 María Alvarado
María Alvarado Antonio J. Martín-Galiano
Antonio J. Martín-Galiano María J. Ferrándiz
María J. Ferrándiz Ángel Zaballos
Ángel Zaballos Adela G. de la Campa
Adela G. de la Campa- 1Unidad de Genética Bacteriana, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Madrid, Spain
- 2Unidad de Genómica, Instituto de Salud Carlos III, Madrid, Spain
- 3Presidencia, Consejo Superior de Investigaciones Científicas, Madrid, Spain
We characterized the mechanism of fluoroquinolone-resistance in two isolates of Streptococcus pseudopneumoniae having fluoroquinolone-efflux as unique mechanism of resistance. Whole genome sequencing and genetic transformation experiments were performed together with phenotypic determinations of the efflux mechanism. The PatAB pump was identified as responsible for efflux of ciprofloxacin (MIC of 4 μg/ml), ethidium bromide (MICs of 8–16 μg/ml) and acriflavine (MICs of 4–8 μg/ml) in both isolates. These MICs were at least 8-fold lower in the presence of the efflux inhibitor reserpine. Complete genome sequencing indicated that the sequence located between the promoter of the patAB operon and the initiation codon of patA, which putatively forms an RNA stem-loop structure, may be responsible for the efflux phenotype. RT-qPCR determinations performed on RNAs of cultures treated or not treated with subinhibitory ciprofloxacin concentrations were performed. While no significant changes were observed in wild-type Streptococcus pneumoniae R6 strain, increases in transcription were detected in the ciprofloxacin-efflux transformants obtained with DNA from efflux-positive isolates, in the ranges of 1.4 to 3.4-fold (patA) and 2.1 to 2.9-fold (patB). Ciprofloxacin-induction was related with a lower predicted free energy for the stem-loop structure in the RNA of S. pseudopneumoniae isolates (−13.81 and −8.58) than for R6 (−15.32 kcal/mol), which may ease transcription. The presence of these regulatory variations in commensal S. pseudopneumoniae isolates, and the possibility of its transfer to Streptococcus pneumoniae by genetic transformation, could increase fluoroquinolone resistance in this important pathogen.
Introduction
Streptococcus pneumoniae (the pneumococcus) is a main human pathogen causing community-acquired pneumonia, meningitis, bacteremia, and otitis media (Obaro, 2000). It has been estimated that one million children 5 years of age and under die annually of pneumococcal infections (World Health Organization, 2007). Pneumococcal resistance to antibiotics normally used to treat pneumococcal infections, such as beta-lactams and macrolides has spread worldwide (Liñares et al., 2010). Nowadays, the fluoroquinolones (FQs) levofloxacin (LVX) and moxifloxacin (MXF) are indicated for the treatment of adult patients with pneumonia (Mandell et al., 2007). Although FQ-resistance in S. pneumoniae is low (<3%) in Europe (Riedel et al., 2007; Domenech et al., 2014), it is higher in Asia (10.5%, Ip et al., 2007), and in Canada (7.3%, Adam et al., 2009). However, an increase in resistance may occur if the use of these drugs increases (Chen et al., 1999). The intracellular targets of the FQs are the DNA topoisomerase enzymes gyrase and topoisomerase IV, which control DNA topology, solving topological problems associated with replication, transcription and recombination (Champoux, 2001). In S. pneumoniae, most FQs act on topoisomerase IV as a primary target (Muñoz and de la Campa, 1996; Morrissey and George, 1999). FQ-resistance in this bacterium occurs mainly by the alteration of their DNA topoisomerases. This can occur either by point mutation or by intraspecific (Stanhope et al., 2005), or interspecific recombination with the genetically related streptococci of the mitis group, commensal microorganisms of the oral cavity (Ferrándiz et al., 2000; Balsalobre et al., 2003; Stanhope et al., 2005). Some FQ-resistant (FQ-R) mutations impose a fitness cost to S. pneumoniae (Rozen et al., 2007; Balsalobre and de la Campa, 2008). Compensation of this cost in isolates carrying recombinant topoisomerase genes (Balsalobre et al., 2011) would envisage a future spread of these resistant isolates. These recombinant isolates are originated by genetic interchange with chromosomal DNA of other streptococci of the mitis group. Among them, the species most genetically close to S. pneumoniae are Streptococcus mitis, Streptococcus oralis, and Streptococcus pseudopneumoniae (Arbique et al., 2004). Homologous recombination among streptococci of the mitis group plays a role in the evolution of antibiotic resistance in these bacteria. Recombination had played a role in antibiotic resistance in S. pneumoniae. Recombinant genes in the topoisomerase genes (Ferrándiz et al., 2000; Balsalobre et al., 2003), in the genes of penicillin-binding proteins (Dowson et al., 1990) and in the rpoB genes of rifampicin-resistant strains (Ferrándiz et al., 2005) have been identified. Recombination and the spread of a few international clones are central factors generating antibiotic resistance in S. pneumoniae.
Alternatively, pumping of FQs out of the cell has also a role on resistance in S. pneumoniae. FQ-efflux is present in S. pneumoniae clinical isolates (Brenwald et al., 1998; de la Campa et al., 2004, 2009) and also in streptococci of the mitis group (Ferrándiz et al., 1999). PmrA, which belongs to the major facilitator superfamily (MFS), was the first pump described in S. pneumoniae, whose inactivation in a FQ-R isolate derived from the laboratory strain R6, restored FQ sensitivity (Gill et al., 1999). Further studies pointed out the existence of other FQ-efflux pumps (Pestova et al., 2002; Brenwald et al., 2003). The role of PmrA in FQ-resistance in clinical isolates has not been proved, since no correlation has been observed between pmrA expression and FQ-efflux (Piddock et al., 2002). A systematic deletion of 11 putative efflux pumps in S. pneumoniae TIGR4 showed an association between deletion of the ABC transporter PatAB and hypersusceptibility to ciprofloxacin (CPX), acriflavine, and ethidium bromide (Robertson et al., 2005). The same phenotype was observed in FQ-R clinical isolates (Garvey et al., 2011). The patA and patB genes showed CPX-induced overexpression in both a FQ-R laboratory mutant (Marrer et al., 2006), and in clinical isolates (El Garch et al., 2010). Further studies associated mutations in a RNA stem-loop structure located upstream of patAB with both the overexpression of these genes and FQ-resistance (Baylay and Piddock, 2015). Likewise, the homolog of PatAB in the zoonotic pathogen Streptococcus suis, named SatAB, has been shown to be constitutively overexpressed and to confer CPX resistance to a clinical isolate (Escudero et al., 2011). The satAB genes are regulated by the satR repressor, which is located upstream of these genes and cotranscribed with them (Escudero et al., 2013). These studies stablished a role for PatAB in FQ-resistance in clinical isolates of S. pneumoniae. In addition, a role in LVX and MXF efflux has been stablished for the multi-antimicrobial extrusion family (MATE) DinF transporter in a pneumococcal laboratory strain (Tocci et al., 2013).
Among the FQ-R (considering those with CPX MICs ≥4 μg/ml) S. pneumoniae isolates characterized in our laboratory in two epidemiological studies, one performed during 1991 to 2001 (Ferrándiz et al., 2000; Balsalobre et al., 2003), and the other in 2002 (de la Campa et al., 2004), we selected 14 isolates carrying DNA topoisomerase genes with high nucleotide sequence variations (>4%). Seven out of these 14 isolates were further classified as S. pseudoneumoniae (Balsalobre et al., 2013). Two of them, 5305 and CipR-71, which did not carry mutations in the quinolone-resistance determining regions of their topoisomerase genes, showed efflux as a single mechanism of FQ resistance. As far as we know, there are no other descriptions of FQ-resistant S. pseudoneumoniae. In this study we aimed to identify the efflux mechanisms of FQ-resistance of these isolates. Whole genome sequencing and genetic transformation experiments were performed together with phenotypic determination of FQ-efflux.
Materials and Methods
Bacterial Strains and Conditions for Growth and Transformation
Streptococcus pneumoniae and Streptococcus pseudopneumoniae strains were grown as static cultures either in a casein hydrolysate based medium (AGCH) supplemented with 0.3% sucrose and 0.2% yeast extract (A+SY) (Lacks et al., 1986) or in Tod-Hewitt (Becton Dickinson) with 0.5% of yeast extract (THY). MICs of ethidium bromide, acriflavine and FQs were determined by agar-dilution (for S. pseudopneumoniae 5305) or by macrodilution following the methods of the CLSI (Clinical Laboratory Standards Institute, 2008) using 2-fold dilutions in A+SY liquid medium, with an inoculum of 5 × 105 cells per well. The efflux inhibitor reserpine (Gill et al., 1999; Garvey et al., 2011) was used at a final concentration of 10 μg/ml, which has been shown to inhibit ethidium bromide, acriflavine, and CPX efflux in streptococci of the mitis group (Ferrándiz et al., 1999). S. pneumoniae R6 was used as the recipient in transformation experiments. Competent cultures containing 9 × 106 CFU/ ml were incubated with 0.1 μg/ ml of DNA during 40 min at 30°C and then at 37°C for 90 min before plating. Colonies were counted after 24 h of growth at 37°C in a 5% CO2 atmosphere in A+SY media plates containing 1% agar and CPX at 2 μg/ ml. No transformation refers to experiments in which no colonies were observed when 100 μl of the transformation mixture was plated, which gave a frequency <1 × 101. DNA used as control was the chromosome of T5305 that yielded 1.1 × 103 transformants/ml.
Whole-Genome Sequencing
Two S. pseudopneumoniae clinical isolates, 5305 and CipR71, and two S. pneumoniae strains, an R6 transformant with 5305 chromosomal DNA, named S. pneumoniae T5305 and our laboratory S. pneumoniae R6 strain, were subjected to whole genome sequencing. Genomic DNA samples were prepared from mid-log phase cultures using the CTAB protocol (Wilson, 1994). Sequencing was performed either in a 454 Life Sciences GS-FLX+ system, Roche (5305 and CipR71) or in an Illumina NextSeq 500 system (T5305 and our laboratory R6). DNA for 454/Roche library preparation was fragmented by nebulization and posterior quantification, emulsion PCR and sequencing performed as recommended by the manufacturer. Libraries for Illumina sequencing were obtained following the transposon-mediated Nextera XT library preparation kit (Illumina) and paired-end sequenced with a 2 × 150 protocol. De novo or reference aided (S. pneumoniae R6) assemblies were performed using Newbler 3.0 (Roche) with default settings. sff files from 454/Roche and fastq files from Illumina were used as input sequences, respectively. Assembly metrics are shown in Table 1. Coverage was calculated by dividing total bases used in the computation by the size of the final alignment following the Lander/Waterman equation (Lander and Waterman, 1988).
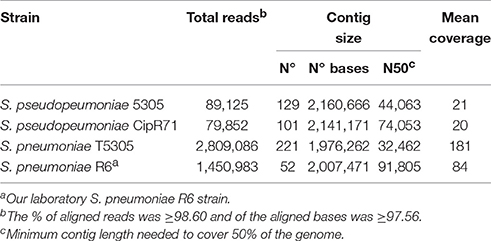
Table 1. Characteristics of the whole genome sequences.
Gene and Protein Analysis
Coding sequences were predicted by Prodigal V.2.6.3 with default settings (Hyatt et al., 2010). S. pneumoniae pump gene sequences (Tocci et al., 2013) were detected in S. pseudopneumoniae genomes by BLASTN using S. pneumoniae TIGR4 sequences as queries and applying bidirectional thresholds of ≥80% identity and an alignment length over ≥80% of the protein sequences. Promoter-containing sequences of S. pseudopneumoniae isolates were extracted as the 100 upstream nucleotides of the pump gene, or, in the case of genes in operons, the 100 upstream nucleotides of the first gene of the operon. The RNA stem-loop structure was predicted by using the RNA-fold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi). Gene organization of operons was downloaded from the DOOR database (Mao et al., 2009), using the S. pseudoneumoniae IS7493 strain as reference. Protein and nucleotide sequences were aligned with Clustal Omega (Sievers et al., 2011).
Nucleotide Sequence Accession Numbers
The sequencing data have been deposited at the whole genome sequencing database linked to the BioProject ID PRJNA386436.
PCRs, RNA Extraction and Quantitative Real Time PCR (RT-qPCR) Procedures
Primers used for PCR amplifications and sequencing are shown in Table 2. Platinum® Taq High Fidelity (Invitrogen), was used for PCR amplifications in the following conditions: an initial cycle of 2 min denaturation at 94°C; 30 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s and extension at 68°C for 1 min per kb of PCR product. Primers used for PCR amplifications and sequencing are shown in Table 2. To quantify the relative expression of hexA, patA, and patB, a procedure previously described (El Garch et al., 2010) was used. Briefly, R6-derivative strains were grown overnight at 37°C in 5% CO2 in Mueller-Hinton agar plates supplemented with 5% defibrinated sheep blood (Becton Dickinson). Bacteria were suspended in 5 ml of THY, grown to OD620nm = 0.2, diluted 10-fold into 25 ml of THY and grown to OD620nm = 0.2 to 0.4 (2–4 × 108 cells). At this point, cultures were split into two parts; one part was grown during 4 h in the presence of CPX at 0.5 × MIC for each strain, and the other was grown in its absence. Cells were collected by centrifugation and their total RNA was extracted with an RNeasy mini kit (Qiagen), with 3 DNase I (Qiagen) treatments, according to the manufacturer's instructions. Synthesis of cDNAs were carried out in 20-μl reactions containing 0.5–5 μg of RNA, 0.5 mM of each deoxynucleoside triphosphate, 2 pmol of hexanucleotide random primers, 40 U of the RNaseOUT RNase inhibitor (Invitrogen), 200 U of SuperScript IV reverse transcriptase (Invitrogen), and 2 U of RNase H− (Invitrogen), in the buffer recommended by the manufacturer. These cDNAs were used in RT-qPCR amplifications (Light cycler 480, Roche). The total volume in each reaction was 20 μl and contained 2 μl of cDNA, 0.4 μM of each primer, and 10 μl of Sso Advanced Universal SYB Green Supermix (BioRad). The program had one initial denaturation cycle of 4 min at 94°C followed by 42 cycles of three steps: denaturation (30 s at 94°C), annealing (30 s at 54°C), and elongation (30 s at 68°C). Oligonucleotide pairs used were: PatARTF/PatARTR (patA); PatBRTF/ PatBRTR (patB), HexARTF/HexARTR (hexA). The amount the RT-PCR products was calculated with a standard curve for each amplicon. Assays were performed in triplicate and normalized against the signal from a 16S rDNA internal control (oligonucleotide pair rDNAF/rDNAR).
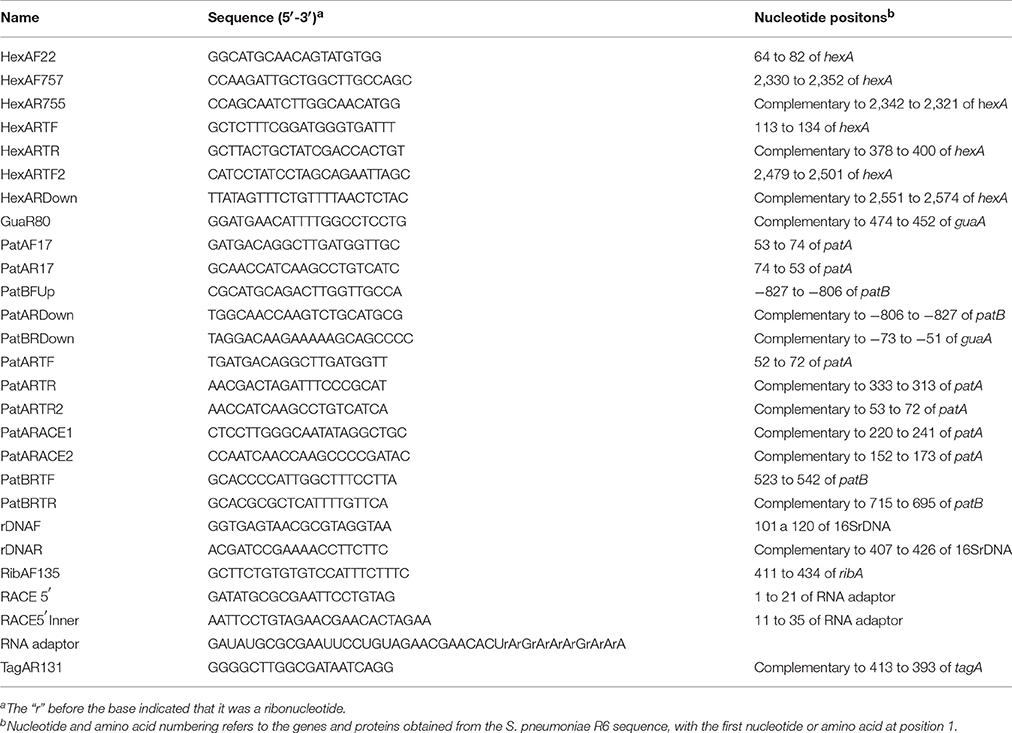
Table 2. Oligonucleotides used in this work.
Transcription Start Site Determination
5′ RACE (rapid amplification of cDNA ends) method was performed using total RNA extracted from strain TCipR71.2 treated with CIP at 0.5 × MIC as template. A total of 12 μg of RNA were used, 6 μg were treated with Tobacco acid pyrophosphatase (TAP) to hydrolyze specifically 5′-triphosphate from primary transcripts, and to subsequently be ligated to the 3′-hydroxyl group of an RNA oligonucleotide (RNA adaptor, Table 2). The remaining 6 μg of total RNA were not treated with TAP and followed the same post-treatment. cDNAs were synthesized from these RNAs using SuperScript IV Reverse Transcriptase (Invitrogen) following manufacturer recommendations and used as templates in PCR reactions with primers PatARACE1 and RACE5′ (Table 2) as detailed in PCRs, RNA Extraction and Quantitative Real Time PCR (RT-qPCR) Procedures. PCR products were purified with MinElute PCR purification kit (Quiagen) and used as templates for a second PCR reaction using primers PatARACE2 and RACE5′Inner (Table 2). PCR products were separated in a 2% Metaphor (FMC Bioproducts) agarose gel and the band that appeared in the sample treated with TAP, and did not appear in the untreated sample, was isolated from the gel with MinElute Gel Extraction Kit (Qiagen). DNA was sequenced using primer PatARACE2 to determine the transcription start site, which was the first base 5′ immediately after the adaptor sequence.
Results
Efflux Phenotypes of S. pseudopneumoniae Isolates
The MICs of isolates 5305 and CipR71 to FQs and some dyes in the presence and absence of the efflux pump inhibitor reserpine were compared to three efflux-negative control strains: S. pneumoniae R6, S. pseudopneumoniae CCUG48465, and the S. pseudopneumoniae type strain CCUG49455T. No significant differences in the MICs of LVX, CPX, ethidium bromide and acriflavine in the presence or absence of reserpine were observed in the control strains. However, the two S. pseudopneumoniae clinical isolates, which showed 8- to 32-fold higher MICs of these drugs in the absence of reserpine than the control strains, essentially behave like the control strains in the presence of the pump inhibitor (Table 3). Reserpine it is assumed not to affect growth of the strains at the concentration used (10 μg/ ml), since the MICs of reserpine for S. pneumoniae have been described to be 60 μg/ml (Garvey and Piddock, 2008). These results suggested the existence of an efflux pump in S. pseudopneumoniae isolates 5305 and CipR71.
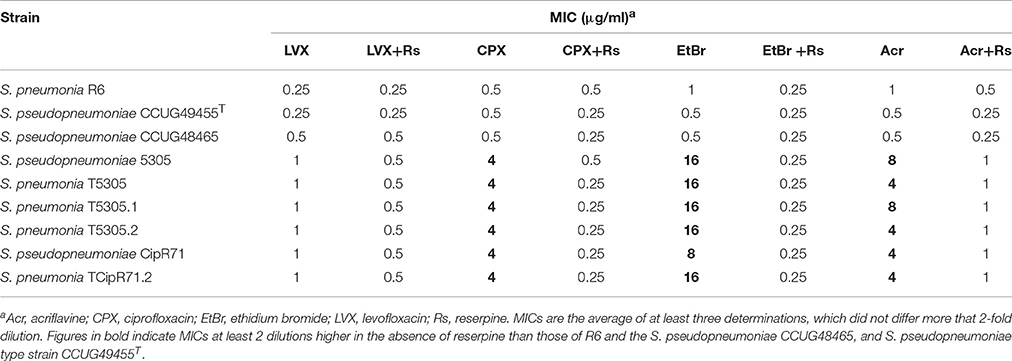
Table 3. Susceptibilities of strains to fluoroquinolones and efflux pump substrates.
Genome Sequence of S. pseudopneumoniae Ciprofloxacin-Resistant Strains
We analyzed the genome sequences of the 5305 and CipR71 resistant strains and compared them with those of S. pneumoniae and S. pseudopneumoniae isolates, paying special attention to those coding for the 11 pneumococcal antimicrobial pumps (Tocci et al., 2013). S. pseudopneumoniae isolates carried nine of these pumps, coded by 14 genes, given that they are constituted by complexes in some cases. Amino acid sequences of the permease subunits and the sequences covering the 100 nucleotides upstream of the coding sequence (or of the first coding sequence of the operon) were compared (Table 4). Subtle differences were detected in the sequences of these pumps. However, simultaneous alterations, i.e., alterations common to both S. pseudopneumoniae isolates with an efflux phenotype (CipR71 and 5305) respect to the efflux-negative S. pseudopneumoniae type strain ATCC BAA-960, were found only in the coding sequences of five of these genes (affecting four pumps), which essentially ruled out the possible involvement of the nine remaining genes in the efflux phenotype. The most remarkable efflux-linked determinants were the coding sequence of the ABC transporter SP_1357 (11 residue changes) and the regulatory sequence of the patAB operon. Both proteins are part of the Drug Exporter-4 family of the Transport Classification Database (TCDB id: 3.A.1.135). The enhanced CPX efflux of isolates 5305 and CipR71 could be attributed either to higher substrate affinity/enhanced activity produced by the residue changes in SP_1357, by an up-regulation of the PatAB pump, or by a combination of both. To discern between these possibilities, genetic transformation experiments were carried out as described below.
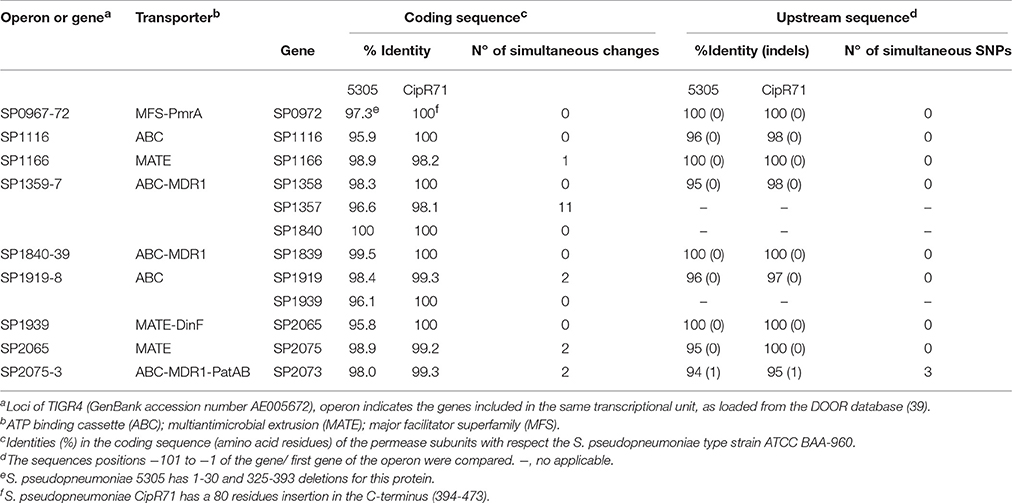
Table 4. Comparison of coding and regulatory sequences of S. pseudopneumoniae homologs to S. pneumoniae pumps associated to drug efflux.
Genome Sequence of the R6 Transformant (T5305) Revealed Two Recombination Regions
Streptococcus pneumoniae R6 was transformed with chromosomal DNA of S. pseudopneumoniae 5305. One transformant, T5305, was selected. T5305 had a CPX MIC of 4 μg/ml, this MIC was equivalent to that of S. pseudopneumoniae 5305. In addition, 5305 and T5305 exhibited the same efflux phenotype to CPX, ethidium bromide and acriflavine (Table 3). The analysis of the S. pneumoniae T5305 genomic sequence showed 420 nucleotide changes with respect to the sequence of S. pneumoniae R6 present in the NCBI data base (Accession number: NC_003098.1). To discard spurious mutations, our laboratory R6 strain was also sequenced, showing 21 SNPs with respect to the NCBI sequence (data not shown). These changes were not considered, then, a total of 399 changes were attributed to the recombination event. These changes were unevenly distributed, 353 of them were located in a 7184 bp region (4.8% of variation) covering the spr1884 to spr1888 genes (guaA-patB-patA-hexA); 45 in a 3540 bp region (1.2% of variation) covering the spr0162 to spr0166 genes (ribA-ribC-ruvA-tag); and 1 point mutation in spr1903 (galU). These results demonstrate that the recombination event yielding T5305 occurred in two regions: spr0162-0166 and spr1884-1888 (Figure 1). This was very likely due to its close spatial proximity considering both recombined regions are at the same distance to the chromosome replication origin, a factor that may increment inter-replicore gene transference.
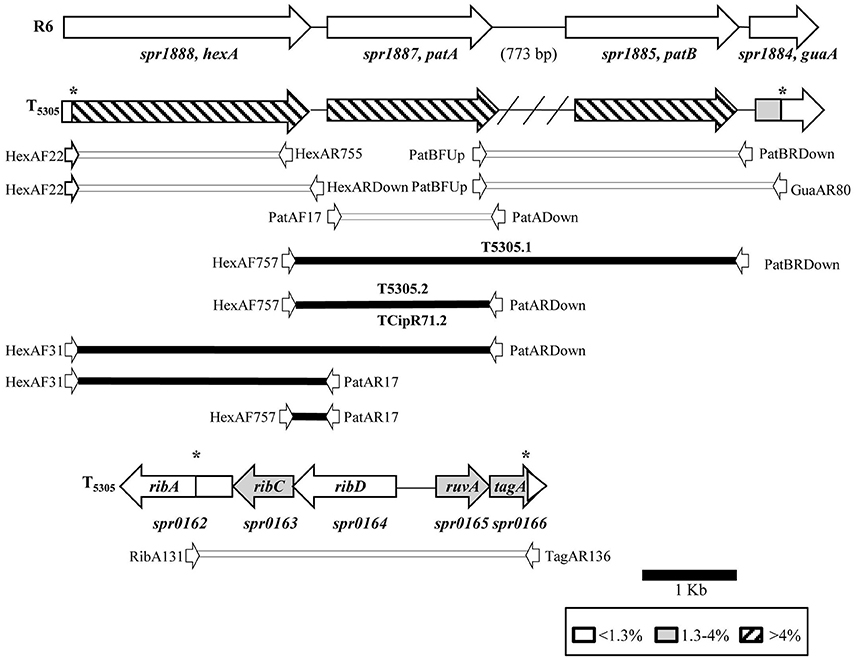
Figure 1. Genomic map of the recombination regions of S. pneumoniae T5305. Genes are represented as arrows, indicating the level of nt divergence with respect SPN R6. PCR fragments used in transformation experiments, together with their corresponding primers (white arrows, not draw to scale), are indicated. The intergenic region between patA and patB is 773-bp in S. pneumoniae, but is absent in S. pseudopneumoniae isolates (indicated by a crosshatched line). Black boxes and white boxes indicate PCR fragments yielding or not yielding CPX-R transformants, respectively. Asterisks indicate recombination points.
The Region Involved in FQ Efflux Includes the Upstream patA Region
To reveal the role of the nucleotide changes present in T5305 in FQ-efflux, diverse DNA fragments were amplified from this strain by PCR with specific oligonucleotides (Table 2). These fragments were used as donor DNAs in transformation experiments with R6 as receptor, and transformants selected at 2 μg/ml of CPX. Transformation experiments were performed as described under Materials and Methods, being the frequencies of transformation in the range of 1 to 7 × 103 transformants/ ml. DNA fragments covering the spr0162-0166 genes or the galU mutation did not result in transformants at elevated CPX concentrations, discarding the role of these genes in CPX-resistance. In contrast, two overlapping PCR fragments from the spr1884-1888 region transformed R6 with high efficiency (Figure 1). One of the amplicons covers the C-terminus of hexA and the intergenic hexA-patA region plus patA, whereas the other covers the same regions but including patB. In accordance, PCR products including most of the hexA and patA sequences were also able to transform to CPX resistance. The smallest fragment able to transform R6 to CPX resistance was obtained by amplification with oligonucleotides HexAF757 and PatAR17. Likewise, PCR products obtained from isolate CipR71 with the same oligonucleotide pairs also transformed R6 with high efficiency. In contrast, DNA fragments containing hexA, patA, patB, or guaA coding sequences, but not the hexA-patA intergenic region, were unable to transform R6 to CPX-resistance. In addition, a fragment covering hexA from residue position 22 to the stop codon, which was obtained from S. pseudopneumoniae strains 5305 and CipR71, did not transform to CPX-resistance (Figure 1). Three resistant transformants, two obtained with PCR products from 5305 (T5305.1 and T5305.2) and one from CipR71 (TCipR71.2) were selected for further studies. Their nucleotide sequences were identical to that of the donor 5305 and CipR71 strains and their MICs were equivalent to that of their parental DNA donors (Table 3). All these results demonstrated that the region involved in FQ-efflux in the S. pseudopneumoniae strains T5305 and CipR71 was that included between oligonucleotides HexARDown and PatAR17 (Figure 2A). This region contains the intergenic hexA-patA region and the first 74 nucleotides of patA. No amino acid changes were predicted in PatA between R6, 5305 and CipR71 or the derived R6-transformants. The intergenic hexA- patA region has been previously stablished to be involved in FQ-efflux in SPN (Baylay and Piddock, 2015). A promoter with a canonical −35 box and an extended −10 sequence was observed in all strains. We determined the transcription start site by RACE 5′ PCR as described in materials and methods, which was located eight nucleotides downstream of extended −10 sequence (Figure 2B). The main differences were found between the extended −10 sequence and the patA initiation codon, in which the two S. pseudopneumoniae FQ-resistant strains CipR71 and 5305 showed a 3-nt insertion (GCT) or a 3-nt deletion (GGA), respectively. A stem-loop structure could be predicted by RNA-fold in this region with a free energy of −15.32 kcal/mol for R6 (Figure 2C). The nucleotide changes observed in CipR71 and 5305 CPX-resistant isolates are predicted to lower the free energies of these structures to −8.58 kcal/mol for 5305 and −13.82 kcal/mol for CipR71. These changes in the stability of the stem-loop structures could cause alterations in the transcription of the downstream patA and/ or patB genes.
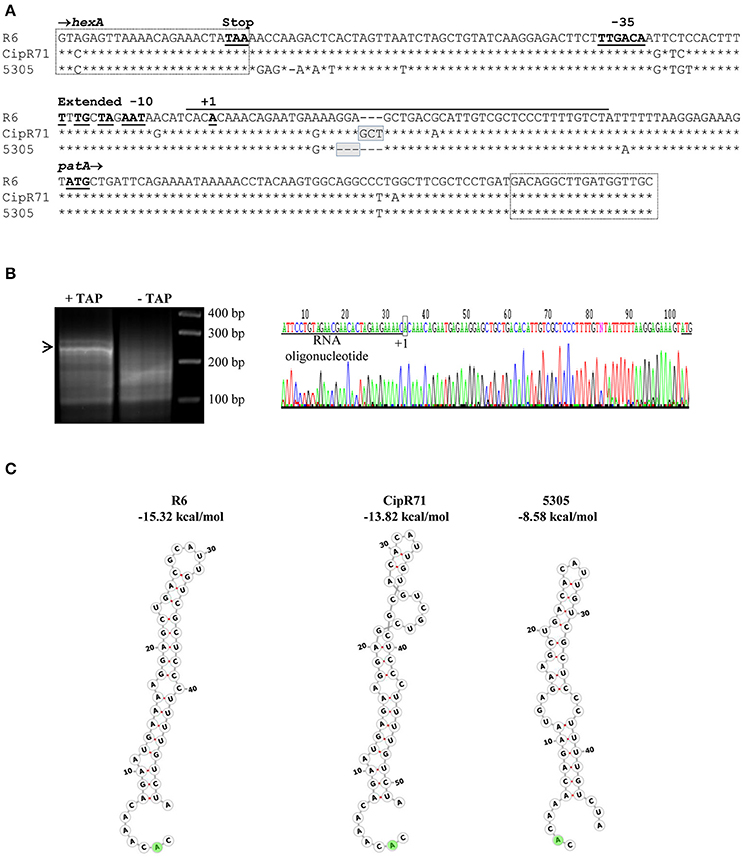
Figure 2. Sequence of the region involved in CPX resistance in S. pseudopneumoniae isolates, determination of the patAB transcription star site, and putative structures in the patA upstream RNAs. (A) Sequence included between oligonucleotides HexARDown and PatAR17, which are boxed, is shown in full for S. pneumoniae R6. For S. pseudopneumoniae strains CipR71 and 5305, only nucleotide changes are indicated, with nucleotides identical to R6 indicated by asterisks, and deletions by dashes. (B) 5′ RACE PCR method. Agarose gel electrophoresis of PCR products of samples treated (+) or non-treated (–) with tobacco acid pyrophosphatase (TAP). The band indicated by an arrowhead was extracted and sequenced. The chromatogram, sequence of the RNA oligonucleotide, star site, and initiation codon of patA are indicated. (C) Structures of the regions marked with a black line in (A) as predicted by RNA-fold. The A shadowed in green indicates the start site.
Transcription of patA Is Induced by CPX
To quantify the induction produced by CPX, RT-qPCR experiments were performed with RNAs of cultures treated or not treated with a subinhibitory CPX concentration. These assays showed that the expression of both patA and patB in the R6-derivative strains was induced by CPX. While no significant changes were observed in R6, increases in patA and patB were detected in all strains with an efflux phenotype. These increases were in the range of 1.4 to 3.4-fold for patA, and of 2.1 to 2.9-fold for patB. However, the transcription of hexA did not show significant differences between R6 and its transformant-derived strains, being in the range of 0.7 to 1.3-fold (Figure 3).
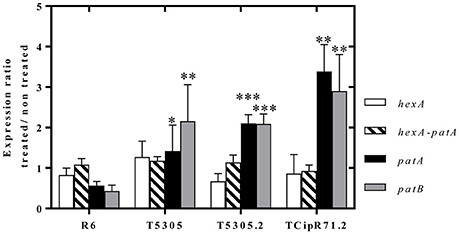
Figure 3. Expression of hexA, patA and patB in R6-derivative S. pneumoniae strains cultures exposed for 4 h to 0.5 × MIC of CPX. Data are represented as ratios of expression under treated and non-treated cultures. Values are mean ±SD of triplicates from three independent experiments. Statistical significance with respect to R6 values by Student T-test: *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001.
Discussion
In this study we have characterized the FQ-efflux phenotype of two S. pseudopneumoniae isolates, 5305 and CipR-71, which do not have mutations in their DNA topoisomerase genes. Changes in the regulatory sequences of the patAB operon were found. The presence of these changes in two genetically diverse isolates is significant. CipR71 is genetically closer to the S. pseudopneumoniae reference strain than to 5305, as inferred from the higher identity between pump sequences (Table 4) and the fact they share three out of seven alleles (ddl, gki and recP) used in multilocus sequence typing, whereas 5305 shares none. These changes would have been acquired by horizontal transfer and/or convergent evolution processes. In addition, these sequence features were absent or very unusual in S. pseudopneumoniae as corroborated in the only complete genome fully sequenced in this species—IS7493—and the 29 S. pseudopneumoniae strains present in the whole-genome shotgun contig database of the NCBI that were screened. In addition, other pumps, whose function has not been determined, were detected in the CPX-efflux isolates (data not shown).
The analysis of the T5305 sequence, together with transformation experiments, showed that the region involved in CPX-efflux, which has the greatest variation by far with respect to R6, was a 7184 bp region covering spr1884 to spr1888 genes (guaA-patB-patA-hexA). The region involved was further reduced to 247-bp, independently of the donor DNA strain (5305 or CipR71). This region contains the last 24 nt of hexA, the intergenic hexA-patA region, and the first 74 nt of patA. No amino acid changes were predicted in HexA or PatA between R6, 5305 and CipR71, and therefore we conclude that the intergenic hexA- patA region previously established to be involved in FQ-efflux in S. pneumoniae (Baylay and Piddock, 2015), is also involved in S. pseudoneumoniae. Then, the enhanced CPX efflux of isolates 5305 and CipR71 could be attributed to the regulation of the expression of the patAB operon, and not to residue changes in the proteins of the pump. The main difference was found in the sequence between the extended−10 region and the patA initiation codon, in which a 3-nt insertion (GCT) in the case of CipR71, or a 3-nt deletion in the case of 5305, were observed. These indels were not found in the patAB upstream regions of the reference genome or in a set of S. pseudoneumoniae genomes in the whole-genome shotgun database. Since this genome set include diverse isolates from disparate geographical origins and sample sources, these indels—and subsequently the efflux phenotype that provides—can be considered as an unusual feature selected by the pressure exerted by the presence of FQs. These indels would affect the stability of the stem-loop structure located downstream of the −10 extended box of the promoter and the mRNA start site. In fact, the predicted free energies for this stem-loop structure were lower for S. pseudopneumoniae isolates showing FQ efflux than for R6. The presence of these alternative structures may influence the amount of the patAB transcript under CPX-induction, as showed by RT-qPCR. Differences between 5305 PatAB and S. suis SatAB protein sequences were actually observed (84–86% similarity), where non-conservative changes are concentrated in the two first transmembrane alpha-helices (data not shown). This variability may account for subtle differences in the FQ specificity of homologs of this protein family potentially found in other streptococci.
The genome of S. pneumoniae (de la Campa et al., 2017) and of other streptococci (Martín-Galiano et al., 2017) is organized in supercoiling domains that react coordinately to changes in supercoiling. Transcriptomic studies of S. pneumoniae R6 showed that patA and patB are up-regulated in response to DNA relaxation with novobiocin, an inhibitor of DNA gyrase (Ferrándiz et al., 2010) and also by hyper-negative supercoiling resulting from the inhibition of DNA topoisomerase I by seconeolitsine (Ferrándiz et al., 2016a). Then, patA and patB genes are located in chromosomal domains that are up-regulated by global changes in DNA supercoiling. This fact may reflect the high functional weight of this pump under threatening conditions for the chromosome supercoiling state caused by xenobiotics, such as FQs. Both ethidium bromide and acriflavine, two substrates of the PatAB pump, are DNA intercalants that alter the supercoiling of the DNA, causing DNA relaxation (Ladoulis and Gill, 1970; Funatsuki et al., 1997). In addition, acriflavine has been shown to inhibit the DNA binding of gyrase (Mraheil et al., 2013). The effect of the treatment with either of these compounds would cause DNA relaxation, and, as in the case of inhibition of gyrase B with novobiocin, the transcription of the patAB operon would be enhanced. Although FQs do not change global supercoiling in S. pneumoniae, local supercoiling changes have been shown to alter the transcriptome of cultures treated with either LVX (Ferrándiz and de la Campa, 2014) or MXF (Ferrándiz et al., 2016b). Transcriptomic changes were different with these two different FQs, but, in no case, the transcription of patAB was affected. However, the transcriptional response to CPX, which also differed from that of LVX or MOX, involves the up-regulation of patAB (Marrer et al., 2006). We proposse that local changes in DNA supercoiling would affect the stability of the stem-loop structure located upstream patA (Glucksmann et al., 1992; Lionnet et al., 2006), regulating the amount of the transcripts of the downstream genes, either by regulation of transcription or by processing of the mRNA. The role of a transcriptional regulator recognizing the hairpin would be compatible with this hypothesis.
The mutations described in this study did not confer clinical FQ resistance by itself, since only conferred resistance to CPX at 4 μg/ml, in agreement with previous studies (Zhanel et al., 2004; El Garch et al., 2010; Lupien et al., 2013). However, it has been described that the presence of CPX efflux pumps favors the acquisition of further mutations in the topoisomerase genes, and that the combination of efflux and first-step mutations can lead to MICs associated with treatment failure (Jumbe et al., 2006). Then, the genetically close species S. pneumoniae and S. pseudopneumoniae, shared the same mechanisms of FQ resistance. Besides mutations in the quinolone-determining regions of their topoisomerase genes, they also shared the up-regulation of the PatAB pump as a mechanism of FQ-efflux. In the same way as mutations in the topoisomerase genes can be transferred between commensal streptococci of the mitis group and S. pneumoniae (Ferrándiz et al., 2000; Balsalobre et al., 2003), the mutations in the regulatory region of patA could be exchanged, as shown in this study, and could therefore contribute to a future increase of FQ resistance in S. pneumoniae. These results reinforce the notion that S. pseudopneumoniae can turn into an exchanger of resistance determinants to S. pneumoniae, since both organisms share ecological niche (they inhabit for long-term in the nasopharynx, Sakwinska et al., 2014), and show high nucleotide identity and sinteny of flanking regions, which would favor horizontal transfer of their genetic material.
Author Contributions
MA and MF performed the experimental work. AM and AZ analyzed genome sequences. AdlC participated in study conception, data interpretation and manuscript writing. All authors participated in data interpretation and manuscript corrections.
Funding
This study was supported by grant BIO2014-55462-R from Plan Nacional de I+D+I of the Ministry of Economy and Competitiveness. AM is the recipient of a Miguel Servet contract from the Spanish Ministry of Economy and Competitiveness.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Monica Amblar (Centro Nacional de Microbiología, ISCIII, Madrid, Spain) for critical reading of the manuscript.
References
Adam, H. J., Hoban, D. J., Gin, A. S., and Zhanel, G. G. (2009). Association between fluoroquinolone usage and a dramatic rise in ciprofloxacin-resistant Streptococcus pneumoniae in Canada, 1997-2006. Int. J. Antimicrob. Agents 34, 82–85. doi: 10.1016/j.ijantimicag.2009.02.002
Arbique, J. C., Poyart, C., Trieu-Cuot, P., Quesne, G., Carvalho Mda, G., Steigerwalt, A. G., et al. (2004). Accuracy of phenotypic and genotypic testing for identification of Streptococcus pneumoniae and description of Streptococcus pseudopneumoniae sp. nov. J. Clin. Microbiol. 42, 4686–4696. doi: 10.1128/JCM.42.10.4686-4696.2004
Balsalobre, L., and de la Campa, A. G. (2008). Fitness of Streptococcus pneumoniae fluoroquinolone-resistant strains with topoisomerase IV recombinant genes. Antimicrob. Agents Chemother. 52, 822–830. doi: 10.1128/AAC.00731-07
Balsalobre, L., Ferrándiz, M. J., de Alba, G., and de la Campa, A. G. (2011). Nonoptimal DNA topoisomerases allow maintenance of supercoiling levels and improve fitness of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 55, 1097–1105. doi: 10.1128/AAC.00783-10
Balsalobre, L., Ferrándiz, M. J., Li-ares, J., Tubau, F., and de la Campa, A. G. (2003). Viridans group streptococci are donors in horizontal transfer of topoisomerase IV genes to Streptococcus pneumoniae. Antimicrob. Agents Chemother. 47, 2072–2081. doi: 10.1128/AAC.47.7.2072-2081.2003
Balsalobre, L., Ortega, M., and de la Campa, A. G. (2013). Characterization of recombinant fluoroquinolone-resistant pneumococcus-like isolates. Antimicrob. Agents Chemother. 57, 254–260. doi: 10.1128/AAC.01357-12
Baylay, A. J., and Piddock, L. J. (2015). Clinically relevant fluoroquinolone resistance due to constitutive overexpression of the PatAB ABC transporter in Streptococcus pneumoniae is conferred by disruption of a transcriptional attenuator. J. Antimicrob. Chemother. 70, 670–679. doi: 10.1093/jac/dku449
Brenwald, N. P., Appelbaum, P., Davies, T., and Gill, M. J. (2003). Evidence for efflux pumps, other than PmrA, associated with fluoroquinolone resistance in Streptococcus pneumoniae. Clin. Microbiol. Infect. 9, 140–143. doi: 10.1046/j.1469-0691.2003.00482.x
Brenwald, N. P., Gill, M. J., and Wise, R. (1998). Prevalence of a putative efflux mechanism among fluoroquinolone-resistant clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 42, 2032–2035.
Champoux, J. J. (2001). DNA topoisomerases: structure, function, and mechanism. Annu. Rev. Biochem. 70, 369–413. doi: 10.1146/annurev.biochem.70.1.369
Chen, D. K., McGeer, A., de Azavedo, J. C., and Low, D. E. (1999). Decreased susceptibility of Streptococcus pneumoniae to fluoroquinolones in Canada. New Engl. J. Med. 341, 233–239. doi: 10.1056/NEJM199907223410403
Clinical and Laboratory Standards Institute (2008). Performance Standards for Antimicrobial Susceptibility Testing; 18th Informational Supplement. CLSI document M100-S18. Wayne, PA: Clinical and Laboratory Standards Institute.
de la Campa, A. G., Ardanuy, C., Balsalobre, L., Pérez-Trallero, E., Marimón, J. M., Fenoll, A., et al. (2009). Changes in fluoroquinolone-resistant Streptococcus pneumoniae after 7-valent conjugate vaccination, Spain. Emerg. Infect. Dis. 15, 905–911. doi: 10.3201/eid1506.080684
de la Campa, A. G., Balsalobre, L., Ardanuy, C., Fenoll, A., Pérez-Trallero, E., and Li-ares, J. (2004). Fluoroquinolone resistance in penicillin-resistant Streptococcus pneumoniae clones, Spain. Emerg. Infect. Dis. 10, 1751–1759. doi: 10.3201/eid1010.040382
de la Campa, A. G., Ferrándiz, M. J., Martín-Galiano, A. J., García, M. T., and Tirado-Vélez, J. M. (2017). The transcriptome of Streptococcus pneumoniae induced by local and global changes in supercoiling. Front. Microbiol. 8:1447. doi: 10.3389/fmicb.2017.01447
Domenech, A., Tirado-Vélez, J. M., Fenoll, A., Ardanuy, C., Yuste, J., Li-ares, J., et al. (2014). Fluoroquinolone-resistant pneumococci: dynamics of serotypes and clones in Spain in 2012 compared with those from 2002 and 2006. Antimicrob. Agents Chemother. 58, 2393–2399. doi: 10.1128/AAC.02669-13
Dowson, C. G., Hutchinson, A., Woodford, N., Johnson, A. P., George, R. C., and Spratt, B. G. (1990). Penicillin-resistant viridans streptococci have obtained altered penicillin-binding protein genes from penicillin-resistant strains of Streptococcus pneumoniae. Proc. Natl. Acad. Sci. U.S.A. 87, 5858–5862. doi: 10.1073/pnas.87.15.5858
El Garch, F., Lismond, A., Piddock, L. J., Courvalin, P., Tulkens, P. M., and Van Bambeke, F. (2010). Fluoroquinolones induce the expression of patA and patB, which encode ABC efflux pumps in Streptococcus pneumoniae. J. Antimicrob. Chemother. 65, 2076–2082. doi: 10.1093/jac/dkq287
Escudero, J. A., San Millan, A., Gutierrez, B., Hidalgo, L., La Ragione, R. M., AbuOun, M., et al. (2011). Fluoroquinolone efflux in Streptococcus suis is mediated by SatAB and not by SmrA. Antimicrob. Agents Chemother. 55, 5850–5860. doi: 10.1128/AAC.00498-11
Escudero, J. A., San Millan, A., Montero, N., Gutierrez, B., Martínez-ovejero, C., Carrilero, L., et al. (2013). SatR is a repressor of fluoroquinolone efflux pump SatAB. Antimicrob. Agents Chemother. 57, 3430–3433. doi: 10.1128/AAC.00515-13
Ferrándiz, M. J., Ardanuy, C., Li-ares, J., García-Arenzana, J. M., Cercenado, E., Fleites, A., et al. (2005). New mutations and horizontal transfer of rpoB among rifampin-resistant Streptococcus pneumoniae from four Spanish hospitals. Antimicrob. Agents Chemother. 49, 2237–2245. doi: 10.1128/AAC.49.6.2237-2245.2005
Ferrándiz, M. J., and de la Campa, A. G. (2014). The fluoroquinolone levofloxacin triggers the transcriptional activation of iron transport genes that contribute to cell death in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 58, 247–257. doi: 10.1128/AAC.01706-13
Ferrándiz, M. J., Fenoll, A., Li-ares, J., and de la Campa, A. G. (2000). Horizontal transfer of parC and gyrA in fluoroquinolone-resistant clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 44, 840–847. doi: 10.1128/AAC.44.4.840-847.2000
Ferrándiz, M. J., Martín-Galiano, A. J., Arnanz, C., Camacho-Soguero, I., Tirado-Vélez, J. M., and de la Campa, A. G. (2016a). An increase in negative supercoiling in bacteria reveals topology-reacting gene clusters and a homeostatic response mediated by the DNA topoisomerase I gene. Nucl. Acids Res. 44, 7292–7303. doi: 10.1093/nar/gkw602
Ferrándiz, M. J., Martín-Galiano, A. J., Arnanz, C., Zimmerman, T., and de la Campa, A. G. (2016b). Reactive oxygen species contribute to the bactericidal effects of the fluoroquinolone moxifloxacin in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 60, 409–417. doi: 10.1128/AAC.02299-15
Ferrándiz, M. J., Martín-Galiano, A. J., Schvartzman, J. B., and de la Campa, A. G. (2010). The genome of Streptococcus pneumoniae is organized in topology-reacting gene clusters. Nucl. Acids Res. 38, 3570–3581. doi: 10.1093/nar/gkq106
Ferrándiz, M. J., Oteo, J., Aracil, B., Gómez-Garcés, J. L., and de la Campa, A. G. (1999). Drug efflux and parC mutations are involved in fluoroquinolone resistance in viridans group streptococci. Antimicrob. Agents Chemother. 43, 2520–2523.
Funatsuki, K., Tanaka, R., Inagaki, S., Konno, H., Katoh, K., and Nakamura, H. (1997). AcrB mutation located at carboxyl-terminal region of gyrase B subunit reduces DNA binding of DNA gyrase. J. Biol. Chem. 272, 13302–13308. doi: 10.1074/jbc.272.20.13302
Garvey, M. I., Baylay, A. J., Wong, R. L., and Piddock, L. J. (2011). Overexpression of patA and patB, which encode ABC transporters, is associated with fluoroquinolone resistance in clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 55, 190–196. doi: 10.1128/AAC.00672-10
Garvey, M. I., and Piddock, L. J. (2008). The efflux pump inhibitor reserpine selects multidrug-resistant Streptococcus pneumoniae strains that overexpress the ABC transporters PatA and PatB. Antimicrob. Agents Chemother. 52, 1677–1685. doi: 10.1128/AAC.01644-07
Gill, M. J., Brenwald, N. P., and Wise, R. (1999). Identification of an efflux pump gene, pmrA, associated with fluoroquinolone resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 43, 187–189.
Glucksmann, M. A., Markiewicz, P., Malone, C., and Rothman-Denes, L. B. (1992). Specific sequences and a hairpin structure in the template strand are required for N4 virion RNA polymerase promoter recognition. Cell 70, 491–500. doi: 10.1016/0092-8674(92)90173-A
Hyatt, D., Chen, G. L., Locascio, P. F., Land, M. L., Larimer, F. W., and Hauser, L. J. (2010). Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 11:119. doi: 10.1186/1471-2105-11-119
Ip, M. C. S., Chi, F., Cheuk, E. S., Ma, H., Lai, R. W., and Chan, P. K. (2007). Longitudinally tracking of fluoroquinolone resistance and its determinants in penicillin-susceptible and -nonsusceptible Streptococcus pneumoniae isolates in Hong Kong, 2000 to 2005. Antimicrob. Agents Chemother. 51, 2192–2194. doi: 10.1128/AAC.00139-07
Jumbe, N. L., Louie, A., Miller, M. H., Liu, W., Deziel, M. R., Tam, V. H., et al. (2006). Quinolone efflux pumps play a central role in emergence of fluoroquinolone resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 50, 310–317. doi: 10.1128/AAC.50.1.310-317.2006
Lacks, S. A., López, P., Greenberg, B., and Espinosa, M. (1986). Identification and analysis of genes for tetracycline resistance and replication functions in the broad-host-range plasmid pLS1. J. Mol. Biol. 192, 753–765. doi: 10.1016/0022-2836(86)90026-4
Ladoulis, C. T., and Gill, T. J. (1970). Physical chemical studies on the specific interaction of acriflavine-phosphotungstic acid complex with double-stranded nucleic acids. J. Cell Biol. 47, 500–511. doi: 10.1083/jcb.47.2.500
Lander, E. S., and Waterman, M. S. (1988). Genomic mapping by fingerprinting random clones: a mathematical analysis. Genomics 2, 231–239. doi: 10.1016/0888-7543(88)90007-9
Liñares, J., Ardanuy, C., Pallarés, R., and Fenoll, A. (2010). Changes in antimicrobial resistance, serotypes and genotypes in Streptococcus pneumoniae over a 30-year period. Clin. Microbiol. Infect. 16, 402–410. doi: 10.1111/j.1469-0691.2010.03182.x
Lionnet, T., Dawid, A., Bigot, S., Barre, F. X., Saleh, O. A., Heslot, F., et al. (2006). DNA mechanics as a tool to probe helicase and translocase activity. Nucleic Acids Res. 34, 4232–4244. doi: 10.1093/nar/gkl451
Lupien, A., Billal, D. S., Fani, F., Soualhine, H., Zhanel, G. G., Leprohon, P., et al. (2013). Genomic characterization of ciprofloxacin resistance in a laboratory-derived mutant and a clinical isolate of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 57, 4911–4919. doi: 10.1128/AAC.00418-13
Mandell, L. A., Wunderink, R. G., Anzueto, A., Bartlett, J. G., Campbell, G. D., Dean, N. C., et al. (2007). Infectious diseases society of America/American thoracic society consensus guidelines on the management of community-acquired pneumonia in adults. Clin. Infect. Dis. 44(Suppl. 2), S27–S72. doi: 10.1086/511159
Mao, F., Dam, P., Chou, J., Olman, V., and Xu, Y. (2009). DOOR: a database for prokaryotic operons. Nucleic Acids Res. 37, D459–D463. doi: 10.1093/nar/gkn757
Marrer, E., Satoh, A. T., Johnson, M. M., Piddock, L. J., and Page, M. G. (2006). Global transcriptome analysis of the responses of a fluoroquinolone-resistant Streptococcus pneumoniae mutant and its parent to ciprofloxacin. Antimicrob. Agents Chemother. 50, 269–278. doi: 10.1128/AAC.50.1.269-278.2006
Martín-Galiano, A. J., Ferrándiz, M. J., and de la Campa, A. G. (2017). Bridging chromosomal architecture and pathophysiology of Streptococcus pneumoniae. Genome Biol. Evol. 9, 350–361. doi: 10.1093/gbe/evw299
Morrissey, I., and George, J. (1999). Activities of fluoroquinolones against Streptococcus pneumoniae type II topoisomerases purified as recombinant proteins. Antimicrob. Agents Chemother. 43, 2579–2585.
Mraheil, M. A., Heising, A., and Heising, P. (2013). An improved assay for the detection of alterations in bacterial DNA supercoiling in vivo. Pharmazie 68, 541–548. doi: 10.1691/ph.2013.6520
Muñoz, R., and de la Campa, A. G. (1996). ParC subunit of DNA topoisomerase IV of Streptococcus pneumoniae is a primary target of fluoroquinolones and cooperates with DNA gyrase A subunit in forming resistance phenotype. Antimicrob. Agents Chemother. 40, 2252–2257.
Obaro, S. K. (2000). Confronting the pneumococcus: a target or bullet change? Vaccine 19, 1211–1217. doi: 10.1016/S0264-410X(00)00262-0
Pestova, E., Millichap, J. J., Siddiqui, F., Noskin, G. A., and Peterson, L. R. (2002). Non-PmrA-mediated multidrug resistance in Streptococcus pneumoniae. J. Antimicrob. Chemother. 49, 553–556. doi: 10.1093/jac/49.3.553
Piddock, L. J., Johnson, M. M., Simjee, S., and Pumbwe, L. (2002). Expression of efflux pump gene pmrA in fluoroquinolone-resistant and -susceptible clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 46, 808–812. doi: 10.1128/AAC.46.3.808-812.2002
Riedel, S., Beekmann, S. E., Heilmann, K. P., Richter, S. S., García-de-Lomas, J., Ferech, M., et al. (2007). Antimicrobial use in Europe and antimicrobial resistance in Streptococcus pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 26, 485–490. doi: 10.1007/s10096-007-0321-5
Robertson, G. T., Doyle, T. B., and Lynch, A. S. (2005). Use of an efflux-deficient Streptococcus pneumoniae strain panel to identify ABC-class multidrug transporters involved in intrinsic resistance to antimicrobial agents. Antimicrob. Agents Chemother. 49, 4781–4783. doi: 10.1128/AAC.49.11.4781-4783.2005
Rozen, D. E., McGee, L., Levin, B. R., and Klugman, K. P. (2007). Fitness costs of fluoroquinolone resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 51, 412–416. doi: 10.1128/AAC.01161-06
Sakwinska, O., Schmid, V. B., Berger, B., Bruttin, A., Keitel, K., Lepage, M., et al. (2014). Nasopharyngeal microbiota in healthy children and pneumonia patients. J. Clin. Microbiol. 52, 1590–1594. doi: 10.1128/JCM.03280-13
Sievers, F., Wilm, A., Dineen, D., Gibson, T. J., Karplus, K., Li, W., et al. (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7:539. doi: 10.1038/msb.2011.75
Stanhope, M. J., Walsh, S. L., Becker, J. A., Italia, M. J., Ingraham, K. A., Gwynn, M. N., et al. (2005). Molecular evolution perspectives on intraspecific lateral DNA transfer of topoisomerase and gyrase loci in Streptococcus pneumoniae, with implications for fluoroquinolone resistance development and spread. Antimicrob. Agents Chemother. 49, 4315–4326. doi: 10.1128/AAC.49.10.4315-4326.2005
Tocci, N., Iannelli, F., Bidossi, A., Ciusa, M. L., Decorosi, F., Viti, C., et al. (2013). Functional analysis of pneumococcal drug efflux pumps associates the MATE DinF transporter with quinolone susceptibility. Antimicrob. Agents Chemother. 57, 248–253. doi: 10.1128/AAC.01298-12
Wilson, K. (1994). “Preparation of genomic DNA from bacteria,” in Current Protocols in Molecular Biology, ed F. M. Ausbel, R. Brent, R. E. Kingston,D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (New York, NY: Wiley, J. and sons, Inc.), 2.4.1.
World Health Organization (2007). Pneumococcal conjugate vaccine for childhood immunization-WHO position paper. Wkly. Epidemiol. Rec. 82, 93–104.
Keywords: DNA topoisomerases, efflux pumps, fluoroquinolones, transcription regulation, Streptococcus pseudopneumoniae
Citation: Alvarado M, Martín-Galiano AJ, Ferrándiz MJ, Zaballos Á and de la Campa AG (2017) Upregulation of the PatAB Transporter Confers Fluoroquinolone Resistance to Streptococcus pseudopneumoniae. Front. Microbiol. 8:2074. doi: 10.3389/fmicb.2017.02074
Received: 01 August 2017; Accepted: 10 October 2017;
Published: 26 October 2017.
Edited by:
Yuji Morita, Aichi Gakuin University, JapanReviewed by:
Françoise Van Bambeke, Université Catholique de Louvain, BelgiumBruno Gonzalez-Zorn, Complutense University of Madrid, Spain
Markus Seeger, University of Zurich, Switzerland
Copyright © 2017 Alvarado, Martín-Galiano, Ferrándiz, Zaballos and de la Campa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adela G. de la Campa, agcampa@isciii.es