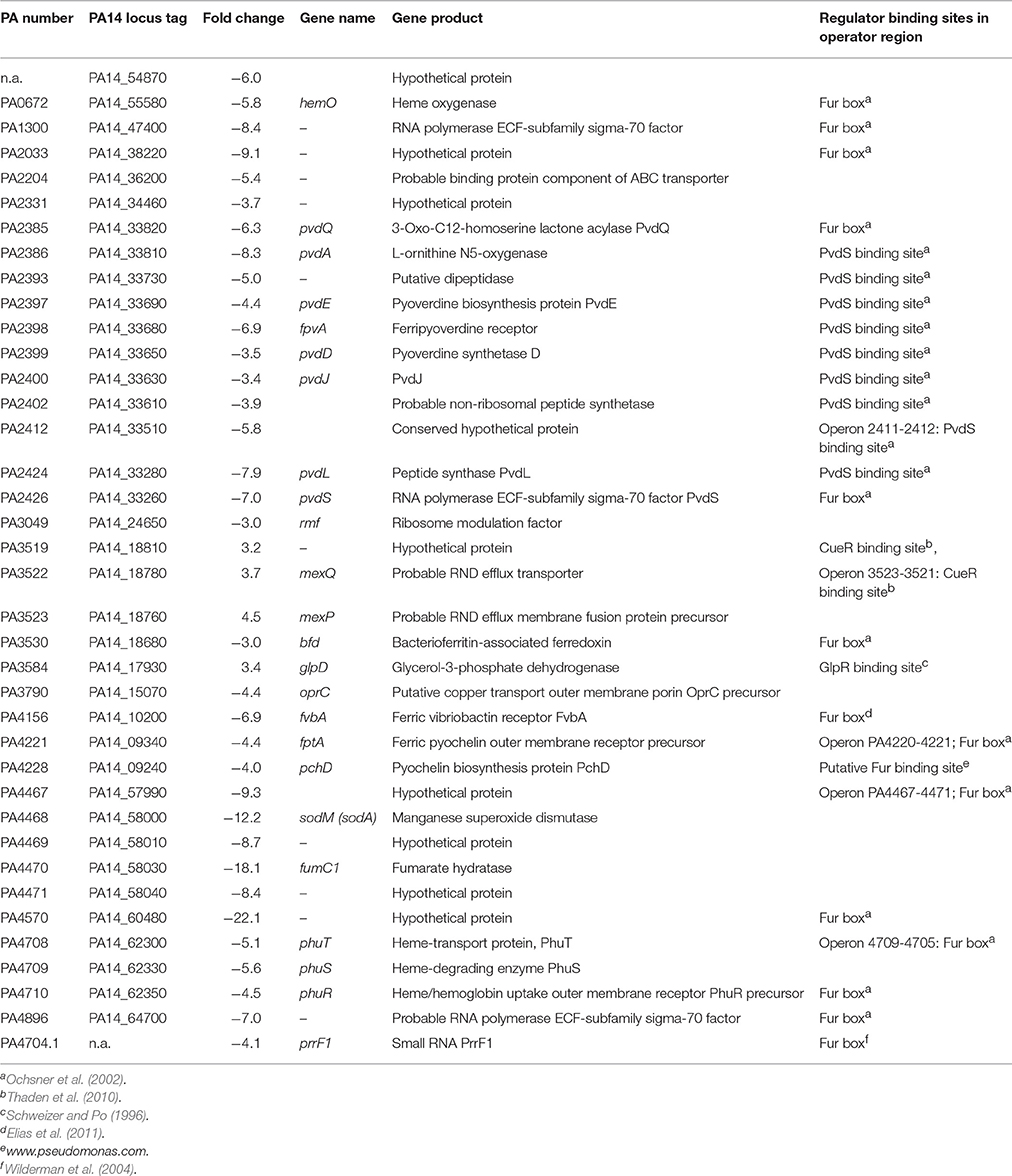
Table 2. Genes of P. aeruginosa PA14 lasB::Tn with decreased expression in presence of HodC.
Because PQS exhibits iron-chelating properties and therefore sequesters iron from the culture medium (Bredenbruch et al., 2006; Diggle et al., 2007), we assumed that the observed downregulation of genes involved in iron metabolism by HodC is due to an increase in available iron in the culture medium as a result of PQS cleavage. This hypothesis is supported by the finding that in chemically defined medium, biofilm formation of the P. aeruginosa lasB mutant was significantly enhanced by the addition of 50 μM FeSO4 or FeCl3 (Figure 6).
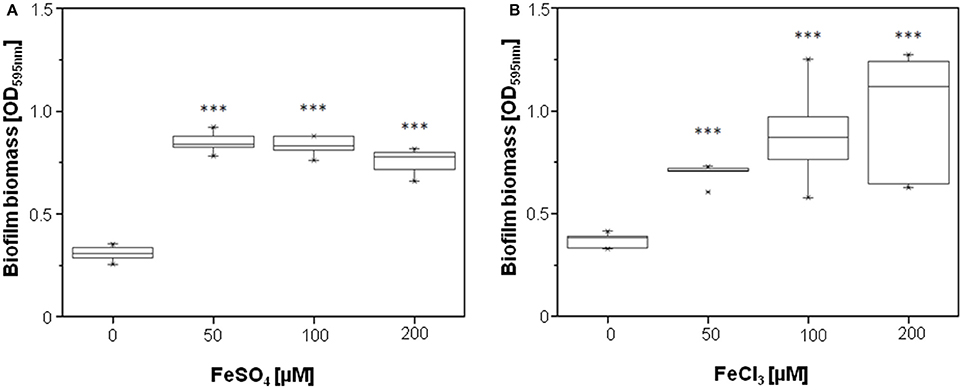
Figure 6. Effect of FeSO4 and FeCl3 on biofilm formation of P. aeruginosa PA14 lasB::Tn. Overnight cultures (in LB) were diluted (10−2) in BM2 medium, and supplemented with FeSO4 (A) or FeCl3 (B) as indicated. After incubation for 24 h at 37°C, cells were stained with 0.1% (w/v) crystal violet and quantified by measuring OD595nm. Experiments were done in triplicates with 6 individual repeats per measurement (n = 18). Statistical analyses were performed with the Mann-Whitney U-test. The highest and lowest outliers are indicated by “X.” ***p ≤ 0.001.
Discussion
Interference with bacterial quorum sensing systems by inactivation of signal molecules has been discussed as a promising anti-virulence strategy, because the signal can hardly become unsusceptible to the quorum quenching enzyme without losing its activity, and because a quorum quenching agent addressing an extracellular target may exert less selection pressure than an agent that acts intracellularly (García-Contreras et al., 2013). However, some drawback of using enzymes to inactivate signal molecules is their susceptibility to denaturation by adverse physicochemical conditions and to proteolytic degradation. In P. aeruginosa, LasB elastase represents one of the major secreted proteins, participating in the proteolytic inactivation of numerous extracellular components of the innate and adaptive immune system of the eukaryotic host (Leduc et al., 2007). LasB also was found to be the basic cause of rapid inactivation of the PQS-cleaving dioxygenase HodC in P. aeruginosa cultures. Thus, to be applicable as a quorum quenching enzyme, the protein needs to be stabilized by enzyme engineering, or protected by immobilization or encapsulation.
HodC-mediated inactivation of PQS quenches the production of virulence factors such as pyocyanin, rhamnolipids, and lectinA, whose production is upregulated in response to PQS signaling (Pustelny et al., 2009). However, HodC-catalyzed cleavage of PQS turned out to stimulate P. aeruginosa biofilm development. This stimulatory effect appears to be due to the fact that PQS cleavage removes not only a signal, but also an iron trap. PQS has been reported to form 2:1 and 3:1 chelate complexes with ferric ions (Diggle et al., 2007). The transcriptome profile of P. aeruginosa in response to PQS revealed a marked induction of genes related to iron acquisition and oxidative stress response; for most genes this differential regulation was due to the iron-chelating effect of PQS (Bredenbruch et al., 2006). Considering that fresh LB medium contains about 6 μM iron (Diggle et al., 2007) and PQS levels (at least in in planktonic cultures) can reach up to ~25 μM, PQS likely induces a limitation of readily bioavailable iron ions.
Both iron limitation and iron excess can adversely affect biofilm formation in P. aeruginosa, with iron excess promoting biofilm dispersal (Singh et al., 2002; Singh, 2004; Banin et al., 2005; Musk et al., 2005; Yang et al., 2007; Patriquin et al., 2008). Interestingly, growth yields of P. aeruginosa biofilm were reported to be affected to a greater extent by iron limitation than planktonic P. aeruginosa cultures (Patriquin et al., 2008). The HodC-mediated increase in viable biofilm biomass as well as the downregulation of a set of iron-controlled genes is in line with the notion that the enzyme-catalyzed PQS cleavage increases iron availability. Under the conditions tested, the growth-promoting effect of iron, released from the PQS-iron complex by degradation of the PQS ligand, overrides the HodC-mediated quenching of the stimulatory effect PQS may have on biofilm development. Thus, under conditions of iron limitation, degradation of PQS is contraindicated for combating P. aeruginosa PA14 biofilms.
Author Contributions
JO, SF, and GB conceived the experiments. CN purified HodC proteins and performed enzyme activity and stability assays. BT and AN performed biofilm experiments and gene expression profiling. AD analyzed the Illumina sequencing data and FK performed the chemical synthesis of N-octanoylanthranilic acid. JO and SF wrote the paper. All authors contributed to the final version of the manuscript, and all authors approved the final manuscript.
Funding
We gratefully acknowledge financial support by the BioInterfaces in Technology and Medicine (BIFTM) Program of the Karlsruhe Institute of Technology (KIT) in the Helmholtz Association, and support by the Deutsche Forschungsgemeinschaft within GRK1409 and grant no. FE 383/25-1.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
SF and CN thank Philip Weyrauch for the construction of pET23a-hodC and Almut Kappius for technical assistance. JO and AD thank Olivier Armant for excellent assistance with the Illumina sequencing.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01978/full#supplementary-material
References
Anders, S., Pyl, P. T., and Huber, W. (2015). HTSeq - a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169. doi: 10.1093/bioinformatics/btu638
Banin, E., Vasil, M. L., and Greenberg, E. P. (2005). Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. U.S.A. 102, 11076–11081. doi: 10.1073/pnas.0504266102
Beermann, B., Guddorf, J., Boehm, K., Albers, A., Kolkenbrock, S., Fetzner, S., et al. (2007). Stability, unfolding, and structural changes of cofactor-free 1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase. Biochemistry 46, 4241–4249. doi: 10.1021/bi0622423
Bredenbruch, F., Geffers, R., Nimtz, M., Buer, J., and Häussler, S. (2006). The Pseudomonas aeruginosa quinolone signal (PQS) has an iron-chelating activity. Environ. Microbiol. 8, 1318–1329. doi: 10.1111/j.1462-2920.2006.01025.x
Breidenstein, E. B. M., Janot, L., Strehmel, J., Fernandez, L., Taylor, P. K., Kukavica-Ibrulj, I., et al. (2012). The Lon protease is essential for full virulence in Pseudomonas aeruginosa. PLoS ONE 7:e49123. doi: 10.1371/journal.pone.0049123
Caballero, A. R., Moreau, J. M., Engel, L. S., Marquart, M. E., Hill, J. M., and O'Callaghan, R. J. (2001). Pseudomonas aeruginosa protease IV enzyme assays and comparison to other Pseudomonas proteases. Anal. Biochem. 290, 330–337. doi: 10.1006/abio.2001.4999
Déziel, E., Gopalan, S., Tampakaki, A. P., Lépine, F., Padfield, K. E., Saucier, M., et al. (2005). The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-L-homoserine lactones. Mol. Microbiol. 55, 998–1014. doi: 10.1111/j.1365-2958.2004.04448.x
Diggle, S. P., Matthijs, S., Wright, V. J., Fletcher, M. P., Chhabra, S. R., Lamont, I. L., et al. (2007). The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem. Biol. 14, 87–96. doi: 10.1016/j.chembiol.2006.11.014
Diggle, S. P., Winzer, K., Chhabra, S. R., Worrall, K. E., Cámara, M., and Williams, P. (2003). The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol. 50, 29–43. doi: 10.1046/j.1365-2958.2003.03672.x
Elias, S., Degtyar, E., and Banin, E. (2011). FvbA is required for vibriobactin utilization in Pseudomonas aeruginosa. Microbiology 157, 2172–2180. doi: 10.1099/mic.0.044768-0
Fetzner, S. (2015). Quorum quenching enzymes. J. Biotechnol. 201, 2–14. doi: 10.1016/j.jbiotec.2014.09.001
Frangipani, E., Visaggio, D., Heeb, S., Kaever, V., Cámara, M., Visca, P., et al. (2014). The Gac/Rsm and cyclic-di-GMP signalling networks coordinately regulate iron uptake in Pseudomonas aeruginosa. Environ. Microbiol. 16, 676–688. doi: 10.1111/1462-2920.12164
Frerichs-Deeken, U., Ranguelova, K., Kappl, R., Hüttermann, J., and Fetzner, S. (2004). Dioxygenases without requirement for cofactors and their chemical model reaction: compulsory order ternary complex mechanism of 1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase involving general base catalysis by histidine 251 and single-electron oxidation of the substrate dianion. Biochemistry 43, 14485–14499. doi: 10.1021/bi048735u
Friedman, L., and Kolter, R. (2004). Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol. Microbiol. 51, 675–690. doi: 10.1046/j.1365-2958.2003.03877.x
García-Contreras, R., Maeda, T., and Wood, T. K. (2013). Resistance to quorum-quenching compounds. Appl. Environ. Microbiol. 79, 6840–6846. doi: 10.1128/aem.02378-13
Gellatly, S. L., and Hancock, R. E. W. (2013). Pseudomonas aeruginosa: new insights into pathogenesis and host defenses. Path. Dis. 67, 159–173. doi: 10.1111/2049-632x.12033
Guo, Q., Kong, W. N., Jin, S., Chen, L., Xu, Y. Y., and Duan, K. M. (2014). PqsR-dependent and PqsR-independent regulation of motility and biofilm formation by PQS in Pseudomonas aeruginosa PAO1. J. Basic Microbiol. 54, 633–643. doi: 10.1002/jobm.201300091
Hänsch, G. M., Prior, B., Brenner-Weiss, G., Obst, U., and Overhage, J. (2014). The Pseudomonas quinolone signal (PQS) stimulates chemotaxis of polymorphonuclear neutrophils. J. Appl. Biomater. Funct. Mater. 12, 21–26. doi: 10.5301/jabfm.5000204
Hassett, D. J., Howell, M. L., Ochsner, U. A., Vasil, M. L., Johnson, Z., and Dean, G. E. (1997). An operon containing fumC and sodA encoding fumarase C and manganese superoxide dismutase is controlled by the ferric uptake regulator in Pseudomonas aeruginosa: fur mutants produce elevated alginate levels. J. Bacteriol. 179, 1452–1459. doi: 10.1128/jb.179.5.1452-1459.1997
Häussler, S., and Becker, T. (2008). The Pseudomonas quinolone signal (PQS) balances life and death in Pseudomonas aeruginosa populations. PLoS Pathog. 4:e1000166. doi: 10.1371/journal.ppat.1000166
Herigstad, B., Hamilton, M., and Heersink, J. (2001). How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 44, 121–129. doi: 10.1016/s0167-7012(00)00241-4
Hooi, D. S. W., Bycroft, B. W., Chhabra, S. R., Williams, P., and Pritchard, D. I. (2004). Differential immune modulatory activity of Pseudomonas aeruginosa quorum-sensing signal molecules. Infect. Immun. 72, 6463–6470. doi: 10.1128/iai.72.11.6463-6470.2004
Hutchison, M. L., and Govan, J. R. W. (1999). Pathogenicity of microbes associated with cystic fibrosis. Microbes Infect. 1, 1005–1014. doi: 10.1016/S1286-4579(99)80518-8
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Leduc, D., Beaufort, N., de Bentzmann, S., Rousselle, J.-C., Namane, A., Chignard, M., et al. (2007). The Pseudomonas aeruginosa LasB metalloproteinase regulates the human urokinase-type plasminogen activator receptor through domain-specific endoproteolysis. Infect. Immun. 75, 3848–3858. doi: 10.1128/iai.00015-07
Lépine, F., Déziel, E., Milot, S., and Rahme, L. G. (2003). A stable isotope dilution assay for the quantification of the Pseudomonas quinolone signal in Pseudomonas aeruginosa cultures. Biochim. Biophys. Acta 1622, 36–41. doi: 10.1016/s0304-4165(03)00103-x
Lesic, B., Lépine, F., Déziel, E., Zhang, J., Zhang, Q., Padfield, K., et al. (2007). Inhibitors of pathogen intercellular signals as selective anti-infective compounds. PLoS Pathog. 3:e126. doi: 10.1371/journal.ppat.0030126
Li, J., Kleintschek, T., Rieder, A., Cheng, Y., Baumbach, T., Obst, U., et al. (2013). Hydrophobic liquid-infused porous polymer surfaces for antibacterial applications. ACS Appl. Mater. Interfaces 5, 6704–6711. doi: 10.1021/am401532z
Liberati, N. T., Urbach, J. M., Miyata, S., Lee, D. G., Drenkard, E., Wu, G., et al. (2006). An ordered, nonredundant library of Pseudomonas aeruginosa strain PA14 transposon insertion mutants. Proc. Natl. Acad. Sci. U.S.A. 103, 2833–2838. doi: 10.1073/pnas.0511100103
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lu, C., Maurer, C. K., Kirsch, B., Steinbach, A., and Hartmann, R. W. (2014). Overcoming the unexpected functional inversion of a PqsR antagonist in Pseudomonas aeruginosa: an in vivo potent antivirulence agent targeting pqs quorum sensing. Angew. Chem. Int. Ed. Engl. 53, 1109–1112. doi: 10.1002/anie.201307547
Mashburn, L. M., and Whiteley, M. (2005). Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 437, 422–425. doi: 10.1038/nature03925
Murray, T. S., Okegbe, C., Gao, Y., Kazmierczak, B. I., Motterlini, R., Dietrich, L. E., et al. (2012). The carbon monoxide releasing molecule CORM-2 attenuates Pseudomonas aeruginosa biofilm formation. PLoS ONE 7:e35499. doi: 10.1371/journal.pone.0035499
Musk, D. J., Banko, D. A., and Hergenrother, P. J. (2005). Iron salts perturb biofilm formation and disrupt existing biofilms of Pseudomonas aeruginosa. Chem. Biol. 12, 789–796. doi: 10.1016/j.chembiol.2005.05.007
Müsken, M., Di Fiore, S., Dötsch, A., Fischer, R., and Häussler, S. (2010). Genetic determinants of Pseudomonas aeruginosa biofilm establishment. Microbiology 156, 431–441. doi: 10.1099/mic.0.033290-0
Ochsner, U. A., Wilderman, P. J., Vasil, A. I., and Vasil, M. L. (2002). GeneChip® expression analysis of the iron starvation response in Pseudomonas aeruginosa: identification of novel pyoverdine biosynthesis genes. Mol. Microbiol. 45, 1277–1287. doi: 10.1046/j.1365-2958.2002.03084.x
Overhage, J., Bains, M., Brazas, M. D., and Hancock, R. E. W. (2008). Swarming of Pseudomonas aeruginosa is a complex adaptation leading to increased production of virulence factors and antibiotic resistance. J. Bacteriol. 190, 2671–2679. doi: 10.1128/jb.01659-07
Patriquin, G. M., Banin, E., Gilmour, C., Tuchman, R., Greenberg, E. P., and Poole, K. (2008). Influence of quorum sensing and iron on twitching motility and biofilm formation in Pseudomonas aeruginosa. J. Bacteriol. 190, 662–671. doi: 10.1128/jb.01473-07
Pesci, E. C., Milbank, J. B. J., Pearson, J. P., McKnight, S., Kende, A. S., Greenberg, E. P., et al. (1999). Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 96, 11229–11234. doi: 10.1073/pnas.96.20.11229
Pustelny, C., Albers, A., Büldt-Karentzopoulos, K., Parschat, K., Chhabra, S. R., Cámara, M., et al. (2009). Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem. Biol. 16, 1259–1267. doi: 10.1016/j.chembiol.2009.11.013
Rahme, L. G., Stevens, E. J., Wolfort, S. F., Shao, J., Tompkins, R. G., and Ausubel, F. M. (1995). Common virulence factors for bacterial pathogenicity in plants and animals. Science 268, 1899–1902. doi: 10.1126/science.7604262
Rajan, S., and Saiman, L. (2002). Pulmonary infections in patients with cystic fibrosis. Semin. Respir. Infect. 17, 47–56. doi: 10.1053/srin.2002.31690
Schweizer, H. P., and Po, C. (1996). Regulation of glycerol metabolism in Pseudomonas aeruginosa: characterization of the glpR repressor gene. J. Bacteriol. 178, 5215–5221. doi: 10.1128/jb.178.17.5215-5221.1996
Singh, P. K. (2004). Iron sequestration by human lactoferrin stimulates P. aeruginosa surface motility and blocks biofilm formation. Biometals 17, 267–270. doi: 10.1023/B:BIOM.0000027703.77456.27
Singh, P. K., Parsek, M. R., Greenberg, E. P., and Welsh, M. J. (2002). A component of innate immunity prevents bacterial biofilm development. Nature 417, 552–555. doi: 10.1038/417552a
Skindersoe, M. E., Zeuthen, L. H., Brix, S., Fink, L. N., Lazenby, J., Whittall, C., et al. (2009). Pseudomonas aeruginosa quorum-sensing signal molecules interfere with dendritic cell-induced T-cell proliferation. FEMS Immunol. Med. Microbiol. 55, 335–345. doi: 10.1111/j.1574-695X.2008.00533.x
Sonnleitner, E., and Haas, D. (2011). Small RNAs as regulators of primary and secondary metabolism in Pseudomonas species. Appl. Microbiol. Biotechnol. 91, 63–79. doi: 10.1007/s00253-011-3332-1
Storz, M. P., Maurer, C. K., Zimmer, C., Wagner, N., Brengel, C., de Jong, J. C., et al. (2012). Validation of PqsD as an anti-biofilm target in Pseudomonas aeruginosa by development of small-molecule inhibitors. J. Am. Chem. Soc. 134, 16143–16146. doi: 10.1021/ja3072397
Tettmann, B., Dötsch, A., Armant, O., Fjell, C. D., and Overhage, J. (2014). Knockout of extracytoplasmic function sigma factor ECF-10 affects stress resistance and biofilm formation in Pseudomonas putida KT2440. Appl. Environ. Microbiol. 80, 4911–4919. doi: 10.1128/aem.01291-14
Thaden, J. T., Lory, S., and Gardner, T. S. (2010). Quorum-sensing regulation of a copper toxicity system in Pseudomonas aeruginosa. J. Bacteriol. 192, 2557–2568. doi: 10.1128/jb.01528-09
Wade, D. S., Calfee, M. W., Rocha, E. R., Ling, E. A., Engstrom, E., Coleman, J. P., et al. (2005). Regulation of Pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa. J. Bacteriol. 187, 4372–4380. doi: 10.1128/JB.187.13.4372-4380.2005
Wells, I. C., Elliott, W. H., Thayer, S. A., and Doisy, E. A. (1952). Ozonization of some antibiotic substances produced by Pseudomonas aeruginosa. J. Biol. Chem. 196, 321–330.
Wilderman, P. J., Sowa, N. A., FitzGerald, D. J., FitzGerald, P. C., Gottesman, S., Ochsner, U. A., et al. (2004). Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. U.S.A. 101, 9792–9797. doi: 10.1073/pnas.0403423101
Williams, P., Winzer, K., Chan, W. C., and Cámara, M. (2007). Look who's talking: communication and quorum sensing in the bacterial world. Philos. Trans. R. Soc. London Ser B 362, 1119–1134. doi: 10.1098/rstb.2007.2039
Winsor, G. L., van Rossum, T., Lo, R., Khaira, B., Whiteside, M. D., Hancock, R. E. W., et al. (2009). Pseudomonas Genome Database: facilitating user-friendly, comprehensive comparisons of microbial genomes. Nucl. Acids Res. 37, D483–D488. doi: 10.1093/nar/gkn861
Xiao, G. P., Déziel, E., He, J. X., Lépine, F., Lésic, B., Castonguay, M. H., et al. (2006). MvfR, a key Pseudomonas aeruginosa pathogenicity LTTR-class regulatory protein, has dual ligands. Mol. Microbiol. 62, 1689–1699. doi: 10.1111/j.1365-2958.2006.05462.x
Yang, L., Barken, K. B., Skindersoe, M. E., Christensen, A. B., Givskov, M., and Tolker-Nielsen, T. (2007). Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology 153, 1318–1328. doi: 10.1099/mic.0.2006/004911-0
Keywords: Pseudomonas aeruginosa, Pseudomonas quinolone signal, quorum sensing, quorum quenching, biofilm, PQS dioxygenase
Citation: Tettmann B, Niewerth C, Kirschhöfer F, Neidig A, Dötsch A, Brenner-Weiss G, Fetzner S and Overhage J (2016) Enzyme-Mediated Quenching of the Pseudomonas Quinolone Signal (PQS) Promotes Biofilm Formation of Pseudomonas aeruginosa by Increasing Iron Availability. Front. Microbiol. 7:1978. doi: 10.3389/fmicb.2016.01978
Received: 21 July 2016; Accepted: 25 November 2016;
Published: 09 December 2016.
Edited by:
Janice Lorraine Strap, University of Ontario Institute of Technology, CanadaReviewed by:
Akos T. Kovacs, University of Jena, GermanyAnton Hartmann, Helmholtz Zentrum München, Germany
Copyright © 2016 Tettmann, Niewerth, Kirschhöfer, Neidig, Dötsch, Brenner-Weiss, Fetzner and Overhage. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joerg Overhage, joerg.overhage@kit.edu
†Susanne Fetzner and Joerg Overhage are joint senior authors.