 Aline Moser
Aline Moser Daniel Wüthrich
Daniel Wüthrich Rémy Bruggmann
Rémy Bruggmann Elisabeth Eugster-Meier
Elisabeth Eugster-Meier Leo Meile
Leo Meile Stefan Irmler
Stefan Irmler- 1Agroscope, Bern, Switzerland
- 2Laboratory of Food Biotechnology, Institute of Food, Nutrition and Health, ETH Zurich, Zurich, Switzerland
- 3Interfaculty Bioinformatics Unit, University of Bern and Swiss Institute of Bioinformatics, Bern, Switzerland
- 4School of Agricultural, Forest and Food Sciences HAFL, Bern University of Applied Sciences, Zollikofen, Switzerland
The advent of massive parallel sequencing technologies has opened up possibilities for the study of the bacterial diversity of ecosystems without the need for enrichment or single strain isolation. By exploiting 78 genome data-sets from Lactobacillus helveticus strains, we found that the slpH locus that encodes a putative surface layer protein displays sufficient genetic heterogeneity to be a suitable target for strain typing. Based on high-throughput slpH gene sequencing and the detection of single-base DNA sequence variations, we established a culture-independent method to assess the biodiversity of the L. helveticus strains present in fermented dairy food. When we applied the method to study the L. helveticus strain composition in 15 natural whey cultures (NWCs) that were collected at different Gruyère, a protected designation of origin (PDO) production facilities, we detected a total of 10 sequence types (STs). In addition, we monitored the development of a three-strain mix in raclette cheese for 17 weeks.
Introduction
Lactobacillus helveticus belongs to the group of lactic acid bacteria (LAB) that are characterized by their ability to produce lactic acid through metabolizing lactose and other carbohydrates (Hammes and Hertel, 2006). A previous study has shown that L. helveticus is one of the predominant species in the natural whey cultures (NWCs) that are used for the production of Gruyère, a protected designation of origin (PDO) cheese (Moser et al., 2017). Gruyère PDO, produced in specific regions of Switzerland, is a smear-ripened, hard-textured cheese made from raw cow's milk (Eugster-Meier et al., 2017). It is likely that L. helveticus plays an important role in this cheese's ripening process, as has been shown for other cheese varieties produced using NWCs (Gatti et al., 2014). Desirable effects in cheese production, such as faster ripening and enhanced flavor development, have been associated with specific strains of L. helveticus (Drake et al., 1997; Jensen M. P. et al., 2009; Jensen and Ardö, 2010). In addition, biotechnologically important characteristics can differ considerably between different strains of L. helveticus (Fortina et al., 1998). Due to the biotechnological importance of this species, it is an advantage to differentiate and characterize the various strains, as this allows for a better understanding of the functional and ecological significance of this species.
Several molecular typing methods, such as randomly amplified polymorphic DNA (RAPD) PCR, repetitive sequence PCR (rep-PCR), pulsed field gel electrophoresis (PFGE), ribotyping, and multilocus sequence typing (MLST) have been used in the past to examine the diversity of L. helveticus isolates (Giraffa et al., 1998; Jenkins et al., 2002; Jensen M. et al., 2009; Sun et al., 2015). All these techniques require cultivation. This can lead to a skewed microbial profile, as the culture medium and incubation could favor strains that are well adapted to these cultivation conditions. We aimed to deploy a culture-independent approach in the analysis to learn more about the population diversity and dynamics of L. helveticus strains in a dairy ecosystem; to our knowledge, such a method has not yet been developed for these strains.
In this study, we report on a culture-independent PCR-based method for the strain typing of L. helveticus. By exploiting the genomic sequence data of L. helveticus strains, a single copy gene of the core genome that encodes a surface layer protein was found to display high genetic heterogeneity and could be used for strain typing. We established an amplicon based high-throughput sequencing method to assess the composition of L. helveticus strains in NWCs and to monitor the development of L. helveticus strains during cheese ripening. The findings clearly show that the method is a useful tool to assess L. helveticus diversity in various habitats.
Materials and Methods
DNA Preparation
Genomic DNA (gDNA) was extracted from bacterial cultures, NWCs, and cheese samples as described by Moser et al. (2017). DNA quantity was determined fluorometrically using the Qubit dsDNA BR Assay Kit (Thermo Fisher Scientific, Baar, Switzerland).
Target Detection
The genomes of 57 L. helveticus strains from the Agroscope culture collection and that of one L. helveticus strain from the Direct Vat Set culture LH-32 from Christian Hansen (Copenhagen, Denmark) were sequenced (Supplemental Table S1). The “TruSeq DNA PCR-Free LT Library Prep” (FC-121-3003, Insert size option: 350 bp) was used to prepare the DNA libraries. The libraries were indexed and pooled, before being sequenced in one lane on an Illumina HiSeq 3000 instrument to produce paired-end reads (151 × 151). Trimmomatic (version 0.33, options: SLIDINGWINDOW:4:8 MINLEN:127; Bolger et al., 2014) was used to trim the raw reads. The remaining reads were assembled using SPAdes (version 3.6.1, options: —careful—mismatch-correction—k 21, 33, 55, 77, 99, 127) and the assembly was scaffolded using SSPACE (version 3.0, default options; Boetzer et al., 2011). The scaffolds were selected for length and coverage as follows: scaffolds that were shorter than 200 bp, and those with a lower median coverage than 20% of the median read-depth of all scaffolds larger than 5,000 bp, were excluded. The resulting assemblies were annotated using Prokka (version 1.11; Seemann, 2014). To determine the orthologous gene clusters (OGCs) between the genomes, all predicted protein-coding sequences were translated and compared using BLASTP (version 2.2.29+, default parameters; Altschul et al., 1990) and clustered using Ortho-MCL (version 2.0.9, default parameters; Li et al., 2003). Single copy OGCs that were present in all genomes were ranked according to the number of unique sequences per OGC. Afterwards the corresponding nucleotide sequences were extracted and aligned using Clustal Omega (version 1.2.1). The alignments were then inspected and not aligned sequences were removed manually using the CLC Main Workbench software (Version 7.5.1, Qiagen, Switzerland). Finally, the remaining aligned sequences were checked by eye for nucleotide polymorphisms. Thereby, a CDS encoding a surface layer protein, named slpH in this report, was found to carry a high amount of polymorphisms and could be used as a target for DNA-based strain typing.
For the assignment of sequence types (STs), additional orthologous genes were retrieved from the NCBI GenBank database. Based on the nucleotide polymorphism present in these genes, the nucleotide sequences were assigned to STs (Table 1). SlpH genes with newly identified polymorphisms were deposited in the NCBI database and their accession numbers are listed in Table 1. The sequences of all of the different slpH genes are presented in the nucleotide sequence alignment in Figure S1. The nucleotide sequences of the ORFs from all STs were translated using the CLC Main Workbench version 7.5.1 (CLC bio, Aarhus, Denmark) and the resulting amino acid sequences were analyzed using InterProScan (Jones et al., 2014). Furthermore, the genomic context of the slpH gene was analyzed in 10 complete L. helveticus genomes from the NCBI database (CP002081, CP000517, CP003799, CP002429, CP002427, CP009907, CP011386, CP012381, CP016827, and CP020029). Therefore, the genomes were reannotated using Prokka (version 1.11; Seemann, 2014) and the orthologous genes were determined using Ortho-MCL (Li et al., 2003). The slpH genes and the surrounding genes of all of the 10 strains were visualized using the package ggplot2 in R (Wickham, 2009).
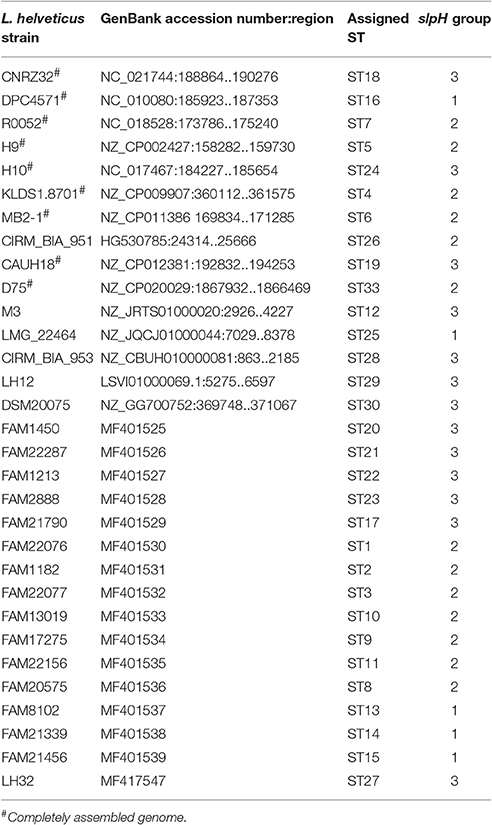
Table 1. slpH genes used in this study and their assignment to sequence types (STs).
Amplification of the slpH Locus
The slpH sequence from L. helveticus CNRZ32 (Table 1) served as reference sequence for primer design. Based on the reference sequence, the primer pair LHslpF (5′-CAAGGAGGAAAGACCACATGA-3′) and LHslpR (5′-TGTACTTGCCAGTTGCCTTG-3′) that amplifies a 1,116-bp region was designed. Primers were designed using Primer3 (version 0.4.0; Untergasser et al., 2012). PCR was carried out on a Veriti® Thermal Cycler (Thermo Fisher Scientific, Baar, Switzerland) with the following conditions: 95°C for 2 min followed by 30 cycles of 95°C for 20 s, 60°C for 10 s, and 70°C for 30 s, and a final extension of 70°C for 7 min. If not otherwise specified, each PCR (25 μL) contained 50 ng of gDNA for cheese or NWC samples or 1 ng of gDNA for pure cultures, 5 pmol of each primer, 5 nmol of each dNTP, 31.25 nmol Mg2SO4, 0.02 U of KOD Hot Start Polymerase (Merck), and 1X buffer for KOD Hot Start Polymerase. PCR products were analyzed using the Agilent DNA 7500 kit on an Agilent 2100 bioanalyzer (Agilent Technologies, Waldbronn, Germany).
Culture-Dependent Typing
An NWC sample (100 μL, sample p in Table 2) was plated on modified MRS plates containing 20 g L−1 of lactose instead of glucose (De Man et al., 1960). After incubation at 42°C for 48 h, 96 colonies were picked from the agar plates and suspended individually in 100 μL of TE buffer (pH 8.0) containing 10 mM Tris-HCl and 10 mM EDTA. A heat treatment (100°C, 8 min) was used to extract gDNA. After the heat treatment, the suspensions with the lysed cells were centrifuged at 5,000 g for 10 min. The supernatant containing the DNA was diluted tenfold in 10 mM Tris-HCl (pH 8.0) and examined by quantitative real-time PCR (qPCR) to identify colonies of L. helveticus. DNA from L. helveticus positive colonies was further examined by slpH PCR as described above. The PCR products were sent to Microsynth (Balgach, Switzerland) for amplicon purification and Sanger sequencing.
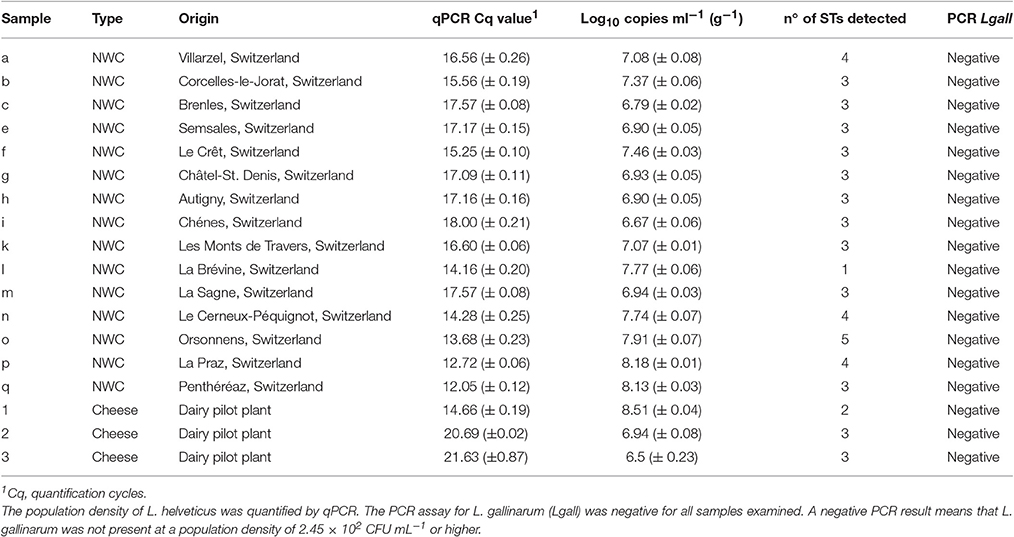
Table 2. Natural whey culture (NWC) and cheese samples used for slpH amplicon sequencing.
PCR Assays for L. helveticus and L. gallinarum Species Confirmation
Identification of L. helveticus isolates and quantitation of the population density of L. helveticus representatives in NWCs and cheese was performed by a qPCR method targeting the single copy pheS gene that encodes the alpha-subunit of the phenylalanyl-tRNA synthetase as described elsewhere (Moser et al., 2017). Briefly, a plasmid containing the target sequence is used for absolute quantification. The equation of the standard curve was used to estimate the copy number for cheese (gene equivalent per g) and NWCs (gene equivalent per mL). Similarly, the pheS gene of Lactobacillus gallinarum was used as a target to develop a PCR-based detection method for this species. Based on the pheS nucleotide sequence from L. gallinarum DSM 10532 (NCBI GenBank Accession Number AZEL01000044:72250…73299), the primers 5′-TCAGGACCTTGTACTACCTTGTAA-3′ and 5′-TGCTACTAAGGCTGAAATCGT-3′ were designed, which enable amplification of a 180-bp fragment. The assay was used to analyze NWC samples for the absence or presence of L. gallinarum. PCR assays were conducted in a final volume of 25 μL containing 300 nM of each primer, 200 μM of each dNTP, 1.5 mM Mg2SO4, 0.02 U of KOD Hot Start Polymerase (Merck), and 1X buffer for KOD Hot Start Polymerase (Merck). Each PCR contained 50 ng of genomic DNA for NWC samples and 1 ng of DNA for pure cultures. The amplicons were amplified under the following conditions: 95°C for 2 min, followed by 30 cycles of 95°C for 20 s, 59°C for 10 s, and 70°C for 4 s, and a final extension of 70°C for 5 min. The PCR products were examined with an Agilent DNA 1000 kit on an Agilent 2,100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany). To evaluate the lower limit of detection, DNA was extracted from a ten-fold dilution series of a pure culture from L. gallinarum DSM 10532 that had been grown overnight in MRS broth at 37°C and for which the population density had been determined by plate-counting.
slpH Gene Semiconductor Sequencing
After slpH-specific PCR, amplicons were purified using the Qiaquick PCR Purification kit (Qiagen, Hombrechtikon, Switzerland). Purified amplicons were fragmented by sonication of 1 μg of DNA in 130 μL nuclease-free water at 50 W, 200 cycles/burst, and 20°C for 90 s on a Covaris M220 instrument (Covaris, Brighton, U.K.). The fragmented DNA was end-repaired and adapter-ligated using the Ion Xpress Plus Fragment Library Kit (Thermo Fisher Scientific) according to the manufacturer's instruction. Samples were barcoded using the Ion Xpress Barcode Adapter 1-16 kit (Thermo Fisher Scientific). After barcoding, the DNA was size-selected for 400 bp reads using an E-Gel® SizeSelect™ Agarose Gel (Thermo Fisher Scientific). The libraries were quantified by qPCR using the Ion Library Quantitation Kit (Thermo Fisher Scientific). Before preparing the template-positive ion sphere particles with the Ion PGM Hi-Q OT2 kit (Thermo Fisher Scientific), each library was diluted to 100 pM in 10 mM Tris-HCl (pH 8.0) and 0.1 mM EDTA, before being pooled. The sequencing was performed using either an Ion 314™ or 316™ chip and the Ion PGM Hi-Q Sequencing kit on an Ion Torrent sequencer (Thermo Fisher Scientific).
Sequence Data Analysis
First, fastq files were generated via the Torrent Suite Software (Thermo Fisher Scientific) using the default parameters; then, the reads were quality trimmed with Trimmomatic (version 0.36, options: SLIDINGWINDOW:4:20 MINLEN:101; Bolger et al., 2014). Finally, reads containing the identifying subsequences for slpH group1 (GGCTACACT, GATCAATTAA, AGTGTAGCC, and TTAATTGATC), for slpH group2 (CCTTAATGTA, CTGACGATGT, TACATTAAGG, and ACATCGTCAG), and for slpH group 3 (ATTGGTTCAG, GGTGTTGCTA, CTGAACCAAT, and TAGCAACACC) were extracted, trimmed, grouped based on 100% sequence identity, and mapped against a reference database containing known STs. Sequences that were not identical to one of the reference sequences were assigned to a new ST. For this procedure, a Python script was developed. The script, its usage and the database for the slpH STs are available at: https://github.com/danielwuethrich87/helveticus_strain_typing.
Validation
The gDNA from 10 different L. helveticus STs (Table 3) were pooled in equal amounts. The amplicons obtained using the slpH-specific PCR were subsequently sequenced on the Ion Torrent sequencer as described above. The experiment was repeated three times.
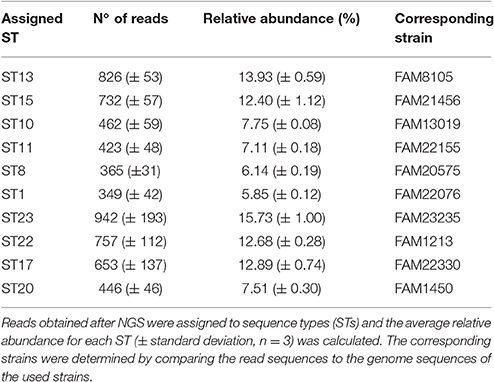
Table 3. The developed strain typing approach was tested with a mixture of 10 L. helveticus strains.
Results
Characteristics of the slpH Locus
We manually inspected the single copy orthologs of 58 own-sequenced genomes of L. helveticus. One of the gene clusters encoding a putative surface layer protein, named slpH in this study, stood out from the others in terms of nucleotide sequence diversity and differentiated 15 STs. Other OGCs differentiated between 8 and 11 STs and were excluded from further analysis. The primer pair LHslpF/R was designed to amplify the slpH locus in L. helveticus. The specificity of the primer pair was tested using the gDNA of 18 L. helveticus strains and 20 other species of LAB (Table 4). By using the primer pair LHslpF/R, an amplicon with a size of approximately 1,252 bp (±60 bp) was obtained for all the L. helveticus and L. gallinarum tested. No amplicon was observed using gDNA from Streptococcus thermophilus and other LAB, including the more closely related Lactobacillus kefiranofaciens kefiranofaciens DSM 5016, L. kefiranofaciens kefirgranum DSM 10550, Lactobacillus crispatus DSM 10532, Lactobacillus acidophilus DSM 20079, and Lactobacillus amylovorus DSM 20531 (Table 4).
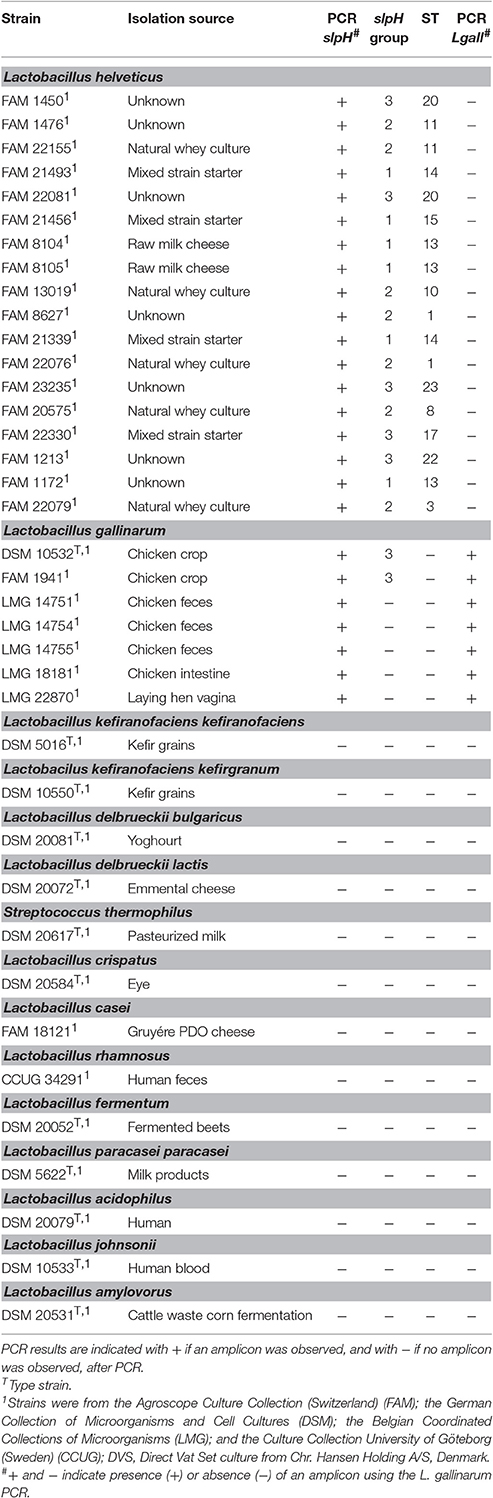
Table 4. Bacterial strains used for the PCR studies.
The nucleotide sequences of the L. helveticus amplicons were determined by Sanger sequencing and subsequently compared to slpH sequences that had been extracted from 79 bacterial genomes consisting of 58 genomes of own-sequenced strains and 21 from the NCBI GenBank. We found that the sequences were identical to the one derived from the illumina read assembly (data not shown) and that 30 STs could be unambiguously discriminated (Supplemental Figure S1). Additionally, the amplicon obtained from the L. gallinarum DSM 10532 was determined. A BLAST search revealed that the amplified region showed 99% identity to the lgsB gene (GenBank accession number AY597262) of L. gallinarum, which encodes a surface layer protein.
A pairwise sequence comparison of 30 STs showed that the slpH sequences from L. helveticus clustered into three groups (Figure 1). Strains that clustered within a group shared, on average, 91.4 (±4.6)% sequence identity, whereas the identity between groups was, on average, 57.1 (±1.7)%.
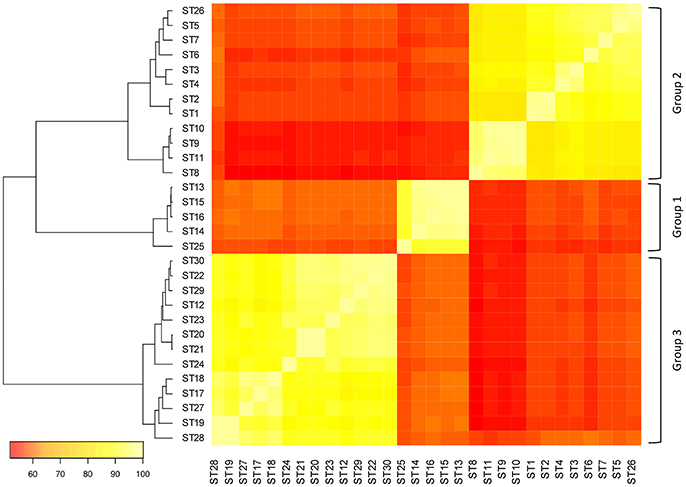
Figure 1. Pairwise comparison of the different sequence types (STs). The DNA sequences of the in silico derived amplicons from the different STs were compared to each other and the percent sequence identity was calculated for each pair. The color of a square between two STs indicates the percent sequence identity of the two STs. The color gradient ranges from white (100% sequence identity) to red (0% sequence identity).
When the deduced amino acid sequences were compared, the primary sequences of ST1 and ST2, ST18 and ST27, and ST19 and ST28, respectively, were identical. All other STs could still be discriminated based on amino acid sequence and especially a high level of variation was observed between the three slpH groups (Figure 2). When the primary sequences were analyzed for domains, InterProScan analysis assigned all STs but ST8, ST9, ST10, and ST11, which belong to the slpH2 group, to the Lactobacillus surface layer protein family (InterPro accession number PIRSF037863) using the PIRSF family classification system (not shown). Furthermore, InterProScan identified in all STs the eight motif fingerprints of Lactobacillus surface layer protein family (InterPro accession number PR01729) based on the PRINTS database for protein fingerprints (blue bars, Figure 2). Additionally, the analysis predicted the presence of a tandem SLAP domain (InterPro accession number IPR024968) in all STs (red bars, Figure 2). Finally, a signal peptide with a length of 30 amino acids was predicted for all the STs (green bars, Figure 2).
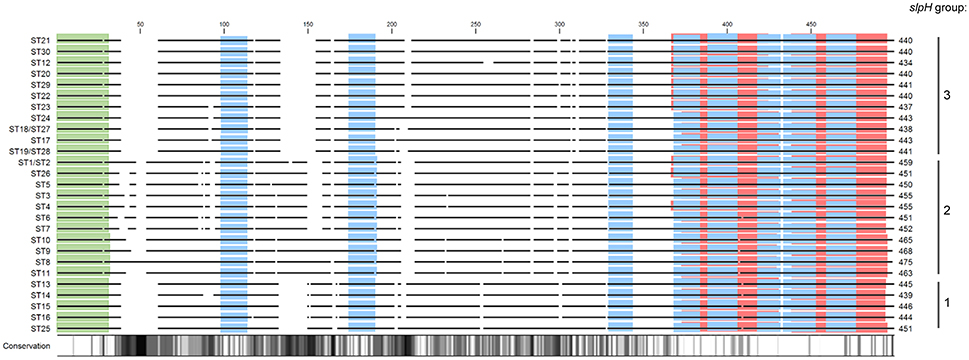
Figure 2. Alignment of the deduced amino acid sequences from the slpH genes from all of the different sequence types (STs). The solid lines represent aligned amino acid sequences. Gaps between the lines represent gaps in the alignment. The length of the amino acid sequence is given for all of the STs. The degree of amino acid conservation among the different sequences is shown with a gradient ranging from white to black. White regions indicate 100% conservation, whereas black regions indicate 0% conservation. The proteins were compared to each other in silico using InterProScan. The red horizontal bars indicate the tandem SLAP domain (InterPro accession number IPR024968). The blue and green bars represent conserved motifs (PRINTS) of the Lactobacillus surface layer protein family (InterPro accession number PR01729) and the signal peptide domains, respectively.
When we compared the surrounding context of the slpH locus in 10 strains possessing different STs and slpH groups, we found that up- and downstream various OGCs were co-localized with the slpH locus between the strains (Figure 3). Nine of the genes surrounding slpH were present in all of the analyzed strains (Figure 3).

Figure 3. Comparison of the slpH gene (14) and surrounding genes of 10 L. helveticus strains. The complete genome sequences were retrieved from GenBank. Orthologous gene clusters were determined using Ortho-MCL. Genes belonging to the same orthologous gene cluster are depicted as arrows with the same number. The different colors represent different gene functions.
Evidence for the Absence of L. gallinarum in Dairy Products
Since the primer pair LHslpF/R can also amplify a region from the gDNA of L. gallinarum strains, we established a PCR to assay dairy products for the presence of this species. A primer pair targeting the pheS gene of L. gallinarum showed species-specificity, as only a 180-bp amplicon was amplified from the gDNA of L. gallinarum, but not from closely related LAB (Table 4). The assay's limit of detection was determined by analyzing gDNA extracted from a dilution series of L. gallinarum DSM 10532. The lower limit of detection was determined to be at 2.45 × 102 colony forming units per mL broth. The method was generally applied to all samples used for semiconductor sequencing to confirm the absence of L. gallinarum.
Experimental Validation
A mixture of 10 L. helveticus gDNAs, each of which represented a different ST (Table 3), was prepared. The amplicons obtained after slpH amplification were sequenced on the Ion Torrent PGM sequencer. After extraction of the reads containing the slpH group specific identifying subsequences, the proportion of reads assigned to the 10 known STs ranged from 5.85 (±0.12)% to 15.73 (±1.00)% (Table 3). Each slpH group also comprised reads that were not assigned to the known STs and indicate the presence of new STs. The abundance of those reads per new ST did not exceed 1.7 (±0.3)% of all reads per slpH group. Based on this observation, we decided that at least 3% of all reads within a slpH group must be assigned to an ST to be considered a true positive result.
Comparison Culture-Dependent Vs. Culture-Independent Typing
We also assessed the applicability of the slpH locus for typing, by comparing the slpH loci obtained after strain isolation using the culture-dependent approach with those obtained with the culture-independent approach. To effect this, the NWC from La Praz (sample p in Table 2) was plated on MRS agar plates containing lactose as a carbohydrate source. Of the 96 colonies picked, 81 were identified as L. helveticus using a species-specific qPCR. The slpH locus was amplified and sequenced from each L. helveticus isolate. In addition to two previously known STs, we identified two new STs, 31 and 32. The four STs were distributed as follows: 54.3% of the colonies were assigned to ST13 (slpH group 1), 8.6% were assigned to ST31 (slpH group 2), and 34.6 and 2.5% were assigned to ST17 and ST32, respectively (both slpH group 3; Figure 4).
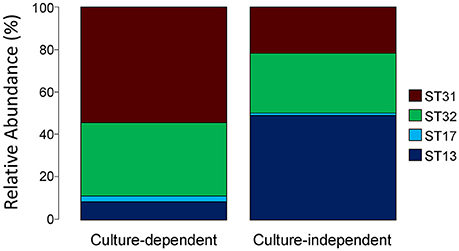
Figure 4. Comparison of the culture-dependent and culture-independent approaches. A natural whey culture (NWC) sample from La Praz, Switzerland was analyzed by plating. The slpH gene sequences were determined from 81 L. helveticus colonies by Sanger sequencing. The same NWC sample was examined using amplicon-based semiconductor sequencing without prior bacterial isolation. The stacked barplots show the relative abundance of each sequence type (ST) found in the sample for both methods.
This result was then compared to the sequence analysis obtained from the culture-independent approach. We identified the same STs as found by the culture-dependent approach. All assigned reads were distributed as follows: 21.66% were assigned to ST13, 49.04% were assigned to ST31, and 28.23 and 1.07% were assigned to ST17 and ST32, respectively (Figure 4).
Distribution of L. helveticus Strains in NWCs
The diversity of L. helveticus strains present in 15 NWCs, collected at variously located Gruyère cheesemaking factories (Table 2), was analyzed using our established culture-independent method. The quantification of L. helveticus using qPCR revealed an average population density of 4.42 (±3.79) × 107 cells per mL of NWCs according to pheS copy numbers. The presence of L. gallinarum was not detected by the specific PCR assay. When we analyzed the slpH loci using the culture-independent approach, we detected a total of 10 STs in the 15 NWC samples (Figure 5). The presence of STs ranged from one to five per NWC. Therefore, for example, the NWC from La Brévine contained only one strain, whereas the specimen from Orsonnens contained five STs. The majority (10 NWCs) contained three different STs of L. helveticus. Remarkably, ST13 was present in all the NWC samples (Figure 5, blue bar).
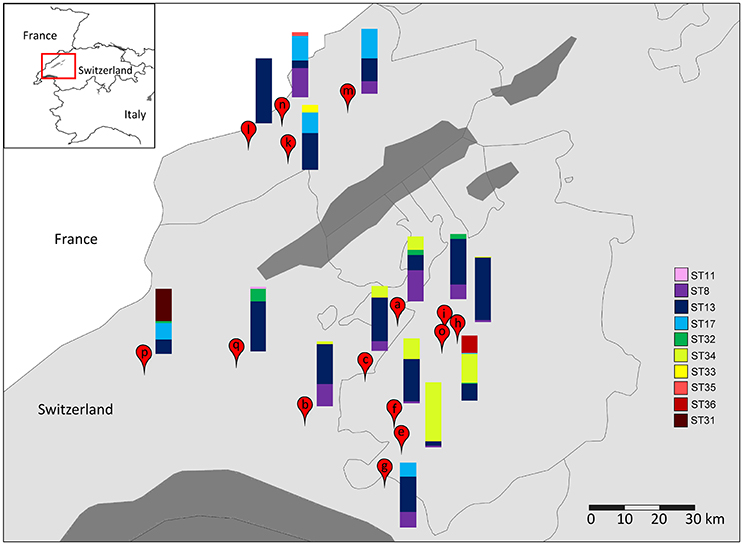
Figure 5. Geographical distribution of L. helveticus strains. The diversity of slpH loci present in NWC samples collected from cheese factories in (a) Villarzel, (b) Corcelles-le-Jorat, (c) Brenles, (e) Semsales, (f) Le Crêt, (g) Châtel-St. Denis, (h) Autigny, (i) Chénens, (k) Les Monts de Travers, (l) La Brévine, (m) La Sagne, (n) Le Cerneux-Péquignot, (o) Orsonnens, (p) La Praz, and (q) Penthéréaz were analyzed by amplicon-based semiconductor sequencing. Borders between provinces and countries are indicated by black lines. Dark gray areas indicate lakes. The relative abundance (%) of each sequence type (ST) is represented by the height of the relative color in the stacked barplot for each sample.
Development of L. helveticus Strains in Raclette Cheeses
We also monitored the development of L. helveticus strains during cheese ripening by analyzing samples collected from a cheese that had been manufactured with three L. helveticus STs, namely ST10 (FAM13019), ST 23 (FAM23236), and ST13 (FAM23237). QPCR estimated the population density of L. helveticus to be 108 copies g−1 cheese after 24 h of ripening and 106 copies g−1 cheese after 80–120 days (Table 2). By using the typing method, we detected only two of the three added L. helveticus STs in the cheese ripened for 24 h (Figure 6). After 80 and 120 days, all three of the STs used were detected. The relative abundance of ST13 increased from 27.36% after 24 h of ripening to 31.42% after 80 days and 67.47% after 120 days. For ST23, the relative abundance decreased from 72.64% after 24 h of ripening to 56.34% after 80 days and 25.92% after 120 days. ST10 was not detected in the cheese after 24 h of ripening and had a relative abundance of 12.24% after 80 days, which decreased to 6.6% after 120 days (Figure 6).
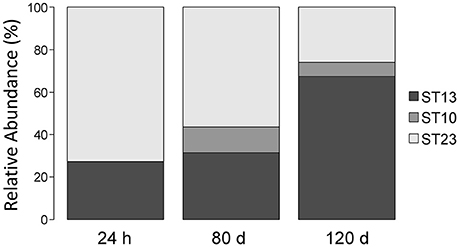
Figure 6. Relative abundance of L. helveticus strains during cheese ripening. Raclette cheese was made with three different sequence types (STs) of L. helveticus; samples were taken after 24 h, 80 d, and 120 d of ripening. The relative abundance of each ST was determined for each sample using the herein developed culture-independent approach for strain typing.
Discussion
The use of high-throughput sequencing techniques enables DNA-based surveys on the biodiversity of food ecosystems without the need to cultivate bacteria. To our knowledge, we present the first study to analyze L. helveticus strain diversity occurring in dairy products using next-generation sequencing. By analyzing the genomic sequences of 79 L. helveticus strains—58 own-sequenced genomes and 21 genomes taken from the GenBank database—, we found that the slpH gene exhibits a high amount of nucleotide polymorphisms and could be a suitable target for amplicon based high-throughput sequencing.
The nucleotide sequence heterogeneity of this locus in L. helveticus has already been described by other researchers (Ventura et al., 2000; Gatti et al., 2005; Waśko et al., 2014). Since Waśko et al. (2014) reported they could not detect the slpH gene in five L. helveticus strains, the question arises if this gene is ubiquitous in L. helveticus. Unfortunately, the authors neither explained how the species of these strains were determined nor did they show experimental evidence for the lack of the slpH gene in their paper. In contrast, we found that the slpH gene is part of the L. helveticus core genome using the 79 genomic sequences.
With regard to the nucleotide sequence heterogeneity present in the surface layer protein encoding genes, Gatti et al. (2005) found that the gene sequences clustered in two groups. The gene showed either similarity to the slpH1 gene encoding a surface layer protein (NCBI accession number X9119) or to the prtY gene encoding a putative cell surface proteinase (NCBI accession number AB026985). The slpH1 and prtY group correspond to the slpH groups 3 and 2 identified in this study (Figure 1). Additionally, we found a third group named slpH group 1 in this report. BLAST searches with the nucleotide sequences of this group resulted in putative surface layer proteins (data not shown). Despite these considerable sequence variations with ambiguous BLAST search results, we think that all sequences are alleles of the same locus. First, bioinformatics analyses showed that the deduced amino acid sequences of the three slpH groups clustered in the same OGC. Second, analysis of the genomic context of the slpH locus revealed conserved gene neighborhood. Finally, the search for domains revealed the presence of conserved amino acid motifs of the Lactobacillus surface layer protein family in all of the analyzed sequences. It is noteworthy that the protein sequence analysis of lactobacilli surface layer proteins was proposed as a method for strain typing (Podleśny et al., 2011). The authors demonstrated that the primary sequence variability present in the surface layer proteins could be determined with LC-MS/MS. Consequently, the method was suitable for lactobacilli strain identification within the L. acidophilus group.
The polymorphisms present in the slpH gene allowed us to differentiate 30 of the 79 strains used in this study. Conserved nucleotide sequences enabled us the design of a primer pair, which amplified a part of all known slpH loci. These primers were not totally species-specific, since an amplicon was also observed in L. gallinarum, the closest relative of L. helveticus. Although L. gallinarum has been detected in cheese by Van Hoorde et al. (2008), this species is usually associated with animals and has been isolated mainly from chicken guts (Hagen et al., 2003; Hammes and Hertel, 2006). Therefore, the occurrence of L. gallinarum in dairy products is probably of minor importance, since it is related to contamination. Further studies, such as metagenomics analyses of cheese or the use of L. gallinarum and L. helveticus in cheese experiments, will clarify whether dairy products are actually a habitat for L. gallinarum. Despite these shortcomings, we developed a species-specific PCR assay to analyze dairy products for the presence of L. gallinarum.
Due to the next-generation sequencing technology used is this study, we had to breakdown the amplicons that ranged between 1,104 and 1,230 bp. Consequently, only reads possessing the identifying subsequences could be used to detect STs (Supplemental Figure S1). Using this information we still differentiate of 24 STs of the 79 study strains and detected two new STs in the NWC of La Praz. New developments in next-generation sequencing technologies with larger read lengths will enable the use of the complete amplicon sequence for typing.
The applicability of the method was tested with a defined L. helveticus mixture, NWCs and cheese. In case of the L. helveticus mixture, all expected strains were identified. With regard to the NWC, no differences in the strain composition were found when a culture-dependent analysis was performed. A remarkable result was that in most cases, the L. helveticus population in NWCs was composed of more than one L. helveticus strain, often with representatives from all three slpH groups. Also in the cheese experiment all L. helveticus strains used for production were found. Again it was remarkable that the three strains coexisted after 120 days of ripening (Figure 4).
We assume that these strains are different in the phenotype. The coexistence in cheese could result from nutrient or physicochemical gradients present in this habitat. Possible reasons for strain diversity in whey could be caused by variations in whey composition over time or by the fact that some cheesemakers of Gruyère cheese use a mixture of whey cultures that are cultivated under different conditions. Another explanation for the stable coexistence of several strains is the reciprocal loss of metabolic genes resulting in inter-dependencies between strains, as discussed by Ellegaard and Engel (2016). Furthermore, bacteriophages might play an important role in the maintenance of bacterial strain diversity (Ellegaard and Engel, 2016). Currently, the drivers for intra-species diversity are not fully understood and need further investigations. NWCs or cheeses can serve as model microbial ecosystems to study the evolution of microbial diversity, as has been suggested by Wolfe and Dutton (2015).
Conclusions
Various cheese types (e.g., Italian-hard cheeses, Gruyère PDO cheese) are produced using NWCs. These starter cultures are undefined and L. helveticus has been shown to be one of the predominant species. The method presented herein was suitable for determining the biodiversity of L. helveticus present in NWCs, without the need to isolate single strains. It was also used to study the development of L. helveticus strains during cheese ripening. Cheesemakers of Swiss cheese varieties often assume that L. helveticus is the cause of unwanted openings, such as splits and cracks, in cheese. Our established culture-independent approach can be used to verify this hypothesis by studying the development of L. helveticus strains from the beginning of the cheesemaking process until the end of cheese ripening. Since L. helveticus is also widely used as starter culture in cheese making, the method can be used to monitor strains during cheese ripening and reveal relationships of certain strains with desired cheese properties. Therefore, we consider that the method is useful for studying the development and diversification of L. helveticus strain communities in cheese and to better understand its influence on cheese quality.
Author Contributions
AM, SI, LM, and EE: Conceived and designed the study; AM: Performed experiments; DW and RB: Performed bioinformatics analyses; AM, SI, and DW: wrote the manuscript; LM and EE: contributed to the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Hélène Berthoud for her critical reviewing of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01380/full#supplementary-material
References
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Boetzer, M., Henkel, C. V., Jansen, H. J., Butler, D., and Pirovano, W. (2011). Scaffolding pre-assembled contigs using SSPACE. Bioinformatics 27, 578–579. doi: 10.1093/bioinformatics/btq683
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
De Man, J., Rogosa, D., and Sharpe, M. E. (1960). A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135. doi: 10.1111/j.1365-2672.1960.tb00188.x
Drake, M., Boylston, T., Spence, K., and Swanson, B. (1997). Improvement of sensory quality of reduced fat Cheddar cheese by a Lactobacillus adjunct. Food Res. Int. 30, 35–40. doi: 10.1016/S0963-9969(96)00054-3
Ellegaard, K. M., and Engel, P. (2016). Beyond 16S rRNA community profiling: intra-species diversity in the gut microbiota. Front. Microbiol. 7:1475. doi: 10.3389/fmicb.2016.01475
Eugster-Meier, E., Fröhlich-Wyder, M. T., Jakob, E., and Wechsler, D. (2017). “Le Gruyère PDO-Switzerland,” in Global Cheesemaking Technology: Cheese Quality and Characteristics, eds P. Papademas, and T. Bintsis (Hoboken, NJ: John Wiley & Sons, Inc.).
Fortina, M. G., Nicastro, G., Carminati, D., Neviani, E., and Manachini, P. (1998). Lactobacillus helveticus heterogeneity in natural cheese starters: the diversity in phenotypic characteristics. J. Appl. Microbiol. 84, 72–80. doi: 10.1046/j.1365-2672.1997.00312.x
Gatti, M., Bottari, B., Lazzi, C., Neviani, E., and Mucchetti, G. (2014). Invited review: microbial evolution in raw-milk, long-ripened cheeses produced using undefined natural whey starters. J. Dairy Sci. 97, 573–591. doi: 10.3168/jds.2013-7187
Gatti, M., Rossetti, L., Fornasari, M. E., Lazzi, C., Giraffa, G., and Neviani, E. (2005). Heterogeneity of putative surface layer proteins in Lactobacillus helveticus. Appl. Environ. Microbiol. 71, 7582–7588. doi: 10.1128/AEM.71.11.7582-7588.2005
Giraffa, G., De Vecchi, P., Rossi, P., Nicastro, G., and Fortina, M. (1998). Genotypic heterogeneity among Lactobacillus helveticus strains isolated from natural cheese starters. J. Appl. Microbiol. 85, 411–416. doi: 10.1046/j.1365-2672.1998.853464.x
Hagen, K. E., Tannock, G. W., Korver, D. R., Fasenko, G. M., and Allison, G. E. (2003). Detection and identification of Lactobacillus species in crops of broilers of different ages by using PCR-denaturing gradient gel electrophoresis and amplified ribosomal DNA restriction analysis. Appl. Environ. Microbiol. 69, 6750–6757. doi: 10.1128/AEM.69.11.6750-6757.2003
Hammes, W. P., and Hertel, C. (2006). “The genera Lactobacillus and Carnobacterium” in The Prokaryotes, eds M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (New York, NY: Springer), 320–403.
Jenkins, J., Harper, W., and Courtney, P. (2002). Genetic diversity in Swiss cheese starter cultures assessed by pulsed field gel electrophoresis and arbitrarily primed PCR. Lett. Appl. Microbiol. 35, 423–427. doi: 10.1046/j.1472-765X.2002.01212.x
Jensen, M., Ardö, Y., and Vogensen, F. K. (2009). Isolation of cultivable thermophilic lactic acid bacteria from cheeses made with mesophilic starter and molecular comparison with dairy-related Lactobacillus helveticus strains. Lett. Appl. Microbiol. 49, 396–402. doi: 10.1111/j.1472-765X.2009.02673.x
Jensen, M. P., and Ardö, Y. (2010). Variation in aminopeptidase and aminotransferase activities of six cheese related Lactobacillus helveticus strains. Int. Dairy J. 20, 149–155. doi: 10.1016/j.idairyj.2009.09.007
Jensen, M. P., Vogensen, F. K., and Ardö, Y. (2009). Variation in caseinolytic properties of six cheese related Lactobacillus helveticus strains. Int. Dairy J. 19, 661–668. doi: 10.1016/j.idairyj.2009.04.001
Jones, P., Binns, D., Chang, H.-Y., Fraser, M., Li, W., McAnulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Li, L., Stoeckert, C. J., and Roos, D. S. (2003). OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res. 13, 2178–2189. doi: 10.1101/gr.1224503
Moser, A., Berthoud, H., Eugster, E., Meile, L., and Irmler, S. (2017). Detection and enumeration of Lactobacillus helveticus in dairy products. Int. Dairy J. 68, 52–59. doi: 10.1016/j.idairyj.2016.12.007
Podleśny, M., Jarocki, P., Komón, E., Glibowska, A., and Targoński, Z. (2011). LC-MS/MS analysis of surface layer proteins as a useful method for the identification of Lactobacilli from the Lactobacillus acidophilus group. J. Microbiol. Biotechnol. 21, 421–429. doi: 10.4014/jmb.1009.09036
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sun, Z., Liu, W., Song, Y., Xu, H., Yu, J., Bilige, M., et al. (2015). Population structure of Lactobacillus helveticus isolates from naturally fermented dairy products based on multilocus sequence typing. J. Dairy Sci. 98, 2962–2972. doi: 10.3168/jds.2014-9133
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3 – new capabilities and interfaces. Nucleic Acids Res. 40:e115 doi: 10.1093/nar/gks596
Van Hoorde, K., Verstraete, T., Vandamme, P., and Huys, G. (2008). Diversity of lactic acid bacteria in two Flemish artisan raw milk Gouda-type cheeses. Food Microbiol. 25, 929–935. doi: 10.1016/j.fm.2008.06.006
Ventura, M., Callegari, M. L., and Morelli, L. (2000). S-layer gene as molecular marker for identification of Lactobacillus helveticus. FEMS Microbiol. Lett. 189, 275–279. doi: 10.1111/j.1574-6968.2000.tb09243.x
Waśko, A., Polak-Berecka, M., Kuzdraliński, A., and Skrzypek, T. (2014). Variability of S-layer proteins in Lactobacillus helveticus strains. Anaerobe 25, 53–60. doi: 10.1016/j.anaerobe.2013.11.004
Keywords: strain typing, semiconductor sequencing, Lactobacillus helveticus, natural whey culture, population composition
Citation: Moser A, Wüthrich D, Bruggmann R, Eugster-Meier E, Meile L and Irmler S (2017) Amplicon Sequencing of the slpH Locus Permits Culture-Independent Strain Typing of Lactobacillus helveticus in Dairy Products. Front. Microbiol. 8:1380. doi: 10.3389/fmicb.2017.01380
Received: 28 March 2017; Accepted: 07 July 2017;
Published: 20 July 2017.
Edited by:
Danilo Ercolini, University of Naples Federico II, ItalyReviewed by:
Pierre Renault, Institut National de la Recherche Agronomique (INRA), FranceMonica Gatti, University of Parma, Italy
Copyright © 2017 Moser, Wüthrich, Bruggmann, Eugster-Meier, Meile and Irmler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stefan Irmler, stefan.irmler@agroscope.admin.ch