 Iker A. Sevilla
Iker A. Sevilla Elena Molina
Elena Molina Maitane Tello
Maitane Tello Natalia Elguezabal
Natalia Elguezabal Ramón A. Juste
Ramón A. Juste Joseba M. Garrido
Joseba M. Garrido- 1Animal Health Department, NEIKER-Instituto Vasco de Investigación y Desarrollo Agrario, Bizkaia Science and Technology Park 812L, Derio, Spain
- 2SERIDA-Servicio Regional de Investigación y Desarrollo Agrario, Carretera de Oviedo, Villaviciosa, Spain
Mycobacteria include obligate and opportunistic pathogens that cause significant human and animal disease. The burden of tuberculosis has been largely reduced in developed territories but remains a huge problem worldwide. The significance of nontuberculous mycobacteria is growing considerably, especially in developed regions with higher life expectancy and more therapy-related immunosuppressed individuals. Due to their robustness mycobacteria can contaminate animal products by direct transmission from infected individuals or by environmental contamination during processing. The situation at market level is poorly known. Most studies analyzing commercially available foods are limited to a small or local scale and mainly focused on a particular mycobacterial species. There is a need to investigate if animal products that have passed the established controls to be for sale at main supermarkets could represent a route of contact with any mycobacteria. Thus, our goal was to study the prevalence of mycobacteria in these foods to assess if this could represent a source of human exposure. Five stores from the main supermarket chains in Spain were selected. 138 dairy and 119 meat products were purchased. All were processed using culture and multiplex real-time PCR methods. Additional molecular methods were used to specifically identify any positive result. Mycobacterium avium subsp. hominissuis (2), M. avium subsp. avium (1), and M. fortuitum (1) were isolated from powdered infant formula and ground beef, chicken sausage, and mortadella cold cut, respectively. Mycobacterial DNA (M. avium, M. tuberculosis complex and other nontuberculous mycobacteria) was detected in 15% of dairy products and 2% of meat products. These results show that the prevalence of viable mycobacteria in foods of animal origin obtained at the supermarket was not substantial although a considerable proportion of them contained mycobacterial DNA. Contact with mycobacteria through this route could be ensured over time. Further investigation is necessary to determine the real impact of foodborne mycobacterial exposure on human health and identify critical points in the food production system to enable setting up more stringent control measures.
Introduction
The genus Mycobacterium includes important obligate and opportunistic infectious microorganisms that cause human and animal disease. M. tuberculosis complex together with M. avium complex and other nontuberculous mycobacteria (NTM) like M. kansasii, M. malmoense, M. chelonae-abscessus group, and M. fortuitum complex stand out among the rest of mycobacterial species in terms of medical and veterinary significance (Biet and Boschiroli, 2014; Falkinham, 2016). M. tuberculosis complex encompasses the etiological agents of human and zoonotic animal tuberculosis (TB), diseases of great health and socioeconomic impact as indicated in annual reports of the World Health Organization and manuals of the World Organization for Animal Health. M. avium subspecies are responsible for paratuberculosis (PTB), avian TB, infections in swine and other domestic and wild species, and cervical lymphadenitis, respiratory disease and disseminated or focal infections in humans (Biet and Boschiroli, 2014; Falkinham, 2016). M. kansasii is responsible for human disseminated infections and lung disease almost identical to typical TB (Griffith et al., 2007) and is commonly found in lymph node and tissue lesions in cattle in some regions (Biet and Boschiroli, 2014). The incidence of some ubiquitous fast growing NTM species (e.g., M. fortuitum complex and M. chelonae-abscessus) considered opportunistic pathogens for humans (Griffith et al., 2007) and animals (Bercovier and Vincent, 2001; Biet and Boschiroli, 2014) that originate diverse infections, has increased notably during the last decades (García-Martos and García-Agudo, 2012). In regions with developed health care resources the number of individuals susceptible to mycobacterial infection, especially with NTM, is growing due to the aging of the population, the maintenance of natural immune deficiencies and chronic diseases and the use of immunosuppressive therapies (Lake et al., 2016). Many mycobacterial infections originate from airborne exposure. However, other routes are also important, including the oral route.
Reports on foodborne (excluding water) mycobacterial transmission are mainly limited to zoonotic TB cases derived from the consumption of untreated or not properly treated M. bovis-contaminated foods. In developed territories infection through this route is not as frequent as in developing ones as a consequence of milk treatment and tuberculosis eradication programs (Pérez-Lago et al., 2014). But other mycobacterial infections can be contracted through oral exposure apart from TB. NTM cervical lymphadenitis affects mainly infants and there is circumstantial evidence indicating that infection originates from oral exposure (Kasperbauer and Huitt, 2013), most likely from water (Falkinham, 2003), and thus contaminated drinks and foods should not be discarded as possible vehicles. In most cases of NTM disseminated infection mycobacteria are introduced through the lungs or the digestive tract (Kasperbauer and Huitt, 2013). An association between NTM lung infection and gastroesophageal disorders has been previously recognized, being aspiration of contaminated water a probable means of bacterial introduction (Thomson et al., 2013). An etiologic role has been attributed to M. avium subsp. paratuberculosis in Crohn's disease but it remains controversial (Sechi and Dow, 2015). If this link is finally demonstrated, consumption of contaminated milk and dairy products could represent the most likely source of M. avium subsp. paratuberculosis (Grant, 2010). In ruminants the oral route is the main route of entry of this bacterium as is the case of M. avium subsp. avium in avian TB (Tell et al., 2001), and thus it could also represent a means of contact for humans.
The term environmental for NTM is determined by their presence in water, pipe systems, soil, dust and other ecosystems (Falkinham, 2016). Tuberculous mycobacteria and also NTM can contaminate foods of animal origin passing directly from infected individuals (including also their fluids and feces) to the product, but environmental contamination during the processing and storage is also possible (Grant, 2010; Ghodbane et al., 2014; Klanicova-Zalewska and Slana, 2014). In addition, as a result of their robustness mycobacteria can resist adverse conditions and survive to some of the disinfection or decontamination procedures applied to foods and drinks. In terms of microbiological safety and hygiene the presence of mycobacteria in foods represents a potential biological hazard that should be prevented. There is a lack of information with reference to supermarket foods that have passed the established controls. Many studies report on the detection of mycobacteria in animal tissues and raw milk and thus support that foods of animal origin may act as vehicles of mycobacteria transmission to humans (Kaneene et al., 2014; Klanicova-Zalewska and Slana, 2014; Pérez-Lago et al., 2014; Waddell et al., 2016). Several surveys have found mycobacterial DNA and viable cells in commercially available foodstuffs but the number of such reports is limited and most of them are mainly focused on the detection of M. avium subspecies or M. bovis (Grant, 2010; Klanicova et al., 2011; Faria et al., 2014; Lorencova et al., 2014; Pereira-Suarez et al., 2014; Savi et al., 2015; Cezar et al., 2016). According to the bibliography, animal products in which mycobacteria have been identified include raw and pasteurized milk, milk powder, powdered infant milk formula, cheese, meat, ground meat, fermented meat, ham, salami and sausages obtained from the main livestock species. Under these circumstances and taking into account the apparent increasing vulnerability of developed populations to some mycobacterial infections, we centered our attention on retail animal products available at the supermarket to determine whether they could represent a source of contact with mycobacteria for humans. For this purpose, processed and raw dairy and meat products purchased at different supermarkets were screened for the presence of members of the genus Mycobacterium using culture and PCR-based methods.
Materials and Methods
Dairy and Meat Products
Five stores belonging to 5 of the main supermarket chains operating in Spain were selected. A total of 138 dairy and 119 meat products of different types and brands were collected at two sampling times. Samples covered top-selling brands along with own-label brands. Dairy foodstuffs included milk, milk powder, powdered infant formula, cream, butter, cheese and yogurt, and meat products consisted of packed ground meat, hamburger patties, fresh and cooked sausages, cold cuts, fresh Spanish chorizo and pâté. Most products were collected at both sampling times, thus two different batches of the same brand could be analyzed. All of them are frequently consumed products produced, processed, bottled or packed in Spain (89.9%), France (6.6%), Ireland (0.8%), Italy (0.4%), Germany (0.8%), Netherlands (0.8%), and Switzerland (0.8%). The number and type of products analyzed is summarized in Table 1. Brand names are not indicated to preserve the anonymity of the survey.
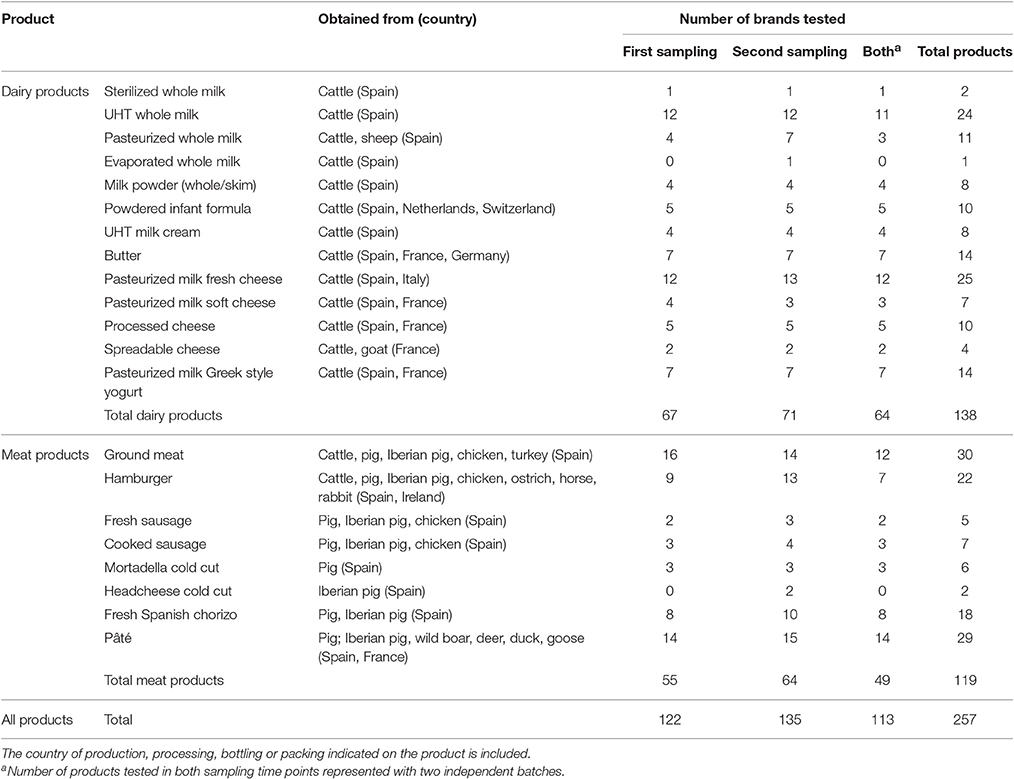
Table 1. List of food types and number of brands and items analyzed.
Culture
All samples were cultured in Lowenstein-Jensen with pyruvate (Difco, Francisco Soria Melguizo SA, Madrid, Spain), Coletsos (Difco), modified Middlebrook 7H10 (Becton, Dickinson and Company, MD, USA) with egg yolk and mycobactin J (IDvet, Grabels, France) prepared as previously described (Whittington et al., 2011) and in BBL Mycobacteria growth indicator tubes (MGIT) supplemented with BACTEC MGIT growth supplement and BBL MGIT PANTA (Becton, Dickinson and Company). Cultures on solid media were incubated at 37°C for 6 months and periodically inspected for the presence of colonies. MGIT cultures were incubated for 3 months in a BACTEC MGIT 960 System and time to detection (TTD) values recorded. Colonies on slants and MGIT cultures with a TTD value were submitted to PCR for verification and identification purposes. Different culture procedures were performed according to the sample type.
Milk, Milk Powder, and Powdered Infant Formula
Powdered material was reconstituted with sterile distilled water following manufacturer's instructions and then treated like liquid milk. For solid cultures 30 ml of milk was centrifuged at 8,000 × g for 20 min. The pelleted material and the cream were homogenized and resuspended in 12 ml of 0.75% (wt/vol) RonaCare Cetylpyridinium Chloride (Merck, Darmstadt, Germany). Tubes were incubated on a rotating wheel at room temperature for 5 h (Donaghy et al., 2008). Samples were then centrifuged at 3,500 × g for 15 min. The intermediate liquid phase was discarded and the pellet and the remaining upper creamy fraction thoroughly vortexed with 1.5 ml of phosphate-buffered saline (PBS) containing 0.05% (wt/vol) Tween 20 (Sigma-Aldrich Co. Ltd., St. Louis, MO, USA). Solid media tubes were inoculated with 0.2 ml of this suspension. An additional 30 ml aliquot of samples was used for liquid culture. Following centrifugation at 8,000 × g for 20 min, the pellet and cream fractions were thoroughly homogenized in 10 ml of PBS-Tween 20. Afterwards, samples were processed with BD BBL MycoPrep kit (Becton, Dickinson and Company) following the instructions of the manufacturer and the resulting material was inoculated in appropriately supplemented MGIT tubes.
Cream, Butter, Cheese, and Yogurt
Ninety ml of pre-warmed diluent containing 0.5% (wt/vol) sodium chloride, 2% (wt/vol) sodium citrate and 1% (wt/vol) Bacto Casitone (Difco, Becton, Dickinson and Company) were added to either 10 g of butter, 10 g of cheese or 10 g of yogurt contained in filter stomacher bags. For cream 30 ml of this product and 60 ml of diluent were used. Filter bags were introduced in a stomacher lab blender and suspensions homogenized for 3 min at high speed. After incubation at 56°C for 1 h on an orbital shaker platform, the homogenization step in the stomacher was repeated. Sixty ml of these suspensions were processed for solid (30 ml) and liquid (30 ml) culture following the steps previously described for milk samples. Ten milliliter of the same suspensions were used for DNA extraction.
Meat Products
Filter stomacher bags were used to weigh 2 g of meat product. Following addition of freshly prepared RonaCare Cetylpyridinium Chloride (38 ml), samples were thoroughly homogenized in a lab blender and the suspension transferred to 50 ml centrifuge tubes. Tubes were then spun at 100 × g for 1 min and 15 ml of the supernatant containing no gross material was transferred to new centrifuge tubes. After an overnight decontamination, samples were centrifuged at 3,500 × g for 15 min and pellets resuspended in 1.5 ml of PBS-Tween 20. This was the suspension used to inoculate solid media tubes. An additional portion of 2 g was homogenized in a filter stomacher bag containing 10 ml of sterile distilled water. The homogenate was collected in 50 ml tubes and treated for MGIT culture as indicated before.
DNA Extraction
Dairy Products
Adiapure milk kit (Adiagèn, bioMérieux, Marcy l'Etoile, France) was used as per manufacturer's instructions to extract DNA from milk and dairy product suspensions previously prepared for culture purposes (Donaghy et al., 2011). Briefly, a mixture consisting of 10 ml of sample and lysis buffer was vortexed and magnetic beads added. Tubes were gently mixed on a rotating wheel during 30 min for mycobacteria capture. Magnetic beads were separated from the liquid phase using a 15/50 ml tube magnet (Qiagen GmbH, Hilden, Germany), resuspended in the appropriate reagent and disrupted with glass beads for 10 min at maximum speed in a Tissue Lyser II (Qiagen) to lyse cells bound to magnetic beads. Beads were precipitated by centrifugation and supernatants treated with the buffer containing proteinase K.
Meat Products
A modified protocol of the Speedtools Tissue DNA extraction kit (Biotools B&M Labs S. A., Madrid, Spain) was used (Sevilla et al., 2015). Meat product samples (2 g) were stomached with 10 ml of sterile distilled water in stomacher filter bags to give a homogeneous suspension. The material suspended in 1.25 ml was pelleted and the supernatant removed. Zirconia/silica beads of 0.1 mm diameter (Bio Spec Products Inc., Bartlesville, OK, USA), lysis buffer and proteinase K was added to tubes. Samples were incubated at 56°C with agitation in a ThermoMixer (Eppendorf, San Diego, CA, USA) until the suspension was completely clear. Incubation was extended with additional fresh proteinase K if this was not achieved at the first attempt. Subsequent processing was as the standard protocol of the kit indicated except for the inclusion of one bead-beating step in a Tissue Lyser II (Qiagen).
Positive Cultures
One milliliter of MGIT broth was centrifuged at 16,000 × g for 3 min and the pellets washed with sterile distilled water. To extract DNA from these pellets and from the colonies grown on solid media the modified Speedtools Tissue DNA extraction kit (Biotools B&M Labs S. A.) protocol described above was employed.
All DNA extracts were stored at −20°C if not used immediately.
Tetraplex Real-Time PCR Screening
DNA from food products and cultures was tested using an improved version of a real-time PCR able to simultaneously detect the genus Mycobacterium, the M. avium subspecies, the M. tuberculosis complex and an internal amplification control (Sevilla et al., 2015). Modifications involved oligonucleotide changes in M. avium and M. tuberculosis complex PCR components to increase performance and accommodate the method to the use of different PCR mastermixes and platforms. The new protocol is currently under evaluation by an external party with commercial interests. Using the unmodified assay would yield similar results if performed under the same conditions reported earlier for the original method (Sevilla et al., 2015). Samples were tested in duplicate and all assays were validated based on the results obtained for internal amplification control, DNA extraction negative and positive controls, no template PCR controls and PCR positive controls. Amplification was carried out in a 7,500 Real-Time PCR instrument (Applied Biosystems, Foster City, CA, USA) under universal TaqMan assay conditions and SDS software v. 1.5.1 (Applied Biosystems) was used to calculate valid threshold cycle (CT) and baseline. CT values can be used as a rough reference of the estimated numbers of mycobacteria that could be present in samples (Sevilla et al., 2015).
Confirmation and Further Identification of Samples and Cultures Positive to the Tetraplex Real-Time PCR Screening
Samples Positive to M. avium
Samples positive for M. avium were submitted to IS900-ISMap02 real-time PCR (Sevilla et al., 2014) and other conventional (Bartos et al., 2006) or real-time (Slana et al., 2010) PCR methods to detect IS1245, IS901, and IS901-flanking region (FR300). M. avium subspecies were identified on the basis of the presence (+) or absence (−) of the specified genomic targets as follows: M. avium subsp. paratuberculosis is IS900+, ISMap02+, IS1245−, IS901−, and FR300+; M. avium subsp. avium is IS900−, ISMap02−, IS1245+, IS901+ and FR300−; M. avium subsp. silvaticum is IS900−, ISMap02−, IS1245+, IS901+, and FR300+, M. avium subsp. hominissuis is IS900−, ISMap02−, IS1245+, IS901−, and FR300+ (Bartos et al., 2006).
Samples Positive to M. tuberculosis Complex
To confirm the positive results and distinguish between the species grouped in the M. tuberculosis complex, these DNA samples were analyzed by spoligotyping (Kamerbeek et al., 1997) and a panel of conventional singleplex PCR assays using primers and conditions previously described to detect the regions of difference (RD) 1, 4, 9, and 12 of M. tuberculosis (Halse et al., 2011). The RD signature patterns used to differentiate between M. tuberculosis complex members were those specified earlier (Halse et al., 2011). Briefly, M. tuberculosis is positive to all RDs; M. canettii is RD1+, RD4+, RD9+, and RD12−; M. africanum and pinnipedii are RD1+, RD4+, RD9−, and RD12+; M. microti is RD1−, RD4+, RD9−, and RD12+; M. caprae is RD1+, RD4+, RD9−, and RD12−; M. bovis is RD1+, RD4−, RD9−, and RD12−.
Samples Positive to Mycobacterium sp. but Negative to M. avium and M. tuberculosis Complex
Species or complex identification and confirmation of samples only positive to the genus Mycobacterium was attempted by PCR and sequence analysis of 16S-23S rRNA internal transcribed spacer (ITS) (Richter et al., 1999) or by sequencing the short amplicon obtained for the ITS component (genus) in the screening PCR (Sevilla et al., 2015). Sequencing primers were the same used for amplification in both cases. Sequencing reactions were carried out using BigDye Terminator chemistry on a 3,130 Genetic Analyzer (Applied Biosystems). Sequences were inspected, edited and aligned with Sequencing Analysis 5.2 (Applied Biosystems) and Vector NTI (Informax Inc., Bethesda, MD, USA) software assistance and compared with other publicly available sequences using online BLAST (NCBI, NLM, Bethesda, MD, USA) analysis.
Results
There was no agreement between PCR and culture positive results, no sample tested positive for both. Four dairy products that were positive in the direct PCR were also positive to the same PCR component when DNA extracted from MGIT culture was used, but viability could not be demonstrated as explained below.
Culture Results and Identification of Isolates
Colonies grown on solid media and MGIT cultures with positive TTD readouts on the BACTEC instrument were screened for the presence of mycobacterial DNA using the tetraplex PCR. All but six MGIT and two Middlebrook 7H10 cultures tested negative (see Table 2). However, four of these MGIT cultures yielded high CT values (Table 2) and subculture attempts failed to demonstrate actual cell viability.
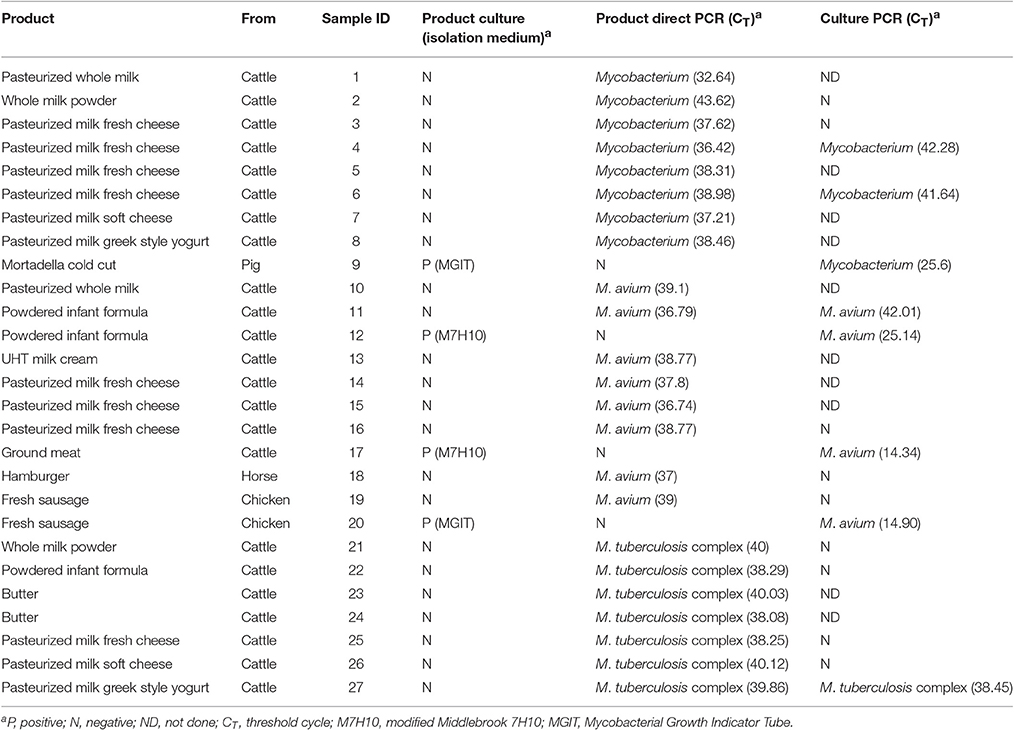
Table 2. Culture and screening tetraplex real-time PCR results of positive samples (detection of M. avium, M. tuberculosis complex and genus Mycobacterium).
Identification of isolates is shown in Table 3. M. avium subsp. hominisssuis was isolated from a powdered infant formula (one colony on 7H10) as well as from one ground beef sample (one colony on 7H10). M. avium subsp. avium and M. fortuitum complex grew in MGIT cultures from a fresh chicken sausage and one mortadella sample, respectively. The ITS sequence obtained for the M. fortuitum complex isolate displayed a homology of 99% with M. senegalense strain MF-417 (82% query coverage) in BLAST analysis. No viable bacteria belonging to the M. tuberculosis complex were isolated.
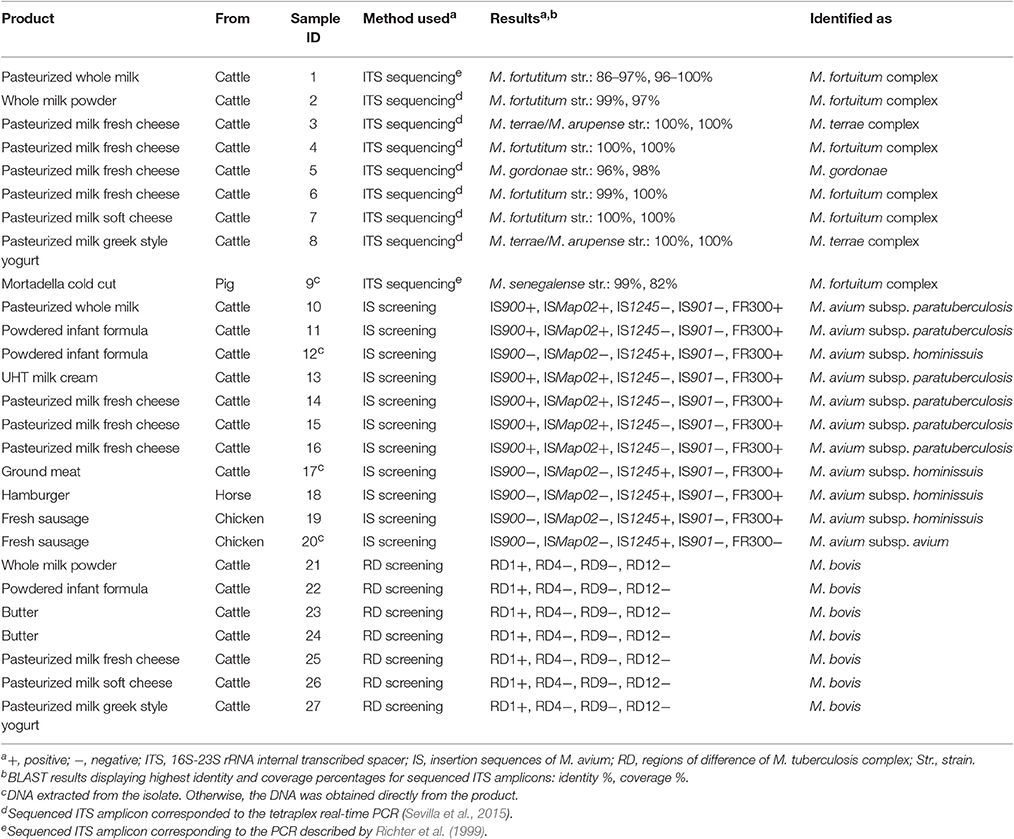
Table 3. Confirmation and identification of samples positive to either culture or screening tetraplex real-time PCR.
Direct PCR Detection in Products and Identification of Positive Samples
Mycobacterial DNA was detected in a total of 23 samples that accounted for the 15.22% of dairy products and 1.68% of meat products analyzed (Table 2). According to the tetraplex PCR results, 34.78% of positives corresponded to M. avium, 30.43% to M. tuberculosis complex and the remaining 34.78% to mycobacteria other than M. avium and M. tuberculosis complex. In meat foods only M. avium was detected. All but one product PCR positive samples displayed CT values above 36 indicating the presence of very low amounts of detectable mycobacterial DNA. Regardless of these high CT values, all positive results could be confirmed by the repetition of the screening PCR as well as by PCR testing of M. avium insertion sequences or M. tuberculosis complex RDs and mycobacterial ITS amplicon sequencing (see Table 3).
In this survey M. tuberculosis complex DNA was identified only in dairy products, including whole milk powder, infant formula, butter, cheese and yogurt. Samples recorded as M. tuberculosis complex positive were inspected for the presence of RD 1, 4, 9, and 12, and only RD1 was detected in all cases. This signature was suggestive of the presence of M. bovis. Spoligotyping failed to confirm this due to the absence of any perceptible hybridization signal.
DNA from M. avium subsp. paratuberculosis and M. avium subsp. hominissuis was identified in 6 dairy foods (pasteurized whole milk, infant formula, cream and fresh cheese) and 2 meat products (hamburger and sausage), as assessed by the presence or absence of the insertion sequences investigated. Samples deemed Mycobacterium sp. positive in the tetraplex PCR were finally classified as mycobacteria compatible with M. fortuitum complex (5), M. terrae complex (2) and M. gordonae (1) according to the sequences obtained.
With regard to foods represented by two product batches, only two displayed a positive result for both. However, the mycobacteria identified differed between batches. One powdered infant formula resulted positive to M. tuberculosis complex (M. bovis) in the first sampling and positive to M. avium (subsp. paratuberculosis) in the second. The other product was a fresh chicken sausage with batches positive to M. avium subsp. hominissuis DNA in one case and to M. avium subsp. avium culture in the other.
Discussion
Direct PCR revealed the presence of mycobacteria in 9% of products, a proportion increased in dairy foodstuffs (15%) if compared with meat products (2%). All samples displayed very high CT values close to or above those obtained at the limit of detection of the unmodified technique with artificially inoculated samples (100 CFU per gram) (Sevilla et al., 2015). The different nature of samples as well as the use of the modified method might account for a slightly different association between CT values and limit of detection. In spite of this, the high CT values were not PCR artifacts as was demonstrated by the confirmatory tests performed and therefore, low bacterial concentrations were to be expected in samples. The lack of agreement observed between PCR and culture can be explained by the heterogeneous distribution and low concentration of viable mycobacteria (probably below the limit of detection of PCR) in the products and the fact that DNA detection does not necessarily imply viability. Other factors that could account for PCR negative but culture positive results include a lower sensitivity of the DNA extraction and detection method or the presence of cell clumps.
M. avium subsp. paratuberculosis DNA was present in 4% of dairy foods. This figure is consistent with the lowest percentages of positives reported elsewhere (Grant, 2010; Waddell et al., 2016). In spite of this, we found less M. avium subsp. paratuberculosis than we anticipated based on the high prevalence and spread of PTB. In comparison to other reports on the culture of viable M. avium subsp. paratuberculosis from raw and pasteurized milk products (Waddell et al., 2016), culture did not produce any isolates. Our survey did not include any raw milk products and the number of pasteurized liquid milk samples was reduced. The sensitivity of DNA extraction and culture could represent an additional limitation that may be improved using novel methods (Botsaris et al., 2016). Despite this, a colony of M. avium subsp. hominissuis was isolated from a powdered infant formula. Little is known about the prevalence of this subspecies in dairy products. Among other infections, it is responsible for cervical lymphadenitis in infants and the most likely route of infection is oral (Falkinham, 2003; Kasperbauer and Huitt, 2013). Water is considered the main source of human exposure to this microbe (Falkinham, 2015) but infant formula could also represent a way of contact for children.
On the other hand, the detection of M. tuberculosis complex DNA in 5% of dairy products was completely unexpected. All cases were identified as M. bovis by RD screening. Spoligotyping was negative presumably because very small amounts and proportion of target DNA was present in DNA specimens obtained from food products. DNA extracted from these complex matrices can harbor a high concentration of nontarget DNA and other elements that may interfere with spoligotyping. The performance of spoligotyping can be limited also when applied directly to clinical samples (Milián Suazo et al., 2010; Ereqat et al., 2013). Our results did not seem to reflect the current PTB and TB situation in developed countries with high PTB prevalence and ongoing TB eradication and surveillance programs (Lombard, 2011; Schiller et al., 2011). It can be speculated that contaminated milk or byproducts imported from regions with high TB burden was used to produce these foods. M. bovis has been detected in milk, cheese and other products (Kaneene et al., 2014; Pereira-Suarez et al., 2014; Pérez-Lago et al., 2014; Cezar et al., 2016), mainly in regions where animal TB still represents a major issue.
All samples positive only to Mycobacterium sp. DNA corresponded to dairy products. This could be related to a higher exposure of milk to environmental NTM due to the way it is collected, transported and processed prior to being commercialized. The mycobacterial DNA detected displayed sequences highly compatible with M. fortuitum complex, M. terrae complex, and M. gordonae. These NTM are only occasionally considered pathogenic (Griffith et al., 2007). M. fortuitum complex is the only rapid growing mycobacterial group identified in our survey. However, it was detected in 4% of dairy samples, including one pasteurized milk with the highest estimated bacterial concentration (CT = 32.64). The detection of these mycobacterial groups in unpasteurized milk is not unusual, but only few studies report on the isolation from pasteurized milk (Sgarioni et al., 2014), consistent with thermal inactivation profiles of M. fortuitum in milk (Grant et al., 1996).
PCR and further methods revealed M. avium subsp. hominissuis DNA in 2% of meat foodstuffs. Culture retrieved one M. avium subsp. avium isolate from a fresh chicken sausage, one M. avium subsp. hominissuis isolate from ground beef and one M. senegalense isolate from mortadella. These data diverged from other surveys on M. avium subspecies detection showing quite high proportions of PCR positives but no isolate recovery (Klanicova et al., 2011; Lorencova et al., 2014). The outcome of our market situation study may fit best with other reports (Jaravata et al., 2007; Savi et al., 2015). M. avium isolates were grown from uncooked meat products. M. senegalense strain was isolated from a cold cut originated from cooked pork meat and fat, suggesting an environmental contamination during further processing. M. avium subsp. avium is an obligate pathogen rarely isolated from environmental samples unrelated to birds (Turenne et al., 2008). The subspecies avium is the main cause of typical avian TB in domestic birds and can infect cattle, deer and wild boar, and humans more sporadically (Dvorska et al., 2004). Since these infections are more common in older animals, we find more likely that this sausage contained meat from culled laying hens instead of young broilers specifically bred for meat and deduce that improved control measures are desirable. Raw meats are generally intended for being consumed cooked, but eating undercooked meats is quite common and mycobacterial inactivation during cooking is time and temperature dependent (Hammer et al., 2013). In addition, cooked products can become cross-contaminated through direct or indirect contact with raw meat as a consequence of improper food handling. Meat could still pose a risk of exposure to viable M. avium subspecies and other mycobacteria if not produced hygienically and cooked properly (Klanicova-Zalewska and Slana, 2014).
Mycobacteria are present infecting animals and in our environment, water and food being likely means of contact for humans. Zoonotic TB and NTM infection have been of great importance in human health over time. The prevalence of zoonotic TB was dramatically reduced owing to the implementation of abattoir inspection, milk pasteurization and other measures, but the problem is still considerable in many regions (Kaneene et al., 2014). With respect to NTM, much effort has been devoted to the understanding of human infection and thus different mechanisms leading to susceptibility and predisposition consisting in immunological flaws and conditions that compromise physical barriers to infection have been identified (Lake et al., 2016). Exposure to attenuated or inactivated mycobacteria can modulate the immune response resulting in a favorable effect (Beltran-Beck et al., 2014; Cardona et al., 2015; Kaufmann et al., 2015; Chambers et al., 2017). On the other hand, genetic susceptibility to chronic inflammatory disorders coupled with heavy exposure to M. avium subsp. paratuberculosis has been pointed out as a potential etiological factor in Crohn's and other chronic inflammatory diseases (Sechi and Dow, 2015). Moreover, it has been hypothesized that it could drive an autoimmune response by molecular mimicry (Sechi and Dow, 2015). Unfortunately, a consensus opinion on the potential role of this bacterium in Crohn's disease has not yet been reached (Van Kruiningen, 2011; Waddell et al., 2016). Since few bacteria are detected in Crohn's disease patients it could be speculated that M. avium subsp. paratuberculosis might not need to be active to trigger a deleterious response. A latent, dormant or even dead condition could be sufficient as showed by the necrotizing colitis induced in mice after transanal injection of M. avium subsp. paratuberculosis antigens (Momotani et al., 2012). However, our results do not indicate that a heavy foodborne exposure occurs in populations where the main source of food of animal origin is processed food.
In summary, our results show that the prevalence of viable mycobacteria in packed products of animal origin available at Spanish supermarkets was not substantial although a considerable proportion of them contained mycobacterial DNA. These figures are probably translatable to other European countries as a result of market globalization and common food safety legislation. This survey indicates that viable mycobacteria and mycobacterial components are present in a range of products and at a frequency that could ensure repeated exposure over time during the lifetime of any individual. Consequently, setting up more stringent control measures should be considered. Further research is necessary to identify critical points in the food production system and determine the real impact of foodborne mycobacterial exposure on human health either directly, causing infection, or indirectly, modifying individual's immune status and susceptibility to other diseases.
Author Contributions
IS, RJ, and JG conceived of the study. EM, MT, IS, and NE carried out the laboratory work and compiled and analyzed the data. IS collated the results and wrote the original draft. IS, EM, MT, NE, RJ, and JG participated in the review and the editing of the original draft. All authors read and approved the final manuscript.
Funding
The work of IS, EM, MT, NE, and JG was supported by the Department of Economic Development and Competitiveness of the Basque Government. MT holds a fellowship from the same Department. We also acknowledge this Department and the Department of Education, Universities and Research of the Basque Government for funding Research Projects 32-2016-00030 and PI2011-50.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was included in the Food Safety Research Plan of the Basque Country (2011-2016) conceived by the Directorate for Food Quality Control and Industries (Department of Economic Development and Competitiveness) of the Basque Government.
References
Bartos, M., Hlozek, P., Svastova, P., Dvorska, L., Bull, T., Matlova, L., et al. (2006). Identification of members of Mycobacterium avium species by Accu-Probes, serotyping, and single IS900, IS901, IS1245 and IS901-flanking region PCR with internal standards. J. Microbiol. Methods 64, 333–345. doi: 10.1016/j.mimet.2005.05.009
Beltran-Beck, B., de la Fuente, J., Garrido, J. M., Aranaz, A., Sevilla, I., Villar, M., et al. (2014). Oral vaccination with heat inactivated Mycobacterium bovis activates the complement system to protect against tuberculosis. PLoS ONE 9:e98048. doi: 10.1371/journal.pone.0098048
Bercovier, H., and Vincent, V. (2001). Mycobacterial infections in domestic and wild animals due to Mycobacterium marinum, M. fortuitum, M. chelonae, M. porcinum, M. farcinogenes, M. smegmatis, M. scrofulaceum, M. xenopi, M. kansasii, M. simiae and M. genavense. Rev. Sci. Tech. 20, 265–290. doi: 10.20506/rst.20.1.1269
Biet, F., and Boschiroli, M. L. (2014). Non-tuberculous mycobacterial infections of veterinary relevance. Res. Vet. Sci. 97, S69–S77. doi: 10.1016/j.rvsc.2014.08.007
Botsaris, G., Swift, B. M., Slana, I., Liapi, M., Christodoulou, M., Hatzitofi, M., et al. (2016). Detection of viable Mycobacterium avium subspecies paratuberculosis in powdered infant formula by phage-PCR and confirmed by culture. Int. J. Food Microbiol. 216, 91–94. doi: 10.1016/j.ijfoodmicro.2015.09.011
Cardona, P., Marzo-Escartin, E., Tapia, G., Diaz, J., Garcia, V., Varela, I., et al. (2015). Oral administration of heat-killed Mycobacterium manresensis delays progression toward active tuberculosis in C3HeB/FeJ mice. Front. Microbiol. 6:1482. doi: 10.3389/fmicb.2015.01482
Cezar, R. D., Lucena-Silva, N., Borges, J. M., Santana, V. L., and Pinheiro Junior, J. W. (2016). Detection of Mycobacterium bovis in artisanal cheese in the state of Pernambuco, Brazil. Int. J. Mycobacteriol. 5, 269–272. doi: 10.1016/j.ijmyco.2016.04.007
Chambers, M. A., Aldwell, F., Williams, G. A., Palmer, S., Gowtage, S., Ashford, R., et al. (2017). The effect of oral vaccination with Mycobacterium bovis BCG on the development of tuberculosis in captive European badgers (Meles meles). Front. Cell Infect. Microbiol. 7:6. doi: 10.3389/fcimb.2017.00006
Donaghy, J. A., Johnston, J., and Rowe, M. T. (2011). Detection of Mycobacterium avium ssp. paratuberculosis in cheese, milk powder and milk using IS900 and f57-based qPCR assays. J. Appl. Microbiol. 110, 479–489. doi: 10.1111/j.1365-2672.2010.04905.x
Donaghy, J. A., Rowe, M. T., Rademaker, J. L., Hammer, P., Herman, L., De Jonghe, J. V., et al. (2008). An inter-laboratory ring trial for the detection and isolation of Mycobacterium avium subsp. paratuberculosis from raw milk artificially contaminated with naturally infected faeces. Food Microbiol. 25, 128–135. doi: 10.1016/j.fm.2007.06.007
Dvorska, L., Matlova, L., Bartos, M., Parmova, I., Bartl, J., Svastova, P., et al. (2004). Study of Mycobacterium avium complex strains isolated from cattle in the Czech Republic between 1996 and 2000. Vet. Microbiol. 99, 239–250. doi: 10.1016/j.vetmic.2004.01.008
Ereqat, S., Nasereddin, A., Levine, H., Azmi, K., Al-Jawabreh, A., Greenblatt, C. L., et al. (2013). First-time detection of Mycobacterium bovis in livestock tissues and milk in the West Bank, Palestinian Territories. PLoS Negl. Trop. Dis. 7:e2417. doi: 10.1371/journal.pntd.0002417
Falkinham, J. O. III. (2016). Current epidemiologic trends of the nontuberculous Mycobacteria (NTM). Curr. Environ. Health Rep. 3, 161–167. doi: 10.1007/s40572-016-0086-z
Falkinham, J. O. (2003). The changing pattern of nontuberculous mycobacterial disease. Can. J. Infect. Dis. 14, 281–286. doi: 10.1155/2003/323058
Falkinham, J. O. III. (2015). Environmental sources of nontuberculous mycobacteria. Clin. Chest Med. 36, 35–41. doi: 10.1016/j.ccm.2014.10.003
Faria, A. C., Schwarz, D. G., Carvalho, I. A., Rocha, B. B., De Carvalho Castro, K. N., Silva, M. R., et al. (2014). Short communication: viable Mycobacterium avium subspecies paratuberculosis in retail artisanal Coalho cheese from Northeastern Brazil. J. Dairy Sci. 97, 4111–4114. doi: 10.3168/jds.2013-7835
García-Martos, P., and García-Agudo, L. (2012). Infections due to rapidly growing mycobacteria. Enferm. Infecc. Microbiol. Clin. 30, 192–200. doi: 10.1016/j.eimc.2011.09.017
Ghodbane, R., Mba, M. F., Lepidi, H., Nappez, C., and Drancourt, M. (2014). Long-term survival of tuberculosis complex mycobacteria in soil. Microbiology 160, 496–501. doi: 10.1099/mic.0.073379-0
Grant, I. R. (2010). “Mycobacterium avium subsp. paratuberculosis in animal-derived foods and the environment,” in Paratuberculosis: Organism, Disease, Control, eds M. A. Behr and D. M. Collins (Oxfordshire: CABI), 29–39.
Grant, I. R., Ball, H. J., and Rowe, M. T. (1996). Thermal inactivation of several Mycobacterium spp. in milk by pasteurization. Lett. Appl. Microbiol. 22, 253–256. doi: 10.1111/j.1472-765X.1996.tb01154.x
Griffith, D. E., Aksamit, T., Brown-Elliott, B. A., Catanzaro, A., Daley, C., Gordin, F., et al. (2007). An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 175, 367–416. doi: 10.1164/rccm.200604-571ST
Halse, T. A., Escuyer, V. E., and Musser, K. A. (2011). Evaluation of a single-tube multiplex real-time PCR for differentiation of members of the Mycobacterium tuberculosis complex in clinical specimens. J. Clin. Microbiol. 49, 262–2567. doi: 10.1128/JCM.00467-11
Hammer, P., Walte, H. G., Matzen, S., Hensel, J., and Kiesner, C. (2013). Inactivation of Mycobacterium avium subsp. paratuberculosis during cooking of hamburger patties. J. Food Prot. 76, 1194–1201. doi: 10.4315/0362-028X.JFP-12-474
Jaravata, C. V., Smith, W. L., Rensen, G. J., Ruzante, J., and Cullor, J. S. (2007). Survey of ground beef for the detection of Mycobacterium avium paratuberculosis. Foodborne. Pathog. Dis. 4, 103–106. doi: 10.1089/fpd.2006.54
Kamerbeek, J., Schouls, L., Kolk, A., van Agterveld, M., van Soolingen, D., Kuijper, S., et al. (1997). Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J. Clin. Microbiol. 35, 907–914.
Kaneene, J. B., Miller, R., Steele, J. H., and Thoen, C. O. (2014). Preventing and controlling zoonotic tuberculosis: a One Health approach. Vet. Ital. 50, 7–22. doi: 10.12834/VetIt.1302.08
Kasperbauer, S., and Huitt, G. (2013). Management of extrapulmonary nontuberculous mycobacterial infections. Semin. Respir. Crit Care Med. 34, 143–150. doi: 10.1055/s-0033-1333576
Kaufmann, S. H. E., Bloom, B., Brosch, R., Cardona, P. J., Dockrell, H., Fritzell, B., et al. (2015). Developing whole mycobacteria cell vaccines for tuberculosis: workshop proceedings, Max Planck Institute for Infection Biology, Berlin, Germany, July 9, 2014. Vaccine 33, 3047–3055. doi: 10.1016/j.vaccine.2015.03.056
Klanicova, B., Slana, I., Vondruskova, H., Kaevska, M., and Pavlik, I. (2011). Real-time quantitative PCR detection of Mycobacterium avium subspecies in meat products. J. Food Prot. 74, 636–640. doi: 10.4315/0362-028X.JFP-10-332
Klanicova-Zalewska, B., and Slana, I. (2014). Presence and persistence of Mycobacterium avium and other nontuberculous mycobacteria in animal tissues and derived foods: a review. Meat Sci. 98, 835–841. doi: 10.1016/j.meatsci.2014.08.001
Lake, M. A., Ambrose, L. R., Lipman, M. C., and Lowe, D. M. (2016). “Why me, why now?” Using clinical immunology and epidemiology to explain who gets nontuberculous mycobacterial infection. BMC Med. 14:54. doi: 10.1186/s12916-016-0606-6
Lombard, J. E. (2011). Epidemiology and economics of paratuberculosis. Vet. Clin. North Am. Food Anim Pract. 27, 525–535. doi: 10.1016/j.cvfa.2011.07.012
Lorencova, A., Vasickova, P., Makovcova, J., and Slana, I. (2014). Presence of Mycobacterium avium subspecies and hepatitis E virus in raw meat products. J. Food Prot. 77, 335–338. doi: 10.4315/0362-028X.JFP-13-252
Milián Suazo, F., Harris, B., Arriaga-Díaz, C., Thomsen, B., Stuber, T., González-Suárez, D., et al. (2010). Sensitivity and specificity of nested PCR and spoligotyping as quick diagnostic tests for bovine tuberculosis in fresh tissue. Rev. Mex. Cienc. Pecu. 1, 403–415. doi: 10.22319/rmcpv%251494. Available online at: http://cienciaspecuarias.inifap.gob.mx/index.php/Pecuarias/article/view/1494
Momotani, E., Ozaki, H., Hori, M., Yamamoto, S., Kuribayashi, T., Eda, S., et al. (2012). Mycobacterium avium subsp. paratuberculosis lipophilic antigen causes Crohn's disease-type necrotizing colitis in mice. Springerplus 1:47. doi: 10.1186/2193-1801-1-47
Pereira-Suarez, A. L., Estrada-Chavez, Y., Zuniga-Estrada, A., Lopez-Rincon, G., Hernandez, D. U., Padilla-Ramirez, F. J., et al. (2014). Detection of Mycobacterium tuberculosis complex by PCR in fresh cheese from local markets in Hidalgo, Mexico. J. Food Prot. 77, 849–852. doi: 10.4315/0362-028X.JFP-13-389
Pérez-Lago, L., Navarro, Y., and García-de-Viedma, D. (2014). Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis: a review. Res. Vet. Sci. 97, S94–S100. doi: 10.1016/j.rvsc.2013.11.008
Richter, E., Niemann, S., Rusch-Gerdes, S., and Hoffner, S. (1999). Identification of Mycobacterium kansasii by using a DNA probe (AccuProbe) and molecular techniques. J. Clin. Microbiol. 37, 964–970.
Savi, R., Ricchi, M., Cammi, G., Garbarino, C., Leo, S., Pongolini, S., et al. (2015). Survey on the presence of Mycobacterium avium subsp. paratuberculosis in ground beef from an industrial meat plant. Vet. Microbiol. 177, 403–408. doi: 10.1016/j.vetmic.2015.03.013
Schiller, I., Waters, W. R., Vordermeier, H. M., Jemmi, T., Welsh, M., Keck, N., et al. (2011). Bovine tuberculosis in Europe from the perspective of an officially tuberculosis free country: trade, surveillance and diagnostics. Vet. Microbiol. 151, 153–159. doi: 10.1016/j.vetmic.2011.02.039
Sechi, L. A., and Dow, C. T. (2015). Mycobacterium avium ss. paratuberculosis Zoonosis - the hundred year war - beyond crohn's disease. Front Immunol. 6:96. doi: 10.3389/fimmu.2015.00096
Sevilla, I. A., Garrido, J. M., Molina, E., Geijo, M. V., Elguezabal, N., Vázquez, P., et al. (2014). Development and evaluation of a novel multicopy-element-targeting triplex PCR for detection of Mycobacterium avium subsp. paratuberculosis in feces. Appl. Environ. Microbiol. 80, 3757–3768. doi: 10.1128/AEM.01026-14
Sevilla, I. A., Molina, E., Elguezabal, N., Perez, V., Garrido, J. M., and Juste, R. A. (2015). Detection of mycobacteria, Mycobacterium avium subspecies, and Mycobacterium tuberculosis complex by a novel tetraplex real-time PCR assay. J. Clin. Microbiol. 53, 930–940. doi: 10.1128/JCM.03168-14
Sgarioni, S. A., Hirata, R. D., Hirata, M. H., Leite, C. Q., de Prince, K. A., de Andrade Leite, S. R., et al. (2014). Occurrence of Mycobacterium bovis and non-tuberculous mycobacteria (NTM) in raw and pasteurized milk in the northwestern region of Parana, Brazil. Braz. J. Microbiol. 45, 707–711. doi: 10.1590/S1517-83822014000200046
Slana, I., Kaevska, M., Kralik, P., Horvathova, A., and Pavlik, I. (2010). Distribution of Mycobacterium avium subsp. avium and M. a. hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR. Vet. Microbiol. 144, 437–443. doi: 10.1016/j.vetmic.2010.02.024
Tell, L. A., Woods, L., and Cromie, R. L. (2001). Mycobacteriosis in birds. Rev. Sci. Tech. 20, 180–203. doi: 10.20506/rst.20.1.1273
Thomson, R., Tolson, C., Sidjabat, H., Huygens, F., and Hargreaves, M. (2013). Mycobacterium abscessus isolated from municipal water - a potential source of human infection. BMC Infect. Dis. 13:241. doi: 10.1186/1471-2334-13-241
Turenne, C. Y., Collins, D. M., Alexander, D. C., and Behr, M. A. (2008). Mycobacterium avium subsp. paratuberculosis and M. avium subsp. avium are independently evolved pathogenic clones of a much broader group of M. avium organisms. J. Bacteriol. 190, 2479–2487. doi: 10.1128/JB.01691-07
Van Kruiningen, H. J. (2011). Where are the weapons of mass destruction - the Mycobacterium paratuberculosis in Crohn's disease? J. Crohns. Colitis. 5, 638–644. doi: 10.1016/j.crohns.2011.08.004
Waddell, L., Rajic, A., Stark, K., and McEwen, S. A. (2016). Mycobacterium avium ssp. paratuberculosis detection in animals, food, water and other sources or vehicles of human exposure: a scoping review of the existing evidence. Prev. Vet. Med. 132, 32–48. doi: 10.1016/j.prevetmed.2016.08.003
Keywords: nontuberculous mycobacteria, Mycobacterium tuberculosis complex, dairy products, meat products, food contamination, prevalence
Citation: Sevilla IA, Molina E, Tello M, Elguezabal N, Juste RA and Garrido JM (2017) Detection of Mycobacteria by Culture and DNA-Based Methods in Animal-Derived Food Products Purchased at Spanish Supermarkets. Front. Microbiol. 8:1030. doi: 10.3389/fmicb.2017.01030
Received: 07 March 2017; Accepted: 23 May 2017;
Published: 09 June 2017.
Edited by:
Giovanna Suzzi, University of Teramo, ItalyReviewed by:
Benjamin Michael Connor Swift, University of Nottingham, United KingdomNigel Cook, Jorvik Food and Environmental Virology Ltd., United Kingdom
Iva Slana, Veterinary Research Institute, Czechia
Copyright © 2017 Sevilla, Molina, Tello, Elguezabal, Juste and Garrido. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Iker A. Sevilla, isevilla@neiker.eus