 Mikkel Stokholm-Bjerregaard†
Mikkel Stokholm-Bjerregaard† Simon J. McIlroy†
Simon J. McIlroy† Marta Nierychlo
Marta Nierychlo Søren M. Karst
Søren M. Karst Mads Albertsen
Mads Albertsen Per H. Nielsen*
Per H. Nielsen*- Center for Microbial Communities, Department of Chemistry and Bioscience, Aalborg University, Aalborg, Denmark
Understanding the microbiology of phosphorus (P) removal is considered essential to knowledge-based optimization of enhanced biological P removal (EBPR) systems. Biological P removal is achieved in these systems by promoting the growth of organisms collectively known as the polyphosphate accumulating organisms (PAOs). Also considered important to EBPR are the glycogen accumulating organisms (GAOs), which are theorized to compete with the PAOs for resources at the expense of P removal efficiency. Numerous studies have sought to identify the PAOs and their GAOs competitors, with several candidates proposed for each over the last few decades. The current study collectively assessed the abundance and diversity of all proposed PAOs and GAOs in 18 Danish full-scale wastewater treatment plants with well-working biological nutrient removal over a period of 9 years using 16S rRNA gene amplicon sequencing. The microbial community structure in all plants was relatively stable over time. Evidence for the role of the proposed PAOs and GAOs in EBPR varies and is critically assessed, in light of their calculated amplicon abundances, to indicate which of these are important in full-scale systems. Bacteria from the genus Tetrasphaera were the most abundant of the PAOs. The “Candidatus Accumulibacter” PAOs were in much lower abundance and appear to be biased by the amplicon-based method applied. The genera Dechloromonas, Microlunatus, and Tessaracoccus were identified as abundant putative PAO that require further research attention. Interestingly, the actinobacterial Micropruina and sbr-gs28 phylotypes were among the most abundant of the putative GAOs. Members of the genera Defluviicoccus, Propionivibrio, the family Competibacteraceae, and the spb280 group were also relatively abundant in some plants. Despite observed high abundances of GAOs (periodically exceeding 20% of the amplicon reads), P removal performance was maintained, indicating that these organisms were not outcompeting the PAOs in these EBPR systems. Phylogenetic diversity within each of the PAOs and GAOs genera was observed, which is consistent with reported metabolic diversity for these. Whether or not key traits can be assigned to sub-genus level clades requires further investigation.
Introduction
Wastewater treatment using the enhanced biological phosphorus removal (EBPR) process is capable of achieving low effluent phosphorus (P) concentrations without addition of chemical precipitants. EBPR systems achieve this by utilizing the ability of polyphosphate accumulating organisms (PAOs) to take up and store excessive amounts of P. Hence, compared to chemical precipitation, EBPR is a sustainable, effective, and economical process (Seviour et al., 2003). In addition, P is an essential nutrient used in fertilizers and is a limited non-renewable resource (Gilbert, 2009). Thus, P-rich sludge from EBPR plants is seen as a high quality, sustainable, and economical alternative source of P (Jeanmaire and Evans, 2001; Molinos-Senante et al., 2011). As EBPR systems are at times subject to failure, or periods of low efficiency, an understanding of the microbiology that underpins the process is important for knowledge-based optimization. As such, many studies on the identity, diversity, and physiology of PAOs and their proposed competitors, the glycogen accumulating organisms (GAOs), have been carried out (Oehmen et al., 2007).
Early PAO metabolic models were proposed to explain the key transformations of the dynamic EBPR environment, where biomass is cycled between carbon rich anaerobic (feast) and carbon deficient aerobic (famine) zones (Comeau et al., 1986; Wentzel et al., 1986; Mino et al., 1987). Volatile fatty acids (VFAs) are stored as polyhydroxyalkanoates (PHAs) under anaerobic conditions, where aerobically stored polyphosphate and glycogen provide the energy and reducing equivalents required. In the subsequent aerobic phase, stored PHAs are utilized for growth and regeneration of glycogen and polyphosphate stores (Oehmen et al., 2007). Substantial effort has been made to identify organisms that conform to these proposed models. Early culture-based approaches identified several activated sludge isolates, which accumulated excess polyphosphate. Of these isolates, members of the genus Acinetobacter received considerable research attention but were never shown to conform to PAO models (Jenkins and Tandoi, 1991; Tandoi et al., 1998a; Seviour et al., 2003), and their low abundance in full-scale systems (Wagner et al., 1994) appears to be maintained by immigration with the influent rather than active growth (Saunders et al., 2016). The betaproteobacterial isolate Lampropedia hyalina was shown to behave according to the proposed PAO models, with anaerobic storage of acetate as PHA, although associated P cycling was much lower than for EBPR biomass (Stante et al., 1997) and its abundance in full-scale EBPR systems is not known. The role of other isolates has not been assessed further than their ability to store polyphosphate. These include Tessarococcus bendigoensis (Maszenan et al., 1999b), Friedmaniella spp. (Maszenan et al., 1999a), Quatrionicoccus australiensis (Maszenan et al., 2002), Gemmatimonas aurantiaca (Zhang et al., 2003), and Malikia granosa (Spring et al., 2005).
The advent of molecular methods revolutionized our understanding of EBPR microbiology generally. In particular, fluorescence in situ hybridization (FISH) is applied to assess in situ abundances and, when coupled with other methods, key metabolic traits can be demonstrated (Wagner and Haider, 2012). Typically, FISH has been applied with microautoradiography (MAR; Nierychlo et al., 2015) and fluorescent staining methods (Serafim et al., 2002) to demonstrate in situ uptake of carbon and P and, PHA and polyphosphate storage, respectively. Raman-spectroscopy and novel staining methods should also allow in situ analyses of glycogen (Huang et al., 2007; Majed and Gu, 2010; Mesquita et al., 2013). Such methods have been applied to at least partially demonstrate the PAO phenotype in situ for “Candidatus Accumulibacter spp.” (Hesselmann et al., 1999; Crocetti et al., 2000; Kong et al., 2004), Tetrasphaera spp. (Kong et al., 2005; Nguyen et al., 2011), Microlunatus spp. (Kawaharasaki et al., 1999; Beer et al., 2006), Dechloromonas spp. (Kong et al., 2007; Günther et al., 2009), Pseudomonas spp. (Günther et al., 2009), and “Candidatus Accumulimonas spp.” (formerly “Candidatus Halomonas phosphatis”; Nguyen et al., 2012).
Changing the way we define PAOs in EBPR, the characterization of the actinobacterial genus Tetrasphaera found that these organisms cycle polyphosphate, but without PHA accumulation, which is not consistent with the original PAO models. Instead, they seem to mainly utilize sugars and amino acids through a fermentative metabolism, where polyphosphate supplements anaerobic energy requirements (Kong et al., 2005; Nguyen et al., 2011; Kristiansen et al., 2013). Members of the related actinobacterial genus Microlunatus also appear to possess a similar physiology (Nakamura et al., 1995a,b; Santos et al., 1999; Kawakoshi et al., 2012). Therefore, in the context of EBPR, the PAO is now more broadly defined to cover any organism utilizing aerobically stored polyphosphate to energize anaerobic carbon uptake (Seviour and McIlroy, 2008).
The GAO phenotype model is similar to that of the traditional “Ca. Accumulibacter” PAO phenotype, except polyphosphate is not cycled, which is compensated by a reliance on glycogen as energy source under anaerobic conditions (Liu et al., 1994; Mino et al., 1995). As they compete with the PAO for resources without contributing to P removal, their proliferation is widely considered to be at the expense of EBPR efficiency (Seviour et al., 2003; Oehmen et al., 2007). As with the PAO, early culture-based studies sought to identify organisms conforming to the proposed GAO model. Because of the frequent association of EBPR failure and the proliferation of tetrad-forming organisms (TFOs), there was a focus on organisms with this morphology accumulating PHAs and glycogen but not polyphosphate—originally referred to as the “G-bacteria,” named for their suggested affinity for glucose assimilation and defined by their TFO morphology (Cech and Hartman, 1993; Seviour et al., 2000). Several isolates from poorly performing systems were suggested. Of these isolates, the GAO phenotype has only been confirmed for Defluviicoccus vanus (Wong and Liu, 2007). Amaricoccus kaplicensis, the original G-bacteria, has not been shown to accumulate substrates under anaerobic conditions, which is key to the GAO phenotype (Falvo et al., 2001). Little is known of the potential importance of other isolates, including the actinobacterial Nakamurella multipartita (Yoshimi et al., 1996), Micropruina glycogenica (Shintani et al., 2000), and Kineosphaera limosa (Liu et al., 2000, 2002).
The application of molecular methods has also allowed the study of the putative GAO in situ where the phenotype has been at least partially confirmed for: the actinobacterial genus Micropruina and closely related sbr-gs28 taxon (Kong et al., 2001), the gammaproteobacterial family Competibacteraceae (Kong et al., 2006) and CCM19a phylotype (probe Gam445) (Kong et al., 2007), the alphaproteobacterial genus Defluviicoccus (Wong et al., 2004; Meyer et al., 2006; Burow et al., 2007; Wong and Liu, 2007; McIlroy et al., 2010), and the betaproteobacterial genera Propionivibrio (Albertsen et al., 2016), and Spb280 (probe Bet65; Kong et al., 2007).
The PAO and GAO phenotypes appear to encompass both phylogenetically and metabolically diverse organisms. Importantly, most studies into the PAO and GAO are based on lab-scale studies with VFAs, such as acetate, as the sole carbon source. In these systems, the “Ca. Accumulibacter” PAO, and the Competibacteraceae and Defluviicoccus GAO, are almost exclusively found, and are therefore widely considered to be the most important groups to EBPR. However, it is still unclear which of the putative PAOs and GAOs are actually the key organisms in full-scale plants, noting that these systems will harbor a much higher phylogenetic and phenotypic diversity relative to the lab-scale environment. For example, large-scale FISH surveys have revealed that Tetrasphaera spp. are the most abundant genus in Danish full-scale EBPR systems (Mielczarek et al., 2013), which brings into question the belief that the “Ca. Accumulibacter” are the most important PAO phylotype. Developments in protocols and technology for the amplicon sequencing of the 16S rRNA gene now allow high-throughput analyses of the microbial community composition in full-scale systems (Zhang et al., 2012; Saunders et al., 2016). The on-going Microbial Database of Activated Sludge (MiDAS) initiative implements this technology to survey the bacterial communities of 18 Danish wastewater treatment plants (WWTPs) over a period of 9 years1 (McIlroy et al., 2015b).
The current study will, for the first time, collectively assess the relative abundance and distribution of all suggested PAO and GAO in Danish full-scale EBPR WWTPs, using the extensive MiDAS survey data. In this article, the available literature for each suggested PAO and GAO is critically assessed, along with their relative amplicon-based abundance, to indicate their potential importance in full-scale systems. In doing so, the study identifies key organisms and questions for the focus of future studies into the microbiology of EBPR.
Materials and Methods
Treatment Plants and Sampling
Sampling and investigation of the microbial populations in the activated sludge biomass from 18 plants with EBPR was done within the MiDAS project (McIlroy et al., 2015b). The plants were sampled up to four times a year from 2006 to 2014 giving a total of 414 samples. They were sampled February, May, August, and October. All samples were taken from the aeration tank and sent by over night mail to our laboratory for processing.
The WWTPs investigated included the following variations in design and operation: +/- primary settling, +/- digester, alternating/recirculation flow, +/- return sludge sidestream hydrolysis (RSS), different industrial loading, and different type and amount of external carbon source added (Table 1). All plants were municipal WWTPs located in Denmark treating primarily household wastewater with contributions from industry of 5–75% of the influent chemical oxygen demand (COD). All plants had EBPR and supplementary chemical P removal. Median temperature was similar in all plants and ranged from 9°C in the winter and 18°C in the summer. pH was stable at 7.2 (±0.3). The median suspended solids (SS) was 4.9 g L-1 with some seasonal variation from 4.4 g L-1 in summer to 5.1 g L-1 in winter. Median influent total COD, total nitrogen (N), and total P after primary treatment (if present) were 528, 42, and 7.3 mg L-1.
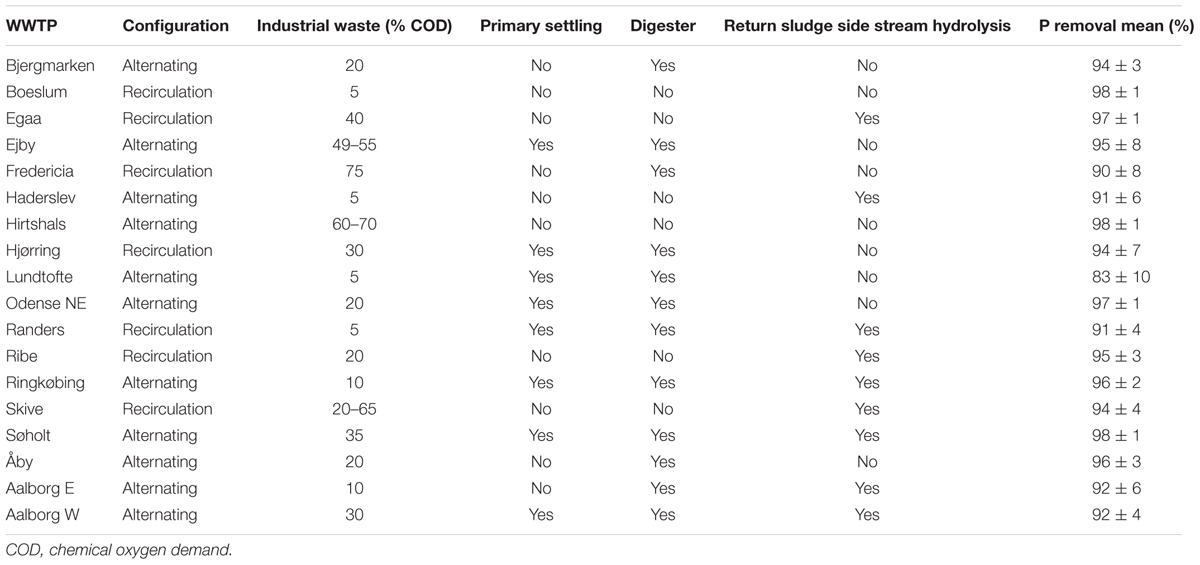
TABLE 1. Design and operation of wastewater treatment plants (WWTPs) included in the survey.
All plants were operated under the same minimum effluent quality regulations of 75 mg L-1 total COD, 8 mg L-1 total N, and 1.5 mg L-1 total P. The actual effluent quality was, however, better, with median effluent COD, N, and P of 25, 4, and 0.3 mg L-1, respectively. P removal efficiency was calculated as the percentage of the removed total P by comparing monthly mean values of influent and effluent concentrations.
DNA Extraction and Amplicon Sequencing
DNA extraction was conducted using the FastDNA spin kit for soil (MP Biomedicals) according to the manufactures instructions, except the bead beating was increased to 4 × 40 s at 6 m/s using a FastPrep FP120 (MP Biomedicals). The procedure for bacterial 16S rRNA amplicon sequencing targeting the V1-3 variable region was modified from Caporaso et al. (2010). Briefly, 10 ng of extracted DNA was used as template and the polymerase chain reaction (PCR) (25 μL) contained dNTPs (400 nM of each), MgSO4 (1.5 mM), Platinum® Taq DNA polymerase high fidelity (HF) (2 mU), 1× Platinum® High Fidelity buffer (Thermo Fisher Scientific) and a pair of barcoded library adaptors (400 nM). V1-3 primers: 27F AGAGTTTGATCCTGGCTCAG and 534R ATTACCGCGGCTGCTGG (Chen et al., 2010). Thermo cycler settings: initial denaturation at 95°C for 2 min, 30 cycles of 95°C for 20 s, 56°C for 30 s, 72°C for 60 s, and final elongation at 72°C for 5 min. All PCR were run in duplicate and pooled afterward. The amplicon libraries were purified using the Agencourt® AMpure XP bead protocol (Beckmann Coulter, Brea, CA, USA) with the following exceptions: the sample/bead solution ratio was 5/4 and the purified DNA was eluted in 33 μL nuclease-free water. Library concentration was measured with Quant-iTTM HS DNA Assay (Thermo Fisher Scientific) and quality validated with a Tapestation 2200 using D1K ScreenTapes (Agilent). Based on library concentrations and calculated amplicon sizes the samples were pooled in equimolar concentrations and diluted to 4 nM. The library pool was sequenced on a MiSeq (Illumina) using a MiSeq Reagent kit v3 [2 × 300 paired end (PE)] following the procedure in Caporaso et al. (2012), with exception of 10% PhiX control library (Illumina) spike-in and final library loading concentration of 20 pM. A complete detailed DNA extraction and amplicon sequencing protocol can be obtained from2 and the optimization of the protocols applied in this study is detailed in Albertsen et al. (2015).
16S rRNA Data Processing, Analysis, and Visualization
All sequenced sample libraries were subsampled to 50,000 raw reads and low quality reads removed using Trimmomatic v. 0.32 with the settings SLIDINGWINDOW:1:3 and MINLEN:275 (Lohse et al., 2012). Forward and reverse reads were merged using FLASH v. 1.2.7 (Magoč and Salzberg, 2011), with the settings -m 25 -M 200 and afterward merged reads smaller than 425 bp or larger than 525 bp were discarded. All merged reads were screened for PhiX contamination using usearch v. 7.0.1090 (Edgar, 2010), with standard settings and all matching reads removed. The potential PhiX contamination is due to the use of an un-indexed PhiX as a quality control, which can result in index-carryover from nearby clusters with indexes. The merged reads were de-replicated and formatted for use in the UPARSE workflow (Edgar, 2013). The merged reads were clustered into operational taxonomic units (OTUs) (97% similarity) using the usearch v. 7.0.1090-cluster_otus with default settings. OTU abundance was estimated using the usearch v. 7.0.1090–usearch_global with –id 0.97.
Taxonomy was assigned using the Ribosomal Database Project (RDP) classifier (Wang et al., 2007) as implemented in the parallel_assign_taxonomy_rdp.py script in Quantitative Insights into Microbial Ecology (QIIME) (Caporaso et al., 2010) using MiDAS taxonomy version 2.1. All data analysis and visualizations were conducted using R (R Core Team, 2014) through the Rstudio IDE3. OTU counts and associated taxonomic assignments were imported and merged to a phyloseq object (McMurdie and Holmes, 2013) and analyzed using the ampvis R package4. The median abundance of the OTUs was determined from the 18 WWTPs with most samples in the database (n = 414). The raw V1-3 16S rRNA amplicon sequences are part of the MiDAS dataset5 available in European Nucleotide Archive (ENA) with the project ID PRJEB19518. All processed data is included in the ampvis R package as a data object: data (MiDAS_1.20).
It is worth noting that these 16S rRNA gene counts are abundance estimations, and not directly comparable to the true biomass fraction due to DNA extraction biases, primer biases, variation in cell size, and 16S rRNA gene copy number between species (Albertsen et al., 2015).
The genera proposed to behave as PAO and GAO in EBPR systems are listed in Tables 2, 3 and their phylogeny is shown in Figure 1. Classification is based on the MiDAS taxonomy release 2.1 (McIlroy et al., 2015b), which is a manually curated version of the SILVA 123 NR99 database (Quast et al., 2013), updated to include abundant organisms in wastewater and activated sludge treatment systems not covered by commonly applied public databases. Putative PAO and GAO not covered by other databases include the “Ca. Accumulimonas” PAO and the Competibacteraceae, sbr-gs28, spb280, and CCM19a GAO phylotypes.
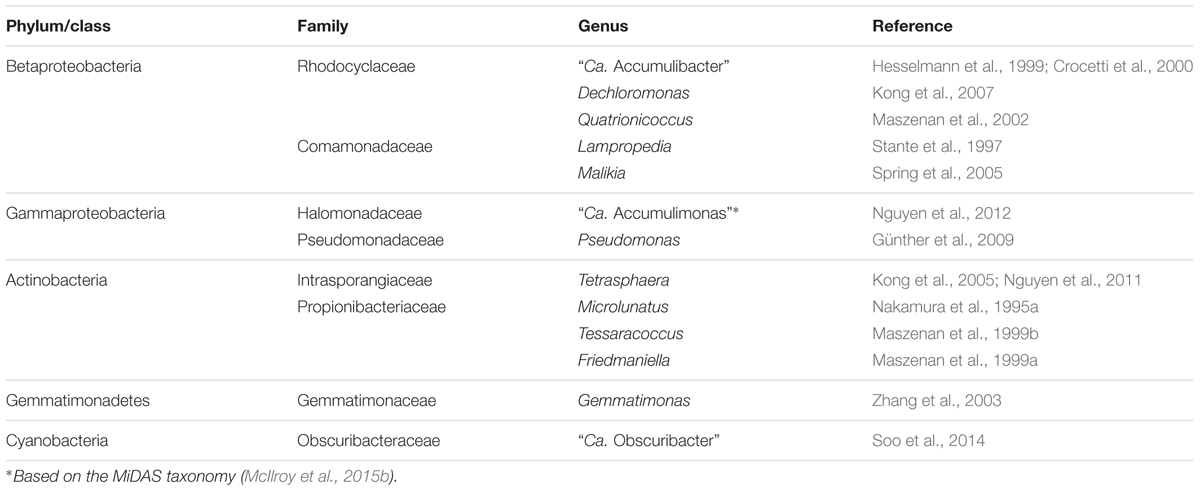
TABLE 2. Summary of the putative PAO.
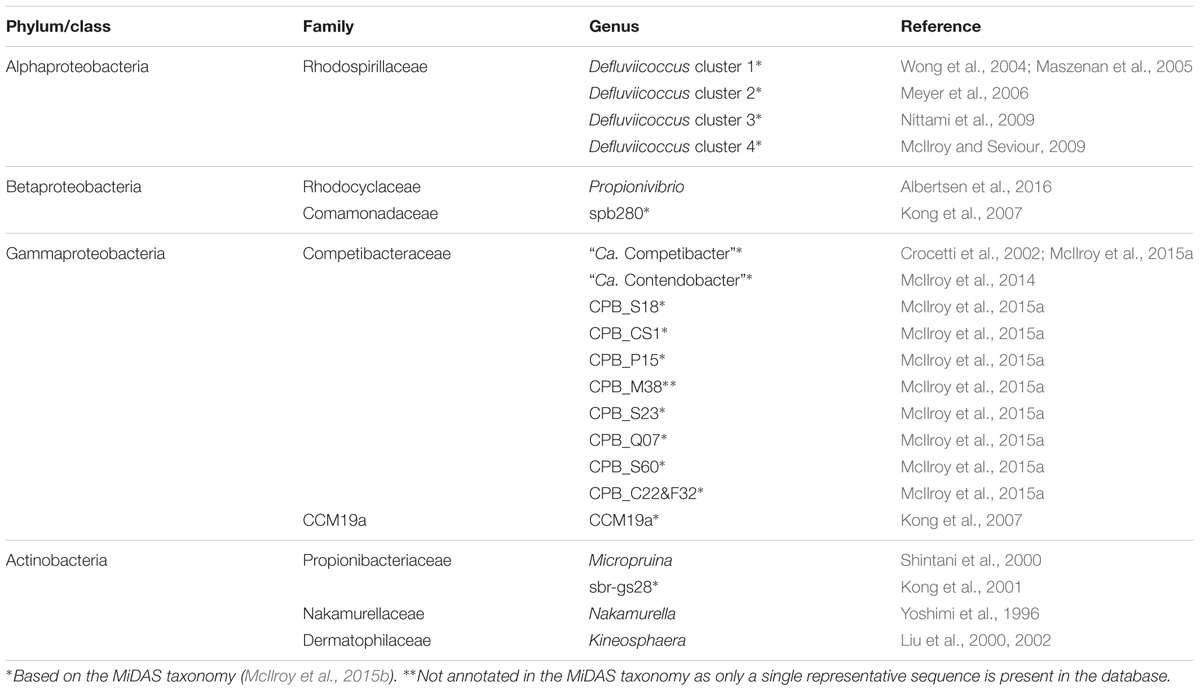
TABLE 3. Summary of the putative GAO.
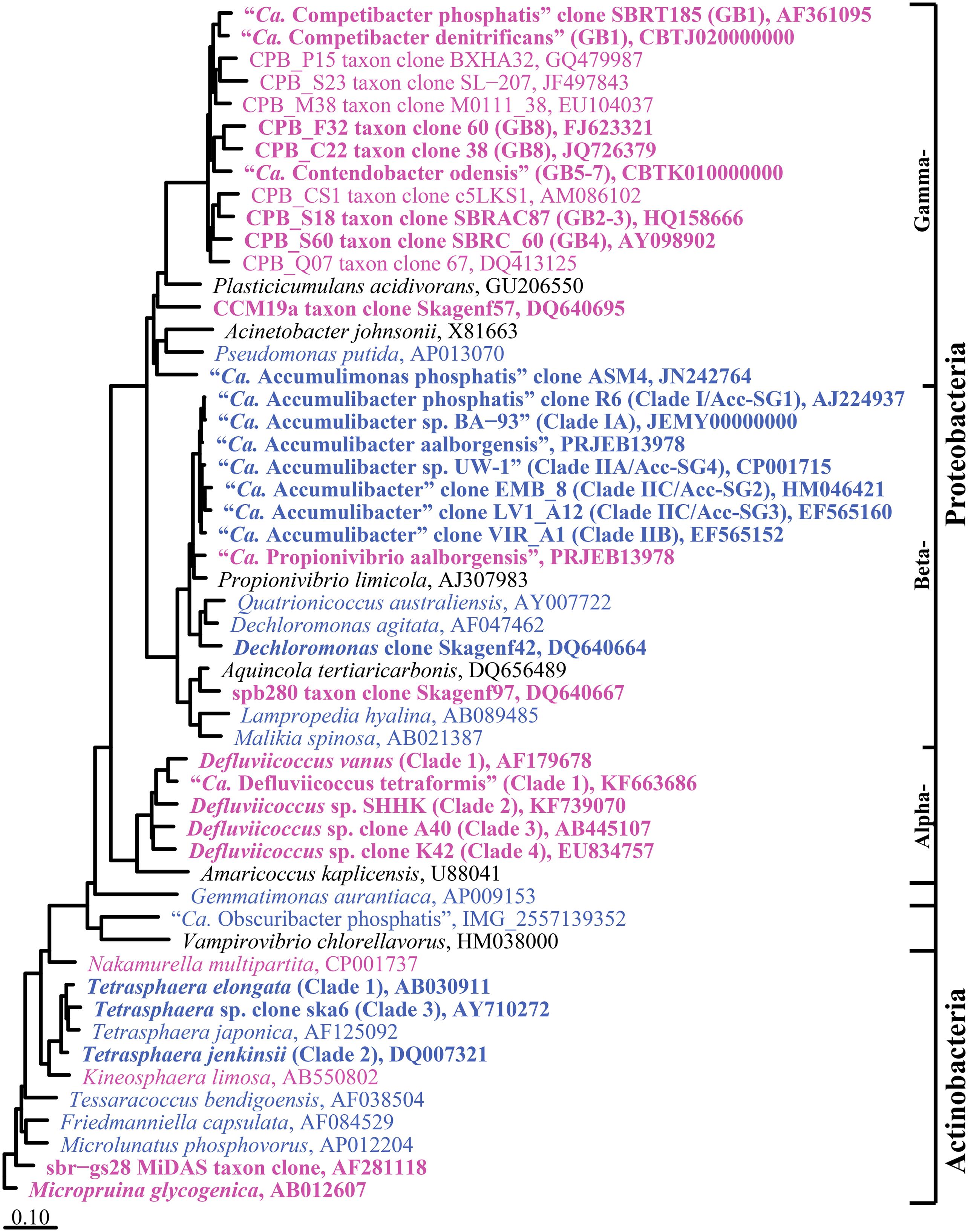
FIGURE 1. Maximum-likelihood (PhyML) 16S rRNA gene phylogenetic tree for suggested PAO and GAO in EBPR activated sludge wastewater treatment plants (WWTPs). Putative PAO sequences are in blue and putative GAO in magenta. Bold typeface indicates the PAO or GAO physiology is supported by in situ evidence. Sub-group and clade classifications for the “Ca. Accumulibacter spp.” (He et al., 2007; Kim et al., 2010, 2013), Competibacteraceae (Kong et al., 2002; Kim et al., 2011; McIlroy et al., 2015a), and Defluviicoccus spp. (Wong et al., 2004; Meyer et al., 2006; McIlroy and Seviour, 2009; Nittami et al., 2009) are taken from previous studies and are given in parenthesis. Brackets to the right indicate the phylogenetic classification of sequences. The tree was prepared using the ARB software (Ludwig et al., 2004) from the MiDAS database (version 2.1) (McIlroy et al., 2015b). Sequences were aligned in the ARB software, trimmed and variable regions excluded with a custom filter (filter by base frequency, 20–100%) leaving 1137 aligned positions. Sequences <1200 bp were added after calculation of the tree with the ARB insert sequences function (AY710272; KF663686). The scale bar represents substitutions per nucleotide base.
Results and Discussion
Overall Composition of the Microbial Communities
The overall microbial populations in the Danish WWTPs are visualized in a redundancy analysis (RDA) plot for the entire period from 2006 to 2014 (Figure 2A). Despite a large shared community of abundant genera among the plants (Saunders et al., 2016), clustering of samples from each system is evident and is largely due to differences in the relative abundance of these shared genera. Interestingly, this suggests that the community composition of each plant was relatively stable throughout the 9 years—making this study, to our knowledge, the first to document such long-term stability of full-scale WWTPs.
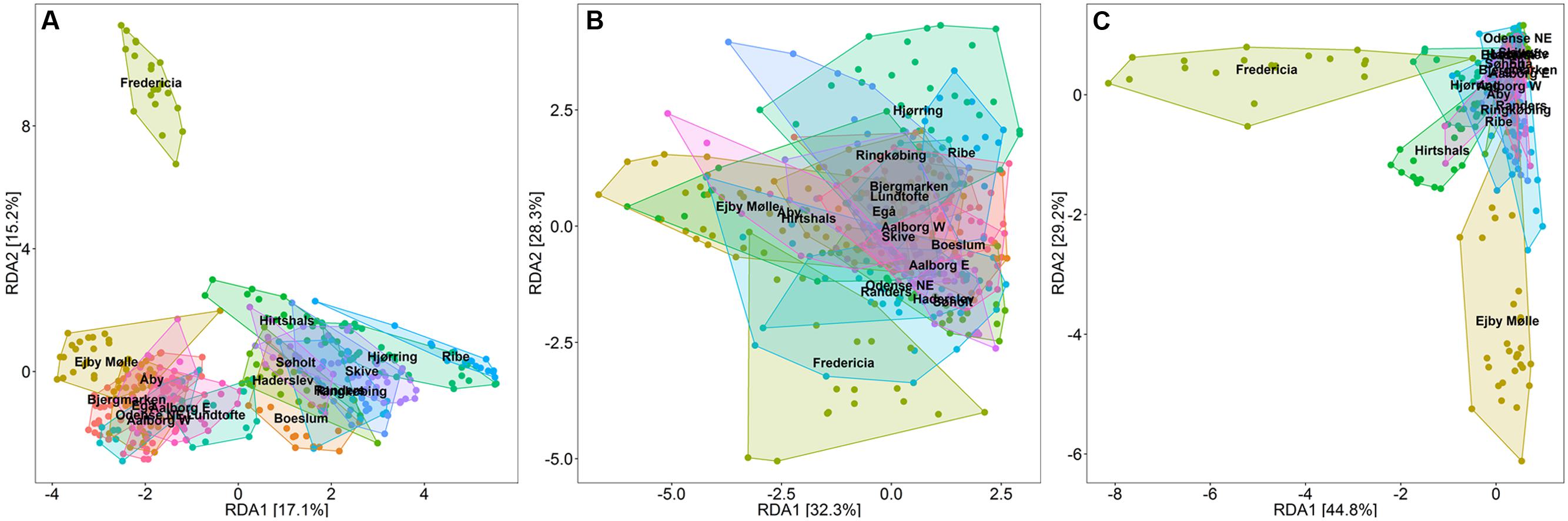
FIGURE 2. Redundancy analysis (RDA) plots, constrained to visualize the variance in OTU abundance, explained by sample plant origin. Axes show the percent of the total variation described by each principal component. (A) RDA plot of the overall bacterial community of 18 Danish full-scale EBPR WWTPs over a period of 9 years showing profiles of each plant. (B) RDA plot of the PAO part of the community showing similar community composition across the WWTPs, and (C) GAO part of the community showing dissimilarities among the WWTPs with several unique plant profiles.
Principal component analysis (PCA) analyses of the putative PAO and GAO populations in these plants indicate that these communities were relatively similar both within and between the plants (Figures 2B,C). Noted exceptions were plants receiving a high industrial load (>50%, e.g., Fredericia), primarily due to differences in the GAO populations. It is worth noting that despite relatively high levels of putative GAO in some plants (>20%), all plants have had stable P removal over several years, as previously reported (Mielczarek et al., 2013). PAO populations in general appeared to be more stably present (Figure 2B and Supplementary Material), where the intermittently high abundances of GAO were likely related to fluctuations in the influent COD: P ratio—where the GAO utilize excess COD (Tu and Schuler, 2013; Law et al., 2016). Though EBPR failure in full-scale systems has been related to high GAO numbers (Saunders et al., 2003), it is yet to be shown that it is directly the result of the GAO outcompeting the PAO for resources. Thus, more work is required to assess the competition hypothesis, which will importantly involve surveys that also cover inefficient and failed EBPR plants.
In the current study, population sizes of each of the proposed PAO and GAO genera varied greatly (Figures 3–6). Some were consistently observed at high abundances, i.e., the Tetrasphaera PAO, while for others high abundance was more transient, i.e., the Competibacteraceae GAO (Figures 3, 4). Time series of all PAOs and GAOs in the individual plants are shown in the Supplementary Material. Several putative PAO and GAO genera were rarely detected. In light of these calculated amplicon-based abundances, evidence for a potential role in EBPR for these genera is discussed.
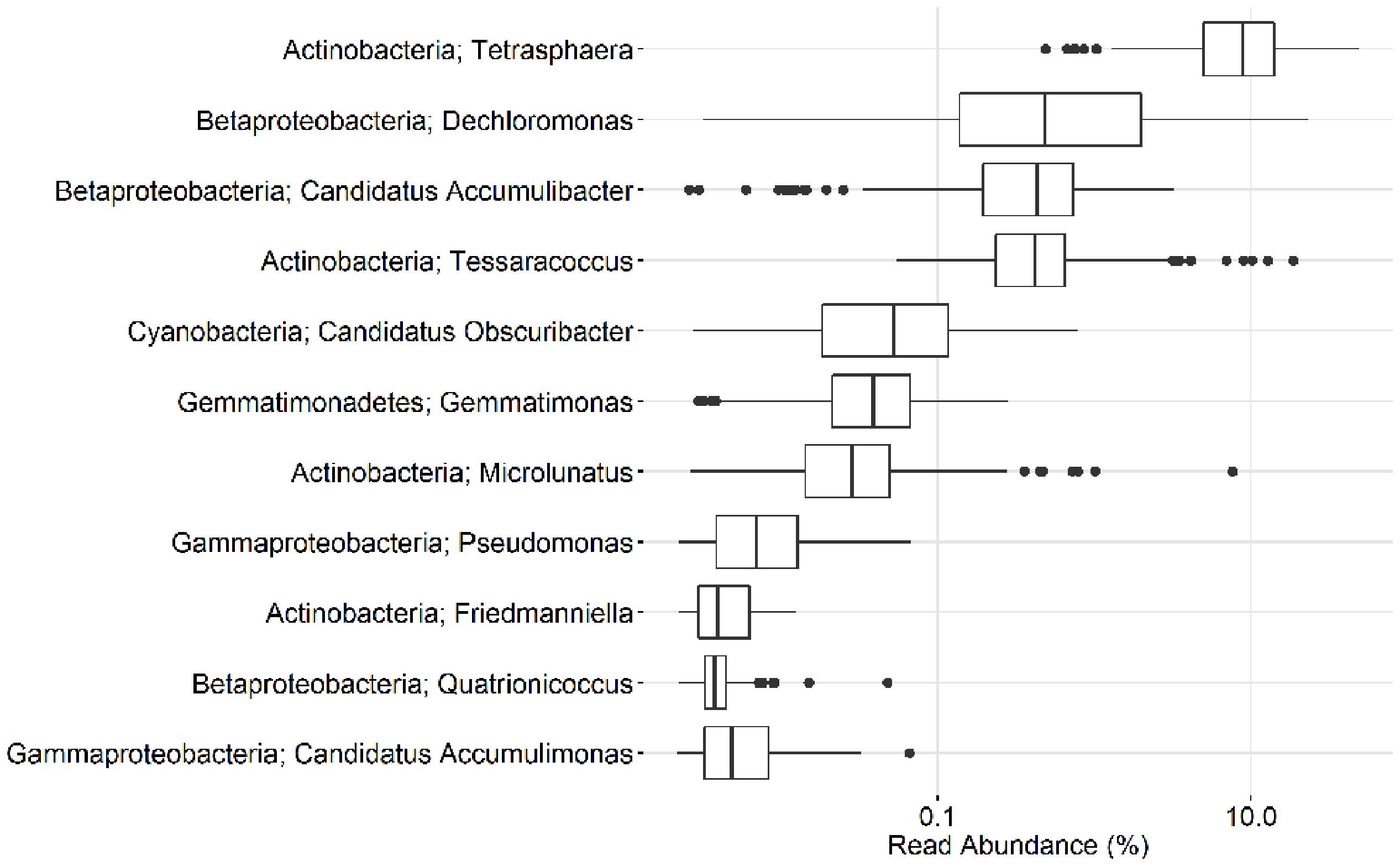
FIGURE 3. Overview of the abundances of putative PAO. Each box shows the data for all samples collected from 18 Danish WWTPs during the years 2006–2014. Malikia and Lampropedia were never detected. Phylum and genus are shown. Small constant (0.01) was added to all the abundance values in order to enable visualization of 0 values on the log scale.
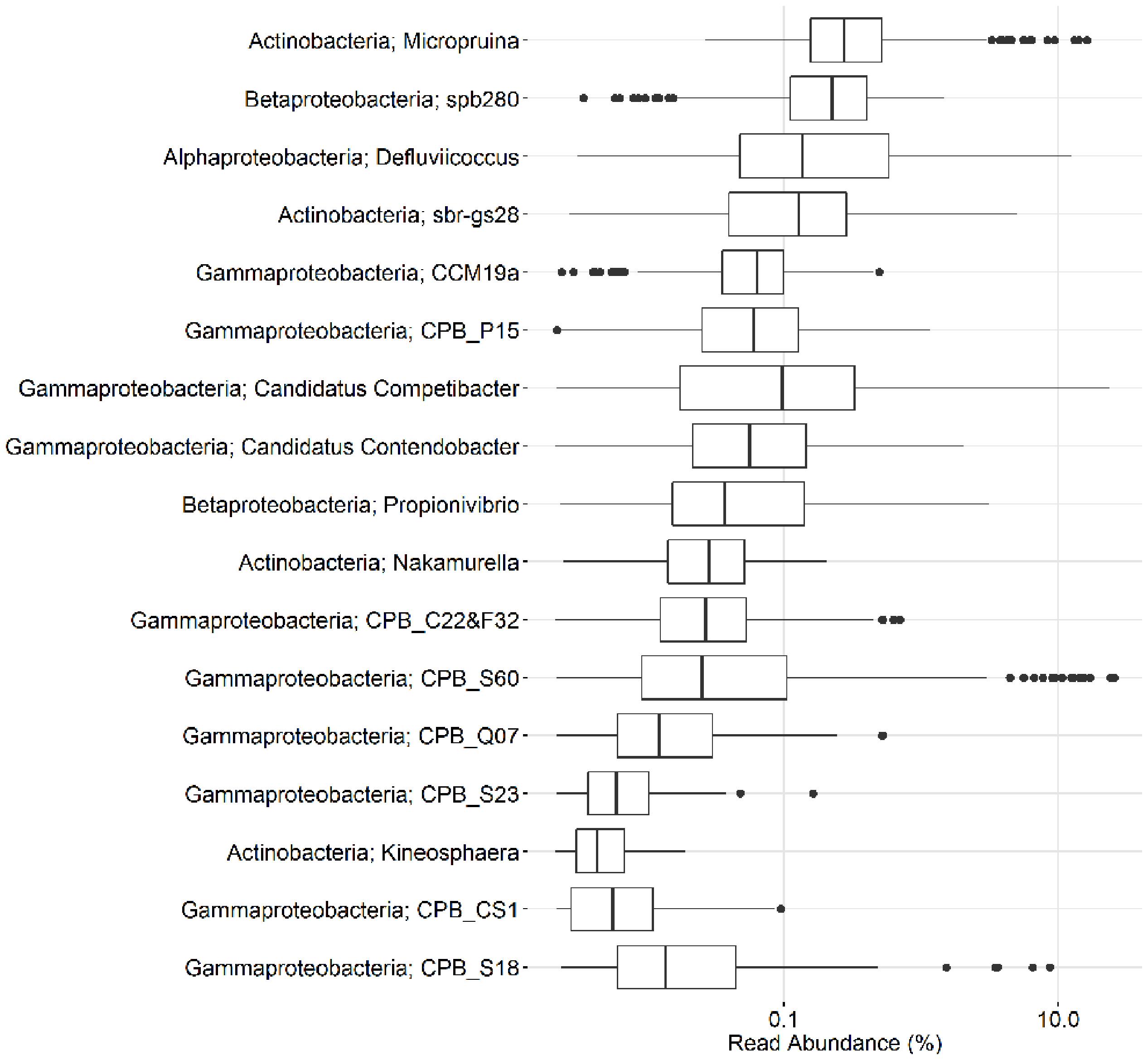
FIGURE 4. Overview of the abundances of putative GAO. Each box shows the data for all samples collected in 18 Danish WWTPs during the years 2006–2014. CPB_M38 is not included as it is not covered by the MiDAS taxonomy. Phylum and genus are shown. Small constant (0.01) was added to all the abundance values in order to enable visualization of 0 values on the log scale.
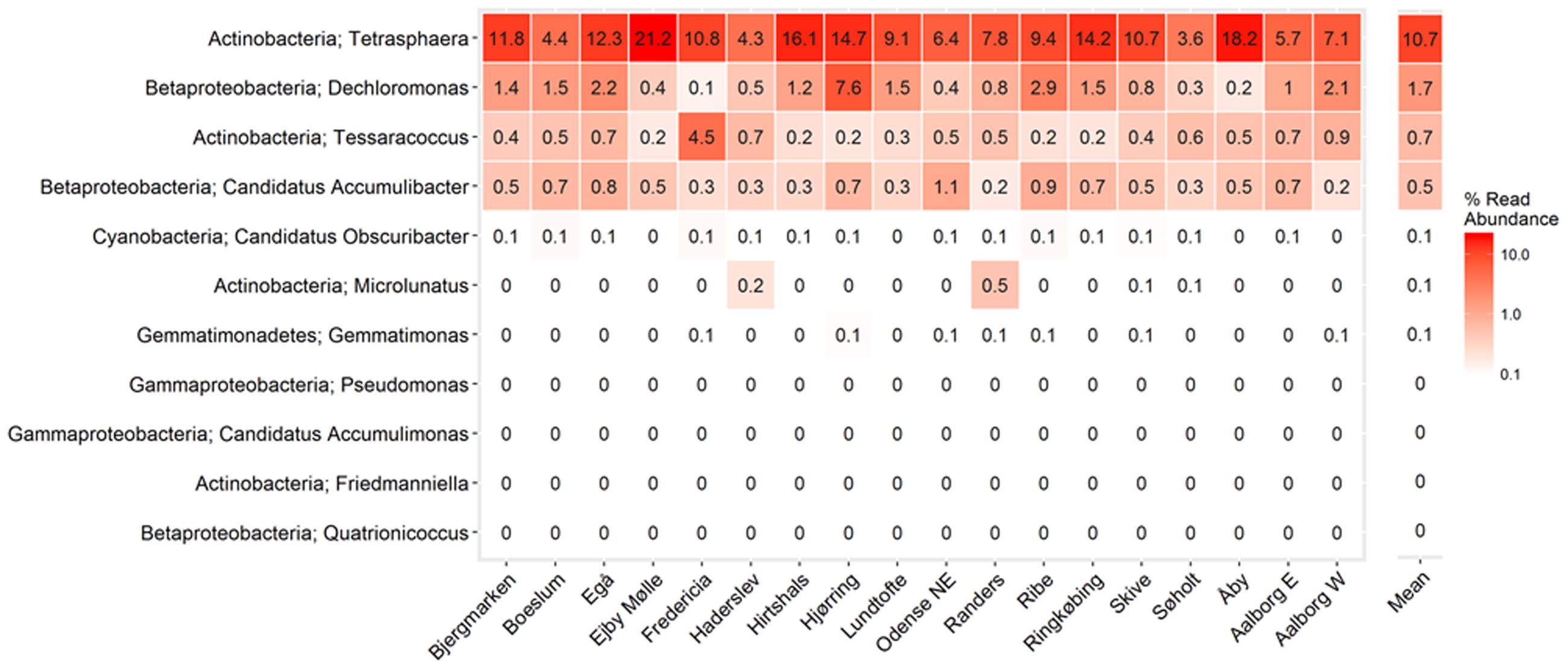
FIGURE 5. Heatmap showing mean read abundance of putative PAOs in percent of all amplicon reads. The numbers represent the average read abundance for given WWTP. The survey was performed in the years 2006–2014 with up to four samples per year for each plant. Phylum and genus are shown.
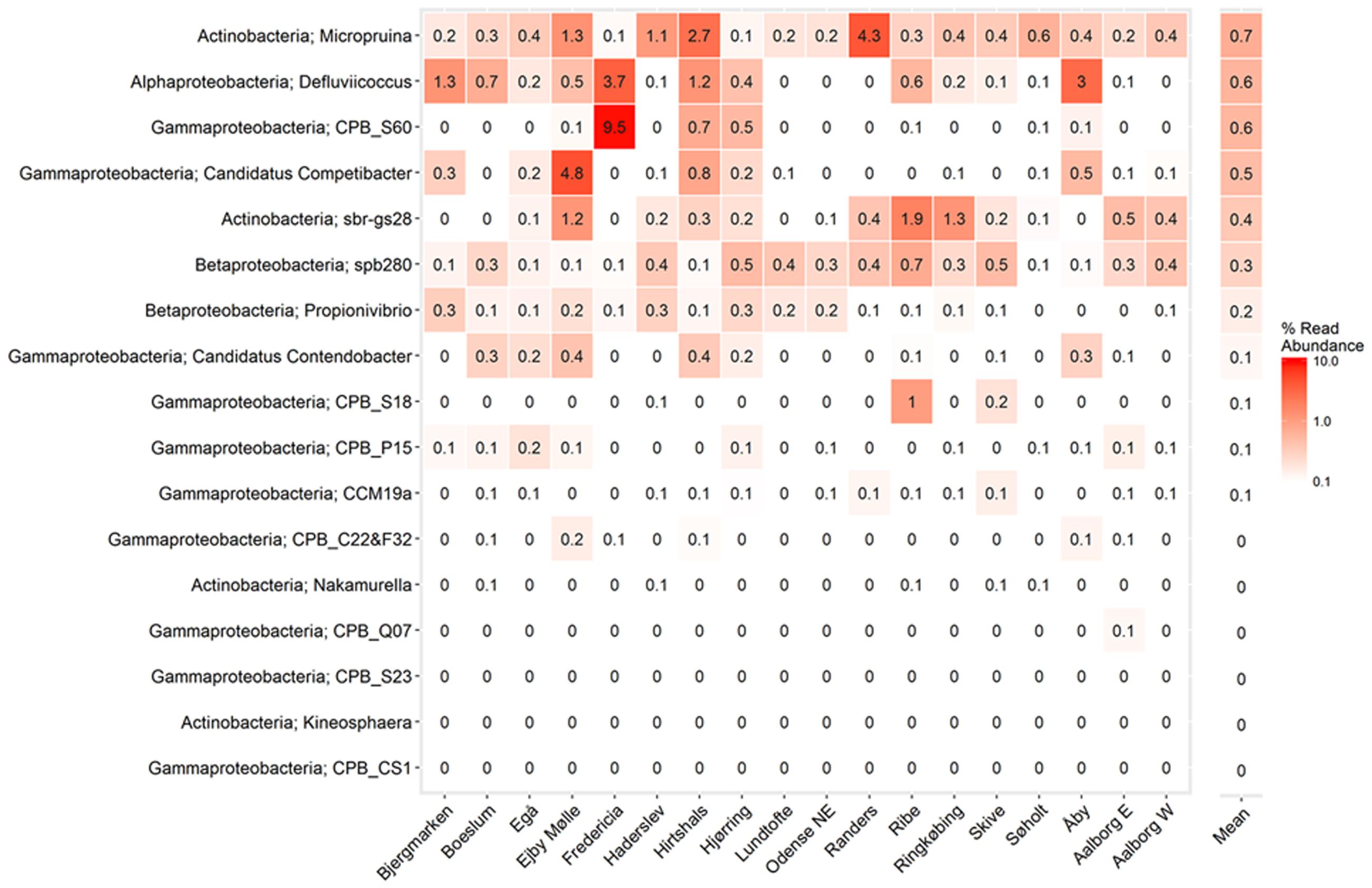
FIGURE 6. Heatmap showing mean read abundance of putative GAOs in percent of all amplicon reads. The numbers represent the average read abundance for the given WWTP. The survey was performed in the years 2006–2014 with up to four samples per year for each plant. Phylum and genus are shown.
The Putative PAOs
The Betaproteobacterial PAO
The uncultured “Ca. Accumulibacter” is the most studied of the PAO genera and is often assumed to be the most important PAO. The classical PAO phenotype has been demonstrated for members of the genus with both in situ methods (Kong et al., 2004) and in sequencing batch reactor (SBR) enrichments, where they can be enriched up to 96% of the biomass (Lu et al., 2006). For a detailed review of their physiology, see He and McMahon (2011). In this study, “Ca. Accumulibacter” had an average and median read abundance of 0.5 and 0.4%, respectively (Figures 3, 5). This is in contrast to previous FISH-based studies, including those assessing the same Danish plants, which recorded values of 3–8% (Mielczarek et al., 2013). A lack of probe specificity for the commonly applied PAOmix probe (see later) may contribute to this discrepancy (Albertsen et al., 2016). In addition, the “Ca. Accumulibacter” often possess relatively large cells (i.e., 2–3 μm (Kim et al., 2010), which results in relatively higher biovolume values in quantitative FISH (qFISH) analyses.
There were three MiDAS OTUs (>97% sequence similarity) classified to the “Ca. Accumulibacter” genus co-existing at similar abundances across the full-scale plants of this study. Several studies have investigated diversity of the genus (Martín et al., 2006; Flowers et al., 2013; Kim et al., 2013; Oyserman et al., 2015; Skennerton et al., 2015; Camejo et al., 2016). The resolution of the 16S rRNA gene to provide meaningful delineation of the genus is questionable and the polyphosphate kinase gene (ppk) has instead been suggested to provide this (McMahon et al., 2007). Two “Ca. Accumulibacter” types (I and II) are further delineated into 13 clades (IA-E and IIA-H) (He et al., 2007; Peterson et al., 2008; Mao et al., 2015). Clades reportedly vary in their morphology, carbon source, and denitrification genes (Flowers et al., 2009; Skennerton et al., 2015). Different “Ca. Accumulibacter” species have been suggested to vary in their level of reliance on polyphosphate storage and carbon uptake kinetics, where the predominance of a particular ppk-clade type may be associated with P removal efficiency (Slater et al., 2010; Welles et al., 2015, 2016). Whether or not important traits can be consistently associated with a ppk-type is yet to be demonstrated. Interestingly, several studies have shown that the “Ca. Accumulibacter” PAO can adopt the GAO phenotype under certain conditions, such as P limitation (Barat et al., 2006, 2008; Zhou et al., 2008; Welles et al., 2015, 2016), and the presence of “Ca. Accumulibacter” cells in full-scale EBPR plants not accumulating polyphosphate has also been reported widely (Zilles et al., 2002a,b; Kong et al., 2004; Wong et al., 2005; Beer et al., 2006). Such findings complicate the use of monitoring PAO phylotype abundances as a measure of EBPR health. Moreover, results of FISH studies should be assessed with caution, given the commonly applied FISH probe sets for the “Ca. Accumulibacter” (PAOmix) also target the Propionivibrio GAO that can be present in comparable abundances (Albertsen et al., 2016).
The second most abundant putative PAO genus in this study was Dechloromonas, with an average plant read abundance of 1.7%, median of 0.5%, and maximum of over 20% for some samples (Figures 3, 5). Contrary to the situation with the closely related “Ca. Accumulibacter,” Dechloromonas appears to be overestimated by amplicon sequencing, where values can be as much as 10-fold higher than with FISH-based quantification (McIlroy et al., 2016; Ziegler et al., 2016). A recent isolate of the genus, from a full-scale nutrient removal system, was shown to accumulate relatively high levels of polyphosphate in axenic culture, although no further characterization was reported (Terashima et al., 2016). Members where shown in situ to accumulate polyphosphate and PHA, and take up carbon under anaerobic conditions (Kong et al., 2007). While some members of the genus Dechloromonas have been shown to behave according to the PAO phenotype, others have been shown to potentially behave as their GAO competitors (Ahn et al., 2007; McIlroy et al., 2016). What proportion of the genus are behaving as PAO, and whether or not this can be confidently assigned to any of the three abundant OTUs of this study, is not known.
The Putative Gammaproteobacterial PAO
Members of the gammaproteobacterial class have also long been suggested as PAO in full-scale systems (Beer et al., 2006). Members of the genus Pseudomonas were identified as being among the most abundant PAO in an SBR EBPR system by applying a polyphosphate staining method with fluorescence activated cell sorting (FACS; Günther et al., 2009). The low abundance of the genus in the present study (Figure 3) is supported by previous FISH surveys of Danish systems and they are unlikely important PAO in full-scale plants (Nguyen et al., 2012).
Also within the class Gammaproteobacteria, the uncultured “Ca. Accumulimonas” store carbon as PHA under anaerobic conditions and polyphosphate under subsequent aerobic conditions, which is consistent with a PAO phenotype (Nguyen et al., 2012). However, they were not abundant in the MiDAS survey (Figures 3, 5), despite FISH surveys reporting their consistent abundance up to 6% of the biovolume (Nguyen et al., 2012). The V1-3 primer set applied in this study was shown to give relatively poor coverage of the Gammaproteobacteria (Albertsen et al., 2015). However, a previous survey of Danish WWTPs applying the V4 primer set also failed to detect the genus in abundance (Saunders et al., 2016)—despite the primer set covering the database sequences of the “Ca. Accumulimonas spp.” well. Further work is required to verify their abundance, and subsequent importance in EBPR systems.
The Actinobacterial PAO
Consistent with previous qFISH-based studies, Tetrasphaera had the highest abundance of the PAO, with an average and median read abundance of 10.7 and 8.9%, respectively, across all plants and present in excess of 40% of the biomass in some plants. Relatively good support for the Tetrasphaera spp. as PAOs has been demonstrated for members of the genus in pure culture and in situ with polyphosphate cycling coupled to anaerobic carbon uptake (Kong et al., 2005; Nguyen et al., 2011; Kristiansen et al., 2013). However, the Tetrasphaera genus has demonstrated diverse physiology, including an ability for fermentative growth, and to accumulate fermentation by-products and amino acids intracellularly (Nguyen et al., 2015). Although numerically the most abundant PAO, key questions need to be answered to establish the contribution members of the genus make to P removal, i.e., how much P is accumulated relative to the “Ca. Accumulibacter” and how varied this is across the abundant Tetrasphaera spp. Three FISH probe-defined clades have been characterized in situ revealing differences in morphology and carbon source uptake (Nguyen et al., 2011). However, analyses of these FISH probes against the current SILVA database suggest that they are unable to separate the different clades with reasonable specificity (data not shown). Although a single dominant OTU made up most of the genus across all plants, several OTUs classified to the genus Tetrasphaera (Figure 7). This is consistent with the observations of Saunders et al. (2016). Due to limitations in the resolution of the short 16S rRNA amplicons, these OTUs could not be related to the proposed three clades of Nguyen et al. (2011).
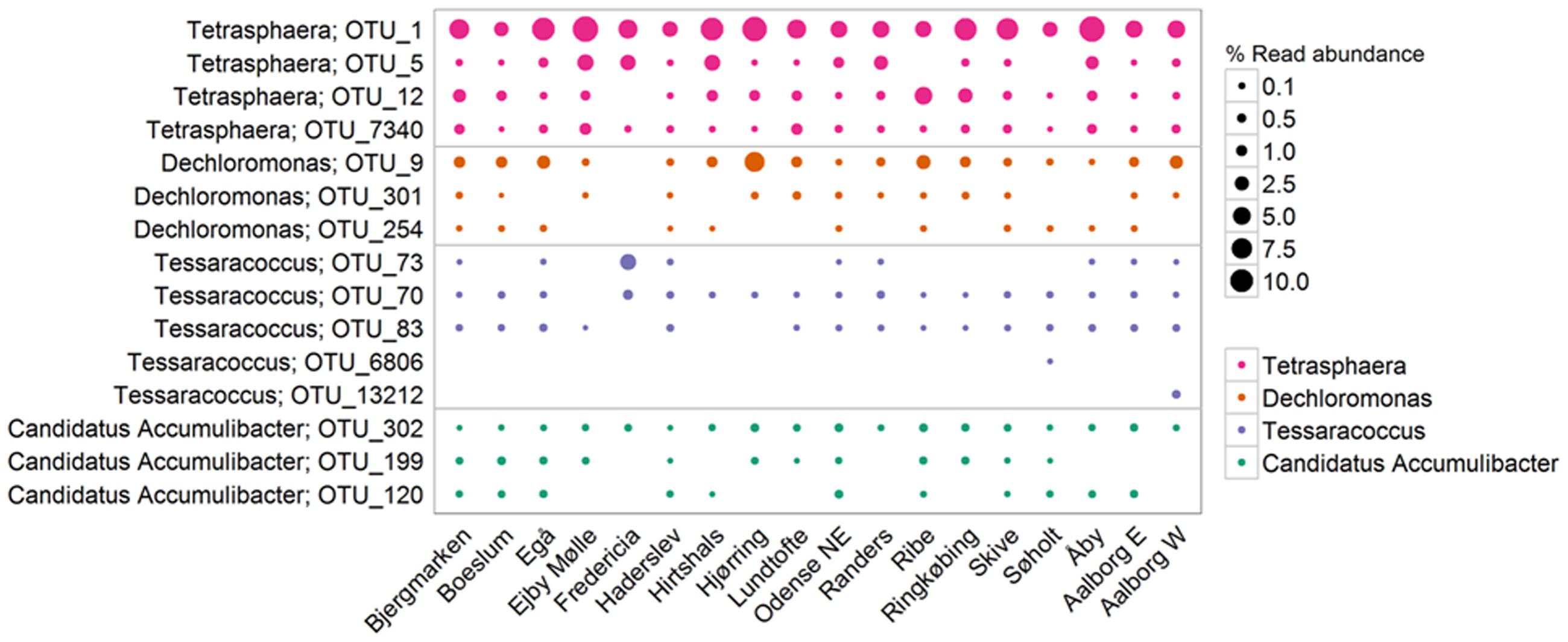
FIGURE 7. The most abundant OTUs within each PAO-related genus. Percentages shown represent the average read abundance of all samples collected in 18 Danish WWTPs during the years 2006–2014.
The genus Microlunatus had a median read abundance of only <0.1%, but was seen as high as 7% in some plants (Figure 3) and has also been detected at abundances up to 9% with qFISH (Beer et al., 2006). Microlunatus may thus be important for the EBPR process in some plants. Microlunatus phosphovorus was isolated from activated sludge and shown to accumulate large amounts of polyphosphate aerobically, and release it anaerobically with a concomitant uptake of glucose (Nakamura et al., 1991, 1995b) and glutamate (Ubakata and Takii, 1994, 1998). Furthermore, in situ strains stain positive for polyphosphate storage (Kawaharasaki et al., 1999; Beer et al., 2006). As with the Tetrasphaera, in pure culture the M. phosphovorus can ferment sugars, and accumulate fermentation products and amino acids anaerobically (Santos et al., 1999). Although some unidentified storage polymer was postulated, neither PHA nor glycogen cycling occurred and P and carbon metabolism were not exclusively coupled (Santos et al., 1999). The type strain reportedly lacks the genes for PHA synthesis (Kawakoshi et al., 2012), however, it has been reported to synthesize the storage polymer (Akar et al., 2006). The physiology of Microlunatus spp. is likely quite similar to that of the Tetrasphaera spp. but requires further investigation.
Members of the actinobacterial genus Tessaracoccus were found to be relatively abundant in some plants with a median read abundance of 0.4% and up to 19% of the reads (Figure 3). Evidence for their role in EBPR rests solely on the ability of T. bendigoensis to accumulate polyphosphate in pure culture (Maszenan et al., 1999b) and nothing is known of their in situ physiology. Isolated members of the genus are facultative anaerobes and thus likely have a fermentative metabolism in wastewater systems (Maszenan et al., 1999b; Finster et al., 2009). Their high abundance appears to be maintained by immigration from the influent wastewater, suggesting a low growth and activity rate (Saunders et al., 2016). It is therefore unknown whether or not they play any substantial role in EBPR systems.
It has also been suggested that members of the genus “Candidatus Microthrix” were responsible for the bulk of P removal in a lab-scale EBPR system, which was based on positive staining for intracellular polyphosphate stores and the observed low abundance of “Ca. Accumulibacter” and Tetrasphaera spp. (Wang J. et al., 2014). Polyphosphate accumulation has been demonstrated for pure culture and in situ species, although it is generally thought to be a response to cell stress (Erhart et al., 1997; Tandoi et al., 1998b) and does not appear to be linked to anaerobic: aerobic carbon cycling (Andreasen and Nielsen, 2000). The genome of “Ca. Microthrix parvicella RN1” also lacks the low-affinity inorganic phosphate transporter (Pit) (McIlroy et al., 2013), which is theoretically essential to the PAO phenotype (Saunders et al., 2007; McIlroy et al., 2014). Its contribution to P removal is therefore uncertain; noting that regardless of any impact it may have on P removal, its presence in the system is undesirable given its proliferation is linked to the serious operational problems of bulking and foaming (Rossetti et al., 2005).
Other Putative PAO Isolates
Members of the genera representing putative PAO isolates, Gemmatimonas, Friedmaniella, Quatrionicoccus, Malikia, and Lampropedia were rarely detected in the Danish plants (Figures 3, 5) and are unlikely numerically important to the EBPR process. It should also be noted that, with the exception of L. hyalina (Stante et al., 1997), the only evidence suggesting their role as PAO is an ability to store visible polyphosphate granules.
A recent study hinted at the involvement of the uncultured cyanobacteria “Candidatus Obscuribacter phosphatis” in EBPR, based on the annotation of its genome (Soo et al., 2014). The genome possesses the low-affinity inorganic phosphate transporter (Pit), which is not in itself evidence for the PAO phenotype (McIlroy et al., 2014), given that it is also found in non-PAO organisms (e.g., Defluviicoccus GAO; Wang Z. et al., 2014). It is generally difficult to identify putative PAOs or GAOs based on their genome, given that the storage of polyphosphate, glycogen, and PHA are widely phylogenetically dispersed traits (Oyserman et al., 2015). In situ work or gene expression studies are required to assess the activities of this genus in EBPR, noting that it is also not numerically important in full-scale EBPR systems in Denmark (Figures 3, 5).
Distribution of the Putative GAOs
The Gammaproteobacterial GAO
What was for many years considered as the genus “Candidatus Competibacter” has recently been suggested to be the family Competibacteraceae to better reflect the phylogenetic diversity of the group (with >89% 16S rRNA gene sequence similarity between members) (McIlroy et al., 2015a). The family is delineated by 14 FISH probe-defined clades and includes the genera “Ca. Competibacter,” “Candidatus Contendobacter,” and Plasticicumulans. The Plasticicumulans do not show the GAO phenotype and are rarely detected in full-scale plants with amplicon or qFISH analyses (McIlroy et al., 2015a), although a related phylotype was suggested to outcompete the “Ca. Accumulibacter” PAO in a novel EBPR system with continuous aeration (Schroeder et al., 2009). The most abundant of the groups in this study were the “Ca. Competibacter” and CPB_S60, which is consistent with qFISH-based studies (McIlroy et al., 2015a). Most of the genera had median abundances of less than 0.1%, yet periodically reached relatively high abundances of up to 25% in some plants (Figures 4, 6)—mainly those receiving high industrial loads (>50%) (i.e., Fredericia and Ejby Mølle; Table 1). The CPB_C22&F32, CPB_CS1, CPB_Q07, and CPB_S23 clades were never observed in read abundances greater than 1% and are unlikely important in Danish EBPR plants. The CPB_M38 clade was not assessed in this study as it is not covered by the MiDAS taxonomy given that it is represented by a single 16S rRNA gene sequence.
The Competibacteraceae are most frequently studied with family-level FISH probes and the classical GAO physiology has been confirmed with enrichment cultures (Lemos et al., 2007; Saunders et al., 2007; Oehmen et al., 2010) and in situ (Kong et al., 2006). Anaerobic carbon storage of VFAs as PHA in the absence of P cycling, consistent with the GAO phenotype, has also been demonstrated in situ for the “Ca. Competibacter” (formerly GB1), “Ca. Contendobacter” (GB5-7), CPB_S18 (GB2-3), CPB_S60 (GB4), and CPB_C22&F32 (GB8) phylotypes (Kong et al., 2002, 2006; Kim et al., 2011). All appear to have an affinity for VFAs with known differences between the phylotypes including carbon substrates type, fermentation, and denitrification (Kong et al., 2006; McIlroy et al., 2014).
Also within the Gammaproteobacteria are members of the uncultured CCM19a phylotype. These organisms were shown to store carbon as PHA under anaerobic conditions in the absence of polyphosphate storage, which is consistent with the GAO phenotype (Kong et al., 2007). In this study, they were rarely detected, with an average abundance of <0.1% and never in excess of 0.5% in any sample (Figure 4). Similarly, low abundances were also found with qFISH surveys, where they were up to 4% but typically <1% (Kong et al., 2007).
The Alphaproteobacterial GAO
In the current study, the genus Defluviicoccus had a median abundance of only 0.1%, but was observed at up to 13% of the amplicon reads (Figure 4). Four probe-defined sub-groups currently delineate the Defluviicoccus genus (Wong et al., 2004; Wong and Liu, 2007; McIlroy and Seviour, 2009; Nittami et al., 2009). Interestingly, clusters 1, 2, and 4 have a TFO morphology, while cluster 3 is filamentous and has been implicated in sludge settleability problems known as bulking (Nittami et al., 2009). Clusters 1 and 2 are the most studied and have been shown to conform to the GAO model with enrichment cultures (Dai et al., 2007; Lemos et al., 2007; Burow et al., 2008, 2009; Oehmen et al., 2010) and in situ (Wong et al., 2004; Meyer et al., 2006; Burow et al., 2007; Wong and Liu, 2007). GAO phenotype has also been demonstrated for the sole isolate, D. vanus, a member of cluster 1 (Maszenan et al., 2005; Wong and Liu, 2007). Anaerobic carbon storage as PHA has also been shown in situ for clusters 3 and 4 (McIlroy et al., 2010). In this study, OTUs representing clusters 2 and 3 made up the majority (>90%) of amplicon reads assigned to the genus, which is consistent with FISH analyses of full-scale systems (Figure 8; Burow et al., 2007; McIlroy and Seviour, 2009). The original report of an alphaproteobacterial Sphingomonas-related GAO (Beer et al., 2004) was later shown to be incorrect and the FISH probes were shown to be binding to members of the Defluviicoccus cluster 1 (McIlroy et al., 2011).
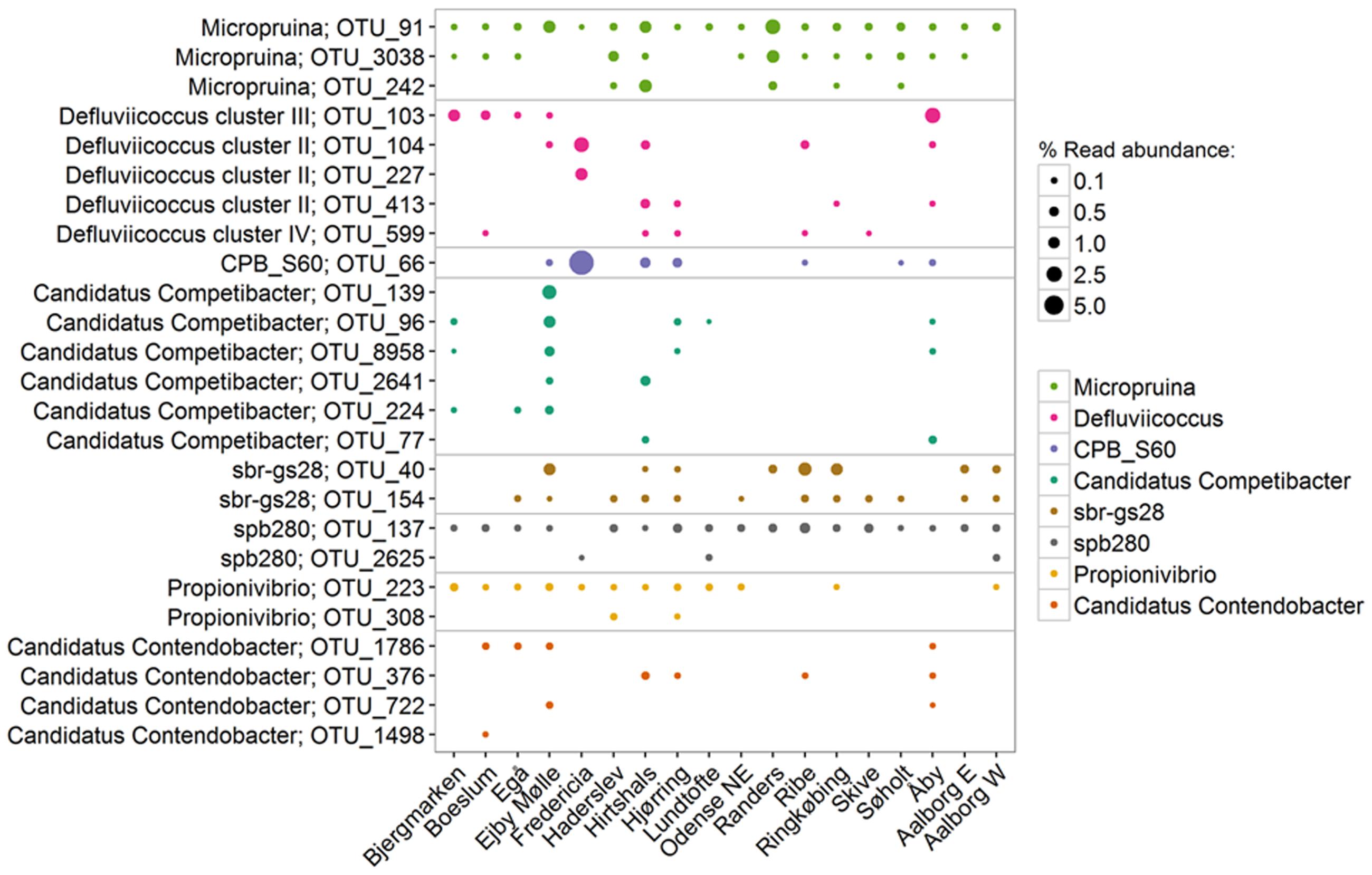
FIGURE 8. Most abundant OTUs within each GAO-related genus. Percentages shown represent the average read abundance of all samples collected in 18 Danish WWTPs during the years 2006–2014.
The Betaproteobacterial GAO
The newly discovered Propionivibrio GAO is closely related to the “Ca. Accumulibacter” and is covered by the PAOmix probe set (Crocetti et al., 2000) commonly applied to target the former (Albertsen et al., 2016). In a lab-scale SBR fed with VFAs as the sole carbon source, they reached 35% of the biovolume and were shown to cycle PHA without polyphosphate storage, consistent with a GAO phenotype. In the current study, the Propionivibrio GAO was detected in a median read abundance of <0.1% and was present in most plants reaching abundances of up to 3% (Figure 4). Amplicon-based assessment of their abundance in full-scale systems has been shown to be comparable to qFISH (Albertsen et al., 2016).
The uncultured betaproteobacterial spb280, similar to the CCM19a phylotype, was also identified in full-scale systems at qFISH abundances up to 6% and the GAO phenotype partially shown in situ (Kong et al., 2007). The phylotype was consistently present with the second highest median abundance and maximum amplicon read abundances of 1.5% (Figure 4). Like the CCM19a phylotype, nothing more has been determined for the phylotype since its initial description.
The Actinobacterial GAO
A few isolates of the phylum Actinobacteria have been reported as potential competitors of the PAO. These include N. multipartita (Yoshimi et al., 1996), M. glycogenica (Shintani et al., 2000), and K. limosa (Liu et al., 2000, 2002), all shown to accumulate carbohydrates anaerobically without any polyphosphate storage. Of the genera these represent, only the Micropruina, along with the closely related uncultured sbr-gs28 phylotype, have been shown in situ to store both acetate and glucose as PHA without polyphosphate storage, consistent with the GAO phenotype (Kong et al., 2001).
In the current study, only the Micropruina and sbr-gs28, both members of the family Propionibacteriaceae, were found in Danish WWTPs. These had median abundances of 0.3 and 0.1%, respectively, but were present up to 16 and 5% of amplicon reads, respectively (Figure 4). Interestingly, the Micropruina were the most abundant of all putative GAOs considered in this study, based on both median and mean values (Figures 4, 6). Their high abundance in full-scale systems was also supported by qFISH surveys in Japan where they constituted up to 8% of the biovolume (Wong et al., 2005). Despite their relatively high abundances, little is known of their in situ physiology. As such, investigating their role in EBPR systems should therefore be a priority and is currently underway (McIlroy et al., unpublished).
Implications of Metabolic Diversity among the PAO and GAO
The ability of the GAO to compete with the PAO for resources is central to their perceived importance in EBPR. As such, it is worth noting that the putative co-existing PAOs and GAOs discussed here are metabolically diverse and will not all compete for the same niche—with prominent differences related to carbon source type taken up and their ability to denitrify and/or ferment. For example, the Defluviicocus GAO will likely utilize the by-products of the fermentative Tetrasphaera PAO, rather than compete for resources. More work is required to determine the nature and importance of competition across the niches in which it occurs.
The genus is often assumed to represent a metabolically coherent group, however, several studies have shown that substantial functional diversity also exist among members of the same genus (see earlier), which can have implications for the way the microbial community composition is interpreted. The co-existence of often several closely related species of PAOs and GAOs is possibly maintained by metabolic differences supporting niche differentiation. In this study, such diversity is evident, with the abundance of each genus being made up of multiple OTUs. The level of diversity for each genus varies. For example, the “Ca. Accumulibacter” had three OTUs that where present in most plants at similar abundances where the genus Tetrasphaera had a single OTU dominating across all plants (Figure 7). Whether or not important traits can be associated with such 16S rRNA gene-defined OTUs in a genus is unclear and needs to be investigated further. It appears possible to identify the filamentous Defluviicoccus cluster 3, but not for the “Ca. Accumulibacter”—where the ppk-gene is instead suggested for phylogenetic resolution (McMahon et al., 2007). This becomes particularly important when utilizing total PAO abundances to assess EBPR health—where it suggested that some members of the genus might instead be behaving according to the GAO phenotype or fermenting without polyphosphate storage.
Conclusion
The abundance and diversity of all proposed PAOs and GAOs were assessed in 18 Danish full-scale wastewater treatment plants with EBPR over a period of 9 years using 16S rRNA gene amplicon sequencing. The plants seem to exhibit relatively high temporal stability in the overall microbial community composition over the 9-year period. Members of the genera Tetrasphaera, “Ca. Accumulibacter”, and Dechloromonas are confirmed to represent the important known PAOs in full-scale EBPR systems, although abundances of the latter two appear to be heavily biased with amplicon-based methods. Potential roles for the Microlunatus, “Ca. Accumulimonas”, and Tessaracoccus are currently unclear and need further work. All other putative PAO considered in this study appear to be numerically unimportant in EBPR systems. For the GAOs, substantial evidence is available for the physiology of the Defluviicoccus and Competibacteraceae GAO only. Future work should focus on the characterization of the actinobacterial Micropruina, sbr-gs28, betaproteobacterial spb280, and the Propionivibrio phylotypes. Other phylotypes were never detected in abundance and thus are unlikely relevant in EBPR. Importantly, in this study, we only consider “known” PAOs and GAOs, which have largely been identified in lab-scale systems fed acetate or through isolation of cultures accumulating phosphate and/or carbon. Large-scale whole community surveys like MiDAS, followed by the systematic characterization of the in situ physiology of all abundant phylotypes in the complex full-scale EBPR systems, will likely uncover novel abundant PAOs and GAOs in the future. Phylogenetic diversity within each of the PAOs and GAOs genera varied, which is consistent with reported metabolic diversity for those tested. Whether or not key traits can be assigned to sub-genus level clades requires further investigation.
Author Contributions
PN planned sample collection; MA, SK, and PN planned the experimental work; MA, SK, and MN performed the experimental work; MS-B, MN, MA, and SM performed the data analyses; SM, MS-B, and PN drafted the manuscript. All authors were involved in revising the manuscript.
Funding
The project was supported by the Danish Research Council for Strategic Research (EcoDesign, grant no. 09-067230) and the Poul Due Jensen (Grundfos) Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We want to acknowledge the many plant operators at the Danish WWTPs for providing samples.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00718/full#supplementary-material
Footnotes
- ^ http://midasfieldguide.org
- ^ www.midasfieldguide.org/en/protocols
- ^ http://www.rstudio.com/
- ^ https://github.com/MadsAlbertsen/ampvis
- ^ www.midasfieldguide.org
References
Ahn, J., Schroeder, S., Beer, M., McIlroy, S., Bayly, R. C., May, J. W., et al. (2007). Ecology of the microbial community removing phosphate from wastewater under continuously aerobic conditions in a sequencing batch reactor. Appl. Environ. Microbiol. 73, 2257–2270. doi: 10.1128/AEM.02080-06
Akar, A., Akkaya, E. U., Yesiladali, S. K., Çelikyilmaz, G., Çokgor, E. U., Tamerler, C., et al. (2006). Accumulation of polyhydroxyalkanoates by Microlunatus phosphovorus under various growth conditions. J. Ind. Microbiol. Biotechnol. 33, 215–220. doi: 10.1007/s10295-004-0201-2
Albertsen, M., Karst, S. M., Ziegler, A. S., Kirkegaard, R. H., and Nielsen, P. H. (2015). Back to basics–the influence of DNA extraction and primer choice on phylogenetic analysis in activated sludge communities. PLoS ONE 10:e0132783. doi: 10.1371/journal.pone.0132783
Albertsen, M., McIlroy, S. J., Stokholm-Bjerregaard, M., Karst, S. M., and Nielsen, P. H. (2016). “Candidatus Propionivibrio aalborgensis”: a novel glycogen accumulating organism abundant in full-scale enhanced biological phosphorus removal plants. Front. Microbiol. 7:1033. doi: 10.3389/fmicb.2016.01033
Andreasen, K., and Nielsen, P. H. (2000). Growth of Microthrix parvicella in nutrient removal activated sludge plants: studies of in situ physiology. Water Res. 34, 1559–1569.
Barat, R., Montoya, T., Borrás, L., Ferrer, J., and Seco, A. (2008). Interactions between calcium precipitation and the polyphosphate accumulating bacteria metabolism. Water Res. 42, 3415–3424. doi: 10.1016/j.watres.2008.05.003
Barat, R., Montoya, T., Borrás, L., Seco, A., and Ferrer, J. (2006). Calcium effect on enhanced biological phosphorus removal. Water Sci. Technol. 53, 29–37. doi: 10.2166/wst.2006.403
Beer, M., Kong, Y. H., and Seviour, R. J. (2004). Are some putative glycogen accumulating organisms (GAO) in anaerobic:aerobic activated sludge systems members of the α-Proteobacteria? Microbiology 150, 2267–2275. doi: 10.1099/mic.0.26825-0
Beer, M., Stratton, H. M., Griffiths, P. C., and Seviour, R. J. (2006). Which are the polyphosphate accumulating organisms in full-scale activated sludge enhanced biological phosphate removal systems in Australia? J. Appl. Microbiol. 100, 223–243. doi: 10.1111/j.1365-2672.2005.02784.x
Burow, L. C., Kong, Y., Nielsen, J. L., Blackall, L. L., and Nielsen, P. H. (2007). Abundance and ecophysiology of Defluviicoccus spp., glycogen-accumulating organisms in full-scale wastewater treatment processes. Microbiology 153, 178–185. doi: 10.1099/mic.0.2006/001032-0
Burow, L. C., Mabbett, A. N., Borras, L., and Blackall, L. L. (2009). Anaerobic central metabolic pathways active during polyhydroxyalkanoate production in uncultured cluster 1 Defluviicoccus enriched in activated sludge communities. FEMS Microbiol. Lett. 298, 79–84. doi: 10.1111/j.1574-6968.2009.01695.x
Burow, L. C., Mabbett, A. N., McEwan, A. G., Bond, P. L., and Blackall, L. L. (2008). Bioenergetic models for acetate and phosphate transport in bacteria important in enhanced biological phosphorus removal. Environ. Microbiol. 10, 87–98. doi: 10.1111/j.1462-2920.2007.01432.x
Camejo, P. Y., Owen, B. R., Martirano, J., Ma, J., Kapoor, V., Santo Domingo, J., et al. (2016). Candidatus Accumulibacter phosphatis clades enriched under cyclic anaerobic and microaerobic conditions simultaneously use different electron acceptors. Water Res. 102, 125–137. doi: 10.1016/j.watres.2016.06.033
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 6, 1621–1624. doi: 10.1038/ismej.2012.8
Cech, J. S., and Hartman, P. (1993). Competition between polyphosphate and polysaccharide accumulating bacteria in enhanced biological phosphate removal systems. Water Res. 27, 1219–1225. doi: 10.1016/0043-1354(93)90014-9
Chen, T., Yu, W.-H., Izard, J., Baranova, O. V., Lakshmanan, A., and Dewhirst, F. E. (2010). The human oral microbiome database: a web accessible resource for investigating oral microbe taxonomic and genomic information. Database (Oxford). 2010:baq013. doi: 10.1093/database/baq013
Comeau, Y., Hall, K. J., Hancock, R. E. W., and Oldham, W. K. (1986). Biochemical model for enhanced biological phosphorus removal. Water Res. 20, 1511–1521. doi: 10.1016/0043-1354(86)90115-6
Crocetti, G. R., Banfield, J. F., Keller, J., Bond, P. L., and Blackall, L. L. (2002). Glycogen-accumulating organisms in laboratory-scale and full-scale wastewater treatment processes. Microbiology 148, 3353–3364. doi: 10.1099/00221287-148-11-3353
Crocetti, G. R., Hugenholtz, P., Bond, P. L., Schuler, A., Keller, J., Jenkins, D., et al. (2000). Identification of polyphosphate-accumulating organisms and design of 16S rRNA-directed probes for their detection and quantitation. Appl. Environ. Microbiol. 66, 1175–1182. doi: 10.1128/AEM.66.3.1175-1182.2000
Dai, Y., Yuan, Z., Wang, X., Oehmen, A., and Keller, J. (2007). Anaerobic metabolism of Defluviicoccus vanus related glycogen accumulating organisms (GAOs) with acetate and propionate as carbon sources. Water Res. 41, 1885–1896. doi: 10.1016/j.watres.2007.01.045
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Erhart, R., Bradford, D., Seviour, R. J., Amann, R., and Blackall, L. L. (1997). Development and use of fluorescent in situ hybridization probes for the detection and identification of Microthrix parvicella in activated sludge. Syst. Appl. Microbiol. 20, 310–318. doi: 10.1016/S0723-2020(97)80078-1
Falvo, A., Levantesi, C., Rossetti, S., Seviour, R. J., and Tandoi, V. (2001). Synthesis of intracellular storage polymers by Amaricoccus kaplicensis, a tetrad forming bacterium present in activated sludge. J. Appl. Microbiol. 91, 299–305. doi: 10.1046/j.1365-2672.2001.01384.x
Finster, K. W., Cockell, C. S., Voytek, M. A., Gronstal, A. L., and Kjeldsen, K. U. (2009). Description of Tessaracoccus profundi sp. nov., a deep-subsurface actinobacterium isolated from a Chesapeake impact crater drill core (940 m depth). Antonie Van Leeuwenhoek 96, 515–526. doi: 10.1007/s10482-009-9367-y
Flowers, J. J., He, S., Malfatti, S., del Rio, T. G., Tringe, S. G., Hugenholtz, P., et al. (2013). Comparative genomics of two “Candidatus Accumulibacter” clades performing biological phosphorus removal. ISME J. 7, 2301–2314. doi: 10.1038/ismej.2013.117
Flowers, J. J., He, S., Yilmaz, S., Noguera, D. R., and McMahon, K. D. (2009). Denitrification capabilities of two biological phosphorus removal sludges dominated by different “Candidatus Accumulibacter” clades. Environ. Microbiol. Rep. 1, 583–588. doi: 10.1111/j.1758-2229.2009.00090.x
Gilbert, N. (2009). Environment: the disappearing nutrient. Nature 461, 716–718. doi: 10.1038/461716a
Günther, S., Trutnau, M., Kleinsteuber, S., Hause, G., Bley, T., Röske, I., et al. (2009). Dynamics of polyphosphate-accumulating bacteria in wastewater treatment plant microbial communities detected via DAPI (4’,6’-diamidino-2-phenylindole) and tetracycline labeling. Appl. Environ. Microbiol. 75, 2111–2121. doi: 10.1128/AEM.01540-08
He, S., Gall, D. L., and McMahon, K. D. (2007). “Candidatus Accumulibacter” population structure in enhanced biological phosphorus removal sludges as revealed by polyphosphate kinase genes. Appl. Environ. Microbiol. 73, 5865–5874. doi: 10.1128/AEM.01207-07
He, S., and McMahon, K. D. (2011). Microbiology of “Candidatus Accumulibacter” in activated sludge. Microb. Biotechnol. 4, 603–619. doi: 10.1111/j.1751-7915.2011.00248.x
Hesselmann, R. P., Werlen, C., Hahn, D., van der Meer, J. R., and Zehnder, A. J. (1999). Enrichment, phylogenetic analysis and detection of a bacterium that performs enhanced biological phosphate removal in activated sludge. Syst. Appl. Microbiol. 22, 454–465. doi: 10.1016/S0723-2020(99)80055-1
Huang, W. E., Stoecker, K., Griffiths, R., Newbold, L., Daims, H., Whiteley, A. S., et al. (2007). Raman-FISH: combining stable-isotope Raman spectroscopy and fluorescence in situ hybridization for the single cell analysis of identity and function. Environ. Microbiol. 9, 1878–1889. doi: 10.1111/j.1462-2920.2007.01352.x
Jeanmaire, N., and Evans, T. (2001). Technico-economic feasibility of P-recovery from municipal wastewaters. Environ. Technol. 22, 1355–1361.
Jenkins, D., and Tandoi, V. (1991). The applied microbiology of enhanced biological phosphate removal–accomplishments and needs. Water Res. 25, 1471–1478. doi: 10.1016/0043-1354(91)90177-R
Kawaharasaki, M., Tanaka, H., Kanagawa, T., and Nakamura, K. (1999). In situ identification of polyphosphate-accumulating bacteria in activated sludge by dual staining with rRNA-targeted oligonucleotide probes and 4’,6-diamidino-2-phenylindol (DAPI) at a polyphosphate-probing concentration. Water Res. 33, 257–265.
Kawakoshi, A., Nakazawa, H., Fukada, J., Sasagawa, M., Katano, Y., Nakamura, S., et al. (2012). Deciphering the genome of polyphosphate accumulating actinobacterium Microlunatus phosphovorus. DNA Res. 19, 383–394. doi: 10.1093/dnares/dss020
Kim, J. M., Lee, H. J., Kim, S. Y., Song, J. J., Park, W., and Jeon, C. O. (2010). Analysis of the fine-scale population structure of “Candidatus Accumulibacter phosphatis” in enhanced biological phosphorus removal sludge, using fluorescence in situ hybridization and flow cytometric sorting. Appl. Environ. Microbiol. 76, 3825–3835. doi: 10.1128/AEM.00260-10
Kim, J. M., Lee, H. J., Lee, D. S., and Jeon, C. O. (2013). Characterization of the denitrification-associated phosphorus uptake properties of “Candidatus Accumulibacter phosphatis” clades in sludge subjected to enhanced biological phosphorus removal. Appl. Environ. Microbiol. 79, 1969–1979. doi: 10.1128/AEM.03464-12
Kim, J. M., Lee, H. J., Lee, D. S., Lee, K., and Jeon, C. O. (2011). Identification of a novel subgroup of uncultured gammaproteobacterial glycogen-accumulating organisms in enhanced biological phosphorus removal sludge. Microbiology 157, 2287–2296. doi: 10.1099/mic.0.047779-0
Kong, Y., Nielsen, J. L., and Nielsen, P. H. (2004). Microautoradiographic study of Rhodocyclus-related polyphosphate-accumulating bacteria in full-scale enhanced biological phosphorus removal plants. Appl. Environ. Microbiol. 70, 5383–5390. doi: 10.1128/AEM.70.9.5383-5390.2004
Kong, Y., Nielsen, J. L., and Nielsen, P. H. (2005). Identity and ecophysiology of uncultured actinobacterial polyphosphate-accumulating organisms in full-scale enhanced biological phosphorus removal plants. Appl. Environ. Microbiol. 71, 4076–4085. doi: 10.1128/AEM.71.7.4076-4085.2005
Kong, Y., Ong, S. L., Ng, W. J., and Liu, W.-T. (2002). Diversity and distribution of a deeply branched novel proteobacterial group found in anaerobic-aerobic activated sludge processes. Environ. Microbiol. 4, 753–757. doi: 10.1046/j.1462-2920.2002.00357.x
Kong, Y., Xia, Y., Nielsen, J. L., and Nielsen, P. H. (2006). Ecophysiology of a group of uncultured Gammaproteobacterial glycogen-accumulating organisms in full-scale enhanced biological phosphorus removal wastewater treatment plants. Environ. Microbiol. 8, 479–489. doi: 10.1111/j.1462-2920.2005.00914.x
Kong, Y., Xia, Y., Nielsen, J. L., and Nielsen, P. H. (2007). Structure and function of the microbial community in a full-scale enhanced biological phosphorus removal plant. Microbiology 153, 4061–4073. doi: 10.1099/mic.0.2007/007245-0
Kong, Y. H., Beer, M., Seviour, R. J., Lindrea, K. C., and Rees, G. N. (2001). Structure and functional analysis of the microbial community in an aerobic: anaerobic sequencing batch reactor (SBR) with no phosphorus removal. Syst. Appl. Microbiol. 24, 597–609. doi: 10.1078/0723-2020-00075
Kristiansen, R., Nguyen, H. T. T., Saunders, A. M., Nielsen, J. L., Wimmer, R., Le, V. Q., et al. (2013). A metabolic model for members of the genus Tetrasphaera involved in enhanced biological phosphorus removal. ISME J. 7, 543–554. doi: 10.1038/ismej.2012.136
Law, Y., Kirkegaard, R. H., Cokro, A. A., Liu, X., Arumugam, K., Xie, C., et al. (2016). Integrative microbial community analysis reveals full-scale enhanced biological phosphorus removal under tropical conditions. Sci. Rep. 6:25719. doi: 10.1038/srep25719
Lemos, P. C., Dai, Y., Yuan, Z., Keller, J., Santos, H., and Reis, M. A. (2007). Elucidation of metabolic pathways in glycogen-accumulating organisms with in vivo 13C nuclear magnetic resonance. Environ. Microbiol. 9, 2694–2706. doi: 10.1111/j.1462-2920.2007.01382.x
Liu, W.-T., Hanada, S., Marsh, T. L., Kamagata, Y., and Nakamura, K. (2002). Kineosphaera limosa gen. nov., sp. nov., a novel Gram-positive polyhydroxyalkanoate-accumulating coccus isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 52, 1845–1849. doi: 10.1099/00207713-52-5-1845
Liu, W.-T., Mino, T., Matsuo, T., and Nakamura, K. (2000). Isolation, characterization and identification of polyhydroxyalkanoate-accumulating bacteria from activated sludge. J. Biosci. Bioeng. 90, 494–500. doi: 10.1016/S1389-1723(01)80029-3
Liu, W.-T., Mino, T., Nakamura, K., and Matsuo, T. (1994). Role of glycogen in acetate uptake and polyhydroxyalkanoate synthesis in anaerobic-aerobic activated sludge with a minimized polyphosphate content. J. Ferment. Bioeng. 77, 535–540. doi: 10.1016/0922-338X(94)90124-4
Lohse, M., Bolger, A. M., Nagel, A., Fernie, A. R., Lunn, J. E., Stitt, M., et al. (2012). RobiNA: a user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 40, W622–W627. doi: 10.1093/nar/gks540
Lu, H., Oehmen, A., Virdis, B., Keller, J., and Yuan, Z. (2006). Obtaining highly enriched cultures of Candidatus Accumulibacter phosphatis through alternating carbon sources. Water Res. 40, 3838–3848.
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res. 32, 1363–1371. doi: 10.1093/nar/gkh293
Magoč, T., and Salzberg, S. L. (2011). FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963. doi: 10.1093/bioinformatics/btr507
Majed, N., and Gu, A. Z. (2010). Application of raman microscopy for simultaneous and quantitative evaluation of multiple intracellular polymers dynamics functionally relevant to enhanced biological phosphorus removal processes. Environ. Sci. Technol. 44, 8601–8608. doi: 10.1021/es1016526
Mao, Y., Graham, D. W., Tamaki, H., and Zhang, T. (2015). Dominant and novel clades of Candidatus Accumulibacter phosphatis in 18 globally distributed full-scale wastewater treatment plants. Sci. Rep. 5:11857. doi: 10.1038/srep11857
Martín, H. G., Ivanova, N., Kunin, V., Warnecke, F., Barry, K. W., McHardy, A. C., et al. (2006). Metagenomic analysis of two enhanced biological phosphorus removal (EBPR) sludge communities. Nat. Biotechnol. 24, 1263–1269. doi: 10.1038/nbt1247
Maszenan, A. M., Seviour, R. J., Patel, B. K., Janssen, P. H., and Wanner, J. (2005). Defluvicoccus vanus gen. nov., sp. nov., a novel Gram-negative coccus/coccobacillus in the Alphaproteobacteria from activated sludge. Int. J. Syst. Evol. Microbiol. 55, 2105–2111. doi: 10.1099/ijs.0.02332-0
Maszenan, A. M., Seviour, R. J., Patel, B. K., Schumann, P., Burghardt, J., Webb, R. I., et al. (1999a). Friedmanniella spumicola sp. nov. and Friedmanniella capsulata sp. nov. from activated sludge foam: Gram-positive cocci that grow in aggregates of repeating groups of cocci. Int. J. Syst. Bacteriol. 49(Pt 4), 1667–1680. doi: 10.1099/00207713-49-4-1667
Maszenan, A. M., Seviour, R. J., Patel, B. K., Schumann, P., and Rees, G. N. (1999b). Tessaracoccus bendigoensis gen. nov., sp. nov., a Gram-positive coccus occurring in regular packages or tetrads, isolated from activated sludge biomass. Int. J. Syst. Bacteriol. 49(Pt 2), 459–468. doi: 10.1099/00207713-49-2-459
Maszenan, A. M., Seviour, R. J., Patel, B. K. C., and Schumann, P. (2002). Quadricoccus australiensis gen. nov., sp. nov., a β-proteobacterium from activated sludge biomass. Int. J. Syst. Evol. Microbiol. 52, 223–228. doi: 10.1099/00207713-52-1-223
McIlroy, S., and Seviour, R. J. (2009). Elucidating further phylogenetic diversity among the Defluviicoccus–related glycogen-accumulating organisms in activated sludge. Environ. Microbiol. Rep. 1, 563–568. doi: 10.1111/j.1758-2229.2009.00082.x
McIlroy, S. J., Albertsen, M., Andresen, E. K., Saunders, A. M., Kristiansen, R., Stokholm-Bjerregaard, M., et al. (2014). “Candidatus Competibacter”-lineage genomes retrieved from metagenomes reveal functional metabolic diversity. ISME J. 8, 613–624. doi: 10.1038/ismej.2013.162
McIlroy, S. J., Kristiansen, R., Albertsen, M., Karst, S. M., Rossetti, S., Nielsen, J. L., et al. (2013). Metabolic model for the filamentous “Candidatus Microthrix parvicella” based on genomic and metagenomic analyses. ISME J. 7, 1161–1172. doi: 10.1038/ismej.2013.6
McIlroy, S. J., Nittami, T., Kanai, E., Fukuda, J., Saunders, A. M., and Nielsen, P. H. (2015a). Re-appraisal of the phylogeny and fluorescence in situ hybridization probes for the analysis of the Competibacteraceae in wastewater treatment systems. Environ. Microbiol. Rep. 7, 166–174. doi: 10.1111/1758-2229.12215
McIlroy, S. J., Saunders, A. M., Albertsen, M., Nierychlo, M., McIlroy, B., Hansen, A. A., et al. (2015b). MiDAS: the field guide to the microbes of activated sludge. Database 2015, 1–8. doi: 10.1093/database/bav062
McIlroy, S. J., Nittami, T., Seviour, E. M., and Seviour, R. J. (2010). Filamentous members of cluster III Defluviicoccus have the in situ phenotype expected of a glycogen-accumulating organism in activated sludge. FEMS Microbiol. Ecol. 74, 248–256. doi: 10.1111/j.1574-6941.2010.00934.x
McIlroy, S. J., Starnawska, A., Starnawski, P., Saunders, A. M., Nierychlo, M., Nielsen, P. H., et al. (2016). Identification of active denitrifiers in full-scale nutrient removal wastewater treatment systems. Environ. Microbiol. 18, 50–64. doi: 10.1111/1462-2920.12614
McIlroy, S. J., Tillett, D., Petrovski, S., and Seviour, R. J. (2011). Non-target sites with single nucleotide insertions or deletions are frequently found in 16S rRNA sequences and can lead to false positives in fluorescence in situ hybridization (FISH). Environ. Microbiol. 13, 38–47. doi: 10.1111/j.1462-2920.2010.02306.x
McMahon, K. D., Yilmaz, S., He, S., Gall, D. L., Jenkins, D., and Keasling, J. D. (2007). Polyphosphate kinase genes from full-scale activated sludge plants. Appl. Microbiol. Biotechnol. 77, 167–173. doi: 10.1007/s00253-007-1122-6
McMurdie, P. J., and Holmes, S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 8:e61217. doi: 10.1371/journal.pone.0061217
Mesquita, D. P., Leal, C., Cunha, J. R., Oehmen, A., Amaral, A. L., Reis, M. A. M., et al. (2013). Prediction of intracellular storage polymers using quantitative image analysis in enhanced biological phosphorus removal systems. Anal. Chim. Acta 770, 36–44. doi: 10.1016/j.aca.2013.02.002
Meyer, R. L., Saunders, A. M., and Blackall, L. L. (2006). Putative glycogen-accumulating organisms belonging to the Alphaproteobacteria identified through rRNA-based stable isotope probing. Microbiology 152, 419–429. doi: 10.1099/mic.0.28445-0
Mielczarek, A. T., Nguyen, H. T., Nielsen, J. L., and Nielsen, P. H. (2013). Population dynamics of bacteria involved in enhanced biological phosphorus removal in Danish wastewater treatment plants. Water Res. 47, 1529–1544. doi: 10.1016/j.watres.2012.12.003
Mino, T., Arun, V., Tsuzuki, Y., and Matsuo, T. (1987). “Effect of phosphorus accumulation on acetate metabolism in the biological phosphorus removal process,” in Biological Phorphorus Removal from Wastewaters, ed. R. Ramadori (Oxford: Pergamon Press), 27–38.
Mino, T., Liu, W.-T., Kurisu, F., and Matsuo, T. (1995). Modelling glycogen storage and denitrification capability of microorganisms in enhanced biological phosphate removal processes. Water Sci. Technol. 31, 25–34. doi: 10.1016/0273-1223(95)00177-O
Molinos-Senante, M., Hernández-Sancho, F., Sala-Garrido, R., and Garrido-Baserba, M. (2011). Economic feasibility study for phosphorus recovery processes. Ambio 40, 408–416. doi: 10.1007/s13280-010-0101-9
Nakamura, K., Hiraishi, A., Yoshimi, Y., Kawaharasaki, M., Masuda, K., and Kamagata, Y. (1995a). Microlunatus phosphovorus gen. nov., sp. nov., a new Gram-positive polyphosphate-accumulating bacterium isolated from activated sludge. Int. J. Syst. Bacteriol. 45, 17–22. doi: 10.1099/00207713-45-1-17
Nakamura, K., Ishikawa, S., and Kawaharasaki, M. (1995b). Phosphate uptake and release activity in immobilized polyphosphate-accumulating bacterium Microlunatus phosphovorus strain NM-1. J. Ferment. Bioeng. 80, 377–382. doi: 10.1016/0922-338X(95)94207-8
Nakamura, K., Masuda, K., and Mikami, E. (1991). Isolation of a new type of polyphosphate accumulating bacterium and its phosphate removal characteristics. J. Ferment. Bioeng. 71, 258–263. doi: 10.1016/0922-338X(91)90278-O
Nguyen, H. T., Le, V. Q., Hansen, A. A., Nielsen, J. L., and Nielsen, P. H. (2011). High diversity and abundance of putative polyphosphate-accumulating Tetrasphaera-related bacteria in activated sludge systems. FEMS Microbiol. Ecol. 76, 256–267. doi: 10.1111/j.1574-6941.2011.01049.x
Nguyen, H. T., Nielsen, J. L., and Nielsen, P. H. (2012). “Candidatus Halomonas phosphatis”, a novel polyphosphate-accumulating organism in full-scale enhanced biological phosphorus removal plants. Environ. Microbiol. 14, 2826–2837. doi: 10.1111/j.1462-2920.2012.02826.x
Nguyen, H. T. T., Kristiansen, R., Vestergaard, M., Wimmer, R., and Nielsen, P. H. (2015). Intracellular accumulation of glycine in polyphosphate-accumulating organisms in activated sludge, a novel storage mechanism under dynamic anaerobic-aerobic conditions. Appl. Environ. Microbiol. 81, 4809–4818. doi: 10.1128/AEM.01012-15
Nierychlo, M., Nielsen, J. L., and Nielsen, P. H. (2015). “Studies of the ecophysiology of single cells in microbial communities by (Quantitative) microautoradiography and fluorescence In Situ hybridization (MAR-FISH),” in Hydrocarbon and Lipid Microbiology Protocols, Springer Protocols Handbooks, eds T. J. McGenity, K. N. Timmis, and B. Nogales Fernández (Berlin: Springer-Verlag).
Nittami, T., McIlroy, S., Seviour, E. M., Schroeder, S., and Seviour, R. J. (2009). Candidatus Monilibacter spp., common bulking filaments in activated sludge, are members of Cluster III Defluviicoccus. Syst. Appl. Microbiol. 32, 480–489. doi: 10.1016/j.syapm.2009.07.003
Oehmen, A., Lemos, P. C., Carvalho, G., Yuan, Z., Keller, J., Blackall, L. L., et al. (2007). Advances in enhanced biological phosphorus removal: from micro to macro scale. Water Res. 41, 2271–2300. doi: 10.1016/j.watres.2007.02.030
Oehmen, A., Lopez-Vazquez, C. M., Carvalho, G., Reis, M. A., and van Loosdrecht, M. C. (2010). Modelling the population dynamics and metabolic diversity of organisms relevant in anaerobic/anoxic/aerobic enhanced biological phosphorus removal processes. Water Res. 44, 4473–4486. doi: 10.1016/j.watres.2010.06.017
Oyserman, B. O., Noguera, D. R., Del Rio, T. G., Tringe, S. G., and McMahon, K. D. (2015). Metatranscriptomic insights on gene expression and regulatory controls in Candidatus Accumulibacter phosphatis. ISME J. 10, 810–822. doi: 10.1038/ismej.2015.155
Peterson, S. B., Warnecke, F., Madejska, J., McMahon, K. D., and Hugenholtz, P. (2008). Environmental distribution and population biology of Candidatus Accumulibacter, a primary agent of biological phosphorus removal. Environ. Microbiol. 10, 2692–2703. doi: 10.1111/j.1462-2920.2008.01690.x
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
R Core Team (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rossetti, S., Tomei, M. C., Nielsen, P. H., and Tandoi, V. (2005). “Microthrix parvicella”, a filamentous bacterium causing bulking and foaming in activated sludge systems: a review of current knowledge. FEMS Microbiol. Rev. 29, 49–64. doi: 10.1016/j.femsre.2004.09.005
Santos, M. M., Lemos, P. C., Reis, M. A., and Santos, H. (1999). Glucose metabolism and kinetics of phosphorus removal by the fermentative bacterium Microlunatus phosphovorus. Appl. Environ. Microbiol. 65, 3920–3928.
Saunders, A. M., Albertsen, M., Vollertsen, J., and Nielsen, P. H. (2016). The activated sludge ecosystem contains a core community of abundant organisms. ISME J. 10, 11–20. doi: 10.1038/ismej.2015.117
Saunders, A. M., Mabbett, A. N., McEwan, A. G., and Blackall, L. L. (2007). Proton motive force generation from stored polymers for the uptake of acetate under anaerobic conditions. FEMS Microbiol. Lett. 274, 245–251. doi: 10.1111/j.1574-6968.2007.00839.x
Saunders, A. M., Oehmen, A., Blackall, L. L., Yuan, Z., and Keller, J. (2003). The effect of GAOs (glycogen accumulating organisms) on anaerobic carbon requirements in full-scale Australian EBPR (enhanced biological phosphorus removal) plants. Water Sci. Technol. 47, 37–43.
Schroeder, S., Petrovski, S., Campbell, B., McIlroy, S., and Seviour, R. J. (2009). Phylogeny and in situ identification of a novel gammaproteobacterium in activated sludge. FEMS Microbiol. Lett. 297, 157–163. doi: 10.1111/j.1574-6968.2009.01676.x
Serafim, L. S., Lemos, P. C., Levantesi, C., Tandoi, V., Santos, H., and Reis, M. A. M. (2002). Methods for detection and visualization of intracellular polymers stored by polyphosphate-accumulating microorganisms. J. Microbiol. Methods 51, 1–18. doi: 10.1016/S0167-7012(02)00056-8
Seviour, R. J., Maszenan, A. M., Soddell, J. A., Tandoi, V., Patel, B. K. C., Kong, Y., et al. (2000). Microbiology of the “G-bacteria” in activated sludge. Environ. Microbiol. 2, 581–593. doi: 10.1046/j.1462-2920.2000.00153.x
Seviour, R. J., and McIlroy, S. (2008). The microbiology of phosphorus removal in activated sludge processes- the current state of play. J. Microbiol. 46, 115–124. doi: 10.1007/s12275-008-0051-0
Seviour, R. J., Mino, T., and Onuki, M. (2003). The microbiology of biological phosphorus removal in activated sludge systems. FEMS Microbiol. Rev. 27, 99–127. doi: 10.1016/S0168-6445(03)00021-4
Shintani, T., Liu, W. T., Hanada, S., Kamagata, Y., Miyaoka, S., Suzuki, T., et al. (2000). Micropruina glycogenica gen. nov., sp. nov., a new Gram-positive glycogen-accumulating bacterium isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 50(Pt 1), 201–207. doi: 10.1099/00207713-50-1-201
Skennerton, C. T., Barr, J. J., Slater, F. R., Bond, P. L., and Tyson, G. W. (2015). Expanding our view of genomic diversity in Candidatus Accumulibacter clades. Environ. Microbiol. 17, 1574–1585. doi: 10.1111/1462-2920.12582
Slater, F. R., Johnson, C. R., Blackall, L. L., Beiko, R. G., and Bond, P. L. (2010). Monitoring associations between clade-level variation, overall community structure and ecosystem function in enhanced biological phosphorus removal (EBPR) systems using terminal-restriction fragment length polymorphism (T-RFLP). Water Res. 44, 4908–4923. doi: 10.1016/j.watres.2010.07.028
Soo, R. M., Skennerton, C. T., Sekiguchi, Y., Imelfort, M., Paech, S. J., Dennis, P. G., et al. (2014). An expanded genomic representation of the phylum Cyanobacteria. Genome Biol. Evol. 6, 1031–1045. doi: 10.1093/gbe/evu073
Spring, S., Wagner, M., Schumann, P., and Kämpfer, P. (2005). Malikia granosa gen. nov., sp. nov., a novel polyhydroxyalkanoate- and polyphosphate-accumulating bacterium isolated from activated sludge, and reclassification of Pseudomonas spinosa as Malikia spinosa comb. nov. Int. J. Syst. Evol. Microbiol. 55, 621–629. doi: 10.1099/ijs.0.63356-0
Stante, L., Cellamare, C. M., Malaspina, F., Bortone, G., and Tilche, A. (1997). Biological phosphorus removal by pure culture of Lampropedia spp. Water Res. 31, 1317–1324. doi: 10.1016/S0043-1354(96)00351-X
Tandoi, V., Majone, M., May, J. W., and Ramadori, R. (1998a). The behaviour of polyphosphate accumulating Acinetobacter isolates in an anerobic-aerobic chemostat. Water Res. 32, 2903–2912. doi: 10.1016/S0043-1354(98)00079-7
Tandoi, V., Rossetti, S., Blackall, L. L., and Majone, M. (1998b). Some physiological properties of an italian isolate of “Microthrix parvicella”. Water Sci. Technol. 37, 1–8. doi: 10.1016/S0273-1223(98)00077-8
Terashima, M., Yama, A., Sato, M., Yumoto, I., Kamagata, Y., and Kato, S. (2016). Culture-dependent and -independent identification of polyphosphate-accumulating Dechloromonas spp. predominating in a full-scale oxidation ditch wastewater treatment plant. Microbes Environ. 31, 449–455. doi: 10.1264/jsme2.ME16097
Tu, Y., and Schuler, A. J. (2013). Low acetate concentrations favor polyphosphate-accumulating organisms over glycogen-accumulating organisms in enhanced biological phosphorus removal from wastewater. Environ. Sci. Technol. 47, 3816–3824. doi: 10.1021/es304846s
Ubakata, Y., and Takii, S. (1994). Induction method of excess phosphate removing bacteria isolated from anaerobic/aerobic activated sludge. Water Sci. Technol. 30, 221–227.
Ubakata, Y., and Takii, S. (1998). Some physiological characteristics of phosphate-removing bacterium, Microlunatus phosphovorus, and a simplified isolation and identification method for phosphate-removing bacteria. Water Sci. Technol. 38, 149–157.
Wagner, M., Erhart, R., Manz, W., Amann, R., Lemmer, H., Wedi, D., et al. (1994). Development of an rRNA-targeted oligonucleotide probe specific for the genus Acinetobacter and its application for the in situ monitoring in activated sludge. Appl. Environ. Microbiol. 60, 792–800.
Wagner, M., and Haider, S. (2012). New trends in fluorescence in situ hybridization for identification and functional analyses of microbes. Curr. Opin. Biotechnol. 23, 96–102. doi: 10.1016/j.copbio.2011.10.010
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07
Wang, J., Qi, R., Liu, M., Li, Q., Bao, H., Li, Y., et al. (2014). The potential role of ‘Candidatus Microthrix parvicella’ in phosphorus removal during sludge bulking in two full-scale enhanced biological phosphorus removal plants. Water Sci. Technol. 70, 367–375. doi: 10.2166/wst.2014.216
Wang, Z., Guo, F., Mao, Y., Xia, Y., and Zhang, T. (2014). Metabolic characteristics of a glycogen-accumulating organism in Defluviicoccus cluster II revealed by comparative genomics. Microb. Ecol. 68, 716–728. doi: 10.1007/s00248-014-0440-3
Welles, L., Lopez-Vazquez, C. M., Hooijmans, C. M., van Loosdrecht, M. C. M., Brdjanovic, D., Acevedo, B., et al. (2016). Prevalence of “Candidatus Accumulibacter phosphatis” type II under phosphate limiting conditions. AMB Express 6, 44. doi: 10.1186/s13568-016-0214-z
Welles, L., Tian, W. D., Saad, S., Abbas, B., Lopez-Vazquez, C. M., Hooijmans, C. M., et al. (2015). Accumulibacter clades Type I and II performing kinetically different glycogen-accumulating organisms metabolisms for anaerobic substrate uptake. Water Res. 83, 354–366. doi: 10.1016/j.watres.2015.06.045
Wentzel, M. C., Lotter, L. H., Loewenthal, R. E., and Marais, G. V. R. (1986). Metabolic behaviour of Acinetobacter spp. in enhanced biological phosphorus removal - a biochemical model. Water SA 12, 209–224.
Wong, M.-T., and Liu, W.-T. (2007). Ecophysiology of Defluviicoccus-related tetrad-forming organisms in an anaerobic-aerobic activated sludge process. Environ. Microbiol. 9, 1485–1496. doi: 10.1111/j.1462-2920.2007.01267.x
Wong, M. T., Mino, T., Seviour, R. J., Onuki, M., and Liu, W.-T. (2005). In situ identification and characterization of the microbial community structure of full-scale enhanced biological phosphorous removal plants in Japan. Water Res. 39, 2901–2914. doi: 10.1016/j.watres.2005.05.015
Wong, M.-T., Tan, F. M., Ng, W. J., and Liu, W.-T. (2004). Identification and occurrence of tetrad-forming Alphaproteobacteria in anaerobic-aerobic activated sludge processes. Microbiology 150, 3741–3748. doi: 10.1099/mic.0.27291-0
Yoshimi, Y., Hiraishi, A., and Nakamura, K. (1996). Isolation and characterization of Microsphaera multipartita gen. nov., sp. nov., a polysaccharide-accumulating Gram-positive bacterium from activated sludge. Int. J. Syst. Evol. Microbiol. 46, 519–525. doi: 10.1099/00207713-46-2-519
Zhang, H., Sekiguchi, Y., Hanada, S., Hugenholtz, P., Kim, H., Kamagata, Y., et al. (2003). Gemmatimonas aurantiaca gen. nov., sp. nov., a Gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int. J. Syst. Evol. Microbiol. 53, 1155–1163. doi: 10.1099/ijs.0.02520-0
Zhang, T., Shao, M.-F., and Ye, L. (2012). 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 6, 1137–1147. doi: 10.1038/ismej.2011.188
Zhou, Y., Pijuan, M., Zeng, R. J., Lu, H., and Yuan, Z. (2008). Could polyphosphate-accumulating organisms (PAOs) be glycogen-accumulating organisms (GAOs)? Water Res. 42, 2361–2368. doi: 10.1016/j.watres.2008.01.003
Ziegler, A. S., McIlroy, S. J., Larsen, P., Albertsen, M., Hansen, A. A., Heinen, N., et al. (2016). Dynamics of the fouling layer microbial community in a membrane bioreactor. PLoS ONE 11:e0158811. doi: 10.1371/journal.pone.0158811
Zilles, J. L., Peccia, J., Kim, M. W., Hung, C. H., and Noguera, D. R. (2002a). Involvement of Rhodocyclus-related organisms in phosphorus removal in full-scale wastewater treatment plants. Appl. Environ. Microbiol. 68, 2763–2769. doi: 10.1128/AEM.68.6.2763-2769.2002
Keywords: 16S rRNA amplicon sequencing, Accumulibacter, activated sludge, EBPR, GAO, PAO, Micropruina, Tetrasphaera
Citation: Stokholm-Bjerregaard M, McIlroy SJ, Nierychlo M, Karst SM, Albertsen M and Nielsen PH (2017) A Critical Assessment of the Microorganisms Proposed to be Important to Enhanced Biological Phosphorus Removal in Full-Scale Wastewater Treatment Systems. Front. Microbiol. 8:718. doi: 10.3389/fmicb.2017.00718
Received: 09 December 2016; Accepted: 06 April 2017;
Published: 27 April 2017.
Edited by:
Kartik Chandran, Columbia University, USAReviewed by:
Cindy J. Smith, University of Glasgow, UKJose M. Bruno-Barcena, North Carolina State University, USA
Copyright © 2017 Stokholm-Bjerregaard, McIlroy, Nierychlo, Karst, Albertsen and Nielsen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Per H. Nielsen, phn@bio.aau.dk
†These authors have contributed equally to this work.